
Larval Mortality Factors of Spodoptera Littoralis in the Azores
Upload
independentCategory
view
0download
0
J. Asia-Pacific Entomol. 8 (4): 397-41 0 (2005)www.entomology.or.kr
CHEMICAL CONTROL
Monitoringand Characterization ofInsecticideResistance intheCotton Leafworm,Spodoptera Iittoralis (Boisd.) (Lepidoptera: Noctuidae)Gamal E. Abo JElghar*, Zeinab A. Elbermawy, Adel G. Yousef and Hany K. Abd Elhady
Department of Pesticides, Faculty of Agriculture, Menoufiya University, Shebin EI-Kom 32511, Egypt
Abstract Resistance to several classes of insecticideswas diagnosed in the cotton leafworm (CL W),Spodoptera littoralis (Boisd.), from cotton fields inthe Nile Delta Egypt through 2002-2004 seasons.Two types of laboratory bioassays were used forthe detection of insecticide resistance. Leaf-dip larvalbioassay and discriminating concentration (LC99 forsusceptible laboratory strain) technique revealed thepresence of resistance to insecticides tested. Both larvaland adult stages showed relatively similar responseto most insecticides tested. However, the glass vialdiscriminating concentration technique is particularlymore useful when resistance is related to the target-site insensitivity rather than to increased metabolism.The pattern of reversion of resistance to five insecticides was determined in two field strains that havebeen released from continuous insecticide applicationand then reared under laboratory conditions for 6-8generations in the absence of insecticide pressure.It was considered that losing of resistance to cypermethrin in both resistant field strains seemed to bequicker than other insecticides tested, in which resistance ratios decreased from 47.7-fold (F0- parents)to 17.8- fold (F6 generation) in MNF- strain, and from38.5- fold (Fs-parents) to 10.7-fold (F6 generation)in KFR-strain. On the other hand, biochemical assaysindicated that both MNF- and KFR-field strains, expressed higher levels of acetylcholinesterase (AChE)activity by 13.2- and 8A-times, respectively, than thatof the susceptible strain. AChE activity was sharplydecreased to 1.01- and 1.92- times in MNF- and KFRstrains, respectively, compared to that of susceptiblestrain, following insect rearing for six successivegenerations without insecticide pressure. Esterasesactivity was also decreased 2.02-times (F0) to 1.41(F6) in MNF-strain and from 2.0-times (Fo) to 1.21-times (F6) in KFR-strain, compared to that of susceptible strain. On the other hand, analysis of esterases by native polyacrylamide gel electrophoresis
·Corresponding author.E-mail: [email protected]: +20-2-5769495; Fax: +20-2-5769495
(Received September 9, 2005; Accepted December 22, 2005)
(PAGE) confirmed the association between esterasesactivity and resistance to insecticides tested in bothfield strains.
Key words acetylcholinesterase, discriminating vialtechnique, esterase, insecticide resistance, resistancereversion, Spodoptera littoralis
Introduction
The Egyptian cotton leafworm, Spodoptera littoralis(Boisd.), has a distribution in northern Africa, MiddleEast, Mediterranean countries, and Near East. It iswell known as one of the most destructive agriculturallepidopterous pests within its subtropical and tropicalrange. It can attack numerous economically importantcrops all the year round. On cotton, the pest maycause considerable damage by feeding on the leaves,fruiting points, flower buds and occasionally on bolls(Avidov and Harpaz, 1969). It is a true generalistspecies with a seemingly promiscuous feeding strategy.Feeding has been observed on at least 84 plant specieswithin 40 families (Brown and Dewhurst, 1975). Thechemical control of S. littoralis has been extensivelyreported in relation especially to cotton in Egypt. Theoverlapping of crops that serve as hosts of this insectthroughout the annual cropping cycle encourages highpopulation densities. As a result of this and the intensive use of chemical control measures to manage thispest in particular on cotton has resulted in the development of resistance to almost all classes of insecticidesused. Until 1968, S. littoralis was held in check bymethyl-parathion, but then resistance to this compounddeveloped. Since then, numerous other organophosphorus,synthetic pyrethroid and other insecticides have beenused, with appearance of resistance and cross resistance in many cases (Issa et al., 1986; EI-Guindy etal., 1989;Abdallah, 1991;Rashwan et al., 1992).However,compulsory limitation of the application of syntheticpyrethroids to one per year on cotton in Egypt hasstopped the appearance of new resistance (Sawicki,

398 J. Asia-Pacific Entomol. Vol. 8 (2005)
1986). Moreover, some levels of resistance to insectgrowth regulators (IGRs), as unconventional insecticides, have been documented in this species for methoprene [juvenile hormone analogue, (JHA)] (El-Guindyet al., 1989), and also for benzoylphenylureas (BPUs)(chitin synthesis inhibitors) (El-Guindy et al., 1983;Abo-Elghar et al., 1992). In Egypt, management strategyagainst this pest has been settled upon the last decadeon cotton to preserve and extend insecticide efficaciousbased on rotating various insecticides including organophosphates, carbamates, insect growth regulators,and pyrethroids every year (Sawicki and Denholm,1987; Temerak, 2002). Given the history of resistanceto various insecticides of insect pests of cotton, a needcontinues to develop baseline data, to monitor forchanges in tolerance, and to detect any problems thatmay occur (Sparks, 1981; Staetz, 1985). Therefore,resistance monitoring is fundamentally important toevery resistance management program. Continuousmonitoring of the populations for changes in resistancefrequencies allows decisions regarding effective management strategies to be made (Prabhaker et al., 1996).Provided that a monitoring survey is sufficientlyextensive, patterns ofresistance can be detected withina region or between regions. Ideally, current resistancemonitoring data should be communicated to growersand pest control advisors who are interested in participating in a resistance management program. With thisinformation, they could alternate to a different insecticide class or alternative control strategy to avoidincreasing selection pressure to populations (Prabhakeret al., 1996). Successful resistance monitoring dependson the availability of simple and reliable methods ofmonitoring. Biochemical and bioassay methods arethe primary means of testing for insecticide resistance.Recent interest has focused on adapting these techniques, especially in lepidopterous insect pests to providefield-based monitoring of insecticide resistance (Brewerand Trumble, 1989; McCutchen et al., 1989). Biochemical techniques are advantageous because they testfor activity that is directly linked to a resistance mechanism (Pasteur and Georghiou, 1981; Miyata, 1986).Unfortunately, such techniques are currently restrictedto the examination of esterase activity and intensiveacetylcholinesterase, which limits the use of organophosphate and carbamate pesticides (Brewer andTrumble, 1989). Although monitoring for lepidopterous adults, e.g. tobacco budworm, Heliothis virescens(F.), resistance indicated the presence of pyrethroidresistance in different cotton-producing areas, thetechnique is not appropriate for determining resistancein individual fields or in larvae, the actual pest stage(Roush and Luttrell, 1987). The adult vial tests becomea standard method for monitoring insecticide resistancein cotton pests. The procedure was initially developed to monitor for resistance to pyrethroids in adulttobacco budworm, H. virescens (F.) (Plapp et al.,
1987) and later modified by Kanga and Plapp (1992).On the other hand, biochemical assays may also beused to detect and monitor resistance. Biochemicalassays measure changes in sensitivity at the pesticide's target site or measure metabolic activities of thepest that are responsible for, or at least linked to,resistance (Bush et al., 1993). Biochemical assaysmay be a better choice for detecting resistance at lowergene frequencies than bioassays (Brown and Brogdon,1987; ffrench-Constant and Roush, 1990). A biochemical assay used by Carlini et al. (1991) showed elevatednonspecific esterase activity linked to azinphos-methylresistance in adults of the tufted apple bud moth,Platynota idaeusalis (Walker). In another tortricid pestof apple, the light brown apple moth Epiphyaspostvittana (Walker), Armstrong and Suckling (1990)demonstrated a positive correlation between nonspecificesterases, glutathione transferases, and azinphos-methylresistance. If the biochemical bases of resistance canbe determined, highly sensitive monitoring techniquescan be devised; these techniques are key factors indeveloping successful resistance management programs(ffrench-Constant and Roush, 1990). Rapid biochemical assays coupled with biological assays are potential tools for estimating both the intensity and thefrequency of resistance in the field (Brown and Brogdon, 1987; Devonshire et al., 1986).
The objectives of this study were (I) to furtherdescribe the expression of resistance to different insecticides in both larvae and adults of field populationsof the cotton leafworm (CLW) collected from cottonfields through three successive seasons (2002-2004)in the Nile Delata, (2) to standardize a rapid andsimple monitoring system that is versatile enough toaccommodate most, ifnot all, insecticides used againstcotton leafworm, (3) to determine the pattern ofreversion of insecticide resistance in field populations,and (4) to investigate the activities of esterases andacetylcholinesterase in both resistant and susceptiblestrains.
Materials and M'ethods
Insects
A susceptible strain of S. littoralis was obtained fromthe Central Agricultural Pesticides Laboratory, Dokki,Egypt, where it has been reared on fresh castor bean(Ricinus communis) leaves for several generationswithout exposure to insecticides. It served as the baseline reference strain for the comparisons between thestudied strains. Two S. littoralis field strains werecollected at the commercial cotton fields located inMenoufia (MNF) and Kafr-Elsheikh (KFR), Nile Delta

area in August throughout three successive seasons(2002-2004). These populations were collected fromcotton fields which had been exposed to the annualroutine chemical control program which directed byEgyptian Agriculture Ministry (EAM) for controllingcotton pests from mid-June to mid-August. Egg massesof S. littoralis strains were collected at these cottonfields in August after the last insecticide applications,where numerous S. littoralis field control failures withseveral insecticides belonging to different groups, i.e.organophosphates, carbamates, pyrethroids, and insectgrowth regulators, were documented. Eggs of eachstrain were allowed to hatch at room temperature, atwhich larvae were reared on fresh castor bean, Ricinuscommunis, leaves as described by Eldefrawi et al.(1964).
Chemicals
Formulated insecticides used in our experiments were:chlorpyrifos (Dursban 48% emulsifiable concentrate[Ee], DowElanco Co.); profenofos (Curacron 72%EC, Ciba-Geigy AG); methomyl (Lannate 90% SL[soluble powder] E. I. du Pont de Nemours & Co.);carbaryl (Sevin 85% WP [wettable powder], RhonePoulenc); thiodicarb (Larvin 80% DF [dust flowable],Union Carbide Agricultural Co., Inc.); esfenvalerate(Sumi-Gold 20% EC, Sumitomo Chemical Co. Ltd.);fenpropathrin (Danitol 30% EC, Sumitomo ChemicalCo. Ltd.); cypermethrin (Cymbush 10% EC, Ciba-GeigyAG); hexaflumuron (Cosult 10% EC, DowElanco Co.);flufenoxuron (Cascade 5% EC, American CyanamidCo.); spinosad (Success 12% SC [suspension concentrate],Dow AgroSciences); and Beauveriabassiana (Naturalis-07.16% EC , Troy BiosciencesInc.).All other chemicalsused in biochemical assays were purchased fromSigma Chemical Company as analytical standards.
Leaf dip bioassay
Leaf dip technique was used for larval bioassays todetermine the responses of the CLW of field strainsto several classes of selected insecticides, i.e. organophosphates, carbamates, pyrethroids, chitin synthesisinhibitors, and biopesticides, used for controlling CLWin cotton fields in Nile Delta area. Stock solution ofeach insecticide formulation was prepared using thetap water, and then six to seven serially diluted concentrations were prepared. The castor bean leaveswere dipped into tap water (control) or insecticidewater solution for 20-30 s, removed and allowed todry. At least six concentrations and three replicationswere used to estimate each concentration-mortalityline. Ten fourth-instar larvae (I-day-old) were transferredto 250-ml glass jar. Treated leaf was placed over the
Insecticide resistance of Spodoptera littoralis 399
jar and covered by a piece of muslin. All jars wereinverted and held at 27"C ± 2, 65-70% RH, and aphotoperiod of 14:10 (L:D) h. Mortality was assessed24-72 h after treatment. Larvae were considered deadif unable to move in a coordinated way when proddedwith a blunt probe. Control mortality was never> 15%;data were corrected by Abbott's (1925) formula. Thedata were analyzed by probit analysis (Finney, 1971).Resistance factors were calculated as the resistantLCso/susceptible LCso.
Discriminating (Diagnostic) concentration bioassay
The diagnostic (discriminating) concentration techniquewas used for monitoring the resistance of the larvaeand adults ofCLW field strains. The glass-vial technique(Plapp et al., 1987) was modified and used to determineresistance levels of field populations of S. littoralis.Glass scintillation vials (20 ml) were treated withdiscriminating concentrations (Magaro and Edelson,1990) of each insecticide dissolved in acetone. Thediscriminating concentration was the LC99 estimatedfrom the leaf dip test with the susceptible-laboratory(S) strain. The LC99 and associated 95% fiducial limitsof the S strain were estimated by probit analysis. Basedon considerations discussed by Roush and Miller (1986)and Halliday and Burnham (1990), we sought to estimate a concentration that would reliably cause-99%mortality of the S strain, but not so high that it wouldfail to detect resistance. Data for three or four testsfor each insecticide at each of four concentrations,chosen according to the suggestions of Robertsonet al. (1984), were used to estimate a diagnostic concentration. We selected concentrations that were equalto or slightly greater than the median LC99 estimates,and after some preliminary trials, increased the concentration as needed. After the diagnostic concentration was estimated for the S strain, the actual mortalityat the diagnostic concentration was evaluated two orthree times for each insecticide against the S and fieldstrain. The mortality response at a single diagnosticconcentration was also corrected for control mortality(Abbott, 1925). One ml of each insecticide solutionwas pipetted into a scintillation vial and the vial wasrolled for approximately 1 min to ensure that all surfacesreceived treatment. All remaining solution was thendrained. Vials were air dried (upright) in a fume hoodat 25"C for 24 h before use. Control vials were precoated with acetone only.
For larval bioassays, the second- and fourth-instarswere collected from KFR-field strain and subjectedto discriminating concentration bioassay for monitoringof insecticide resistance. Seven larvae of 2nd instarsor five larvae of 4th instars were placed in each vial(replicate) with leaf disk treated with the discriminating concentration of tested insecticide. Cotton

400 1. Asia-Pacific Entomol. Vol. 8 (2005)
swabbing was used to plug the vial opening. Eachtreatment was replicated five times. Vials with larvaewere placed in a controlled temperature chamber at25°C, and mortality was assessed for 24 h. Criteriafor determining mortality was the same as describedin the leaf dip bioassay. Percentage mortality for eachfield populations was compared with that for thesusceptible laboratory colony.
For adult bioassay, the glass-vial technique describedby Plapp et at. (1987) was modified and used formonitoring the resistance of CLW moths. Male mothswere taken from the susceptible laboratory strain andtested to estimate the discriminating concentration(LC99) ofeach insecticide tested. One moth was placedin a 20-ml glass scintillation vial pre-coated with eachinsecticide dilution as described above. Five to sixconcentrations (10-15 moths per concentration) wereused for each adult vial test. Vials treated with acetonewere used as controls. For each vial, a small pieceof cotton soaked with 10 % sugar solution was added.Vials containing moths were stored on their sides atroom temperature. Mortality was determined 24 hafter exposure. Adults that were unable to fly wereconsidered dead. Control mortality was never> 10%;data were corrected with Abbott's (1925) formula andsubjected to probit analysis (Finney, 1971). For monitoring the insecticide resistance of the field strain,adults were placed in vials treated with discriminatingconcentrations. At least ten replicates and one mothper replicate were used to predict which insecticidewould result in the best control of the population andan insecticide that would not control the populationas well. Resistance percentages were determined byusing the following formula (McCutchen et al., 1989):Resistance Percentage = 1OO-(MF/MS X 100); where,MF = % mortality at discriminating concentration infield-collected larvae (or adults) and MS = % mortality(constant) at discriminating concentration in susceptiblelarvae (or adults).
Reversion of resistance to insecticides infield strains
A colony of CLW population was collected fromcommercial cotton fields in KFR during August 2002at which received 4-5 applications of different insecticides through the normal routine program for cottonpest control directed by EAM before field collection.Larvae collected from these fields showed resistancelevels against chlorpyrifos, methomyl, esfenvalerate,fenpropathrin, and cypennethrin. These colonies wereestablished in the laboratory. Monitoring was performed with fourth-larval instars using the leaf diptechnique mentioned above. The reversion study wasreported for monitoring of resistance stability of eachcolony through 6-8 successive generations.
Biochemical aSSclYS
Acetylcholinesterase (AChE) assay, AChE assays weremeasured using CLW adult head homogenates following the method of Ellman et at. (1961). Ten adultsfrom each strain were decapitated and homogenizedin 10 ml (1 ml for each head) of ice-cold buffer (0.1M phosphate buffer, pH 8.0) with Teflon-pestle for30 s. The homogenate was centrifuged at 10,000 gfor 15 min. Resulting supernatants were held in iceand used for enzyme assays within 30 min of preparation. In each test, the reaction mixture consistedof 2.5 ml 0.1 M phosphate buffer (pH 8.0), 100 fL1color reagent (0.01 M dithiobisnitrobenzoic acid, DlNB),100 fL1 substrate (0.03 M acetylthiocholine iodide,ATC), and 500 f-Ll insect homogenate. Thiocholine,the product of the hydrolysis of substrate reacts withDTNB to produce a yellow anion 5-thio-2-nitrobenzoicacid. The rate of change in absorbance during 2 minwas measured at 412 nm. Data were corrected usingblank test without substrate in the mixture reaction.Protein content was determined using the method ofBradford (1976). The AChE activity was determinedusing an extinction coefficient of 13.6 mM-J cm-I forthionitrobenzoic acid.
Esterase assays. Esterase activity towards cx-NA wasquantified using the assay method of Van Asperen(1962). After dissecting five midguts from fifth-instarCLW larvae of each strain, each midgut was splitlongitudinally, rinsed with ice-cold phosphate buffer(0.04 M, pH 7.0) to remove gut contents, and washedin ice-cold 1.15 % KCI. The midguts were masshomogenized with Teflon-pestle for 30 s in ice-cold0.04 M sodium phosphate buffer, pH 7.0, and thenfiltered through the cheesecloth. The homogenate wascentrifuged at 10,000 g for 15 min at 4°C. The supernatants were used as enzyme sources. The esteraseassay was done measuring the production of a-naphtholfrom a- naphthyl acetate substrate. Each sample wasadded to a total volume of 3.0 ml reaction mixturecontaining: 25 f-Ll of0.03 M a- naphthyl acetate, 2.475ml of 0.04 M sodium phosphate buffer (pH 7.0) and0.5 ml tissue sample. The reaction mixture wasincubated at 27°C for 30 min. After incubation, thereaction was stopped, and color was developed byaddition of 0.5 ml Diazoblue B salt (0.3%) in 3.5%aqueous sodium lauryl sulfate mixture (DBLS). Theresulting complex was measured, 10 min later, colorimetrically, at 600 nm, with an enzyme blank. Theconcentration of hydrolyzed substrate was determinedfrom a standard curve of a-naphthol. Esterase activitywas presented relative to protein concentration, whichwas measured using the assay of Bradford (1976) withbovine serum albumin (BSA) as the standard. Theesterase activity was expressed as fL mol a-naphtholproduced/min/mg protein.

Electrophoretic analysis of esterases. Non-denaturingpolyacrylamide gel electrophoresis (PAGE) of the ex:
-NA hydrolyzing esterases from different CLW strainswas carried out using MiniProtean vertical minigelapparatus (Bio-Rad Hercules, CA) (Williams and Reisfold, 1964). Batches of 10 midguts from fifth-larvalinstars of each strain were dissected as describedabove and mass homogenized in ice-cold 0.04 Msodium phosphate buffer, pH 7.0, and then filteredthrough the cheesecloth. The homogenate was centrifuged at 10,000 g for 15 min' at 4°C. The supernatantswere used as enzyme sources for PAGE analysis. Eachlane was loaded with same protein amount to allowcomparison between samples. The gel (4 and 7.5%acrylamide in stacking and separating gels, respectively)was run at two steps: firstly, at 100 V for 60 min,and then, at 200 V for 140 min. After electrophoresis,esterase activity was detected by incubation of thegel in O.I-M sodium phosphate buffer (pH 6.5) containing 3.2 mM a- or b-naphthyl acetate and 2.4 mM
Insecticide resistance of Spodoptera littoralis 401
Fast Blue BB salt for 60 min in the dark at 37°C.The intensities of the bands on each gel were thenscanned with a computing densitometer at absorbanceof 6J3 nm and results were graphically depicted withImage Phoretic (IDA) Quantifier software (PhoretixInternational, London). The peak heights, Rf , intensityand area were used as a relative comparison of bandintensity between zygomorphs on the same gel.
Results and Discussion
Resistance monitoring in field populations
Responses of fourth-larval instars from the Labsusceptible and field strains collected during the cottonseason of 2002-2004 to selected insecticides werevaried (Tables 1-3). Data in Table I showed that tests
Table 1. Responses of fourth-instar S. littoralis larvae from field strains collected during 2002-2004 seaons to selectedorgaonophosphate and carbamate insecticides as assayed by leaf dip technique bioassay
2002 season 2003 season 2004 seasonInsecticide Strain LC
50o,b Slope
RRc LC50
o,b SlopeRRc LC50
G,b Slope RRc
(95% FL) (SE) (95% FL) (SE) (95% FL) (SE)
L-SUS 78.55 1.45 78.55 1.45 78.55 1.45(53.2-104.3) (0.23) (53.2-104.3) (0.23) (53.2-104.3) (0.23)
Profenofos MNF237.5 2.0
3.02295.5 2.01
3.76 536.2 2.33 6.83(175.0-299.4) (0.28) (226.6-367.5) (0.28) (433.9-640.8) (0.29)
KFR 553.13 1.68 7.04 428.4 2.41 5.45 626.93 2.12 7.98(431.6-763.0) (0.30) (339.6-514.7) (0.30) (506.6-756.9) (0.28)
L-SUS 27.49 1.71 27.49 1.71 27.49 1.71(19.6-35.1) (0.25) (19.6-35.1 ) (0.25) (19.6-35.1 ) (0.25)
Chlorpyrifos MNF 423.81 1.87 15.42 697.65 2.04 25.38 651.06 1.30 23.68(335.6-536.2) (0.31) (569.1-896.5 ) (0.36) (467.6-932.2) (0.23)
KFR 481.46 2.2617.51
642.0 1.98 23.35 679.76 1.98 24.73(382.3-579.9) (0.29) (524.7-815.4) (0.29) (551.5-875.9) (0.35)
L-SUS24.78 2.27 24.78 2.27 24.78 2.27
(20.5-29.8) (0.27) (20.5-29.8) (0.27) (20.5-29.8) (0.27)
Methomyl MNF867.8 3.23
35.02 988.3 3.5339.88
758.07 3.1530.59(763.3-1001.8) (0.41) (873.5-1144.2) (0.50) (664.4-868.9) (0.39)
KFR 568.86 3.2122.96 920.4 3.38 37.15 822.3 3.25 33.18(494.1-663.1 ) (0.45) (812.7-1062.1) (0.43) (723.8-843.9) (0.41)
L-SUS 127.63 127.63 2.35 127.63 2.35(104.0-153.0) (104.0-153.0) (0.36) (104.0-153.0) (0.36)
Thiodicarb MNF d 2300.6 2.1118.03 2354.25 2.66 18.45(1912.0-3114.7) (0.45) (2019.2-2963.4) (0.47)
KFR d 1508.8 2.6711.82
1644.9 3.01 12.89(1292.3-1841.0) (0.48) (1425.8-1995.7) (0.50)
L-SUS112.31 2.34 112.31 2.34 112.31 2.34
(90.7-134.4) (0.36) (90.7-134.4) (0.36) (90.7-134.4) (0.36)
Carbaryl MNF3010.8 2.30
26.813134.93 2.97 27.91(2549.5-3721.6) (0.43) (2746.1-3694.2) (0.48)
KFR d 1756.5 2.36 15.64 2030.7 2.41 18.08(1458.9-2129.7) (0.36) (1699.1-2496.3) (0.38)a Expressed as ppm; h 24 h after larval feeding on treated leaves.c Resistance ratio (RR) calculated by dividing the LCso for the field strain by the LCso for the Lab-susceptible strain.d Data not available due to loss of strain.

402 J. Asia-Pacific Entomol. Vol. 8 (2005)
Table 2. Responses of fourth-instar S. littoralis larvae from field strains collected during 2002-2004 seaons to selectedpyrethroid insecticides as assayed by leaf dip technique bioassay
2002 season 2003 season 2004 seasonInsecticide Strain LCso
G•b Slope RRc LCso"·b Slope
RF~cLCso",b Slope RRc
(95% FL) (SE) (95% FL) (SE) (95% FL) (SE)
L-SUS12.30 1.35 12.30 1.35 12.30 1.35
(8.88-16.61 ) (0.16) (8.88-16.61 ) (0.16) (8.88-16.61 ) (0.16)
Esfenvalerate MNF207.2 1.99 16.92 317.6 2.33
25.93242.1 2.22
19.77(161.4-255.3) (0.34) (262.3-383.5) (0.34) (191.1-298.4) (0.35)
KFR 207.27 1.99 183.57 2.39 236.56 2.32(161.4-255.4) (0.34) (141.2-222.8) (0.35) (189.1-284.6) (0.34)
L-SUS 38.40 2.63 38.40 2.63 38.40 2.63(32.9-45.0) (0.35) (32.9-45.0) (0.35) (32.9-45.0) (0.35)
Fenpropathrin MNF 436.9 1.85 11.38 361.0 1.93 9.40286.1 1.92
7.45(354.1-575.7) (0.33) (294.1-451.1 ) (0.32) (227.7-350.8) (0.31)
KFR 436.98 1.85 398.58 2.01 361.32 2.39(354.1-575.7) (0.33) (327.3-502.6) (0.33) (304.6-431.6) (0.35)
L-SUS 14.54 1.25 14.54 1.25 14.54 1.25(10.2-21.2) (0.16) (10.2-21.2) (0.16) (10.2-21.2) (0.16)
Cypermethrin MNF 559.0 1.8238.45 1431.5 2.56
98.46803.0 2.09
55.23(422.8-702.2) (0.24) (1201.9-1830.1) (0.42) (637.0-980.3) (0.37)
KFR 559.08 1.82 38.45 1879.0 2.21129.24 1438.8 2.20
98.96(422.8-702.2) (0.24) (1551.4-2325.7) (0.39) (1153.5-1740.2) (0.37)
" Expressed as ppm; b 24 h after larval feeding on treated leaves.
c Resistance ratio (RR) calculated by dividing the LC,o for the field strain by the LC,o for the Lab-susceptible strain.
with organophosphates revealed an increase in resistance levels to chlorpyrifos in MNF-strain from 15.4fold in 2002 to 25.4- and 23.7-fold in 2003 and 2004seasons, respectively. However, the resistance towardprofenofos appeared to increase slowly from 3.0-foldin 2002 to 6.8-fold in 2004. For the KFR-strain, theresistance levels toward both organophosphates testedappeared to be similar, in general, to those in MNFstrain. Tests with carbamates showed that resistancelevels in both field strains (MNF and KFR) followeda consistent pattern. Resistance levels for methomylin both MNF strain (30.5-fold to 39.8-fold) and KFRstrain (22.9-fold to 37. I-fold) were greater than thosefor thiodicarb and carbaryl.
Tests with pyrethroids (Table 2) revealed higherresistance levels especially toward cypermethrin inboth MNF- and KFR-strains. Resistance to cypermethrinwas highly extended from 38.4-fold in 2002 to 98.4-fold in 2003 and reached to 55.2-fold in 2004. Similartrend of resistance pattern toward cypermethrin wasalso demonstrated in KFR strain, but the resistancelevels were much more higher than those in MNFstrain, at which increased from 38.4-fold in 2002 tol29.2-fold in 2003 and to 98.9-fold in 2004. Resistance levels to esfenvalerate during the season of20022004 ranged from l6.9-fold to 25.9-fold in MNFstrain, and from l4.9-fold to 19.3-fold in KFR strain.The lowest levels of resistance (7.4-fold to l1.3-fold)were reported toward fenpropathrin in both fieldstrains.
Data in Table 3 revealed high levels of resistanceto biopesticide, B. bassiana, in MNF strain with resistance ratios (RR) ranged from 41.9-fold in 2003 to58. l-fold in 2004, but KFR showed higher levels ofresistance at which RR ranged from 60.9-fold (2003)to 69.7-fold (2004). However, the two field populationsdid not exhibit any progression in tolerance towardthe other biopesticide tested, spinosad, through theseasons of 2003-2004, at which RR did not exceed0.5-fold. On the other hand, resistance monitoring forselected chitin synthesis inhibitors showed that resistance levels to hexaflumuron were increased slightlyin MNF strain from 4.4-fold to 8.2-fold, and from2.0-fold to 5.7-fold in KFR strain, in 2003 and 2004,respectively. However, resistance levels to flufenoxuronin both strains were considerably low at which didnot exceed 3.8-fold or 2.9-fold in MNF and KFRstrains.
Diagnostic concentration determination
Based on estimates LC99 and associated fiducial limits(Table 4) and visual examination of the concentration-response data, we selected a concentration of eachinsecticide tested for using as a diagnostic (or discriminating) concentration to determine resistance levelsin KFR field populations collected during the seasonsof 2003-2004. The diagnostic concentrations selectedagainst second-larval instars were: 228.1, 78.3, 327.2,
Insecticide resistance of Spodoptera littoralis 403
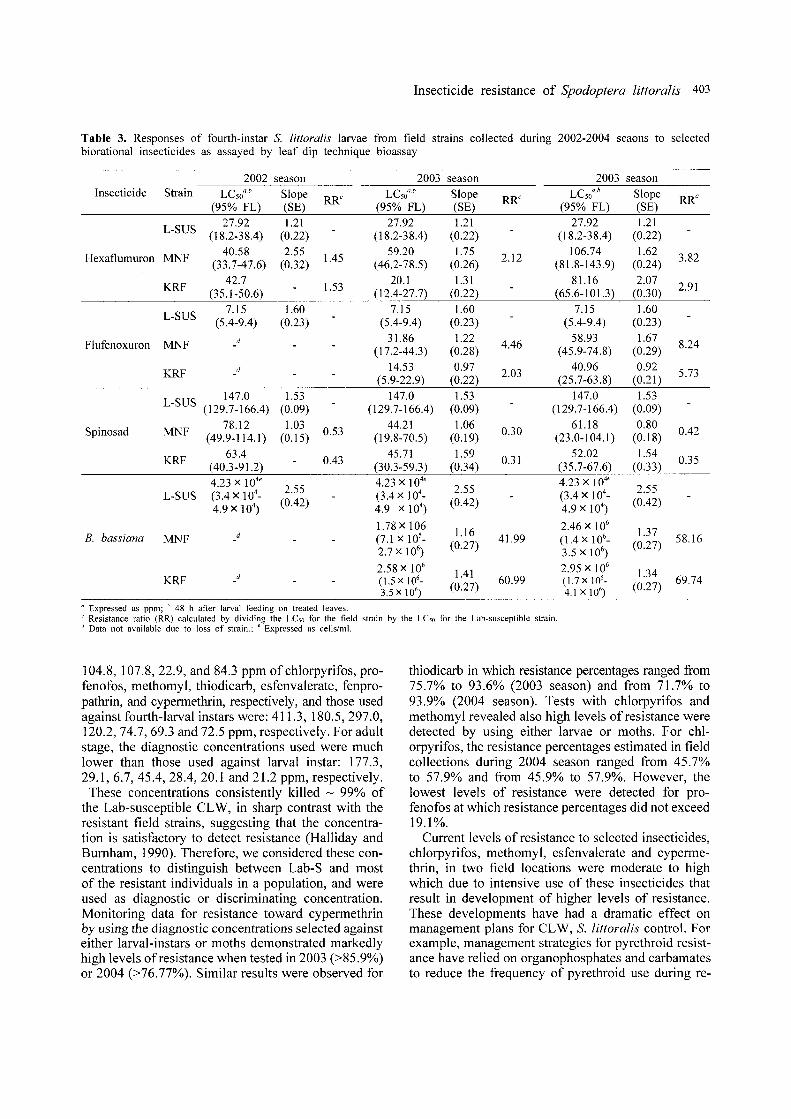
Table 3. Responses of fourth-instar S. littoralis larvae from field strains collected during 2002-2004 seaons to selectedbiorational insecticides as assayed by leaf dip technique bioassay
2002 season 2003 season 2003 seasonInsecticide Strain LCsoa,b Slope
RRc LCsO",b Slope RRcLC
50a,b Slope
RRc
(95% FL) (SE) (95% FL) (SE) (95% FL) (SE)
L-SUS 27.92 1.21 27.92 1.21 27.92 I.21(18.2-38.4) (0.22) (18.2-38.4) (0.22) (18.2-38.4) (0.22)
Hexaflumuron MNF40.58 2.55 1.45 59.20 1.75 2.12 106.74 1.62 3.82(33.7-47.6) (0.32) (46.2-78.5) (0.26) (81.8-143.9) (0.24)
KRF42.7 1.53 20.1 1.31 81.16 2.07 2.91(35.1-50.6) (12.4-27.7) (0.22) (65.6-101.3) (0.30)
L-SUS 7.15 1.60 7.15 1.60 7.15 1.60(5.4-9.4) (0.23) (5.4-9.4) (0.23) (5.4-9.4) (0.23)
Flufenoxuron MNF d 31.86 1.22 4.46 58.93 1.67 8.24(17.2-44.3) (0.28) (45.9-74.8) (0.29)
KRF d 14.53 0.97 2.03 40.96 0.92 5.73(5.9-22.9) (0.22) (25.7-63.8) (0.21)147.0 1.53 147.0 1.53 147.0 1.53
L-SUS (129.7-166.4) (0.09) (129.7-166.4) (0.09) (129.7-166.4) (0.09)
Spinosad MNF 78.12 1.03 0.53 44.21 1.06 0.30 61.18 0.80 0.42(49.9-114.1) (0.15) (19.8-70.5) (0.19) (23.0-104.1 ) (0.18)
KRF63.4 0.43 45.71 1.59 0.31 52.02 1.54 0.35(40.3-91.2) (30.3-59.3) (0.34) (35.7-67.6) (0.33)
4.23 x 104'
2.554.23 x 104e
2.554.23 x 104
'2.55
L-SUS (3.4 x 104_ (3.4 x 104_ (3.4 X 104
_
4.9 X 104)
(0.42) 4.9 x 104)
(0.42) 4.9 x 104)
(0.42)
1.78x 106 1.16 2.46 x 106
1.37B. bassiana MNF d (7.1 x 105_ 41.99 (1.4 x 106_ 58.162.7 x 106
)(0.27)
3.5 xI06)
(0.27)
2.58 x 106
1.41 2.95 x 106
1.34KRF d (1.5 xI06
- 60.99 (1.7x 106_ 69.743.5 x 106
)(0.27) 4.1 X 106
)(0.27)
" Expressed as ppm; 48 h after larval feeding on treated leaves.c Resistance ratio (RR) calculated by dividing the LC,o for the field strain by the LCso for the Lab-susceptible strain.J Data not available due to loss of strain.; e Expressed as cells/ml.
104.8,107.8,22.9, and 84.3 ppm ofchlorpyrifos, profenofos, methomyl, thiodicarb, esfenvalerate, fenpropathrin, and cypermethrin, respectively, and those usedagainst fourth-larval instars were: 411.3, 180.5,297.0,120.2,74.7,69.3 and 72.5 ppm, respectively. For adultstage, the diagnostic concentrations used were muchlower than those used against larval instar: 177.3,29.1,6.7,45.4,28.4,20.1 and 21.2 ppm, respectively.
These concentrations consistently killed ~ 99% ofthe Lab-susceptible CLW, in sharp contrast with theresistant field strains, suggesting that the concentration is satisfactory to detect resistance (Halliday andBurnham, 1990). Therefore, we considered these concentrations to distinguish between Lab-S and mostof the resistant individuals in a population, and wereused as diagnostic or discriminating concentration.Monitoring data for resistance toward cypermethrinby using the diagnostic concentrations selected againsteither larval-instars or moths demonstrated markedlyhigh levels ofresistance when tested in 2003 (>85.9%)or 2004 (>76.77%). Similar results were observed for
thiodicarb in which resistance percentages ranged from75.7% to 93.6% (2003 season) and from 71.7% to93.9% (2004 season). Tests with chlorpyrifos andmethomyl revealed also high levels of resistance weredetected by using either larvae or moths. For chlorpyrifos, the resistance percentages estimated in fieldcollections during 2004 season ranged from 45.7%to 57.9% and from 45.9% to 57.9%. However, thelowest levels of resistance were detected for profenofos at which resistance percentages did not exceed19.1%.
Current levels of resistance to selected insecticides,chlorpyrifos, methomyl, esfenvalerate and cypermethrin, in two field locations were moderate to highwhich due to intensive use of these insecticides thatresult in development of higher levels of resistance.These developments have had a dramatic effect onmanagement plans for CLW, S. littoralis control. Forexample, management strategies for pyrethroid resistance have relied on organophosphates and carbamatesto reduce the frequency of pyrethroid use during re-

404 J. Asia-Pacific Entomol. Vol. 8 (2005)
Table 4. Responses of S. littoralis larvae and adults from KFR- field strain (2003-2004) to different insecticides as assayedby using vial-discriminating concentration technique
Disciminating Resistance percentages (RPYInsecticide Stage concentration a.b 2003 season 2004 season
(95% FL) % MF RP % MF RP
2nd 228.1 (125.7-572.8) 99.0 0.0 80.7 18.52
Profenofos 4th 411.3 (213.3-792.2) 98.0 1.01 80.0 19.19
Adult 177.3 (68.1-640.3) 94.0 5.05 92.0 7.07
2nd 78.3 (46.5-204.4) 46.3 53.20 41.7 57.91
Chlorpyrifos 4th 180.5 (89.8-719.1) 56.0 43.34 53.5 45.96
Adult 29.1 (13.7-183.2) 52.0 47.47 46.0 53.54
2nd 327.2 (101.9-662.7) 53.0 46.50 52.7 46.80
Methomyl 4th 297.0 (104.2-947.3) 52.0 47.50 65.5 33.84
Adult 6.7 (1.90- 16.5) 36.0 63.64 30.0 69.70
2nd 104.8 (63.8-249.4) 6.3 93.60 6.0 93.94
Thiodicarb 4th 120.2 (67.4-365.7) 10.0 89.90 22.0 77.80
Adult 45.4 (19.8-85.4) 24.0 75.76 28.0 71.72
2nd 107.8 (46.1-644.7) 69.3 29.97 34.0 65.66
Esfenvalerate 4th 74.7 (40.16-235.20) 30.5 69.19 27.0 72.73
Adult 28.4 (8.96-68.7) 76.0 23.30 62.0 37.73
2nd 22.9 (13.6- 52.1) 64.7 34.68 60.7 38.72
Fenpropathrin 4th 69.3 (27.7-149.1) 65.5 33.84 82.5 16.67
Adult 20.1(8.94-143.4) 74..0 25.30 78.0 21.20
2nd 84.3 (38.4-400.1) 10.7 89.23 23.0 76.77
Cypermethrin 4th 72.5 (39.6-212.2) 7.0 92.93 17.5 82.32
Adult 21.2 (7.97-38.95) 14.0 85.90 20.0 79.80o Concentration of an insecticide that kills 99% of the susceptible strain. expressed as ppm.b 24 h-toxicity data.C Resistance percentage (RP)=I OO-(MF/MS X 100), where MF~% mortality at discriminating concentration in field-collected insects, and MS= % mortality(constantJ
at discriminating concentration in susceptible insects.
cent years (Graves et al., 1988; Plapp et al., 1990), soexposure of insect to these compounds has increased.The cotton insect management plan recommended byBaldwin and Graves (1991) for Louisiana seems tobe reliable to apply in Egypt. In essence, the windowstrategy, based on the use of a single class of insecticide on discrete generations of an insect pest is stilladvocated. However, the carbamates are recommendedduring the early season, pyrethroids are recommendedfrom 1 July to 15 August, and organophosphates arethen recommended during the late season. Based onour current study, we are in agreement with recommendations of Baldwin and Graves (1991) whichencourage the use ofbiological insecticides, especiallyduring the early season. All plans should be modifiedto stress strategies that involve conservation of allinsecticides used against the pest. Research on thefeasibility of using biological insecticides for controlof these pests should be emphasized. Our presentstudy further describes the expression of insecticideresistance in larvae and adults of S. littoralis by using
a rapid technique that uses a single diagnostic concentration of each test insecticide that can discriminatebetween susceptible and resistant individuals. Roushand Miller (1986) have shown the advantages ofusingdiagnostic concentrations rather than a comparison ofLCso estimates. Therefore, in this part we used a glassvial technique using larvae and adult stage to selectthe life stage that is more appropriate to use forresistance monitoring study. Our data on first instal'S(2nd- and 4th-larval instal'S) are important becausecontrol is usually directed at this stage. Adult dataare also reported because of their importance inmonitoring for resistance and similarity in resistancespectrum to neonates (Campanhola and Plapp, 1989).We have indicated that both larval and adult stagesfrom all different field collections showed similarresponses to most selected insecticides. We demonstrated also that both monitoring techniques, i.e. larvalbioassays (leaf dip technique) and larval (or adult)-discriminating concentration technique, indicate thepresence of resistance to selected insecticides in the
CLW. And, both adult and larval vial testing maybe appropriate methods to detect the resistance. Theglass vial technique is particularly useful whenresistance is related to target-site insensitivity ratherthan to increased metabolism, and is exposed in alldevelopmental stages (Kanga et al., 1995). Wood andBishop (1981) suggested that metabolic differencesmight account for the differential levels in larval andadult stages. Most insecticide resistance studies havebeen focused on each developmental stage of theinsect pest that is the target for chemical control. Butcomparative studies on the development of resistancein different developmental stages could provide aninsight into the resistance mechanisms involved andthe ontogeny and evolution of resistance (Tabashniket al., 1988).Also, the National Research Council (1986)reported that the rate of resistance could be loweredby direct treatment with insecticides against stages
Insecticide resistance of Spodoptera littoralis 405
such as the adults and early instars because later instarshave greater enzymatic activity. We also conclude thatthe similarity of resistance in adults and larval stages(2nd and 4th instars) provides the evidence that adultmonitoring is a useful tool in predicting the resistancein neonates, the life stage at which most control isaimed (Campanhola and Plapp, 1989).
Reversion of insecticide resistance in fieldpopulations
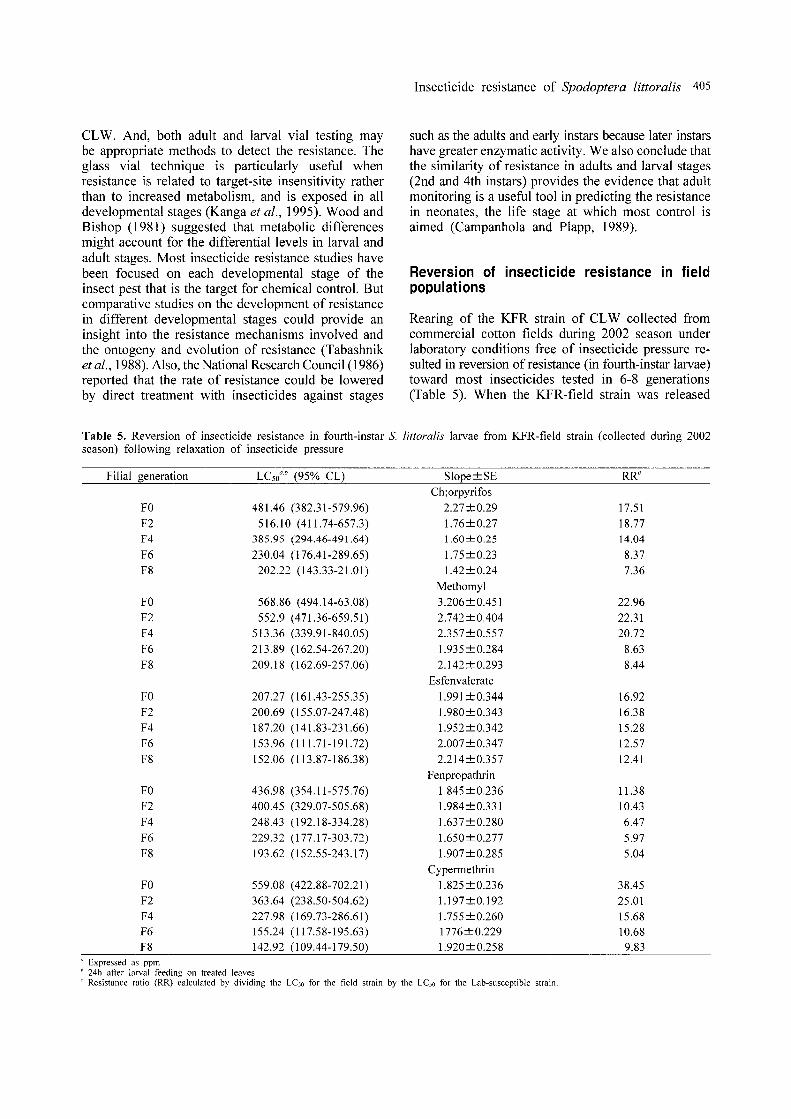
Rearing of the KFR strain of CLW collected fromcommercial cotton fields during 2002 season underlaboratory conditions free of insecticide pressure resulted in reversion of resistance (in fourth-instar larvae)toward most insecticides tested in 6-8 generations(Table 5). When the KFR-field strain was released
Table 5. Reversion of insecticide resistance in fourth-instar S. littoralis larvae from KFR-field strain (collected during 2002season) following relaxation of insecticide pressure
Filial generation LCsoa b (95% CL) Slope±SE RRc
Ch;orpyrifosFO 481.46 (382.31-579.96) 2.27±0.29 17.51F2 516.10 (411.74-657.3) l.76±0.27 18.77F4 385.95 (294.46-491.64) 1.60±0.25 14.04
F6 230.04 (176.41-289.65) l.75±0.23 8.37F8 202.22 (143.33-21.01) 1.42±0.24 7.36
MethomylFO 568.86 (494.14-63.08) 3.206±0.451 22.96F2 552.9 (471.36-659.51) 2.742±0.404 22.31F4 513.36 (339.91-840.05) 2.357±0.557 20.72F6 213.89 (162.54-267.20) 1.935±0.284 8.63F8 209.18 (162.69-257.06) 2.142±0.293 8.44
EsfenvalerateFO 207.27 (161.43-255.35) 1.991±0.344 16.92F2 200.69 (155.07-247.48) 1.980±0.343 16.38F4 187.20 (141.83-231.66) 1.952±0.342 15.28F6 153.96 (111.71-191.72) 2.007±0.347 12.57F8 152.06 (113.87-186.38) 2.214±0.357 12.41
FenpropathrinFO 436.98 (354.11-575.76) 1.845 ±0.236 11.38F2 400.45 (329.07-505.68) 1.984±0.331 10.43F4 248.43 (192.18-334.28) 1.637±0.280 6.47F6 229.32 (177.17-303.72) 1.650±0.277 5.97F8 193.62 (152.55-243.17) 1.907±0.285 5.04
CypermethrinFO 559.08 (422.88-702.21) 1.825±0.236 38.45F2 363.64 (238.50-504.62) 1.197±0.192 25.01F4 227.98 (169.73-286.61) 1.755±0.260 15.68F6 155.24 (117.58-195.63) 1776±0.229 10.68F8 142.92 (109.44-179.50) 1.920 ± 0.258 9.83
" Expressed as ppmb 24h after larval feeding on treated leaves .• Resistance ratio (RR) calculated by dividing the LCso for the field strain by the LCso for the Lab-susceptible strain.

406 1. Asia-Pacific Entomol. Vol. 8 (200S)
from selection pressures, they lost the resistance tocypermethrin quicker than other selected insecticidesin which RR decreased from 38.5-fold (parents) to9.8-fold after eight generations. Similar trend wasobserved for methomyl at which the RR valuesdecreased from 22.9-fold (parents) to 8.4-fold in eightgenerations. For esfenvalerate, the data showed thatthe KFR strain exhibited slow reversion of resistanceto this compound in eight generations at which RRvalues decreased gradually from 16.9-fold (parents)to 12.4-fold (Fs). Also, similar trend was observedwith chlorpyrifos tests at which the strain lost itsresistance to this compound gradually through eightgenerations where RR decreased from 17.5-fold (Fo)to 7.3-fold (Fg) .
It is obvious generally that the KFR field-strainused in this study appeared to be not homogenousfor resistance to five tested insecticides representingmajor classes of insecticides, i.e. organophosphates,carbamates and pyrethroids. Metcalf (1955) and Crow(1957) reported that the reversion of resistance inhouse fly, Musca domestica L. populations that arereturned conditions free of insecticides is a characteristic of partially resistant and not homozygouslyresistant strains. Sawicki et al. (1980) reported thateven highly resistant clones of the green peach aphid,Myzus persicae (Sulzer) lost resistance when selectionpressure was relaxed. Abedi and Brown (1980) attributedthe reversion to susceptibility after the initial phaseof selection of the yellow fever mosquito, Aedesaegypti (L.), to the lower biotic potential of theresistant individuals. They also postulated that onceresistance is stabilized, the RR individuals are nolonger at an advantage under insecticide-freeconditions.In similar study conducted by Osman et al. (1991),they found that rearing of two field strains (Mama,Yama) of pink bollworm, Pectinophora gosspiella(Saunders), from Arizona under conditions free ofinsecticides resulted in reversion of resistance in fourand five generations, respectively, to levels close tothat found in the susceptible laboratory strain, per-
methrin resistance in these field strains was unstableand was apparently in its early phase of development.Likely, our present study demonstrated that resistanceto the test insecticides in KFR-field strain was generallyunstable and reverted slowly to the tolerance level«IO-fold resistance) when reared for 6-8 generationsunder laboratory conditions without any insecticidalpressure. Therefore, our study is in agreement withthe findings of Osman et al. (1991) who suggestedthat monitoring of the pink bollworm resistance infield strains reared in the laboratory should be performed in the F I generation. On the other hand, ifthe rate of relaxation of resistance toward insecticidesis slow or stable and resistance does not revert toits initial level of susceptibility, the insecticide mustnot be reused (EI-Guindy, 1999). Where, the quickand fast rates of relaxation indicate that the insecticideto be repute again in practice is likely possible. Itshould be emphasized that if certain insecticides areto be reused due to their fast of relaxation, they mustnot be frequently applied against pests because it iswell known now that an old type of resistance thathas been relaxed due to cessation ofchemical pressurefor a certain period can always be reestablished morequickly with continuous selection in subsequentgenerations.
AChE and esterase activities
Insects from resistant field populations collected eitherfrom MNF (Fa, parents) or KFR (Fa, parents) cottonfields, in 2002 season, expressed higher levels ofAChE activity than the reference (Lab-susceptible)strain, in which AChE activity was 13.2- and 8.4-foldhigher than that from the Lab-susceptible strain,respectively (Table 6). However, AChE activity wassharply decreased following laboratory rearing ofbothMNF and KFR indicating that these field-collectedstrains appeared to loss their resistance in the absenceof insecticide pressure. The activity in F6 generation
Table 6. Levels of acetylcholinesterase (AChE) and esterase activities in laboratory-susceptible strain, field-resistant (Fo), andF6-reversion strains of S. littoralis
Esterases activity"Strain
Lab-Sus
Fo-MNF
F6-MNF
Fo-KFR
F6-KFR
AChE activity"
nmol/min/mg protein R/S (activity ratio)
8S.S ± 1.43d 1.00
1131.7±9.42a 13.24
86.3±3.17d 1.01
719.1 ±8.67b 8.41
163.0±3.4Sc 1.92
nmol/min/mg protein
6I.S±4.:5b
113.S±12.2a
94.0±4.7ab
117.7±8.7a
101.0±4.5a
R/S (activity ratio)
1.00
1.84
1.53
1.91
1.64
a AChE activity was measured in heads of CL W moths toward acetylcholine iodide and expressed as nmol/min/mg ptotein.b Esterases activity were measured in midgut homogenate from CLW larvae toward ex-naphthyl acetate (ex-NA) and expressed asnmol ex-naphthol/min/mg protein.
Values by the same letter within each vertical column are not significantly different (Turkey's P=O.05).

Insecticide resistance of Spodoptera Iittoralis 407
Fig. 1. Nondenaturating polyacrylmide gel electrophoresis ofthe -naphthyl acetate (NA)-hydrolyzing esterases from theLab-S, KFR- and MNF-resistant field strains (Fo) and F6reversion strains. Each lane was loaded with the same proteinamount, and the gel was stained for the enzyme activity usinga-NA as substrate.
susceptible strain and had five additional denselystaining (Fig. I). The results showed also that lossof insecticides resistance in KFR-field collection appeared to be correlated with disappearing of all additional bands of esterases that were document inFO-resistant field collection. The data also concludedthat biochemical studies indicated that the CLW resistant-field collections had a greatly increased AChE andesterase (toward a -NA) activities compared tosusceptible, the electrophoresis gels showing bandsthat were not present in the susceptible strain.Enhanced levels of esterase activities are associatedwith insecticide resistance in other lepidopteranspecies, e.g. tobacco budworm, Heliothis virescens(McCaffery, 1998). In previous studies, increasedesterase activities were measured in larvae that areresistant to OPs such as methyl parathion, profenofos,and azinophosmethyl (Goh et al., 1995, Harlod andOttea, 2000). Moreover, Goh et al. (1995) concludedthat substantially increased esterase activities (towardI-NA) might be involved in resistance in a carbamate,thiodicarb-resistant strain. Enhanced esterase activitiesalso confer pyrethroid resistance to H virescens (Graveset al., 1991; Goh et al., 1995). Detecting and monitoringinsecticide resistance and associated mechanisms ininsect populations are possible using biochemicalassays with model substrates (Brown and Brogdon,1987; Devonshire, 1987). High frequencies of OP,profenofos resistance were detected collected in larvalof H virescens from II field strains collected in
of MNF strain was almost similar to that in theLab-susceptible strain, whereas insects from MNFand KFR-F 6 generation showed somewhat an increasein AChE activity by 1.9-fold higher than that in Labsusceptible strain. Therefore, our data demonstratedthat loss of insecticide resistance in both two fieldcollected strains seems to be correlated with sharpdecrease in AChE activity, following laboratory rearingof these strains for six generations in the absence ofinsecticide pressure. This also may be particularlyimportant for understanding AChE insensitivity as aninsecticide resistance mechanism, which has beenreported in a number of insect species (Soderlund andBloomquist, 1990; Harold and Ottea, 1997).
Esterase activities toward a -naphthyl acetate (a-NA) in CLW larvae from both MNF- and KFRfield strain, were higher than those measured in Labsusceptible strain (Table 6). Similar esterase activities were reported in Fo-generation of both MNF andKFR strains, and increased by 1.84- and 1.9l-tirnes,respectively, than those in Lab-susceptible strain. Dataalso showed that esterase activities appeared to decreasefollowing laboratory rearing of both field strains.Where, after six generations (F6) under laboratoryrearing, in the absence of insecticide pressure, esteraseactivities measured in MNF and KFR strains wereslightly increased by 1.53- and 1.64-times, respectively,than those in Lab-susceptible strain.
Native electrophoresis PAGE of esterases
The results of the electrophoretic examination ofCLWlarval homogenates of Lab-susceptible, field-collected,and relaxed strains were presented in Fig. 1. GelsofLab-susceptible CLW stained for a -naphthyl acetatehydrolyzing activity showed three major esteraseactivity bands and three other indistinct bands. MNFfield strain (Fs) collected during 2002-late cottonseason, showing all the major bands detected in Labsusceptible strain, had six additional densely stainingbands, suggesting that increased l-naphthyl acetatehydrolysis was due to increased amounts of enzyme.Such enhanced levels of esterase activities seemedto be associated with insecticide resistance in CLW.
Our data showed also that loss of resistance in MNFstrain after relaxation by laboratory rearing, for sixgenerations, in the absence of insecticide pressure,resulted in disappearing all the additional esterasebands (except band found at R, = 0.570) which foundin F0-field strain (Fig. 1) indicating reversion of resistant field-strain toward the susceptibility. This confirmsour findings that the esterases have an important rolein insecticide resistance in CLW. The same trend wasobviously apparent for KFR field strain (Fo) possessing all major esterase bands which detected in Lab-
Ii:I
ILZ:E
cI
ILZ:E
Ii:I
a::IL~
Ii:I
a::IL~

408 J. Asia-Pacific Entomol. Vol. 8 (2005)
Louisiana, USA during 1995, and there was a strongcorrelation between profenofos resistance rations andesteraseactivities toward 1-NA(Harold and Ottea, 1997).Staining electrophoretic gels with model substrates(e.g. 1- and 2-NA) can be used to visualize quantitative and qualitative differences in esterases associatedwith resistance. In a previous study, Harold and Ottea(1997) demonstrated that one esterase was consistentlyover expressed and a second band under-expressedin OP resistant H virescens from both laboratoryselected and field-collected strains.
In conclusion, early detection of insecticide resistance provides a basis for the management of resistantpest populations (Brewer and Trumble, 1991), anddepends on development of sensitive test methods tomonitor resistance. Rapid biochemical assays coupledwith biological assays are potential tools for estimatingboth the intensity and the frequency of resistance inthe field (Brown and Brogdon, 1987; Devonshireet 01., 1986). For example, measurement of AChEsensitivity to inhibition by insecticides or resistance-associated esterases with naphthyl acetate estersprovides a rapid indication of resistance mechanismsexpressed in insecticide-resistant pests (Byrne andDevonshire, 1991; Brown et 01., 1996). However, useof non-insecticide substrates as monitoring tools isimprudent without establishing a relationship betweenenzyme activities with model substrates and insecticide susceptibility (Ibrahim and Ottea, 1995). Harlodand Ottea (1997) revealed that high frequencies ofresistance to profenofos were evident among the fieldpopulations tested and resistance was unstable in theabsence of insecticides. They added that resistanceto OP, profenofos was highly correlated with esteraseactivity (toward a -NA), suggesting an associationbetween these enzymes and profenofos resistance.Similar relationships have been also established inother insects between esterase activities and resistanceto OP (Devonshire and Moores, 1982) and pyrethroid(Riskallah et 01., 1983; Gunning et 01., 1996) insecticides,although positive correlations are not always found(Carlini et 01., 1995). Thus, the utility ofthis esteraseassay as a sensitive and rapid biochemical markerfor monitoring insecticide resistance in field populationsof CLW is being explored further.
Literature Cited
Abbott, W.S. 1925. A method for computing the effectivenessof an insecticide. J. Econ. Entomol. 18: 265-267.
Abdallah, M.D. 1991. A general view ofthe resistance problemof cotton pests in Egypt. Resistant-Pest-Management. 3:22-25.
Abedi, Z.H. and A.W.A. Brown. 1980. Development andreversion of DDT-resistance in Aedes aegypti. Can. J.
Genet. Cytol. 2: 252-261.Abo-Elghar, G.E., M.l-I. Rashwan, Z.A. EI-Bermawy, H.S.
Radwan and A.H. Hussein. 1992. Monitoring for resistancein the cotton leafworm, Spodoptera littoralis (Boisd.),against insecticide mixtures with benzoylphenyl ureas.Bull. Entomol. Soc. Egypt, Eco. Ser. 19: 249-259.
Armstrong, K.F. and D.M. Suckling. 1990. Correlation andinheritance of azinphos-methyl resistance in the lightbrown apple moth (Lepidoptera: Tortricidae). Pestic.Biochem. Physiol. 36: 281-289.
Avidov, Z. and 1. Harpaz, 1969. Plant pests of Israel.Universities Press, Jerusalem. 549 p.
Baldwin, J.L. and J.B. Graves. 1991. Cotton insect pestmanagement. LA. Coop. Ext. Servo Bull. 1829.
Bradford, M.M. 1976. A rapid and sensitive method for thequantification of microgram quantities of protein utilizingthe principle of protein-dye binding. Anal. Biochem. 62:294- 254.
Brewer, MJ. and J.T. Trumble. 1989. Field monitoring forinsecticide resistance in beet armyworm (Lepidoptera:Noctuidae). 1. Ecan. Entomol. 82: 1520-1526.
Brewer, M.l. and J.T. Trumble. 1991. Classifying resistanceseverity in field populations. Sampling inspection plansfor an insecticide resistance monitoring program. J. Econ.Entomol. 84: 379.
Brown, TM. and W.G. Brogdon. 1987. Improved detectionof insecticide resistance through conventional and molecular techniques. Annu. Rev. Entomol. 32: 145-162.
Brown, E.S. and C.F. Dewhurst. 1975. The genus Spodoptera(Lepidoptera, Noctuidae) in Africa and the Near East. Bull.Entomol. Res. 65: 221-262.
Brown TM., P.K. Bryson, F. Arnette, M. Roof, l.L B. Mallett,J.B. Graves and S..r. Nemec. 1996. Surveillance of resistantacetylcholinesterase in Heliothis virescens. pp.149-15 7, inMolecular genetics and evolution of pesticide resistance,Ed. TM. Brown. Washington, D.C. American ChemicalSociety Books.
Bush, M.R., Y.A.I. Abdel-All, K. Saito and G.e. Rock. 1993.Azinphosmethyl resistance in the tufted apple bud moth(Lepidoptera: Tortricidae): reversion, diagnostic concentrations, associated esterases, and glutathione transferases.J. Econ. Entomol. 86: 213-225.
Byrne, F.J. and A.L. Devonshire. 1991. In vivo inhibition ofesterase and acetylcholinesterase activities by profenofos
. treatments in the tobacco whitefly, Bemisia tabaci (Genn.):Implications for routine biochemical monitoring of theseenzymes. Pestic. Biochem. Physiol. 40: 198-204.
Campanhola, e. and F.W. Plapp. 1989. Pyrethroid resistancein the tobacco budworm (Lepidoptera: Nocutidae): insecticide bioassays and field monitoring. J. Econ. Entomol.82: 22-28.
Carlini, EJ., B.A. Mcl'heron, C.M. Felland and L.A. Hull.1991. Elevated esterase activity in resistant tufted applebud moth, Platynota idaeusalis (Walker) (Lepidoptera:Tortricidae). Compo Biochem. Physiol. C99: 375-377.
Carlini, EJ., B.A. Mcl'heron, C.M. Felland and L.A. Hull.1995.Biochemical mechanismsofazinphosmethyl resistancein the tufted apple bud moth Platynota idaeusalis. Pestic.Biochem. Physiol. 51: 38-47.
Crow, J.F. 1957. Genetics of insect resistance to chemicals.Annu. Rev. Entomol. 2: 252-261.
Devonshire, A.L. 1987. Biochemical studies of organopgosphorus and carbamate resistance in house flies and aphids.Combating Resistance to Xenobiotics: Biological andChemical Approaches (Eds., M. G. Ford et al.). Weinheim,Federal Republic of Germany. pp. 239-355.

Devonshire, A.L. and G.D. Moores. 1982. A carboxyleserasewith broad substrate specificity causes organophosphorous,carbamate and pyrethroid resistance in peach-potato aphids(Myzus persicae). Pestic. Biochem. Physiol. 30: 235-246.
Devonshire, A.L., G.D. Moores and RH. ffernch-Constant.1986. Detection of insecticide resistance by immunologicalestimation of carboxylesterase activity in Myzus persicae(Sulzer) and cross reaction of the antiserum with Phordanhumuli (Schrank) (Hemiptera: Aphididae). Bull. Entomol.Res. 76: 97-107.
Eldefrawi, M.E., A. Tappozada, N. Mansouer and M. Zied.1964. Toxicological studies on the Egyptian cotton leafworm, Prodenia litura: susceptibility of different larvalinstar of P. litura to insecticides. J. Econ. Entomol. 57:591-593.
EI-Guindy, M.A. 1999. Pesticide resistance management andoptimization of integrated pest management programs inEgyptian cotton. Adv. Agric. Res. Egypt. 1: 1-52.
EI-Guindy, M.A., A.A. EI-Refai and M.M. Abd-Elsattar. 1983.The pattern of cross resistance to insecticides and juvenilehormone analogues in a diflubenzuron-resistant strain ofthe cotton leafworm, Spodoptera littoralis (Boisd.). Pestic.Sci. 14: 235.
EI-Guindy, M.A., M.E. Keddis, M.M. Abd-Elsattar and Y.F.Ghoneim. 1989. Status of resistance in cotton leafwormSpodoptera littoralis (Boisd.) under the present Egyptiancotton pest control programme. Proc. 1st Int. Conf. Econ.Entomol. Vol. II: 453- 462.
Ellman, G.L., K.D. Coutney, V. Andres and R.M. Featherstone.1961. A new and rapid colorimetric determination ofacetylcholinesterase activity. Biochem. Pharm. 7: 88-95.
ffrench-Constant, R.H. and R.T. Roush. 1990. Resistance detection and documentation: the relative roles of pesticidaland biochemical assays. pp. 4-38, in Pesticide resistancein arthropods. Eds. R.T. Roush and B.E. Tabashnik.Chapman and Hall, New York.
Finney, 0.1. 1971. Probit Analysis, 3rd ed. Cambridge University press, Cambridge University, Press, London, UK.
Goh, D.K.S., D.D. Anspaugh, N. Motoyama, G.C. Rockand RM. Roe. 1995. Isolation and characterization of aninsecticide-resistance associated esterase in the tobaccobudworm, Heliothis virescens (F.). Pestic. Biochem.Physiol. 51: 192-204.
Graves, J.B., B.R. Leonard, A.M. Pavloff, G. Burris, K.Ratchford and S. Micinski. 1988. Monitoring pyrethroidresistance in tobacco budworm in Louisiana during 1987:Resistance management implications. J. Agric. Entomol.5: 109-115.
Graves, J.B., B.R. Leonard, S. Micinski and S.H. Martin. 1991.Situation on tobacco budworm budworm resistance topyrethroids in Louisiana during 1991. Proc. Beltwide CottonProd. Res. Conf. Memphis, Tenn.: National Cotton Councilof America. 2: 743-746.
Gunning, R.V., G.D. Moores and A.L. Devonshire. 1996. Esterases and esfervalerate resistance in Australian Helicoverpaarmigera (Hubner) (Lepidoptera: Noctuidae). Pestic. Biochem.Physiol. 54: 12-23.
Halliday, W.R. and K.P. Burnham. 1990. Choosing the optimaldiagnostic dose for monitoring insecticide resistance. J.Econ. Entomol. 83: 1151-1159.
Harold, J.A. and J. A. Ottea. 1997. Toxicological significanceof enzyme activities in profenofos-resistant tobacco budworm Heliothis virescens (F.). Pestic. Biochem. Physiol.58: 23-33.
Harold, J.A. and J.A. Ottea. 2000. Characterization of esterasesassociated with profenofos resistance in the tobacco
Insecticide resistance of Spodoptera littoralis 409
budworm, Heliothis virescens (F.). Arch. Insect Biochem.Physiol. 45: 47-59.
Ibrahim, S.A. and J.A. Ottea. 1995. Biochemical and toxicological studies with laboratory and field populations ofHeliothis virescens (F.), Pestic. Biochem. Physiol. 53:116-128.
Issa, Y.H., M.E. Keddis, F.A. Ayad, M.M. Abd-Elsattar andM.A. El-Guindy. 1986. Survey of resistance to organophosphorous insecticides in field strains of the cottonleafworm Spodoptera littoralis (Boisd.) during 1980-1984cotton growing seasons. Bull. Entomol. Soc. Egypt Econ.Ser. 14: 399-404.
Kanga L.H. and F.W. Plapp, Jr. 1992. Development of a glassvial technique for monitoring resistance to organophosphate and carbamate insecticides in the tobacco budwormand boll weevil, pp. 731-734. in Proc. Beltwide CottonProd. and Research Conf., National Cotton Council,Memphis, TN.
Kanga L.H., F.W. Plapp, Jr., G.W. Elzen, M.L. Wall and J.D.Lopez, Jr. 1995. Monitoring for resistance to organophosphorus, carbamate and cyclodiene insecticides in tobaccobudworm adults (Lepidoptera:Noctuidae). J. Econ. Entomol.88: 1144-1149.
Magaro, J.1. and J.V. Edelson. 1990. Diamondback moth(Lepidoptera: Plutellidae) in south Texas: a technique forresistance monitoring in the field. J. Econ. Entomol. 83:1201-1206.
McCaffery, A.R. 1998. Resistance to insecticides in HeliothineLepidoptera: A global view. Phil. Trans. R Soc. 8. 353:1735-1750.
McCutchen, B.F., P.W. Plapp, Jr., S.1. Nemec and C.Campanho1a. 1989. Development ofdiagnostic monitoringtechniques for larvae pyrethroid resistance in Heliothisspp. (Lepidoptera: Noctuidae) in cotton. J. Econ. Entomol.82: 1502-1507.
Metcalf, R.L. 1955. Physiological basis for insect resistanceto insecticides. Physiol. Rev. 35: 197-232.
Miyata, T. 1986. Detection and monitoring methods for resistance in arthropods based on biochemical characteristics.pp. 99-116, in Pest resistance to pesticides. Eds. G.P.Georghiou and T. Saito. Plenum, New York.
National Research Council. 1986. Pesticide resistance strategies and tactics for management. National Academy ofSciences Press, Washington, D.C.
Osman, A.A., T.F. Watson and S. Sivasupramaniam. 1991.Reversion of permethrin in field strains and selection forazinphosmethyl and permethrin resistance in pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 84:353-357.
Pasteur, N. and G.P. Georghiou. 1981. Filter paper test forrapid determination of phenotypes with high esteraseactivity in organophosphate resistant mosquito. Mosq.News 41: 181-183.
Plapp, F.W., Jr., G.M. McWhorter and W.H. Vance. 1987.Monitoring for pyrethroid resistance in the tobacco budworm in Texas-1986. pp. 324-326, in Proceedings ofbeltwide cotton production research conferences. NationalCotton Council of America, Memphis, TN.
Plapp, F.W., Jr., C. Campanhola, R.D. Bagwell and B.F.McCutchen. 1990. Management of pyrethroid-resistanttobacco budworm on cotton in the United States. pp.237-260, in Pesticide resistance in arthropods. Eds. R.T.Roush and B.E. Tabashnik. Chapman & Hall, New York.
Prabhaker, N., N.C. Toscano, T.J. Hennebrry, S.l. Castle andD. Weddle. 1996. Assessment of two bioassay techniquesfor resistance monitoring of silverleaf whitefly (Homoptera:

410 1. Asia-Pacific Entomol. Vol. 8 (2005)
Aleyrodidae) in California. 1. Econ. Entomol. 89: 805-815.Rashwan, M.B., Z.A. EI-Bermawy, A.A. El-Shiekh and H.S.A.
Radwan. 1992. The onset of organophosphates and carbamates resistance among lower Egypt population of thecotton leafworm, Spodoptera littoralis (Boisd.). Bull. Ent.Soc. Egypt, Econ. Ser. 19: 211-220.
Riskallah, M.R., S.F. Abd-Elghafar, M.R. Abo-Elghar andM.E. Nassar. 1983. Development of resistance and crossresistance in fenvalerate and deltamethrin selected strainsof Spodoptera littoralis (Boisd.). Pestic. Sci. 14: 508-512.
Robertson, J., K.C. Smith, N.E. Savin and RJ. Lavigne. 1984.Effects of dose selection and sample size on precisionof lethal dose estimates in dose-mortality regression. J.Econ. Entomol. 77: 833-837.
Roush, R.T. and RG. Luttrell. 1987. The phenotypic. expressionof pyrethroid resistance in Heliothis and. implications forresistance management,pp. 220-224. in Proceedings,Dallas,National Cotton Council of America, Memphis, TN.
Roush, R.T. and G.L. Miller. 1986. Considerations for designof insecticide resistance monitoring programs. J. Econ.Entomol. 79: 293-298.
Sawicki, R.M. 1986. Resistance to synthetic pyrethroids canbe countered successfully. Agribusiness-Worldwide. 8:22-25.
Sawicki, RM. and 1. Denholm. 1987. Management of resistanceto pesticides in cotton pests. Tropical Pest Management33: 262-272.
Sawicki, R.M., A.L. Devonshire, R.W. Payne and S.M.Petzing. 1980. Stability of insecticide resistant in the
peach-photo aphid, Myzus persicae (Sulzer). Pestic. Sci.11: 33-42.
Soderlund, D.M. and J.R. Blopmquist. 1990. Molecular Mechanisms of insecticide resistance. pp. 58-96, in Pesticideresistance in arthropods. Eds. R.T. Roush and B.E.Tabashnik. Chapman and Hall, New York.
Sparks, T.e. 1981. Development of insecticide resistance inHeliothis zea and Heliothis virescens in North America.Bull. Entomol. Soc. Am. 27: 186-192.
Staetz, C.A. 1985. Susceptibility of Heliothis virescens (p.)(Lepidoptera: Noctuidae) to permethrin from across thecotton belt: a five-year study. J. Econ. Entomol. 78:505-510.
Tabashnik, B.E., M.D. Rethwisch and M.W. Johnson. 1988.Variation in adult mortality and knockdown caused byinsecticides among populations of diamondback moth(Lepidoptera: Plutellidae). J. Econ. Entomol. 81 101-105.
Temerak S.A. 2002. Historical records of field cotton leafworm(Spodoptera littoralis) resistance to conventional insecticidesas influenced by the resistance programs in Egypt.Resistant Pest Management Newslett. 12: 7-10.
van Asperen, K. 1962. A study of housefly esterase by meansof a sensitive colorimetric method. J. Insect Physiol. 5:401-416.
Williams, D.F. and n.r. Reisford. 1964. Disc electrophoresisin poly-aery lamide: gels: extension to new conditions ofpH and buffer. Ann. N.Y. Acad. Sci. 121: 373-381.
Wood, R. and 1. Bishop. 1981. Insecticide resistance: populations and evolution. in The genetic basis of man-madechange. Eds. lA. Bishop and L.M. Cook. Academic Press.
Copyright © 2022 FDOKUMEN




















