
Monitoring and action threshold determination for the obscure mealybug Pseudococcus viburni...
6
Monitoring and action threshold determination for the obscure mealybug Pseudococcus viburni (Signoret) (Hemiptera: Pseudococcidae) using pheromone-baited traps Pride Mudavanhu * , Pia Addison, L. Pringle Ken University of Stellenbosch, Department of Conservation Ecology & Entomology, Faculty of AgriSciences, Private Bag X1, Matieland 7602, South Africa article info Article history: Received 12 November 2010 Received in revised form 10 February 2011 Accepted 11 February 2011 Keywords: Insect pest monitoring Pheromone-baited traps Visual sampling Correlation Mealybug Pome fruit abstract The absence of effective pest monitoring has resulted in an increase in populations of the obscure mealybug Pseudococcus viburni (Signoret) (Hemiptera: Pseudococcidae) in pome fruit orchards in the Western Cape Province of South Africa recently. The sex pheromone for P. viburni was recently identified and synthesized. Flight activity of adult male P. viburni was monitored by placing and servicing three, evenly spaced phero- mone-baited traps per orchard in three pome fruit growing regions used for the study. Fortnightly fruit sampling was done by randomly picking three fruits per tree for the duration of each fruit season, dissecting and noting mealybug infestation. The biweekly male P. viburni trap catch information and fruit infestation data collected over two seasons were correlated. There was a positive and significant relationship between the fruit infestation and number of P. viburni adult males caught in pheromone-baited traps (r 2 ¼ 0.454, P < 0.001). The action threshold level was estimated to be 2.5 male P. viburni caught per trap per fortnight at an economic threshold of 2% fruit infestation. This monitoring method was less labor intensive, more accurate and quicker than the current visual sampling and monitoring techniques. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction In South Africa, Pseudococcus viburni is being controlled primarily with insecticides (Myburgh et al., 1975; Swart, 1977; Van der Merwe, 2000). Despite concerted efforts to keep this pest from reaching damaging levels, increases in mealybug infestations have been ascribed to the inadequate, improper and incomplete use of these pesticides. Geiger and Daane (2001) and Walton (2003) argued that the challenges facing chemical control of mealybug range from poor insecticide coverage under dense leaf cover or bark, where this pest often resides, to insecticide run off over the mealybug’s waxy body secretion (Walton et al., 2004). For these reasons, and in line with the principles of sustainable integrated pest management, the use of insecticides must be exercised only when needed and limited to target applications against the most vulnerable life stage. The importance of timely applications of insecticides necessi- tates the use of a species-specific monitoring programme that can quickly determine pest presence and density (Walton et al., 2004). Visual sampling of mealybug is a laborious and time consuming process while, examination of trees in orchards has led to assess- ments which do not accurately reflect its pest status (Myburgh et al., 1975). Inspection of culled fruit in packhouses has also had its challenges. Not all infestations are detected during the sampling process which has led to under-estimation of the severity of infestation potential to be expected during ensuing seasons. P. viburni has a typical clumped distribution and cryptic habit during much of the year, similar to that of a closely related species, the vine mealybug (Planococcus ficus) (Signoret) (Walton et al., 2004). This renders visual monitoring methods ineffective since late in the summer, when mealybugs occur in high densities, most damage will have already been done. Mealybug damage is secondary in nature in that they go into fruit calyx where they produce honeydew on which black sooty mould grows thereby making the fruit unsightly. Furthermore their presence is also of phytosanitary concern in some countries. Walton et al. (2004) and Brown and Pringle (2006) stated that an effective monitoring system will provide information early in the season and at low mealybug densities in order to target control actions and appropriately schedule insecticide applications. These actions may subsequently help to prevent infestations of fruit, limit insecticide residues as well as prevent outbreaks in ensuing seasons. Trapping consists of some of the most vital sampling methods for insect surveys (Pedigo, 1999). Monitoring programs * Corresponding author. Tel.: þ27 83 315 5009; fax: þ27 21 808 2546. E-mail addresses: [email protected], [email protected] (P. Mudavanhu). Contents lists available at ScienceDirect Crop Protection journal homepage: www.elsevier.com/locate/cropro 0261-2194/$ e see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.cropro.2011.02.034 Crop Protection 30 (2011) 919e924
Transcript of Monitoring and action threshold determination for the obscure mealybug Pseudococcus viburni...

lable at ScienceDirect
Crop Protection 30 (2011) 919e924
Contents lists avai
Crop Protection
journal homepage: www.elsevier .com/locate/cropro
Monitoring and action threshold determination for the obscure mealybugPseudococcus viburni (Signoret) (Hemiptera: Pseudococcidae) usingpheromone-baited traps
Pride Mudavanhu*, Pia Addison, L. Pringle KenUniversity of Stellenbosch, Department of Conservation Ecology & Entomology, Faculty of AgriSciences, Private Bag X1, Matieland 7602, South Africa
a r t i c l e i n f o
Article history:Received 12 November 2010Received in revised form10 February 2011Accepted 11 February 2011
Keywords:Insect pest monitoringPheromone-baited trapsVisual samplingCorrelationMealybugPome fruit
* Corresponding author. Tel.: þ27 83 315 5009; faxE-mail addresses: [email protected]
(P. Mudavanhu).
0261-2194/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.cropro.2011.02.034
a b s t r a c t
The absence of effective pest monitoring has resulted in an increase in populations of the obscure mealybugPseudococcus viburni (Signoret) (Hemiptera: Pseudococcidae) in pome fruit orchards in the Western CapeProvince of South Africa recently. The sex pheromone for P. viburniwas recently identified and synthesized.Flight activity of adult male P. viburni was monitored by placing and servicing three, evenly spaced phero-mone-baited traps per orchard in three pome fruit growing regions used for the study. Fortnightly fruitsampling was done by randomly picking three fruits per tree for the duration of each fruit season, dissectingand noting mealybug infestation. The biweekly male P. viburni trap catch information and fruit infestationdata collected over two seasons were correlated. There was a positive and significant relationship betweenthe fruit infestation and number of P. viburni adult males caught in pheromone-baited traps (r2 ¼ 0.454,P< 0.001). The action threshold level was estimated to be 2.5 male P. viburni caught per trap per fortnight ataneconomic thresholdof 2% fruit infestation. Thismonitoringmethodwas less labor intensive,more accurateand quicker than the current visual sampling and monitoring techniques.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
In South Africa, Pseudococcus viburni is being controlled primarilywith insecticides (Myburgh et al., 1975; Swart,1977; Van derMerwe,2000). Despite concerted efforts to keep this pest from reachingdamaging levels, increases in mealybug infestations have beenascribed to the inadequate, improper and incomplete use of thesepesticides. Geiger and Daane (2001) and Walton (2003) argued thatthe challenges facing chemical control of mealybug range from poorinsecticide coverage under dense leaf cover or bark, where this pestoften resides, to insecticide run off over the mealybug’s waxy bodysecretion (Walton et al., 2004). For these reasons, and in linewith theprinciples of sustainable integrated pest management, the use ofinsecticides must be exercised only when needed and limited totarget applications against the most vulnerable life stage.
The importance of timely applications of insecticides necessi-tates the use of a species-specific monitoring programme that canquickly determine pest presence and density (Walton et al., 2004).Visual sampling of mealybug is a laborious and time consuming
: þ27 21 808 2546.k, [email protected]
All rights reserved.
process while, examination of trees in orchards has led to assess-ments which do not accurately reflect its pest status (Myburghet al., 1975). Inspection of culled fruit in packhouses has also hadits challenges. Not all infestations are detected during the samplingprocess which has led to under-estimation of the severity ofinfestation potential to be expected during ensuing seasons.P. viburni has a typical clumped distribution and cryptic habitduring much of the year, similar to that of a closely related species,the vine mealybug (Planococcus ficus) (Signoret) (Walton et al.,2004). This renders visual monitoring methods ineffective sincelate in the summer, when mealybugs occur in high densities, mostdamage will have already been done. Mealybug damage issecondary in nature in that they go into fruit calyx where theyproduce honeydew on which black sooty mould grows therebymaking the fruit unsightly. Furthermore their presence is also ofphytosanitary concern in some countries.
Walton et al. (2004) and Brown and Pringle (2006) stated thatan effective monitoring system will provide information early inthe season and at lowmealybug densities in order to target controlactions and appropriately schedule insecticide applications. Theseactions may subsequently help to prevent infestations of fruit, limitinsecticide residues as well as prevent outbreaks in ensuingseasons. Trapping consists of some of the most vital samplingmethods for insect surveys (Pedigo, 1999). Monitoring programs

Fig. 1. Weighted regression of Logit (z) on Ln (n) where z is the proportion of infestedfruit and n¼ number of male Pseudococcus viburni/trap/fortnight.
P. Mudavanhu et al. / Crop Protection 30 (2011) 919e924920
based on the use of pheromone-baited traps offer a convenientmonitoring method. This monitoring tool may also be extremelyuseful in helping producers determine whether or not their cropshould be considered for export to countries inwhich P. viburni is ofphytosanitary concern. The sex pheromone of the obscuremealybug P. viburni has recently been identified and synthesized(Millar et al. 2005). Zaviezo et al. (2007) described its field use.According to Romoser and Stoffolano (1998) pest damage can berelated to the number of insects caught in a trap and managementdecisions can be made accordingly. The abundance of P. ficus malescaught on sticky pheromone traps was correlated with steminfestations (Walton et al., 2004) making it possible to use trapcatch information to predict when stem inspections shouldcommence (de Villiers and Pringle, 2007). Increasing malemealybug trap counts correlated well with increasing crop infes-tation levels according to findings for P. ficus in vineyards in Cal-ifornia and South Africa (Walton et al., 2004).
The objective of this study was to investigate the relationshipbetween P. viburni males caught in sticky pheromone-baited trapsand fruit infestation levels as recorded by visual sampling methods.We also aim to establish an action threshold based on pheromone-baited trap counts, which corresponds to the 2% fruit infestationeconomic threshold proposed by Swart (1977), which is based onorchard cull analysis. This will in future help producers make moreinformed decisions regarding fruit destined for various markets.
2. Materials and methods
2.1. Sites
Two orchards per site, each approximately 1 ha in area wereinspected fortnightly in each of three different pome fruit growing
Table 1Between-region average (�SEM) of male trap counts and percent fruit infestation for 2numbers in brackets).
Orchard region Season 2007-08
Trap counts % Fruit infest
S’bosch 1(TF) 45.7 � 4.25 a 1.61 � 0.99 aS’bosch 2 (TF&P) 30.1 � 4.14 a 3.1 � 0.97 aElgin 1 (EW48) 8.5 � 4.14 b 0.42 � 0.97 aElgin 2 (EG9) 10.7 � 4.14 b 1.81 � 0.97 aCeres 1 (CA) 0.9 � 4.14 b 0.28 � 0.97 aCeres 2 (CP) 0.4 � 4.14 b 0.28 � 0.97 a
*Within each column, means followed by different letters are significantly different (Tu*TF¼ Orchard TF (Forelle pears), TF&P¼ Orchard TF&P (Forelle and Packham pears), EW4CA ¼ Orchard CA (Royal Gala apples), CP ¼ Orchard CP (Beurre Hardy pears).
areas in the Western Cape Province during the period November2007 to June 2009. The three sites were Elgin (34.16S, 19.05E,Elevation: 312 m) (Oak Valley Farm: Granny Smith apples plantedin both orchards), Ceres (33.34S, 19.57E, Elevation: 1046 m)(Lakenvlei Farm: Royal Gala apples and Beurre Hardy pears) andStellenbosch (33.90S, 18.86E, Elevation: 183 m) (Timberlea Farm:Forelle pears and a mixture of Forelle and Packham pears). In eachorchard block six evenly spaced rows with six trees per row (36trees in total per experimental block) were selected for sampling.The same trees were sampled for the duration of this study. Theorchard blocks were at least 100m away from each other in all sites.
2.2. Monitoring of male P. viburni using traps
The flight activity of adult male P. viburni was monitored byplacing and servicing three, evenly spaced pheromone-baited trapsin each of the orchards in all three pome fruit growing regions usedfor the study. Monitoring was also done fortnightly fromNovember2007 to June 2009.
P. viburni pheromone lures consisting of grey rubber septaloaded with 0.1 mg dose of racemic synthetic pheromone in hexane(Millar et al., 2005) were placed ontowhite sticky pads with a blackgrid-lined surface (total sticky surface area ¼ 320 cm2). The pher-omone lure was obtained directly from Jocelyn G. Millar, Universityof California Riverside. The lure and sticky pad were placed inyellow delta traps (Chempack�, Simondium Paarl, South Africa) andhung in the tree canopy at head height. The traps were at least 50mapart in a diagonal orientation, running across each orchard (twoon opposite edges and one in the centre). The sticky pads andpheromone lures were checked and replaced on each field visitevery twoweeks. All adult male P. viburni caught on the sticky trapswere counted in the laboratory using a stereo microscope.
2.3. Correlation between trap counts and fruit infestation
The abundance of male P. viburni captured on pheromone-bai-ted traps was correlated with data obtained from fruit infestationassessments, as determined by visual monitoring. The visualmonitoringmethod usedwas based onmethodologies described byVan der Merwe (2000) and Walton (Unpublished research report:Deciduous Fruit Producers Trust, 2006). Three fruits per tree wererandomly picked and labelled from each of 36 trees per trial blockdescribed above every fortnight from November until harvest.Fruits were taken to the laboratory, dissected to expose the calyxand ovary. Any presence of live or dead mealybugs, ovisacs, waxfilaments, sooty mold and/or honeydew on the surface, stem end,calyx or ovary of the fruit was noted as mealybug infestation andabsence was noted as un-infested (Van der Merwe, 2000).
007e08 and 2008e09 growing seasons for three pome fruit growing areas (Block
Season 2008-09
ation Trap counts % Fruit infestation
45.2 � 3.86 a 2.86 � 0.91 a26.15 � 3.95 b 2.95 � 0.93 a
6.5 � 3.95 c 1.09 � 0.93 a2.6 � 3.95 c 0.46 � 0.93 a0.8 � 3.95 c 0.21 � 0.93 a0.2 � 3.95 c 0.25 � 0.93 a
key HSD test, P < 0.05).8¼ OrchardW48 (Granny Smith apples), EG9¼ Orchard EG9 (Granny Smith apples),

Table 2Between-season differences in male trap counts and percent fruit infestation withinthree pome fruit growing regions during the 2007e2008 and 2008e2009 growingseasons.
Orchard region Pheromone trapcounts (d.f. ¼ 1, 40)
% Fruit infestation(d.f. ¼ 1, 40)
S’bosch 1(TF) F ¼ 0.0018, P ¼ 0.966 F ¼ 0.3191, P ¼ 0.575S’bosch 2 (TF&P) F ¼ 0.3803, P ¼ 0.541 F ¼ 0.0048, P ¼ 0.945Elgin 1 (EW48) F ¼ 1.1467, P ¼ 0.291 F ¼ 1.3667, P ¼ 0.249Elgin 2 (EG9) F ¼ 12.818, P ¼ 0.00092 F ¼ 3.6042, P ¼ 0.065Ceres 1 (CA) F ¼ 0.0497, P ¼ 0.825 F ¼ 0.1105, P ¼ 0.741Ceres 2 (CP) F ¼ 1.3160, P ¼ 0.258 F ¼ 0.0168, P ¼ 0.898
P. Mudavanhu et al. / Crop Protection 30 (2011) 919e924 921
2.4. Data analysis
The pheromone-baited trap data from the three traps in eachorchard for each sampling date were averaged and transformed tonatural logs. The fruit infestation data were transformed toempirical logits and regressed on the natural log of the trap countsusing a weighted regression with the reciprocal of the variance ofthe empirical logits as the weighting co-efficient (Cox, 1970):
Zj ¼ ln
Rj þ 1=2
nj � Rj þ 1=2
!; j ¼ 1;2;3.N samples; (1)
and,
Fig. 2. Adult male Pseudococcus viburni trap counts (left e hand scale) and percent fruit infes1 (a) and Stellenbosch 2 (b).
Vj ¼�nj þ 1
��nj þ 2
�nj�Rj þ 1
��nj � Rj þ 1
�; j ¼ 1;2;3.N samples; (2)
where,Zj is the empirical logit for the jth sample,Rj ¼ number of infested fruit in the jth sample,nj ¼ total number of fruit in the jth sample,Vj is the variance of the empirical logit for the jth sample.The weighted regression had the form:
zðwÞ ¼ aþ ðbÞln�nT�; (3)
where, z(w) are the weighted empirical logits, a and b are theregression constants and ln (nT) are the natural logs of the trapcounts. There were instances wheremales were caught in the traps,but therewas no fruit infestation. These points were included in theweighted regression. However, in this regression model the furthera point is from 50% infestation, the lower the weighting. Therefore,these points at 0% infestation carry low weightings and do notcontribute as much to the trend in the regression line as the rest ofthe points.
Dummy variable regression (Gujarati, 1970a,, 1970b; Neter andWasserman, 1974; Draper and Smith, 1981) was used to test forpossible differences in the regression constants in (1) between thethree sites. The proportion of infested fruit, p, could be estimatedfor any number, nT, of males trapped using Cox (1970),
tations (right e hand scale) for each of two pear orchard blocks sampled in Stellenbosch
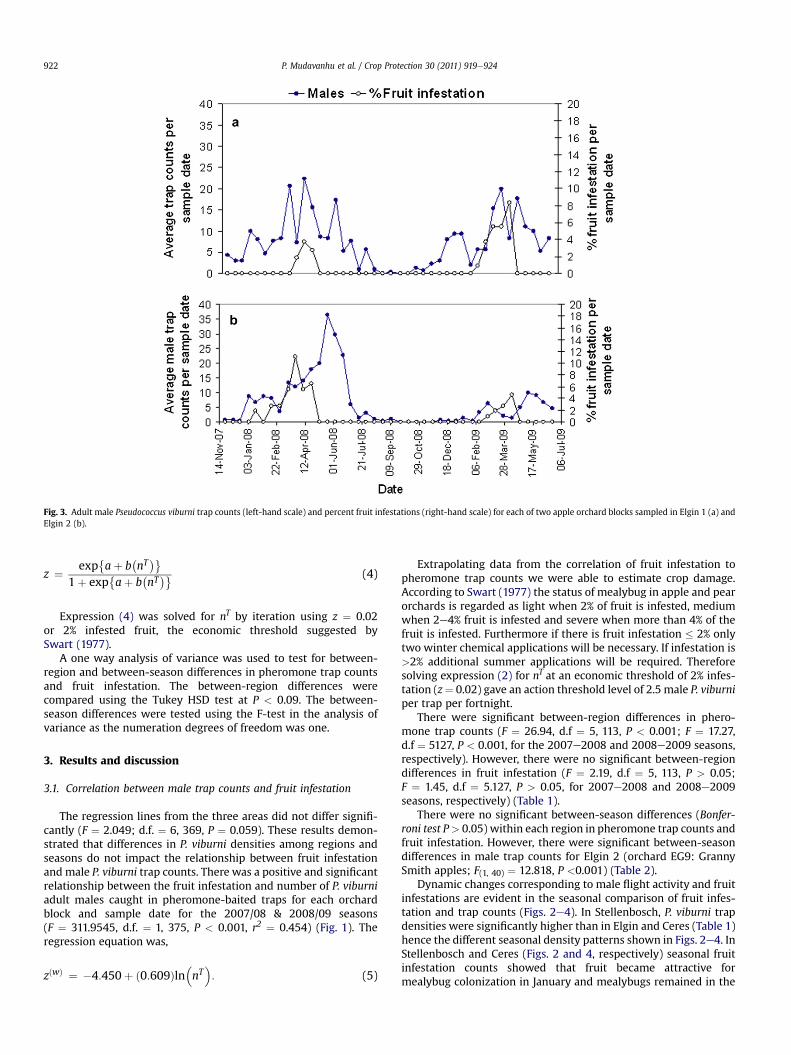
Fig. 3. Adult male Pseudococcus viburni trap counts (left-hand scale) and percent fruit infestations (right-hand scale) for each of two apple orchard blocks sampled in Elgin 1 (a) andElgin 2 (b).
P. Mudavanhu et al. / Crop Protection 30 (2011) 919e924922
z ¼ exp aþ b nT� �T�� (4)
� � ��1þ exp aþ b n
Expression (4) was solved for nT by iteration using z ¼ 0.02or 2% infested fruit, the economic threshold suggested bySwart (1977).
A one way analysis of variance was used to test for between-region and between-season differences in pheromone trap countsand fruit infestation. The between-region differences werecompared using the Tukey HSD test at P < 0.09. The between-season differences were tested using the F-test in the analysis ofvariance as the numeration degrees of freedom was one.
3. Results and discussion
3.1. Correlation between male trap counts and fruit infestation
The regression lines from the three areas did not differ signifi-cantly (F ¼ 2.049; d.f. ¼ 6, 369, P ¼ 0.059). These results demon-strated that differences in P. viburni densities among regions andseasons do not impact the relationship between fruit infestationand male P. viburni trap counts. There was a positive and significantrelationship between the fruit infestation and number of P. viburniadult males caught in pheromone-baited traps for each orchardblock and sample date for the 2007/08 & 2008/09 seasons(F ¼ 311.9545, d.f. ¼ 1, 375, P < 0.001, r2 ¼ 0.454) (Fig. 1). Theregression equation was,
zðwÞ ¼ �4:450þ ð0:609Þln�nT�: (5)
Extrapolating data from the correlation of fruit infestation topheromone trap counts we were able to estimate crop damage.According to Swart (1977) the status of mealybug in apple and pearorchards is regarded as light when 2% of fruit is infested, mediumwhen 2e4% fruit is infested and severe when more than 4% of thefruit is infested. Furthermore if there is fruit infestation � 2% onlytwo winter chemical applications will be necessary. If infestation is>2% additional summer applications will be required. Thereforesolving expression (2) for nT at an economic threshold of 2% infes-tation (z¼ 0.02) gave an action threshold level of 2.5 male P. viburniper trap per fortnight.
There were significant between-region differences in phero-mone trap counts (F ¼ 26.94, d.f ¼ 5, 113, P < 0.001; F ¼ 17.27,d.f ¼ 5127, P < 0.001, for the 2007e2008 and 2008e2009 seasons,respectively). However, there were no significant between-regiondifferences in fruit infestation (F ¼ 2.19, d.f ¼ 5, 113, P > 0.05;F ¼ 1.45, d.f ¼ 5.127, P > 0.05, for 2007e2008 and 2008e2009seasons, respectively) (Table 1).
There were no significant between-season differences (Bonfer-roni test P> 0.05) within each region in pheromone trap counts andfruit infestation. However, there were significant between-seasondifferences in male trap counts for Elgin 2 (orchard EG9: GrannySmith apples; F(1, 40) ¼ 12.818, P <0.001) (Table 2).
Dynamic changes corresponding to male flight activity and fruitinfestations are evident in the seasonal comparison of fruit infes-tation and trap counts (Figs. 2e4). In Stellenbosch, P. viburni trapdensities were significantly higher than in Elgin and Ceres (Table 1)hence the different seasonal density patterns shown in Figs. 2e4. InStellenbosch and Ceres (Figs. 2 and 4, respectively) seasonal fruitinfestation counts showed that fruit became attractive formealybug colonization in January and mealybugs remained in the

Fig. 4. Adult male Pseudococcus viburni trap counts (left-hand scale) and percent fruit infestations (right-hand scale) for each of two orchard blocks sampled in Ceres apples (a) andCeres pears (b).
P. Mudavanhu et al. / Crop Protection 30 (2011) 919e924 923
fruit until harvest in March. In Elgin, where both orchards wereplanted with the late maturing Granny Smith apple cultivar, blockElgin 1 (EW48) (Fig. 3a) had a mixture of Golden Delicious andyounger Granny Smiths though only the Granny Smith trees weresampled. The period of infestation was from February to April inblock EG9 (Fig. 3b) and March to May for EW48 (Fig. 3a).
In all areas, trap counts generally showed that male flight activityis apparent almost throughout the year except for the one Ceres pearorchard where P. viburni density was very low (Fig. 4b). In Stellen-bosch, Elgin 1 and Ceres apples (Figs. 2e4, respectively) trap countsfollowed a similar trend of a steady increase to a peak from lateOctober to January through February coinciding with an increase infruit infestation then temporarily declining fromMarch toMay. Trapcounts then briefly rose from late May to early July before decliningwith the progression of the winter and tree dormancy. A differentseasonal flight pattern was observed in the granny smith appleorchard EG9 (Elgin 2) (Fig. 3b) during both seasons where the peakmale mealybug flight period was during the late summer to earlywinter (March to June), compared to the other orchards.
Data collected from P. viburni pheromone trap counts showedpeaks and troughs of male flight activity which may have repre-sented changes in mealybug seasonal population age structurerather than density. In addition this variation in trap catches couldalso be due to climatic conditions during that trapping session. Thiswas similar to observations by Walton et al. (2004) who investi-gated the male flight activity of a related mealybug species, P. ficus.The summer peak flight periods coincidedwith the fruit season andwere the most important sampling dates, hence this analysisprovided a good fit to the data set (r2 ¼ 0.454) (Fig. 1).
4. Conclusion
The results from this study showed that pheromone-baited trapscould be a useful tool for use by farmers to aid in decision makingwith regards to fruit destined for various markets and targeting ofcontrol actions against P. viburni to prevent future infestations inensuing seasons. In some instances the traps caught male P. viburniwhere female mealybugs were not detected by the visual moni-toring procedure (Fig. 1). Therefore, the traps appear to be moresensitive than visual monitoring and can provide early warning ofP. viburni infestation. Furthermore the similar response acrossa variety of crops where traps have been used for monitoring, in twoyears, suggests that these results are robust and broadly applicable.
Acknowledgements
The research was financially supported by the Deciduous FruitProducers Trust (DFPT) and Technology and Human Resources forIndustry Programme (THRIP). We also thank Mathew Addison,Professor Daane Nel, Dr. Marelize de Villiers, Casper Nyamu-kondiwa, Archbold Sasa and Precious Mudavanhu for their valuablecontribution to this study.
References
Brown, L., Pringle, K.L., 2006. Monitoring System for Pests on Pome Fruit. DeciduousFruit Producers Trust/Syngenta-Stellenbosch University Department ofConservation and Entomology Pamphlet, 4 pp.
Cox, D., 1970. Analysis of Binary Data: Monographs on Statistics and AppliedProbability. Chapman and Hall, USA.

P. Mudavanhu et al. / Crop Protection 30 (2011) 919e924924
de Villiers, M., Pringle, K.L., 2007. Seasonal occurrence of vine pests in commerciallytreated vineyards in the Hex River valley in the Western Cape Province, SouthAfrica. Afr. Entomol. 15, 241e260.
Draper, N.R., Smith, H., 1981. Applied Regression Analysis, second ed.. John Wileyand Sons, New York.
Geiger, C.A., Daane, K.M., 2001. Seasonal movement and sampling of grapemealybug, Pseudococcus maritimus (Ehrhorn) (Homoptera; Pseudococcidae), inSan Joaquin valley vineyards. J. Econ. Entomol. 94, 291e301.
Gujarati, D., 1970a. Use of dummy variables in testing for equality between sets ofcoefficients in two linear regressions: a note. Am. Stat. 24, 50e53.
Gujarati, D., 1970b. Use of dummy variables in testing for equality between sets ofcoefficients in two linear regressions: a generalization. Am. Stat. 24, 18e22.
Millar, J.G., Midland, S.L., Mcelfresh, J.S., Daane, K.M., 2005. (2,3,4,4-tetrame-thylcyclopentyl) methyl acetate, a sex pheromone from the obscure mealybug:first example of a new class of monoterpenes. J. Chem. Ecol. 31, 2999e3005.
Myburgh, A.C., Rust, D.J., Stubbings, D., 1975. Mealybugs on apples and pears.Deciduous Fruit Grow. 25, 176e179.
Neter, J., Wasserman, W., 1974. Applied Linear Statistical Models. Richard D. IrwinInc., Homewood, Illinois.
Pedigo, L.P., 1999. Entomology and Pest Management, third ed.. Prentice Hall, Inc.,New Jersey.
Romoser, W.S., Stoffolano, J.G., 1998. The Science of Entomology, fourth ed..McGraw-Hill, Boston.
Swart, P.L., 1977. Mealybugs: more economic management on apples and pears.Deciduous Fruit Grow. 27, 186e191.
Van der Merwe, F., 2000. Is mealybug on pome fruit under control? (1) Pest status.Deciduous Fruit Grow. 50, 1e6.
Walton, V.M., 2003. Development of an integrated pest management system forvine mealybug, Planococcus ficus (Signoret), in vineyards in the Western CapeProvince, South Africa. PhD Thesis, Univ. of Stellenbosch, Stellenbosch, SouthAfrica, 127p.
Walton, V.M., Daane, K.M., Pringle, K.L., 2004. Monitoring Planococcus ficus inSouth African vineyards with sex pheromone-baited traps. Crop Prot. 23,1089e1096.
Zaviezo, T., Nunez, J., Millar, J.G., 2007. Pheromone-baited traps for monitoringPseudococcus viburni (Hemiptera: Pseudococcidae) in table grapes. AnnualMeeting of the Entomological Society of America. http://www.esa.confex.com/esa/2007/techprogram/paper_30496.htm.12.03.09.




















