
effect of parasite infestation on the - University Of Nigeria ...

12 Springer
Dear Author: Please find attached the final pdf file of your contribution, which can be viewed using the Acrobat Reader, version 3.0 or higher. We would kindly like to draw your attention to the fact that copyright law is also valid for electronic products. This means especially that: • You may not alter the pdf file, as changes to the published contribution are
prohibited by copyright law.
• You may print the file and distribute it amongst your colleagues in the scientific community for scientific and/or personal use.
• You may make your article published by Springer-Verlag available on your personal home page provided the source of the published article is cited and Springer-Verlag is mentioned as copyright holder. You are requested to create a link to the published article in Springer's internet service. The link must be accompanied by the following text: The original publication is available at http://link.springer.de or at http://link.springer-ny.com . Please use the appropriate URL and/or DOI for the article. Articles disseminated via SpringerLink are indexed, abstracted and referenced by many abstracting and information services, bibliographic networks, subscription agencies, library networks and consortia.
• Without having asked Springer-Verlag for a separate permission your institute/your company is not allowed to place this file on its homepage.
• Please address any queries to the production editor of the journal in question, giving your name, the journal title, volume and first page number.
Yours sincerely, Springer-Verlag

ORIGINAL PAPER
Molecular diagnosis of Chagas’ disease and use of an animalmodel to study parasite tropism
Published online: 16 January 2003� Springer-Verlag 2003
Abstract Chagas’ disease, which is an important healthproblem in humans, is caused by the protozoan Try-panosoma cruzi. The cellular and molecular mechanisms,involved in the selective tropism of T. cruzi to differentorgans remain largely unknown. In this study we de-signed a PCR-based molecular diagnosis method in or-der to study the tropism and growth kinetics of T. cruziin a murine model infected with parasites isolated froman endemic area of Mexico. The growth kinetics andparasite tropism of T. cruzi were also evaluated in theblood and other tissues. We observed that T. cruzi iso-lates from the Western Mexico showed a major tropismto mouse heart and skeletal muscles in this murinemodel.
Introduction
Chagas’ disease in humans is caused by the protozoanTrypanosoma cruzi. It is a major health problem inCentral and South America. Chagas’ disease affectsmainly rural areas, and is usually associated with con-ditions of poverty, poor sanitation and drainage(Velasco et al. 1991; Pinto et al. 1996). It affects 16–18million people in 17 countries and approximately 80–100million people are exposed to the infection (Pinto et al.1996). Infection with T. cruzi may occur as an acute orchronic phase. The acute phase is generally asymptom-
atic with the presence of parasites in the blood. In thechronic phase, digestive and cardiac alterations can de-velop, with fatal consequences as a result of progressivedestruction of the invaded organ. Morbidity, which hasan important socioeconomic impact, is also common inthe chronic phase (Velasco et al. 1991).T. cruzi is aparasite closely related to other species of trypanosom-atides (Leshmania, and Trypanosoma rangeli), which arevery variable in their clinical manifestation, possessinga great capacity for infection and variable virulence.T. cruzi genetic variability may be a determinant ofdifferential tissue tropism and clinical forms of the dis-ease (Macedo and Pena 1998). The diagnosis of Chagas’disease is difficult, due to the low number of parasites inthe circulating blood. During the acute phase, diagnosisis carried out by microscopic detection of parasites in theblood or by the presence of antibodies (Kirchhoffet al.1996). However, in the chronic phase, parasites arescarce and are detected only by indirect and time-con-suming methods such as xenodiagnosis and hemoculture(Pinto et al.1996), which both have low sensitivity andspecificity (70%) (Krautz et al.1996). This fact is in partdue to cross-reactions with antigens from other parasites(Kirchhoff et al.1996). Engman et al. (1989) havedetected a 24-kDa T. cruzi flagellar protein (Tc-24) thathas been identified as an antigenic, highly expressedprotein evoking a strong humoral immune response inhumans and experimental animals (Piras et al. 1988).New techniques using blood samples have been devel-oped in order to detect T. cruzi in all developmentstages, particularly in the vertebrate infective stage(trypomastigote) (Kirchhoff et al. 1996). Some investi-gators have demonstrated high efficiency and sensitivityin the detection of T. cruzi by polymerase chain reaction(PCR) (Piras et al. 1988; Britto et al. 1995; Guevara et al.1996). However, primer design represents a critical step,since T. cruzi presents cross-reactivity with other similarspecies. The detection of the gene encoding for the Tc-24protein by PCR is more sensitive than the detection ofother parasite sequences (Guevara et al. 1996). Also, theidentification of the TcZ sequence, corresponding to the
Parasitol Res (2003) 89: 480–486DOI 10.1007/s00436-002-0787-0
J.M. Vera-Cruz Æ E. Magallon-Gastelum
G. Grijalva Æ A.R. Rincon Æ C. Ramos-Garcıa
J. Armendariz-Borunda
J.M. Vera-Cruz Æ E. Magallon-Gastelum Æ G. GrijalvaA.R. Rincon Æ J. Armendariz-Borunda (&)Institute for Molecular Biology in Medicine and Gene Therapy,CUCS, University of Guadalajara, Apdo. Postal 2-123, 44281,Guadalajara, Jalisco, MexicoE-mail: [email protected].: +52-33-36174159Fax: +52-33-36174159
C. Ramos-GarcıaInstituto Nacional de Salud Publica,Cuernavaca, Morelos, Mexico

highly variable region of the kinetoplast minicircle of T.cruzi, has also been reported to be useful in the identi-fication of different natural clones (Hart et al. 1981).Based on the above studies, we establish a moleculardiagnosis methodology based on PCR amplification ofthe Tc-24 gene. Another major aim of this work was tostudy the tropism and growth kinetics of T. cruzi in amurine model infected with parasites isolated from anendemic area of Mexico. In addition, we compared PCRamplification with ELISA immunoassay as a way toidentify the presence of the parasite Tc-24 gene ininhabitants of the same endemic zone.
Materials and methods
Parasites and infection in Balb/C mice
Isolates (JALGO) of T. cruzi, were obtained from the feces of tri-atomines collected in San Martin Hidalgo, Jalisco, Mexico. Feceswere collected by pressing the posterior of the stomach of triato-mines and placed on a slide. Samples were considered positive if asingle flagellated parasite was observed by microscopic analysis.Feces were resuspended in 1 ml of 0.9% saline solution and inoc-ulated intraperitoneally into 20 g Balb/C mice. These animals werethen used as reservoirs of parasites. Blood was drawn from the miceto further increase the parasite number by direct inoculation ofinfected blood into naive Balb/C mice to obtain viable trypom-astigotes. Briefly, 1·105 T. cruzi trypomastigotes from each infectedmouse were inoculated intraperitoneally (i.p) into 6-week-old Balb/c mice (n=20). These 20 animals were used to determine survival.Those that survived were killed at the end of the study (360 days).During the course of the infection we also determined mortalityduring the acute and chronic phases.
Kinetic curves and tropism analysis
Another 30 mice (6 weeks old) were inoculated i.p. with 1· 105
trypomastigotes, and blood samples were obtained from the mousetail vein every 3rd day until parasites were microscopically ob-served. Then, the number of parasites per ml of blood was deter-mined in a hemocytometer (Hart et al. 1981) to obtain the growthkinetics. For tropism analysis, mice were killed at 4, 15, 20,28 and120 days postinfection (PI) and their organs were observedmacroscopically. Also, DNA was isolated from the blood anddifferent tissues (heart, liver, kidney, duodenum, ileum, spleen,colon and skeletal muscle) using microanalysis methodology.
Histopathological analysis
Mice were killed on days 18 and 180 PI and tissue samples wereobtained from the liver, kidney, duodenum, ileum, spleen, colon,skeletal muscle and heart. The samples were fixed in formaldehyde(10%) in 100 mM phosphate buffer (pH 7.2). Tissue samples wereembedded in paraffin and stained with hematoxylin and eosin.
Extraction and purification of DNA
We standardized a micro-method for DNA extraction since onlysmall blood volumes could be obtained from the mouse tail vein. Atotal of 50 ll of heparinized blood and homogenized organ samplesfrom non-infected and infected mice were immediately mixed withfive volumes of lysis buffer (10 mM Tris-HCl pH 7.6, 10 mMNaCl, 0.5% SDS and 300 lg of proteinase K). The mixture was
incubated at 50�C (heparinized blood for 2 h and ground organsfor 18 h). A total of 100 ll of a mix of phenol-chloroform-isoamylalcohol (25:24:1, v/v/v) was added to this mixture, which was thenvortexed for 1 min and spun for 4 min at 12,000 g. The aqueousphase was recovered and DNA was precipitated with ethanol andsodium acetate (3 M, pH 5.5), vortexed and centrifuged at 12,000 gfor 10 min. The pellet was washed with 70% ethanol and resus-pended in Tris-EDTA solution (pH 8.0) and stored at )70�C untilPCR was performed. Tissue samples were ground up in lysis bufferusing a Polyton homogeneizer (Tissue Tearor, model 985370,Bartlesville, Okla., USA).
Detection of T. cruzi by PCR
Detection of the DNA sequence encoding the Tc-24 protein of Tcruzi was used for the identification of the parasite in blood andtissues. Amplification of Tc-24 sequences was performed using oneset of primers for a first PCR as reported by Ouaissi et al (1992).The first PCR mix was prepared as follows: 5 ll of 10· Taqpolymerase reaction buffer, 2 ll of 2.5 mM dNTP’s solution, 3 llof 50 mM MgCl2 solution, 1 lg of DNA sample and 7 ll of eachpair of 3 lM oligonucleotides (sense primer 5¢GAC GGC AAGAAC GCC AAG GAC3¢ and antisense primer 5¢TCA CGC GCTCTC CGG CAC GTT GTC3¢), 1 IU of Taq DNA polymerase andwater up to 50 ll. We designed an internal primer sequence of Tc-24 to perform nested PCR to improve the specificity of the assay.This reaction was performed as above, using 2 ll of the first PCRamplification product and 7 ll of each pair of 3 lM oligonucleo-tides (sense 5¢ AAG AAG TTC GAC AAG AAC GA 3¢ and an-tisense 5¢ AAA CTC GTC GAA CGT CAC GG 3¢) in a 50 llreaction. The reaction mix was subjected at one cycle of 5 min at94�C and 35 cycles for the first PCR (94�C for 1 min, 62�C for1 min and 72�C for 2 min) and 25 cycles for nested PCR (94�C for1 min, 50�C for 1 min and 72�C for 1 min), and one cycle of 7 minat 72�C. The PCR was performed in a DNA thermal cycle (Perkin-Elmer, Calif.). PCR products of 550 bp and 336 bp for the first andnested PCR, respectively, were detected in 2% agarose gel elec-trophoresis. PCR products were stained with ethidium bromideand photographed. The intensities were evaluated by densitometricanalysis using Kodak Digital Science 1D software. All results wereconfirmed by three subsequent PCR analyses.
Collection of human blood samples
This study involved 39 people (30% males, 70% females with anage range of 14–35 years); all lived in San Martin Hidalgo (endemiczone of Jalisco for T. cruzi) and had been in contact with triato-mines in the past. Although 94.8% of the participants were as-ymptomatic for Chagas’ disease, two of them presented apathology of megaesophagus and cardiomegaly. Negative controlsera were obtained from ten healthy individuals (25–30 years)without a history of contact with the vector. Informed consent toparticipate in this study was obtained from both groups.
Extraction of DNA and PCR amplification
Human DNA blood samples were prepared and detection of theDNA sequence encoding for the Tc-24 protein of T. cruzi wasperformed in the same way as described for the mouse samples.
Detection of antibodies against T. cruzi
Circulating antibodies against T. cruzi were determined with acommercial kit kindly provided by Hemagen Diagnostics (Hema-gen Chagas’s Kit, Waltham, Mass., USA). Serum samples werediluted 1:26 and the procedure was carried out following themanufacturer’s instructions. Absorbance values were obtained byusing an ELISA plate reader (Dynatech, Alexandria, Va., USA) at
481

OD 450 nm. Samples in which the optical density was more than10% above the cut-off value were considered positive. The positivecut-off for assays was determined from the mean value±3 SD ofnegative values. All serum samples were analyzed in quadruplicate.
Results
T. cruzi kinetics of growth
The growth kinetics of T. cruzi (JALGO) in infectedmice is shown in Fig. 1A. Parasites were observed inthe blood of all mice (n=30) by the 7th day and werepresent up to the 45th day. The average number of
trypomastigotes increased exponentially by the 3rdweek. A 15% mortality (3/20) was observed during thecourse of 21 days, correlating with the maximum num-ber of parasites in the blood. By the end of the study(12 months) the mortality rate had reached 50% (10/20)(Fig. 1B). The JALGO isolate showed a mild virulenceagainst mice, which became stronger in the chronicphase. All mice displayed a ruffled hair coat and im-paired reflexes. Moreover, hindquarter paralysis andhumpy appearance were observed in the mice which diedduring the course of the study, but not in mice whichwere killed.
Morphological and histopathological analysisof T. cruzi infected mice
Abnormal growth was observed mostly in the spleen andcolon of mice (4 months PI) (Fig. 1C). Histopathologi-cal analyses demonstrated minor damage in all organs inthe acute phase (18th day) ( data not shown). Thisdamage was characterized by cellular infiltrates in allorgans. Furthermore, heart and skeletal muscle pre-sented clusters of amastigotes. After 4 months, chronicdamage was manifested along with a higher numberof inflammatory cells in all organs. The main changesobserved were macrophages with parasites in the spleen
Fig. 1A–D Biological characteristics of Trypanosoma cruzi. AParasite growth kinetics. Twenty Balb/C mice were intraperitonallyinoculated with 100,000 trypomastigotes and monitored for thepresence of T. cruzi in blood by PCR for up to 12 months. Parasiteswere observed in blood from 7th to the 43rd day. B Survival curve.Around the 21st day we observed a 15% mortality (3/20). At theend of the study, seven more deaths had occurred, reaching amortality rate of 50% (10/20). C Macroscopic analysis of infectedmice. Abnormal growth of colon and spleen in infected mice wasobserved (right photo) compared with uninfected animals (leftphoto). D Histopathological studies of different organs frominfected animals. Infected mice were killed on day 18 and at6 months after infection. Chronic damage included cell infiltrationin the colon and clusters of amastigotes in the heart and skeletalmuscle
482

and colon (Fig. 1D), kidneys with glomerulonephritis,ileum with ganglional cell necrosis of the mesentericplexus and steatosis was observed in the liver (data notshown). In the chronic phase, skeletal muscles presentedclusters of amastigotes around nerves and the necrosis ofmuscle fibers of the skeletal and heart muscle (myo-carditis) (Fig. 1D).
Detection of T. cruzi in blood by microscopic analysisand PCR amplification in infected mice
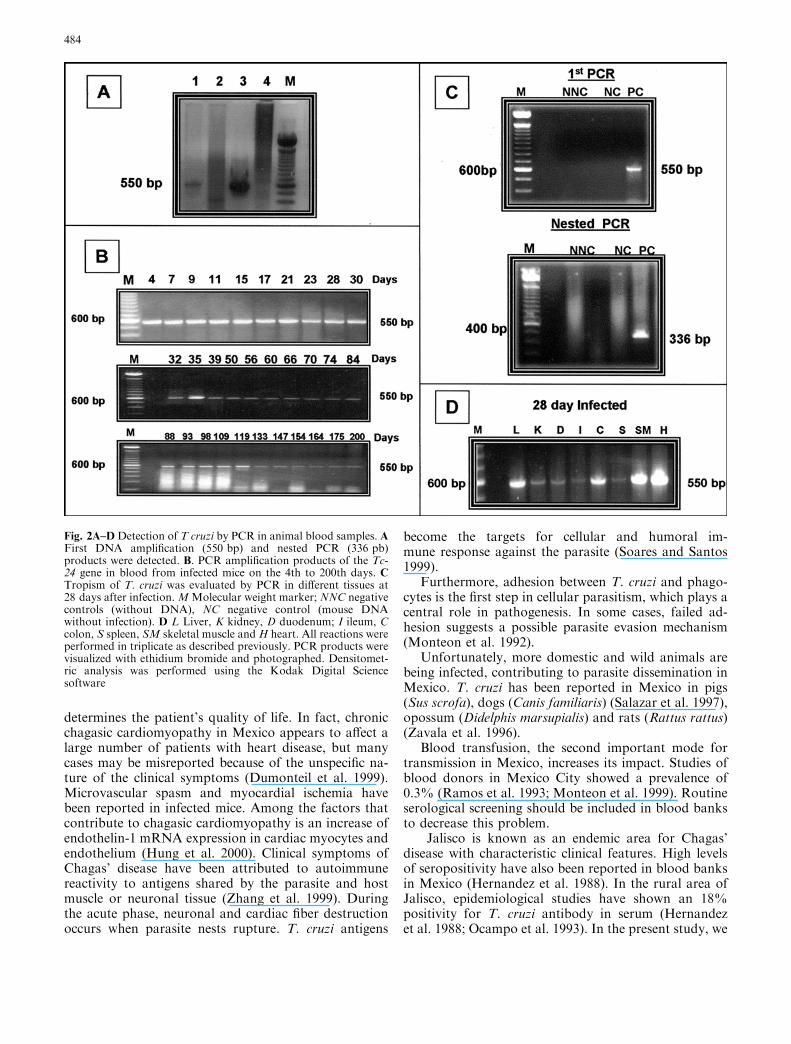
Mouse blood samples (n=30 mice) were analyzed bymicroscopic observation and parasites were detectedfrom the 7th day PI to the 39th day. On the other hand,parasite genomic DNA was detected throughout thestudy, beginning the 4th day and up to 200th day PI,demonstrating a higher sensitivity than microscopicobservation, (compare Figs. 1A and 2B). It is notewor-thy that the intensity of the PCR bands at day 21(Fig 2B) correlated with the peak observed in growthkinetic (Fig 1A). In order to improve and to verify thespecificity of our PCR detection methodology, we alsoemployed a nested PCR to the Tc-24 gene of T. cruziamplifying a 336 bp band which is also specific for theTc-24 gene (Fig. 2C, lower panel).
Parasite tropism in mice infected with T. cruzi
Whereas parasites were detected by PCR in the tissues atdifferent times, the growth kinetics showed the highestnumber of parasites and the PCR bands were mostclearly observed at 28 day PI (Fig. 2C). Skeletal muscleand heart had the highest intensity of PCR amplifiedproducts, suggesting the presence of large numbers ofparasites. These results were confirmed by histologicalanalysis for the presence of clusters of amastigotes. Al-though parasites were present in other organs, they weremostly localized within macrophages and no extensivetissue damage was observed in comparison with heartand skeletal muscle.
To rule out the possibility of blood contamination,we perfused the tissues with saline solution before DNAextraction. Comparison of PCR products from perfusedand non-perfused mouse tissues showed the same re-sults, confirming that only skeletal and heart muscle hadintracellular amastigotes (data not shown).
Detection of T. cruzi by PCR in infected human blood
A total of 39 blood samples from individuals living in anendemic area of Mexico were collected and subjected toPCR amplification and immunoassay analysis in orderto detect the presence of T. cruzi. All of the tests wereperformed in triplicate and positive and negative con-trols were included. In order to evaluate the specificity ofPCR detection of T. cruzi DNA in human samples, we
collected blood samples from ten normal subjects, whohad never been in contact with the vector, to be used asnegative controls. These controls were negative for thePCR amplification products tested (Fig. 3A). A repre-sentative PCR of the blood of inhabitants of the en-demic area is shown in Fig. 3B. As a positive control weincluded blood from infected mice. Also, a nested PCRwas used to demonstrate the specificity to PCR with theTc-24 gene of T. cruzi (336 bp) (Fig. 3C).
Comparison of ELISA and PCR in detectingT. cruzi in human blood
When PCR and immunological assays were compared,we observed that 21 of 39 were positive by ELISA(53.8%) and 33 of 39 were positive by PCR (84.6%).More importantly, we extended our results since 17 outof 18 seronegatives for ELISA were proved to be posi-tive when tested by PCR. On the other hand, 16 patientsout of 21 who had been positive by ELISA were alsoPCR-positive and the five remaining were PCR negative.Based on these results, the ELISA immunoassay usedshowed only 48% sensitivity and 17% specificity for thedetection of antibodies to T. cruzi in infected people. It isclear then, that the PCR reaction could be considered asa ‘‘gold standard’’ procedure for monitoring T. cuzichronic infections in humans.
Discussion
The genetic and biological characteristics of NorthAmerican isolates of T. cruzi have not been completelystudied. Recently, 17 Mexican isolates were character-ized by RFLP analysis using an rRNA gene spacer as amolecular probe and HinfI digestion patterns of totalDNA (Zavala et al. 1996). This analysis allowed agrouping of the isolates that clearly correlated with thegeographical origin of the stocks. In that study, fiveMexican isolates were genetically and biologicallycharacterized, with distinct degrees of virulence anddifferent patterns with restriction enzymes to T. cruzi.Two isolates produced high mortality and three lowmortality in infected mice. T. cruzi virulence manifests agreat diversity of medical and biological properties.These depend on the geographical distribution and ge-netic variation between strains (Espinoza et al. 1998).The T. cruzi used in this study produced low mortality(15%) in mice and was initially only mildly virulent.Mortality increased (to 50%) by the end of the study.
During the acute phase, parasites are present in dif-ferent organs, but in the chronic phase they destroyspecific organs, e.g. the heart. T. cruzi manifests geneticheterogeneity among isolates and stocks which couldexplain the degree of tropism to different organs. In thisstudy, the JALGO isolate was more likely to occur inskeletal and heart muscle. Obviously, the involvementof heart muscle is more harmful, since this organ
483
determines the patient’s quality of life. In fact, chronicchagasic cardiomyopathy in Mexico appears to affect alarge number of patients with heart disease, but manycases may be misreported because of the unspecific na-ture of the clinical symptoms (Dumonteil et al. 1999).Microvascular spasm and myocardial ischemia havebeen reported in infected mice. Among the factors thatcontribute to chagasic cardiomyopathy is an increase ofendothelin-1 mRNA expression in cardiac myocytes andendothelium (Hung et al. 2000). Clinical symptoms ofChagas’ disease have been attributed to autoimmunereactivity to antigens shared by the parasite and hostmuscle or neuronal tissue (Zhang et al. 1999). Duringthe acute phase, neuronal and cardiac fiber destructionoccurs when parasite nests rupture. T. cruzi antigens
become the targets for cellular and humoral im-mune response against the parasite (Soares and Santos1999).
Furthermore, adhesion between T. cruzi and phago-cytes is the first step in cellular parasitism, which plays acentral role in pathogenesis. In some cases, failed ad-hesion suggests a possible parasite evasion mechanism(Monteon et al. 1992).
Unfortunately, more domestic and wild animals arebeing infected, contributing to parasite dissemination inMexico. T. cruzi has been reported in Mexico in pigs(Sus scrofa), dogs (Canis familiaris) (Salazar et al. 1997),opossum (Didelphis marsupialis) and rats (Rattus rattus)(Zavala et al. 1996).
Blood transfusion, the second important mode fortransmission in Mexico, increases its impact. Studies ofblood donors in Mexico City showed a prevalence of0.3% (Ramos et al. 1993; Monteon et al. 1999). Routineserological screening should be included in blood banksto decrease this problem.
Jalisco is known as an endemic area for Chagas’disease with characteristic clinical features. High levelsof seropositivity have also been reported in blood banksin Mexico (Hernandez et al. 1988). In the rural area ofJalisco, epidemiological studies have shown an 18%positivity for T. cruzi antibody in serum (Hernandezet al. 1988; Ocampo et al. 1993). In the present study, we
Fig. 2A–DDetection of T cruzi by PCR in animal blood samples. AFirst DNA amplification (550 bp) and nested PCR (336 pb)products were detected. B. PCR amplification products of the Tc-24 gene in blood from infected mice on the 4th to 200th days. CTropism of T. cruzi was evaluated by PCR in different tissues at28 days after infection.MMolecular weight marker; NNC negativecontrols (without DNA), NC negative control (mouse DNAwithout infection). D L Liver, K kidney, D duodenum; I ileum, Ccolon, S spleen, SM skeletal muscle and H heart. All reactions wereperformed in triplicate as described previously. PCR products werevisualized with ethidium bromide and photographed. Densitomet-ric analysis was performed using the Kodak Digital Sciencesoftware
484

demonstrated by PCR assays the presence of Chagas’disease in a rural area of Jalisco. Tropism of T. cruzi toskeletal muscle observed in the mouse model of thisstudy could explain, in part, some asymptomatic pa-tients who were in contact with the vector and did notmanifest the characteristic clinical symptoms of thisdisease.
At present, the techniques employed for Chagas’ di-agnosis are the microscopic examination of blood andthe detection of antibodies against T. cruzi. These con-ventional techniques are not very reliable due to theirlow sensitivity, specificity and the presence of cross re-actions with other parasites. The use of purified T. cruzirecombinant molecules could overcome the problemof specificity. In recent studies, the utility of a 24-kDa(Tc-24) T. cruzi recombinant antigen for serodiagnosishas been shown (Godsel et al. 1995; Taibi et al. 1996).Tc-24 recombinant protein was employed in Westernblot assays showing that this protein was expressed in allT. cruzi strains obtained in different endemic countries
(Guevara et al. 1996). This problem might be overcomewith the use of PCR techniques.
Because of T. cruzi genetic variability, different pairsof primers have been used to detect the parasite by PCRtechniques. PCR reaction using oligonucleotides to theTc-24 gene protein was more sensitive than that basedon the amplification of T. cruzi kinetoplastid DNA(Guevara et al. 1996; Taibi et al. 1995).
Finally, this study shows that PCR amplificationcould be used as a diagnostic test for the early detection(acute phase) of Chagas’ disease and/or later during thechronic phase of infection when the parasite is notlonger in circulating blood. Moreover, PCR could beemployed in autopsy sample analysis to detect whetherT.cruzi was the cause of death, especially in myocardiuminfarct.
ELISA showed 17% specificity, and 48% sensitivity.In the population used in this study, PCR showed 85%(33/39) prevalence, data that are consistent with theparasite presence in endemic areas. These findings sug-gest that the PCR detection of T. cruzi represents thebest method to detect this parasite during all stages ofthe disease.
This easy and sensitive PCR test will prove to beuseful in epidemiological studies by providing an alter-native to other time consuming techniques. PCR is agood tool for epidemiological studies and at the sametime can provide an understanding of the etiology ofChagas’ disease and parasite tropism.
Fig. 3A–C Detection of T. cruzi by PCR amplification in humanblood samples. A Representative PCR amplification products fromthe blood of negative control human subjects who had never beenin contact with parasite (lanes A–F). PC Positive control, NCnegative control. B Representative PCR products of inhabitants ofan endemic zone (lanes 1–10) (San Martin Hidalgo, Jalisco).Patients 1 and 2 were PCR negative. NC Negative control, PCpositive control. The molecular size of the amplified DNAfragment was 550 bp. C Nested PCR (336 pb). HGDNA Humangenome DNA, M molecular marker
485

References
Britto C, Cardoso MA, Monteiro CM, Hasslocher, Xavier SS,Oelemann W et al. (1995) Polymerase chain reaction detectionof Trypanosome cruzi: human blood samples as a tool for di-agnosis and treatment evaluation. Parasitology 110:241–247
Dumonteil E (1999) Update on Chagas’ disease in Mexico. SaludPubl Mex 41:322–327
Engman DM, Kranse KH, Blumin JH, Kirchhoff LV, Donelson JE(1989) A novel flagelar Ca+2-binding-protein in trypanosomes.J Biol Chem 264:18627–18631
Espinoza B, Vera CJM, Gonzalez H, Ortega E, Hernandez R(1998) Genotype and virulence correlation within Mexicanstocks of Trypanosoma cruzi isolated from patients. Acta Trop70:63–72
Godsel LM,Tibbetts RS, Olson CL, Chaudoir BM, Engman D(1995) Utility of recombinant flagellar calcium-binding proteinfor serodiagnosis of Trypanosoma cruzi infection. J ClinMicrobiol 33:2082–2085
Guevara EA, Taibi A, Bilaut MO, Ouaissi A (1996) PCR-baseddetection of Trypanosma cruzi useful for specific diagnosis ofhuman Chagas’ disease. J Clin Microbiol 34:485–486
Hart DT, Vickerman K, Coombs GH (1981) A quick, simplemethod for purifying Leishmania mexicana amastigotes in largenumbers. Parasitology 82:345-355
Hernandez GRJ, Lozano KF, Martınez OC, Trujillo CF, Ruval-caba ES (1988) Estudio epidemiologico de la prevalencia de laenfermedad de Chagas en el estado de Jalisco, Mexico. RevAssoc Med Jal 3:28–33
Hung H, Petkova SB, Pestell RG, Bouzahzah B, Chan J, MagazineH, et al. (2000) Trypanosoma cruzi infection (Chagas’ disease)of mice causes activation of the mitogen-activated proteinkinase cascade and expression of endothelin-1 in the myocar-dium. J Cardiovasc Pharmacol 36:S148–150
Kirchhoff LV, Votava JR, Diane E, Moser OR, Moser DR (1996)Comparison of PCR and microscopic methods for detectingTrypanosoma cruzi. J Clin Microbiol 31:1171–1175
Krautz GM, Galvao MC, Camcado JR, Guevara EA, Ouaissi A,Rettli (1996) Use of a 24 kilodalton Trypanosoma cruzirecombinant protein to monitor cure of human Chagas’ disease.J Clin Microbiol 33:2086–2090
Macedo AM, Pena SDJ (1998) Genetic variability of T. cruzi:implications for the pathogenesis of Chagas disease. ParasitolToday 14: 91–96
Monteon PVM, Nekrassov V, Zepeda A, Aranda A, Reyes PA(1992) Interaction of a mixture of epimastigotes and trypom-astigotes from a culture of a Mexican isolate of Trypanosomacruzi with mouse macrophages. Rev Latinoam Microbiol34:43–48
Monteon PVM, Hernandez BN, Guzman BC, Rosales EJL, ReyesLPA (1999) American trypanosomiasis (Chagas’ disease)and blood banking in Mexico City: seroprevalence and itspotential transfusional transmission risk. Arch Med Res30:393–398
Ocampo GO, Lopez y Lopez J, Galvez GB, Trujillo CF, Cova-rrubias PA (1993) Megaesofago Chagasico con serologıa yxenodiagnostico positivos. Rev Gastroenterol Mex 58:36–38
Ouaissi A, Aguirre T, Plumas MB, Piras M, Shoneck R, Gras MH,et al. (1992) Cloning and sequencing of a 24 K-Da Trypanosomacruzi specific antigen released in association with membranevesicles and defined by a monoclonal antibody. Biol Cell 75:11–17
Pinto JC (1996) Las enfermedades tropicales y el enfoque de gen-ero. Biol Oficina Saint Panam 12:261–281
Piras R, Osomo de Rodrıguez OO, Martınez JC, Aguirre T, PirasMM, Henrıquez D (1988). Identificacion de antıgenos de Try-panosoma cruzi-especifico para su aplicacion en serodiagnosti-co. Acta Cient Venez 39:35
Ramos EAA, Monteon PVM, Reyes LPA (1993) Detection ofantibodies against Trypanosoma cruzi in blood donors. SaludPubl Mex 35:56–64
Salazar SPM, Bucio MI, Cabrera M, Bautista J (1997). First caseof natural infection in pigs. Review of Trypanosoma cruzi res-ervoirs in Mexico. Mem Inst Oswaldo Cruz 92:499–502
Soares MB, Santos RR (1999) Immunopathology of cardiomyop-athy in the experimental Chagas’ disease. Mem Inst OswaldoCruz 94:257–262
Taibi A, Guevara EA, Ouassi A (1995) Trypanosoma cruzi: analysisof cellular and humoral response against a protective recom-binant antigen during experimental Chagas’ disease. ImmunolLett 48:193–200
Taibi A, Guevara EA, Schoneck, Yahiaoui B, Ouaissi A (1996)Improved specificity of Trypanosoma cruzi identification bypolymerase chain reaction using an oligonucleotide derivedfrom the amino-terminal sequence of a Tc-24 protein. Parasi-tology 114:437
Velasco O, Guzman C, Cruz J, Gonzalez F (1991) La enfermedadde Chagas. Publicacion Tecnica del INDRE no. 8, MexicoCity
Zavala VJ, Barrera PM, Rodrıguez FME, Guzman ME, Pina H(1996) Infection by Trypanosoma cruzi in mammals in Yucatan,Mexico: a serological and parasitological study. Rev Inst MedTrop Sao Paulo 38:289–292
Zhang L, Tarleton RL (1999) Parasite persistence correlates withdisease severity and localization in chronic Chagas’ disease.J Infect Dis 180:480–486
486
Copyright © 2022 FDOKUMEN




















