
Morphological and Ecological Differences Between Shallow- and Deep-water Lake Trout in Lake...
14
J. Great Lakes Res. 33:156–169 Internat. Assoc. Great Lakes Res., 2007 156 Morphological and Ecological Differences Between Shallow- and Deep-water Lake Trout in Lake Mistassini, Quebec Mara S. Zimmerman 1,* , Charles C. Krueger 2 , and Randy L. Eshenroder 2 1 Department of Fisheries and Wildlife Michigan State University Lansing, Michigan 48823 2 Great Lakes Fishery Commission 2100 Commonwealth Blvd, Suite 100 Ann Arbor, Michigan 48105 ABSTRACT. Lake trout (Salvelinus namaycush) in Lake Mistassini, Quebec, were investigated to determine whether they resembled the lean and siscowet morphotypes of Lake Superior and Great Slave Lake. Lake trout caught in deep water were predicted to resemble the siscowet morphotype and to be bet- ter adapted for vertical migration (i.e., low percent buoyancy) than those caught in shallow water. The research objectives were to 1) identify groups based on shape, and 2) determine whether shape was asso- ciated with other morphological traits (fin length, buoyancy, color), ecology (habitat depth, diet), and life history (size at adulthood). Eighty-five lake trout were collected from three depth zones. At least two phe- notypes exist in Lake Mistassini. A shallow-water form (< 50-m depth), identified by its streamlined shape, was dark in color and high in percent buoyancy. A deep-water form (> 50-m depth), identified by a deep anterior-body profile, was light in color and lower in percent buoyancy than the shallow-water form. Absolute buoyancies were relatively high in both forms; therefore, the deep-water form did not appear well-adapted for vertical migration. Opossum shrimp (Mysis relicta) were more frequent and abundant in stomachs of deep-bodied trout. All deep-bodied trout (minimum 32-cm SL) had reached adulthood, whereas immature streamlined individuals were as long as 49 cm in SL. The deep-bodied form resembled humper lake trout, a lesser-known third morphotype from Lake Superior. A humper-like mor- photype in Lake Mistassini, and the apparent absence of a siscowet-like morphotype, challenges the pre- viously-held hypothesis that humpers resulted from an introgression of leans and siscowets. INDEX WORDS: Adaptation, buoyancy regulation, humper, lean, phenotypic diversity, siscowet. * Corresponding author. E-mail: [email protected] Address correspondence to Great Lakes Fishery Commission, 2100 Commonwealth Blvd., Suite 100, Ann Arbor, MI 48105-1563 INTRODUCTION An abundance of available niches is hypothesized to have facilitated morphological divergence in fish species colonizing recently deglaciated lakes (Sku- lason and Smith 1995, Smith and Skulason 1996). In at least eight fish families, resource polymor- phism is attributed to divergent selection in benthic and pelagic habitats (Robinson and Wilson 1994, Smith and Skulason 1996). A less-recognized eco- logical variable, habitat depth, is also associated with the radiation of freshwater fishes (Eshenroder et al. 1999, Turgeon et al. 1999, Power et al. 2005). For example, lake trout ( Salvelinus namaycush) phenotypic diversity in the Laurentian Great Lakes once included shallow- and deep-water forms in Lakes Superior, Michigan, Huron, and Erie (Brown et al. 1981, Goodier 1981, Krueger and Ihssen 1995). However, this diversity was lost from Lakes Michigan, Huron, and Erie during the past century. Some of the original phenotypic diversity remains in Lake Superior (maximum depth ~400 m) in forms known as lean, humper, and siscowet lake trout (Khan and Qadri 1970, Lawrie and Rahrer 1973, Moore and Bronte 2001). The origin and maintenance of lake trout pheno- typic diversity has been difficult to investigate due to the loss of diversity from most of the Laurentian Great Lakes and the highly disturbed nature of re- maining communities. Therefore, recent lake trout
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Morphological and Ecological Differences Between Shallow- and Deep-water Lake Trout in Lake...

J. Great Lakes Res. 33:156–169Internat. Assoc. Great Lakes Res., 2007
156
Morphological and Ecological Differences Between Shallow- andDeep-water Lake Trout in Lake Mistassini, Quebec
Mara S. Zimmerman1,*, Charles C. Krueger2, and Randy L. Eshenroder2
1Department of Fisheries and WildlifeMichigan State UniversityLansing, Michigan 48823
2Great Lakes Fishery Commission2100 Commonwealth Blvd, Suite 100
Ann Arbor, Michigan 48105
ABSTRACT. Lake trout (Salvelinus namaycush) in Lake Mistassini, Quebec, were investigated todetermine whether they resembled the lean and siscowet morphotypes of Lake Superior and Great SlaveLake. Lake trout caught in deep water were predicted to resemble the siscowet morphotype and to be bet-ter adapted for vertical migration (i.e., low percent buoyancy) than those caught in shallow water. Theresearch objectives were to 1) identify groups based on shape, and 2) determine whether shape was asso-ciated with other morphological traits (fin length, buoyancy, color), ecology (habitat depth, diet), and lifehistory (size at adulthood). Eighty-five lake trout were collected from three depth zones. At least two phe-notypes exist in Lake Mistassini. A shallow-water form (< 50-m depth), identified by its streamlinedshape, was dark in color and high in percent buoyancy. A deep-water form (> 50-m depth), identified bya deep anterior-body profile, was light in color and lower in percent buoyancy than the shallow-waterform. Absolute buoyancies were relatively high in both forms; therefore, the deep-water form did notappear well-adapted for vertical migration. Opossum shrimp (Mysis relicta) were more frequent andabundant in stomachs of deep-bodied trout. All deep-bodied trout (minimum 32-cm SL) had reachedadulthood, whereas immature streamlined individuals were as long as 49 cm in SL. The deep-bodied formresembled humper lake trout, a lesser-known third morphotype from Lake Superior. A humper-like mor-photype in Lake Mistassini, and the apparent absence of a siscowet-like morphotype, challenges the pre-viously-held hypothesis that humpers resulted from an introgression of leans and siscowets.
INDEX WORDS: Adaptation, buoyancy regulation, humper, lean, phenotypic diversity, siscowet.
*Corresponding author. E-mail: [email protected] correspondence to Great Lakes Fishery Commission, 2100Commonwealth Blvd., Suite 100, Ann Arbor, MI 48105-1563
INTRODUCTION
An abundance of available niches is hypothesizedto have facilitated morphological divergence in fishspecies colonizing recently deglaciated lakes (Sku-lason and Smith 1995, Smith and Skulason 1996).In at least eight fish families, resource polymor-phism is attributed to divergent selection in benthicand pelagic habitats (Robinson and Wilson 1994,Smith and Skulason 1996). A less-recognized eco-logical variable, habitat depth, is also associatedwith the radiation of freshwater fishes (Eshenroderet al. 1999, Turgeon et al. 1999, Power et al. 2005).
For example, lake trout (Salvelinus namaycush)phenotypic diversity in the Laurentian Great Lakesonce included shallow- and deep-water forms inLakes Superior, Michigan, Huron, and Erie (Brownet al. 1981, Goodier 1981, Krueger and Ihssen1995). However, this diversity was lost from LakesMichigan, Huron, and Erie during the past century.Some of the original phenotypic diversity remainsin Lake Superior (maximum depth ~400 m) informs known as lean, humper, and siscowet laketrout (Khan and Qadri 1970, Lawrie and Rahrer1973, Moore and Bronte 2001).
The origin and maintenance of lake trout pheno-typic diversity has been difficult to investigate dueto the loss of diversity from most of the LaurentianGreat Lakes and the highly disturbed nature of re-maining communities. Therefore, recent lake trout

Phenotypic Diversity of Lake Trout in Lake Mistassini 157
studies have been conducted on other large, deepNorth American lakes. Descriptions of lake troutcommunities in Great Bear (maximum depth ~450m) and Great Slave (maximum depth ~600 m) lakeshave provided a broader perspective on the associa-tion of lake trout phenotypes with ecological vari-ables such as habitat depth. In Great Slave Lake, forexample, a siscowet-like trout with high-fat-contenttissue recently was caught in deep-water habitatsmore than 50 m in depth (Zimmerman et al. 2006).In Great Bear Lake, deep-water forms have notbeen identified; however, shallow-water trout areexceptionally diverse in their morphology (Blackieet al. 2003, Alfonso 2004). Further, inter-lake com-parisons of large, deep lakes would provide a betterunderstanding of lake trout phenotypic diversityand its association with deep-water habitat.
The siscowet phenotype from Lake Superior hashigh-lipid tissue (Eschmeyer and Phillips 1965),which may be an adaptation for buoyancy regula-tion in deep-water habitats (Thurston 1962, Hender-son and Anderson 2002). The compressed swimbladder of a vertically migrating fish, when the fishhas descended, is ineffective at regulating buoyancy(Alexander 1972, 1993). High-lipid content lowersthe density of body tissue in water and, conse-quently, minimizes the energetic costs of swimmingwith a compressed swim bladder (Henderson andAnderson 2002). Diel vertical migrations have beenobserved in siscowet predators (Hrabik et al. 2006,Jensen et al. 2006) as well as in deep-water lacus-trine prey such as bloater Coregonus hoyi (Eshen-roder et al. 1998, TeWinkel and Fleischer 1999) andopossum shrimp Mysis relicta (Beeton 1960, Bow-ers 1988). The ubiquity of a siscowet-like pheno-type in large, deep North American lakes wouldsuggest that this form is best adapted to deep-waterliving and to foraging on vertically migrating preysuch as deep-water coregonines.
A second deep-water form, the humper trout, isfar less abundant than siscowets in Lake Superior(Peck 1975). Humpers have been considered to bea “form” (Eschmeyer and Phillips 1965) or a “mor-phological variant” (Lawrie and Rahrer 1973, Burn-ham-Curtis and Bronte 1996) of the lean trout andhave been hypothesized to have resulted from theintrogression of lean and siscowet trout during a pe-riod of fluctuating water levels (Burnham-Curtis1993, Burnham-Curtis and Smith 1994). If humper-like trout occur as distinct populations outside LakeSuperior in the absence of the siscowet form, theexisting hypothesis on their origin would need to bereconsidered.
The purpose of this study was to quantitativelydescribe the phenotypic diversity of lake trout inLake Mistassini, a large, deep lake (maximumdepth > 150 m) in central Quebec. The abundanceof deep-water habitat in this lake was proportion-ally comparable to other lakes where lake trout phe-notypic diversity was known; however, this lakewas smaller in surface area (2,150 km2) than LakeSuperior (82,100 km2) or Great Slave Lake (28,568km2) (Beeton 1984). Two hypotheses were tested inthis study. First, lake trout occupying deep-waterhabitat are predicted to have lower percent buoy-ancy, and thus be better adapted for vertical migra-tion, than those occupying shallow-water habitat.Second, deep-water forms, if they exist, will likelyresemble the shape and buoyancy of siscowet troutfrom Lake Superior. The research objectives wereto 1) identify lake trout groups based on shape, and2) determine whether lake trout shape was associ-ated with other morphological traits (fin length,buoyancy, color), ecology (habitat depth, diet), andlife history (size of adulthood).
MATERIAL AND METHODS
Study Site
Lake Mistassini (50º40′N 73º20′W) is the largestnatural lake (2,150 km2) in the Province of Quebec(Fig. 1) and is located east of all other lakes wherecoexisting lake trout morphotypes are known. Thelake has east and west arms separated by a long is-land chain. Each arm has steep, rocky shorelinesand abundant deep-water habitat (> 50-m depths).The food web includes typical lake trout prey suchas opossum shrimp (Dadswell 1974), cisco Core-gonus artedi, and shortjaw cisco C. zenithicus. Rep-resentative Coregonus specimens are held in theUniversity of Michigan Museum of Zoology FishDivision (C. artedi Cat. No. 247005, C. zenithicusCat. No. 247006). Sport fishing in Lake Mistassiniis supported by abundant lake trout, brook trout (S.fontinalis), walleye (Sander vitreus), and northernpike (Esox lucius) populations. Additional humanimpact is minimized by a provincial wildlife re-serve that surrounds most of the lake (Fraser andBernatchez 2005).
Field Procedures
Lake trout were caught with gill-nets set in theeast and west arms of Lake Mistassini during Au-gust 2003 (Fig. 1). Gill-net mesh sizes ranged from64-114-mm stretch measure. Of the 12 gill-net sets,

158 Zimmerman et al.
four were in water depths of 0-50 m, five in 50–100m, and three in 100–150 m. Sets were overnightand approximated a 24-hour soak time. Depth stratawere established based on the known depths of laketrout forms in Lake Superior (Moore and Bronte2001, Bronte et al. 2003). The data collected in thefield from each individual included a full-body pho-tograph, air weight, water weight with swim blad-der deflated (for estimating buoyancy), sex, andreproductive state. Stomachs were collected andtheir contents fixed in 10% formalin.
The lateral image of each lake trout was capturedin a full-body photograph as described in Zimmer-man et al. (2006). Digital-image files were used toquantify color, fin lengths, and body shape. Percentbuoyancy was measured as water weight divided byair weight (Zimmerman et al. 2006). Each fish wassuspended from a Pesola spring scale (Jennings
1989) with a hook and weighed to the nearest gramin air and then in water. Prior to measuring waterweight, any remaining swim-bladder gas was ex-pelled through a longitudinal incision. The buoy-ancy measure accounted for the differences in softand hard tissues that affect the specific gravity oflake trout tissue (Alexander 1972). As lipids havelower specific gravity than water, tissue with highlipid content will have a low percent buoyancy.
All trout were assessed for sex and reproductivestate. Juvenile trout had undeveloped (i.e., imma-ture) gonads. Adult trout were those whose gonadseither were greatly enlarged or were in a so-calledresting state from a previous spawning event. Rest-ing females had discernable though small (< 1.0mm in diameter) ova, whereas immature females(with which they could be confused) had ova dis-cernable only under magnification. Resting-stage
FIG. 1. Location of 12 sites where lake trout were collected from Lake Mistassini, Quebec.

Phenotypic Diversity of Lake Trout in Lake Mistassini 159
males had testes that were slightly enlarged,whereas the entire length of the testis was undevel-oped in immature fish.
Measures of Color, Fins, Shape, and Diet
Brightness, one measure of body coloration, wasquantified from each image using a 255-unit grayscale (0 = black, 255 = white) and ImageJ v. 1.32software (http://rsb.info.nih.gov/ij). Brightness wasaveraged for six measures from each lake trout’slateral image (cheek, three dorsal, two ventral)based on a 40 × 40 pixel square of each region.Lake trout were excluded from color analysis iftheir body coloration became mottled or bleachedafter collection.
Fin-length measurements (Fig. 2a) were identicalto those used by Zimmerman et al. (2006) on laketrout from Great Slave Lake; comparable measuresfrom Lake Superior were not available. Body shapewas measured using 16 landmarks and four semi-landmarks (Fig. 2b). The 16 landmarks were con-sistent with lake trout studies on Great Slave Lake(Zimmerman et al. 2006) and Lake Superior(Moore and Bronte 2001). The four semi-landmarksdescribed the belly curvature at 20, 30, 40, and 50%standard length (SL) (Sampson et al. 1996, Langer-hans et al. 2003). Although semi-landmarks are nothomologous, and thus less valuable than landmarksfor shape comparisons (Zelditch et al. 2004), theywere included because they described a variable re-gion between the pectoral and pelvic fins where ho-mologous landmarks do not exist.
The landmark-based geometric morphometricmethod was used to quantify body shape (Book-stein 1991, Zelditch et al. 2004). This method cal-culated shape variables from digitized coordinates(Zimmerman et al. 2006). To better approximatehomology, semi-landmarks were further adjusted by“sliding” along the contour defined by the foursemi-landmarks and two landmarks representingstandard length (1 and 10, Fig. 2b) (Zelditch et al.2004). Sliding minimized the Euclidean distancebetween corresponding semi-landmarks (Sampsonet al. 1996). Lake trout were excluded from theanalysis if fins were bent or broken or if the bodywas bent or otherwise distorted in the photographs.
The x,y coordinates that captured fin length andbody shape were digitized using tpsDIG software(http://life.bio.sunysb.edu/morph). Fin lengths werecalculated using TMorphGen software, shape vari-ables were calculated using CoordGen software,and semi-landmarks were adjusted using SemiLand
software. TMorphGen, CoordGen, and SemiLandare part of a series of Integrated Morphometric Pro-grams (IMP) produced in MatLab6 (Mathworks2000) and used for morphometric analysis(http://www2.canisius.edu/~sheets/morphsoft.html).
Stomach data included prey identity and preyabundance. Identification of diet items was made tothe lowest taxonomic level possible based on thecondition of partially digested prey. For the purposeof analysis, prey were classified as terrestrial insect,aquatic invertebrate, or fish. Fish were combined
FIG. 2. Fin lengths (a) and landmarks (b) usedto compare lake trout from Lake Mistassini.Length (a) was measured from the insertion pointto the tip of the first fin ray for the dorsal (1), anal(3), pelvic (4), and pectoral (5) fins. Caudal fin (2)was measured as the longest dorsal ray. Land-marks (b) were anterior tip of the snout (1), poste-rior tip of the maxilla (2), center of the eye (3), topof the cranium (4), posterior of neurocraniumabove top of opercle (5), anterior insertion of dor-sal fin (6), posterior insertion of dorsal fin (7),anterior insertion of adipose fin (8), dorsal inser-tion of caudal fin (9), midpoint of hypural plate(10), ventral insertion of caudal fin (11), posteriorinsertion of anal fin (12), anterior insertion ofanal fin (13), insertion point of pelvic fin (14),insertion point of pectoral fin (15), and connec-tion between gill covers (16), and 17-20 are semi-landmarks (*) representing belly curvature at 20,30, 40, and 50% of lake trout standard length.

160 Zimmerman et al.
into a single category for analysis because the di-gested state of most prey fish obscured an accurateidentification to the genera level and thus loweredthe sample size for statistical comparison.
Identification of Lake Trout Phenotype Groups
An unweighted-pair-group method with arith-metic mean (UPGMA) cluster analysis based onshape variables was inspected for groups (Objective1; MVSP v. 3.1, Kovach Computing Services 2003).Prior to analysis, shape variables were adjusted to acommon size (using Standard6, a program in theIMP series). Shape variables were partial warpscores that have the correct number of degrees offreedom for multivariate tests (Zelditch et al. 2004).Groupings identified in the cluster analysis wereevaluated with a canonical variate analysis (CVA).Robustness of the groups was evaluated with a jack-knifing procedure that randomly removed 10% ofthe sample, recalculated the canonical function, re-assigned individuals to groups based on the recalcu-lated function, and computed the rate of correctlyclassified individuals over 500 trials (Nolte andSheets 2005). The CVA was performed usingCVAGen, a program in the IMP series.
Phenotypic Measures Associated withLake Trout Shape
Two approaches, univariate and multivariate,were used to describe how shape was associatedwith other phenotypic measures (Objective 2). Theunivariate approach, through a series of parametricand nonparametric tests, provided an empirical de-scription of differences between identified shapegroups. The multivariate approach, a two-block par-tial-least-squares analysis (2B-PLS), identifiedwhether and how shape covaried with multiplemeasures of phenotype (Rohlf and Corti 2000,Ruber and Adams 2001).
Lake trout groups, identified in the cluster analy-sis based on shape measures, were tested for differ-ences in buoyancy, coloration, fin lengths, capturedepth, diet, and reproductive maturity. These mea-sures were selected because they were known todiffer among co-existing shallow- and deep-watermorphotypes in Lake Superior and Great SlaveLake. An analysis of covariance (ANCOVA) testedwhether buoyancy and brightness differed betweengroups. A multivariate analysis of covariance(MANCOVA) tested whether fin lengths (dorsal,caudal, anal, pelvic, and pectoral) differed between
groups. Group and sex were the explanatory factorsand standard length was the covariate in theseanalyses. Fin and standard lengths were log10 trans-formed prior to analysis. Except where noted in theresults, models met the assumption of homoge-neous slopes. Significant multivariate effects wereinterpreted from standardized-canonical-functioncoefficients derived from a discriminant functionanalysis. A G-test of independence determinedwhether lake trout groups differed in depth distribu-tion and in the number of empty versus full stom-achs. Separate G-tests for goodness-of-fit testedwhether the presence of terrestrial insect, aquaticinvertebrate, and fish prey differed between forms.Separate Mann-Whitney U tests determinedwhether the abundance of terrestrial insects, aquaticinvertebrates, and fish differed between groups. AG-test of independence examined whether the dis-tribution of juvenile and adult trout differed be-tween groups, and a two-way analysis of variance(ANOVA) tested whether size (SL) of adult laketrout differed between groups or sexes.
Two two-block partial-least-squares analyseswere performed. The first analysis tested the co-variation of shape with buoyancy, brightness, andcapture depth. The second analysis tested the co-variation between shape and fin lengths. In bothanalyses, Block 1 included shape variables stan-dardized to a common centroid size. In the firstanalysis, Block 2 included buoyancy, brightness,and capture-depth measures. In the second analysis,Block 2 included all five fin lengths. Block 2 mea-sures were size-adjusted (except capture depth) andstandardized to a z-distribution. Size-adjusted mea-sures were the residuals from each measure re-gressed on SL (fin lengths and SL were log10transformed prior to the regression). The 2B-PLScalculated vector pairs that best explained the co-variation between the two blocks and that were de-scribed based on the weighting of each contributingvariable (similar to interpretation of principal com-ponent axes). Permutation tests determined whetherthe identified vector pairs explained more covari-ance between blocks than expected by chance andwhether the existing correlations within identifiedvector pairs were less likely to occur than random.The 2B-PLS analysis was conducted with PLS-Maker, a program in the IMP series.
Unless specifically mentioned, statistical analy-ses were performed using SPSS 12.0 (SPSS 2003).Results were considered significant when p < 0.05.

Phenotypic Diversity of Lake Trout in Lake Mistassini 161
RESULTS
A total of 85 lake trout were analyzed from the12 gill-nets set in the three depth zones (Mean, 10trout/set). Forty-nine lake trout, ranging in lengthfrom 28 to 71-cm SL (Mean ± 1 SD, 45.2 ± 6.9),were caught in the 0-50-m depth stratum. Twenty-seven lake trout, ranging in length from 34 to 45-cm SL (Mean ± 1 SD, 39.6 ± 3.1), were caught inthe 50-100-m depth stratum. In the 100-150-mdepth stratum, nine trout were caught and ranged inlength from 32 to 43-cm SL (Mean ± 1 SD, 37.9 ±3.9). Two individuals were length outliers (> 3 stan-dard deviations from the mean) and at least 12-cmlonger than any other trout collected. These individ-uals were removed from the statistical analyses be-cause they biased the size-adjustment procedure
and consequently affected other results of thisstudy.
Groupings of Lake Trout Phenotypes
Two major groups were identified from theUPGMA cluster analysis (Fig. 3). Group 1 had astreamlined body profile, a long upper jaw, and aneye positioned centrally on the head. Group 2 had adeep anterior-body profile, a narrow caudal pedun-cle, a short upper jaw, and an eye positioned dor-sally high on the head (Canonical variate 1: λ =0.18, χ2 = 109, df = 36, P < 0.001). The canonicalfunction discriminating the groups correctly classi-fied 100% of individuals originally assigned toGroup 1 and 98% of those originally assigned to
FIG. 3. Dendrogram produced by a UPGMA cluster analysis based on shapevariables calculated for Lake Mistassini lake trout. Each terminal line representsan individual fish. Outlines were drawn from vector plots of the first canonicalaxis, which discriminated between Groups 1 and 2. Shapes were standardized to 42-cm standard length. Images are representative lake trout from each group.

162 Zimmerman et al.
Group 2. Eight-six percent of individuals were cor-rectly classified when a jack-knifing procedure wasapplied to the canonical function. Representativesof the deep-bodied form were placed in the Univer-sity of Michigan Museum of Zoology (Cat. No.247004).
Phenotypic Measures Associated withLake Trout Shape
Brightness differed between forms (F1,47 = 19.6,P < 0.001). On average, a 42-cm SL lake trout ofthe deep-bodied (Group 2) form was 45% lighter incolor than the streamlined (Group 1) form (Table1). Brightness did not differ between males and fe-males in either group (F1,47 = 0.6, P = 0.4), and theinteraction between sex and form was not signifi-cant (F1,47 = 0.8, P = 0.4). Within each form, bodycoloration darkened with length (F1,47 = 13.3, P =0.001).
Buoyancy differed between forms (F1,51 = 4.8, P= 0.03). However, this difference was slight and therange of buoyancies represented in each groupoverlapped substantially (Group 1 = 4.2–7.1%,Group 2 = 4.5–6.5%). On average, a 42-cm laketrout of the deep-bodied form was just 0.3% lighter(i.e., % buoyancy was lower) than a similar-sizedstreamlined form (Table 1). Buoyancy of males andfemales did not differ in either group (F1,51 = 2.1, P= 0.2), and the interaction between sex and formwas not significant (F1,51 = 0.7, P = 0.9). Withineach form, percent buoyancy increased with length(F1,51 = 11.4, P = 0.001).
Size-adjusted fin lengths differed between thetwo lake trout groups (F5,62 = 4.5, P = 0.002) andsex (F5,62 = 4.6, P = 0.001). The function that dis-criminated between forms was heavily weighted by
the caudal fin (1.48), which was longer in thestreamlined form, and by the anal fin (-0.82), whichwas longer in the deep-bodied form (Wilks λ =0.76, χ2 = 18, df = 5, P = 0.003). The magnitude ofdifference was small; caudal and anal fins differedless than 0.3 cm between streamlined and deep-bodied trout (Table 1). In comparison to the caudaland anal fins, group differences were minimally in-fluenced by dorsal, pelvic, and pectoral fin lengths;standardized-canonical-function coefficients were–0.50, –0.35, and 0.26 respectively. Fin lengthswere size-adjusted prior to analysis because the re-lationships between fin and standard length differedfor male and female trout (sex by SL interaction:F5,58 = 5.9, P < 0.001). Small males had shorter finsthan similar-sized females and large males hadlonger fins than large females. Fin lengths wereequivalent (slopes intersected) at approximately 42cm (SL). The size-adjustment was performed usingresiduals from the regression of each fin length ver-sus standard length calculated separately for maleand female trout. Residuals from these regressionswere added to the predicted fin lengths of a 42-cmlake trout (Reist 1985).
The deep-bodied form occupied deeper habitatsthan the streamlined form (Table 1). The stream-lined form (Group 1) represented a higher propor-tion of those caught in the 0–50-m depth stratumwhereas the deep-bodied form (Group 2) repre-sented a higher proportion of those caught in the50–100-m and the 100–150-m depth strata (Fig. 4,G = 24.4, df = 2, P < 0.001).
Opossum shrimp occurred more frequently (G =4.07, df = 1, P = 0.04) and were more abundant (U = 161, P = 0.007) in the stomachs of the deep-bodied than in the stomachs of streamlined laketrout (Fig. 5). Terrestrial insects did not differ be-tween forms in percent occurrence (G = 2.47, df = 1, P = 0.12) or abundance (U = 204, P = 0.06).Similarly, fish prey did not differ in occurrence (G = 0.73, df = 1, P = 0.39) or abundance (U = 237,P = 0.34) in the stomachs of the two lake troutforms. Empty stomachs were more common in thestreamlined (35%) than the deep-bodied (11%)form (G = 4.87, df = 1, P = 0.03). Of particularnote, terrestrial insects were found in the stomachsof 13 deep-bodied trout; ten of which were caughtin water deeper than 80 m. Terrestrial insects in thediet included Hymenoptera and Coleoptera. Aquaticinvertebrates in the diet were primarily opossumshrimp; one stomach also contained Ephemeropteranymphs. Fish prey included sculpins (Cottidae),
TABLE 1. Phenotypic differences between thestreamlined (Group 1) and deep-bodied (Group 2)forms of lake trout in Lake Mistassini. Brightness,buoyancy, caudal fin, and anal fin measures arecalculated for a 42-cm SL lake trout. Data aremeans (± 1 S.E.).
Group 1 Group 2Measure (streamlined) (deep-bodied)
Brightness 85.2 (6.2) 123.5 (3.4)Buoyancy 5.7 (0.1) 5.4 (0.1)Caudal fin 8.0 (0.1) 7.7 (0.1)Anal fin 5.8 (0.1) 5.9 (0.1)Depth (m) 38.8 (3.5) 72.3 (4.3)Adult size (cm SL) 43.3 (0.8) 41.3 (0.7)

Phenotypic Diversity of Lake Trout in Lake Mistassini 163
sticklebacks (Pungitius pungitius), ciscoes (Core-gonus spp.), and charr (Salvelinus spp.).
Catch of the streamlined form included propor-tionally more juveniles (i.e., immature) than thedeep-bodied form (Fig. 6, G = 14.2, df = 1, P <0.001) even though the size ranges for deep-bodied(34–51-cm SL) and streamlined (28-53-cm SL) laketrout caught in the gillnets were similar. Of the tenjuveniles in the streamlined group, the longest was49 cm (SL). All lake trout in the deep-bodied group
were adults, including the 34-cm smallest individ-ual. On average, lake trout in the deep-bodied group(all adults) were 2-cm shorter than adults in thestreamlined group (F1,70 = 4.6, P = 0.04, Table 1).Length did not differ between adult male and fe-male trout in either group (F1,70 = 0.1, P = 0.8), andno interaction existed between sex and group (F1,70= 3.1, P = 0.08). Of note, length comparisons didnot include the two outlier lake trout which wereboth longer than 65 cm (SL).
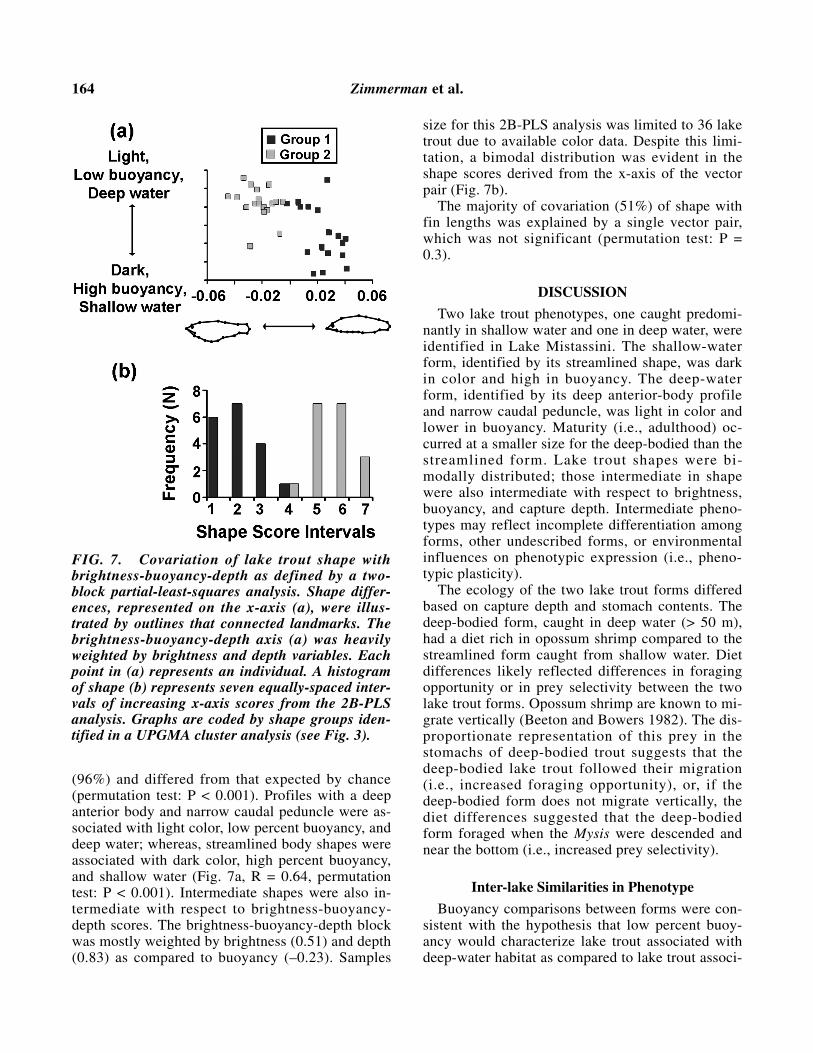
Covariation of shape with brightness, buoyancy,and depth was explained by a single vector pair
FIG. 4. Relative abundance of lake trout formsin three depth strata in Lake Mistassini. Graphshows the proportion of each form caught in eachdepth strata. The streamlined (Group 1) and deep-bodied (Group 2) forms were identified by aUPGMA cluster analysis (see Fig. 3).
FIG. 5. Frequency of occurrence (a) and abundance (b) of prey items inthe stomachs of streamlined (Group 1) and deep-bodied (Group 2) laketrout.
FIG. 6. Reproductive status of lake trout in threelength categories. Streamlined (Group 1) anddeep-bodied (Group 2) forms are plotted sepa-rately. Graph shows the proportion of adult tototal fish in each length category.
164 Zimmerman et al.
(96%) and differed from that expected by chance(permutation test: P < 0.001). Profiles with a deepanterior body and narrow caudal peduncle were as-sociated with light color, low percent buoyancy, anddeep water; whereas, streamlined body shapes wereassociated with dark color, high percent buoyancy,and shallow water (Fig. 7a, R = 0.64, permutationtest: P < 0.001). Intermediate shapes were also in-termediate with respect to brightness-buoyancy-depth scores. The brightness-buoyancy-depth blockwas mostly weighted by brightness (0.51) and depth(0.83) as compared to buoyancy (–0.23). Samples
size for this 2B-PLS analysis was limited to 36 laketrout due to available color data. Despite this limi-tation, a bimodal distribution was evident in theshape scores derived from the x-axis of the vectorpair (Fig. 7b).
The majority of covariation (51%) of shape withfin lengths was explained by a single vector pair,which was not significant (permutation test: P =0.3).
DISCUSSION
Two lake trout phenotypes, one caught predomi-nantly in shallow water and one in deep water, wereidentified in Lake Mistassini. The shallow-waterform, identified by its streamlined shape, was darkin color and high in buoyancy. The deep-waterform, identified by its deep anterior-body profileand narrow caudal peduncle, was light in color andlower in buoyancy. Maturity (i.e., adulthood) oc-curred at a smaller size for the deep-bodied than thestreamlined form. Lake trout shapes were bi-modally distributed; those intermediate in shapewere also intermediate with respect to brightness,buoyancy, and capture depth. Intermediate pheno-types may reflect incomplete differentiation amongforms, other undescribed forms, or environmentalinfluences on phenotypic expression (i.e., pheno-typic plasticity).
The ecology of the two lake trout forms differedbased on capture depth and stomach contents. Thedeep-bodied form, caught in deep water (> 50 m),had a diet rich in opossum shrimp compared to thestreamlined form caught from shallow water. Dietdifferences likely reflected differences in foragingopportunity or in prey selectivity between the twolake trout forms. Opossum shrimp are known to mi-grate vertically (Beeton and Bowers 1982). The dis-proportionate representation of this prey in thestomachs of deep-bodied trout suggests that thedeep-bodied lake trout followed their migration(i.e., increased foraging opportunity), or, if thedeep-bodied form does not migrate vertically, thediet differences suggested that the deep-bodiedform foraged when the Mysis were descended andnear the bottom (i.e., increased prey selectivity).
Inter-lake Similarities in Phenotype
Buoyancy comparisons between forms were con-sistent with the hypothesis that low percent buoy-ancy would characterize lake trout associated withdeep-water habitat as compared to lake trout associ-
FIG. 7. Covariation of lake trout shape withbrightness-buoyancy-depth as defined by a two-block partial-least-squares analysis. Shape differ-ences, represented on the x-axis (a), were illus-trated by outlines that connected landmarks. Thebrightness-buoyancy-depth axis (a) was heavilyweighted by brightness and depth variables. Eachpoint in (a) represents an individual. A histogramof shape (b) represents seven equally-spaced inter-vals of increasing x-axis scores from the 2B-PLSanalysis. Graphs are coded by shape groups iden-tified in a UPGMA cluster analysis (see Fig. 3).

Phenotypic Diversity of Lake Trout in Lake Mistassini 165
ated with shallow-water habitat. Correlationsamong buoyancy, body depth, and capture depth inLake Mistassini paralleled observations from LakeSuperior and Great Slave Lake; however, buoyancydifferences between Lake Mistassini forms wereslight. In Lake Superior, high-fat, deep-bodied sis-cowet trout were present in deeper water than low-fat, streamlined lean trout, and humper trout wereintermediate in fat content and depth (Eschmeyerand Phillips 1965, Moore and Bronte 2001, Bronteet al. 2003). A similar trend was reported fromGreat Slave Lake; low-percent-buoyancy deep-bodied trout were caught in deeper water than high-percent-buoyancy streamlined trout (Zimmerman etal. 2006). The water-depth and body-shape associa-tion evident in all three lakes was also reported forLakes Michigan and Huron in the 19th and early20th centuries (Brown et al. 1981, Goodier 1981,Eshenroder et al. 1995).
Buoyancy and fin length differences between theshallow- and deep-water forms in Lake Mistassiniwere slight in comparison with the differences ob-served in lake trout from Great Slave Lake (Zim-merman et al. 2006). The buoyancy differencebetween streamlined and deep-bodied forms inGreat Slave Lake was 11 times larger in magnitudethan in Lake Mistassini. The lightest deep-bodiedindividual in Great Slave Lake was almost neutrallybuoyant (buoyancy = 0.5%). In Lake Mistassini, thelightest deep-bodied individual was negativelybuoyant (3.8%) and near the upper buoyancy limitof the deep-water phenotype in Great Slave Lake.The pectoral-fin length difference between stream-lined and deep-bodied forms reported from GreatSlave Lake was not observed in Lake Mistassini.Existing caudal and anal fin differences betweenthe Lake Mistassini forms were small enough inmagnitude that they were unlikely to influenceswimming performance.
The deep-bodied form in Lake Mistassini was notwell adapted for daily vertical migration behaviorin comparison to deep-water trout in Lake Superioror Great Slave Lake. Low percent buoyancy, ob-served in Great Slave Lake and Lake Superior, andlong paired-fins, observed in Great Slave Lake, canfunction as buoyancy aids when the swim bladder iscompressed after rapid descents (Henderson andAnderson 2002). However, low percent buoyancy(i.e., high lipid content) has a high energetic costand, in the absence of vertical migratory behavior,buoyancy regulation is more efficiently accom-plished by changes in swim bladder volume(Alexander 1972, Henderson and Anderson 2002).
Based on tissue density and fin data, we expect thatthe deep-bodied form in Lake Mistassini does notundergo extensive vertical migrations, or, if it does,its migrations are more energetically costly thanthose of siscowets in Lake Superior or siscowet-liketrout in Great Slave Lake. Restriction of the deep-bodied form to deep waters was further supportedby the lack of local traditional ecological knowl-edge regarding the existence of the deep-water form(Andrew Coon, Kenny Longchamp, and Eric Coon-Come, Council of the Cree Nation of Mistissini,personal communications).
Given that morphological and buoyancy datasuggest the deep-bodied form was not well adaptedfor vertical migration, the presence of terrestrial in-sects in their stomachs was perplexing. Terrestrialinsects might indicate surface feeding, and surfacefeeding required vertical movements of greater than80 m for most of the deep-bodied trout. Alterna-tively, deep-bodied trout may capture insects, aftersinking, at or near the lakebed. Further study of theswimming movements and trophic position of thelake trout forms in Lake Mistassini is needed to bet-ter understand the foraging behavior and the ener-getic trade-offs unique to these deep-water laketrout.
Distinctiveness of Lake Mistassini Forms
The size and brightness of the streamlined dark-colored form in Lake Mistassini were distinct fromlean-like trout occupying similar depth strata inLake Superior and Great Slave Lake. In size(length), they were more similar to planktivorouspopulations in inland lakes than to piscivorous leantrout observed in large lakes (Martin 1966, Konkleand Sprules 1986). An exception to this observationwere the two large lean-like trout (> 65-cm SL) re-moved from the statistical analyses as length out-liers. Both of these trout had the robust appearanceof piscivorous trout found in the Great Lakes andmay represent a cannibalistic morph of the lean-liketrout (Martin and Olver 1980). The dark color ofthe streamlined form, while not unique to LakeMistassini (Hallock 1877, Goode 1884, Koelz1926, Gallagher 2002), was more difficult to inter-pret as numerous variables contribute to color vari-ation in fishes. Background matching is awell-recognized occurrence in fishes (van der Salmet al. 2005) but not a likely explanation in LakeMistassini; the differences between forms were op-posite from the theoretically expected light-in-shal-low-water and dark-in-deep-water patterns that

166 Zimmerman et al.
were empirically reported for lake trout in GreatSlave Lake, NWT (Zimmerman et al. 2006) and forArctic charr (Salvelinus alpinus) in Gander Lake,Newfoundland (O’Connell and Dempson 2002,O’Connell et al. 2005). Color can be linked to re-productive behavior (Houde and Endler 1990,McLennan 1996, Foote et al. 2004); however,brightness did not differ between males and femaleswithin the Lake Mistassini forms. Skin colorationcan also be affected by diet (Kalinowski et al.2005), water chemistry (i.e., tannin staining), and/orgenetic make-up (Brooks and Endler 2001, Shikano2005)—three possibilities that, with further study,could provide explanations for the differences re-ported here.
The morphology and life history characteristicsof the deep-bodied form in Lake Mistassini weremore similar to humper trout from Lake Superiorthan to siscowet-like phenotypes from Great SlaveLake and Lake Superior. The small difference inbuoyancy between Lake Mistassini forms was simi-lar to that between leans and humpers but not be-tween leans and siscowets in Lake Superior(Thurston 1962, Eschmeyer and Phillips 1965) orbetween lean- and siscowet-like trout in GreatSlave Lake (Zimmerman et al. 2006). With their an-teriorly distributed body depth and narrow caudalpeduncle, the deep-bodied form in Lake Mistassiniresembled the profile of humper trout in Lake Supe-rior but did not resemble the deep head, mid-body,and caudal profiles of siscowet trout in Lake Supe-rior (Khan and Qadri 1970, Moore and Bronte2001) and of siscowet-like trout in Great SlaveLake (Zimmerman et al. 2006). Although the deep-bodied form in Lake Mistassini lacked the thin ab-dominal wall that characterizes humper trout(Lawrie and Rahrer 1973, Burnham-Curtis andBronte 1996, Moore and Bronte 2001), this featureis not consistent across humper populations (Peck1975). The “pot-belly” appearance of the Lake Mis-tassini deep-bodied form also was not characteristicof Lake Superior humpers (Moore and Bronte2001); in this respect, the deep-bodied form in LakeMistassini was distinctive in appearance. In termsof life history, the deep-bodied form appeared tomature at smaller sizes than did the streamlinedform. All deep-bodied individuals caught were ca-pable of reproducing at a length of 32 cm (SL),whereas immature streamlined individuals were aslong as 49 cm (SL). Small maturation size of thedeep-bodied form was similar to humper trout inLake Superior, which reach adulthood at shorter
lengths than lean and siscowet trout (Rahrer 1965,Burnham-Curtis and Bronte 1996).
Origins of Sympatric Shallow- and Deep-water Phenotypes
The availability of shallow- and deep-waterniches may be a key ecological variable contribut-ing to lake trout phenotypic differentiation in LakeMistassini. Indeed, deep-water habitat is a featureof all four lakes where lake trout phenotypes havebeen observed. In comparison with Lake Superior,Great Slave Lake, and Great Bear Lake, Lake Mis-tassini is young, shallow, and small in surface area(Beeton 1984, Dyke and Priest 1987, Evans 2000).As such, the presence of two sympatric phenotypesin Lake Mistassini is important for understandingthe environment necessary for phenotypic differen-tiation to occur. A genetic basis for these differentphenotypes awaits further study, but is hypothe-sized based on the genetic distinctiveness of leanand siscowet trout in Lake Superior (Burnham-Cur-tis and Smith 1994, Page et al. 2004) and on aknown genetic basis for two traits associated withdeep-water living—lipid content (Eschmeyer andPhillips 1965) and swim-bladder gas retention(Ihssen and Tait 1974). Ecological trade-offs con-tributing to phenotypic differentiation also awaitfurther study. The competitive trade-off proposedfor high-lipid, vertically migrating siscowet-liketrout (Zimmerman et al. 2006) did not necessarilyapply to the deep-water phenotype in Lake Mis-tassini, which more closely resembled humper troutwhose ecology is poorly understood.
The existence of a humper-like phenotype inLake Mistassini challenges the hypothesis that thehumper form was produced from an introgressionof lean and siscowet forms (Burnham-Curtis 1993,Burnham-Curtis and Smith 1994). If a siscowet-likeform never inhabited Lake Mistassini, the existingdeep-bodied form might either be the descendent ofa colonizing form (presumably the lean form), haveoriginated from dispersal after deglaciation of ahumper-like progenitor, or be one of two environ-mentally induced phenotypes in the lake (i.e., phe-notypic plasticity). If siscowet-like trout havehistorically inhabited or presently inhabit LakeMistassini, the form is either extinct or in low abun-dance. If siscowet-like trout were once present andparallel introgression events occurred in Lake Supe-rior and Lake Mistassini, the lakes differ remark-ably in the long-term abundances of humper versussiscowet phenotypes.
A humper-like phenotype is the dominant form in

Phenotypic Diversity of Lake Trout in Lake Mistassini 167
deep-water habitats in Lake Mistassini and theirabundance contrasts with Lake Superior and GreatSlave Lake where deep-water habitats are domi-nated by the siscowet phenotype. In Lake Superior,humper trout occur in isolated populations on off-shore humps similar to sea mounts. In Great SlaveLake, a comparable humper-like phenotype was notfound in two survey years (2001 and 2002) of deep-water habitat (Zimmerman et al. 2006) indicatingthat it is either absent or in low abundance and lo-cally distributed in this lake as it is in Lake Supe-rior. Based on available data, Lake Mistassini isunique in that it demonstrates that, under certainconditions, humper-like trout have the capacity tobe ecologically dominant in the deep waters oflarge lakes.
ACKNOWLEDGMENTS
Michael Prince and Andrew Coon (Council ofthe Cree Nation of Mistissini) assisted with permit-ting and study logistics. Kenny Longchamp andEric Coon-Come (Mistissini, Quebec) and MarkEbener (Chippewa/Ottawa Resource Authority) as-sisted with field work under demanding weatherconditions. Allison Niggemyer and Sean Sislerhelped with the diet analysis in laboratory spaceprovided by the Great Lakes Science Center(USGS, Ann Arbor, MI). Julia Cameron and Kath-leen Weesies (Michigan State University) con-structed the map of sampling localities. H. DavidSheets (Canisius College) produced software usedfor geometric morphometric analyses. Funding forthis research was provided by the Fishery ResearchProgram of the Great Lakes Fishery Commission.
REFERENCESAlexander, R.M. 1972. The energetics of vertical migra-
tion by fishes. In The effects of pressure on livingorganisms. Symposia of the Society for ExperimentalBiology., eds. M.A. Sleigh and A.G. MacDonald, pp.273–294. New York, New York: Academic Press.
———. 1993. Buoyancy. In The physiology of fishes, ed.D.H. Evans, pp. 75–97. Boca Raton, Florida: CRCPress.
Alfonso, N.R. 2004. Evidence for two morphotypes oflake charr, Salvelinus namaycush, from Great BearLake, Northwest Territories, Canada. Env. Biol. Fish.71:21–32.
Beeton, A.M. 1960. The vertical migration of Mysisrelicta in lakes Huron and Michigan. J. Fish. Res.Board Can. 17:517–539.
———. 1984. The world’s great lakes. J. Great LakesRes. 10:106–113.
——— , and Bowers, J.A. 1982. Vertical migration ofMysis relicta Loven. Hydrobiologia 93:53–61.
Blackie, C.T., Weese, D.J., and Noakes, D.L.G. 2003.Evidence for resource polymorphism in the lake charr(Salvelinus namaycush) population of Great BearLake, Northwest Territories, Canada. Ecoscience10:509–514.
Bookstein, F.L. 1991. Morphometric tools for landmarkdata. New York, New York: Cambridge UniversityPress.
Bowers, J.A. 1988. Diel vertical migration of the Opos-sum Shrimp Mysis relicta in Lake Superior—observa-tions and sampling from the Johnson-Sea-Link II sub-mersible. Bull. Mar. Sci. 43:730–738.
Bronte, C.R., Ebener, M.P., Schreiner, D.R., DeVault,D.S., Petzold, M.M., Jensen, D.A., Richards, C., andLozano, S.J. 2003. Fish community change in LakeSuperior, 1970–2000. Can. J. Fish. Aquat. Sci.60:1552–1574.
Brooks, R., and Endler, J.A. 2001. Direct and indirectsexual selection and quantitative genetics of maletraits in guppies (Poecilia reticulata). Evolution55:1002–1015.
Brown, E.H., Eck, G.W., Foster, N.R., Horrall, R.M.,and Coberly, C.E. 1981. Historical evidence for dis-crete stocks of lake trout (Salvelinus namaycush) inLake Michigan. Can. J. Fish. Aquat. Sci.38:1747–1758.
Burnham-Curtis, M.K. 1993. Intralacustrine speciationof Salvelinus namaycush in Lake Superior: an investi-gation of genetic and morphological variation andevolution of lake trout in the Laurentian Great Lakes.Ph. D. dissertation. University of Michigan, AnnArbor, Michigan.
———, and Bronte, C.R. 1996. Otoliths reveal a diverseage structure for humper lake trout in Lake Superior.Trans. Am. Fish. Soc. 125:844–851.
———, and Smith, G.R. 1994. Osteological evidence ofgenetic divergence of lake trout (Salvelinus namay-cush) in Lake Superior. Copeia 1994:843–850.
Dadswell, M.J. 1974. Distribution, ecology, and post-glacial dispersal of certain crustaceans and fishes ineastern North America. National Museum of Canada,Ottawa, Canada. Publications in Zoology, No. 11.
Dyke, A.S., and Priest, V.K. 1987. Late Wisconsinanand Holocene history of the Laurentide ice sheet.Geogr. Phys. Quat. 41:237–263.
Eschmeyer, P.H., and Phillips, A.M. 1965. Fat content ofthe flesh of siscowets and lake trout from Lake Supe-rior. Trans. Am. Fish. Soc. 94:62–74.
Eshenroder, R.L., Payne, N.R., Johnson, J.E., Bowen, C.,and Ebener, M. P. 1995. Lake trout rehabilitation inLake Huron. J. Great Lakes Res. 21 (Suppl. 1):108–127.
———, Argyle, R.L., and TeWinkel, L.M. 1998. Evi-dence for buoyancy regulation as a speciation mecha-

168 Zimmerman et al.
nism in Great Lakes ciscoes. Arch. Hydrobiol. Sp.Issues Adv. Limnol. 50:207–217.
———, Sideleva, V.G., and Todd, T.N. 1999. Functionalconvergence among pelagic sculpins of Lake Baikaland deepwater ciscoes of the Great Lakes. J. GreatLakes Res. 25:847–855.
Evans, M.S. 2000. The large lake ecosystems of northernCanada. Aquat. Ecosystem Health Management3:65–79.
Foote, C.J., Brown, G.S., and Hawryshyn, C.W. 2004.Female colour and male choice in sockeye salmon:implications for the phenotypic convergence ofanadromous and nonanadromous morphs. Anim.Behav. 67:69–83.
Fraser, D.J., and Bernatchez, L. 2005. Adaptive migra-tory divergence among sympatric brook charr popula-tions. Evolution 59:611–624.
Gallagher, C.P. 2002. Traditional knowledge and geneticdiscimination of Lake Nipigon lake trout stocks. M.Sc. thesis. University of Manitoba, Manotiba.
Goode, G.B. 1884. Section I. Natural history of usefulaquatic animals. U.S. Government Printing Office,Washington, D.C.
Goodier, J.L. 1981. Native lake trout (Salvelinus namay-cush) stocks in the Canadian waters of Lake Superiorprior to 1955. Can. J. Fish. Aquat. Sci. 38:1724–1737.
Hallock, C. 1877. The game animals, birds and fishes ofNorth America: their habits and various methods ofcapture. New York, New York: “Forest and Stream”Publishing Company.
Henderson, B.A., and Anderson, D.M. 2002. Phenotypicdifferences in buoyancy and energetics of lean andsiscowet lake charr in Lake Superior. Environ. Biol.Fishes 64:203–209.
Houde, A.E., and Endler, J.A. 1990. Correlated evolutionof female mating preferences and male color patternsin the guppy Poecilia reticulata . Science248:1405–1408.
Hrabik, T.R., Jensen, O.P., Martell, S.J.D., Walters, C.J.,and Kitchell, J.F. 2006. Diel vertical migration in theLake Superior pelagic community. I. Changes in ver-tical migration of coregonids in response to varyingpredation risk. Can. J. Fish. Aquat. Sci.63:2286–2295.
Ihssen, P., and Tait, J.S. 1974. Genetic differences inretention of swimbladder gas between two popula-tions of lake trout (Salvelinus namaycush). J. Fish.Res. Board Can. 31:1351–1354.
Jennings, M.R. 1989. Use of spring scales for weighinglive fish in the field. N. Am. J. Fish. Man. 9:509–511.
Jensen, O.P., Hrabik, T.R., Martell, S.J.D., Walters, C.J.,and Kitchell, J.F. 2006. Diel vertical migration in theLake Superior pelagic community. II. Modeling trade-offs at an intermediate trophic level. Can. J. Fish.Aquat. Sci. 63:2296–2307.
Kalinowski, C.T., Robaina, L.E., Fernandez-Palacios,H., Schuchardt, D., and Izquierdo, M.S. 2005. Effect
of different carotenoid sources and their dietary levelson red porgy (Pagrus pagrus) growth and skin colour.Aquaculture 244:223–231.
Khan, N.Y., and Qadri, S.U. 1970. Morphological differ-ences in Lake Superior lake char. J. Fish. Res. BoardCan. 27:161–167.
Koelz, W. 1926. Fishing industry of the Great Lakes:Appendix XI to the report of the U. S. Commissionerof fisheries for 1925. Bureau of Fisheries, U. S.Department of Commerce, Washington, D. C. Bureauof Fisheries Document No. 1001.
Konkle, B.R., and Sprules, W.G. 1986. Planktivory bystunted lake trout in an Ontario lake. Trans. Am. Fish.Soc. 115:515–521.
Krueger, C.C., and Ihssen, P.E. 1995. Review of geneticsof lake trout in the Great Lakes: History, moleculargenetics, physiology, strain comparisons, and restora-tion management. J. Great Lakes Res. 21:348–363.
Langerhans, R.B., Layman, C.A., Langerhans, A.K., andDewitt, T.J. 2003. Habitat-associated morphologicaldivergence in two Neotropical fish species. Biol. J.Linnean Soc. 80:689–698.
Lawrie, A.H., and Rahrer, J.H. 1973. Lake Superior: acase history of the lake and its fisheries. Great LakesFishery Commission, Ann Arbor, Michigan. Techni-cal Report No. 19.
Martin, N.V. 1966. The significance of food habits in thebiology, exploitation, and management of AlgonquinPark, Ontario, lake trout. Trans. Am. Fish. Soc.95:415–422.
———, and Olver, C.H. 1980. The lake charr, Salvelinusnamaycush. In Charrs: salmonid fishes of the genusSalvelinus, ed. E.K. Balon, pp. 205–277. The Hague,The Netherlands: Dr. W. Junk Publishers.
Mathworks. 2000. Matlab v. 6.2. Natick, Massachusetts.McLennan, D.A. 1996. Integrating phylogenetic and
experimental analyses: The evolution of male andfemale nuptial coloration in the stickleback fishes(Gasterosteidae). Syst. Biol. 45:261–277.
Moore, S.A., and Bronte, C.R. 2001. Delineation of sym-patric morphotypes of lake trout in Lake Superior.Trans. Am. Fish. Soc. 130:1233–1240.
Nolte, A.W., and Sheets, H.D. 2005. Shape based assign-ments tests suggest transgressive phenotypes in nat-ural sculpin hybrids (Teleostei, Scorpaeniformes, Cot-tidae). Frontiers in Zoology 2:11.
O’Connell, M.F., and Dempson, J.B. 2002. The biologyof Arctic charr, Salvelinus alpinus, of Gander Lake, alarge, deep, oligotrophic lake in Newfoundland,Canada. Environ. Biol. Fishes 64:115–126.
———, Dempson, J.B., and Power, M. 2005. Ecologyand trophic relationships of the fishes of Gander Lake,a large, deep, oligotrophic lake in Newfoundland,Canada. Int. Rev. Hydrobiol. 90:486–510.
Page, K.S., Scribner, K.T., and Burnham-Curtis, M.2004. Genetic diversity of wild and hatchery lake

Phenotypic Diversity of Lake Trout in Lake Mistassini 169
trout populations: Relevance for management andrestoration in the Great Lakes. Trans. Am. Fish. Soc.133:674–691.
Peck, J.W. 1975. Brief life history accounts of five com-mercial salmonid fishes in Lake Superior. MichiganDepartment of Natural Resources Fisheries Division,Lansing, Michigan. Research Report No. 1821.
Power, M., O’Connell, M.F., and Dempson, J.B. 2005.Ecological segregation within and among Arctic charmorphotypes in Gander Lake, Newfoundland. Envi-ron. Biol. Fishes 73:263–274.
Rahrer, J.F. 1965. Age, growth, maturity, and fecundityof “humper” lake trout, Isle Royale, Lake Superior.Trans. Am. Fish. Soc. 94:75–83.
Reist, J.D. 1985. An empirical evaluation of several uni-variate methods that adjust for size variation in mor-phometric data. Can. J. Zool. 63:1429–1439.
Robinson, B.W., and Wilson, D.S. 1994. Characterrelease and displacement in fishes: a neglected litera-ture. Am. Nat. 144:596–627.
Rohlf, F.J., and Corti, M. 2000. Use of two-block partialleast-squares to study covariation in shape. Syst. Biol.49:740–753.
Ruber, L., and Adams, D.C. 2001. Evolutionary conver-gence of body shape and trophic morphology in cich-lids from Lake Tanganyika. J. Evol. Biol. 14:325–332.
Sampson, P.D., Bookstein, F.L., Sheehan, F.H., and Bol-son, E.L. 1996. Eigenshape analysis of left ventricularoutlines from constrast ventriculograms. In Advancesin morphometrics, L.F. Marcus, M. Corti, and A. Loy,eds., pp. 211–233. New York, New York: Plenum.
Shikano, T. 2005. Marker-based estimation of heritabil-ity for body color variation in Japanese flounder Par-alichthys olivaceus. Aquaculture 249:95–105.
Skulason, S., and Smith, T.B. 1995. Resource polymor-phisms in vertebrates. Trends Ecol. Evol. 10:366–370.
Smith, T.B., and Skulason, S. 1996. Evolutionary signifi-cance of resource polymorphisms in fishes, amphib-ians, and birds. Annu. Rev. Ecol. Syst. 27:111–133.
SPSS. 2003. v. 12.0 for Windows. SPSS Inc, Chicago,Illinois.
TeWinkel, L.M., and Fleischer, G.W. 1999. Verticalmigration and nighttime distribution of adult bloaters inLake Michigan. Trans. Am. Fish. Soc. 128:459–474.
Thurston, C.E. 1962. Physical characteristics and chemi-cal composition of two subspecies of lake trout. J.Fish. Res. Board Can. 19:39–44.
Turgeon, J., Estoup, A., and Bernatchez, L. 1999. Speciesflock in the North American Great Lakes: Molecularecology of Lake Nipigon Ciscoes (Teleostei: Corego-nidae: Coregonus). Evolution 53:1857–1871.
van der Salm, A.L., Spanings, F.A.T., Gresnigt, R.,Bonga, S.E.W., and Flik, G. 2005. Background adap-tation and water acidification affect pigmentation andstress physiology of tilapia, Oreochromis mossambi-cus. Gen. Comp. Endocrinol. 144:51–59.
Zelditch, M.L., Swiderski, D.L., Sheets, H.D., and Fink,W.L. 2004. Geometric morphometrics for biologists:a primer. New York, New York: Elsevier AcademicPress.
Zimmerman, M.S., Krueger, C.C., and Eshenroder, R.L.2006. Phenotypic diversity of lake trout in Great SlaveLake: differences in morphology, buoyancy, and habi-tat depth. Trans. Am. Fish. Soc. 135:1056–1067.
Submitted: 15 May 2006Accepted: 6 December 2006Editorial handling: Carol A. Stepien




















