
Biological Cycle of Podocoryna Exigua (Cnidaria: Hydrozoa) from a Sandy Bottom of the Ligurian Sea

Pesticide Biochemistry and Physiology 101 (2011) 198–205
Contents lists available at SciVerse ScienceDirect
Pesticide Biochemistry and Physiology
journal homepage: www.elsevier .com/locate /pest
Monitoring of beet armyworm Spodoptera exigua (Lepidoptera: Noctuidae)resistance to chlorantraniliprole in China
Tiancai Lai a, Jia Li a, Jianya Su a,b,⇑a College of Plant Protection, Nanjing Agricultural University, Jiangsu 210095, Chinab Jiangsu Key Laboratory of Pesticide Science, Nanjing Agricultural University, Nanjing 210095, China
a r t i c l e i n f o a b s t r a c t
Article history:Received 22 May 2011Accepted 8 September 2011Available online 22 September 2011
Keywords:Spodoptera exiguaChlorantraniliproleInsecticide resistanceMixed-function oxidaseEsteraseGlutathione-S-transferaseSynergism assay
0048-3575/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.pestbp.2011.09.006
⇑ Corresponding author at: College of Plant ProtUniversity, Jiangsu 210095, China.
E-mail address: [email protected] (J. Su).
Beet armyworm, Spodoptera exigua is a major insect pest of vegetables in China, and has been reported todevelop resistance to many broad-spectrum insecticides. Recently registered chlorantraniliprole providesa novel option for control of this pest resistant to other conventional insecticides. The susceptibilities offield collected populations were measured by diet incorporation assay with neonate, obvious variation ofsusceptibility was observed among the 18 field populations with LC50 values varying from 0.039 to0.240 mg/liter. Moderate resistant level was discovered in 8 of 18 field populations, other 8 populationshad become low level tolerance to chlorantraniliprole, and only one population in all the field coloniesremained susceptible. Biochemical assays were performed to determine the potential mechanismsinvolved in tolerance variation. Field populations displayed varied detoxification enzyme activities, butthe regression analysis between chlorantraniliprole toxicities and enzyme activities demonstrated eachfield population might have specific biochemical mechanisms for tolerance. Artificial selection in labora-tory with chlorantraniliprole was carried out, 23 generations of continuous selections resulted in11.8-fold increase in resistance to chlorantraniliprole, and 3.0-fold and 3.7-fold increases in mixed func-tion oxidase and esterase, respectively. Compared with the susceptible strain kept in laboratory the selec-tion strain had developed 128.6-fold resistance to this insecticide. Synergism assays showed thedetoxification enzymes might not involved in the resistance observed in field collected populationsand the selected strain.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
The beet armyworm, Spodoptera exigua (Hübner) (Lepidoptera:Noctuidae), is a widely distributed polyphagous pest [1]. In Chinait seriously damages many cultivated crops including cabbage, cow-pea, soybean, onion, cotton and others [2,3], insecticide applicationis the most practical way to prevent its damages, however, manyinsecticides have been reported to fail to control S. exigua due tothe insecticide resistance, and the resistance cases to various typesof traditional insecticides have been documented in China [4–7] andother countries [8–13], some populations have developed resis-tance to newer insecticides such as spinosad [14–16], tebufenozide[17] and Bacillus thuringiensis cry toxins [18] in the fields. Therefore,environmental friendly alternatives for S. exigua control are re-quired for use in the development of integrated pest management(IPM) programs.
Chlorantraniliprole, the first commercialized ryanodine receptorinsecticide from anthranilic diamides which belongs to insecticide
ll rights reserved.
ection, Nanjing Agricultural
resistance action committee (IRAC) mode of action class 28 [19],has exceptional insecticidal activity on a range of Lepidopteranpests and other orders, such as Coleoptera, Diptera, Isoptera andHemiptera [20,21]. Chlorantraniliprole activates the unregulatedrelease of internal calcium stores leading to Ca2+ depletion, feedingcessation, lethargy, muscle paralysis and finally insect death [22].Additionally, because of hasn’t been found to exhibit cross-resistance with other commercial insecticides [23–25] and low eco-toxicology [26,27], chlorantraniliprole is an excellent alternative tobroad-spectrum insecticides in Integrated Pest Management (IPM)programs where commercial standards are no longer effective be-cause of resistance.
As a newly developed insecticide, it is critical to establish thesusceptibility levels of various populations at the outset even be-fore the widespread use of chlorantraniliprole. Additionally, con-tinuous monitoring of S. exigua populations to document shifts insensitivity after the commercial introduction of chlorantraniliprolewill aid in implementing resistance management strategies beforecontrol failures occur. The objectives of this study were to investi-gate the susceptibility of S. exigua field populations to chlorantra-niliprole in China, and to evaluate the detoxification activities ofmixed function oxidases (MFO), glutathione-S-transferases (GST)

T. Lai et al. / Pesticide Biochemistry and Physiology 101 (2011) 198–205 199
and esterases (EST), as well as to determine the synergism effectsof synergists to help explain possible mechanisms involved in theresistance to chlorantraniliprole.
2. Materials and methods
2.1. Insects
The susceptible strain (Lab-Sus) of S. exigua, was used as the ref-erence population which was provided by Wuhan Kernel Bio-pesticide Company, Hubei, China, in May 2001. Lab-Sus has beenmaintained in the laboratory without exposure to any insecticide.Eighteen field populations of S. exigua were collected from 13 dif-ferent geographical regions of China. About 200 3rd to 5th-instarlarvae were collected from each sampling site (Fig. 1, Table 1).
They were mass mated and their F1 larvae were used for bioas-says. Adults were fed 10% sugar solution. All stages were kept inthe same standard conditions of 27 ± 1 �C, 60–70% RH and14:10 h light: dark photoperiod. Newly laid eggs were sterilizedwith 5% formaldehyde to prevent viral pathogens, and larvae werereared with artificial diet [5].
2.2. Chemicals
Chlorantraniliprole (Rynaxypyr™ 200 g/liter SC, Coragen; Du-Pont Agricultural Chemicals Ltd., Shanghai, China) was commer-cially available. NADPH, p-nitrophenol (PNP), Glutathione (GSH)and a-naphthol were purchased from Bio Basic Inc., USA. Piperonylbutoxide (PBO) was purchased from Koch-Light Laboratories Co.,Ltd, UK. Diethylmaleate (DEM), 4-nitroanisole (PNA), triphenylphosphate (TPP), Coomassie Brilliant Blue G-250 and a-naphthylacetate (a-NA) were obtained from Shanghai Chemical Factory,China. 2,4-Dinitrochlorobenzene (CDNB) and EDTA were pur-chased from Sangon Biotech (Shanghai) Co., Ltd. DTT was fromBeijing Solarbio Science and Technology Co., Ltd, China. Bovine ser-
Fig. 1. Sampling sites of S. exigu
um albumin (BSA), PMSF and S,S,S-tributylphosphorothioate (DEF),were obtained from Sino-American Biotechnology Co., Ltd, China.
2.3. Bioassays
Diet incorporation bioassay: the chlorantraniliprole susceptibil-ity of beet armyworm from field populations and selected strainwere assayed with neonate larvae (newly hatched larvae <10 hold) of S. exigua by diet incorporation method. Seven concentra-tions of chlorantraniliprole using twofold dilutions were preparedwith distilled water. After preparing the diet, a quantity of 5 mL ofdiluted chlorantraniliprole was mixed thoroughly with 45 mL ofartificial diet in glass cup (below 54 �C), then approximately2.5 mL of the diet was dispensed into cylindrical identical tissueculture tubes (2 � 8 cm). Five neonate larvae were placed in eachof culture tube, and 20 tubes were prepared for each concentrationwhich a total of 100 larvae were used for each concentration. Thenall the tubes were closed with cotton pads and placed in an incu-bator (27 ± 1 �C, 60–70% RH, and 14:10 h light: dark photoperiod).Larval mortality was evaluated at 3 d. Larvae were recorded asdead if they did not respond with head movements or peristalticcontractions when touched with camel’s-hair brush. To ensure thatoffspring of many females were assayed, larvae hatching from anygiven egg mass were systematically distributed among variousconcentrations (i.e., a maximum of about 5 larvae per egg masswere exposed to every concentration).
Leaf dip bioassay: the third-instar larvae (5–7 mg body weight)were exposed to chlorantraniliprole using the leaf-dip techniquerecommended by the Insecticide Resistance Action Committee(IRAC). Serial dilutions of chlorantraniliprole were prepared usingdistilled water containing 0.1% Triton X-100, and 5 cm cabbage,Brassica aleracea L., leaf discs were cut and dipped into the testsolutions for 15 s with gentle agitation. Control discs were treatedwith 0.1% Triton X-100 solution only. They were dried at roomtemperature for 2–3 h. Three larvae were released onto each leaf
a field population in China.

Table 1Locations, sampling dates, host plants and developmental stages of S. exigua collected from fileds.
Strains Location Collection date Map ref. # Sites Host plants Developmental stage
SZ09 Shengzhen, Guangdong Nov. 2009 1 22.69�N, 114.13�E Allium fistulosum L. var. giganteum Makino 3rd–5th LarvaeSZ10 Shengzhen, Guangdong Oct. 2010 1 22.69�N, 114.13�E Allium fistulosum L. var. giganteum Makino 3rd–5th LarvaeDG10 Dongguang, Guangdong Oct. 2010 2 22.84�N, 113.75�E Allium fistulosum L. var. giganteum Makino 3rd–5th LarvaeNN10 Nanning, Guangxi Jun. 2010 3 22.85�N, 108.25�E Vigna unguiculata (Linn) Walp. 3rd–5th LarvaeHZ10 Huizhou, Guangdong Oct. 2010 4 23.03�N, 114.36�E Allium fistulosum L. var. giganteum Makino 3rd–5th LarvaeZZ10 Zhangzhou, Fujian Oct. 2010 5 24.51�N, 117.67�E Brassica chinensis L. 3rd–5th LarvaeCS10 Changsha, Hunan Sep. 2010 6 28.18�N, 113.09�E Brassica oleracea L. var. capitata 3rd–5th LarvaeWH09 Wuhan, Hubei Aug. 2009 7 30.52�N, 114.32�E Brassica oleracea L. var. capitata 3rd–5th LarvaeSH08 Minhang, Shanghai Sep. 2008 8 31.10�N, 121.38�E Brassica chinensis L. 3rd–5th LarvaeSH10 Minhang, Shanghai Aug. 2010 8 31.10�N, 121.38�E Brassica chinensis L. 3rd–5th LarvaeHX09 Hexian, Anhui Oct. 2009 9 31.72�N, 118.41�E Brassica chinensis L. 3rd–5th LarvaeHF10 Hefei, Anhui Aug. 2010 10 31.85�N, 117.23�E Brassica chinensis L. 3rd–5th LarvaeNJ08 Nanjing, Jiangsu Sep. 2008 11 32.13�N, 118.97�E Brassica chinensis L. 3rd–5th LarvaeNJ09A Nanjing, Jiangsu Jul. 2009 11 32.05�N, 118.94�E Brassica chinensis L. 3rd–5th LarvaeNJ09B Nanjing, Jiangsu Sep. 2009 11 32.05�N, 118.94�E Brassica chinensis L. 3rd–5th LarvaeNJ10 Nanjing, Jiangsu Sep. 2010 11 32.06�N, 118.97�E Brassica chinensis L. 3rd–5th LarvaeXX10 Xinxiang, Henan Aug. 2010 12 35.31�N, 113.93�E Brassica chinensis L. 3rd–5th LarvaeTA08 Tai’an, Shandong Sep. 2008 13 36.23�N, 117.09�E Brassica chinensis L. 3rd–5th Larvae
200 T. Lai et al. / Pesticide Biochemistry and Physiology 101 (2011) 198–205
disc and then kept at 27 ± 1 �C, 60–70% RH with a 14:10 h light:dark photoperiod. For each concentration, at least 30 3rd-instarlarvae were exposed to each concentration. The mortalities wererecorded after 96 h of exposure. Larvae were considered dead if un-able to move in a coordinated way when prodded with a fine-haired brush. The same numbers of leaf discs per treatment weredipped into distilled water as an untreated control.
2.4. Selection for resistance
SH08 strain collected from Shanghai, China in 2008 was takenas starting colony for selection with chlorantraniliprole. The con-centration of chlorantraniliprole used to select each subsequentgeneration was the LC70 based on the results of bioassays fromthe previous generation. The number of neonate larvae used foreach generation varied (5–8 larvae per tube, total 5000–10000 lar-vae) depending on availability. After 3 d of exposure, surviving lar-vae were transferred to untreated artificial diet and reared in thelaboratory under conditions described above.
2.5. Synergism assays
For analysis of the effects of synergists on toxicity of chlorantra-niliprole, toxicity with a concentration range of synergists was firstdetermined to choose a suitable concentration that would have noeffect on larval mortality. Concentrations up to 100 mg/liter ofDEM, TPP, DEF and PBO had no effect on larval mortality. Cabbageleaves treated with that concentration of each synergist were ap-plied to third instars for 12 h, and then larvae were assayed for tox-icity against chlorantraniliprole as described above. The synergismratio (SR) was determined by dividing the LC50 of insecticide aloneby the LC50 of insecticide with synergist.
2.6. Enzyme preparation
The 3rd-instar larvae (5–7 mg) were separated and starved for3 h to remove digested food particles. The whole larvae werehomogenized on ice in homogenization buffer (0.1 M phosphatebuffer, pH 7.6, containing 1 mM EDTA, 1 mM DTT, 1 mM PTU,1 mM PMSF and 20% glycerol). Insects were chilled on ice beforehomogenization. Sixteen insects were homogenized in 2 mL buffer,the homogenate was centrifuged at 4 �C, 10, 000 g for 15 min, thesolid debris and cellular material were discarded. The supernatantwas decanted into a clean eppendorf tube, placed on ice and usedimmediately for mixed function oxidase (MFO),
glutathione-S-transferase (GST), esterase (EST) assays and total pro-tein content assays.
2.7. Enzyme assays
The assay of mixed function oxidase (MFO) was conductedusing the procedures developed by Rose et al. [28]. 100 lL of2 mM p-nitroanisole solution and 90 lL enzyme were added toeach well of transparent 96-well Costar microplate (Corning LifeSciences, Lowell, MA). The microplate was incubated for 3 min at27 �C and the reaction initiated by the addition of 10 lL of9.6 mM NADPH. The absorbance was read in a TECAN microplatereader (Infinite™ M200) at 405 nm and 27 �C for 15 min. The activ-ity of MFO was calculated from p-nitrophenol standard curve andpresented as nanomoles of p-nitrophenol per minute per milligramof protein.
Glutathione-S-transferase (GST) activity was measured using1-chloro-2, 4-dinitrobenzene (CDNB) as substrate on transparent96-well Costar microplates according to the methods of Yang etal. [29]. 10 lL of the enzyme solution (diluted 10-fold in 0.1 MpH 7.6 sodium phosphate buffer) was mixed with 100 lL 1.2 mMCDNB and 100 lL 6 mM glutathione (GSH) in each well. Enzymeactivity was calculated in a TECAN microplate reader at 340 nmand 27 �C using the kinetic mode for 10 min. The activity of GSTwas determined using the extinction coefficient of 9.6 mM�1 cm�1
for CDNB. The results were expressed as nanomoles glutathioneconjugate per minute per milligram of protein.
Total esterase (EST) activity was determined using a-naphthylacetate (a-NA) as a substrate on transparent 96-well Costar micro-plates according to the methods of Yang et al. [29]. 10 lL of en-zyme solution (diluted 10-fold in 0.1 M pH 7.6 sodium phosphatebuffer) was mixed with 200 lL of substrate solution (5 mL 0.2 MpH 6.0 phosphate buffer, 10 mg Coomassie Brilliant Blue G-250and 0.1 mL 100 mM a-NA) in each well. Enzyme activity was mea-sured at 450 nm and 27 �C using a TECAN microplate reader in thekinetic mode for 10 min. The activity of the esterase was extrapo-lated from a-naphthol standard curve and expressed as nanomolesof a-naphthol per minute per milligram of protein.
Total protein content was determined by the Bradford [30]method using bovine albumin as a standard.
2.8. Statistical analysis
Lethal concentration values (LC50) and their 95% fiducial limits(FL) were estimated using POLO-Plus program [31]. Significantdifferences of LC50 were determined by non-overlapping 95%

T. Lai et al. / Pesticide Biochemistry and Physiology 101 (2011) 198–205 201
fiducial limits (FL). Resistance factors (RF) were estimated at the LC50
level as RF = LC50 of collected strain/LC50 of Lab-Sus strain. Insecti-cide resistance level was described by using RFs as reported by Keid-ing [32] and WHO [33]: Susceptibility (RF = 1), Decreasedsusceptibility (RF = 3–5), Low resistance (RF = 5–10), Moderateresistance (RF = 10–40), High resistance (RF = 40–160), and Veryhigh resistance (RF > 160). The activity of MFO, GST and EST was sub-jected to an ANOVA analysis followed by t-test (a < 0.05). Signifi-cance was accept at a = 0.05 in all statistical tests used in this study.
3. Results
3.1. Susceptibility of S. exigua to chlorantraniliprole
Chlorantraniliprole susceptibility of 18 S. exigua field popula-tions collected from China during 2008–2010 and a susceptiblestrain (Lab-Sus) were assayed (Table 2). The Lab-Sus strain hadthe highest susceptibility to this compound (LC50 = 0.014 mg/L)among all the tested populations. The field populations collectedfrom different locations had varied susceptibility to this insecticide(LC50 = 0.039–0.240 mg/L), there was 6.2-fold difference in LC50 val-ues between DG10 and WH09 strain. DG10 strain exhibited 17.1-fold resistance to this insecticide which had reached moderateresistance level compared with the Lab-Sus strain. Other field pop-ulations with moderate resistance included DG10, HZ10, SZ10,ZZ10, SH08, SH10, HF10 and TA08, 8 populations showed low resis-tance level, decreased susceptibility was observed in NN10 popula-tion and only WH09 remained susceptible to this compound.
Although obvious variation of susceptibility among differentgeographic populations (6.2-fold) were observed, no differenceswere found in populations collected from the same location in dif-ferent years (2008–2010). Four strains from Nanjing during2008–1010 displayed similar tolerant level to chlorantraniliprole,LC50s were in the range of 0.090–0.117 mg/L. No difference was de-tected from Shanghai strains collected in 2008 (SH08) and 2010(SH10). Similar tolerant levels were observed in strains from Shen-gzheng, Dongwan, Huizhou and Zhangzhou, which are very closeon geographical distance, except the SZ09 strain collected in 2009.
3.2. Resistance selection
SH08 strain was taken as the starting population for chlorantra-niliprole selection. Selection at LC70 produced significant increase
Table 2Susceptibility to chlorantraniliprole of S. exigua collected from the fields in China.
Strains na LC50:mg/L (95% FL) Slope ± SE v2 (df) RFb
Lab-Sus 675 0.014 (0.010–0.017) 1.770 ± 0.239 2.695(4) 1.0NN10 600 0.068 (0.057–0.078) 3.206 ± 0.341 0.919(3) 4.8SH08 600 0.152 (0.128–0.177) 2.540 ± 0.269 2.960(3) 10.9SH10 450 0.141 (0.115–0.170) 2.528 ± 0.400 1.228(3) 10.1TA08 500 0.140 (0.117–0.161) 3.524 ± 0.578 1.946(3) 10.0NJ08 600 0.112 (0.090–0.135) 2.012 ± 0.207 1.802(3) 8.0NJ09A 700 0.117 (0.090–0.145) 1.791 ± 0.200 3.257(4) 8.4NJ09B 700 0.090 (0.069–0.113) 1.578 ± 0.160 0.688(4) 6.4NJ10 540 0.101 (0.083–0.121) 2.537 ± 0.335 2.331(3) 7.2HF10 375 0.163 (0.140–0.185) 4.433 ± 0.577 0.135(3) 11.7HX09 600 0.104 (0.088–0.123) 2.089 ± 0.215 0.329(3) 7.4SZ09 500 0.137 (0.116–0.158) 2.746 ± 0.280 1.370(3) 9.8SZ10 700 0.211 (0.176–0.247) 2.407 ± 0.227 3.489(4) 15.1DG10 600 0.240 (0.203–0.278) 2.331 ± 0.206 1.078(3) 17.1HZ10 600 0.212 (0.176–0.255) 2.201 ± 0.221 0.927(3) 15.1ZZ10 600 0.162 (0.137–0.189) 2.410 ± 0.221 0.728(3) 11.6XX10 375 0.124 (0.105–0.143) 3.337 ± 0.378 0.821(2) 8.9CS10 500 0.119 (0.100–0.139) 2.746 ± 0.351 0.232(2) 8.5WH09 600 0.039 (0.030–0.048) 1.643 ± 0.174 2.922(3) 2.8
a Number of neonate larvae used.b RF = LC50 of a population/LC50 of the Lab-Sus strain.
in tolerance to this insecticide. After 23 generations of selection,the LC50 value had increased to 1.801 mg/L (Table 3). These dataindicated the selection had resulted in 11.8-fold increase of resis-tance towards chlorantraniliprole compared with 0.152 mg/L forthe initial population. Compared with the reference strain Lab-Sus the selection strain had developed 128.6-fold resistance to thischemical.
The 128.6-fold resistance of SH-Ryn strain was calculated fromthe results of neonate larvae by diet incorporation assay, the resis-tance levels were verified with 3rd-larvae by leaf-dip bioassay onSH-Ryn strain and two field collected populations with higherresistance to chlorantraniliprole. Two different assay methods pro-duced similar results on SH-Ryn, DG10 and SZ10 strains (Table 4).
3.3. Enzyme assays
The detoxification enzyme activities were measured in 15 fieldcollected populations, one laboratory susceptible strain (Lab-Sus)and the 10th and 23rd generation of selection strain (SH-Ryn).There were narrow variations of detoxification enzyme activitiesamong field populations. Maximum 3.35-fold difference in MFOactivities was observed from field populations (Fig. 2A), the highestMFO activity was found in HF10 strain (0.332 nmol/min/mg pro-tein), the lowest activity in SZ09 (0.099 nmol/min/mg protein)which was lower than that of Lab-Sus strain (0.162 nmol/min/mgprotein). The highest MFO activity was not observed in the mosttolerant populations, likewise, the lowest MFO activity didn’t ap-pear in the most susceptible populations. The correlation betweenMFO activity and toxicity of chlorantraniliprole on beet armywormdidn’t fit to the linear regression models (P = 0.478, R2 = 0.039) (Fig2D).
Maximum 4.8-fold difference in esterase activities was ob-served among the field populations, the highest esterase activitywas from ZZ10 strain (241.19 nmol/min/mg protein) (Fig. 2B),the lowest one from SZ09 strain (49.86 nmol/min/mg protein)which was lower than that of Lab-Sus strain(64.96 nmol/min/mgprotein). There was a maximum 3.5-fold difference in glutathi-one-S-transferase activity among field populations. The highestand the lowest activities were from SZ10 (1979.70 nmol/min/mgprotein) and HX10 (572.72 nmol/min/mg protein) respectively.The GST activity of HX10 was also lower than that of Lab-Sus(Fig. 2C). The correlation between EST, GST activities and toxicitiesof chlorantraniliprole on S. exigua didn’t reach a good fit to the lin-ear regression model (P = 0.021 and 0.032, R2 = 0.327 and 0.288)(Fig. 2E and Fig. 2F). These data demonstrate that the field popula-tions collected from different geographic areas perhaps harbor dis-crepant biochemical mechanism of tolerance to chlorantraniliproledue to different histories of insecticide use.
In selection experiment SH08 was taken as initial colony. After10 generations of selection, the activity of MFO and esterase in-creased obviously. 2.9-Fold and 3.0-fold increases were measuredin 10th generation of SH-Ryn for MFO and esterase activities,respectively. Continuous selection for 13 generations from G10 toG23 lead to obvious increase in esterase activity (3.7-fold), howeveronly slight increase in MFO activity. No obvious changes in GSTactivities were observed in SH-Ryn strain during the course ofselection (Table 5).
The changes of detoxification enzyme activities during theselection course may reflect the shift of resistance mechanism ofbeet armyworm to chlorantraniliprole.
3.4. Synergism tests
In order to analyze the biochemical mechanisms of resistance inbeet armyworm to chlorantraniliprole the mixed function oxidaseinhibitor PBO, esterase inhibitor DEF, glutathione-S-transferase
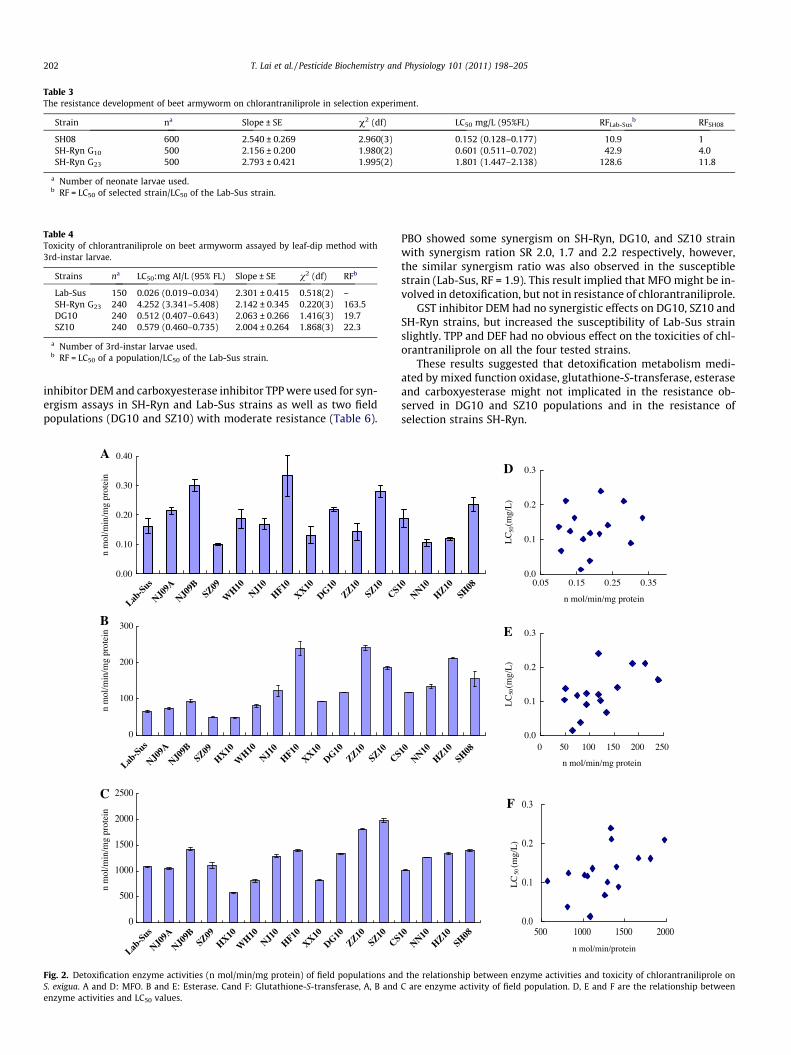
Table 3The resistance development of beet armyworm on chlorantraniliprole in selection experiment.
Strain na Slope ± SE v2 (df) LC50 mg/L (95%FL) RFLab-Susb RFSH08
SH08 600 2.540 ± 0.269 2.960(3) 0.152 (0.128–0.177) 10.9 1SH-Ryn G10 500 2.156 ± 0.200 1.980(2) 0.601 (0.511–0.702) 42.9 4.0SH-Ryn G23 500 2.793 ± 0.421 1.995(2) 1.801 (1.447–2.138) 128.6 11.8
a Number of neonate larvae used.b RF = LC50 of selected strain/LC50 of the Lab-Sus strain.
Table 4Toxicity of chlorantraniliprole on beet armyworm assayed by leaf-dip method with3rd-instar larvae.
Strains na LC50:mg AI/L (95% FL) Slope ± SE v2 (df) RFb
Lab-Sus 150 0.026 (0.019–0.034) 2.301 ± 0.415 0.518(2) –SH-Ryn G23 240 4.252 (3.341–5.408) 2.142 ± 0.345 0.220(3) 163.5DG10 240 0.512 (0.407–0.643) 2.063 ± 0.266 1.416(3) 19.7SZ10 240 0.579 (0.460–0.735) 2.004 ± 0.264 1.868(3) 22.3
a Number of 3rd-instar larvae used.b RF = LC50 of a population/LC50 of the Lab-Sus strain.
202 T. Lai et al. / Pesticide Biochemistry and Physiology 101 (2011) 198–205
inhibitor DEM and carboxyesterase inhibitor TPP were used for syn-ergism assays in SH-Ryn and Lab-Sus strains as well as two fieldpopulations (DG10 and SZ10) with moderate resistance (Table 6).
A
0.00
0.10
0.20
0.30
0.40
Lab-Sus
NJ09A
NJ09BSZ09
WH10NJ10
HF10XX10DG10ZZ10
SZ10
CS
n m
ol/m
in/m
g pr
otei
n
B
0
100
200
300
Lab-Sus
NJ09A
NJ09BSZ09HX10
WH10NJ10HF10XX10DG10ZZ10SZ10C
n m
ol/m
in/m
g pr
otei
n
C
0
500
1000
1500
2000
2500
Lab-Sus
NJ09A
NJ09BSZ09HX10
WH10NJ10HF10XX10DG10ZZ10SZ10CS
n m
ol/m
in/m
g pr
otei
n
Fig. 2. Detoxification enzyme activities (n mol/min/mg protein) of field populations anS. exigua. A and D: MFO. B and E: Esterase. Cand F: Glutathione-S-transferase, A, B andenzyme activities and LC50 values.
PBO showed some synergism on SH-Ryn, DG10, and SZ10 strainwith synergism ration SR 2.0, 1.7 and 2.2 respectively, however,the similar synergism ratio was also observed in the susceptiblestrain (Lab-Sus, RF = 1.9). This result implied that MFO might be in-volved in detoxification, but not in resistance of chlorantraniliprole.
GST inhibitor DEM had no synergistic effects on DG10, SZ10 andSH-Ryn strains, but increased the susceptibility of Lab-Sus strainslightly. TPP and DEF had no obvious effect on the toxicities of chl-orantraniliprole on all the four tested strains.
These results suggested that detoxification metabolism medi-ated by mixed function oxidase, glutathione-S-transferase, esteraseand carboxyesterase might not implicated in the resistance ob-served in DG10 and SZ10 populations and in the resistance ofselection strains SH-Ryn.
10NN10HZ10SH08
D
0.0
0.1
0.2
0.3
0.05 0.15 0.25 0.35
n mol/min/mg protein
LC
50(m
g/L
)
S10NN10HZ10SH08
E
0.0
0.1
0.2
0.3
0 50 100 150 200 250
n mol/min/mg protein
LC
50(m
g/L
)
10NN10HZ10SH08
F
0.0
0.1
0.2
0.3
500 1000 1500 2000
n mol/min/protein
LC
50(m
g/L
)
d the relationship between enzyme activities and toxicity of chlorantraniliprole onC are enzyme activity of field population. D, E and F are the relationship between
Table 5The changes of detoxification enzyme activities of S. exigua after selection with chlorantraniliprole.
Strains MFO EST GST
Activitya Rateb Activitya Rateb Activitya Rateb
SH08 0.236 ± 0.024a – 155.971 ± 20.380a – 1403.590 ± 19.699a –SH-Ryn G10 0.688 ± 0.033b 2.9 472.584 ± 35.467b 3.0 1477.672 ± 30.927a 1.1SH-Ryn G23 0.711 ± 0.028b 3.0 573.539 ± 32.892c 3.7 1489.520 ± 20.335a 1.1
Means (±SE) followed by different lowercase letters within column are significantly difference (a < 0.05).a Enzyme activity was showed as means ± SE (n mol/min/mg protein).b Rate = the enzyme activity in selected strain SH-Ryn/the activity in SH08 strain.
Table 6Synergistic effect of PBO, DEM, TPP and DEF on the toxicity of chlorantraniliprole to DG10, SZ10 and SH-Ryn strains of S. exigua.
Strains Treatment na LC50:mg/L (95% FL) Slope ± SE v2 (df) SRb
Lab-Sus Chlorantraniliprole 150 0.026 (0.019–0.034) 2.301 ± 0.415 0.518(2) –+PBO 150 0.014 (0.011–0.019) 2.165 ± 0.403 0.337(2) 1.9+DEM 150 0.015 (0.011–0.019) 2.550 ± 0.431 0.800(2) 1.7+TPP 180 0.024 (0.018–0.032) 1.884 ± 0.296 2.839(3) 1.1+DEF 180 0.026 (0.020–0.034) 2.247 ± 0.322 1.556(3) 1.0
SH-Ryn G23 Chlorantraniliprole 240 4.252 (3.341–5.408) 2.142 ± 0.345 0.220(2) –+PBO 240 2.078 (1.624–2.649) 2.104 ± 0.342 1.078(2) 2.2+DEM 200 3.358 (2.609–4.149) 2.247 ± 0.356 1.319(2) 1.3+TPP 240 3.193 (2.619–3.899) 2.472 ± 0.296 2.267(3) 1.3+DEF 240 3.762 (2.969–4.689) 2.296 ± 0.355 0.092(2) 1.1
DG10 Chlorantraniliprole 240 0.512 (0.407–0.643) 2.063 ± 0.266 1.416(3) –+PBO 240 0.301 (0.248–0.367) 2.545 ± 0.302 0.721(3) 1.7+DEM 240 0.481 (0.374–0.617) 1.841 ± 0.253 1.719(3) 1.1+TPP 240 0.547 (0.447–0.673) 2.384 ± 0.290 2.769(3) 0.9+DEF 240 0.398(0.318–0.492) 2.224 ± 0.278 0.700(3) 1.3
SZ10 Chlorantraniliprole 240 0.579 (0.460–0.735) 2.004 ± 0.264 1.868(3) –+PBO 240 0.260 (0.212–0.319) 2.405 ± 0.289 0.609(3) 2.2+DEM 240 0.541 (0.430–0.684) 2.032 ± 0.265 1.648(3) 1.1+TPP 200 0.451 (0.357–0.566) 2.039 ± 0.266 2.355(3) 1.3+DEF 240 0.387 (0.304–0.483) 2.074 ± 0.270 0.634(3) 1.5
a Number of 3rd-instar larvae used.b SR (synergism ratio) = LC50 of a strain treated with chlorantraniliprole alone divided by LC50 of the same strain treated with chlorantraniliprole plus a synergist.
T. Lai et al. / Pesticide Biochemistry and Physiology 101 (2011) 198–205 203
4. Discussion
S. exigua has become a major pest in various vegetable cropsand cotton in China. Various kinds of insecticides have been re-ported fail to control this species. The current study establishedthe baseline susceptible of S. exigua to chlorantraniliprole in China,this was the first time that geographical differences in susceptibil-ity of S. exigua to chlorantraniliprole had been recorded and obvi-ous variation of tolerance in field populations was observed. Theresults of the present study clearly demonstrated that several geo-graphical populations of S. exigua had developed moderate resis-tance to this insecticide. And the data should be useful forresistance monitoring and management in the future.
Our finding indicated that populations from different geograph-ical regions displayed varied tolerance or resistance level to chlo-rantraniliprole (6.2-fold variation). Most populations tested hadbecome less susceptible to this compound, and the resistance ofseveral populations had reached to moderate level compared withsusceptible strain kept in laboratory. Only one (WH09) in 18 pop-ulations remained susceptible. The highest resistance level wasfound in DG10 (RF = 17.1), SZ10 (RF = 15.1) and HZ10 (RF = 15.1)population which were collected from very close geographicallocations in southeast of China where the S. exigua had long dam-age period, and received frequent insecticidal exposures in a year.The resistance prevalence in beet armyworm from different areaswhere this insecticide was just introduced into market only for ashort time or had never been used for insect control in the past im-plied that the resistance might be a consequence of past insecticideapplications.
The susceptibility data of beet armyworm from field popula-tions was assayed by diet incorporation with neonate in this
research. Moulton et al. [18] pointed that the diet incorporation as-say with neonate was less sensitive for resistance detection oftebufenozide and methoxyfenozide than leaf-dip assay with3rd-instars. In current research the sensitivity difference betweentwo bioassays were not obvious based upon comparisons of RFsderived from dose–responses of DG10, SZ10 and SH-Ryn strainsversus the Lab-Sus strain (Table 2 and Table 4), the leaf-dip assayof chlorantraniliprole against 3rd-instars was only slightly moresensitive (RF = 19.7 versus 17.1 for DG10, RF = 22.3 versus 15.1for SZ10 and RF = 163.5 versus 128.6 for SH-Ryn strain) in detect-ing differences in susceptibility than was the diet incorporationassay. Although the leaf-dip bioassay was slightly more sensitivein discriminating the susceptibility of beet armyworm to chloran-traniliprole than diet incorporation, the diet incorporation assaywith neonate was much simpler and more timesaving than leaf-dip assay, so the diet incorporation assay with neonate was feasi-ble in resistance monitoring programme of chlorantraniliprole.
The susceptibility variation in field populations of four insectspecies to chlorantraniliprole has been reported recently. Narrowvariations in chlorantraniliprole susceptibility were observed inpopulations of Plutella xylostella (about 5-fold) [24], Cnaphalocrocismedinalis (5.8-fold) [34] and oriental fruit moth Grapholita molesta[35], However, significant variation in susceptibility existed infield-collected Choristoneura rosaceana populations with 39-foldresistance in the WEB populations[26]. Selection experimentshowed S. exigua might develop resistance to chlorantraniliproleunder continuous exposures [36]; therefore, the resistance risk tochlorantraniliprole should not be overlook.
Understanding the biochemical mechanisms conferring insecti-cide resistance has been shown to be necessary for resistance man-agement tactics. One of the most important factors of insect

204 T. Lai et al. / Pesticide Biochemistry and Physiology 101 (2011) 198–205
resistance is the increase in metabolic processes leading to higherdetoxification of insecticides by enzymes such as MFOs, ESTs andGSTs [37]. Beet armyworm resistance to pyrethroid and OP insecti-cides were related with the increases of MFOs activity [12,38–40]and EST activity [41,42], resistance to spinosad was also related withincrease of MFOs activity [43]. The oxidases are important in detox-ification of teburenozide in S. exigua [44]. Recently Cao et al.reported the increase in EST and GST activities of Helicoverpaarmigera larvae could be induced by chlorantraniliprole challenge[24]. Therefore, the enzyme activities of MFOs, EST and GSTs weremeasured to investigate metabolic resistance in the field popula-tions in comparison with the susceptible strain. The activities ofMFO, EST and GST varied significantly among the field populations;however when the correlation between toxicities of chlorantranili-prole on field populations and their activities of MFOs, ESTs andGSTs were plotted and analyzed respectively, no regression correla-tion was found between susceptibility and enzyme activities (Fig. 2).This result means that each field population might have differentbiochemical mechanisms for its tolerance to chlorantraniliprole.
When the field population SH08 was subjected to continuousexposures to chlorantraniliprole for 23 generation, the activitiesof MFOs and ESTs increased rapidly with 3.0-fold and 3.7-fold rise,respectively (Table 5). The boost in enzyme activities were incurredmainly in the first phase (1–10 generations), there was only aslightly increase in MFO activities during the latter phase of selec-tion of 11–23 generations. The activities of ESTs had obvious in-crease in the latter phase selection. Chlorantraniliprole selectiondid not have obvious effects on the activities of GSTs in S. exigua lar-vae. Significant increase of EST activity in the chlorantraniliprole se-lected colony of obliquebanded leafroller was also observed [45].The increase of MFOs and ESTs in selected resistance strain mightforebode the participation of MFOs and ESTs in the detoxificationon chlorantraniliprole, however, synergism assays with PBO, DEFand TPP didn’t demonstrate their involvement in resistance to thisinsecticide. The resistance in DG10 and SZ10 populations couldn’tbe blocked by synergist. Smirle et al. [46] discovered that the mech-anisms associated with resistance of C. rosaeana to azinphosmethylwas dependent on the area, the mechanisms responsible for resis-tance to the same chemical may vary from one population to an-other. Therefore, detoxification mechanisms responsible forchlorantraniliprole resistance observed in selected strain in thisstudy may not be the same as those existing in field populations.
The induction of MFOs and ESTs activities in S. exigua by chloran-traniliprole draws attention on the cross resistance and insecticideswhich couldn’t be rotated or mixed with chlorantraniliprole. Cao etal. [24] speculated chlorantraniliprole cannot rotate or mix withabamectin, b-cypermethrin and tebufenozide, and it cannot be usedto manage the resistance of temephos, malathion and pyrethioids,because chlorantraniliprole challenge may induce higher EST andGST activities in larvae and the resistance of temephos, malathionand pyrethroids were related with the increase of EST or GST activ-ity in H. armigera. Therefore, the spectrum of cross resistance withchlorantraniliprole in S. exigua should be clarified.
The fact that S. exigua has the potential risk to evolve resistanceto chlorantraniliprole [36], tolerant status of S. exigua from fieldcollected populations reported in this study should serve as awarning of vulnerability of chlorantraniliprole to resistance devel-opment in beet armyworm. Scientific strategies should belaunched as early as possible to avoid/delay the further develop-ment of resistance.
Acknowledgments
We thank J.X. Jiang, A.P. Zheng, M. Wang, Y.J. Liu and Y. Qiu forassistance in collecting field colonies of S. exigua. This study was
sponsored by The National Science Foundation of China (30671369 and 30971946).
References
[1] CAB, Distribution maps of pests, Spodoptera exigua (Hübner), CommonwealthAgricultural Bureau, London, Series A Map 302 (1972).
[2] W.C. Wang, L. Li, H.F. Jin, S.Y. Li, Preliminary observation of serious damage ofresistant cotton created by beet armyworm, China Cotton 10 (2002) 27–28.
[3] X.F. Jiang, L.Z. Luo, Outbreak of Spodoptera exigua (Hb.) and its control, PlantProt. 25 (1999) 32–34.
[4] S. Wu, Y. Gu, Z. Shen, H. Zheng, Monitoring of insecticide resistance andchemical control in beet armyworm, J. Plant Prot. 22 (1995) 95–96.
[5] Y.J. Liu, J.L. Shen, Monitoring for four group of insecticides resistance inSpodoptera exigua Lepidoptera: Noctuidae, Cotton Science 14 (2002) 356–360.
[6] S.A. Aldosari, T.F. Watson, S. Sivasupramaniam, A.A. Osman, Susceptibility offield populations of beet armyworm (Lepidoptera: Noctuidae) to cyfluthrin,methomyl, and profenofos, and selection for resistance to cyfluthrin, J. Econ.Entomol. 89 (1996) 1359–1363.
[7] K.Y. Wang, X.Y. Jiang, M.Q. Yi, B.K. Chen, X.M. Xia, Insecticide resistance ofSpodoptera exigua, Acta Phytoph. Sin. 29 (2002) 229–234.
[8] M.J. Brewer, J.T. Trumble, Field monitoring for insecticide resistance in beetarmyworm (Lepidoptera: Noctuidae), J. Econ. Entomol. 82 (1989) 1520–1526.
[9] M.J. Brewer, J.T. Trumble, Beet armyworm resistance to fenvalerate andmethomyl: resistance variation and insecticide synergism, J. Agric. Entomol. 11(1994) 291–300.
[10] B.K. Chen, K.Y. Wang, X.Y. Jiang, M.Q. Yi, Studies and surveys on the insecticideresistance of Spodoptera exigua, Acta Phytoph. Sin. 29 (2002) 366–370.
[11] D.L. Kerns, J.C. Palumbo, T. Tellez, Resistance of field strains of beet armyworm(Lepidoptera: Noctuidae) from Arizona and California to carbamateinsecticides, J. Econ. Entomol. 91 (1998) 1038–1043.
[12] M. Ahmad, M.I. Arif, Resistance of beet armyworm Spodoptera exigua(Lepidoptera: Noctuidae) to endosulfan Organophosphorus and pyrethroidinsecticides in Pakistan, Crop Prot. 29 (2010) 1428–1433.
[13] C.J. Pérez, P. Alvarado, C. Narváez, F. Miranda, L. Hernández, H. Vanegas, A.Hruska, A.M. Shelton, Assessment of insecticide resistance in five insect pestsattacking field and vegetable crops in Nicaragua, J. Econ. Entomol. 93 (2000)1779–1787.
[14] J.K. Moulton, D.A. Pepper, T.J. Dennehy, Beet armyworm (Spodoptera exigua)resistance to spinosad, Pest Manag. Sci. 56 (2000) 842–848.
[15] A. Osorio, A.M. Martínez, M.I. Schneider, O. Díaz, J.L. Corrales, M.C. Avilés, G.Smagghe, S. Pineda1, Monitoring of beet armyworm resistance to spinosad andmethoxyfenozide in Mexico, Pest Manag. Sci. 64 (2008) 1001–1007.
[16] V.J. Mascarenhas, J.B. Graves, B.R. Leonard, E. Burris, Dosage mortalityresponses of third-instars of beet armyworm (Lepidoptera: Noctuidae) toselected insecticides, J. Agric. Entomol. 15 (1998) 125–140.
[17] J.K. Moulton, D.A. Pepper, R.K. Jansson, T.J. Dennehy, Pro-active management ofbeet armyworm (Lepidoptera: Noctuidae) resistance to tebufenozide andmethoxyfenozide: baseline monitoring, risk assessment, and isolation ofresistance, J. Econ. Entomol. 95 (2002) 414–424.
[18] W.J. Moar, M. Pusztai-Carey, H. Van Faassen, D. Bosch, R. Frutos, C. Rang, K.Luo, M.J. Adang, Development of Bacillus thuringiensis CryIC resistance bySpodoptera exigua (Hübner) (Lepidoptera: Noctuidae), Appl. Environ.Microbiol. 61 (1995) 2086–2092.
[19] Insecticide Resistance Action Committee, IRAC Mode of Action Classification, v.6.3, IRAC Mode of Action Working Group, (2009) http://www.irac-online.org/documents/MoA%20classification_v6.3.3_28july09.pdf.]
[20] G.P. Lahm, D. Cordova, J.D. Barry, New and selective ryanodine receptoractivators for insect control, Bioorg. Med. Chem. Lett. 17 (2009) 4127–4133.
[21] D.B. Sattelle, D. Cordova, T.R. Cheek, Insect ryanodine receptors: moleculartargets for novel pest control chemicals, Invert. Neurosci. 8 (2008) 107–119.
[22] G.P. Lahm, T.P. Selby, J.H. Freudenberger, T.M. Stevenson, B.J. Myers, G.Seburyamo, B.K. Smith, L. Flexner, C.E. Clark, D. Cordova, Insecticidalanthranilic diamides: a new class of potent ryanodine receptor activators,Bioorg. Med. Chem. Lett. 15 (2005) 4898–4906.
[23] X.L. Wang, X.Y. Li, A.D. Shen, Y.D. Wu, Baseline susceptibility of diamondbackmoth (Lepidoptera: Plutellidae) to chlorantraniliprole in China, J. Econ.Entomol. 103 (2010) 843–848.
[24] G. Cao, Q. Lu, L. Zhang, F. Guo, G. Liang, K. Wu, K.G. Wyckhuys, Toxicity ofchlorantraniliprole to CryAc-susceptible and resistant strain of Helicoverpaarmigera, Pestic. Biochem. Physiol. 98 (2010) 99–103.
[25] A.A. Sial, J.F. Brunner, M.D. Doerr, Susceptibility of Choristoneura rosaceana(Lepidoptera: Tortricidae) to two new reduced-risk insecticides, J. Econ.Entomol. 103 (2010) 140–146.
[26] G.P. Lahm, T.M. Stevenson, T.P. Selby, J.H. Freudenberger, D. Cordova, L.Flexner, C.A. Bellin, C.M. Dubas, B.K. Smith, K.A. Hughes, J. Gary Hollingshaus,C.E. Clark, E.A. Benner, Rynaxypyr™: a new insecticidal anthranilic diamidethat acts as a potent and selective ryanodine receptor activator, Bioorg. Med.Chem. Lett. 17 (2007) 6274–6279.
[27] Dupont, Rynaxypyr™ insect control technical bulletin (2007). http://www2.dupont.com/Production_Agriculture/en_US/assets/downloads/pdfs/Rynaxypyr_Tech_Bulletin.pdf).
[28] R. Rose, L. Barbhaiya, R. Roe, G. Rock, E. Hodgson, Cytochrome P-450-associated insecticide resistance and the development of biochemical

T. Lai et al. / Pesticide Biochemistry and Physiology 101 (2011) 198–205 205
diagnostic assays in Heliothis virescens, Pestic. Biochem. Physiol. 51 (1995)178–191.
[29] Y. Yang, Y. Wu, S. Chen, G.J. Devine, I. Denholm, P. Jewess, G.D. Moores, Theinvolvement of microsomal oxidases in pyrethroid resistance in Helicoverpaarmigera from Asia, Insect Biochem. Mol. Biol. 34 (2004) 763–773.
[30] M.M. Bradford, A rapid and sensitive method for the quantification ofmicrogram quantities of protein using the principle of protein-dye binding,Anal. Biochem. 726 (1976) 48–254.
[31] LeOra software, 2002. POLO-Plus, A use’s guide to probit or logit analysis,LeOra Software, Berkeley, CA.
[32] J. Keiding, Development of resistance to pyrethroids in field populations ofDanish house flies, Pestic. Sci. 7 (1976) 283–291.
[33] WHO, Status of resistance in houseflies, Musca domestica, Document VBC/EC/80.7, (1980), World Health Organization, Geneva.
[34] X. Zheng, X, Ren, J. Su, Insecticide susceptibility of Cnaphalocrocis medinalis(Lepidoptera: Pyralidae) in China, J. Econ. Entomol 104 (2011) 653–658.
[35] M.M. Jones, J.L. Robertson, R.A. Weinzierl, Susceptibility of oriental fruit moth(Lepidoptera: Tortricidae) larvae to selected reduced-risk insecticides, J. Econ.Entomol. 103 (2010) 1815–1820.
[36] T. Lai, J. Su, Assessment of resistance risk in Spodoptera exigua (Hübner)(Lepidoptera: Noctuidae) to chlorantraniliprole. Pest Manag Sci, 2011,10.1002/ps.2201.
[37] M. Mohan, G.T. Gujar, Local variation in susceptibility of the diamondbackmoth, Plutella xylostella (Linnaeus) to insecticides and role of detoxificationenzymes, Crop Prot. 22 (2009) 495–504.
[38] Y.J. Liu, J.L. Shen, Biochemical mechanism and genetics of resistance tolambdacyhalothrin in the beet armyworm, Spodoptera exigua, and the relativefitness of the resistant strain, Acta Entomol. Sin. 46 (2003) 567–572.
[39] Y.J. Liu, J.L. Shen, X.D. Zhao, P.J. Xu, H.R. Shu, Relationship betweenmultifunction oxidases, the resistance to lambda-cyhalothrin in beetarmyworm, Spodoptera exigua Chinese, J. Pestic. Sci. 7 (2005) 19–23.
[40] K. Natsuhara, K. Shimada, T. Tanaka, T. Miyata, Phenobarbital induction ofpermethrin detoxification and phenobarbital metabolism in susceptible andresistant strains of the beet armyworm Spodoptera exigua (Hübner), Pestic.Biochem. Physiol. 79 (2004) 33–41.
[41] E. Garza-Urbina, A.P. Teran-Vargas, Beet armyworm (Spodoptera exigua)resistance mechanisms to insecticides in Southern Tamaulipas, Mexico. In:Proceedings of Beltwide Cotton Conferences, (1998) pp. 1343–1344.
[42] R. Delorme, D. Fournier, J. Chaufaux, A. Cuany, J.M. Bride, D. Auge, D.W. Berge,Esterase metabolism and reduced penetration are causes of resistance todeltamethrin in Spodoptera exigua Hübner (Noctuidae: Lepidoptera), Pestic.Biochem. Physiol. 32 (1988) 240–246.
[43] W. Wang, J. Mo, J. Cheng, P. Zhuang, Z. Tang, Selection and characterization ofspinosad resistance in Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae),Pestic. Biochem. Physiol. 84 (2006) 180–187.
[44] G. Smagghe, T.S. Dhadialla, S. Derycke, L. Tirry, D. Degheele, Action of theecdysteroid agonist tebufenozide in susceptible and artificially selected beetarmyworm, Pestic. Sci. 54 (1998) 27–34.
[45] A.A. Sial, J.F. Brunner, S.F. Garczynski, Biochemical characterization ofchlorantraniliprole and spinetoram resistance in laboratory-selectedobliquebanded leafroller, Choristoneura rosaceana (Harris) (Lepidoptera:Tortricidae), Pestic. Biochem. Physiol. 99 (2011) 274–279.
[46] M.J. Smirle, C. Vincent, C.L. Zurowski, B. Rancourt, Azinphosmethyl resistancein the obliquebanded leafroller, Choristoneura rosaceana: reversion in theabsence of selection and relationship to detoxification enzyme activity, Pestic.Biochem. Physiol. 61 (1998) 183–189.
Copyright © 2022 FDOKUMEN




















