
MOLECULAR PHYLOGENY OF CONGENERIC MONOGENEAN PARASITES (DACTYLOGYRUS): A CASE OF INTRAHOST...
18
1001 q 2004 The Society for the Study of Evolution. All rights reserved. Evolution, 58(5), 2004, pp. 1001–1018 MOLECULAR PHYLOGENY OF CONGENERIC MONOGENEAN PARASITES (DACTYLOGYRUS): A CASE OF INTRAHOST SPECIATION ANDREA S ˇ IMKOVA ´ , 1,2,3 SERGE MORAND, 4,5 EDOUARD JOBET, 6,7 MILAN GELNAR, 1,8 AND OLIVIER VERNEAU 2,9 1 Department of Zoology and Ecology, Faculty of Science, Masaryk University, Kotla ´r ˇska ´ 2, 61137 Brno, Czech Republic 2 Parasitologie Fonctionnelle et Evolutive, UMR 5555 CNRS-UP, Universite ´, 52 Avenue Paul Alduy, 66860 Perpignan Cedex, France 3 E-mail: [email protected] 4 Centre de Biologie et de Gestion des Populations, Campus international de Baillarguet, CS 30016, Montferrier sur Lez cedex, 34988 Montpellier, France 5 E-mail: [email protected] 6 Laboratoire Ge ´nome et De ´veloppement des Plantes, UMR 5096 CNRS-UP, Universite ´, 52 Avenue Paul Alduy, 66860 Perpignan Cedex, France 7 E-mail: [email protected] 8 E-mail: [email protected] 9 E-mail: [email protected] Abstract. Dactylogyrus species (Dactylogyridae: Monogenea) are a group of monogenean gill parasites that are highly specific to freshwater fish of the family Cyprinidae. Dactylogyrus species were sampled from 19 cyprinids and one percid collected in Europe. Using partial 18S rDNA and ITS1 sequences, a phylogeny of 51 Dactylogyrus species was reconstructed to investigate the patterns of parasite speciation and diversification. Three main Dactylogyrus lineages were recognized from all phylogenetic trees, that is, analysis of 18S rDNA alone and combined 18SrDNA and ITS1. The first lineage associates the Dactylogyrus species of Cyprinus carpio and Carassius auratus of the Cyprininae; the second associates Dactylogyrus species of the Gobioninae, Pseudorasbora parva of the Rasborinae, and Ctenophar- yngodon idella of the Cyprininae; and the third associates Dactylogyrus species of the Leuciscinae and Alburninae and Barbus barbus of the Cyprininae. Our results suggest that the genus Dactylogyrus is of quite recent origin and that these three lineages separated from each other in a very short period of time. Host subfamily mapping onto the parasite tree inferred from analysis of the combined dataset showed that the Cyprininae could be plesiomorphic hosts for Dactylogyrus. Dactylogyrus parasites would have secondarily colonized the Percidae and representatives of the Leuciscinae, Alburninae, Gobioninae, and Rasborinae. Comparison of host and parasite phylogenetic relationships indicated that a very high number of parasite duplications occurred within two of the three Dactylogyrus lineages. Dactylogyrus diversification can be mainly explained by sympatric intrahost speciation events that seem to be correlated to strict host specificity. Moreover, the present study shows that the congeneric parasites speciating within one host tend to occupy niches within hosts differing at least in one niche parameter. Key words. Cyprinidae, Dactylogyrus, intrahost speciation, molecular phylogeny, Monogenea, niche preference. Received October 20, 2003. Accepted January 8, 2004. The search for patterns and processes of evolution to un- derstand speciation and diversification of parasites has been one of the main subjects in evolutionary biology (Brooks and McLennan 1993). As suggested by Brooks and MacLennan (1993), one important point is the investigation of host spec- ificity that can be the basis of parasite speciation. Parasite speciation may occur in allopatry, that is, on two distinct geographically isolated host species, or sympatry, that is, either on distinct host species by a host shift or in the same host species by duplication. Cospeciation in host-parasite assemblages was first re- ported for pocket gophers and their chewing lice (Hafner and Nadler 1988; Hafner et al. 1994). In that case, allopatric parasite speciation was involved, following geographical iso- lation of their hosts. Thus, when host lineages speciate, par- asites on the descendant host species may also speciate, which is illustrated by congruent host and parasite phylogenies. The presence of sister species in small areas (e.g., isolated islands or lakes) might imply sympatric speciation (Coyne and Price 2000), which can present an outcome of competition for re- sources (Dieckmann and Doebeli 1999). When considering only parasites, sympatric speciation might be the result of association of the host with particular ecological conditions (Klassen and Beverley-Burton 1987; Gue ´gan and Agne `se 1991), thus facilitating host switches. In that case, phylo- genetically closely related parasite species infesting different host species are exemplified by incongruent host and parasite phylogenies. However, sympatric speciation may also be con- sidered when a single host species is infested by a mono- phyletic parasite lineage. In that case, it represents intrahost speciation or parasite duplication, that is, the parasite spe- ciates without a corresponding host cospeciation event and this leads to two or more lineages of parasites being present on single host species (Paterson and Gray 1997). Vickery and Poulin (1998) suggested that this could explain the occur- rence of congeneric parasite species in the same host species. Finally, it is sometimes difficult to distinguish between sym- patric and allopatric speciation when parasite extinction has occurred during host evolution (Page et al. 1998; Paterson and Banks 2001). In that case, the phylogenetic patterns of parasites cannot be easily explained, even in the light of host relationships. The Monogenea, as a group of ectoparasites with a direct life cycle that predominantly live on the gills and skin of fish, seem to be an appropriate model for studying the process of parasite diversification, mainly because of their high spe- cies richness and morphological and ecological diversity (Poulin 2002). Moreover, monogeneans are highly host (re-
Transcript of MOLECULAR PHYLOGENY OF CONGENERIC MONOGENEAN PARASITES (DACTYLOGYRUS): A CASE OF INTRAHOST...

1001
q 2004 The Society for the Study of Evolution. All rights reserved.
Evolution, 58(5), 2004, pp. 1001–1018
MOLECULAR PHYLOGENY OF CONGENERIC MONOGENEAN PARASITES(DACTYLOGYRUS): A CASE OF INTRAHOST SPECIATION
ANDREA SIMKOVA,1,2,3 SERGE MORAND,4,5 EDOUARD JOBET,6,7 MILAN GELNAR,1,8 AND OLIVIER VERNEAU2,9
1Department of Zoology and Ecology, Faculty of Science, Masaryk University, Kotlarska 2, 61137 Brno, Czech Republic2Parasitologie Fonctionnelle et Evolutive, UMR 5555 CNRS-UP, Universite, 52 Avenue Paul Alduy,
66860 Perpignan Cedex, France3E-mail: [email protected]
4Centre de Biologie et de Gestion des Populations, Campus international de Baillarguet, CS 30016, Montferrier sur Lez cedex,34988 Montpellier, France
5E-mail: [email protected] Genome et Developpement des Plantes, UMR 5096 CNRS-UP, Universite, 52 Avenue Paul Alduy,
66860 Perpignan Cedex, France7E-mail: [email protected]: [email protected]
9E-mail: [email protected]
Abstract. Dactylogyrus species (Dactylogyridae: Monogenea) are a group of monogenean gill parasites that are highlyspecific to freshwater fish of the family Cyprinidae. Dactylogyrus species were sampled from 19 cyprinids and onepercid collected in Europe. Using partial 18S rDNA and ITS1 sequences, a phylogeny of 51 Dactylogyrus specieswas reconstructed to investigate the patterns of parasite speciation and diversification. Three main Dactylogyrus lineageswere recognized from all phylogenetic trees, that is, analysis of 18S rDNA alone and combined 18SrDNA and ITS1.The first lineage associates the Dactylogyrus species of Cyprinus carpio and Carassius auratus of the Cyprininae; thesecond associates Dactylogyrus species of the Gobioninae, Pseudorasbora parva of the Rasborinae, and Ctenophar-yngodon idella of the Cyprininae; and the third associates Dactylogyrus species of the Leuciscinae and Alburninaeand Barbus barbus of the Cyprininae. Our results suggest that the genus Dactylogyrus is of quite recent origin andthat these three lineages separated from each other in a very short period of time. Host subfamily mapping onto theparasite tree inferred from analysis of the combined dataset showed that the Cyprininae could be plesiomorphic hostsfor Dactylogyrus. Dactylogyrus parasites would have secondarily colonized the Percidae and representatives of theLeuciscinae, Alburninae, Gobioninae, and Rasborinae. Comparison of host and parasite phylogenetic relationshipsindicated that a very high number of parasite duplications occurred within two of the three Dactylogyrus lineages.Dactylogyrus diversification can be mainly explained by sympatric intrahost speciation events that seem to be correlatedto strict host specificity. Moreover, the present study shows that the congeneric parasites speciating within one hosttend to occupy niches within hosts differing at least in one niche parameter.
Key words. Cyprinidae, Dactylogyrus, intrahost speciation, molecular phylogeny, Monogenea, niche preference.
Received October 20, 2003. Accepted January 8, 2004.
The search for patterns and processes of evolution to un-derstand speciation and diversification of parasites has beenone of the main subjects in evolutionary biology (Brooks andMcLennan 1993). As suggested by Brooks and MacLennan(1993), one important point is the investigation of host spec-ificity that can be the basis of parasite speciation. Parasitespeciation may occur in allopatry, that is, on two distinctgeographically isolated host species, or sympatry, that is,either on distinct host species by a host shift or in the samehost species by duplication.
Cospeciation in host-parasite assemblages was first re-ported for pocket gophers and their chewing lice (Hafner andNadler 1988; Hafner et al. 1994). In that case, allopatricparasite speciation was involved, following geographical iso-lation of their hosts. Thus, when host lineages speciate, par-asites on the descendant host species may also speciate, whichis illustrated by congruent host and parasite phylogenies. Thepresence of sister species in small areas (e.g., isolated islandsor lakes) might imply sympatric speciation (Coyne and Price2000), which can present an outcome of competition for re-sources (Dieckmann and Doebeli 1999). When consideringonly parasites, sympatric speciation might be the result ofassociation of the host with particular ecological conditions(Klassen and Beverley-Burton 1987; Guegan and Agnese
1991), thus facilitating host switches. In that case, phylo-genetically closely related parasite species infesting differenthost species are exemplified by incongruent host and parasitephylogenies. However, sympatric speciation may also be con-sidered when a single host species is infested by a mono-phyletic parasite lineage. In that case, it represents intrahostspeciation or parasite duplication, that is, the parasite spe-ciates without a corresponding host cospeciation event andthis leads to two or more lineages of parasites being presenton single host species (Paterson and Gray 1997). Vickery andPoulin (1998) suggested that this could explain the occur-rence of congeneric parasite species in the same host species.Finally, it is sometimes difficult to distinguish between sym-patric and allopatric speciation when parasite extinction hasoccurred during host evolution (Page et al. 1998; Patersonand Banks 2001). In that case, the phylogenetic patterns ofparasites cannot be easily explained, even in the light of hostrelationships.
The Monogenea, as a group of ectoparasites with a directlife cycle that predominantly live on the gills and skin offish, seem to be an appropriate model for studying the processof parasite diversification, mainly because of their high spe-cies richness and morphological and ecological diversity(Poulin 2002). Moreover, monogeneans are highly host (re-

1002 ANDREA SIMKOVA ET AL.
stricted to one or a few host species) and niche specific (re-stricted to several microhabitats within the same host spe-cies), and it has been shown that congeneric species couldcoexist on the same host because of a low level of interspe-cific competition (Rohde 1977, 1979, 1991; Simkova et al.2000). To date, several models of congeneric monogeneanspecies have been investigated to study the process of parasitediversification using phylogenies (Littlewood et al. 1997;Desdevises et al. 2002; Huyse and Volckaert 2002).
Congeneric gill parasites belonging to Lamellodiscus(Monopisthocotylea), a group of parasites specific to marinefish of the Sparidae, were used as a model by Desdevises etal. (2002). These parasites are either generalists, infestingdifferent host species, or specialists that may infest one hostspecies. Desdevises et al. (2002) suggested that the genuswas old and that intraspecific morphological variability couldexplain their ability to colonize different host species. Theyproposed that the rapid speciation of Lamellodiscus that infesthosts living in sympatry happened without any cospeciationand intrahost speciation. Similarly, no intrahost speciationwas reported for Gyrodactylus (Monopisthocotylea), a genusoccurring in multiple parasite lineages on single host speciesof freshwater and marine fish (Huyse and Volckaert 2002).Zietara and Lumme (2002) proposed the host switch as themodel of Gyrodactylus speciation facilitated by their partic-ular reproductive system.
Polystomes (Polyopisthocotylea) are geographically wide-spread endoparasitic monogeneans infecting mainly amphib-ians and freshwater turtles. These parasites are morpholog-ically very similar, are highly host specific, and congenericspecies can be found on the same host species within turtles,but infecting different niches (i.e., oral cavities, urinary blad-ders, and conjunctival sacs). Littlewood et al. (1997), byinvestigating phylogenetic relationships of six polystomespecies of turtles, showed no intrahost speciation and con-cluded that the occurrence of congeneric species within thesame host species could be the result of either host switchingor cospeciation.
Therefore, one may ask whether parasite speciation mayoccur within a single host. Moreover, within one host species,different congeneric species can occupy different niches, thatis, microhabitats within the host (Rohde 1989). Therefore,one could also ask whether phylogenetically closely relatedcongeneric species parasitize similar or different microhab-itats within hosts. In the case of site-specific polystomemonogeneans, congeneric species infecting the same nichewithin the different host species (even if geographically iso-lated host species) are more closely related than congenersinfecting different sites of the same host species (Littlewoodet al. 1997). Following the hypotheses of Rohde (1991), nichesegregation or reproductive isolation between congenericspecies parasitizing the same host species has been predicted,to prevent competition and to increase intraspecies matingcontacts (Simkova et al. 2000; Morand et al. 2002).
The model of congeneric monogeneans of the genus Dac-tylogyrus (Dactylogyrinae: Dactylogyridae: Monopisthoco-tylea) was investigated in this study for the following reasons:Dactylogyrus is a highly diversified group within the Mono-genea (Gusev 1985) with more than 900 nominal speciesmainly restricted to freshwater fish of the Cyprinidae (Gibson
et al. 1996), although, occasionally reported on the Percidae(Gusev 1985; Valtonen et al. 1990; Cone et al. 1994; Hayward1997). In this genus, a high number of congeneric speciescoexisting on the same host species has been reported (Rohde1989; Kennedy and Bush 1992; Simkova et al. 2000, 2002),and several studies have been conducted on host specificitywithin the Cyprinidae (Guegan and Agnese 1991; Gueganand Lambert 1991; El Gharbi et al. 1994; Lambert and ElGharbi 1995; Simkova et al. 2001). The coexistence of Dac-tylogyrus species on the same host was suggested to be fa-cilitated by niche distances and differing morphology of re-productive apparatus (Simkova et al. 2002; Jarkovsky et al.2004).
Similarly, the Cyprinidae is the most diverse family amongfreshwater fish with about 2000 species (Helfman et al. 1997).Seven subfamilies are recognized, among them six with rep-resentatives occurring in Europe (Acheilognathinae, Albur-ninae, Cyprininae, Gobioninae, Leuciscinae, and Rasbori-nae), the most diverse being the Leuciscinae (Winfield andNelson 1991). It has been believed for a long time that cyp-rinids originated in Asia and later dispersed to Europe, Africa,and North America (Winfield and Nelson 1991). Neverthe-less, it is still questioned whether eastern Asia was an im-portant interchange for cyprinids or a center of speciation(Durand et al. 2002).
The present study carried out phylogenetic analyses, basedon partial 18S, or small subunit, ribosomal RNA gene (rDNA)and ribosomal RNA gene internal transcribed spacer 1 (ITS1)sequences obtained from 51 Dactylogyrus species sampledfrom cyprinids and one percid species of central Europe. Theprocess of parasite speciation and diversification, especiallywhether congeneric parasites coexisting on the same hostspecies have speciated via intrahost speciation and whetherphylogenetically related parasites occupied similar or differ-ent microhabitats, was investigated.
MATERIALS AND METHODS
Parasite Sampling
A total of 51 Dactylogyrus species, including specialist andgeneralist species, were collected from freshwater fish of theMorava River basin in the Czech Republic (central Europe).Among them, 49 species parasitizing fish species belongingto the family Cyprinidae (19 cyprinid species) and two spe-cies parasitizing Gymnocephalus cernuus belonging to thefamily Percidae, were recognized. A total of 44 Dactylogyrusspecies were collected only on one host species, and sevenDactylogyrus species were collected on more than one hostspecies. For generalist species, several individuals of the dif-ferent host species were collected. Dactylogyrus species withtheir hosts are given in Table 1. This sample is representativeof the fauna living in central Europe (71 species of Dactyl-ogyrus and 34 host species are known).
Fish sampling was performed as described in Ergens andLom (1970). Parasites were removed from the gills, placedon slides, covered by a coverslip, and identified using a lightmicroscope equipped with phase contrast, differential inter-ference contrast, and digital image analysis (MicroImage 4.0for Windows, Olympus Optical Co., Hamburg, Germany).The sclerotized parts of the parasite attachment organ (the

1003MOLECULAR PHYLOGENY OF DACTYLOGYRUS
TABLE 1. Dactylogyrus sequences obtained in this study with their host species and GenBank accession numbers for partial 18SrDNAand ITS1. Numbers 1, 2, and 3 on the right column refer to the Dactylogyrus lineages deduced from the parasite neighbor-joining tree.Dactylogyrus species and their hosts investigated for niche preference are indicated by asterisks.
Fish species Dactylogyrus species Accession number Dactylogyrus lineage
Abramis ballerus
Abramis brama*
Abramis sapa
auriculatuschranilowifalcatuswunderi*zandti*propinquus
AJ564112AJ564117AJ564130AJ564164AJ564165AJ564147
333333
Alburnus alburnus*
Aspius aspius
alatus*fraternus*minor*parvus*ramulosustuba
AJ564109AJ564136AJ564143AJ564146AJ564149AJ564158
333333
Barbus barbus*
Abramis bjoerkna*
carpathicus*dykimalleus*cornoides*cornu*distinguendus*sphyrna*
AJ564115AJ564127AJ564142AJ564118AJ564119AJ564125AJ564155
3333333
Carassius auratus* anchoratus*dulkeiti*formosus*inexpectatus*intermedius*vastator*
AJ564111AJ564126AJ564135AJ564138AJ564139AJ564159
111111
Chondrostoma nasus
Ctenopharyngodon idellaCyprinus carpio
chondrostomiergensivistulaelamellatusachmerovianchoratusextensus
AJ564116AJ564128AJ564160AJ564141AJ564108AJ490161AJ564129
3332111
Gobio albipinatusGobio gobioGymnocephalus cernuus
Leuciscus cephalus*
finitimuscryptomeresamphibothriumhemiamphibothriumfallaxfolkmanovae*nanoides*
AJ564133AJ564123AJ564110AJ564137AJ564132AJ564134AJ564144
2233333
Leuciscus idus
prostae*vistulae*vranoviensis*cruciferramulosustubavistulae
AJ564148AJ564161AJ564163AJ564122AJ564150AJ564157AJ564162
3333333
Phoxinus phoxinusPseudorasbora parvaRutilus rutilus*
borealissquameuscaballeroi*crucifer*fallax*nanus*rarissimus*
AJ564113AJ564156AJ564114AJ564120AJ564131AJ564145AJ564151
3233333
Scardinius erythrophthalmus*
rutili*similis*sphyrnacruciferdifformis*difformoides*izjumovae*
AJ564152AJ564153AJ564154AJ564121AJ490160AJ564124AJ564140
3333333
opisthaptor; i.e., the central hooks called anchors, seven pairsof marginal hooks, one connective bar [dorsal in this case]or two connective bars [dorsal and ventral]) and reproductiveorgans (vaginal armaments and copulatory organs) were used
for parasite determination according to Gusev (1985). Someparasite specimens were fixed in a mixture of glycerine andammonium picrate and deposited in the collection of the De-partment of Zoology and Ecology, Masaryk University (Brno,

1004 ANDREA SIMKOVA ET AL.
Czech Republic). Other parasites were preserved in 95% eth-anol before DNA extraction.
DNA Extraction and Polymerase Chain Reaction
Individual parasites were removed from ethanol, dried, andDNA was extracted using the standard phenol-chloroformmethod (Sambrook et al. 1989). For generalist species, in-dividuals from different host species were investigated. TotalDNA obtained from individual parasites was resuspended in25 ml distilled water. Partial 18S rDNA and entire ITS1 re-gion were amplified in one round using primer S1 (59-ATTCCGATAACGAACGAGACT-39) that anneals in theterminal region of the 18S gene (Sinnappah et al. 2001) andH7 (59-GCTGCGTTCTTCATCGATACTCG-39; Sinnappahet al. 2001) or IR8 primers (59-GCTAGCTGCGTTCTTCA-TCGA-39; Simkova et al. 2003) that anneal in the 5.8S rDNA.Each amplification reaction was performed in a final volumeof 25 ml containing 1.5 units of Taq polymerase, 1X buffer,1.5 mM MgCl2, 200 mM of each dNTP, 0.8 mM of eachpolymerase chain reaction (PCR) primer and 3 ml DNA. PCRwas carried out with the following steps: 4 min at 958C fol-lowed by 35 cycles of 1 min at 958C, 1 min at 558C, and 1min 30 sec at 728C, and 10 min of final elongation at 728C.Electrophoresis was performed on a 1% agarose gel stainedwith ethidium bromide for DNA visualization. PCR productswere cut from the gel and purified using siliconized glass-wool. DNA was precipitated and resuspended in sterile water.
Some of the PCR products were sequenced directly. Otherswere ligated into the pGEM-T vector and cloned in Esche-richia coli JM109 competent cells when the amount of pu-rified DNA after PCR was too low for direct sequencing.Recombinant colonies were checked following Sekar’s pro-cedure (1987) and plasmids were purified with the WizardPlus miniprep kit (Promega, Madison, WI). At least threeclones were sequenced per species.
DNA Sequencing
The direct sequencing of purified PCR products as well asrecombinant plasmids was performed using the same primersas for PCR or universal primers (T7 and SP6) supplied byPromega. Moreover, for sequencing species included in theD. minor group (group 3, see Results), two forward primersIF8 (59-AACTGTTCAATCATCGTCGTG-39; Simkova et al.2003) and newly designed IF9 (59-ATCCGCCGACTCT-GACTGGA-39) and two reverse primers IR5 (59-TA-CGGAAACCTTGTTACGAC-39; Sinnappah et al. 2001) andnewly designed IR9 (59-RRGACTCACCCGAAGGGAG-39)were used. For sequencing of species included in the D. an-choratus group (groups 1 and 2, see Results), the same in-ternal primers were used, with the exception of IF9, whichwas replaced by IF10 (59-YMTTCTCCCTTCGGGTGAGT-39) also designed for this study.
Sequences obtained, including partial 18S rDNA, completeITS1, and partial 5.8S rDNA ranged from 962 to 1130 bp.Primers IF8 and IR5 anneal in the 18S rDNA at positions234–254 and 471–490, respectively, of D. difformis sequenceaccession no. AJ490160. The primers IF9 and IR9 anneal inthe ITS1 region at positions 223–243 and 348–367, respec-tively, of D. difformis, and IF10 anneals in the ITS1 region
at positions 274–294 of D. anchoratus accession no.AJ490161. Sequencing was carried out using ABI Prism BigDye Terminator Cycle Sequencing kit (Applied Biosystems,Foster City, CA) and electrophoresis was performed on anautomated sequencer (ABI373 DNA Sequencer). Sequenceswere read and corrected using Sequencher software (GeneCodes Corp., Ann Arbor, MI). New sequences were depositedin EMBL (see Table 1 for their accession numbers).
Phylogenetic Analyses of Dactylogyrus
DNA sequences were aligned using the ED program in theMUST package (Philippe 1993). Gaps and ambiguouslyaligned regions were removed. First, phylogenetic analysisusing partial 18S rDNA alone was performed to polarizeDactylogyrus evolution as the variability of this marker al-lows for unambiguous alignment with the outgroup. For thisanalysis seven other monogenean species belonging to dif-ferent subfamilies of the Dactylogyridae were included, fol-lowing the phylogenetic relationships previously inferred forDactylogyridae (Simkova et al. 2003). Among them two spe-cies, Thaparocleidus vistulensis (accession no. AJ490165) ofthe Ancylodiscoidinae and Cleidodiscus pricei (AJ490168)of the Ancyrocephalinae, were used for rooting the tree. Fiveother species previously found to form a group includingDactylogyrus species were also used (i.e., Pseudodactylogy-roides apogonis, Pseudodactylogyrus anguillae, and P. hazeof the Pseudodactylogyrinae and Thylacicleidus sp. andPseudohaliotrema sphincteropus of the Ancyrocephalinae;AB065115, AJ490162, AB0651141, AJ490169, AJ287568,respectively). For generalist species, only the sequence of theindividual parasitizing the most commonly infested host wasretained, so 58 species were analyzed. Distance trees weregenerated with a neighbor-joining (NJ) algorithm based onKimura two-parameter distances (Kimura 1980) and per-formed with PAUP* 4b10 (Swofford 2002). Support valuesfor internal nodes were estimated using a bootstrap resam-pling procedure with 1000 replicates (Felsenstein 1985).Maximum likelihood (ML) and NJ analyses based on MLdistances were also conducted using PAUP from the bestappropriate model (TrNf 1 G 1 I in this case) selected bythe ModelTest program (Posada and Crandall 1998). Thesearch of the best ML tree was done using a branch-swappingalgorithm (TBR, tree bisection reconnection). One hundredreplicates for ML and 1000 replicates for NJ were calculatedusing the same model as above, using the TBR branch-swap-ping algorithm for ML.
Maximum parsimony (MP) analysis was also performedusing heuristic search with stepwise random addition se-quence running on unweighted informative characters. Sup-port values for internal nodes were estimated after 1000 rep-licates using the TBR branch-swapping algorithm.
Further analyses were performed using combined sequenc-es from partial 18S rDNA and ITS1 without including otherspecies belonging to the Dactylogyridae as ITS1 sequencesof Dactylogyrus species could not be aligned with ITS1 se-quences of the seven species of Dactylogyridae used in theanalysis of 18S rDNA. The analyses of combined 18S rDNAand ITS1 data were done to obtain a better phylogeneticresolution within the Dactylogyrus genus. NJ analysis was

1005MOLECULAR PHYLOGENY OF DACTYLOGYRUS
TABLE 2. Fish species used for phylogenetic analysis, with the exception of Gobio albipinatus, for which the cytochrome b gene hasnot yet been sequenced. Asterisks indicate species for which Dactylogyrus species were recorded. The taxonomic position and GenBankaccession numbers for cytochrome b sequences are indicated for each species.
Order Family Subfamily Species Accession number
Cypriniformes BalitoridaeCharacidaeCyprinidae Cyprininae
Crossostoma lacustreAstyanax mexicanusBarbus barbus*Carassius auratus*Cyprinus carpio*Ctenopharyngodon idella*
M91245AF045997Y10450AF051858X61010AF051860
Gobioninae
RasborinaeLeuciscinae
Gobio albipinatus*Gobio gobio*Pseudorasbora parva*Abramis ballerus*Abramis bjoerkna*
no sequence availableY10452Y10453AY026409Y10442
Abramis brama*Abramis sapa*Aspius aspius*Chondrostoma nasus*Leuciscus cephalus*Leuciscus idus*
Y10441AY026408AY026398AY026402Y10446AY026397
Perciformes PercidaeAlburninae
Phoxinus phoxinus*Rutilus rutilus*Scardinius erythrophthalmus*Alburnus alburnus*Ammocrypta claraEtheostoma kennicotti
Y10448Y10440Y10444Y10443AF045350AF045341
Salmoniformes Salmonidae
Gymnocephalus cernuus*Perca flavencensPerca fluviatilisZingel streberOncorhynchus mykiss
AF045356AJ001521AF045358AF045360L29771
performed as described above from Tamura Nei distancesincluding a proportion of invariable characters and gammadistribution (TrN 1 I 1 G). One thousand bootstrap replicateswere conducted following the same evolutionary model. MLanalyses were also performed on the TrN 1 I 1 G modelselected by ModelTest. One hundred replicates were calcu-lated following the same model and using the same proceduredescribed for the analyses of 18S rDNA alone. MP analysiswas performed as described in the case of 18S rDNA.
Host Phylogeny
The phylogeny of cyprinids has been previously investi-gated using mitochondrial markers (Briolay et al. 1998; Gilleset al. 1998, 2001; Zardoya and Doadrio 1999; Zardoya et al.1999; Durand et al. 2002), but not all species used in ourstudy were included. Thus, we reconstructed the host phy-logeny from cytochrome b sequences retrieved from Gen-Bank, including 18 representatives of the Cyprinidae, one ofthe Balitoridae, one of the Characidae, and six of the Per-cidae. The tree was rooted with Oncorhynchus mykiss fromthe Salmonidae. All species with their accession numbers aregiven in Table 2.
The ModelTest program was used for selecting the bestevolutionary model for ML analysis (TVM 1 I 1 G). Supportvalues for internal nodes were estimated using a bootstrapresampling procedure with 100 replicates following a branch-swapping algorithm (TBR) on the same model. NJ analysiswas performed using evolutionary parameters selected byModelTest. One thousand replicates were calculated follow-
ing the same model. MP analysis was performed as describedfor previous analyses.
Mapping of Host Subfamilies onto the Parasite Tree
The host subfamily for each Dactylogyrus species wasmapped onto the NJ tree inferred from analysis of combineddata on 51 Dactylogyrus species, using MacClade version4.0.1 with Farris optimization (Maddison and Maddison1992). Host species were divided into subfamilies accordingto Nelson (1994) and are given in Table 2. For parasite spe-cies that have been recorded on different host species (gen-eralists) of different host subfamilies, mapping was per-formed with the host subfamily from which parasites wereexamined and analyzed. It should be noted that this hostsubfamily corresponds to the most infested host species.
Comparison of Host and Parasite Phylogenies
TreeMap 1.0 (Page 1994) was used to represent host-par-asite associations using the Dactylogyrus tree inferred fromanalysis of combined data and the cyprinid tree after ex-cluding fish species not infested by Dactylogyrus. Gobio al-bipinatus was added to the fish topology as a sister speciesof G. gobio considered as a monophyletic group (Barus andOliva 1995).
Mapping of Niche Preference
The mapping of niche preference was performed using thesame methodology as for mapping of host subfamilies. For

1006 ANDREA SIMKOVA ET AL.
niche preference, data concerning niche position (i.e., theposition on the gills within the hosts) were recorded for 33Dactylogyrus species obtained from eight different cyprinidspecies (asterisks in Table 1). We could not obtain niche datafor all Dactylogyrus species from all host species investigatedbecause of either low host sample or low parasite sample.Some fish species were found only rarely, and even whenusing a relatively high number of host individuals we ob-tained only a few individual specimens of several Dactylo-gyrus species.
The parasite niche was investigated as previously describedin Simkova et al. (2000). For a description of the gill arch,see Figure 6A. The niche position of each parasite individualwas recorded (i.e., its position on the gill arch, gill segment,gill area), and the position where the maximum number ofparasite individuals was found was considered as the pre-ferred niche. The preferred niche (gill arch, segment, andarea) was mapped onto the phylogenetic tree obtained fromthe NJ analysis using combined 18S rDNA and ITS1 se-quences.
RESULTS
Molecular Divergence within Generalist Species
For each of the seven generalist species investigated, wesequenced at least two individuals recovered from each dif-ferent host species. Whereas no differences were detectedbetween individuals collected from the same host species,differences were observed between individuals collectedfrom different host species. Pairwise comparisons showed0.4% molecular divergence between D. anchoratus specimensfrom Cyprinus carpio and Carassius auratus; no differencebetween D. crucifer specimens from Rutilus rutilus, Scardi-nius erythrophthalmus, and Leuciscus idus; 0.3% moleculardivergence between D. fallax specimens from R. rutilus andLeuciscus cephalus; 1% molecular divergence between D.ramulosus specimens from Aspius aspius and L. idus; 0.6%molecular divergence between D. sphyrna specimens ofAbramis bjoerkna and R. rutilus; and no difference betweenD. tuba specimens of Aspius aspius and L. idus. Finally, 0.2%divergence was calculated between D. vistulae specimensfrom Chondrostoma nasus and L. idus; the same order ofdivergence (1.2%) was found between D. vistulae specimensfrom C. nasus and L. idus with specimens from L. cephalus.The genetic divergence between the two most closely relatedspecies was 1.4% (D. caballeroi and D. crucifer), and thegenetic divergence between other pairs was more than 2.6%.
Phylogenetic Analysis Using Partial 18S rDNA
For the phylogenetic analyses based on partial 18S rDNAalone we used all Dactylogyrus species sequenced for thisstudy and seven other dactylogyrids. The sequence alignmentcomprised 440 unambiguously alignable positions after re-moving gaps and ambiguous aligned regions, of which 150were variable with 106 parsimony informative. The first anal-ysis was performed using NJ with Kimura two-parameterbecause of equal nucleotide base frequencies (about 25%).The tree obtained from the Kimura two-parameter distances
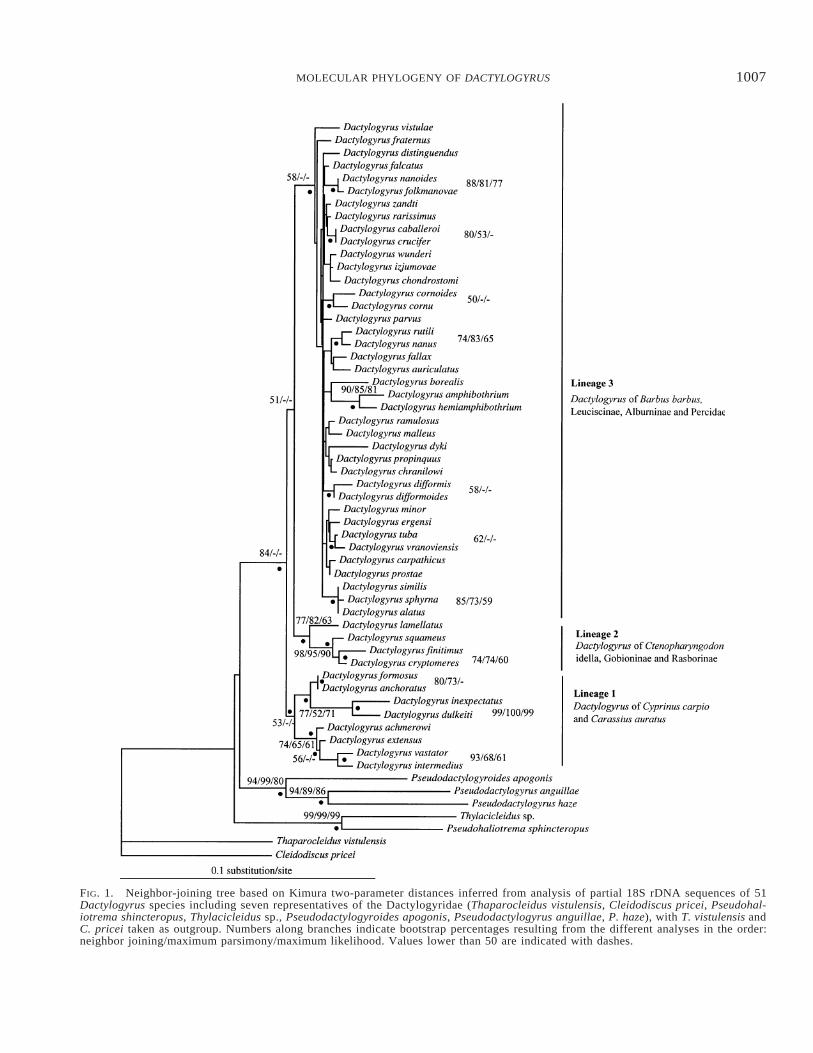
with bootstrap proportions (BP) supported the monophyly ofDactylogyrus (BP 5 84; see Fig. 1).
ML analysis was performed on the TrNf 1 G 1 I model,with the following parameters: substitution rate matrix: A-C5 1.000, A-G 5 4.1211, A-T 5 1.000, C-G 5 1.000, C-T5 5.4533, G-T 5 1.000; proportion of invariable sites 50.4342; and rate heterogeneity approximated by a gammadistribution (four rate categories), a 5 0.5912. Surprisingly,the best ML tree (not shown) shows nonmonophyly of Dac-tylogyrus. Indeed, the group including D. cryptomeres, D.finitimus, D. squameus, and D. lamellatus was positioned asthe sister group of the Pseudodactylogyrinae. Following NJanalysis using TrNf 1 G 1 I, species of Pseudodactylogyr-inae are not nested within Dactylogyrus species. Neverthe-less, the monophyly of Dactylogyrus was again not supported.MP analysis was also performed. In MP analyses, 100 equallyparsimonious trees with 393 steps were retained (consistencyindex, CI 5 0.550, retention index, RI 5 0.639). However,BP values for MP show a very weak phylogenetic resolution.No statistically significant differences among NJ on Kimuratwo-parameter, ML tree, NJ based on ML distance (TrNf 1G 1 I), and the equally most parsimonious 100 trees werefound (Shimodaira-Hasegawa test implemented in PAUP*4b10, P . 0.05).
This incongruence (monophyly vs. nonmonophyly of Dac-tylogyrus) may be due to different evolutionary rates betweenDactylogyrus species and the other species used as outgroup.Li (1997) stated that ML methods may become inconsistentif the rate of evolution is assumed to be uniform when infact it is not. We observed highly variable substitution ratesamong the branch lengths in the tree inferred from the MLapproach, suggesting that this assumption may be violated.Furthermore, we noted that rate heterogeneity was differentbetween Dactylogyrus species and other dactylogyrids. Fi-nally, the high number of parameters for the TrNf 1 G 1 Imodel involves a higher variance of branch-length estimationand therefore may lead to incorrect reconstruction of internalshort branches (Nei and Kumar 2000). For those reasons, weretained the NJ tree based on the Kimura two-parameter dis-tances that shows the monophyly of Dactylogyrus.
Within Dactylogyrus we observe three monophyletic lin-eages, which are moderately supported (BP values). The first(further referenced as clade 1) includes the Dactylogyrus spe-cies from two species of the Cyprininae; the second (clade2) associates one Dactylogyrus species from the Cyprininae,one from the Rasborinae, and two from the Gobioninae; andthe third (clade 3) includes mainly Dactylogyrus species par-asitizing species of the subfamilies Leuciscinae and Albur-ninae, three Dactylogyrus species of Barbus barbus (subfam-ily Cyprininae), and two of the family Percidae. While noclear phylogenetic pattern is observed among the three clades,relationships between species within clades 1 and 2 are betterresolved than relationships between species of clade 3 (seeFig. 1).
Phylogenetic Analysis Using Combined Data(18S rDNA and ITS1)
The partition homogeneity test as implemented in PAUP*4b10 (Swofford 2002) was used to test the congruence of the
1007MOLECULAR PHYLOGENY OF DACTYLOGYRUS
FIG. 1. Neighbor-joining tree based on Kimura two-parameter distances inferred from analysis of partial 18S rDNA sequences of 51Dactylogyrus species including seven representatives of the Dactylogyridae (Thaparocleidus vistulensis, Cleidodiscus pricei, Pseudohal-iotrema shincteropus, Thylacicleidus sp., Pseudodactylogyroides apogonis, Pseudodactylogyrus anguillae, P. haze), with T. vistulensis andC. pricei taken as outgroup. Numbers along branches indicate bootstrap percentages resulting from the different analyses in the order:neighbor joining/maximum parsimony/maximum likelihood. Values lower than 50 are indicated with dashes.

1008 ANDREA SIMKOVA ET AL.
two datasets: 18S and ITS1. As no significant difference wasfound (P 5 0.100), further analyses were performed with the51 Dactylogyrus species examined in this study using com-bined sequences from partial 18S rDNA and ITS1. The align-ment of the partial 18S rDNA and ITS1 sequences comprised712 unambiguously alignable positions after gaps were re-moved, of which 246 were variable with 201 parsimony in-formative. The ML analysis was performed on the model TrN1 I 1 G with the following parameters: substitution ratematrix: A-C 5 1.0000, A-G 5 3.0453, A-T 5 1.0000, C-G5 1.0000, C-T 5 3.7850, G-T 5 1.0000; the proportion ofinvariable sites 5 0.5341; and rate heterogeneity approxi-mated by a gamma distribution (four rate categories), a 50.7423. The topology of the best ML tree was similar to thetopology inferred from NJ analysis on the Tamura Nei dis-tances including one proportion of invariable characters andgamma distribution (TrN 1 I 1 G). MP analysis was alsoperformed and 100 equally parsimonious trees with 951 stepswere retained (CI 5 0.428, RI 5 0.621). The strict consensusdisplays similar topology to the NJ and ML trees, that is, itsupports the clustering of species found from the NJ and MLanalyses with BP of more than 50%. No statistically signif-icant differences among NJ, ML, and trees obtained by MPwere found (Shimodaira-Hasegawa test implemented inPAUP* 4b10, P . 0.05), therefore we only present the NJtree giving BP values obtained from all three analyses (Fig.2).
The phylogenetic relationships inferred from 18S rDNAanalysis (Fig. 1) do not allow us to clearly specify which ofthe three reported lineages diverged first. Therefore, the treeobtained from combined 18S rDNA and ITS1 data was drawnto reflect the phylogenetic relationships described in the anal-ysis of 18S rDNA sequences, that is, three Dactylogyrusgroups. Each of the three Dactylogyrus groups is well sup-ported (BP values more than 90). Moreover, the midpointrooting technique was applied as implemented in PAUP*4b10 and highlights the first Dactylogyrus group as the mostbasal lineage.
Several general trends are inferred from all analyses. Thefirst lineage (group 1, Fig. 2) is divided into two sister sub-groups. The first associates two specialists found on C. carpiothat are related to the two specialists found on C. auratus.The second one associates four species found on C. auratus.However, one of them, D. anchoratus, is also found on C.carpio.
The second lineage (group 2, Fig. 2) is composed of fourspecies. Two Dactylogyrus species found on Gobio speciesare related to D. squameus from P. parva. The fourth species,D. lamellatus from Ctenopharyngodon idella, is the most bas-al. All associations within the first and second lineages arewell supported by BP in all analyses.
The third lineage (group 3, Fig. 2) can be divided in twogroups: the first one associates two Dactylogyrus species ofthe Percidae (Gymnocephalus cernuus), and the second as-sociates all remaining Dactylogyrus species, that is, parasitesof the Leuciscinae, A. alburnus of the Alburninae, and B.barbus of the Cyprininae. Phylogenetic interrelationshipsamong species of this group are in general moderately orpoorly supported or unresolved with the exception of onesubgroup of generalist parasites that associates species of the
Leuciscinae and Alburninae (((D. similis, D. alatus) D. sphyr-na) D. vistulae). In all cases, parasites of the three host sub-families are monophyletic. Conversely, the Dactylogyrus spe-cies of the two species that are the most common hosts forDactylogyrus, L. cephalus and R. rutilus, do not form mono-phyletic groups.
Phylogeny of Cyprinid Species Using Cytochrome b
The alignment of cytochrome b sequences comprised 1140alignable positions, of which 545 were variable with 487parsimony informative. The selected model for ML analysiswas TVM 1 I 1 G with the following parameters: A-C 50.6912, A-G 5 8.1493, A-T 5 0.4777, C-G 5 0.3696, C-T5 8.1493, G-T 5 1.0000; the proportion of invariable sites5 0.4887; and rate heterogeneity approximated by a gammadistribution (four rate categories), a 5 0.8008. The best MLtree is reported in Figure 3. Whereas the monophyly of theCyprinidae is well supported, the monophyly of the differenthost subfamilies is moderately supported. The only repre-sentative of Alburninae is nested within Leuciscinae. TheGobioninae and Rasborinae are sister subfamilies, but onlyone species from these two subfamilies was examined. Fi-nally, monophyly of the Cyprininae was not found, but thedivergence of this subfamily appears to be the most basalevent within the Cyprinidae. NJ using the parameters selectedby ModelTest was also performed. Three equally parsimo-nious trees with 2909 steps were retained (CI 5 0.331, R 50.429), but no statistically significant differences were foundamong the topologies of the trees obtained from ML, NJ, andMP analyses (Shimodaira-Hasegawa test implemented inPAUP* 4b10, P . 0.05), and BP values obtained from ML,NJ, and MP analyses are reported in Figure 3.
Mapping Host Subfamilies onto the ParasitePhylogenetic Tree
The fish subfamilies were mapped onto the parasite phy-logeny inferred from the analysis of the combined dataset(18S rDNA and ITS1). This mapping was done with theassumption that the first Dactylogyrus group has the mostbasal divergence, following the results obtained from theanalysis of combined 18S 1 ITS1 using a midpoint rootingtechnique.
However, following the results using only partial 18SrDNA, where lineages 1 and 2 are not well supported, threehypotheses should be mentioned when considering which lin-eage is basal. When groups 1 (Fig. 4) or 2 (not shown) areconsidered as the most basal taxa, the Cyprininae is hypoth-esized to be a plesiomorphic host subfamily for Dactylogyrus.In that case, the Percidae would have been secondarily col-onized by Dactylogyrus species. However, when group 3 (notshown) is considered as the most basal taxon, the plesiom-orphic host subfamily is equivocal. It could be representativesof the Cyprininae, Leuciscinae, or Percidae. In all cases, par-asites of B. barbus (Cyprininae) and of A. alburnus (Albur-ninae) are derived from host fish of the Leuciscinae.
Comparison of Host and Parasite Phylogenies
Although phylogenetic relationships are not fully resolvedfor the parasites, several cases of parasite duplication within

1009MOLECULAR PHYLOGENY OF DACTYLOGYRUS
FIG. 2. Neighbor-joining tree based on Tamura-Nei distances, including one proportion of invariable characters and gamma distribution,inferred from analysis of combined partial 18S rDNA and ITS1 sequences of 51 Dactylogyrus species. Numbers along branches indicatebootstrap percentages resulting from different analyses in the order: neighbor joining/maximum parsimony/maximum likelihood. Valueslower than 50 are indicated with dashes.

1010 ANDREA SIMKOVA ET AL.
FIG. 3. Maximum-likelihood tree inferred from analysis of cytochrome b sequences of 18 representatives of the Cyprinidae, one of theBalitoridae, one of the Characidae, six of the Percidae, and one of the Salmonidae. Selected model was TVM 1 I 1 G. Numbers alongbranches indicate bootstrap percentages resulting from different analyses in the order: maximum likelihood/maximum parsimony/neighborjoining. Values lower than 50 are indicated with dashes.
host species (i.e., intrahost speciation) can be recognized(Fig. 5). Dactylogyrus duplications were found within thehost species A. alburnus, which is infested by three specialistparasites; within S. erythrophthalmus, which is also infestedby three specialist parasites; within C. nasus, which is in-fested by two specialists; within A. bjoerkna for the threespecialist parasites D. cornoides, D. cornu, and D. distin-guendus; within L. cephalus for the two specialists D. na-noides and D. folkmanovae; within A. brama infested by twospecialists D. zandti and D. wunderi; within B. barbus infestedby two specialists; and within G. cernuus, which is infested
by two specialist parasites. Those duplications were sup-ported by BP of more than 50 except those found within S.erythrophthalmus, A. brama, and B. barbus (see Figs. 2, 5).Two duplication events can be recognized within R. rutilus.The first one is characterized by the two specialist parasitesD. rutili and D. nanus (BP . 70 from all analyses) and thegeneralist parasite D. fallax (BP , 50), and the second oneby the specialist D. caballeroi and the generalist D. crucifer(BP . 90 from all analyses), the two generalist species in-festing predominantly R. rutilus. All those duplication eventsoccurred within lineage 3. Similarly, some duplication events

1011MOLECULAR PHYLOGENY OF DACTYLOGYRUS
FIG. 4. Mapping of host subfamilies onto the parasite neighbor-joining tree inferred from analysis of combined data. The first Dactylogyrusgroup is considered as the most basal lineage within Dactylogyrus following the most likely scenario of Dactylogyrus evolution.
were also reported within lineage 1. Dactylogyrus duplica-tions were found within the host species C. carpio (BP .60), which is infested by two specialist parasites, and withinC. auratus for which two duplication events were reported.The first gave rise to two specialist species (D. vastator andD. intermedius, BP 5 100) and the second (BP . 95) to threespecialist parasites and one generalist, that is, D. anchoratusalso infests C. carpio. No duplication event was reportedwithin lineage 2.
Besides intrahost duplication events, several cospeciationevents (numbers in Fig. 5) can also be recognized from thecomparison of host and parasite phylogenies. In lineage 2,the two sister species D. finitimus and D. cryptomeres para-sitize two congeneric host species, G. albipinatus and G. go-bio respectively (a in Fig. 5). Moreover, Dactylogyrus ofGobio species are a sister group to D. squameus of P. parva(b in Fig. 5). Because Gobio species and P. parva form amonophyletic group, cospeciation events can be suggestedto explain similarities in host and parasite relationships. Sim-ilarly, the parasite species infesting A. alburnus from theAlburninae are closely related to Dactylogyrus species in-
festing S. erythrophthalmus from the Leuciscinae (lineage 3,c in Fig. 5), but this cospeciation is not supported by bootstrap(Fig. 2). Even if the Alburninae are recognized to be poly-phyletic and nested within Leuciscinae, A. alburnus is a sisterspecies of S. erythrophthalmus (Figs. 3, 5). Thus, it is likelythat these two groups of parasites cospeciated with their hostspecies and later diversified within hosts. Dactylogyrus pros-tae would have switched to L. cephalus before intrahost spe-ciation occurred on S. erythrophthalmus.
Finally, several host switches must be invoked to explaindiscordant host and parasite relationships. That is the case,for instance, for Dactylogyrus species parasitizing B. barbusthat are nested within the species of the Leuciscinae.
Mapping of Niche Preference onto the Phylogenetic Tree
The preferred niche of each Dactylogyrus species wasmapped onto their phylogeny. For parasite species found onmore than one host species (D. sphyrna and D. crucifer), theposition on the fish species with the highest parasite abun-dance was chosen. This position, however, was similar to

1012 ANDREA SIMKOVA ET AL.
FIG. 5. Tanglegram of Dactylogyrus and cyprinid species deduced from comparison of the parasite neighbor-joining tree inferred fromanalysis of combined data (18SrDNA and ITS1 sequences) with the topology of a fish phylogeny that includes only species infested byDactylogyrus species. Gobio albipinatus was added to the fish phylogeny as a sister species of Gobio gobio considering that they aremonophyletic. Intrahost duplications are depicted on the trees. Dactylogyrus speciation by intrahost duplication supported by bootstrappercentage greater than 50 are given in bold. Potential coevolution events are indicated by a, b, and c.

1013MOLECULAR PHYLOGENY OF DACTYLOGYRUS
that observed when considering individuals of a given Dac-tylogyrus species obtained from more than one host species.
The first mapping shows the preferred position on the gillarch (Fig. 6B). The position on the second gill arch seemsto be ancestral and several changes toward the first, third,and fourth arches were noted. The evidence for strong chang-es in the arch position was found within the groups of par-asites on C. auratus. The clade including the specialist Dac-tylogyrus species parasitizing B. barbus, L. cephalus, S. er-ythrophthalmus, and A. alburnus shows a shift from the po-sition on the second to the third gill arch with changes towardfirst and second gill arches for parasites of S. erythrophthal-mus.
The preferred segment position on the gill arches wasmapped onto the phylogeny (Fig. 6C). The position on thedorsal segment is found to be the ancestral state with severalchanges toward the medial and ventral segments for the spe-cies in terminal positions. Changes in preferred segment po-sition were found between the branches within monophyleticgroups of the lineages of Dactylogyrus species parasitizingthe same host species.
The preferred area position on the gill arches was mappedonto the phylogeny (Fig. 6D). The position on the distal areais found to be the ancestral state with several changes towardthe central and proximal areas. The same observation as inthe case of preferred segment position was noted, that is, thechanges in preferred areas are found between the brancheswithin the group of Dactylogyrus species parasitizing thesame host species. However, when Dactylogyrus species ofone clade and parasitizing the same host (i.e., recognized asa case of intrahost duplication) were separated by their archposition, then the same segment or area positions are oftenfound in the phylogenetic tree. Looking at changes for allpositions, we can note that no sister groups have identicalniches and at least one of the niche parameters (arch, segment,or area) has changed.
DISCUSSION
The Dactylogyrus-Cyprinidae Model
In general, parasites with direct life cycle and free-livinginfectious stages attaching to the external surface of theirhosts are believed to have a narrow host range (i.e., they arespecialists) and to have coevolved with their hosts, in contrastto parasites with indirect life cycles (Littlewood et al. 1997;Paterson and Poulin 1999). Therefore, highly specific mono-geneans could be considered an appropriate model for study-ing the evolution of host-parasite associations to investigatepatterns and processes of parasite speciation. Nevertheless,different levels of correspondence between patterns of par-asite speciation and host specificity have been shown acrossdifferent models of congeneric monogenean parasites (Lit-tlewood et al. 1997; Bentz et al. 2001; Desdevises et al. 2002;Huyse and Volckaert 2002), suggesting importance of thehost species in the process of parasite diversification.
When considering that evolution of freshwater fish is moreconstrained than that of marine fish (i.e., it is correlated tothe history of freshwater movements and dispersals; Tsigen-opoulos and Berrebi 2000), then freshwater fish may repre-sent more suitable models for studying host-parasite coevo-
lutionary patterns (e.g., case study of marine monogeneanparasites by Desdevises et al. 2002). Therefore, we studiedhighly specific monogenean parasites of the genus Dactylo-gyrus that infest freshwater fish of the Cyprinidae. This studyrepresents a large parasite sampling from central Europeanfish.
Dactylogyrus Origin
On the basis of all phylogenetic analyses using partial 18SrDNA separately or combined 18S rDNA and ITS1 data, threemain lineages of Dactylogyrus species were recognized: (1)the clade of parasites of C. carpio and C. auratus, both rep-resentatives of the Cyprininae; (2) the lineage that includesparasite species of C. idella (Cyprininae), P. parva of theRasborinae (this species is considered as Gobioninae by Chenet al. 1984) and Gobio of the Gobioninae; and (3) the lineagethat includes parasite species of B. barbus (Cyprininae), Leu-ciscinae, Alburninae, and Percidae, all of European origin.Phylogenetic relationships between species within each ofthe three main lineages were better resolved when analyzingcombined data. The 18S rDNA represents a well-conservedgene that evolves relatively slowly (Hillis and Dixon 1991).This explains why it has been widely used for the study ofplathyhelminth relationships (Littlewood and Olson 2001;Olson and Littlewood 2002). However, fast-evolving se-quences, for instance the ITS1 region, could provide usefulphylogenetic information for resolving relationships withingroups of recent origin (Booton et al. 1999). Desdevises etal. (2002) showed in the case of monogenean species be-longing to Lamellodiscus that ITS1 sequences could not bealigned among species, which suggested an older age of thisgroup of parasites than hypothesized from a morphologicalpoint of view. It may also be the result of an increased sub-stitution rate in this genus. Our results inferred from analysisof ITS1 sequences suggest that Dactylogyrus is of quite recentorigin and that the three lineages separated from each otherin a very short period of time. This is confirmed by ouralignment of ITS1 sequences derived from all Dactylogyrusspecies and by the lack of basal resolution in the genus.
Plesiomorphic Host Subfamily
Results inferred from mapping host subfamily onto theparasite phylogenetic tree could suggest three scenarios con-cerning plesiomorphic hosts for Dactylogyrus. Evolutionaryscenarios considering either lineage 1 or 2 as displaying themost basal divergence, both support the hypothesis that Asi-atic Cyprininae could be the ancestral hosts. However, thescenario involving basal divergence of lineage 3 proposes anequivocal solution where either representatives of the Cy-prininae, Leuciscinae, or Percidae would be plesiomorphichosts. In the latter case, if we consider that Dactylogyrus firstevolved within percids, then it would imply that Dactylogyrusspecies became extinct within the Percidae, except in onespecies, G. cernuus, and at least in the Characidae and Bal-itoridae, which are the most basal taxa within Cypriniformes.Indeed, the majority of Dactylogyrus species infest represen-tatives of the Cyprinidae. For these reasons, we consider thispossibility very unlikely. The second solution is to considerthe Leuciscinae as the primitive host subfamily for Dactyl-

1014 ANDREA SIMKOVA ET AL.
FIG. 6. Mapping of niche within hosts preferred by Dactylogyrus species. (A) Gill arch division: gill segments: D, dorsal; M, medial;V, ventral; gill area: d, distal; c, central; p, proximal; gill surface: in, inner; out, outer; gill hemibranch: A, anterior; P, posterior. (B)Gill arch, (C) gill segment, and (D) gill area were mapped onto the parasite phylogeny.
ogyrus. In that case, it would imply that Dactylogyrus orig-inated in the ancestor of the Leuciscinae and secondarilycolonized the Percidae and Cyprininae. This scenario alsoseems unlikely if we consider that east-central Asia is thecenter of speciation of the Cyprininae. How then can weexplain host switches from European to Asian host fish,whereas the divergence of Asian cyprinins appears basalwithin the Cyprinidae (Fig. 3)? Indeed, C. auratus and C.carpio, the hosts for eight parasite species in our study, areconsidered to have originated in Asia and been recently im-ported in Europe. Furthermore, there is no paleontologicalevidence for their presence in Europe during the Pleistocene(Barus and Oliva 1995). However, it should be noted thatthe origin of different subfamilies of the Cyprinidae and dis-persal migration routes are still debated (Durand et al. 2002).When considering the above arguments and host subfamilymapping, we propose that Dactylogyrus originated within theCyprininae and secondarily colonized ancestral hosts of theLeuciscinae and several species of the Percidae. Concerning
the distribution of the latter family, that is, its absence inAfrica and eastern Asia, limited diversification in Europe,and extensive diversity in North America, it is very likelythat host transfer occurred in Europe from fish of the Cy-prininae.
Pattern and Process of Parasite Diversification
The nested position of Dactylogyrus species parasitizingB. barbus within the species of the Leuciscinae suggests hostswitching from host fish of the Leuciscinae. It has been pro-posed that Western Palearctic Barbus would have originatedin eastern Asia and migrated to Europe and Asia during theMiocene–Upper Oligocene (Tsigenopoulos and Berrebi2000). However, many species belonging to the genus Barbusare of European origin (Cunha et al. 2002). It has also beenshown that Barbus divergence is of the same order as theradiation of the Leuciscinae genera and that both are con-sidered to have their center of speciation in Siberia (Durand

1015MOLECULAR PHYLOGENY OF DACTYLOGYRUS
FIG. 6. Continued.
et al. 2002). The biogeography of this species could explainwhen this capture of parasites has occurred. Whether nu-merous host switches can be inferred from the comparisonof host and parasite relationships (Fig. 5), their interpretationswould at the moment be too speculative. However, we canassume that parasites of B. barbus of the Cyprininae and ofA. alburnus of the Alburninae are derived from parasites ofLeuciscinae host fish.
Although several nodes are not well supported, the mo-lecular phylogeny shows a consistent pattern of relationshipsof Dactylogyrus species, as well as a very high number ofintrahost speciation events (i.e., parasite duplications), com-pared to very few cospeciation events. Eight duplicationevents were recognized within lineage 3 across seven hostspecies and three within lineage 1 across two host species.These speciation events gave rise mainly to specialist species,that is, parasite species that infest a single host species. How-ever, there are several exceptions where two duplicationevents gave rise to both specialist and generalist parasitespecies. Within the first parasite clade of R. rutilus (D. ca-
balleroi, D. crucifer, and D. rarissimus), D. crucifer mainlyinfests R. rutilus and occasionally L. idus and S. erythro-phthalmus. Within the second parasite clade of R. rutilus (D.rutili, D. nanus, and D. fallax), D. fallax has been more oftenreported from R. rutilus than from L. cephalus. Within theparasite clade of C. auratus (D. formosus and D. anchoratus),D. anchoratus has also been reported from C. carpio. How-ever, because of low sampling across the two latter host spe-cies, we are currently unable to know which of these twohost species is preferred for D. anchoratus. Nevertheless,sympatric speciation seems to be closely correlated to stricthost specificity as suggested by Vickery and Poulin (1998).Our observations suggest that the majority of Dactylogyrusparasites first speciated by intrahost duplication and second-arily colonized new host species that are closely related. Thisconfirms the suggestion of Gusev (1985) who defined so-called basic host species for many generalist species (hostspecies when Dactylogyrus species occurs in high abundance)and rare host species (other phylogenetically closely relatedspecies). Our results contradict the investigations of Guegan

1016 ANDREA SIMKOVA ET AL.
and Agnese (1991) on congeneric African cyprinids and theirDactylogyrus species. The comparison of Dactylogyrus phy-logeny based on morphological grounds with a host phylog-eny (with fish species belonging only to a single genus) basedon genetic markers revealed no intrahost speciation, but didsuggest cospeciation and host switching events (Guegan andAgnese 1991).
Parasite duplication is a coevolutionary scenario that hasbeen recently considered to explain host-parasite relation-ships (Paterson and Gray 1997; Johnson et al. 2003). Patersonand Poulin (1999) showed that intrahost speciation could bemore likely than cospeciation events within chondracanthidcopepods and their teleost hosts. They suggested that thelarge geographical distances among hosts could explain thispattern of speciation. However, in the case of Dactylogyrusspecies, intrahost speciation is observed within hosts that arenot geographically isolated but living in sympatry.
In the tree reported in Figure 3, four of the several gen-eralist species, D. alatus, D. similis, D. sphyrna, and D. vis-tulae, form a clade of large generalists with similar morpho-logical features (mainly strongly developed central hooks andsimilar shape of the third pair of marginal hooks, which dif-ferentiate this group from other Dactylogyrus species). Al-though two species (D. alatus and D. similis) were sequencedfrom only one host species, their presence on other host spe-cies was recorded during this study (D. alatus on L. idus andD. similis on L. cephalus were not sequenced due to the lownumber of parasite individuals collected from those hosts).According to Gusev (1985) and Moravec (2001), those spe-cies may parasitize a wide host range. Like the generalistsincluded among a majority of specialists from the molecularphylogeny (see above), those four generalist species also oc-cur in high abundance in one host species and only rarely onother host species. Because those species form a clade, itcould be hypothesized that the ancestral parasite was able tocolonize a wide range of hosts with consequent speciation ofnew species within one host species. New parasite speciespreferentially choose this host species and occasionally infestclosely related host species, maybe because of their largebody size. The presence of several large species could pre-clude species coexistence.
Molecular divergence among generalist Dactylogyrus spe-cies collected from the different host species was about 1%.This could suggest that there is low gene flow between dif-ferent populations of the same Dactylogyrus species parasit-izing different host species, which may potentially reduceinterbreeding among the different populations. However,such suggestion needs further investigation in the future.
When comparing the mode of Dactylogyrus species spe-ciation with that of Gyrodactylus (also a monogenean genuswith high number of parasite species), host switch was ac-cepted for Gyrodactylus, and adaptive radiation was sug-gested to be the consequence of host switch to a new family(Zietara and Lumme 2002). Those differences may be ex-plained by different reproductive strategy (viviparous Gy-rodactylus vs. oviparous Dactylogyrus). In the case of Gy-rodactylus species, only one specimen may be requiredachieve a successful switch to a new host if the host defensesystem is tolerated.
As Dactylogyrus species form groups of phylogenetically
closely related species within a host, it could be hypothesizedthat their mode of speciation (i.e., intrahost speciation) shouldbe closely related to the evolution of attachment organs (i.e.,opisthaptor) morphology and reproductive system, as sug-gested by Rohde (1989). However, the morphology of theattachment apparatus of the Dactylogyrus genus is complex,including central and marginal hooks and connective bars.The number of the morphological characters studied so farwould not allow for phylogenetic reconstruction based onmorphology when using a parasite sample as large as the 51species in the present study. The evolution of complex char-acters and the comparison of these with molecular evolutionrequire further studies in the future.
Gusev (1985) suggested that there are some morphologicalfeatures, mainly in the sclerotized parts of opisthaptor andreproductive organs, that differ between Dactylogyrus speciesof Cyprininae and Leuciscinae subfamilies, and some of thesemay be common for several Dactylogyrus species occurringon the same or closely related species. However, there is noevidence that morphological characters help determine thegroups within Dactylogyrus, in contrast to the genus Gyro-dactylus, where the protonephridial system and opisthaptorhave been used (Malmberg 1970). However, the morpholog-ical characters of the attachment apparatus and reproductiveorgans for Dactylogyrus species are sufficient for species dis-crimination, and there is no need to use the molecular markersfor facilitating species discrimination, as suggested Zietaraand Lumme (2002) for Gyrodactylus species.
Our findings on intrahost speciation are closely related tohypotheses explaining species coexistence. Coexistence ofDactylogyrus species on the same host is facilitated by dif-fering morphology of copulatory organs, or niche center dis-tances, which prevent competition and increase intraspecificmating contacts (Simkova et al. 2002). The present studyshows that parasites speciating within one host (intrahostspeciation) tend to occupy niches differing at least in oneparameter (gill arch, segment, or area). The same result wasobtained in a study of Dactylogyrus assemblages coexistingon one fish species (i.e., roach) by Simkova et al. (2000).However, molecular phylogeny indicates that Dactylogyrusspecies parasitizing roach do not form a monophyletic group.In that case, isolation by niche segregation is important forcongeneric species. Moreover, when different Dactylogyrusspecies occupy similar or closely situated niches, reproduc-tive barriers could also reinforce reproductive isolation be-tween congeners (Rohde and Hobbs 1986; Rohde 1991; Mor-and et al. 2002; Jarkovsky et al. 2004).
Finally, the mapping of niche position within the fish gillsindicates that there is an ancestral position (possibly a sitemore protected against water current), from which other po-sitions seem to be derived.
Conclusions
Phylogenetic relationships of Dactylogyrus species com-bined with host subfamily mapping suggest that the genusoriginated recently within fish of the Cyprininae. They wouldhave secondarily evolved by colonizing different host speciesof the subfamilies Rasborinae, Gobioninae, Leuciscinae, andAlburninae and the family Percidae by host switching and

1017MOLECULAR PHYLOGENY OF DACTYLOGYRUS
cospeciation. The main process of Dactylogyrus diversifica-tion corresponds to sympatric speciation (i.e., intrahost spe-ciation), which seems correlated to strict host specificity. Thiskind of speciation was previously very rarely recognized inhost-parasite associations and the model Dactylogyrus-Cy-prinidae presents the first case study where intrahost speci-ation has been recorded for monogenean parasites. Moreover,the present study shows that congeneric parasites speciatingwithin one host tend to occupy niches differing at least inone niche parameter.
ACKNOWLEDGMENTS
This study was supported by Research Project of the Mas-aryk University, Brno, project number J07/98:143100010.AS was founded by the Grant Agency of the Czech Republic,project number 524/03/P108. The stay of AS in France wassupported by a postdoctoral fellowship from the Ministerede la Recherche in France. We would like to thank M. On-drackova, R. Sonek, M. Novakova, and K. Korınkova of Mas-aryk University, Brno, the Czech Republic, for their help incollecting material; P. Jurajda from the Institute of VertebrateBiology, Brno, Czech Republic, in electrofishing. We arevery grateful to C. O. Cunningham from FRS Marine Lab-oratory, Aberdeen, Scotland, for correction of English; J.-F.Martin from Centre de Biologie et de Gestion des Popula-tions, Montferrier, France, and two reviewers for helpfulcomments.
LITERATURE CITED
Barus, V., and O. Oliva. 1995. Petromyzontes and Osteichthyes.Academy of Science of Czech Republic, Prague.
Bentz, S., S. Leroy, L. du Preez, J. Mariaux, C. Vaucher, and O.Verneau. 2001. Origin and evolution of African Polystoma(Monogenea: Polystomatidae) assessed by molecular methods.Int. J. Parasitol. 31:697–705.
Booton, G. C., L. Kaufman, M. Chandler, R. Oguto-Ohwayo, W.Duand, and P. A. Fuerst. 1999. Evolution of the ribosomal RNAInternal transcribed spacer one (ITS-1) in cichlid fishes of theLake Victoria region. Mol. Phylogenet. Evol. 11:273–282.
Briolay, J., N. Galtier, R. M. Brito, and Y. Bouvet. 1998. Molecularphylogeny of Cyprinidae inferred from cytochrome b DNA se-quences. Mol. Phylogenet. Evol. 9:100–108.
Brooks, D. R., and D. A. McLennan. 1993. Parascript: parasitesand the language of evolution. Smithsonian Institution Press,Washington, DC.
Chen, X. L., P. Q. Yue, and R. D. Lin. 1984. Major groups withinthe family Cyprinidae and their phylogenetic relationships. ActaZootaxonomica Sin. 9:424–498.
Cone, D., T. Eurell, and V. Beasley. 1994. A report of Dactylogyrusamphibothrium (Monogenea) on the gills of European ruffe inwestern Lake Superior. J. Parasitol. 80(3):476–478.
Coyne, J. A., and T. D. Price. 2000. Little evidence for sympatricspeciation in island birds. Evolution 54:2166–2171.
Cunha, C., N. Mesquita, T. E. Dowling, A. Gilles, and M. M. Coel-ho. 2002. Phylogenetic relationships of Eurasian and Americancyprinids using cytochrome b sequences. J. Fish Biol. 61:929–944.
Desdevises, Y., S. Morand, O. Jousson, and P. Legendre. 2002.Coevolution between Lamellodiscus (Monogenea: Diplectani-dae) and Sparidae (Teleostei): the study of a complex host-par-asite system. Evolution 56:2459–2471.
Dieckmann, U., and M. Doebeli. 1999. On the origin of species bysympatric speciation. Nature 400:354–357.
Durand, J.-D., C. S. Tsigenopoulos, E. Unlu, and P. Berrebi. 2002.Phylogeny and biogeography of the family Cyprinidae in the
Middle East inferred from cytochrome b DNA: evolutionarysignificance of this region. Mol. Phylogenet. Evol. 22:91–100.
El Gharbi, S., E. Birgi, and A. Lambert. 1994. Monogenes Dac-tylogyridae parasites de Cyprinidae du genre Barbus d’AfriqueNord. Syst. Parasitol. 27:45–70.
Ergens, R., and J. Lom. 1970. Causative agents of fish diseases.(In Czech.) Academia Praha, Prague.
Felsenstein, J. 1985. Confidence limits on phylogenies: an approachusing the bootstrap. Evolution 39:783–791.
Gibson, D. I., T. A. Timofeeva, and P. I. Gerasev. 1996. A catalogueof the nominal species of the monogenean genus DactylogyrusDiesing, 1850 and their host genera. Syst. Parasitol. 35:3–48.
Gilles, A., G. Lecointre, E. Faure, R. Chappaz, and G. Brun. 1998.Mitochondrial phylogeny of the European cyprinids: implica-tions for their systematics, reticulate evolution, and colonizationtime. Mol. Phylogenet. Evol. 10:132–143.
Gilles, A., G. Lecointre, A. Miquelis, M. Loerstcher, R. Chappaz,and G. Brun. 2001. Partial combination applied to phylogeny ofEuropean cyprinids using the mitochondrial control region. Mol.Phylogenet. Evol. 19:22–33.
Guegan, J.-F., and J.-F. Agnese. 1991. Parasite evolutionary eventsinferred from host phylogeny: the case of Labeo species (Te-leostei, Cyprinidae) and their dactylogyrid parasites (Monoge-nea, Dactylogyridae). Can. J. Zool. 69:595–603.
Guegan, J.-F., and A. Lambert. 1991. Dactylogyrids (Platyhelmin-thes: Monogenea) of Labeo (Teleostei: Cyprinidae) from WestAfrican coastal rivers. J. Helminthol. Soc. W. 58:85–99.
Gusev, A. V. 1985. Metazoan parasites. I. Identification key toparasites of freshwater Fish. Vol. 2. O. N. Bauer, ed. Nauka,Leningrad. (In Russian)
Hafner, M. S., and S. A. Nadler. 1988. Phylogenetic trees supportthe coevolution of parasites and their hosts. Nature 332:258–259.
Hafner, M. S., P. D. Sudman, F. X. Villablanca, T. A. Spradling,J. W. Demestes, and S. A. Nadler. 1994. Disparate rates ofmolecular evolution in cospeciating hosts and parasites. Science265:1087–1090.
Hayward, C. J. 1997. Helminth ectoparasites of sillaginid fishes(Perciformes: Percoidei) have low species richness. Folia Par-asitol. 44(3):173–187.
Helfman, G. S., B. B. Collette, and D. E. Facey. 1997. The diversityof fishes. Blackwell Science, Madlen, MA.
Hillis, D. M., and M. T. Dixon. 1991. Ribosomal DNA: molecularevolution and phylogenetic inference. Q. Rev. Biol. 66:411–453.
Huyse, T., and A. M. Volckaert. 2002. Tangled trees: comparingthe phylogenies of Gyrodactylus and its goby host. Pp. 157–175in T. Huyse, ed. Evolutionary associations between Gyrodactylusand its goby host: Bound forever? Ph.D. diss., Katholieke Univ-ersiteit Leuven, Belgium.
Jarkovsky, J., S. Morand, A. Simkova, and M. Gelnar. 2004. Re-productive barriers between congeneric monogenean parasites(Dactylogyrus: Monogenea): attachment apparatus morphologyor copulatory organ incompatibility? Parasitol. Res. 92:95–105.
Johnson, K. P., R. J. Adams, R. D. M. Page, and D. H. Clayton.2003. When do parasite fail to speciate in response to host spe-ciation? Syst. Biol. 52:37–47.
Kennedy, C. R., and A. O. Bush. 1992. Species richness in helminthcommunities: the importance of multiple congeners. Parasitol-ogy 104:189–197.
Kimura, M. 1980. A simple method for estimating evolutionary rateof base substitutions through comparative studies of nucleotidesequences. J. Mol. Evol. 16:111–120.
Klassen, G. J., and M. Beverley-Burton. 1987. Phylogenetic rela-tionship of Ligictaluridus spp. (Monogenea: Ancyrocephalidae)and their ictalurid (Siluriformes) hosts: an hypothesis. Proc. Hel-minthol. Soc. Wash. 54:84–90.
Lambert, A., and S. El Gharbi. 1995. Monogenean host specificityas a biological and taxonomic indicator for fish. Biol. Conserv.72:227–235.
Li, W.-H. 1997. Molecular evolution. Sinauer Associates, Sunder-land, MA.
Littlewood, D. T. J., and P. D. Olson. 2001. Small subunit rDNAand the Platyhelminthes: signal, noise, conflict and compromise.Pp. 262–278 in D. T. J. Littlewood and R. A. Bray, eds. Inter-

1018 ANDREA SIMKOVA ET AL.
relationships of the Platyhelminthes. Taylor and Francis, Lon-don.
Littlewood, D. T. J., K. Rohde, and K. A. Clough. 1997. Parasitespeciation within or between host species? Phylogenetic evi-dence from site-specific polystome Monogeneans. Int. J. Par-asitol. 27:1289–1297.
Maddison, W. P., and D. R. Maddison. 1992. MacClade: analysisof phylogeny and character evolution. Sinauer Associates, Sun-derland, MA.
Malmberg, G. 1970. The excretory systems and the marginal hooksas a basis for the systematics of Gyrodactylus (Trematoda, Mono-genea). Ark. Zool. 23:1–235.
Morand, S., A. Simkova, I. Matejusova, L. Plaisance, O. Verneau,and Y. Desdevises. 2002. Investigating patterns may reveal pro-cesses: evolutionary ecology of monogeneans. Int. J. Parasitol.32:111–119.
Moravec, F. 2001. Checklist of the Metazoan parasites of fishes ofthe Czech Republic and the Slovak Republic. Academia Praha,Prague.
Nei, M., and S. Kumar. 2000. Molecular evolution and phyloge-netics. Oxford Univ. Press, New York.
Nelson, J. S. 1994. Fishes of the world. John Wiley & Sons, NewYork.
Olson, P. D., and D. T. J. Littlewood. 2002. Phylogenetics of theMonogenea: evidence from a medley of molecules. Int. J. Par-asitol. 32:233–244.
Page, R. D. M. 1994. Parallel phylogenies: reconstructing the his-tory of host-parasite assemblages. Cladistics 10:155–173.
Page, R. D. M., P. L. M. Lee, S. A. Becher, R. Griffiths, and D.H. Clayton. 1998. A different tempo of mitochondrial DNA evo-lution in birds and their parasitic lice. Mol. Phylogenet. Evol.9:276–293.
Paterson, A. M., and J. Banks. 2001. Analytical approaches to mea-suring cospeciation of host and parasites: through a glass, darkly.Int. J. Parasitol. 31:1012–1022.
Paterson, A. M., and R. D. Gray. 1997. Host-parasite co-speciation,host switching and missing the boat. Pp. 236–250 in D. H. Clay-ton and J. Moore, eds. Host-parasite evolution: general principlesand avian models. Oxford Univ. Press, Oxford, U.K.
Paterson, A. M., and R. Poulin. 1999. Have chondracanthid cope-pods co-speciated with their hosts? Syst. Parasitol. 44:79–85.
Philippe, H. 1993. MUST, a computer package for managementutilitarians for sequences and trees. Nucleic Acids Res. 21:5264–5272.
Posada, D., and K. A. Crandall. 1998. Modeltest: testing the modelof DNA substitution. Bioinformatics 14:817–818.
Poulin, R. 2002. The evolution of monogenean diversity. Int. J.Parasitol. 32:245–254.
Rohde, K. 1977. A non-competitive mechanism responsible for re-stricting niches. Zool. Anz. 199:164–172.
———. 1979. A critical evaluation of intrinsic and extrinsic factorsresponsible for niche restriction in parasites. Am. Nat. 114:648–671.
———. 1989. Simple ecological systems, simple solutions to com-plex problems? Evol. Theor. 8:305–350.
———. 1991. Intra- and interspecific interactions in low densitypopulations in resource-rich habitats. Oikos 60:91–104.
Rohde, K., and R. Hobbs. 1986. Species segregation: competition
or reinforcement of reproductive barriers? Pp. 189–199 in M.Cremin, C. Dobson, and E. Noorhouse, eds. Parasites lives, pa-pers on parasites, their hosts and their association to Honour.JFA Sprent, University of Queensland Press, St. Lucia.
Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular clon-ing: a laboratory manual. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY.
Sekar, V. 1987. A rapid screening procedure for the identificationof recombinant bacterial clones. Biotechniques 5:11–13.
Simkova, A., Y. Desdevises, M. Gelnar, and S. Morand. 2000. Co-existence of nine gill ectoparasites (Dactylogyrus: Monogenea)parasitizing the roach (Rutilus rutilus L.): history and presentecology. Int. J. Parasitol. 30:1077–1088.
———. 2001. Morphometric correlates of host specificity in Dac-tylogyrus species (Monogenea) parasites of European cyprinidfish. Parasitology 123:169–177.
Simkova, A., M. Ondrackova, M. Gelnar, and S. Morand. 2002.Morphology and coexistence of congeneric ectoparasite species:reinforcement of reproductive isolation? Biol. J. Linn. Soc. 76:125–135.
Simkova, A., L. Plaisance, I. Matejusova, S. Morand, and O. Ver-neau. 2003. Phylogenetic relationships of the DactylogyridaeBychowsky, 1933 (Monogenea: Dactylogyridea): the need forthe systematic revision of the Ancyrocephalinae Bychowsky,1937. Syst. Parasitol. 54:1–11.
Sinnappah, N. D., L. H. S. Lim, K. Rohde, R. Tinsley, C. Combes,and O. Verneau. 2001. A paedomorphic parasite associated witha neotenic amphibian host: phylogenetic evidence suggests arevised systematic position for Sphyranuridae within anuran andturtle polystomatoineans. Mol. Phylogenet. Evol. 18:189–201.
Swofford, D. L. 2002. PAUP*: phylogenetic analysis using parsi-mony and other methods. Version 4.0b10. Sinauer Associates,Sunderland, MA.
Tsigenopoulos, C. S., and P. Berrebi. 2000. Molecular phylogenyof north Mediterranean freshwater barbs (genus Barbus: Cypri-nidae) inferred from cytochrome b sequences: biogeographic andsystematic implications. Mol. Phylogenet. Evol. 14:165–179.
Valtonen, E. T., M. Prot, and R. Rahkonen. 1990. Seasonality oftwo monogeneans from two freshwater fish from an oligotrophiclake in northeast Finland. Int. J. Parasitol. 20:101–107.
Vickery, W. L., and R. Poulin. 1998. Parasite extinction and col-onisation and the evolution of parasite communities: a simulationstudy. Int. J. Parasitol. 28:727–737.
Winfield, I. J., and J. S. Nelson. 1991. Cyprinid fishes: Systematics,biology and exploitation. Fish and Fisheries Series no. 3. Chap-man and Hall, London.
Zardoya, R., and I. Doadrio. 1999. Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprinids.J. Mol. Evol. 49:227–237.
Zardoya, R., P. S. Economidis, and I. Doadrio. 1999. Phylogeneticrelationships of Greek Cyprinidae: molecular evidence for atleast two origins of the Greek cyprinid fauna. Mol. Phylogenet.Evol. 13:122–131.
Zietara, M. S., and J. Lumme. 2002. Speciation by host switch andadaptive radiation in a fish parasite genus Gyrodactylus (Mono-genea, Gyrodactylidae). Evolution 56:2445–2458.
Corresponding Editor: R. Poulin




















