
effect of parasite infestation on the - University Of Nigeria ...
107
1 EFFECT OF PARASITE INFESTATION ON THE BIOCHEMICAL AND HAEMATOLOGICAL CHARACTERISTICS OF SOME CLARIID (SILIRUFORMES) SPECIES OF THE ANAMBRA RIVER SYSTEM OF NIGERIA BY UGBOR, OGECHI NNABUCHI PG/M.Sc./12/64435 SUPERVISOR: DR G. E. ODO DEPARTMENT OF ZOOLOGY AND ENVIRONMENTAL BIOLOGY FACULTY OF BIOLOGICAL SCIENCES UNIVERSITY OF NIGERIA, NSUKKA JANUARY, 2015
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of effect of parasite infestation on the - University Of Nigeria ...

1
EFFECT OF PARASITE INFESTATION ON THE BIOCHEMICAL AND HAEMATOLOGICAL
CHARACTERISTICS OF SOME CLARIID (SILIRUFORMES) SPECIES OF THE ANAMBRA RIVER SYSTEM OF NIGERIA
BY
UGBOR, OGECHI NNABUCHI
PG/M.Sc./12/64435
SUPERVISOR: DR G. E. ODO
DEPARTMENT OF ZOOLOGY AND ENVIRONMENTAL BIOLOGY
FACULTY OF BIOLOGICAL SCIENCES
UNIVERSITY OF NIGERIA, NSUKKA
JANUARY, 2015

2
TITLE PAGE
EFFECT OF PARASITE INFESTATION ON THE BIOCHEMICAL
AND HAEMATOLOGICAL CHARACTERISTICS OF SOME
CLARIID (SILIRUFORMES) SPECIES OF THE ANAMBRA RIVER
SYSTEM, NIGERIA

3
DEDICATION
This thesis is dedicated to God the Father, God the Son and God the Holy Spirit in whom I live,
move and have my being.

4
CERTIFICATION
Ugbor, Ogechi Nnabuchi a postgraduate M.Sc. student in the Department of Zoology and
Environmental Biology, with Registration Number PG/M.Sc./12/64435 has satisfactorily
completed the requirements for the award of the degree of Master of Science (M.Sc.) in Zoology
and Environmental Biology (Fisheries). The content of this Thesis is original and has not been
submitted in part or full for other certificate, diploma, or degree of this or any other University
elsewhere.
__________________________ _____________________
Dr. G. E. Odo Date Supervisor
__________________________ ____________________
Prof. B. O. Mgbenka Date Head of Department
__________________________ _____________________
External Examiner Date

5
ACKNOWLEDGEMENTS
I am grateful to my Supervisor, Dr G. E. Odo for his sustained patience, guidance, fatherly
advice and constructive criticism which led to the success of this thesis. I am grateful to my
lecturers in the Department of Zoology and Environmental Biology for the profound influence
they had on my career. As my teachers, they were a source of inspiration to me and gave me
every necessary encouragement.
My indebtedness is due to my family members for their moral and financial support throughout
the study. May God bless you all.
I am thankful to Dr C. D. Nwani for his academic, moral and financial support. May heaven
reward you abundantly. To my colleagues, Chidinma Agbakwuo, Nonso Eze, Ejiofor Ezeamachi,
Gerald Attamah and so many of you who made my stay in the University of Nigeria, Nsukka
worthwhile, I say a very big thank you.
Ugbor, O. N. PG/M.Sc./12/64435

6
TABLE OF CONTENTS
TITLE PAGES
Title page - - - - - - - - - - - i Dedication - - - - - - - - - - - ii Certification - - - - - - - - - - - iii Acknowledgements - - - - - - - - - - iv Table of contents - - - - - - - - - - v List of figures - - - - - - - - - - - viii List of tables - - - - - - - - - - - ix List of plates - - - - - - - - - - - x Abstract - - - - - - - - - - - xi
CHAPTER ONE: INTRODUCTION AND LITERATURE REVIEW 1.1 Introduction - - - - - - - - - - 1 1.1.1 Justification of the Study - - - - - - - - 4 1.1.2 Objectives of the study - - - - - - - - - 5 1.2 Literature Review - - - - - - - - - - 5 1.2.1 Parasites and their effect on freshwater fish host - - - - - 5 1.2.2 Protozoa - - - - - - - - - - 6 1.2.3 Myxozoa - - - - - - - - - - 8 1.2.4 Trematoda - - - - - - - - - - 9 1.2.5 Cestoda - - - - - - - - - - 10 1.2.6 Nematoda - - - - - - - - - - 11 1.2.7 Acanthocephala - - - - - - - - - - 12 1.2.8 Crustacea - - - - - - - - - - - 13 1.2.9 Hirudinea - - - - - - - - - - 14 1.2.10 Factors affecting parasite assemblages in fish hosts - - - - - 14 1.2.10.1 Fish size (age) - - - - - - - - - 15 1.2.10.2 Fish diet - - - - - - - - - 16 1.2.10.3 Fish sex - - - - - - - - - 16 1.2.10.4 Fish immune status - - - - - - - - - 17 1.2.10.5 Condition of the environment - - - - - - - 18 CHAPTER TWO: MATERIALS AND METHODS 2.1 Study area - - - - - - - - - - - 20 2.2 Fish sampling - - - - - - - - - - 22 2.3 Examination of parasites - - - - - - - - - 22 2.3.1 Examination of ectoparasite - - - - - - - - 22 2.3.2 Examination of endoparasites - - - - - - - - 23

7
2.4 Treatment, preservation and fixation of parasites. - - - - - 23 2.4.1 Microscopic parasites - - - - - - - - - 23 2.4.2 Cestodes: - - - - - - - - - - 23 2.4.3 Nematodes - - - - - - - - - - 23 2.5 Identification of parasites - - - - - - - - - 23 2.6 Haematological examination - - - - - - - - 24 2.6.1 Haemoglobin estimation (Hb) - - - - - - - - 24 2.6.2 Blood haematocrit estimation (PCV) - - - - - - - 25 2.6.3 Red blood cell count (RBC) - - - - - - - - 25 2.6.4 Total white blood cell count (WBC) - - - - - - - 26 2.7 Biochemical Parameters - - - - - - - - - 26 2.7.1 Determination of aspartate transaminase (AST) - - - - - 26 2.7.2 Determination of alanine transaminase (ALT) - - - - - - 27 2.7.3 Determination of alkaline phosphatase (ALP) - - - - - - 27 2.7.4 Determination of blood urea - - - - - - - - 28 2.7.5 Determination of serum creatinine - - - - - - - 28 2.8 Statistical Analysis - - - - - - - - - 29 CHAPTER THREE: RESULTS 3.1 Fish Species - - - - - - - - - - 30 3.2 Parasite Species - - - - - - - - - - 30 3.3 Morphological features of the Parasites Recovered - - - - - - 30 3.4 Comparative Prevalence of Parasites in some Clariid Species of the Anambra River System. - - - - - - - - - - 33 3.5 Parasitic infections of some Clariid fish hosts - - - - - - 35 3.6 Monthly/Seasonal prevalence of parasites of some Clariid species of the Anambra River system. - - -- - - - - - - 37 3.7 Monthly/Seasonal Patterns of Parasitic Infection of Fish Hosts. - - - - 39 3.8 Prevalence of Parasites of Some Clariid Species of the Anambra River System in relation to sex - - - - - - - - -. - 41 3.9 Abundance of Parasites of some Clariid Species of the Anambra River System in Relation to Sex. - - - - - - - - - - 43 3.10 Intensity of Parasites in some Clariid species of the Anambra River System - - - 45 3.11 Parasitic Infestations of Fish Hosts (Clarridae) by Sex - - - - - 47 3.12 Effect of Parasite Intensity on the Condition Factor K, and Hepatosomatic Index, HSI of some Clariid species of the Anambra River System. - - - - 49 3.13 Parasite Infection of Fish Hosts by Body Weight - - - - - 52 3.14 Parasite Infection of Fish Hosts by Total Length of Fish - - - - 54 3.15 Site of Infection of Parasites in the Fish Hosts - - - - - - 56 3.16 Effect of Parasite on the Haematological Profile of some Clariid Species of the

8
Anambra River System - - - - - - - - - 58 3.17 Effect of Parasites on the Biochemical Parameters of some Clariid Species of the Anambra River System - - - - - - - - 60 CHAPTER FOUR: DISCUSSION 4.1 Prevalence, Mean Intensity and Abundance of Parasites - - - - 63 4.1.1 Protozoan - - - - - - - - - - 63 4.1.2 Cestodes - - - - - - - - - - 64 4.1.3 Nematodes - - - - - - - - - - 65 4.2 Influence of Body Weight and Total Length on Parasitic Infection - - - 65 4.3 Influence of Fish Hosts Sex on Parasitism - - - - - - 66 4.4 Influence of Season on the Patterns of Parasite Occurrence - - - - 67 4.5 Effect of Parasite on the Condition Factor and Hepatosomatic Index - - - 68 4.6 Effects of Parasites on the Haematological Profile of Fishes - - - - 69 4.7 Influence of Parasites on the Biochemical Parameters of Clariid Fishes of the Anambra River System - - - - - - - - 71 4.8 Conclusion - - - - - - - - - - 73 4.9 Recommendations - - - - - - - - - 74 REFERENCES

9
LIST OF FIGURES
FIGURES PAGES
1: Map of Anambra River - - - - - - - - 21
2: Correlation graph of condition factor of C. gariepinus and parasite intensity - 50
3: Correlation graph of hepatosomatic index of C. gariepinus and parasite intensity- 50
4: Correlation graph of condition factor of C. anguilaris and parasite intensity - 51
5: Correlation graph of hepatosomatic index of C. anguilaris and parasite intensity- 51

10
LIST OF TABLES
TABLES PAGES
1: Comparative prevalence of parasites in some Clariid species of the
Anambra River system - - - - - - - - 34
2: Parasite Infection of Fish Hosts by Species - - - - - 36
3: Monthly/Seasonal prevalence of parasites of Clariid species of the
Anambra River System. - - - - - - - - 38
4: Monthly/seasonal patterns of parasitic infection of fish host - - - 40
5: Prevalence of Parasites of some Clariid spps. of the Anambra River System
in relation to Sex - - - - - - - - - 42
6: Abundance in some Clariid Species of the Anambra River System - - 44
7: Intensity of Parasites of some Clariid species of the Anambra River System - - 46
8: Parasite infection of fish host by sex - - - - - - 48
9: Parasite Infection of Fish by Body Weight - - - - - 53
10: Parasite Infection of Fish by Total Length of Fish - - - - 55
11: Site Preference of Parasites in Fish Host - - - - - - 57
12: Effect of Parasites on the Haematological Profile of the Infected Clariid fishes - 59
13: Effect of Parasites on the Biochemical Parameters of some Clariid Species
of the Anambra River System - - - - - - - 61

11
LIST OF PLATES
PLATES PAGES
1: Trichodina sp. - - - - - - - - - 31
2: Epistylis sp. - - - - - - - - - - 31
3: Monobothroides woodland - - - - - - - - 31
4: Tail region of Monobothriod woodland - - - - - - 31
5: Polyonchobothrium clarias- - - - - - - - 32
6: Tail region of Polyonchobothrium clarias - - - -- - - 32
7: Head region of Rhabdochona congolensis - - - - - - 32
8: Head region of Procamallanus laevionchus - - - - - 32

12
ABSTRACT
A study based on monthly sampling of catches in Anambra river system (Otuocha station) was carried out between May 2013 and April 2014 to investigate the parasitofauna of wild clariid catfishes with reference to their prevalence, mean intensity and abundance; and also to ascertain the impact of these parasites on the physiology of its fish hosts. A total of 360 fish hosts, comprising of Clarias gariepinus (231) and Clarias anguilaris (129), were examined for parasites. Blood samples were collected from the caudal peduncle for haematological and biochemical enzymes assay using standard procedures. Six parasite species including two protozoans (Trichodina acuta and Epistylis sp.), two cestodes (Polyonchobothrium clarias and Monobothroides woodlandi) and two nematodes (Procamallanus laeviconchus and Rhabdocona congolensis) were recovered. The overall parasite prevalence is 41.1%, with protozoan parasites having the highest prevalence (25.5%), cestode (15.0%). Whereas nematode has the least parasite prevalent, infecting only 4.72% of the fish hosts. The relatively high overall parasite prevalence may be attributed to the relatively high level of domestic effluent into the river. Analysis of prevalence of parasitic infection of fish species by body weight and total length showed that parasite loads increased with increase in body weight and total length fish hosts. The study revealed that male fishes accumulate more parasites (P<0.05) than the female fishes. Monthly/seasonal patterns of parasite occurrence varied from one parasite to another. Whereas some parasites were found throughout the year, others were highly pronounced either in the rainy season or in the dry months. The influence of parasite prevalence on the condition factor (K) and hepatosomatic index (HSI) were highly significant (P<0.05). Whereas, condition factor showed negative correlations with increase in parasite intensity, HSI showed positive correlation with increase in parasite intensity. The study recorded a significant decrease in the mean values of red blood cell count (RBC), Haemoglobin concentration (Hb) and packed cell volume (PCV) (P<0.05) and significant increase in the mean values of white blood count (WBC) of the infected fishes when compared to the uninfected ones (P<0.05). Moreso, significant increase were observed in the mean values of aspartate transaminase (AST), alanine transaminase (ALT), alkaline phosphatase (ALP), urea and creatinine of the infected fishes when compared to the uninfected fishes (P<0.05). The haematological and biochemical alterations observed in the infected fishes reflect anemia and tissue damages caused by parasites invasion.

13
CHAPTER ONE
INTRODUCTION AND LITERATURE REVIEW
1.1 Introduction
Fish is an important and cheap source of protein supply in human diet (Usip et al., 2010; Bichi
and Dawaki, 2010). Its digestibility, amino acid contents, and low cholesterol content rank it
amongst the superior protein foods (Agbamu and Orhorhoro, 2007). According to El-Seify et al.
(2011), people obtain about one-fourth (25%) of their animal protein world-wide from fin and
shell fishes. More than forty percent (40%) of the protein diet of two-third of the global
population is obtained from fish (Agbamu and Orhorhoro, 2007). Fish is especially important in
the diet of people in the developing countries where malnutrition constitutes a major problem
(Omeji et al., 2010). Thus in Nigeria today, fish is the major source of affordable animal protein.
Due to the enormous demand on fish and other limiting factors, the demand and supply gap of
fish for this country stands at about 1 mmt per annum (Atanda, 2007) with a current demand of
1.5 mmt (Abolagba and Omorodion, 2006).
Fishes are important to man as a good source of protein in man’s diet and as a vector of some
human disease pathogens. One of the scientific importance of identifying a fish properly is to tell
to some reliable extent the health condition of the fish, and certain parasitic infections present
with some symptoms that bear on the external treatment of the fish (Ayanda, 2009a). All species
of fish are vulnerable to various parasitic infections depending on the species of fish and the type
of stream inhabited (Edema et al., 2008).
Some of the factors that enhance parasitic infection in fishes include reduced oxygen content of
water, increase in organic matter in the water and poor environmental conditions (Ayanda,
2009b).

14
Parasites are organisms that are metabolically dependent on other organisms (hosts) for their
continual existence (Symth, 1994). They obtain their basic requirements such as nutrients, shelter
and enzymes from their host (Smyth, 1994; Okafor and Ubachukwu, 2009). They are a major
concern not only to maritime but to freshwater fishes all over the world, and of particular
importance in the Tropics (Bichi and Yelwa, 2010; Ekanem et al., 2011). Their effects on fishes
include, nutrient devaluation (Hassan et al., 2010; Ekanem et al., 2011); alteration of biology and
behavior (Lafferty, 2008); lowering of immune capability, morbidity and mortality (Nmor et al.,
2004; Ekanem et al., 2011); growth and fecundity reduction (Nmor et al., 2004) and injuries
depending on the parasite species and load. Studies on the parasites of fishes is a major concern
in Africa presently (Akinsanya et al., 2007), Nigeria inclusive.
In Africa, a checklist of helminth parasites of freshwater fishes has been published by Khalil
(1971) and various reports also exist from different parts (countries) of Africa, highlighting on
intensities, prevalence, epidemiology and pathology of such parasitic infections. Paperna (1996)
reported that the cichlids harbour majority of the infection which includes the adult digenea
infecting different tissues of the body; trematode metacercaria of the family Clinostomidae
encysting in tissue (Echi et al., 2009); and adult monogenea of the families Pousopothocotylidae,
Dactylogyridae and Gyrodactylidae infecting the gills and skin.
The emanating need to culture fishes for protein consumption for the teeming rapidly growing
populations in the developing countries have made it necessary to intensify studies on the
parasite fauna of African freshwater fishes (Akinsanya et al., 2007). There is appreciable
documentation of parasite fauna of catfish in Nigeria. One of the earliest reports in Nigeria in
inland waters concerning fish parasites was that of Awachie (1966) who documented preliminary
information on the parasites of fish in the Kainji Reservoir. He observed that not many fishes

15
were infected. However in a similar study, Ukoli (1969) observed heavy parasitic infection of
fish species from the same reservoir. Similar works have also been done in Nigeria by Yakubu et
al. (2002) in Plateau State, Oniye et al. (2004) in Zaria, Ibiwoye et al. (2004) in Bida, Akinsanya
and Otubanjo (2006) in Lagos, Edema et al. (2008) in Benin city, Ayanda (2008) in Ilorin, Echi
et al. (2009) in Opi lake, Awharitoma and Ehigiator (2012) in Edo and Delta States, Eyo and
Iyaji (2013) in Kogi State and Eyo et al. (2014) at Warri river.
The Anambra River is the largest tributary of the lower Niger River below Lokoja, and often
regarded as a component part of the lower Niger lowlands (Udo, 1975). Thus considering its
importance as commercial fishing center supplying fishes to populace from Southern Nigeria and
beyond, a considerable biological and ecological studies have been undertaken and documented
on some economically important tropical fish fauna from the river basin (Awachie and Hare,
1977; Awachie and Ezenwaji, 1981; Eyo and Mgbenka, 1992; Mgbenka and Eyo, 1992; Nwani,
1998; Ezenwaji, 1999; 2002; Nwani, 2004; 2006; Odo, 2004; Odo et al., 2012).
Azugo (1978) studied the ecology of the helminth parasite of the fishes of Anambra River
system. Ezenwaji et al. (2005) studied the endo-helminth parasites of morchokid fishes of
Anambra river basin. However, there is paucity of information on the impact of parasites on the
haematology and the serum chemistry of the clariid family of the Anambra River System.
Therefore, as part of the study on the improvement of fishery and fish production in Anambra
River basin, more so, because of the importance of catfish in aquaculture industry (Food and
Agricultural Organisation, 2006), a study on the abnormalities of the blood and serum chemistry
of the fishes of the river caused by parasite infestation is necessary.

16
1.1.1 Justification of the Study
Fish health management is the concept of proactively regulating the host, pathogen and
environment to maximize the optimal condition for sustained growth and health. In order to get
better nutrition from fishes, they must be free from diseases and mishandling. Fish diseases may
be due to parasitic or non-parasitic causes. Among the parasites that infect freshwater fishes,
helminthes form the most diversified group (Pinky et al., 2012).
Parasites are a major concern to freshwater and marine fishes all over the world, and of particular
importance in the tropics (Iyaji and Eyo, 2008; Bichi and Dawaki, 2010; Ekanem et al., 2011).
They constitute a major limiting factor to the growth of farmed fish in Nigeria (Bichi and Yelwa,
2010). The effects of parasites on fish include nutrient devaluation (Hassan et al., 2010);
alteration of biology and behaviour (Lafferty, 2008); lowering of immune capability, induction
of blindness (Echi et al., 2009 a, b); morbidity, mortality, growth and fecundity reduction (Nmor
et al., 2004) and mechanical injuries depending on the parasite species and load (Echi et al.,
2009 a, b). Most supply of fish in Nigeria comes from the Riverian ecosystem (Ekanem et al.,
2011). Anambra State, where the Anambra River is located, is a traditional fishing district with a
vast coastal land mass in the eastern area of the Niger River.
However, the increased demand on fish as a ready and safe source of protein to humans has
necessitated the continuous studies on fish fauna and parasites. Therefore, the present study is
necessary to fill the gap in the current knowledge on the parasitofauna of catfishes from
Anambra River with regards to their effects on the haematology and serum chemistry of these
species.

17
1.1.2 Objectives of the Study
The general objective of this study is to investigate the parasitofauna of wild clariid catfishes
harvested from the Anambra River System, Nigeria.
The specific objectives are to:
- investigate the prevalence, intensity and abundance of parasitic infection in the catfishes
of the Anambra River System;
- study the haematological abnormalities in the catfish fauna of Anambra River System
due to parasitic infections;
- compare the biochemical enzyme activities of parasite infected and uninfected catfishes
of Anambra River System and
- study the influence of parasite on the condition factor and hepatosomatic index of
infected catfishes of Anambra River System.
1.2 Literature Review
1.2.1 Parasites and their Effect on Freshwater Fish Host
The effects of parasites on fish hosts in the wild may be difficult to isolate and quantify. However, studies
of fish in captivity or under culture conditions have provided much information about the effects of
parasites on fish survival (Iyaji and Eyo, 2008). It is evident that parasites can act as severe pathogens
causing direct mortality or rendering the fish more vulnerable to predators (Kunz and Pung, 2004). Other
effects of parasites on fish hosts, according to Sindermann (1987) include muscles degeneration, liver
dysfunction, interference with nutrition, interference with respiratory functions, cardiac disruption,
nervous system impairment, castration or mechanical interference with spawning, weight loss and gross
distortion of the body.

18
According to Iyaji and Eyo (2008), the economic important of freshwater parasites are grouped
into microparasites and macroparasites. The microparasites include protozoans – microsporideans
and myxozoans while the macroparasites are comprised of helminthes such as monogenea and the
diageneas, trematodes (flukes), cestodes (tapeworms), nematodes (roundworms) and
acanthocephalans (thorny headed worms). The arthropod parasites are represented mainly by the
copepods (Marcogliese, 2002), while annelid parasites are the leeches.
1.2.2 Protozoa
The Protozoans represent a vast assemblage of eukaryotic unicellular organisms. Protozoans are
the most commonly encountered fish parasites, and can be the easiest to identify and control
(Klinger and Francois-Floyd, 2009). According to Klinger and Francois-Floyd (2009), they can
build up to very high numbers when fish are crowded causing weight loss, debilitation and
mortality. This is because they exhibit a direct life cycle. Studies have established the presence
of ecto- and endo-parasitic protozoans among freshwater fishes in Nigeria (Bichi and Dawaki,
2010; Bichi and Yelwa, 2010; Abidemi-Iromini and Eze, 2011; Omeji et al., 2011; Adeyemo and
Daunemugham, 2012) and other countries (Nikolic and Simonovic, 1996; Molnar et al., 2004;
Dove and O’donoghue, 2005; Hussein et al., 2010). Bichi and Dawaki (2010) reported the
presence of the protozoa Ichthyopthirius and Myxobolus from the survey of ectoparasites of the
gills, fins and skin of Oreochromis niloticus in Bagauda Fish Farm in Kano, Nigeria. Omeji et al.
(2011) in Benue State, Nigeria, reported infestation of Clarias gariepinus in the wild and
cultured environment by the protozoan parasites, Ichthyobodo sp., Ichthyopthirius multifiliis,
Chilodonella sp. Trichodina sp. and Cryptobia iubilans. Adeyemo and Daunemughan (2012)
while, investigating parasites of wild Parachanna obscura in Bayelsa, Nigeria, recorded the
presence of Microsporidian parasite.

19
The main groups of protozoan parasites of freshwater fishes are the ciliates, flagelletes,
coccidians and microsporidia (Klinger and Francois-Floyd, 2009). Ciliates are protozoan
parasites with cilia. Symptoms associated with infection depend on parasite load and species.
Most ciliates do not appear to trouble host until numbers become excessive (Klinger and
Francois-Floyd, 2009). Ciliates affecting freshwater fishes include Ichthyopthirius multifilis,
Chilodonella sp., Tetrahymena sp., Trichodina sp., Apiosoma sp., Ambiphyra (formerly
Scyphidia sp.), Epistylis sp. and Capriniana sp. (Pouder et al., 2011). According to Alvarez-
Pellitero (2004) symptoms of ciliate infestation include spots of different colours on skin,
hyperplasia, degeneration and necrosis of the gills, weakening of fish, loss of appetite,
restlessness and breathing problems. Ciliates have been reported in freshwater fishes in the wild
(Nikolic and Simonovic, 1996; Molnar et al., 2004; Omeji et al., 2010) and cultured (Hoffman et
al., 1975; Abidemi-Iromini and Eze, 2011; Tang et al., 2012) environment. Abidemi-Iromini and
Eze (2011) recovered I. multifiliis and Trichodina acuta from assessment of parasite fauna of
Tilapia zilli and O. niloticus from different water bodies in Akure, Nigeria.
Flagellates are characterized by the presence of one or more flagella. These include Ichthyobodo
spp (also known as Costia) which causes costiasis, a disease of the skin and gills. Fish affected
by Ichthyobodo necator appear lethargic and thin, and may show grey-whitish pellicles on the
skin, epidemic erosion, haemorhages or ulcers, and gill hyperplasia, and oedema (Alvarez-
Pellitero, 2004 ). Cryptobia sp. and Piscinoodinium sp. are other ectoparasitic flagellates of
freshwater fishes. Hexamita (Spironucleus) sp., Trypanosoma sp. and intestinal and
haemoparasitic Cryptobia sp., are some endo-parasitic flagellates.
The coccidian parasites which are mainly intestinal, found in freshwater fishes are members of
the genera Gaussia (Molnar et al., 2004), Eimeria and Cryptosporidium (Eli et al., 2011).

20
Microsporidia are strictly intracellular parasites that utilize host tissue for reproduction. In some
species, the infected cell becomes hypertrophic, accommodating proliferating parasites (xenoma)
(FAO, 1991). The development cycle of microsporidians include merogony and schizogony
(proliferative phase) producing an enormous number of parasites, and sporogomy which
generates mature spores. The major pathological sign is associated with hypertrophy. Adeyemo
and Daunemughan (2012), investigating parasites of wild Parachanna obscura in Bayelsa,
Nigeria, recorded the presence of microsporidian parasite.
1.2.3 Myxozoa
Myxozoa parasites are spore-forming metazoans that have a two-host life cycle. Myxosporidia
are almost exclusively parasites of fish (Fomena et al., 2008). Some species are of economic
importance as they can cause chronic weakening disease (Klinger and Francois-Floyd, 2009).
2,180 myxopsorean species distributed in 62 genera have been established so far (Lom and
Dykova, 2006) and it is not unlikely that many more remain undiscovered. In Africa, more than
135 species are known today, which affect freshwater as well as marine fishes (Kostoingue et al.,
2001; Fomena et al., 2007; Fomena et al., 2008). About a 100 species in freshwater fishes in
Africa belonging to the genera Myxobolus (BUTSHLI, 1882), Henneguya (THELOHAN, 1892),
Myxobilatus (DAVIS, 1994), Chloromyxum (MINGAZZINI, 1890) and Parahenneguya
(SEKITI, 1997), has been established (Fomena et al., 2007). Cichlidae and Cyprinidae are some
common freshwater hosts (Paperna, 1996).
Myxospora cause coelozoic (in internal cavities e.g. urinary bladder) and histozoic infections
(Paperna, 1996). Infections by Myxobolus sp. and Henneguya sp. have been reported in Central,
East and West Africa (Paperna, 1996; Kostoingue et al., 2001; Fomena et al., 2007; Abowei and
Ezekiel, 2011). Fomena et al. (2007) reported three new species of Myxosporea of genus

21
Myxobolus in Cameroon. Bichi and Yelwa (2010) recorded Henneguya sp. from examination of
Clarias gariepinus in Bagauda Fish Farm, Kano, Nigeria.
1.2.4 Trematoda
Trematodes are flatworms also referred to as flukes. The class Trematoda consists of the
Monogenea, Digenea and Aspidogastrea with the last being of little parasitological significance
as most are free living. Monogeneans are mostly ecto-parasitic platyhelminths and typically
parasitize the gills, skins and fins of fishes (Reed et al., 2009). They are highly host-and site-
specific and also exhibit a direct life cycle (Reed et al., 2009). The opisthaptor, a posterior
adhesive apparatus, is the most recognizable morphological character of the group. Monogenea
includes two main groups, Monopistocotylea (with a simple adhesive disc) and
Polyopistocotylea (with a complex adhesive disc) (Olson, 1974). Gyrodactylus sp., Dactylogyrus
sp., Furnestinia sp. and Diplectanum spp. are the most significant monopisthocotyleans species
for cultured fishes (Alvarez–Pellitero, 2004). Polyopisthocotylea includes several species of
pathological concern for fish cultures, and most of them belong to the family Microcotylidae,
and some to Heteraxinidae. Several studies have reported monogeneans in freshwater fishes
(Jean-Francois and Alain, 1991; Mendoza-Franco et al., 1999; Tombi and Bilong, 2004;
Salgado-Maldonado, 2008; Soliman and Ibrahim, 2012). Mendoza-Franco et al. (1999) reported
the presence of monogeneans in cichlid, Pimelodid, characid and Poeciliid fishes. Data about
Monogenea in Nigeria fishes appear to be limited.
Lethargy, anoxia, loss of appetite, excess mucus secretion, scratching and haemorrhage are some
chemical signs associated with monogenean infection (Alvarez-Pellitero, 2004).
Digenean trematodes are endoparasitic platyhelminths with complex (indirect) life cycle
involving a series of hosts (the first of which is almost always a mollusk) (Cox, 1993, Smyth,

22
1994). Gastrointestinal tract, coelom and blood vessels are some sites occupied by the adult
digenean trematodes. Over 50 species of digenetic trematodes from 15 families occur in
freshwater fish species in Africa (Iyaji and Eyo, 2008). Fishes may act as either intermediate or
definitive host depending on the digenean species. Okaka and Akhigbe (1999) reported the
presence of the trematodes (Clinostomum spp., Allocreadium spp. and Diplostomum tragenna) in
freshwater fishes in River Osse in Benin, a Niger Delta area of Nigeria. The trematode
Clinostomum metacercaria had been observed in Parachanna obscura from Lekki Lagoon,
Lagos, Nigeria (Akinsanya et al., 2010).
1.2.5 Cestoda
Cestodes (tapeworms) are endoparasitic flat worms with indirect life cycle involving at least one
intermediate host. There is variation in the life cycle of cestodes with fish acting as the primary,
intermediate or paratenic host. Cestodes parasitize fishes in both culture and wild environments
and are of variable economic importance (Ayanda, 2009a; Bichi and Yelwa, 2010). Adult
cestodes are usually found in the gastrointestinal tract, while larval stages may be isolated from a
variety of organs. Larval cestodes unlike adults, are not highly host-specific.
Most species causing diseases in fish of economic significance fall within three orders:
Caryophyllidea (e.g. Carophyllaeus), Pseudophyllidea (e.g. Bothnocephalus, Ligula and
Diphyllobothrium) and Proteocephalidea (e.g. Proteocephalus) (Alvarez-Pellitero, 2004).
Mechanical damage or obstruction and interference with nutrient absorption may characterize the
presence of adult cestodes in the gastrointestinal tract, but the most serious pathology is caused
by migrating larva. The plerocercoid larvae of cestodes constitute the most damaging parasites of
freshwater fishes; the problem associated with infection results when larvae damage vital organs
such as brain, heart and eye (Klinger and Francis-Floyd, 2009).

23
Several studies have reported the presence of cestodes in freshwater fishes in the wild and
culture environments in Nigeria (Ezeri, 2002; Yakubu et al., 2002; Nmor et al., 2004; Akinsanya
and Otubanjo, 2006; Ayanda, 2009a; Bichi and Yelwa, 2010; Ekanem et al., 2011; Ogbulie et
al., 2011). The presence of the cestode, Polyonchobothrium was reported by Okaka and
Akhigbe (1999), Akinsanya and Otubanjo (2006) and Ayanda (2009a) in freshwater fishes in
Nigeria. Ezeri (2002) reported the presence of the larval cestode, Paradilepsis in cultured
Oreochromis niloticus in Ogun State, Nigeria. Anomotaemia sp., Protoeocephalus sp., Stocksia
pujehuni and Wenyonia acuminate are other species of cestode that have been isolated from
freshwater fishes in Nigeria (Okaka and Akhigbe, 1999; Akinsanya and Otubanjo, 2006;
Ayanda, 2009a).
1.2.6 Nematoda
Nematodes (or round worms) are endoparasites in animals. Most species which infect fish have
indirect life cycle with a single intermediate host and one or more paratenic hosts. Fish may
serve as primary, intermediate or paratenic host (Marina, 2008). They can infect all organs of the
host causing loss of function to damaged area (Klinger and Francois – Floyd, 2009). Adults are
usually found in the gastrointestinal tract and larvae in a variety of organs including skin, body
cavity, gastrointestinal tract and visceral of the fish hosts. Emaciation, anaemia, unthriftness and
reduced vitality are some symptoms of nematode infection in fishes (Klinger and Francois-
Floyd, 2009).
Nematodes have been reported in various freshwater systems in Nigeria (Okaka and Akhigbe,
1999; Nmor et al., 2004; Olofintoye, 2006; Akinsanya et al., 2008; Ayanda, 2009a). Olofintoye
(2006) reported the presence of the nematode, Cuculanus in freshwater fishes in Ekiti State,
Nigeria. Camallanus sp., Procamallanus sp., Spirocamallanus sp., Spinitectus sp., Serradactnitis

24
sp. and Spironoura sp. (nematodes) were isolated by Okaka and Akhigbe (1999) from freshwater
fish in Osse River, Edo State, Nigeria.
1.2.7 Acanthocephala
Acanthocephalan parasites are endoparasitic helminthes with in-direct life cycle involving
arthropod intermediate hosts and vertebrate final host. Fish may act as the definitive host,
harbouring adult worm in its gastrointestinal tract, or as intermediate host harbouring the second
stage larvae (cystecanth) with its retracted proboscis, in other extra-intestinal sites (Lyndon and
Kennedy, 2001). Acanthocephalan parasites are recognized by their recurved hooks-crowned
evaginable proboscis. Pathogenic effects are due to attachment of the adult to the gastrointestinal
tract by means of the proboscis, and also to the encapsulation of the larva stages in the tissues
(Paperna, 1996). Pathological sign depends on the acanthoceplan species and location. Extensive
inflammation, granuloma, peritonitis, and obstruction of the alimentary canal may be observed
(Paperna, 1996). Acanthocephalan parasites have been reported in freshwater fishes in Nigeria
(Okaka and Akigbe, 1999; Nwani et al., 2008; Ayanda, 2009a; Usip et al., 2010). Nwani et al.
(2008) isolated Rhadinorhynchus horridus and Gnathonemus petersi (Acanthocephala) from the
fish, Hyperopisus bebe bebe in Anambra River, Nigeria. Onyedineke et al. (2010) recovered the
acanthocephalan parasites, Pomphorhynchus, Quadrigidae and Neochinorhynchus in freshwater
fishes from River Niger at Illushi, Edo State, Nigeria.
1.2.8 Crustacea
Parasitic crustaceans are ectoparasites on fish and are usually blood feeders on the gills, skin and
fins; and large numbers can cause serious pathogenic effects (Marina et al., 2008). Parasitic
crustaceans are increasingly serious problems in wild and cultured fish (Piasecki et al., 2004;
Klinger and Francis – Floyd, 2009). Depending on the fish species and degree of invasion,

25
parasitic crustaceans may cause fish mortality (Oktener et al., 2008). Oktener et al. (2008)
reported mortality of the fish, Cyprinus carpio and Capoeta trutta due to infestation by
Lamproglena pulchella (Lernaeidae).
Parasitic crustaceans are found mainly in the class Branchiura, Copepoda and Malocostraca
(Marina, 2008); the class Copepoda being a major concern to fresh water fishes (Marcogliese
and Parasitology Module Steering Committee, 2011). Copepoda play major roles in freshwater
ecosystem which includes serving as food for many fish, acting as fish parasites, micropredator
of fish and other organisms, intermediate host of fish parasites, and hosts and vectors of human
diseases (Piasecki et al., 2004). According to Piasecki et al. (2004), they serve useful purpose if
they are properly managed.
Most of the parasitic crustaceans that have been described are copepods and majority of them
constitute a problem to freshwater fish mainly from the families Lernaeidae and Ergasilidae (Ho,
1998; Klinger and Francois-Floyd, 2009). According to Ho (1998), about 110 species of lernaeid
copepods are known from 322 species of freshwater fishes belonging to 161 genera in 41
families. Perez-Bote (2005) and Yuniar et al. (2007) are among studies that had reported
parasitic copepods in fish. Bichi and Yelwa (2010) had reported the presence of the crustacean
Ergasilus sarsi with 24.6% on Clarias gariepinus in Bagauda Fish Farm, Kano, Nigeria.
1.2.9 Huridinea
Leeches are occasionally seen in wild and pond raised fish where they act as ectoparasites
feeding on blood and body fluid of hosts; and pathology is dependent on infestation intensity
(Klinger and Francis Floyd, 2009). Abidemi-Iromini and Eze (2011) had reported the presence of
leeches in freshwater fish in Nigeria.

26
Volonterio et al. (2004) observed the effects of the leech Myzobdella urguayensis infesting the gill
filaments of Hoplias alabaricus, (Characiformes, Erythrinidae) and Rhamdia quelen (Siluriformes,
Pimelodidae), which resulted in hemorrhage with clot formation and fibrin deposition at the attachment
sites. They equally observed inflamed gill filaments which exhibited oedema and infiltration of
mononuclear leukocytes. Damages to skin comprised of bite wounds, hemorrhages, erosion of mucus
membranes, the epidermis and basal hyperplasia (Food and Agricultural Organisation, 1991; Volonterio et
al., 2004). Apart from the mechanical injuries and pathological effects leeches have on fish hosts; they
have also been discovered to be vectors of haemoprotozoans (Food and Agricultural Organisation, 1991).
For example Piscicola geometrea was shown to transmit viral disease to carp (Ahne, 1985). Bleeding
wounds may also become contaminated by opportunistic bacteria and fungi (Kabata, 1985).
1.2.10 Factors Affecting Parasite Assemblages in Fish Hosts
There are numerous biotic and abiotic factors that affect parasite assemblages (Esch, 1982; Kennedy,
1995). These factors include the following: physiological condition of the fish host, host diet, host size,
evolutionary history and environmental factors such as season of the year, size and type of water body,
altitude, temperature, salinity, oxygen and pH (Poulin, 2004; Rolbiecki, 2006; Sinkova et al., 2008;
Alverez – Pellitero, 2008; Iyaji et al., 2009; Lagrue et al., 2011 and Rahman and Saidin, 2011).
Though a number of physical and chemical factors are known to affect a wide range of aquatic
vertebrates and invertebrate’s life cycles, the effect of biotic factors on abundance and prevalence of
parasites has been the major focus of research (Iyaji et al., 2009). The ecological relationship between
hosts and parasites are usually influenced by the organisms inter and intra-specific interactions with
biotic and abiotic components of the environment (Williams and Jones, 1994).

27
1.2.10.1 Fish size (age)
Poulin (2000) stated that in fish population, parasitic infection tends to increase with increasing
host age and size. He argued that older fish have longer time to accumulate parasites than
younger ones and may provide establishment and therefore tend to have heavier worm burdens
because they eat more parasitized prey and offer large surface area for skin-attaching parasites.
Munoz and Crib (2005) reported that larger host has higher parasites richness, abundance and
pattern might be explained by combination of resources, time and prey. In general, large hosts
have more space, more flux of energy (i.e. food) and microhabitats for parasites than small hosts.
Furthermore, large fish are older than smaller individuals of the same species so that they have
more opportunities to become infected (Rhode 1993; Munoz et al., 2002).
Several studies have reported corresponding variation in parasite load and type with increasing
fish size (Olofintoye, 2006; Sinkova et al., 2008; Ekanem et al., 2011; Adeyemo and
Daunemughan, 2012). While Olofintoye (2006) reported an increase in infection with fish
growth in T. zillii, Clarias anguilaris and C. gariepinus examined, Adeyemo and Daunemughan
(2012) observed higher infection among the smaller sizes of Parachanna obscura in his
investigation. Rolbiecki (2006) in his explanation of the reasons for variation in parasite load
with varying fish length was of the view that different length classes of fish differ in their mode
of life and thus their degree of exposure to parasites. Variation in food type and increase in the
quantity of water consumed were among other reasons given as contributing to the observed
size-based parasite variation. He also stated that for parasites that actively penetrate their host
such as the monogeneans Ancyrocephalus paradoxus in Zander and Dactylogyrous
amphibothrium in ruffe, the trematode Posthodiplostomum and the copepod Anchtheres
percanum in Zander, size of the fish is the most important factor facilitating infection. Akinsanya

28
et al. (2007), Akinsanya et al., (2008) and Akinsanya et al. (2010) have, however, reported
parasite burdens that were independent on fish age. Length limits in parasite infection have been
reported (Rolbiecki, 2006). Increase in parasitic infection with increasing length of fish in fresh
water, and decrease in parasites with increasing fish length in marine habitat has been observed
(D’Silva et al., 2012). Fish size as it appears, is not an exclusive determinant of parasite
prevalence and/or intensity. The nature of the parasite and the fish habitat are other contributing
factors.
1.2.10.2 Fish diet
Link exists between fish diet and the flow of trophically transmitted parasites (Lagrue et al.,
2011). Food–borne parasite species are distributed among fish hosts according to food preference
(Rolbiecki, 2006) such that euryphagous (with varied diet) species accommodate wider range of
food borne parasite species, specificity in parasitism being the only restriction, while
stenophagous (with narrow range of diet) fish species are restricted to the parasites associated
with their food choice. Various parasites especially helminthes (e.g. Euplorchis californiensis
and Coitocaecum parvum) utilize trophic interaction among various organisms as a means of
transmission from one host to another. Euplorchis carliforniensis, for example, is transmitted
from killer fish, Fundulus parvipinnis to bird (final host) through trophic interaction (Lafferty,
2008). The transmission of Coitocaexum pervum to the fish Gobiomorphus cotidianus is another
instance ((Lagrue et al., 2011).
1.2.10.3 Fish Sex
Fish sex is one of the parameters that have been widely studied to determine its contribution to
parasite assemblage in fish. Explanations of this sex bias in parasitism have focused mainly on
two factors: variation among male and female fish in reproductive demand and ecological

29
requirements. According to Skarstein et al. (2001) and Simkova et al. (2008) fishes both male
and female invest differently in reproduction; male invest more in mate attraction through the
exhibition of sexual ornamentation while females invest in gamete production. The steroid
hormones (mainly testosterone) required for ornamentation in males is immunosuppressive
(Folstad and Karter, 1992) and thus subjects the male fish to great risk of parasitic infection
during spawning. Female fish are susceptible also to parasite during breeding period. Variation
in ecological requirements between male and female fish within a population may also contribute
to differences in parasite species assemblage (Iyaji et al., 2009).
Observations by several researchers have on the contrary, revealed parasite assemblage not
significantly affected by fish sex (Gbankoto et al., 2001; Akinsanya et al., 2007; Bichi and
Yelwa, 2010; Rahman and Saidin, 2011; Adeyemo and Daunemughan, 2012). Gbankoto et al.
(2001), observed a non-significant difference in the prevalence of Myxobolus sp. in male (20.7%)
and female (20.4%) Sarotherodon melanotheron. Rahman and Saidin (2011) in their conclusion
stated that fish sex play an important role in influencing the susceptibility of fish to parasites
despite the non-significant difference observed in prevalence stratified by sex in fish examined.
1.2.10.4 Fish Immune Status
A system of cells and tissues, biochemical and physiological characteristics of an organism that
ensures its protection against foreign invaders (such as parasites) constitute its immune system.
Because of the negative consequences of the presence of parasites on or in the host, the host has
developed a performing immune system to reduce fitness cost generated by parasitism (Sheldon
and Verhulst, 1996; Lochmiller and Deerenberg, 2000). The parasite conversely, attempt to
circumvent or weaken the host immune defense. These defense and evasion strategies by both

30
the host and the parasite are a necessity for survival in unfavourable condition (Cornet et al.,
2009; Nnadi et al., 2011).
Both innate and adaptive immune responses similar to that found in mammals are mounted by
fish to control parasitic infections (Alvarez – Pellitero, 2008; Rohlenova et al., 2011). According
to Alvarez –Pellitero (2008), innate immune initiation in fish depends on the recognition of
pathogen by pathogen recognizing-receptors (PRRs) (mainly Toll-like receptors). B-lymphocytes
and antibodies have also been implicated in adaptive response. The efficiency of host immune
response and hence its ability to eliminate and/or prevent infection depends on the host genetic,
physiological and environmental conditions such as stress, immaturity or aging, and
malnutrition; and also the evasive strategies of the parasite (Smyth, 1994).
1.2.10.5 The Condition of the Environment
The condition of the environment influence the parasites infecting a fish host (Koskivaara, 1992)
and the symptoms associated with the infection. Abiotic and biotic factors in the environment
such as water temperature, pH, oxygen content, pollution and competitors among other factors
affect fish (Thompson and Larsen, 2004) and fish parasitoses (Kenedy, 1995; Vankara et al.,
2011). Some of these factors affect fish physiology. Water temperature is considered the
strongest abiotic factor which affects fish physiology including immune function (Rohlenova et
al., 2011). Unfavourable temperature, as suggested by Rohlenova et al. (2011), lowers fish
acquired immunity. Pollution of fish habitat, which reduces water oxygen contents and /or alters
its pH level, may offset fish physiology favouring parasite infestation (Kelly et al., 2010). Lewis
and Morris (1986) had drawn attention to the hypnoxic effect to fish of nitrite pollution of their
habitat. Kelly et al. (2010) reported the synergistic effect of glyphosate formulation, a herbicide,
and the trematode parasite Telogaster opisthorchis on the fish Galaxias anomalus; such that the

31
juvenile fish survival which were unaffected by exposure to either glyphosate or I. opiosthorchis
infection alone, was significantly reduced by simultaneous exposure to both.

32
CHAPTER TWO
MATERIALS AND METHODS
2.1 Study Area
The Anambra River has its source in Ankpa highlands of Kogi State of Nigeria about 100 km
North of Nsukka (Azugo, 1978). It lies between latitudes 6010
- and 7
040
- East of the Niger
(Awachie and Hare, 1977). Essentially the river has a southward course crossing the Kogi /
Enugu State boundary, and then meanders through Ogurugu to Otuocha from where it flows
down to its confluence with the Niger at Onitsha. The main river channel, which has a total
length of about 207.40 km (Azugo 1978), has its bank covered by such plants like Echinoclae
species, Salvinia nymnellula, Ludiwigia decurrens, Imperita cylindirica, Andropogon spp,
Jussiaea spp, Pennisetum spp and Cynodon spp (Nwani, 2006). There is a rainy season (April –
September / October) and a dry season (October / November - March). The mean annual rainfall
is between 150 cm and 200 cm (Ilozumba, 1980). From December to January / February, the
basin is influenced by the harmattan but its effect is not well marked. The water temperature and
secchi disc reading in the river ranged from 24o C to 31o C and 5 cm to 85 cm, respectively (Odo,
2004). Agricultural activities are very high and crops such as yam, cassava, rice, millet,
vegetables, groundnut, potatoes, banana, and plantain are produced in large quantities. Fishing
methods in the river basin include bailing out water or pumping out water from ponds with water
pumps, construction of fish fences, the use of “atalla”, hooks and line, set lines, lift nets,
dragnets, beach seines, cast nets, among others (Awachie and Ezenwaji, 1981; Eyo and Akpati
1995). Species of fishes found in the river include Distichodus sp, Alestes sp, Mormyrids,
Clarias sp., Labeo and Heterobranchus sp. among others.

33
Fig. 1: Map of Anambra River
Source: Odo et al. (2009).

34
2.2 Fish Sampling
Fish samples (C. gariepinus and C. anguilaris) were collected monthly from Otuocha River
Station along the Anambra River from artisanal fishermen who used gill nets of mesh sizes
ranging from 150 mm to 200 mm. Baskets, traps and hook and lines were also used. Fish
collected were transported in a plastic container with water to the Hydrobiology and Fisheries
Research Laboratory, Department of Zoology and Environmental Biology, University of Nigeria,
Nsukka for analysis. In the laboratory, preliminary data records were taken. Fish were identified
using keys and catalogues (Olaosebikan and Raji, 1998, Idodo Umeh, 2003), date caught and sex
of the host fish (mature specimens). Total length was measured to the nearest 0.1cm using a
meter rule mounted on a dissecting board, while weight was measured to the nearest 0.1g using
an electronic balance.
2.3 Examination of Parasites
Freshly caught fish were examined for parasites using procedures described by Arthur and Albert
(1994) as follows:
2.3.1 Examination of ectoparasite
The external surfaces – fins, gills and skins were brushed into petri dish containing normal saline
and examined with a hand lens for the presence of ecto parasites. Scrapings from the skin, fins
and gills of each fish were taken and smeared on glass slides for examination of protozoan
parasites and smaller metazoan parasites (Paperna, 1996; Hamish, 2010; Ekanem et al., 2011).
Fish gills were dissected out and each gill filament and arch were examined with a hand lens for
the presence of monogeneans and myxosporidean cysts.

35
2.3.2 Examination of endoparasites
The fishes were dissected to expose the viscera. The visceral cavities and organs were examined
for cysts and larval endoparasites. The guts were removed and placed in petri dishes. Contents of
the guts were flushed with normal saline into beakers and then shaken to loosen mucus and other
intestinal debris. Parasites were recovered from the residue after centrifugation (1000 rpm) and
decanting of supernatant. Recovered parasites were mounted on slides and viewed using
Olympus microscope under high power magnification (×40) and identified. All parasites
recovered were recorded. Fish not examined were refrigerated (-40C) overnight and examined
the following day for parasites.
2.4 Treatment, Preservation and Fixation of Parasites.
2.4.1 Microscopic parasites: microscopic parasites were first stained for about 12h in
Haematoxyline and Eosin and transferred to 45% acetic acid for 2min and placed in methyl
salicyclate for 1min. The parasites were mounted in Canada balsam on clean slides.
2.4.2 Cestodes: cestodes were fixed in 4% neutral formalin and dehydrated in ethanol. They
were then stained with Eosin and mounted in Canada balsam on clean slides.
2.4.3 Nematodes: nematodes were placed in 70% ethyl alcohol and 5% glycerin added for
storage. They were later stained with Eosin and mounted whole in Canada balsam.
2.5 Identification of Parasites
Collected parasites were indentified to species level using appropriate keys (Yamaguti, 1963;
Paperna, 1996; Moravec, 1998; Chambrier and Vaucher, 1999; Pouder et al., 2011).

36
Parasitological indices were calculated following the method described by Jaywant et al. (2010)
as follow:
- Prevalence of infection = infected host x 100/total hosts examined.
- Mean intensity of infection = No. of parasites collected per sample/No of infected host.
- Abundance of infection = No. of parasites collected per sample/total host examined.
2.6 Haematological Examination
Prior to blood collection, the fishes were tranquilized with 150mg/l solution of tricane methane
sulphate (MS222). Blood samples were collected from the caudal artery using 2ml plastic
syringe and needle treated with anti-coagulant into glass tubes containing EDTA. The collected
blood were transferred into sample bottles for blood indices estimation.
2.6.1 Haemoglobin estimation (Hb)
Haemoglobin concentration was determined using the method described by Blaxhall and Daibley
(1973). The graduated tube was filled to the mark 2 on the red graduations with N/10 HCL using
a dropper. Blood sample was sucked into the capillary pipette to 20 cm mark; the end of the
pipette was wiped and blown into the acid in the mixing tube. The pipette was then rinsed by
sucking up the acid which was blown backwards into the tube 3 to 4 times before being allowed
to stand for 10 minutes to allow acid haematin to form. Similarly, the acid haematin was diluted
with distilled water in drop wise manner and mixed thoroughly. The colour was compared with
the standard by holding the comparator towards light. This process was repeated until the colour
of the diluted fluid matched that of the standards in the comparator. From the level of the fluid in
the mixing tube, the Hb was estimated in gram percent.

37
2.6.2 Blood haematocrit estimation (PCV)
This was determined by the microhaematocrit method described by Coles (1986). The coloured
end of a heparinized capillary tube was sealed with plasticin. The tube was filled to
approximately three fourth (3/4) of its length with well-mixed anticoagulated blood. The
capillary tube was then placed into a microhaematocrit centrifuge. Content of the tube was
centrifuged at 10,000 revolutions per minute (rpm) for 5 minutes. The height of the packed cell
in millimeter was measured using the haematocrit reader. The packed cell volume was then
calculated using the formula:
PCV (%) =
2.6.3 Red blood cell count (RBC)
Red blood cell count was estimated according to Backer et al. (1975). The blood was sucked
slowly and carefully up to 0.5 mark in the red blood cell diluting pipette. Immediately, the
pipette was plunged into the diluting fluid and sucked up to 101 mark. Then, the ends of the
pipette were gripped between the finger and the thumb and shaken thoroughly for 3 minutes. The
diluted blood sample was loaded on a Neubauer counting chamber and all red blood cells in the
five groups of 16 small squares in the central area of the Neubauer chamber were counted using
a light microscope at a high magnification of x 100. The number of cells counted for each sample
was calculated using the formula:
Where: dilution factor = 1: 200 and Volume counted = 0.02
Then, RBC (
RBC (

38
2.6.4 Total white blood cell count (WBC)
The White blood cell (WBC) was determined using the method described by Backer et al.
(1975). The blood was sucked slowly and carefully up to 0.5 mark in the red diluting pipette.
Immediately, the pipette was plunged into the diluting fluid and sucked up to 101 marks. Then,
the ends of the pipette were gripped between the finger and the thumb and shaken thoroughly for
3 minutes. The diluted blood sample was loaded on a Neubauer counting chamber and all the
cells in the four corner squares of the Neubauer chamber were counted using a light microscope
at a high magnification of x 100. The number of cells counted for each sample was calculated
using the formula:
Where: dilution factor = 1: 20, and Number of large squares counted = 4
WBC/
2.7 Biochemical Parameters
2.7.1 Determination of aspartate transaminase (AST)
The aspartate transaminase content of serum was determined using the method described by
Reitman and Frankel (1971). Two test tubes were labeled blank and sample respectively.
Distilled water (0.1 ml) was pipetted into the blank test tube, and 0.1 ml of serum added to the
sample test tube. Similarly, 0.5 ml of R1 was added into both test tubes. They were mixed
thoroughly and incubated for 30 minutes at 37oC. Thereafter, 0.5 ml of R2 was added to the test

39
tubes. The contents were mixed again and allowed to stand for 20 minutes at 25oC. NaOH (5 ml)
was added into all the tubes, and their contents were once again mixed and allowed to stand for
another 5 minutes. The absorbance of the samples against the sample blank was thereafter, read
using a spectrophotometer (Spectrumlab 752S, B) at 546 nm. The concentration of AST in the
serum sample was obtained from the standard curve.
2.7.2 Determination of alanine transaminase (ALT)
The alanine transaminase content of serum was determined using the method of Reitman and
Frankel (1971). Two test tubes were labeled blank and sample respectively. A total volume of
0.1 ml distilled water was pipetted into the blank test tubes, and 0.1 ml of serum added to the
sample test tubes. Similarly, 0.5 ml of the Buffer was added into both test tubes, mixed and
incubated for 30 minutes at 37oC. Thereafter, 0.5 ml of 2, 4- dinitrophenylhydrazine was added
into the test tubes. Contents of the test tubes were mixed again and allowed to stand for 20
minutes at 25oC. NaOH (5 ml) was added into the test tubes, and the contents were once again
mixed and allowed to stand for another 5 minutes. Thereafter, the absorbance of the sample
against the blank was read using a spectrophotometer (Spectrumlab 752S, B) at 550 nm. The
concentration of ALT in the serum was obtained from the standard curve.
2.7.3 Determination of alkaline phosphatase (ALP)
Alkaline Phosphatase content of serume was determined by the method of Haussament (1977).
0.02 ml of sample was pipetted into a cuvette followed by 1.0 ml of the reagent and mixed. The
initial absorbance was read at a wavelength of 405 nm after which timing commenced. The
absorbance reading was repeated after 1 minute, 2 minutes and 3 minutes intervals. Thereafter,
the ALP activity was calculated as follows:
Alkaline phosphatase (U/L) = 2760 × ∆ A/min

40
2.7.4 Determination of blood urea
The urea content was determined according to the method of Patton and Crouch (1977). Three
test tubes were labeled blank, sample and standard respectively for each of the samples. A total
volume of 10µl distilled water was pipetted into the blank test tubes, 10µl of serum into the
sample test tubes, and 10µl of the standard into the standard test tubes. Similarly, 100µl of R1
was then added into all the test tubes, mixed thoroughly and incubated for 10 minutes at 37oC.
Thereafter, 2.5 ml of R2 followed by 2.5 ml of R3 were added to all the test tubes. The contents
were mixed immediately and incubated again for 15 minutes at 37oC. The absorbance of the
samples (Asample) and Standard (Astandard) were read against the blank using spectrophotometer
(Spectrumlab 752S, B) at 546 nm. The concentration of Urea in the serum was then calculated
using the formular:
Urea concentration in serum (mg/dl) =
Where: Asample = Absorbance of the sample and Astandard= Absorbance of the standard
2.7.5 Determination of serum creatinine
The creatinine content was determined using the method described by Husdan and Rapoport
(1968). Three test tubes were labeled blank, sample and standard respectively for each of the
samples. A total volume of 100 µl distilled water was pipetted into the blank test tubes, 100 µl of
serum into the sample test tubes, and 100 µl of the standard into the test tubes labeled standard.
Similarly, 1000 µl of working reagent (R1a and R2b) were added into all the test tubes and
mixed. After 30 seconds, the absorbance (A1) of the standard and sample were read at 492 nm.
Exactly 2 minutes later, the absorbance (A2) of the standard and sample were also determined

41
using the same wavelength. The concentration of creatinine in the serum was then calculated
using the following formula: A2 – A1 = ∆ASample or ∆AStandard
Conc. of creatinine in serum (mg/dl) =
Where: A1 = Absorbance of sample or standard after 30 seconds, A2 = Absorbance of sample or
standard after 2 minutes, ∆Asample = change in absorbance of the sample, ∆Astandard = change in
absorbance of the standard, Asample = Absorbance of the sample Astandard = Absorbance of the
standard.
2.8 Statistical Analysis
Results generated were analyzed using Statistical Package for Social Sciences (SPSS) version
20.0. Significant differences in the prevalence of fish parasites were determined by Chi-square
test. One way analysis of variance (ANOVA) was used to compare hematological and
biochemical parameters of infected and uninfected fish samples. Regression analysis was used to
evaluate the length-weight relationship of fish. The length and weight was log- transformed
before subjecting to regression analysis. The condition factor (K) of the fish was calculated using
the formula K = 100W/L3 while the hepatosomatic index (HSI) was calculated using w/W × 100
and regression analysis was used to determine the effect of parasite on the K-factor and the HSI.
All analyses were done at 5% level of significant.

42
CHAPTER THREE
RESULTS
3.1 Fish Species
In the course of the study, a total of 360 clariid fish hosts were sampled and examined. The fish
hosts comprised of two species. The species were Clarias gariepinus, (N = 231) and Clarias
anguilaris, (N = 129). A total of 148 (41.1%) fish hosts were found to be infected while 212
(58.89%) were uninfected
3.2 Parasite Species
In this study, a total of 605 parasites were recovered, comprising the Protozoan ciliates,
Tricodina acuta and Epistylis sp.; the Cestode, Monobothroide woodlandi (Caryophyllidae) and
Polyonchobothrium clarias (Pseudophyllidae); the Nematode, Procamallanus laeviconchus
(Camallanidae), and Rhabdochona congolensis (Rhabdochanidae). Plates 1-6 illustrate the
parasites of some clariid fishes from the Anambra River System.
3.3 Morphological features of the Parasites Recovered
Parasites recovered were identified based on their morphological characteristics. The protozoan
parasite, Trichodina acuta (ciliate) was found on gills and skin of their fish hosts. They were
large with disc shaped body. The adhesive disc is saucer shaped. The parasite is provided with
several rows of cilia at the circular periphery and the inner circle of toothed dentricles. The
macronucleus is horse shoe-shaped and the micronucleus is small and difficult to be seen in
some specimens (Plate 1). Epistylis sp. was isolated from the skin and gills of their fish hosts. It
is a sessile contractile ciliate. It has a long and non contractile stalk. It often formed branched
colonies. The distal end of the organism is surrounded by rapidly moving cilia which appeared as
a blur (Plate 2).

43
Plate 1: Trichodina acuta Plate 2: Epistylis sp.
Plate 3: Monobothroid woodland Plate 4: Tail region of Monobothriod woodlandi

44
Plate 5: Polyonchobothrium clarias Plate 6: Tail region of Polyonchobothrium clarias
Plate 7: Head region of Rhabdochona congolensis Plate 8: Head region of
Procamallanus laeviconchus

45
The unsegmented cestode, Monobothroide woodlandi (Caryophyllidae) was found in the
intestine of the fish hosts. The worm is white in colour and elongated. It has a large rounded or
triangular scolex. The worm has only one bothrium at the anterior end (Plate 3). The segmented
cestode, Polyonchobothrium clarias (Pseudophyllidae) was isolated from glandular stomach of
their fish hosts. The scolex is elongated, triangular in shape and carries one row of hooks and
bears laterally two shallow Bothria (Plate 4). Segmentation begun directly after the scolex with
immature stages, then mature stages. The nematodes (round worms) were diverse in appearance
in this study. Usually, they were elongated and cylindrical, tapering at both ends. In P.
laeviconchus (Camallanidae), the nerve ring surrounding the oesophagus is located towards the
anterior part of the oesophagus. The mouth is oval in shape with a blunt tail in the males and a
tapering pointed tail in the females (Plate 5). Rhabdochona congolensis (Rhabdochanidae)
another common nematode has the nerve ring on the intestine with a funnel like mouth. The tail
of both males and females are pointed (Plate 6).
3.4 Comparative Prevalence of Parasites in some Clariid Species of the Anambra River
System
In the Anambra River System, the overall prevalence of parasites in the clariids (C. gariepinus
and C. anguilaris) does not differ significantly (P> 0.05). However, C. gariepinus had higher
prevalence (44.2%) of infection than C. anguilaris (35.7%) as shown in Table 1.

46
Table 1: Comparative Prevalence of Parasites in Some Clariid Species of the Anambra
River system.
Fish spps No. sampled No. infected (%)
Clarias gariepinus 231 102 (44.2)
Clarias anguilaris 129 46 (35.7)
X2 2.468
P – value 0.120ns
X2 = Chi square, n.s = no significant differences (p>0.05)

47
3.5 Parasitic Infections of some Clariid Fish Hosts
All the six parasite taxa recorded in this study infected all the fish hosts, but with varying rates
(Table 2). Among the parasites that infected the clariid fishes, infection by protozoans was
generally high compared to other parasites. Tricodina acuta recorded the highest infection with
49 fish hosts (C. gariepinus, N=31 and C. anguilaris, N=17) infected. C. gariepinus had a
prevalence of 13.4%, mean intensity of 2.23 and abundance of 0.31 while C. anguilaris had a
prevalence of 13.2%, mean intensity of 2.12 and abundance of 0.28. Whereas, Epistylis sp.
recorded a prevalence of 12.12%, in C. gariepinus, C. anguilaris recorded prevalence values of
12.4% each. The prevalence recorded by other parasites included 3.46%, 2.16%, 8.23% and
7.79% by Procamallanus laevionchus, Rhabdochona congolensis, Polyonchobothrium clarias
and Monobothroide woodlandi respectively for C. gariepinus and a prevalence of 2.33%, 0.78%,
8.53% and 6.20% by Procamallanus laeviconchus, Rhabdochona congolensis,
Polyonchobothrium clarias and Monobothroide woodlandi for C. anguilaris respectively.

48
Table 2: Parasite Infection of Fish Hosts by Species
A= No. of hosts infected, B = No. of parasites recovered, C = % prevalence of parasites, D = Mean intensity of parasites, E = parasite abundance
Parasite
Clarias gariepinus Clarias anguilaris
PROTOZOANS
A B C D E A B C D E
Trichodina acuta
31
72
13.4
2.23
0.31
17
36
13.2
2.12
0.28
Epistylis sp. TOTAL NEMATODES
28 59
95 167
12.12 25.25
3.39 2.83
0.41 0.72
16 33
34 70
12.4 25.58
2.13 2.12
0.26 0.24
P.laeviconchus
8
33
3.46
4.13
0.14
3
18
2.33
6
0.14
R. Congolensis TOTAL CESTODES
5 13
18 51
2.16 5.62
3.6 3.92
0.078 0.21
1 4
3 21
0.78 3.11
3 5.25
0.023 0.16
P. clarias
19
93
8.23
4.9
0.40
11
44
8.53
4
0.34
M. woodland TOTAL
18 37
104 197
7.79 16.06
5.78 5.32
0.45 0.85
8 19
55 99
6.20 14.73
6.9 5.2
0.43 0.77

49
3.6. Monthly/Seasonal Prevalence of Parasites of some Clariid Species of the Anambra
River System.
From Table (3) it is shown that C. gariepinus had the highest prevalence of infection (70% and
60%) in the months of May and April respectively. Whereas C. anguilaris had the highest
prevalence of 60% in January 2014 (dry season) and April 2014 (rainy season). However, there
were no significant differences (P>0.05) in the monthly/seasonal prevalence within the two fish
species studied.

50
Table 3: Monthly/Seasonal Prevalence of Parasites of Clariid Species of the Anambra River
System.
Clarias gariepinus Clarias anguilaris
Month No. sampled No. infected (%) No. sampled No. infected (%)
May 013 20 14(70) 10 5(50)
June 013 20 9(45) 10 4(40)
July 013 20 7(35) 10 2(20)
Aug. 013 19 5(26.3) 11 3(27.3)
Sept. 013 20 7(35) 10 2(20)
Oct. 013 19 5(26.3) 11 2(18.2)
Nov. 013 18 7(38.9) 12 5(41.7)
Dec. 013 16 8(50) 14 4(28.6)
Jan. 014 20 11(55) 10 6(60)
Feb. 014 18 9(50) 12 5(41.7)
March 014 21 8(38.1) 9 2(22.2)
April 014 20 12(60) 10 6(60)
X2 15.664
P–value 0.154n.s
11.475
0.404n.s
X2 = chisquare, n.s = no significant difference (P>0.05)

51
3.7. Monthly/Seasonal Patterns of Parasitic Infection of Fish Hosts.
Table 4 showed the seasonal patterns of parasitic infection of clariid fish hosts of the Anambra
River System. The protozoan infestations were recorded throughout the year. Trichodina acuta
and Epistylis sp. had their peak of infestations on the fish hosts in the months of Jan. and Feb.
(dry season). While Trichodina acuta recorded a prevalence of 23.3% in Jan. and 26.67% in Feb.
Epistylis sp. recorded a prevalence of 20% and 16.67% in the months of Jan. and Feb.
respectively. Whereas Trichodina acuta had the least infestation (6.67%) each in the months of
July and Aug. (rainy season), Epistylis sp. had its least infestation in October (end of rainy
season).
Among the Nematode parasites, infestations by Procamallanus laeviconchus on the fish hosts
were absent in the months of July, Sept. and Oct. (peak of rainy season) and Feb. (dry season).
Infestations by R. congolensis were recorded in the months of May, Oct. and April (rainy season)
and January (peak of dry season). It was observed that infestations by R. congolensis were absent
in the peak of rainy season. Generally, nematode infestations of fish hosts were observed to be
low in the present study.
The cestode, Polyonchobothrium clarias infestation was recorded throughout the year with
highest prevalence of infection of 16.677% and 13.33% in the months of May and April (rainy
season) respectively. M. woodlandi infestation occurred throughout the year except in Dec (onset
of dry season). However, the prevalence (13.33%) of M. woodlandi was highest in the month of
Feb. (peak of dry season).

52
Table 4: Monthly/Seasonal Patterns of Parasitic Infection of Fish Hosts
Variables Months Parasite May
013 June 013
July 013
Aug. 013
Sept. 013
Oct. 013
Nov 013
Dec. 013
Jan. 014
Feb. 014
Mar. 014
Apr. 014
Trichodina sp. Host infected 5 3 2 2 3 3 5 5 7 8 3 6 Parasite recovered
13 9 6 3 8 8 10 9 13 13 7 18
Prevalence 16.67 10 6.67 6.67 10 10 16.67 16.67 23.33 26.67 10 20 Mean intensity 2.6 3 3 1.5 2.67 2.67 2 1.8 1.89 1.63 2.33 3 Abundance 0.43 0.3 0.2 0.1 0.27 0.27 0.33 0.3 0.43 0.43 0.23 0.6 Epistylis sp. Host infected 4 4 2 3 2 1 2 4 6 5 4 3 Parasite recovered
10 12 7 6 9 6 8 11 15 12 9 15
Prevalence 13.33 13.33 6.67 10 6.57 3.33 6.67 13.33 20 16,678 13.33 10 Mean intensity 2.5 3 3.5 2 4.5 6 4 2.75 2.5 2.4 2.5 5 Abundance 0.33 0.4 0.23 0.2 0.3 0.2 0.27 0.37 0.5 0.4 0.3 0.5 P. laevionchus Host infected 2 1 0 1 0 0 1 1 3 0 2 2 Parasite recovered
11 8 0 5 0 0 2 6 6 0 5 11
Prevalence 6.67 3.33 0 3.33 0 0 3.33 3.33 10 0 6.67 6.67 Mean intensity 5.5 8 0 5 0 0 2 6 2 0 2.5 5.5 Abundance 0.367 0.267 0 0.17 0 0 0.067 0.2 0.2 0 0.17 0.37 R. congolensis Host infected 1 0 0 0 0 1 0 0 1 0 1 1 Parasite recovered
4 0 0 0 0 4 0 0 3 0 2 6
Prevalence 3.33 0 0 0 0 3.33 0 0 3.33 0 3.33 3.3 Mean intensity 4 0 0 0 0 4 0 0 3 0 2 6 Abundance 0.133 0 0 0 0 0.133 0 0 0.1 0 0.067 0.2 P. clarias Host infected 5 2 3 1 3 1 2 2 4 3 2 4 Parasite recovered
24 16 9 10 12 13 18 19 21 16 9 17
Prevalence 16.67 6.67 10 3.33 10 3.33 6.67 6.67 13.33 10 6.67 13.33 Mean intensity 4.8 8 3 10 4 13 9 9.5 5.3 5.33 4.5 4.3 Abundance 0.16 0.53 0.3 0.33 0.4 0.43 0.6 0.63 0.7 0.53 0.3 0.57 M. woodlandi Host infected 3 2 2 1 1 1 2 0 2 4 2 3 Parasite recovered
15 12 6 8 7 7 11 0 11 18 8 10
Prevalence 10 6.67 6.67 3.33 3.33 3.33 6.67 0 6.67 13.33 6.67 10 Mean intensity 5 6 3 8 7 7 5.5 0 5.5 4.5 4 3.3

53
3.8 Prevalence of Parasites of Some Clariid Species of the Anambra River System in
Relation to Sex.
Comparison of the overall prevalence of parasites in relation to the sexes of the two clariid
species studied (C. gariepinus and C. anguilaris), showed no significant differences (P>0.05) as
shown in Table (5). However, highlighting the prevalence between sexes, male clariids (with
46.7% and 38.2%) prevalence for C. gariepinus and C. anguilaris respectively had more
parasites than the female with 40.9% and 32.1% prevalence for both C. gariepinus and C.
anguilaris.

54
Table 5: Prevalence of Parasites of some Clariid Species of the Anambra River System in
Relation to Sex.
Clarias gariepinus Clarias anguilaris
Sex No. sampled No. infected (%) No. sampled No. infected (%)
Male 137 64(46.7) 76 29(38.2)
Female 97 38(40.9) 53 17(32.1)
X2 0.769
P – value 0.48n.s
0.503
0.576n.s
n.s no significant differences (P> 0.05) across the vertical row.

55
3.9 Abundance of Parasites of some Clariid Species of the Anambra River System in
Relation to Sex.
The overall parasite abundance in C. gariepinus and C. anguilaris is shown in Table 6. The
values are 1.83 ± 0.15 and 1.44 ± 0.18 respectively. A Comparison of the abundance of parasites
by sexes of the two clariid species studied, show significant differences (p<0.05) between only
sexes of C. gariepinus (Table 6).

56
Table 6: Abundance of Parasites in some Clariid Species of the Anambra River System
Clarias gariepinus Clarias anguilaris
No. sampled Abundance
(mean ± S.E)
No. sampled Abundance
(mean ± S.E)
Overall Sexes 231 1.83±0.15 129 1.44±0.18
Male 137 2.15±0.22 76 1.06±0.24
Female 94 1.38±0.19 53 1.71±0.27
P – value 0.02* 0.088n.s
n.s No significance differences( P>0.05) *Significant differences (P<0.05)

57
3.10 Intensity of Parasites in some Clariid species of the Anambra River System
The overall parasite intensity in C. gariepinus and C. anguilaris are 4.15 ± 1.57 and 4.04 ± 1.51
respectively. A Comparison of the intensity of parasites by sexes for the two species showed
significant variation (P<0.05) between sexes of the two species (Table 7).

58
Table 7: Intensity of Parasites of some Clariid species of the Anambra River System
Clarias gariepinus Clarias anguilaris
No. sampled
No. infected
Intensity (mean±S.E)
No. sampled
No. infected
Intensity (mean ± S.E)
Overall Sexes 231 102 4.15±1.57 129 46 4.04±1.51
Male 137 63 4.57±0.19 76 29 4.48±0.25
Female 94 38 3.37±0.21 53 17 3.29±0.37
P – value 0.0001* 0.0001*
*Significant difference (P<0.05)

59
3.11 Parasitic Infestations of Fish Hosts (Clariidae) by Sex
Chi-square analysis indicated that while sex is an important factor in parasitic infection of many
fish species, some fish species showed no marked influence on sex. Secondly, while some
parasites infected both sexes equally, other parasites infected one sex more heavily than the other
(Table 8).

60
Table 8: Parasite infection of fish host by sex
Host/parasite Male N = 137 Female N = 94
C. gariepinus A B C D E A B C D E Trichodina acuta 22 51 16.1 6.2 0.37 9 21 9.6 10.4 0.22
Epistylis sp. 18 61 13.1 7.6 0.45 10 34 10.6 9.4 0.36
P.laeviconchus 4 17 3.0 8.1 0.12 4 16 4.3 23.4 0.17
R. Congolensis 3 11 2.2 12.5 0.08 2 7 2.1 47 0.07
P. clarias 12 64 8.8 11.4 0.47 6 29 6.4 15.67 0.31
M. woodlandi 11 64 8.0 12.5 0.47 7 40 7.4 13.4 0.43
Male N = 76 Female N = 53
C. anguilaris A B C D E A B C D E
Trichodina acuta 11 23 14.5 6.9 0.30 6 13 11.3 4.1 0.25 Epistylis sp. 8 17 10.5 9.5 0.23 8 13 15.1 4.1 0.25
P.laeviconchus 2 12 2.6 38 0.6 1 6 1.9 8.8 0.11
R. Congolensis 1 3 1.3 76 0.013 0 0 0 0 0
P. clarias 7 28 9.2 10.9 0.37 4 16 7.5 3.3 0.3
M. woodlandi 5 35 6.6 15.2 0.46 3 20 5.7 2.7 0.38
A= No. of host infected, B = No. of parasites recovered, C = % prevalence of parasites, D = Mean intensity of parasites, E = parasite abundance

61
3.12 Effect of Parasite Intensity on the Condition Factor K, and Hepatosomatic Index, HSI
of some Clariid species of the Anambra River System.
The intensity of parasite was found to have highly significant influence (P<0.05) on the
condition factor and hepatosomatic index of both C. gariepinus and C. anguilaris.
C. gariepinus recorded 0.611 and 0.977 correlation coefficient (r) for condition factor and
hepatosomatic index respectively, while 0.548 and 0.965 were recorded by C. anguilaris
Parasite intensity explained the 37.3% and 95.4% variation in the condition factor and
hepatosomtic index of C. gariepinus. In the same vein, parasite intensity had 30.1% and 93.1%
influence on the K-factor and HSI of C. anguilaris respectively (Fig. 2 - 5).
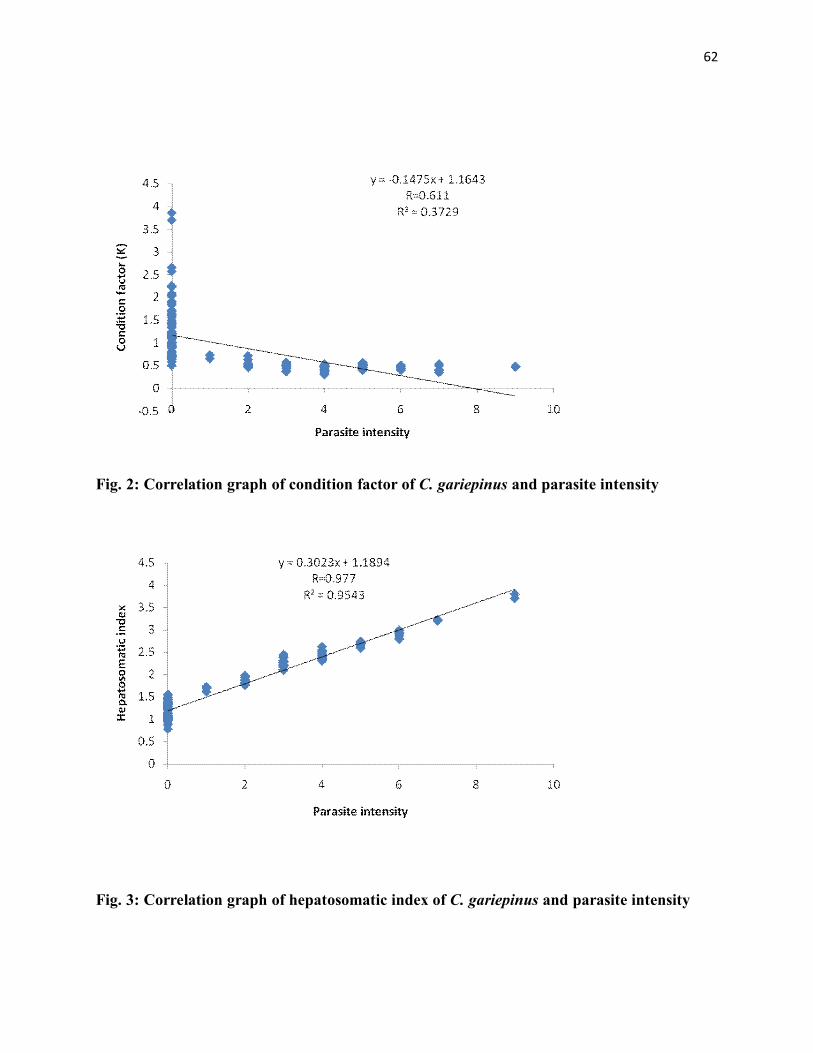
62
Fig. 2: Correlation graph of condition factor of C. gariepinus and parasite intensity
Fig. 3: Correlation graph of hepatosomatic index of C. gariepinus and parasite intensity

63
Fig. 4: Correlation graph of condition factor of C. anguilaris and parasite intensity
Fig. 5: Correlation graph of hepatosomatic index of C. anguilaris and parasite intensity

64
3.13 Parasite Infection of Fish Hosts by Body Weight
Table 9 shows the infection of parasites in the fish species based on body weight.
The protozoan parasites were absent in fishes, with weight less than 75g. However, as the weight
of fish increased from 76g and above, an increase in parasite load was observed in the clariid
species studied. The highest prevalence of infection (25.95% and 24.43%) in Trichodina acuta
and Epistylis sp. were recorded in fish of 126g+ body weight classes.
The numbers of cestode parasites were found to increase with increase in body weight of fish
hosts. The highest prevalence of parasite infection was recorded in 126g+ weight class of fishes.
R. congolensis followed the same trend with cestode infection. However, Procamallanus
laeviconchus was found to be absent in fishes of less than 75g weight class. They recorded their
highest frequency of 6.11% in fish of 126g+ weight class. Generally, it was observed from this
study that parasite load increased with increase in weight of fish hosts (Table 9).

65
Table 9: Parasite Infection of Clariid Fish Hosts by Body Weight
BODY WEIGHT CLASS
PARASITES 0 – 75 g N = 148
76 – 125 g N = 81
126 g+ N = 131
PROTOZOANS
A B C D E A B C D E A B C D E
Trichodina acuta
0
0
0
0
0
14
30
17.28
2.14
0.37
34
78
25.95
2.3
0.60
Espistylis sp. NEMATODES
0 0 0 0 0 12 34 14.80 2.83 0.41 32 93 24.43 2.9 0.71
P.laevionchus 0 0 0 0 0 3 11 3.70 3.67 0.13 8 40 6.11 5 0.31
R. Congolensis CESTODES
1 2 0.68 2 0.014 2 5 2.47 2.5 0.06 3 15 2.29 5 0.11
P. clarias 1 3 0.68 3 0.020 10 31 14.35 3.1 0.40 18 103 13.74 5.72 0.79
M. woodlandi 1 6 0.68 6 0.041 6 38 7.41 6.33 0.47 19 115 14.5 0.88
A= No. of host infected, B = No. of parasites recovered, C = % prevalence of parasites, D = Mean intensity of parasite, E = parasites abundance

66
3.14. Parasite Infection of Clariid Fish Hosts by Total Length of Fish
Table 10 shows that the parasite prevalence for all the fish species increased with increase in
total length. The highest records of prevalence (26%, 23.6%, 7.3%, 2.44%, 13.82% and 16.3%)
for Trichodina acuta, Episylis sp., P. laeviconchus, R. Congolensis, P. Clarias and M. woodlandi
respectively were indicated for fishes at a length class of 30 cm+.

67
Table 10: Parasite Infection of Clariid Fish by Total Length
PARASITE
0 – 15 cm N = 148
16 – 30 cm N = 81
30 cm+ N=123
PROTOZOANS
A B C D E A B C D E A B C D E
Trichodina acuta
0
0
0
0
0
18
34
10.4
1.89
0.2
32
74
26.0
2.3
0.6
Epistylis sp. NEMATODES
0 0 0 0 0 15 29 8.7 1.93 0.17 29 94 23.6 3.2 0.8
P. laeviconchus 0 0 0 0 0 2 9 1.2 4.5 0.05 9 41 7.3 4.56 0.33
R. Congolensis CESTODES
0 0 0 0 0 2 4 1.2 20 0.02 3 16 2.44 5.33 0.13
P. clarias 0 0 0 0 0 11 29 6.4 2.6 0.17 17 105 13.82 6.2 0.90
M. woodlandi 0 0 0 0 0 5 35 2.9 7 0.2 20 118 16.3 5.9 0.95
A= No. of hosts infected, B = No. of parasites recovered, C = % prevalence of parasites, D = Mean intensity of parasites, E = parasites abundance

68
3.15 Site of Infection of Parasites in the Fish Hosts
The results of the sites where parasites were recovered in fish hosts are presented in Table 11.
Results indicated that 75.9% of the Trichodina sp. were recovered from the gills whereas 14.8%
were recovered from the skin. 70.5% of Epistylis sp. were found on the gill while 29.5% were
found on the skin. R.congolensis and M. woodlandi were found to have 100% preference for the
intestine.

69
Table 11: Site Preference of Parasites in Fish Host
Parasites Intestine Stomach Gill Body cavity Skin Total
PROTOZOANS
No
% No % No % No % No %
Trichodina acuta 0 0 0 0 82 75.9 10 9.3 16 14.8 108
Epistylis sp. NEMATODES
0 0 0 0 91 70.5 0 0 38 29.5 129
P. laevionchus 48 94.1 3 5.9 0 0 0 0 0 0 51
R. Congolensis CESTODES
22 100 0 0 0 0 0 0 0 0 22
P. clarias 0 0 137 100 0 0 0 0 0 0 137
M. woodlandi 159 100 1 0 0 0 0 0 0 0 159
A= No. of host infected, B = No. of parasites recovered, C = % prevalence of parasites, D = Mean intensity of parasites, E = parasites abundance

lxx
3.16 Effect of Parasite on the Haematological Profile of some Clariid Species of the
Anambra River System
The results of the haematological indices determined for both infected and uninfected species of
C. gariepinus and C. anguilaris were presented in Table 12. The results indicated vividly that
packed cell volume (PCV), Haemoglobin concentration (Hb) and Red blood cell count (RBC)
were lower in the infected than in the uninfected fishes for both species studied. However, the
WBC values were higher in the infected fishes than in the uninfected ones. Statistical analysis
revealed significant differences (P<0.05) between the heamatological indices of the infected and
uninfected clariid species of the Anambra River System.

lxxi
Table 12: Effect of Parasites on the Haematological Profile of the Infected Clariid Fishes
Clarias gariepinus
Parameters Infected Uninfected P-values
RBC (cells/mm3) 4.16±0.07 7.79±0.08 0.0001*
PCV (%) 24.61±0.32 37.56±0.23 0.0001*
Hb (g/dl) 7.03±0.11 10.86±0.11 0.0001*
WBC (cells/mm3) 1.20x104± 1.53 x 1.02 7.50x103±4.7 x 1.01 0.0001*
Clarias anguilaris
RBC (cells/mm3) 4.16±0.09 7.70±0.10 0.001*
PCV (%) 24.75±0.50 37.78±0.25 0.001*
Hb (g/dl) 7.01±0.17 10.75±0.14 0.001*
WBC (cells/mm3) 1.21x104±2.06 x 1.02 7.57x103±5.32 x 1.01 0.001*
*Significant difference (P<0.05) across the horizontal rows

lxxii
3.17 Effect of Parasites on the Biochemical Parameters of some Clariid Species of the
Anambra River System.
The liver enzyme assayed were aspartate aminotransaminase (AST), Alanine amino transferase
(ALT) and alkaline phosphatase (ALP). The serum chemical compounds analyzed were urea and
creatinine. The activities of AST, ALT and ALP were found to be significantly (P<0.05) higher
in the infected fishes than in the uninfected ones (Table 13). The levels of creatinine and urea
showed higher significant differences (P<0.05) in both infected and uninfected fish species.

lxxiii
Table 13: Effect of Parasites on the Biochemical Parameters of some Clariid Species of the
Anambra River System
Clarias gariepinus
Parameters Infected Uninfected p-values
AST (U/L) 95.85±0.36 73.13±0.38 0.001*
ALT (U/L) 130.0±1.18 93.27±0.52 0.001*
ALP (U/L) 75.92±0.37 53.03±0.32 0.001*
Creatirine (U/L) 1.62±0.02 1.02±0.01 0.001*
Urea (U/L) 38.21±0.23 24.62±0.28 0.001*
Clarias anguilaris
AST (U/L) 95.35±0.57 73.31±0.44 0.001*
ALT (U/L) 129.1±1.56 94.59±0.6 0.001*
ALP (U/L) 75.59±0.50 52.91±0.41 0.001*
Creatirine (U/L) 1.61±0.02 1.02±0.41 0.001*
Urea (U/L) 37.87±0.38 24.48±0.35 0.001*
*significant different (P < 0.05) across the horizontal rows

lxxiv
CHAPTER FOUR
DISCUSSION
The parasites recorded from the investigation are among the parasites listed by Paperna (1996) as
parasites of fishes in Africa. Protozoa, cestode and nematode parasites have been reported in
fishes of several water bodies in Nigeria (Oniye et al., 2004; Ayanda, 2009a; Eyo et al., 2013;
Eyo and Iyayi, 2014; Eyo et al., 2014).
The overall prevalence of parasites (41.1%) was low compared to the 59.2% recorded for fishes
in the Nigeria River at Illushi, Edo State (Onyedineke et al., 2010). In the same vein, the
recorded prevalence of parasites is lower when compared to the 60.3% recorded for Clarotes
laticepts in Rivers Niger-Benue confluence, Lokoja, Nigeria (Eyo and Iyayi, 2014). It is
however, higher when compared with records by other investigators in rivers from Nigeria who
reported overall parasite prevalence of 17.1% in the Osse River, 6.9% in the Okhuo River, 3.3%
in the Great Kwa River and 32.9% in the Warri River (Okaka and Akhigbe, 1999; Edema et al.,
2008; Ekanem et al., 2011; Eyo et al., 2014). These variations in rate of parasitism could be
attributed to abiotic and biotic conditions of the environments where the studies were carried out
(Eyo et al., 2013; Thompson et al., 2014). Unfavorable conditions may offset fish physiology
favoring parasite infestation and invasion. Rohlenova et al. (2011) reported that unfavorable
temperature may alter fish physiology including immune function favoring parasite invasion.
Pollution of the fish environment also contributes to parasitizing of fish significantly (Kelly et
al., 2010). Also, heavy parasitic infection in fish has been linked to environmental contamination
by different pollutants, including heavy metals and hydrocarbons (Schludermann et al., 2003;
Williams and Mackenzie, 2003) and organic enrichment of sediments by domestic sewage
(Marcogliese and Cone, 2001). Khan and Thulin (1991) and Kemp and Spotila (1997) reported

lxxv
that urban effluents promote aquatic pollution, thus, making aquatic organisms vulnerable to
increased incidence of parasites. The relatively high infection rate of fishes from Anambra River
could therefore, be attributed to the contamination of the river by various pollutants from the
numerous market women who often use the river as a means of waste disposal from Otuocha
market, since Anambra River (Otuocha station) where the fishes were sampled lies parallel to
Otuocha market.
4.1 Prevalence, Mean Intensity and Abundance of Parasites
4.1.1 Protozoan
In this study Trichodina acuta and Epistylis sp. were the protozoan parasite species found to be
parasitizing the fishes. Trichodina acuta infected 48 clariid catfishes with infection spreading
across all the species and sexes of fish under investigation. Epistylis sp. infected 44 fish hosts.
Protozoan parasites where found in all the species studied. However, the parasite have relatively
low infection rate (25.55%) when compared to the 69% prevalence recorded by Mahmoud et al.
(2011) in a tropical fresh water in Egypt.
The relatively high infection of the protozoa in the skin and gills of their fish hosts could cause
irritation and breathing problems. According to Klinger and Floyd (2002), the parasites cause
serious skin and gill irritation displayed by flashing, rubbing, rapid breathing and excessive
mucus secretion in gills. The authors also reported that the parasite have direct life cycle and
could build up to high numbers when fish are crowded causing weight loss, debilitation and
mortality. Although some of the fishes examined during this study were infected with many
protozoans with mucus secretion in their gills, they were acceptable in the market.

lxxvi
4.1.2 Cestodes
The cestode parasites recovered from the fishes during the course of this study were the
unsegmented Caryophyllaeidae, Monobothriode woodlandi and the segmented Pseudophyllidae,
Polyonchobothrium clarias. The infections of cestode parasites in the clariid fish species studied
with prevalence of 15.56% were quite variable with the two forms, Caryophyllaeidae and
Pseudophyllidae well represented. The findings had similarities with those reported from other
freshwater bodies in the tropic. Laboni et al. (2012) reported cestode prevalence as high as
83.78% from Bangladash while Barson and Avenant-oldewange (2006) reported 71% prevalence
of cestodes from the Rietvlei Dam, South Africa. However, Ayanda (2009b) reported extremely
low infection prevalence of 5% cestodes from Asa Dam River in Nigeria.
The level of infection of some clariid species of the Anambra River System by cestode parasites
could be due to the ingestion of eggs, copepods and mollusks which served as intermediate hosts
of the larval stages for the cestodes (Paperna, 1996). Clariid species were reported to feed
heavily on insects, copepods, mollusks and smaller fishes (Reed et al., 1967).
However, the fact that Polyonchobothrium clarias physically resisted detachment from the gut
mucosa suggested that the suction created by the bothria and the clasp of the apical hooks could
cause severe pathological effects in heavy infections (Barson and Avenant-oldewege, 2006).
Monobothriode woodlandi parasites were recorvered from hosts intestine. This finding is in
conformity with the observations of Barson and Avenant-Oldewage (2006); Jaywant et al.
(2010), Eyo and Iyaji (2014) and Eyo et al. (2014) who reported cestode parasites prevalence in
the intestines and glandular stomachs of their fish hosts.

lxxvii
4.1.3 Nematode
The nematode parasites recorded in this study were Procamallanus laeviconchus and
Rhabdochona congolensis. The prevalence (3.88%) of the parasites was generally low in the fish
hosts. The low prevalence of nematode parasites in the clariid species maybe as a result of the
association of namatodes with fish hosts that feeds on mud, debris and detritus as reported in
most Synodontis species (Iyayi, 2011).
4.2 Influence of Body Weight and Total Length on Parasitic Infection
The significantly higher prevalence and mean intensity of parasites recorded in fishes of large
weight and length classes (76 g+ and 30 cm+) examined compared to the smaller weight and
length classes indicated increase in parasitism with increase in size which is also related to age.
Positive correlation in parasitism with host size was reported in several studies (Chandler et al.,
1995; Brickle et al., 2003; Oniye et al., 2004; Ayanda, 2009a; Eyo et al., 2013, Eyo and Iyaji
2014; Eyo et al., 2014).
Roberts (1978) reported that bigger fishes provided large surface areas for infection than smaller
ones. Since it is not likely to be the adult stage of the parasites that the fishes would ingest
normally (that is, it is the egg or larval stages that will eventually multiply in numbers in the
fishes); it is therefore, a plausible explanation that the big fish would provide a good ground for
the parasites to multiply over time. An increase in size is a reflection of increase in length which
is considered a measure of age (Torres et al., 1977; Lagler et al., 1979). Catfishes are known to
be herbivorous at the juvenile stage with the tendency of being omnivorous at the adult stage.
The higher parasitism observed in adults over juveniles might be attributed to change in diet of
the fish from weeds, seeds, phytoplankton and zooplankton as juveniles to adult insect and larval

lxxviii
forms, snails, crustaceans, worms and smaller fishes as adulthood is attained (Reed et al., 1967;
Ayanda, 2009b).
Another plausible explanation for the higher parasitism in the adults over juveniles may not be
unconnected with activity. The adult as expected would be more active than the juveniles. As
such, they are able to compete better for food than the other age groups. This will result in more
contact with food and hence a higher tendency of getting infected with parasites.
4.3 Influence of Fish Hosts Sex on Parasitism
Parasitism in fish has been reported to be sex biased, with males suffering greater susceptibility
(Eyo et al., 2014). This sex linked parasitism has been explained as resulting from differences in
reproductive investment by male and female fish (Skarstein et al., 2001; Sinkova et al., 2008).
Immune-suppression by steroid hormone during spawning in males has been suggested as a
major factor contributing to the greater susceptibility of males to parasite invasion (Folstad and
Karter, 1992). Other factors suggested include, competition for mate (Folstad anad Karter, 1992)
and cost of territorial defense (Reinchen, 2001).
In the present study, the overall parasite prevalence in male clariid species studied i.e. C.
gariepinus and C. anguilaris (46.7% and 38.2% respectively) were higher than that of their
female counterparts with 40.7% for C. gariepinus and 32.1% for C. anguilaris. Although the
males had higher prevalence than the females, they were not significantly different (P>0.05).
Whereas, there were no significant differences (P>0.05) in the parasite abundance in both male
and female C. anguilaris, the abundance of parasites in male C. gariepinus (2.15 ± 0.22) was
significantly higher (p<0.05) than that of the infected female C. gariepinus (1.38 ± 0.19).
In this study, the male fishes have higher parasitic infections than the females. These findings are
in accordance with the observation of Folstad and Karter (1992); Poulin (1996); Wedekind and

lxxix
Jacobson (1998). These authors argued that males bear the cost of sexual selection through
competition for mates. Kennedy (1995) summarized that the quantitative difference in parasite
infection between sexes can be expected and may be explained as a consequence of different
habitats occupied by males and females as well as differences in the diets and or physiology.
4.4 Influence of Season on the Patterns of Parasite Occurrence
In this study, analysis of seasonal patterns of parasite species infection showed that while some
parasites had their highest infection rates in the dry months before the rainy season, others had
their highest infection rates during the rainy season. The seasonal pattern of higher abundance of
some parasites in the dry months before the rainy season in tropical areas has been observed in
several studies and several suggestions have been offered to explain the phenomenon. Gee and
Davey (1986) argued that the higher infection in autumn could result from life cycle of the
parasites. They pointed out that the seasonal variation in infection of Mytilicola intestinalis in
blue mussels with higher intensity of infection in late autumn was associated with the occurrence
of egg production in the parasite between June and August. The eggs produced between June and
August were hatched in summer and produced larvae that became infective adults during the
autumn and could result in increase in the intensity during autumn.
It was also argued that higher abundance of some parasites in the dry months before the rainy
season in some tropical areas could be due to an increase in host density and greater overlap of
intermediate and definitive hosts as water bodies shrink (Ezenwaji and Ilozumba, 1992) or due to
pre-spawning congregation of hosts (Gupta et al., 1984), both of which facilitate transmission.
Higher abundance of parasites during the rainy months of the year was also observed in other
studies in the tropics (Granath and Esch, 1983a, b; Marcogliese and Esch, 1989a, b). Zaman

lxxx
(1985) and Zaman and Seng (1989) reported two peaks of parasite abundance with increase in
rainfall and another peak in the dry month of January.
It could be deduced from the above that as much as other factors are important in explaining the
variations in seasonal patterns of parasite occurrence, the most important single factor could be
the life cycle of the parasites. Each of the parasite species found exhibited different patterns of
occurrence which could only be explained by the differences in their life cycle patterns.
4.5 Effect of Parasite on the Condition Factor and Hepatosomatic Index
The condition factor often referred to as “K” provides information on the wellbeing of a fish and
is usually influenced by fish sex, season, maturity stage (Anyanwu et al., 2007). The role of
condition indices as stated by Stevenson and Woods (2006) is to quantify the health of
individuals in a population or to tell whether a population is healthy relative to other populations.
When fish of a given length exhibits higher weight, it means they are in better condition (Anwa-
Udondiah and Pepple, 2011).
In this study, the condition factors of the parasitized clariid fishes were significantly different
(P<0.05) from the non-parasitized ones. It is evident that the correlation matrixes of the influence
of parasite prevalence on the K-factors of the infected fishes were positively and significantly
correlated. The reduction in the condition factor of the parasitized fishes maybe as a result of the
dependent of the parasites on the fish for its nutrition (Mehra, 1991), thereby reducing the
nutrient available for their fish hosts. Parasites are also known to act as stressors to their hosts
(Pinky et al., 2012) and as such, causing discomfort to their hosts. This finding is in agreement
with the observation of Bryan and Luca (2004) who observed a decrease in the condition factor
of bluegill sunfish (Lepomis macrochirus) due to parasite load. They concluded that condition
factor could be one of the best predictors of parasite density.

lxxxi
On the other hand, hepatosomatic index (HSI) is the general measurement of the overall
condition of fish or the growth status of the liver (West, 1990) and can be an excellent predictor
of adverse health condition in fish (Adams and McLean, 1985). In this study, there was a
positive correlation between parasite load and HSI. The HSI of the infected fishes were
significantly higher (p<0.05) than the uninfected fishes. In fact, the correlation matrix plot of the
influence of parasite prevalence on HSI tended towards unity (r = 0.913 and 0.916 for C.
gariepinus and C. anguilaris respectively). This significant increase observed in the liver weight
of the infected fishes may be as a result of the migration of the larvae of the cestode and
nematode worms into the liver of the fish, a finding that corroborated with reports by
Sommerville (1982) and Pinky et al. (2012). Also some parasites are known to utilize their host
tissues and organs for reproduction thereby making the infected cells become hypertrophic and
able to accommodate proliferating parasites (xenoma) (FAO, 1991).
Mahmoud et al. (2011) reported that parasites are known to produce intoxication in their fish
hosts. It may therefore be suggested that the increase in the liver weight of the infected fishes
may also be as a result of the increased activity of hepatic mixed-function oxidase enzyme which
developed an increased ability to metabolize xenobiotic (Fabacher and Bauman, 1985). This is
because the liver is the main and important detoxifying organ in fish and is essential for both the
metabolism and excretion of toxic substances in the body (van Dyk et al., 2007).
4.6 Effects of Parasites on the Haematological Profile of Fishes
Blood is a good bioindicator used to determine the health of an organism (Joshi et al., 2002). It
also acts as a pathological reflector of the whole body. Hence, the haematological parameters are
important in diagnosing the functional status of the fish (host) infested by parasites (Joshi et al.,

lxxxii
2002) and also to evaluate the physiological condition and nutritional state of fish (Chagas and
Val, 2003).
In the present study, all the haematological parameters studied varied significantly (p<0.05)
between the infected and the uninfected fishes. This observation supports the works of many
researchers. Sexena and Chauchan (1993) reported increase in lymphocyte count in
Heteroponeutus fossilis infected with Lucknowia indica. A higher degree of eosinophilia was
observed in Clarias batrachus carrying helminth infections (Sinha, 2010). Shabi et al. (2013)
reported decreased RBC, Hb and PCV from fishes infected with hemoparasites.
The reduction in RBC count, haemoglobin (Hb) value and packed cell volume (PCV) in the
infected fishes may be as a result of the parasitic infestation that often leads to anemia (Martins
et al., 2004). Furthermore, the parasites simply acted as stressors and during the primary stages
of stress, the PCV are altered due to the release of catecholamine (an enzyme) which mobilizes
RBCs to swell as a result of fluid entry into the intracellular compartment (Chiocchia and
Motias, 1989). Similar results were recorded by Hassen (2002) and Ismail (2003) in Clarias
gariepinus that are naturally infected with T. mukasai. Lofty et al. (2003) and Omran and El-
Kholary (2003) also reported that the severe anemia that lead to the reduction of RBC, PCV and
Hb of the infected fishes maybe as a result of chronic liver inflammation which causes
depression of erythrogenesis. On the other hand, significant increase was observed in WBC of
the infected fishes when compared to those of the uninfected fishes. According to Lebola et al.
(2001) and Hassen (2002), the increase in WBCs count occurred as a pathological response since
WBCs play a major role during infestation by stimulating the haemopoietic tissues and immune
system to produce antibodies and chemical substances which work as defense agent against
infection. Pinky et al. (2012) had a similar report.

lxxxiii
4.7 Influence of Parasites on the Biochemical Parameters of Clariid Fishes of the Anambra
River System
Enzymes catalyze physiological reactions by lowering the activation energy level that the
reactant (substrate) must reach for the reaction to occur (Kabir, 2009). The effect of
parasite/infection on enzymatic activity is one of the most important biochemical parameters,
which is affected under stress (Mohmoud et al., 2011). When an organ is diseased, enzymatic
activity appears to be increased or inhibited due to the active site being either denatured or
distorted (Madu et al., 2008). Since enzymes catalyze some steps in the metabolism of
carbohydrates and proteins, they are present in most tissues (Kabir, 2009). The increase or
decrease in their level maybe sufficient to provide information of diagnostic values.
The liver is one of the vital organs of the body, susceptible to various parasites and disease
conditions which affect the total health status of the animal. It is the center of all metabolic
processes and posses’ considerable functional reserve and regenerative capacity (Egbu, 2011).
Alkaline Phosphatase (ALP) is a membrane bound enzyme found in pinocytic vesicle and Golgi
complex. It is present on all cell membranes where active transport occurs (Kori-Sikpere et al.,
2010). Alkaline phosphatase diagnosis is important in bone and hepatobilary diseases and is thus
employed to assess the integrity of plasma membrane and endoplasmic reticulum (Akanji et al.,
1993).
The activity of the serum ALP in the parasite infected clariid species of the Anambra River
System revealed a significant increase when compared to the uninfected fishes. This finding is in
agreement with the observations of EL-Aziz et al. (2002) and Ahmed et al. (2006) who reported

lxxxiv
increase in ALP in Fasciola infected monkey. They argued that the elevation in the ALP activity
may be as a result of the penetration of the parasite into the bile duct.
Aminotransferases are gainfully used in the diagnosis of disease of tissue damage. It functions as
a link between carbohydrate and protein metabolism by catalyzing the inter conversion of
strategic compounds respectively (Martin et al., 1983). They are normally localized within the
cells of the liver, heart, gill, kidney, muscles and other organs. Their presence in the serum may
give information on organ dysfunction (Wells et al., 1986). Alanine aminotransferase is
remarkably specific for liver function since aspartate aminotransferase is mostly present in
kidney (Witthawaskid et al., 2003). The activities of AST and ALT were significantly higher
(P<0.05) in the infected fishes when compared to the uninfected fishes. A similar result was
observed by Younis (1999) who reported that AST, ALT and Urea showed significant increase
in O. niloticus infected with external potrozoa and monogenetic trematodes. Osman et al. (2009)
reported that blood serum AST, ALT enzyme activities, creatinin and Urea values were
increased in Trichodina infected Clarias gariepinus. Mahmoud et al. (2011) reported that the
blood serum AST, ALT, creatinin and urea values were elevated in the infected O. niloticus and
C. gariepinus. The increase in the activity of AST and ALT in the serum of the infected fishes
revealed that the parasites had effect on the parenchymous tissues and skeletal musculature
(Egbu, 2011), which probably may have altered the permeability (Kabri, 2009) and integrity of
cell organelles as reported by Adamu and Iloba (2008).
The pattern of ALT and AST activities observed in this study are biochemical symptoms tending
towards liver cytolysis, indicating disturbance in the structure and integrity of cell organelles like
endoplasmic reticulum and membrane transport system (Dere and Polat, 2001). Alterations in

lxxxv
their activities may have adverse effects on the amino acid metabolism of the tissues and
consequently the intermediates needed for gluconeogenesis (Kabri, 2009).
Serum urea and cretinine levels were observed to be elevated in the present study due to parasitic
infestation of fishes. Kabir and Ovie (2011) reported that creatinine leaves the muscles and enter
the blood where it is a waste product largely from the muscle breakdown. It is removed by
filtration through the glomeruli of the kidney and excreted as urine. The increase in the level of
creatinine in the infected fishes may be as a result of the alteration of the muscles structure of the
infected fishes by parasites (Egbu, 2011). Mahmoud et al. (2011) also reported elevated levels of
creatinine and urea in fishes infected with protozoa parasites.
Urea is a principal end product of protein catabolism. It is a waste product metabolized in the
liver and excreted by the kidney. The increases in the urea values in the infected fishes maybe
due to gill dysfunction (caused by protozoa mainly found in the gills) as urea are mainly excreted
through the gill (Murray et al., 1990). Also, these findings may be attributed to the inflammatory
reactions of intoxication produced by the parasites in the infected fishes (Mehra, 1991;
Mahmoud et al., 2011).
4.8 Conclusion
This study was an effort to contribute to the knowledge of parasitism of tropical fish hosts
particularly, clariid, catfishes and to determine the impact of these parasites on the physiology
and haematology of these fishes. This is also an attempt to disperse the general perception that
there is lack of adequate information on the parasite fauna of tropical fish. Protozoans,
nematodes and cestodes were found in this study. However, ecto parasites (leeches and
copepods) were not encountered during the course of this study. A plausible explanation to this
could be that the fishers might have cleaned them off before they were sold off.

lxxxvi
It could be observed from this study that while some parasites conformed to the general pattern
of higher abundance in the dry months before the rainy season, others had their highest infection
during the rainy months of the year. These differences in patterns of infection could be
influenced by differences in their life cycle patterns.
The entire study revealed that the intensity of parasitic infections maybe responsible for
alteration of the haematology of fish hosts. It was further speculated that mechanical damage
caused by Polyonchobothrium clarias to the host gut mucosa could have caused vitamin B-12
and folic acid deficiency which were otherwise responsible for RBC maturation. The mechanical
injury may have also led to side tracking of iron to affected tissues which is otherwise
responsible for enthropoiesis. The increased number of WBC values maybe associated with the
defense mechanism and immunological responses against infectious diseases caused by
parasites.
From this study, the elevation in enzyme activities of the infected fishes could be attributed to
the destruction of the liver parenchyma by the parasites which led to the leakage of the liver
enzymes into the general circulation. The invasion of the liver cells by parasites caused organ
congestion, sinusoidal blockage and cellular inflammation. When these happen, the parenchyma
(transaminases) and membranous (alkaline phosphatase and gamma glutamyl transpepetidse)
enzymes of the liver leak out into the circulation, leading to increased enzymes activity.
4.9 Recommendations
• It is recommended that studies that cut across parasite groups and several fish species
with emphasis on life cycle be undertaken to improve our understanding of the factors
that affect seasonal patterns of parasite occurrence.

lxxxvii
• The pollution status and heavy metals profile of the Anambra River System should be
investigated so as to improve our understanding of the interplay between pollutants and
incidence of parasite occurrence of the river
• Further studies on the impact of parasites on the histology of the fishes of the river should
be encouraged so as to understand the extent of damage caused by parasites to the
various organs of fishes.

lxxxviii
REFERENCES
Abidemi – Isomini, A. O. and Eze, R. N. (2011). Comparative assessment of parasite infestation of Tilapia in natural and cultured environment. In: Lipings, L. and Fitzsimanons, K. (Editors). Better Science, Better Fish, Better life (Proceedings of the Ninth International Symposium on Tilapia in Aquaculture. Aquafish Collaborative Research Support Program.
Abo-Esa, J. F. K. (2008). Study on some ectoparasitic diseases of catfish, Clarias gariepinus with their control by Zingiber officiale. Mediterranean Aquaculture Journal, 1(1): 1 – 9.
Abolagba, O. J. and Omorodion, G. O. (2006). Field survey of fish pond management practices in Delta State, Nigeria. Tropical Freshwater Biology, 15: 43 – 54.
Abowei, J. F. N. and Ezekiel, E. N. (2011). A review of Myxosporea, Microspora and Monogenea infections in African fish. British Journal of Pharmacology and Toxicology, 2(5): 236 – 250.
Adams, S. M. and McLean, R. B. (1985). Estimation of largemouth bass, Micropterus salmoides, lacepede, growth using liver somatic index and physiological variables. Journal of Fish Biology, 26: 111 – 126.
Adamu, K. M. and Iloba, K. I. (2008). Effect of sublethal concentrations of porland cement powder solution on the amino tranferases of the African catfishes {Clarias gariepinus (Burchell, 1822)}. Acta Zoologica Lituanica, 18(1): 50 - 54.
Adeyemo, A. O. and Daunemughan, T. (2012). Parasitic investigation on wild Parachanna obscura. International Journal of Current Biological and Medical Science, 2(1): 195 – 198.
Agbamu, J. U. and Orhorhoro, W. C. (2007). Adoption of aquaculture management techniques in Delta State. Niger Agriculture Journal, 38: 125 - 134.
Ahmed, M. I., Ambali, A. G. and Baba, S. S. (2006). Haematological and biochemical responses of Balami sheep to experimental Fasciola gigantic infection. Journal of Food, Agriculture and Environment, 4(2): 71 – 74.
Ahne, W. (1985). Argulus foliaceaus L. and Piscicola geometra L. as mechanical vector of Spring Viraemia Carp Virus SVCV. Journal of Fish Diseases, 8: 241 – 242.
Akanji, M. A., Olagoke, O. A. and Oloyede, O. B. (1993). Effect of chronic consumption of metabisulphite on the integrity of the rat kidney cellular system. Toxicology, 81: 173 – 179.

lxxxix
Akinsanya, A. A., Hassan, A. A. and Adeogun, A. O. (2008). Gastrointesitnal helminth parasites of the fish Synodontis clarias (Siluriformes: Mochokidae) from Lekki Lagoon, Lagos, Nigeria. Revista de Biologia Tropical, 56(4): 2021 – 2026.
Akinsanya, B. and Otubanjo O. A. (2006). Helminth parasites of Clarias gariepinus (Clariidae) in Lekki Lagoon, Lagos, Nigeria. Revista de Biologia Tropical, 54(1): 93 - 99.
Akinsanya, B. Otubanjo, O. A. and Ibidapo, C. A. (2007). Helminth bioload of Chrysichthys nigdigitatus (Lace pede 1802) from Lekki Lagoon, Lagos, Nigeria. Turkish Journal of Fisheries and Aquatic Sciences, 7: 83 – 87.
Akinsanya, B., Hassan, A. A. and Ibidapo, C. A. (2010). The parasitic helminth fauna of Parachanna obscura from Lekki Lagoon, Lagos, Nigeria. Researcher, 2(9): 78 – 83.
Alvarez-Pellitero, P. (2004). Report about Fish Parasitic Disease. Consejo Superior de Investigaciones Cientificas (CSIC). Instituto de Acuiculture Torre dela sal, Ribera de Cabanes, Spain.
Alvarez-Pellitero, P. (2008). Fish Immunity and parasite infection: from innate immunity to immunoprophylatic prospects. Veterinary Immunology and Immunopathology, 126: 171 – 198.
Anwa-udondiah, E. P. and Pepple, P. C. G. (2011). Length-weight relationship and condition factor of black chin tilapia (Sarotherodon mealnotheron) cultured in sheltered outdoor tanks. In: Kolo, R. J. and Orire, A. M. (Editors). Proceedings of of the 26th Annual Conference of the Fisheries Society of Nigeria (FISON). Minna, 28th November – 2nd December, 98 – 102.
Anyanwu, P. E., Okoro, P. C., Anyanwu, A. O., Matanmi, M. A., Ebonwu, B. I., Ayabu-Cookey, I. K., Hamzat, M. B., Ihumekpen, F. and Afolabi, S. E. (2007). Length-weight relationship, condition factor and sex ratio of African mud catfish (Clarias gariepinus) reared in indoor water recirculation system tanks. Research Journal of Biological Sciences, 2(7): 780 – 783.
Arthur, J. R. and Albert, E. (1994). A survey of the Greenland halibut Reinhardtius hippoglossiodes caught off Atlantic Canada, with notes on their zoogeograohy in this fish. Canadian Journal of Zoology. 72: 765 – 778.
Atanda, A. N. (2007). Freshwater Fish Seed Resource in Nigeria, pp. 361 – 380. In: Bondad-Reantaso M. G. (Editor). Assessment of Freshwater Fish Seed Resources for Sustainable Aquaculture. Food and Agriculture Organization (FAO) Fisheries Technical Paper. 501. Rome, FAO.

xc
Awachie, J. B. E and Ezenwaji, H. M. G. (1981). The Importance of Clarias species in the Fisheries Development of the Anambra River Basin Nigeria. CIFA Technical Paper.
Awachie, J. B. E. (1966). Preliminary notes on the parasites of fish in the area of Kainji reservoir. In the first scientific report of the Kainji. Biological. Resource Team, 1: 65 - 69.
Awachie, J. B. E. and Hare, L. (1977). The fisheries of Anambra, Ogun and Oshun river systems in Southern Nigeria. CIFA Technical Papers, 5: 170 - 184.
Awharitoma, A. O. and Ehigiator, F. A. R. (2012). Helminth parasites of fishes from some rivers in Southern Nigeria. African Scientist, 3(2): 65 - 69.
Ayanda, O. I. (2009a). Comparative parasitic helminth infection between cultured and wild species of Clarias gariepinus in Ilorin, North-Central Nigeria. Scientific Research and Essay, 4(1): 18 – 21.
Ayanda, O. I. (2009b). Comparison of parasitic health infection between the sexes of Clarias gariepinus from Asa dam Ilorin, north-central Nigeria. Scientific Research and Essay, 4(4): 357 - 360.
Azugo, W. I. (1978). Ecological Studies of the Helminth Parasites of the Fish of Anambra River System. M. Phil Thesis, University of Nigeria Nsukka, 178 pp.
Babatunde, B. O. and Aminu, R. (2004). Field Guide to Nigerian Freshwater Fishes. Federal College of Freshwater Fisheries Technology, New Bussa, Nigeria.
Baker, F. I., Silverton, R. E. and Luck Cock, E. D. (1975). Introduction to Medical Laboratory Technology. Fourth Edition. Butterworth and Co. Ltd London.
Barson, M. and Avenant-Oldemage, A. (2006). On cestode and digenean parasites of Clarias gariepinus (Burchell, 1822) from the Rietvlei Dam, South Africa. Onderstepoort Journal of Veterinary Research, 73: 101 – 110.
Bauer, O. (1959). The influence of environmental factors on the reproduction of fish parasite. Vapr. Ekoi (Izdat, Kiev University), 3: 132 - 141.
Bichi, A. H. and Dawaki, S. S. (2010). A Survey of the ectoparasites on the gills, skin and fins of Oreochromis niloticus at Bagauda fish farm, Kano, Nigeria. Bayero Journal of Pure and Applied Science, 3(1): 83 – 86.
Bichi, A. H. and Yelwa, S. I. (2010). Incidence of piscine parasites on the gill and gastrointestinal tract of Clarias gariepnis (Teugels) at Bagauda fish farm, Kano. Bayero Journal of Pure and Applied Sciences, 3(1): 104 – 107.

xci
Blaxhall, G. and Diabley, N. (1973). Routine haematological methods for use with fish blood. Journal of Fish Biology, 5: 771 – 941.
Brickle, P. Buxton, N. G. and Villalon, E. (2003). Infection of Sphyrion laevigatum (copepod: sphyridae) on Genypterus blacodes (pisces: Ophidiidae) from the Falkland islands, South Atlantic. Parasitology, 89(2): 242 - 244.
Bryan, D. N. and Luca, M. C. (2004). Relationship between condition factors, parasite load and pertanity in bluegill sunfish, Lepomis macrochirus. Environmental Biology of Fishes, 71: 297 – 304.
Burtis, C., Ashwood, E. and Border, B. (2001). Liver functions. In: Tietz A. (Editor). Fundamentals of Clinical Chemistry. 5th Edition. Sunders Company.
Chagas, E. C. and Val, A. L. (2003). Effeito da vitmina C no ganho de peso e em parametros hematologicos de tambaqui. Pesquisa Agropecuaria Brasileira. Brasilia, 38(3): 397 – 402.
Chambrier, A. and Vaucher, C. (1999). Proteocephalidea et Monticelliidea (Eucestoda: Proteocephalidea) parasites de poisons d’eau douce au Paraguay, avec descriptions d’un genre nouveau et dix especes nouvelles {Proteocephalidae and Monticelliidae (Eucestoda: Proteocephalidae parasites of freshwater fishes in Paraguay, with descriptions of a new genus and ten new species}. Revue Suisse De Zoologie, 106: 165 – 240.
Chandler, M., Chapman, L. J. and Chapman, C. A. (1995). Patchiness in the abundance of metacercariae parasitizing Poecilla gilli (Poeclidae) isolated in pools of an intermittent tropic stream. Environmental Biology of Fishes, 42: 313 - 321.
Chocchia, G. and Motais, R. (1989). Effect of catecholamines on deformability of red blood cells from trout: relative roles of cyclic AMP and cell volume. Journal of Physiology, 412, 321 – 332.
Choudhury, M and Dick, T. A. (2000). Richness and diversity of helminth communities in tropical freshwater fishes: empirical evidence. Journal of Biogeography. 27: 935 – 956.
Coles, E. H. (1986). Veterinary Clinical Pathology. Fourth Edition. W. B. Saunders Company, Philadelphia.
Cornet, S., Franceschi, N., Bauer, A., Rigand, T. and Moret, Y. (2009). Immune depression induced by acanthocephalan parasites in their intermediate crustancean host: consequences for the risk of super-infection and link with host behavioural manipulation. International Journal of Parasitology, 39: 221 - 229.
Cox, F. E. G. (1993) Modern Parasitology. Second Edition. Blackwell Science, Oxford, London.

xcii
D’ Silva, J., Bhuyan, A. I. and Bristow, A. G. (2012). Distribution of helminth parasites in size groups and ortgans of hilsa shad, Tenualosa ilisha. Dhaka Univiversity Journal of Biological Science, 21(1): 55 – 65.
Dere, E. and Polat, F. (2001). The effect of paraquat on the activity of some enzymes in different tissues of mice (Mus musculus). Turkish Journal of Biology, 25: 323 – 332.
Dove, A. D. M. and O’donoyhue, P. J. (2005). Trichodinids (Ciliophora: Trichodinidae) from native and exotic Australian Freshwater fishes. Acta Protozoologica, 44: 51 – 60.
Echi, P. C., Eyo, J. E. and Okafor, F. C. (2009a). Co-parasitism and morphormetrics of three clinostomatids Digenea: Clinostomatidae; in Sarotherodon melanotheron from a tropical freshwater lake. Animal research International, 6(2): 982 – 986.
Echi, P. C., Okafor, F. C. and Eyo, J. E. (2009b). Co-parasitism and morphormetrics of three clinostomatids Digenea: Clinostomatidae; in Tilapia guinensis Bleeker, 1862 from Opi lake, Nigeria. Bio-Research, 7(1): 432 – 436.
Egbu, F. M. (2011). Prevalence of Bovine Edema, C. U., Okaka, C. E., Oboh, I. P. and Okogub, B. O. (2008). A preliminary study of parasitic infection of some fishes from Okhuo River, Benin City, Nigeria. Internationl Journal of Biomedical and Health Sciences, 4(3): 107 - 112.
Cysticercosis and Fascioliasis in Slaughtered Cattle in Nsukka Zone and the Haematological, Biochemical and Histopatological Changes in Response to Active Infection. M. Sc. Thesis. University of Nigeria, Nsukka, Nigeria.
Eissa, I. A. M., Viola, H. Z., Nadia, G. M. A. and Mona, S. Z. (2012). Studies on prevailing cestodiasis in wild African catfish, Clarias gariepinus at Kafr El-Sheckh governorate. Life Science Journal, 9(3): 506 – 511.
Ekanem, A. P., Eyo, V. O. and Sampson, A. F. (2011). Parasites of landed fish from create Kwa River, Calabar, Cross River State, Nigeria. International Journal of Fisheries and Aquaculture, 3(12): 225 – 230.
El-Aziz, M. Z. A., Emara, S. A. and Salem, F. S. (2002). Clinopathological studies on fasciolaisis among sheep in Giza province. Egyptian Journal of Veterinary Science, 36: 75 – 86.
Eli, A., Briyai, O. F. and Abowei, J. F. N. (2011). A review of some parasitic diseases of African fish gut lumen Protozoa, Coccidioses, Cryptosporidium infections, Haemoprotozoa, Haemosporidia. Research Journal of Applied Sciences, Engineering and Technology, 4(11): 1438 – 1448.

xciii
El-Seify, M. A., Zaki, M. S., Desouky, A. R. Y., Abbas, H. H., Abdel Hady, O. K. and Zaid, A. A. A. (2011). Seasonal variation and prevalence of some external parasites affecting freshwater fishes reared at Upper Egypt. Life Science Journal, 8(3): 397 – 400.
Esch, G. W. (1982). Abiotic factors: an overview. Pages 279 - 288. In: Mettrick, D. F. and Desser, S. S. (Editors). Parasites; their World and Ours. Proceedings of the Fifth International Congress of Parasitology Toronto, Canada, 7 - 14 August, 1982. Elsevier Biomedical Press, Amsterdam.
Eyo, J. E and Akpati, C. I. (1995). Fishing gears and fishing methods. Pages 143 – 167. In: Ezenwaji, H. M. G., Inyang, N. M. and Orji, E. C. (Editors). Proceeding of the UNDP Assisted Agriculture and Rural Development Programme (Ministry of Agriculture Awka, Anambra State). Training Course on Artisanal Fisheries Development, Held at University of Nigeria, Nsukka, October 29-November 12, 1995.
Eyo, J. E. and Iyaji, F. O. (2014). Parasites of Clarotes laticeps (Ruppell, 1832, Silurifomes, Bagridae) at River Niger-Benue confluence, Lokoja, Nigeria. Journal of Fisheries and Aquatic Science, 3: 1 – 9.
Eyo, J. E. and Iyaji, F. O. (2014). Parasites of Clarotes laticeps (Ruppell, 1832, Silurifomes, Bagridae) at River Niger-Benue confluence, Lokoja, Nigeria. Journal of Fisheries and Aquatic Science, 3: 1 – 9.
Eyo, J. E. and Mgbenka, B. O. (1992). Aspect of the biology of Clarias gariepinus in Anambra river basin. 1. Oocyte diameter, fecundity and Sex ratio. Journal of Agricultural Science Technology, 2(1): 47 - 51.
Eyo, J. E., Ejere, V. C., Aguzie, O. I., Ivoke, N., Eke, F. N., Ezenwaji, N. E. and Onoja, U. S. (2014). Parasitofauna of five freshwater fishes in a Nigerian freshwater ecosystem. Croatian Journal of Fisheries, 72: 1 – 16.
Eyo, J. E., Iyaji, F. O. and Obiekezie, A. I. (2013). Parasitic infestation of Synodontis batensoda (Ruppell, 1832, Silurifomes, Mockokidae) at River Niger-Benue confluence, Nigeria. African Journal of Biotechnology, 12(20): 3029 – 3039.
Ezenwaji, H. M. G. (1999). The abundance and trophic biology of Clarias albopunctatus (Nichols and La Monte, 1953). (Osteichthyes: Clariidae) in the Anambra flood river system. Nigerian Ecological Freshwater, 8: 22 - 25.
Ezenwaji, H. M. G. (2002). The biology of Clarias ebriensis Pellegrin, 1920 (Osteichthyes: Clariidae) in an African rain forest river basin. Fisheries Resources, 54: 235 - 252.

xciv
Ezenwaji, H. M. G. and Ilozumba, P. C. O. (1992). Helminth fauna of four western African small Clarias species (Osteichthyes: Claridae) from Nigeria. Journal of African Zoology, 106: 391 - 400.
Ezenwaji, H. M. G. and Inyang, N. M. (1998). Observations of the biology of Clarias agboyiensis Sydenham. 1980 (Osteichthyes: Clariidae) in the Anambra river flood system. Nigerian Fisheries Resources, 36: 47 - 60.
Ezeri, G. N. O. (2002). Infection by larval cestodes of the genus Paradilepis in Oreochromis niloticus L. Journal of Aquatic Sciences, 17(1): 60 - 62.
Fabacher, D. L. and Baumann, P. C. (1985). Enlarged liver and hepatic microsomal mixed-function oxidase components in tumor-bearing brown bullheads from a chemically contaminated river. Environmental Toxicology and Chemistry, 4: 703 – 710.
Folstad, I. and Karter, A. J. (1992). Parasies, bright males, and immunocompetence handicap. American Naturalist, 139: 603 – 622.
Fomena, A., Folefack, G. B. L. and Bouix, G. (2008). Three new species of Henneguya (Myxozoa: Myxosporea), parasites of freshwater fishes in Cameroon (Central Africa). Journal of Afrotropical Zoology, 4: 93 – 103.
Food and Agriculture Organization of the United Nations (FAO) (1991). Diagnostics, Prevention and Therapy of Fish Diseases and Intoxications. In: Svobodova, Z. and Vykusova, B., (editors). Manual for international Training Course on Fresh-water Fish Diseases and Intoxifications: Diagnostics, Prophylaxis and Therapy. Research Institute of Fish Culture and Hydrobiology Vodrary, Czechodovakia.
Fomena, A., Folefack, G. B. L. and Tang, I. C. (2007). New Species of Myxobolus (myxosporea: Myxobolidae) Parasites of freshwater fishes in Cameroon (Central Africa). Journal of Biological Sciences, 7(7): 1171 – 1178.
Food and Agriculture Organization (2006). The State of World Fisheries and Aquaculture. FAO Fisheries Technical paper, No. 500. http://www.fao.org/docrep/fao. Accessed on 7th July, 2012.
Food and Agriculture Organization of the United Nations (1990). National Review for
Aquaculture Development in Africa. Nigeria FAO Fisheries Circular No. 770.29. Food and Agriculture Organization of the United Nations (1991). Diagnostics, prevention and
therapy of fish diseases and intoxications. In: Svobodova, Z. and Vykusova, B. (Editors). Manual for International Training Course on Fresh-water Fish Diseases and

xcv
Intoxifications: Diagnostics, Prophylaxis and Therapy. Research Institute of Fish Culture and Hydrobiology Vodrary, Czechodovakia.
Gbankoto, A., Pampoulie, C., Marques, A. and Sakiti, G. N. (2001). Occurrence of myxosporean
parasite in the gills of two Tilapia species from Lake Nokoue (Benin, West Africa): effect of host size and sex, and seasonal patterns of infection. Diseases of Aquatic organisms, 44: 217 – 222.
Gee, J. M. and Davey, J. T. (1986). Experimental studies on the infestation of Mytilus edulis (L.) by Mytilicola intestinalis Steuer (Copepoda, Cyclopoida). Journal du Conceil, Conceil International pour 19 Exploration de la Mer, 42: 265 – 271.
Granath, W. O. Jr. and Esch, G. W. (1983a). Seasonal dynamics of Bothriocephalus
acheilognathi in ambient and thermally altered areas of North Carolina cooling reservoir. Proceedings of the Helminthological Society Ofhington, 50: 205 – 218.
Granath, W. O. Jr. and Esch, G. W. (1983b). Temperature and other factors that regulate the
composition and infra population densities of Bothriocephalus acheilognathi (Cestoda) in Gambusia affinis (Pisces). Journal of Parasitology, 69: 1116 – 1124.
Gupta, A. K., Niyogi, A., Naik, M. L. and Argawal, S. M. (1984). Population dynamics of endohelminths of Channa punctatus at Raipur, India. Brazilian Journal of Biology, 63(2): 261 – 268.
Hamish, D. R. (2010). Fish Disease Manual. Vet-Aqua International, Ireland.
Hassan, A. A., Akinsanya, B. and Adegbaju, W. A. (2010). Impacts of helminth parasites on Clarias gariepinus and Synodontis clarias from Lekki Lagoon, Lagos, Nigeria. Report and Opinion, 2(11): 42 – 48.
Hassen, F. E. Z. M. (2002). Studies on Diseases Caused by Henneguya Infection. Ph.D. Thesis, Faculty of Veterinary Medicine, Sues Canal University, Egypt.
Haussament, T. U. (1977). Determination of serum alkaline phosphatase. Clinical Chemistry, 35: 271 – 273.
Ho, J. S. (1998). Cladistic of the Lernaeidae (Cyclopoida), a major family of freshwater fish parasites. Journal of Marine Systems, 15(1-4): 177 – 183.
Hoffman, G. L., Lando, M., Camper, J. E., Coats, D. W., Stookey, J. L. and Burek, J. D. (1975). A disease of freshwater fishes caused by Tetrahymena curlissi Thompson, 1955, and a key for identification of holotrich ciliates of freshwater fishes. The Journal of Parasitology, 61(2) 217 – 233.

xcvi
Husdan, H. and Rapoport, A. (1968). Estimation of creatinine by Jafee reaction. Clinical Chemistry, 14: 222 – 226.
Hussein, A. N. A., Rabie, S. A., Mohammed, N. E. and Hussein, N. M. (2010). Light and Scanning Electron Microscopic studies of trypanosomes parasites infecting freshwater fishes in Qena Governarate, Egypt. Egyptian Academic Journal of Biological Science, 2(1): 17 – 31.
Ibiwoye, T. I. I., Balogun, A. M., Ogunsusi, R. A. and Agbontate, J. J. (2004). Determination of the infection Densities of Eustrongylides in mudfish, Clarias gariepinus and C. anguilaris from Bida Floodplain of Nigeria. Journal of Applied Science and Environmental Management, 8: 39 - 44.
Ibraq, K. and Fayaz, A. (2012). Impact of helminth parasites o haematological profile of fishes of Shallabugh wetland, Kashmir. Trend in Fisheries Research, 1(1): 2319 – 2324.
Idodo-Umeh, G. (2003). Freshwater Fishes of Nigeria (Taxonomy, Ecological Note, Diet and Utilization). Idodo-Umeh Publishers, Edo State, Nigeria.
Ismail, G. A. E. (2003). Histopatological and Physiological Studies on Naturally Infected Catfish, Clarias gariepinus with Trypanosomes. M.Sc. Thesis, Faculty of Science, Cairo University, Egypt.
Iyaji, F. (2011). Parasites of Siluriformes at River Niger-Benue Confluence, Nigeria. Ph.D. Thesis, University of Nigeria, Nsukka, Nigeria.
Iyaji, F. O. and Eyo, J. E. (2008). Parasites and their freshwater fish host. Bio-Research, 6(1): 328 - 338.
Iyaji, F. O., Etim, L. and Eyo, J. E. (2009). Parasite assemblages in fish hosts. Bio-Research, 7(2): 561 - 570.
Jarike, A. E., Emuveyon, E. E. and Idogun, S. F. (2002). Pitfall in the interpretation of liver parenchymal and membranous enzyme results. Nigerian Clinical Medicine, 10(2): 21 – 27.
Jaywant, D., Sushil, J., Somnath, W. and Ramrao, C. (2010). Survey of helminth parasites on freshwater fishes from Marathwada region, MS, India. Journal of Fisheries and Aquaculture, 1(1): 1 - 7.
Jean-Francois, G. and Alain, L. (1991). Dactylogyrid (Platyhelminthes: Monogenea) of Labeo (Teleostei: Cyprinidae) from West African coastal rivers. Journal of Helminthological Society of Washington, 58(1): 85 – 99.

xcvii
Joshi, P. K., Bose, M. and Harish, D. (2002). Changes in certain haematological parameters in suliroid catfish, Clarias batrachus (L.) exposed to cadmium chloride. Pollution Resources, 21(2): 119 – 122.
Kabata, Z. (1985). Parasites and Diseases of Fish Cultured in the Tropics. Taylor and Francis, London.
Kabir, M. A. (2009). Sublethal Effects of tobacco (Nicotiana tobaccum) leave dust on enzymatic activities of Heteroclarias (a hybrid of Heterobranchus bidorsalis and Clarias gariepinus). Jordan Journal of Biological Sciences, 2(4): 151 – 158.
Kabir, M. A. and Ovie, K. (2011). Effects of sublethal concentrations of tobacco (Nicotiana tobaccum) leaf dust on some biochemical parameters of hybrid (Clarias gariepinus and Heterobranchus bidorsalis). Brazilian Archive of Biology and Technology, 54(1): 183 – 196.
Kelly, D. W., Poulin, R., Tompkins, D. M. and Townsend, C. R. (2010). Synergistic effects of glyphosate formulation and parasite infections and survival. Journal of Applied Ecology, 47: 498 - 504.
Kemp, S. J. and Spotila, J. R. (1997). Effect of urbanization on brown trout, Salmo trutta, other fishes and microinvertebrates in valley creek, Valley Forge, Pennsylvania. American Milland Nature, 138: 56 – 67.
Keneddy, C. R. (1995). Richness and diversity of macro parasite communities in tropical eels Anguilla reinhardtii in Queens land, Australia. Parasitology, 3: 233 – 245.
Khalil, L. F. (1971). Checklist of the Helminth Parasites of African Freshwater Fishes. Technical Communication, Number 42, Comm. Inst. Helm, CAB, England.
Khan, R. A. and Thaulin, J. (1991). Influence of pollution on parasites of aquatic animals. Advanced Parasitology, 30: 201 – 238.
Klinger, R. E. and Floyd, R. F. (2002). Introduction to Freshwater Fish Parasites. Florida Cooperative Extension Service. Institute of Food and Agricultural Sciences. University of Florida. http://edis.ifasufl.edu. Accessed 20th May, 2014.
Klinger, R. E. and Francois–Floyd, R. (2009). Introduction to Freshwater Fish parasites. http://edis.ifas.ulf.edu/CIR716. Accessed 12th April, 2014.
Kori-Siakpere, O., Ikomi, R. B. and Ogbe, M. G. (2010). Variation in acid phosphatase and alkaline phosphatase activities in the plasma of the African catfish: Clarias gariepinus exposed to sublethal concentrations of potassium permanganate. Asian Journal of Experimental Biologica Science, 1(1): 170 – 174.

xcviii
Koskivaara, M. (1992). Environmental factors affecting monogeneans parasite on freshwater fishes. Parasitology Today, 8(10): 339 – 342.
Kostoingue, B., Biebakate, C., Faye, N. and Toguebaye, B. S. (2001). Presence of Myxosporidea (Myxozoa: Myxosporea) of the genus Henneguya Thelohan, 1892 in freshwater fishes from Chad (Central Africa). Acta Protozoologica, 40: 117 – 123.
Kunz, A. K. and Pung, O. J. (2004). Effect of Microphallus turgididus (Trematoda: Microphallidae) on predation, behavior and swimming stamina of the grass shrimp Palaemonetes pugio. Journal of Parasitology, 90 (3): 441 – 445.
Laboni, N. N., Chandra, K. J. and Chhanda, M. S. (2012). Effects of Caryophyllaeid cestode infestations on Clarias batrachus (Linn.). Bangladesh Journal of Fisheries Resources, 38(2): 135 – 144.
Lafferty, K. D. (2008). Ecosystem consequences of fish parasites. Journal of Fish Biology, 73: 2083 – 2093.
Lagler, K. F., Bardach, J. E. and Miller, R. R. (1979). Ichthyology. John Wiley, New York.
Lagrue, C., Kelly, D. W., Hicks, A. and Poulin, R. (2011). Factors influencing infection patterns of trophically transmitted parasites among a fish community: host diet, host-parasite compatibility or both? Journal of Fish Biology, 79: 406 – 485.
Lebola, S. L., Saunders, D. K. and Crawford, T. G. (2001). Observation on blood viscosity in striped bass, Morone saxatilis (Walbaum) associated with fish hatchery condition. Kansas Academy of Science, 104: 183 – 184.
Lewis, N. M. and Morris, D. (1986). Toxicity of nitrite: a review. Transactions of the American Fisheries Society, 115: 183 – 195.
Lochmiller, R. L. and Deerenberg, C. (2000). Trade-offs in evolutionary immunology: just what is the cost of immunity? Oikos Journal, 88: 87 – 98.
Lofty, H. S., Mahmoud, S. M. and Abdel-Gawad, M. A. (2003). Some studies on Fascioliasis in Mid-Egypt. Agricultural Research, 81(2): 209 – 227.
Lom, J. and Dykova, I. (2006). Myxozoan genera: definition and notes on taxonomy, life-cycle terminology and organic species. Folia Parasitologica, 53: 1 – 36.
Lyndon, A. R. and Kennedy, C. R. (2001). Colonization and extinction in relation to competition and resource partitioning in acanthocephalans of freshwater fishes of the British Isles, Folia Parasitologica, 48: 37 – 46.

xcix
Mahmoud, A. E., Mona, S. Z., Abdel, R. Y., Dsouky, H. A., Osman, K. A. H. and Ottia, A. A. Z. (2011). Study on clinopathological and biochemical changes in some freshwater fishes infected with external parasites and subjected to heavy metals pollution in Egypt. Life Science Journal, 8(3): 401 – 405.
Mandoza, Franco, E. F., Scholz, T., Vivas-Rodriguez, C. and Vargas-Vazquez, J. (1999). Monogeneans of freshwater fishes from cenotes (sinkholes) of the Yucatan Peninsula, Mexico. Folia Parasitologica, 46: 267 – 273.
Marcogliese, D. J. (2002). Parasites of Fishes in Freshwater. Ecological Monitoring and Assessment Network (EMAN) Parasitology Module Steering Committee, Parasitology Section, Canadian Society of Zoologist. Http:/www.eman.rese.ca/eman/ecotools/ protocols/freshwater/parasites/intro.htm. Accessed on June, 2014.
Marcogliese, D. J. and Cone, D. K. (2001). Structure of Myxozoan communites parasitizing Notropis hudsonius (Cyprinidae) at selected localities of the St. Lawrence River, Quebec: possible effects of urban effluents. Journal of Parasitology, 87: 951 - 956.
Marcogliese, D. J. and Esch, G. W. (1989a). experimental and natural infection of planktonic and benthic copepods by Asian tapeworm, Bothriocephalus acheilognathi. Proceedings of Helminthological Society Ofhington, 56: 151 – 155.
Marcogliese, D. J. and Esch, G. W. (1989b). Alteration in seasonal dynamics of Bothriocephalus acheilognathi in a North Carolina cooling reservoir over a seven year period. Journal of Parasitology, 75: 151 – 155.
Marcogliese, D. J. and Parasitology Module Steering Committee (PMSC) (2011). Protocols for Measuring Biodiversity: Parasites of Fishes in Freshwater (Update). Protocols Manual for Water Quality Sampling in Canada. Ecological Monitoring and Assessment Network (EMAN).
Marina, H. (2008). Parasites of Native and Exotic Freshwater Fishes in the South-West of Western Australia. A Thesis Presented for the Degree of Doctor of Philosophy. Murdoch University, Perth, Western Australia.
Marina, H., Beatty, S. J., Morgan, D. L. Doupe, R. G. and Lymbery, A. J. (2008). An Introduced parasite, Lernaea cyprinacea L., found on native freshwater fishes in the South West of Western Australia. Journal of the Royal Society of Western Australia, 91: 149 – 153.
Martins, M. L., Tavares-Dias, M., Fujimoto, R. Y., Onaka, E. M. and Nomura, D. T. (2004). Haematological alterations of Lepomis macrocephalus (Osteichthyes: Aniskidae) in fish pond. Arq. Brasileiro Medical and Veterinary Zootenecnia, 56: 640 – 646.
Mehra, H. R. (1991). The Fauna of India and the Adjacent Countries. Vol- 1.

c
Mgbenka, B. O. and Eyo, J. E. (1992). Aspects of the biology of Clarias gariepinus in Anambra river basin. 2. Maturation and condition factor of Aquaculture. Science Technology, 2(1): 52 - 55.
Molnar, K., Avenant-Oldewage, A. and Szekely, C. (2004). A Survey of coccidian infection of freshwater fishes in South Africa, with the description of Goussia anopli n. sp. (Apicomplexa. Eimeriidae). Systematic Parasitology, 59: 75 – 80.
Moravec, F. (1998). Nematodes of Freshwater Fishes of the Neotropical Region. Academia, Prague, Czech Republic.
Moyo, D. Z., Chimbria, C. and Yalala, P. (2009). Observations on the helminth parasites of fish in Insukamini Dam, Zimbabwe. Research Journal of Agriculture and Biological Sciences, 5(5): 782 - 785.
Munoz, G. and Cribb, T. H. (2005). Infracommunity structure of parasites of Hemigymnus melapterus (Pisces: Labridea) from Lizard island, Australia: The importance of habitat sand parasite body size. Journal of Parasitology, 91: 38 – 44.
Munoz, G., Valdebenito, V. and George-Nascimento, M. (2002). La deita y la fauna de parasitoz metozoos del torito Bovichthys chilensis egan 1914 (Pisces: Bovichthydae) on la costa de chile centrosur: Variaciones georgraficas y ontogeneticas. Rivista Chilena de HIstoria Natural, 75: 661 - 671.
Murray, R. K. D., Rranne, P. A. M. and Rodwell, V. W. (1990). Harper's Biochemistry. Norwalk, Connecticut/ Los Altos, California.
Nadia, M. (1991). Studies on Round Worms and Cestodes of Some Freshwater Fish. M. V. Sc. Thesis. Faculty of Veterinary Medicine Alexander University.
Nikolic, V. P. and Simonovic, P. D. (1996). Occurrence of parasitic ciliates (Protozoa) on perch (Perca fluviatilis) in Lake Vlasinsko. Annales Zoologici Finnici, 33: 707- 710.
Nmor, J. C., Egwunyenga, A. O. and Ake, J. E. G. (2004). Observation of the intestinal helminths parasites of cichlids in the upper reaches of River Orogodo, a freshwater body in Delta State, Southern Nigeria. Tropical Freshwater Biology, 13: 131 – 136.
Nnadi, E. I., Awi-Waadu, G. D. B., Imafidor, H. O. and Obiajuru, I. O. C. (2011). Association between parasitic infection and fish habitat. Journal of Research in National Development, 9(1): 186 - 190.
Nwani C. D. (1998). Aspects of the Biology of Distichondus sp in Anambra River, Nigeria. M.Sc. Thesis Dept. of Zoology, University of Nigeria Nsukka, p. 133.

ci
Nwani, C. D. (2004). Aspects of the Biology of Mormyrids (Osteichthyes: Mormyridae) in Anambra River, Nigeria. Ph.D Thesis University of Nigeria Nsukka, 194 pp.
Nwani, C. D. (2006). Length-weight relationship and condition factor of Distichodus species of Anambra River. Animal Research International, 3(2): 461 - 465.
Nwani, C. D., Oti, E. E., Odoh, G. E. and Ezenwaji, H. M. G. (2008). Endoparasitic helminthes of four Mormyrid species (Osteichthyes: Mormyridae) from a West African flood river system. Pakistan Journal of Biological Sciences, 11(2): 202 - 207.
Nwita, C. and Nkwengulila, G. (2004). Parasites of Clarias gariepinus (Burchell, 1822) (Pisces: Clariidae) from the Mwanza Gulf, Lake Victoria. Tanzanian Journal of Science, 30(1): 223 – 227.
Odo, G. E. (2004). Studies on the Ecology of Macro Invertebrate Fauna as Fish Food in Anambra River Basin, Nigeria. Ph.D. Thesis. University of Nigeria, Nsukka, Nigeria, 192 Pp.
Odo, G. E., Didigwu, N. C. and Eyo, J. E. (2009). The fish fauna of Anambra River Basin, Nigeria species abundance and morphometry. Revista de Biologia Tropical, 57(1 – 2): 177 – 186.
Odo, G. E., Onoja, S. O. and Onyishi, G. C. (2012). The biology of Parachanna obscura (Osteichthyes: Channidae) in Anambra river, Nigeria. International Journal of Fisheries and Aquaculture, 4(8): 154-169.
Ogbulie, T. E., Nwigwe, H. C. and Anyadoh, S. O. (2011). Comparative assessment of bioload of healthy and diseased Oreochromis niloticus as means of food security. Universitatii din Oredea – Fascicula Biologie, 18(1): 10 – 14.
Okafor, F. C. and Ubachukwu, P. O. (2009). Parasitism and other Animals. Pp. 1 – 10. In: Okafor, F. C. and Okoye, I. C. (Editors). Parasitology for the Health Science. Jolyn Publishers, Enugu, Nigeria.
Okaka, C. E. and Akhigbe, J. E. (1999). Helminth parasites of some tropical freshwater fish from Osse River in Benin, Southern Nigeria. Tropical Freshwater Biology, 8: 41 – 48.
Oktener, A., Egribas, E. and Basusta, N. (2008). A preliminary investigation on serious mortalities of fish in Balikligöl (Halil-ür Rahman Gölü, Şanliurfa). Gazi University Journal of Science, 21(1): 9 - 13.
Olaosebikan B. D. and Raji, A. (1998). Field Guide to Nigeria Freshwater Fishes. Federal College of Freshwater Fisheries Technology, New Bussa.

cii
Olofintoye, L. K. (2006). Parasitofauna in some freshwater fish species in Ekiti State, Nigeria. Pakistan Journal of Nutrition 5(4): 359 – 362.
Olson, O. W. (1974). Animal Parasites. University Park Press, USA.
Omeji, S., Solomon, S. G. and Idoga, E. S. (2011). A Comparative study of the common protozoan parasites of Clarias gariepinus from wild and cultured environments in Benue State, Nigeria. Journal of Parasitology Research, 3: 1 – 8.
Omeji, S., Solomon, S. G. and Obande, R. A. (2010). A Comparative Study of common protozoan parasites of Heterobranchus longifilis from the wild and cultured environment in Benue State. Pakistan Journal of Nutrition, 9(9): 865 – 872.
Omran, H. H. and El-Kholony, K. H. (2003). The influence of Nitroxynil on clinical, biochemical and haematological parameters in Egyptian buffaloes affected with liver flukes in Sharkia governorate. Egyptian Journal of Agricultural Research, 81(2): 739 – 751.
Oniye, S. J., Adebote, D. A. and Ayanda, O. I. (2004). Helminth parasites of Clarias gariepinus in Zaria, Nigeria. Journal of Aquatic Science, 19(2): 71 - 76.
Osman, H. A. M., Ismaiel, M. M., Abbas, T. W. and Ibrahim, T. B. (2009). An approach to the interaction between Trichodiniasis and pollution with Benzo-apyrene in catfish (Clarias gariepinus). World Journal of Fish and Marine Sciences, 1(4): 283 - 289.
Onyedineke, N. E., Obi, U., Ofoegbu, P. U. and Ukogo, I. (2010). Helminth parasites of some freshwater fish from River Niger at Illushi, Edo State, Nigeria. Journal of American Science, 6(3): 16 - 21.
Paperna, I. (1996). Parasites, Infection and Diseases of Fishes in Africa-an Update. Central Institute for Freshwater Aquaculture (CIFA) Technical Paper. No. 31 Rome, FAO.
Patton, C. J. and Crouch, S. R. (1977). Enzymatic calorimetric determination of urea. Annals of Chemistry, 49: 464 – 469.
Perez-Bote, J. L. (2005). First record of Lernaea cyprinaea (Copepoda: Cydopoida) on the Allis Shad. Folia Biological, 53(3 – 4): 197 – 198.
Piasecki, W., Goodwin, A. E., Eiras, J. C. and Nowak, B. F. (2004). Importance of Copepoda in freshwater aquaculture. Zoological Studies, 43(2): 193 – 205.
Pinky, K., Qureshi, T. A., Rekha, S., Susan, M. and Bilal, A. (2012). Histopathological and haematological investigations on Nandus nandus (Ham.) parasitized by metacercariae of Clinostomum complanatum (Rudolphi, 1819). International Journal of Environmental Sciences, 2(3): 1324 – 1330.

ciii
Pinky, K., Rekha, S., Qureshi, T. A., Susan, M., Saroch, J. D. and Shivani, S. (2012). Host parasite relationship: Sengasp (Dollfus 1934) in association with naturally infected freshwater host fish, Channa punctatus Bl. Journal of Chemical, Biological and Physical Science, 2(4): 2175 - 2181.
Pouder, D. B., Curtis, E. W. and Yanong, R. P. E. (2011). Common freshwater fish parasites pictorial guide: Sessile ciliates. http:edis.ifas.ufl.edu/FA-107. Accessed 12th April, 2014.
Pouin, R. (2004). Macro ecological patterns of species riches in parasite assemblages. Basic and Applied Ecology, 5: 423 - 434.
Poulin, R. (2000).Variation in the intraspecific relationship between fish length and intensity of the parasitic infection: biological and statistical causes. Journal of Fish Biology, 56: 123 - 137.
Poulin, R. (2006). Variation in infection parameters among populations within parasite species: intrinsic properties versus local factors. Journal of Parasitology, 36: 877 - 885.
Rahman, W. A. and Saidin, H. (2011). Relationship between sex and parasite intensity in four freshwater fish species from Tasik Merah, Peninsular Malaysia. World Journal of Zoology, 6(4): 370 - 374.
Reed, C. C., Basson, L. and Van As, L. L. (2003). Myxozoans infecting the sharptooth catfish, Clarias gariepinus in the Okavangou River and Delta, Botswana, including descriptions of two new species, Henneguya samochimensis species Myxobolus gariepinus species. Folia Parasitologica, 50: 183 – 189.
Reed, P., Francis-Floyd, R. and Klinger, R. E. (2009). Monogenean parasites of fish. http://edis.fas.edu.FA28. Accessed 13th April, 2014.
Reed, W., Buchard, J., Hopson, A. J., Jennes, J. and Yaro, I. (1967). Fish and Fisheries of Northern Nigeria. Published by Gaskiya. Corporation, Zaria Nigeria.
Reimchen, T. E. (2001). Ecological causes of sex-biased parasitism in threespine stickle back. Biological Journal of the Linnean Society, 73: 51 - 63.
Reimchen, T. E. (2001). Ecological causes of sex-biased parasitism in three spine stickle back. Biological Journal of the Linnaeus Society, 73: 51 - 63.
Reitman, S. and Frankel, S. (1971). Determination of serum glutamic oxalacetic and glutamic pyruvic transaminase. American Journal of Clinical Pathology, 28: 56 – 58.
Rhode, K. (1993). Ecology of Marine Parasites. Second Edition. University of Queens land Press.

civ
Roberts, R. J. (1978). Fish Pathology. A Bailliere Tindall Book, Publ. Cassell Ltd. Rohlenova, K., Morand, S., Hyršl, P., Tolarová, S., Flajšhans, M. and Sinkova, A. (2011). Are
fish immune systems really affected by parasites? And immunological study of common carp (Cyrinus carpio). Parasites and Vectors, 40: 120 – 138.
Rolbiecki, L. (2006). Correlation between the occurrence of parasites and body length of roach carp beam European Perch, zander, and ruffe in the Vistula Lagoon estuary. International Journal of Oceanography and Hydrobiology, 35(3): 257 – 267.
Salgado-Maldonado, G. (2008). Helminth parasites of freshwater fish from Central America. Zootaxa, 1915: 29 – 52.
Schludermann, C., Konecny, R., Laimgruber, S., Lewis, J. W., Schiemer, F., Chovanec, A. and Sures, B. (2003). Fish macroparasites as indicators of heavy metal pollution in river sites in Austria. Parasitology, 126: 62 – 69.
Sexena, K. K. and Chauchan, R. R. S. (1993). Effect of parasitic infection on the blood of Heteropneustus fossilis (Bloch). Bioved, 4: 41 – 43.
Shahi, N., Yousuf, A. R., Rather, M. I., Ahmad, F. and Yaseen, T. (2013). First report of blood parasites in fishes from Kashmir and their effect on the haematological profile. Open Veterinary Journal, 3(2): 89 – 95.
Sheldon, B. C. and Verhulst, S. (1996). Ecological immunology: costly parasite defenses and trade-off in evolutionary ecology. Trends in Ecology and Evolution, 11: 317 – 321.
Simkova, A., Lafond, T., Ondračkova, M., Jurajda, P., Ottova, E. and Morand, S. (2008). Parasitism, life history traits and immune defense in cyprinid fish from Central Europe. BMC Evolutionary Biology, 8: 29 – 40.
Sindermann, C. J. (1987). Effect of parasites on fish population: practical considerations. International Journal of Parasitology, 17: 371 – 382.
Sinha, K. P. (2000). Haematological manifestation in Clarias batrachus carrying helminth infection. Journal of Parasitic Diseases, 24: 167 – 170.
Skarstein, F., Folstad, I. and Liljeda, S. (2001). Whether to reproduce or not: Immune suppression and costs of parasites during reproduction in the Arctic Charr. Canadian Journal of Zoology, 79: 271 – 278.
Smyth, J. D. (1994). Animal Parasitology. Cambridge University Press, Britian.

cv
Soliman, M. F. M. and Ibrahim, M. M. (2012). Monogenean community structure of Oreochoromis niloticus in relation to heavy metal pollution and host reproductive cycle. Journal of the Eygptian Society of Parasitology, 42(1): 11 – 24.
Sommerville, C. (1982). The pathology of Haplorchis pumilio (Looss, 1895) infection in cultured Tilapias. Journal of Fish Disease, 5: 243 – 250.
Stevenson, R. D. and Woods, W. A. (2006). Condition indices for conservation: new uses for evolving tools. Integrative and Comparative Biology, 46(6): 1169 – 1190.
Tang, F., Zhao, Y. and Liu, C. (2012). Two trichodinids of Paratrichodina (Ciliophora, Paritrichida, Trichodinidae) infecting gills of Letalurus punetaus from Chongqing, China. African Journal of Microbiology Research, 6(9): 2145 – 2149.
Thompson, L. C. and Larsen, R. (2014). Fish Habitat in Freshwater Stream. Farm Water Quality Planning FWQP; Reference Sheet 10.3, Publication 8112. University of California, California.
Tombi, J. and Bilong, C. F. B. (2004). Distribution of gill parasites of the freshwater fish Barbus martorelli Roman, 1971 (Teleostei: Cyprinidae) and tendency to inverse intensity evolution between myxosporidia and Monogenea as a function of the host age. Revue de’Élevage et Médecine Vetrinaire des Pays Tropicaux, 57(1 – 2): 71 - 76.
Torres, P. B., Contreras, L., Figuerga, R., Franjola, H., Gonzaleh, R. and Martin, R. (1977). Research on Pseudophyllidea from the South of Chile, I. Preliminary investigation on infection by Plerocercoids of Diphyllobotrium sp. in Salmo gairdnerii from Calatquen lake Chile. Boletin Chileno de Parasitologi. 32: 73 - 80.
Udo, R. K. (1975). Geographical Regions of Nigeria. Heinemann Educational Books, Ltd, London.
Ukoli, F. M. A. (1969). Preliminary report on the helminthic infections in the River Niger at Shagunu (near Kainji). In: Obene, E. E. (Editor). Proceedings Accra Symposium Man Made Lakes. Accra, Ghana, University Press for Ghana Academy of Sciences.
Usip, L. P. E., Udoidiong, O. M., Ekwu, A., Ekpo, I. and Essienibok, M. (2010). Parasite spectrum of cichlid species in the lower Cross River at Itu Akwa Ibom State, Nigeria. World Journal of Applied Science and Technology, 2(1): 59 – 68.
van Dyk, J. C., Pieterse, G. M. and van Vuren, J. H. J. (2007). Histological changes in the liver of Oriochromis mossambicus (Cichlidae) after exposure to cadmium and zinc. Ecotoxicology and Environmental Safety. 66: 432 – 440.

cvi
Vankara, A. P., Mani, G. and Vijayalakshmi, C. (2011). Metazoan parasite infracommunities of the freshwater eel, Mastacembelus armatus Lacepede, 1800 from River Godavari, India: International Journal of Zoological Research, 7(1): 19 – 33.
Volonterio, O., Lopez de Leon, E. and Ponce de Leon, R. (2004). Infestation dynamics and histopathology of two species of fresh water leaches (Hirudinea: Piscicolidae) on teleoist fish from Uruguay. Journal of Comparative Parasitology, 71(1): 21 - 28.
Wedkind, C. and Jacobson, P. J. (1998). Male-biased susceptibility to helminth infections: an experimental test with a copepod. Oikos Journal, 81: 458 - 462.
Wells, R. M. G. and Weber, R. E. (1990). Spleen in hypoxic and exercised rainbow trout. Journal of Experimental Biology, 150: 461 – 466.
West, H. J. (1989). Evaluation of total plasma bile-acid concentrations for the diagnosis of hepatobiliary disease in horses. Research of Veterinary Science, 46(2): 264 – 270.
Williams, H. and Jones A. (1994). Parasitic Worms of Fish. Taylor and Francis, Bristol, UK.
Williams, H. H. and Makenzie, K. (2003). Marine parasites as pollution indicators: an update. Parasitology, 126: 27 – 41.
Withwaskul, P., Panthong, A., Kanjanapothi, D., Taesothikul, T. and Lertprasertsuke, A. (2003). Acute and sub-acute toxicity of saponin mixture isolated from Schefflera leucantha viguuire. Journal of Ethnopharmacology, 89: 115 – 121.
Yakubu, D. P., Omoregie, E., Wade, J. W. and Faringaro, D. U. (2002). A comparative study of gut helminthes of Tilapia zilli and Clarias gariepinus from River Uke, Plateau State, Nigeria. Journal of Aquatic Science, 17(2): 137 – 139.
Yamaguti, S. (1961). Systema Helmithium: The Nematodes of Vertebrates. Interscience Publisher, New York.
Yamaguti, S. (1963). Systema Helminthum: The Acanthocephala. Interscience Publisher, New York.
Younis, A. A. E. (1999). Effect of some ectoparasites on the blood and serum constituents of Oreochromis niloticus fish with referring to treatment. Beni Suif. Veterinary Medical Journal, 9(3): 341 - 351.
Yuniar, A. T., Palm, H. W. and Walter, T. (2007). Crustacean fish parasites from Segara Anakam Lagoon, Java, Indonesia. Parasitology Research, 100: 1193 – 1204.
Zaman, Z. (1985). Parasite Fauna of Paddy Field Catfish (Genus: Clarias) from Kedah and Perak, Peninsula Malaysia. Ph.D Thesis, University of Sains, Minden Malaysia.

cvii
Zaman, Z. and Seng, L. T. (1989). The seasonal abundance of the parasite in Clarias batrachus and C. macrocephalus from two areas (Kedah and Perak) of Malaysia and its relationship to the maturity of the hosts. Bangladesh Journal of Zoology, 17: 47 - 55.




















