
Efficacy of herbicide seed treatments for controlling Striga infestation of Sorghum
7
CROP SCIENCE, VOL. 49, MAY– JUNE 2009 923 RESEARCH S orghum [ Sorghum bicolor (L.) Moench] is a staple food crop for millions of people in Africa, South Asia, and Central America. Worldwide, it is the fifth major cereal crop in terms of production, after maize ( Zea mays L. ssp. mays), wheat (Triticum aestivum L.), rice ( Oryza sativa L.), and barley (Hordeum vulgare L.) (FAOSTAT, http://faostat.org). Sorghum is particularly impor- tant in Africa where production has increased steadily over the past 40 yr from nearly 10 million metric tons to 26 million metric tons on approximately 25 million ha (http.faostat.org). Sorghum production is negatively influenced by abiotic (heat, drought, and low fertility) and biotic stresses including diseases, insects, and weeds. Witchweeds ( Striga spp.) pose a serious and grow- ing threat to sustainable and economic food production in Africa (Ejeta and Butler, 1993). Annual crop losses due to Striga in the savanna regions of Africa are worth at least US$7,000,000,000 and affect more than 100 million people (Ejeta, 2007; de Groote, 2007). Strategies to minimize effects of Striga on crop production have focused on developing resistant cultivars, reducing the Striga seed bank in the soil, limiting Striga seed production, and Efficacy of Herbicide Seed Treatments for Controlling Striga Infestation of Sorghum Mitchell R. Tuinstra,* Souley Soumana, Kassim Al-Khatib, Issoufou Kapran, Aboubacar Toure, Aad van Ast, Lammert Bastiaans, Newton W. Ochanda, Issoufou Salami, Mountaga Kayentao, and Siaka Dembele ABSTRACT Witchweed (Striga spp.) infestations are the greatest obstacle to sorghum [Sorghum bicolor (L.) Moench] grain production in many areas in Africa. The objective of this study was to evalu- ate the efficacy of herbicide seed treatments for controlling Striga infestation of sorghum. Seeds of an acetolactate synthase (ALS) herbicide– tolerant sorghum hybrid were treated with two ALS-inhibiting herbicides. Treatments included three rates of imazapyr (IMI), three rates of met- sulfuron-methyl (MET), and an untreated control group. In greenhouse trials, observations at 32, 46, and 60 d after planting showed that seeds treated with the highest herbicide rates had the fewest Striga attachments and the greatest delay in attachment. All plants in the untreated group died at or before sorghum flowering; how- ever, herbicide seed treatments, particularly metsulfuron, reduced Striga emergence and significantly increased sorghum grain yield and dry matter production. Field studies comparing seed treatments produced similar results with delayed Striga emergence and fewer emerged Striga plants in herbicide-treated plots. These studies indicate that herbicide seed treatments may provide a highly effective tool for managing Striga in sorghum. M.R. Tuinstra, Dep. of Agronomy, Purdue Univ., West Lafayette, IN, 49706; S. Soumana, K. Al-Khatib, and N.W. Ochanda, Dep. of Agron- omy, Kansas State Univ., Manhattan, KS, 66506; I. Kapran and I. Salami, INRAN, Niamey, Niger; A. Toure, M. Kayentao, and S. Dembele, IER, Bamako, Mali; A. van Ast and L. Bastiaans, Crop and Weed Ecology Group, Wageningen Univ., Wageningen, the Netherlands. Contribution no. 08-329-J from the Kansas Agricultural Experiment Station. Received 23 June 2008. *Corresponding author ([email protected]). Abbreviations: ALS, acetolactate synthase; DAP, days after planting; IMI, imazapyr; IR, imidazolinone resistant; MET, metsulfuron-methyl. Published in Crop Sci. 49:923–929 (2009). doi: 10.2135/cropsci2008.06.0357 © Crop Science Society of America 677 S. Segoe Rd., Madison, WI 53711 USA All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Efficacy of herbicide seed treatments for controlling Striga infestation of Sorghum

CROP SCIENCE, VOL. 49, MAY–JUNE 2009 923
RESEARCH
Sorghum [Sorghum bicolor (L.) Moench] is a staple food crop for millions of people in Africa, South Asia, and Central
America. Worldwide, it is the fi fth major cereal crop in terms of production, after maize (Zea mays L. ssp. mays), wheat (Triticum aestivum L.), rice (Oryza sativa L.), and barley (Hordeum vulgare L.) (FAOSTAT, http://faostat.org). Sorghum is particularly impor-tant in Africa where production has increased steadily over the past 40 yr from nearly 10 million metric tons to 26 million metric tons on approximately 25 million ha (http.faostat.org).
Sorghum production is negatively infl uenced by abiotic (heat, drought, and low fertility) and biotic stresses including diseases, insects, and weeds. Witchweeds (Striga spp.) pose a serious and grow-ing threat to sustainable and economic food production in Africa (Ejeta and Butler, 1993). Annual crop losses due to Striga in the savanna regions of Africa are worth at least US$7,000,000,000 and aff ect more than 100 million people (Ejeta, 2007; de Groote, 2007).
Strategies to minimize eff ects of Striga on crop production have focused on developing resistant cultivars, reducing the Striga seed bank in the soil, limiting Striga seed production, and
Effi cacy of Herbicide Seed Treatments for Controlling Striga Infestation of Sorghum
Mitchell R. Tuinstra,* Souley Soumana, Kassim Al-Khatib, Issoufou Kapran, Aboubacar Toure, Aad van Ast, Lammert Bastiaans, Newton W. Ochanda, Issoufou Salami, Mountaga Kayentao, and Siaka Dembele
ABSTRACT
Witchweed (Striga spp.) infestations are the
greatest obstacle to sorghum [Sorghum bicolor
(L.) Moench] grain production in many areas in
Africa. The objective of this study was to evalu-
ate the effi cacy of herbicide seed treatments for
controlling Striga infestation of sorghum. Seeds
of an acetolactate synthase (ALS) herbicide–
tolerant sorghum hybrid were treated with two
ALS-inhibiting herbicides. Treatments included
three rates of imazapyr (IMI), three rates of met-
sulfuron-methyl (MET), and an untreated control
group. In greenhouse trials, observations at 32,
46, and 60 d after planting showed that seeds
treated with the highest herbicide rates had
the fewest Striga attachments and the greatest
delay in attachment. All plants in the untreated
group died at or before sorghum fl owering; how-
ever, herbicide seed treatments, particularly
metsulfuron, reduced Striga emergence and
signifi cantly increased sorghum grain yield and
dry matter production. Field studies comparing
seed treatments produced similar results with
delayed Striga emergence and fewer emerged
Striga plants in herbicide-treated plots. These
studies indicate that herbicide seed treatments
may provide a highly effective tool for managing
Striga in sorghum.
M.R. Tuinstra, Dep. of Agronomy, Purdue Univ., West Lafayette, IN,
49706; S. Soumana, K. Al-Khatib, and N.W. Ochanda, Dep. of Agron-
omy, Kansas State Univ., Manhattan, KS, 66506; I. Kapran and I. Salami,
INRAN, Niamey, Niger; A. Toure, M. Kayentao, and S. Dembele, IER,
Bamako, Mali; A. van Ast and L. Bastiaans, Crop and Weed Ecology
Group, Wageningen Univ., Wageningen, the Netherlands. Contribution
no. 08-329-J from the Kansas Agricultural Experiment Station. Received
23 June 2008. *Corresponding author ([email protected]).
Abbreviations: ALS, acetolactate synthase; DAP, days after planting;
IMI, imazapyr; IR, imidazolinone resistant; MET, metsulfuron-methyl.
Published in Crop Sci. 49:923–929 (2009).doi: 10.2135/cropsci2008.06.0357© Crop Science Society of America677 S. Segoe Rd., Madison, WI 53711 USA
All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher.

924 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, MAY–JUNE 2009
diminishing the spread of Striga to uninfested fi elds (Ejeta and Butler, 1993; Rodenburg et al., 2006; van Ast and Bastiaans, 2006). Although host-plant resistance and cul-tural management strategies such as hand pulling, crop rotation, trap crops, and improved soil fertility contrib-ute to management of this parasite, new technologies and strategies are needed to further enhance Striga control (Ejeta, 2005; van Ast et al., 2005).
Herbicide seed treatments are a relatively new approach to herbicidal control of parasitic weeds (Abayo et al., 1996; Kanampiu et al., 2001, 2003; Diaz-Sanchez et al., 2003; Dembele et al., 2005; de Groote et al., 2007). This approach uses low rates of herbicides and does not rely on specialized application equipment. Seed treatment consists of either coating the seed surface with a herbicide or imbibing or priming seeds in a herbicide solution. Herbicides absorbed into the seed or germinating seedling are subsequently transferred to an attached parasite ( Jurado-Exposito et al., 1999; Diaz-Sanchez et al., 2002) or kill nearby Striga plants through diff usion into the surrounding soil (Kanampiu et al., 2002). Use of herbicide seed treatments for parasitic weed control was fi rst demonstrated by Berner et al. (1994) using imazaquin on cowpea [Vigna unguiculata (L.) Walp.] to control Striga gesnerioides (Willd.) Vatke. Subsequently, Jurado-Exposito et al. (1996, 1997) showed that seed treat-ments with other imidazolinone herbicides improved con-trol of crenate broomrape (Orobanche crenata Forssk.) on broad bean (Vicia faba L.) and pea (Pisum sativum L.).
Success of herbicide seed treatments requires a suffi -cient margin of selectivity between the crop and weed such that the crop is not injured by the herbicide. Development of herbicide-resistant crops that tolerate high levels of her-bicide, such as acetolactate synthase (ALS)–inhibiting her-bicide tolerance in maize (Berner et al., 1997; Kanampiu et al., 2001), was a major breakthrough for using herbi-cide seed treatment technology to manage Striga infesta-tion. Acetolactate synthase herbicide–tolerant maize was developed in the United States by pollen mutagenesis and is marketed as IR (imidazolinone resistant) maize. The IR maize plants have a mutation of Ala133Thr (Greaves et al., 1993; Wright and Penner, 1998). Recent studies in Africa with ALS inhibitors dressed as a drench (Abayo et al., 1996, 1998) or seed coating (Berner et al., 1997) to IR maize seed demonstrated that low doses of herbicides can provide eff ective control of Striga infestation.
Sorghum is susceptible to many ALS-inhibiting her-bicides; however, Tuinstra and Al-Khatib (2007) identi-fi ed a mutant with tolerance to ALS herbicides in 2004. This trait is being incorporated into improved cultivars adapted for production and use in Africa. The objective of this study was to determine whether imazapyr or met-sulfuron-methyl seed treatments could provide eff ective control of Striga hermonthica infestation of ALS herbicide–tolerant sorghum in greenhouse and fi eld conditions.
MATERIALS AND METHODS
Plant Material and Herbicide TreatmentsThe sorghum genotype evaluated in these trials was a F
1 hybrid
produced from a cross between ‘ATx623’ and ‘Tailwind’. ATx623
is an ALS herbicide–susceptible genotype widely used in the trop-
ics for hybrid seed production including production of the com-
mercially important F1 hybrids ‘Hageen-Durra-1’ in Sudan and
‘NAD-1’ in Niger. Tailwind is an ALS herbicide–tolerant sorghum
genotype derived from a wild sorghum accession collected from a
population of wild sorghum plants in a fi eld near Manhattan, KS.
Seeds of this accession were sown in a greenhouse in Manhattan,
KS. Seedlings were sprayed at the V3 growth stage with 140 g ha–1
a.i. nicosulfuron (2-[[[[(4,6-dimethoxy-2-pyrimidinyl)amino]car-
bonyl]amino] sulfonyl]-N,N-dimethyl-3-pyridinecarboxamide)
in the form of Accent Herbicide (E.I. du Pont de Nemours and
Co., Wilmington, DE). Nicosulfuron is a sulfonylurea herbicide
that exhibits high selectivity on many grass and broadleaf weeds.
All of the seedlings of this accession were uniform and equally
tolerant to the foliar herbicide application. A representative plant
showing high levels of tolerance was self-pollinated to produce
seeds for subsequent studies. Tailwind was crossed as a pollinator
to ATx623 to produce an experimental F1 hybrid. This hybrid sur-
vived foliar herbicide applications of nicosulfuron, indicating that
herbicide tolerance was inherited as a dominate trait.
Seeds of the experimental F1 hybrid were treated with varying
rates of two ALS-inhibiting herbicides. Herbicide seed treatments
included imazapyr (IMI) and metsulfuron-methyl (MET). The
IMI treatments were applied as Arsenal Herbicide (BASF Corpo-
ration, Florham Park, NJ), and MET treatments were applied as
Ally XP Herbicide (DuPont, Wilmington, DE). In preliminary
experiments, sorghum seeds were treated with 0, 19, 38, 75, 150,
and 300 μg a.i. IMI seed–1 and 0, 6, 12, 25, 50, and 100 μg a.i.
MET seed–1 to establish tolerance of the experimental hybrid used
in this study to these treatments. Based on results of these experi-
ments, 19, 38, and 75 μg a.i. IMI seed–1 (IMI-low, IMI-medium,
and IMI-high, respectively) and 3, 6, and 12 μg a.i. MET seed–1
(MET-low, MET-medium, and MET-high, respectively) were
evaluated for comparison to an untreated control group.
Greenhouse TrialsThree IMI and three MET herbicide seed treatments and an
untreated control group were evaluated for diff erences in sor-
ghum and Striga growth and development under Striga-infested
and noninfested conditions in pots in a greenhouse at Wagen-
ingen University, Wageningen, the Netherlands, at 32, 46, and
60 d after planting (DAP) and at physiological maturity. The
experiment was conducted using a randomized complete block
design with eight replications per treatment and sampling date.
The greenhouse study was initiated on 30 May 2005. Plants
were grown in 12-L pots that contained a mixture (3:1) of sand
and arable sandy soil collected from the top layer (0–0.25 m) of
a fi eld near Wageningen. Striga-infested pots were prepared by
mixing 30 mg Striga hermonthica seed (4000 viable seeds) collected
in Cinzana, Mali, into the top 10 cm of each pot at a density
of approximately 1 viable Striga seed cm–3. Pots were watered
and then covered with plastic for 10 d to keep the soil moist
and allow conditioning of the Striga seeds. Noninfested pots
were handled similarly except without sowing of Striga seeds.
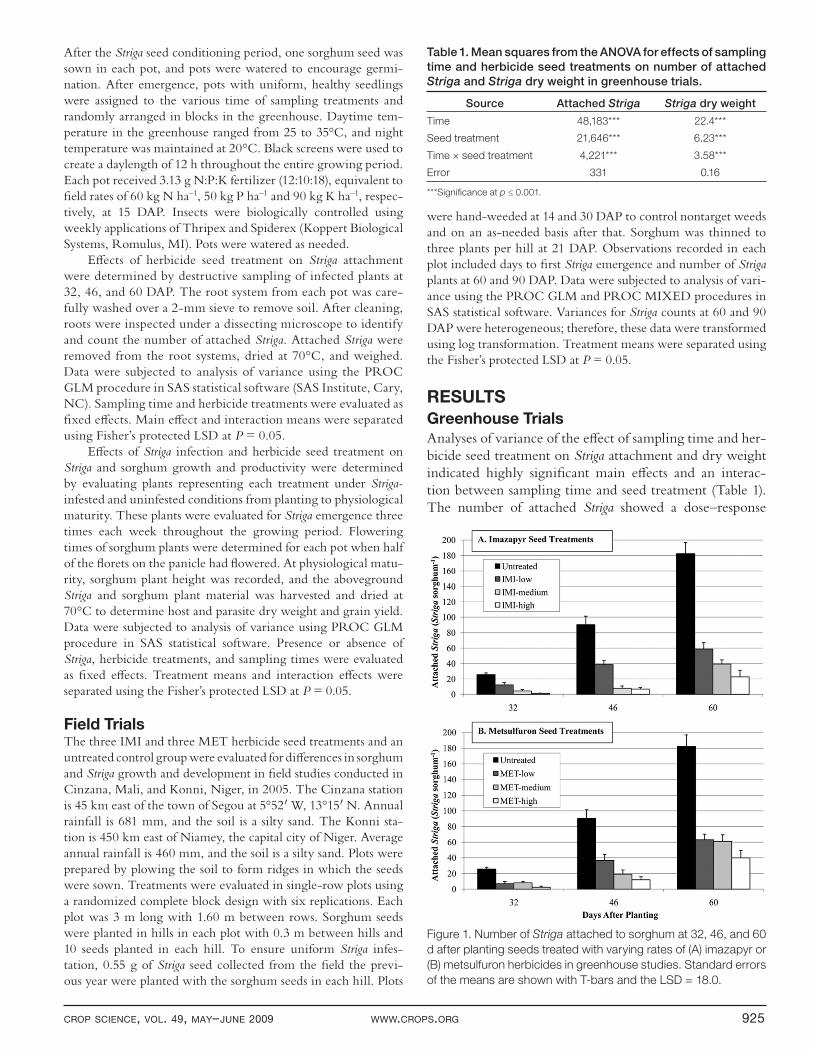
CROP SCIENCE, VOL. 49, MAY–JUNE 2009 WWW.CROPS.ORG 925
were hand-weeded at 14 and 30 DAP to control nontarget weeds
and on an as-needed basis after that. Sorghum was thinned to
three plants per hill at 21 DAP. Observations recorded in each
plot included days to fi rst Striga emergence and number of Striga
plants at 60 and 90 DAP. Data were subjected to analysis of vari-
ance using the PROC GLM and PROC MIXED procedures in
SAS statistical software. Variances for Striga counts at 60 and 90
DAP were heterogeneous; therefore, these data were transformed
using log transformation. Treatment means were separated using
the Fisher’s protected LSD at P = 0.05.
RESULTS
Greenhouse TrialsAnalyses of variance of the eff ect of sampling time and her-bicide seed treatment on Striga attachment and dry weight indicated highly signifi cant main eff ects and an interac-tion between sampling time and seed treatment (Table 1). The number of attached Striga showed a dose–response
After the Striga seed conditioning period, one sorghum seed was
sown in each pot, and pots were watered to encourage germi-
nation. After emergence, pots with uniform, healthy seedlings
were assigned to the various time of sampling treatments and
randomly arranged in blocks in the greenhouse. Daytime tem-
perature in the greenhouse ranged from 25 to 35°C, and night
temperature was maintained at 20°C. Black screens were used to
create a daylength of 12 h throughout the entire growing period.
Each pot received 3.13 g N:P:K fertilizer (12:10:18), equivalent to
fi eld rates of 60 kg N ha–1, 50 kg P ha–1 and 90 kg K ha–1, respec-
tively, at 15 DAP. Insects were biologically controlled using
weekly applications of Thripex and Spiderex (Koppert Biological
Systems, Romulus, MI). Pots were watered as needed.
Eff ects of herbicide seed treatment on Striga attachment
were determined by destructive sampling of infected plants at
32, 46, and 60 DAP. The root system from each pot was care-
fully washed over a 2-mm sieve to remove soil. After cleaning,
roots were inspected under a dissecting microscope to identify
and count the number of attached Striga. Attached Striga were
removed from the root systems, dried at 70°C, and weighed.
Data were subjected to analysis of variance using the PROC
GLM procedure in SAS statistical software (SAS Institute, Cary,
NC). Sampling time and herbicide treatments were evaluated as
fi xed eff ects. Main eff ect and interaction means were separated
using Fisher’s protected LSD at P = 0.05.
Eff ects of Striga infection and herbicide seed treatment on
Striga and sorghum growth and productivity were determined
by evaluating plants representing each treatment under Striga-
infested and uninfested conditions from planting to physiological
maturity. These plants were evaluated for Striga emergence three
times each week throughout the growing period. Flowering
times of sorghum plants were determined for each pot when half
of the fl orets on the panicle had fl owered. At physiological matu-
rity, sorghum plant height was recorded, and the aboveground
Striga and sorghum plant material was harvested and dried at
70°C to determine host and parasite dry weight and grain yield.
Data were subjected to analysis of variance using PROC GLM
procedure in SAS statistical software. Presence or absence of
Striga, herbicide treatments, and sampling times were evaluated
as fi xed eff ects. Treatment means and interaction eff ects were
separated using the Fisher’s protected LSD at P = 0.05.
Field TrialsThe three IMI and three MET herbicide seed treatments and an
untreated control group were evaluated for diff erences in sorghum
and Striga growth and development in fi eld studies conducted in
Cinzana, Mali, and Konni, Niger, in 2005. The Cinzana station
is 45 km east of the town of Segou at 5°52′ W, 13°15′ N. Annual
rainfall is 681 mm, and the soil is a silty sand. The Konni sta-
tion is 450 km east of Niamey, the capital city of Niger. Average
annual rainfall is 460 mm, and the soil is a silty sand. Plots were
prepared by plowing the soil to form ridges in which the seeds
were sown. Treatments were evaluated in single-row plots using
a randomized complete block design with six replications. Each
plot was 3 m long with 1.60 m between rows. Sorghum seeds
were planted in hills in each plot with 0.3 m between hills and
10 seeds planted in each hill. To ensure uniform Striga infes-
tation, 0.55 g of Striga seed collected from the fi eld the previ-
ous year were planted with the sorghum seeds in each hill. Plots
Table 1. Mean squares from the ANOVA for effects of sampling
time and herbicide seed treatments on number of attached
Striga and Striga dry weight in greenhouse trials.
Source Attached Striga Striga dry weight
Time 48,183*** 22.4***
Seed treatment 21,646*** 6.23***
Time × seed treatment 4,221*** 3.58***
Error 331 0.16
***Signifi cance at p ≤ 0.001.
Figure 1. Number of Striga attached to sorghum at 32, 46, and 60
d after planting seeds treated with varying rates of (A) imazapyr or
(B) metsulfuron herbicides in greenhouse studies. Standard errors
of the means are shown with T-bars and the LSD = 18.0.

926 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, MAY–JUNE 2009
relationship with the IMI and MET treatments with the fewest attachments and greatest delay in attachment at the highest rates of herbicide treatment (Fig. 1). The number of attached Striga increased over time. In the untreated control group, approximately 20 attached Striga per sorghum were observed at 32 DAP, and 180 Striga per sorghum plant were observed at 60 DAP (Fig. 1). Striga attachment was much lower in the IMI and MET herbicide seed treatments. At the highest treatment rates, very few attached Striga were found at 32 DAP, and a four- to eightfold reduction in Striga attachment compared with the control was observed for the MET and IMI treatments, respectively, at 60 DAP (Fig. 1). Low and intermediate rates of IMI and MET herbicide treatments also were eff ective at reducing Striga attachment, but the highest rates provided the greatest protection. Simi-
lar trends were found when comparing eff ects of herbicide seed treatments on Striga dry weight (data not shown).
Analyses of variance of the eff ects of sampling time and herbicide treatment on Striga emergence indicated highly signifi cant main eff ects (p < 0.001) and interaction (p < 0.001) between sampling time and herbicide treatment. In pots representing the untreated control group, Striga emergence increased dramatically over time beginning at approximately 25 DAP and reaching a maximum of more than 40 Striga per pot by 71 DAP (Fig. 2). Striga emergence was delayed and reduced in pots with herbicide-treated seed (Fig. 2). The IMI- and MET-treated plants had signifi -cantly fewer emerged Striga than control plants throughout most of the growth cycle. At the highest rates of IMI and MET herbicide treatment, fewer than fi ve Striga plants per pot were observed at 60 DAP; more than 30 Striga plants per pot were observed in the untreated control group. Effi -cacy of herbicide treatments declined later in development, particularly for the IMI treatments.
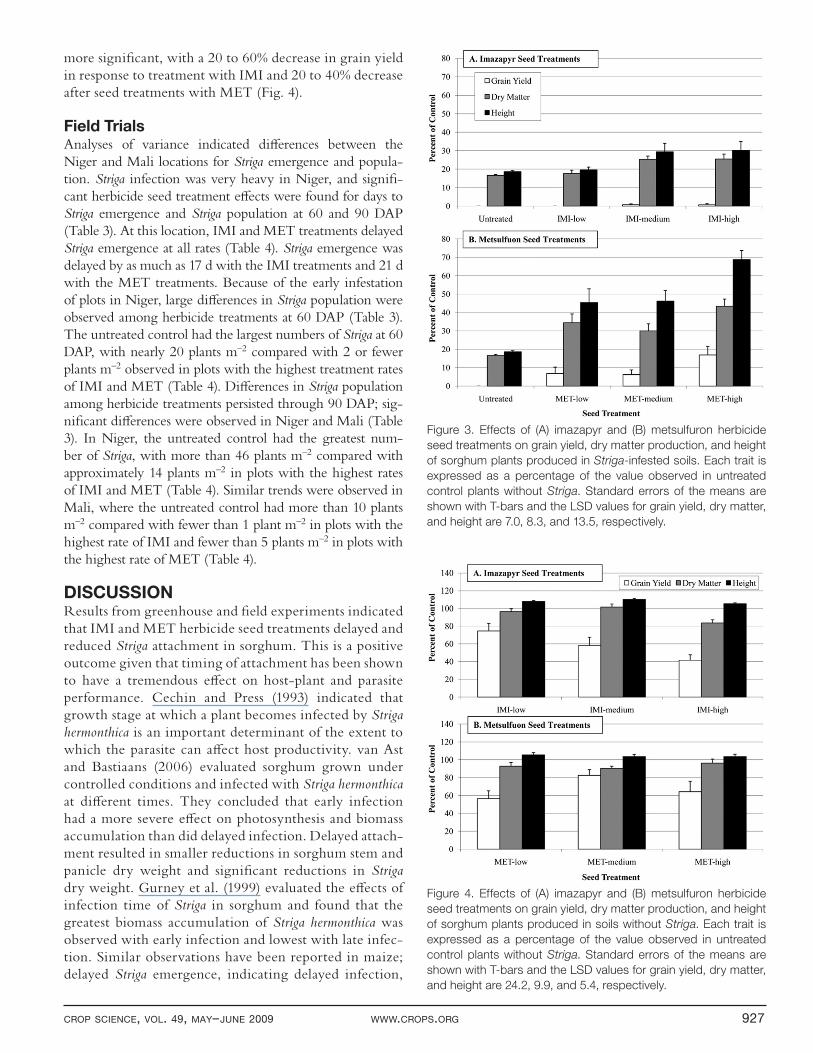
Analyses of variance of the eff ects of Striga infestation and herbicide seed treatment on sorghum grain yield, dry matter accumulation, and plant height indicated highly sig-nifi cant (p < 0.001) main eff ects and an interaction between Striga infestation and herbicide seed treatment (Table 2). Under Striga-infested conditions, plants in the untreated control group died prematurely as a result of the massive parasite infection. These plants produced no grain and only 20% of the dry matter and plant height of untreated plants not infected with Striga (Fig. 3). Plants produced from IMI-treated seed generally survived to maturity and performed better than the untreated control but were not as productive as the MET-treated group (Fig. 3). Under Striga-infested conditions, IMI-treated plants representing the highest treatment rate produced a small amount of grain and 25 to 30% of the dry matter and plant height of untreated plants not infected with Striga (Fig. 3). Striga-induced stunting was noticeable in the MET-treated group; however, plants representing the highest treatment rate survived to physi-ological maturity and produced 15% of the grain yield, 40% of the dry matter, and nearly 70% of the plant height of untreated plants not infested with Striga (Fig. 3).
Some visual characteristics of herbicide damage were noted in young plants representing the various herbicide seed treatments. These characteristics included chlorosis and some stunting for the fi rst 1 to 2 wk after emergence. Analyses of variance of the eff ects of seed treatments on grain yield, dry matter production, and plant height of sorghum not infested with Striga indicated that these eff ects were signifi cant (p < 0.05). Comparisons of treat-ment eff ects showed that changes in plant height and dry matter in response to the IMI and MET seed treatments were small, with treated plants producing slightly lower dry matter yields and similar or greater plant height (Fig. 4). The eff ect of herbicide treatments on grain yield was
Figure 2. Striga emergence in sorghum grown from seeds treated
with (A) imazapyr and (B) metsulfuron herbicides in Striga-infested
soils under greenhouse conditions. Standard errors of the means
are shown with T-bars and the LSD = 7.5.
Table 2. Mean squares from the ANOVA for effects of
Striga infestation and herbicide seed treatments on sor-
ghum grain yield, dry matter production, and plant height
in greenhouse trials.
Source Grain yield Dry matter Plant height
Striga infestation 10,369*** 16,4667*** 1,177,418***
Seed treatment 138*** 574*** 11,334***
Striga infestation ×
seed treatment159*** 789*** 12,887***
Error 28.7 110 956
***Signifi cance at p ≤ 0.001.
CROP SCIENCE, VOL. 49, MAY–JUNE 2009 WWW.CROPS.ORG 927
more signifi cant, with a 20 to 60% decrease in grain yield in response to treatment with IMI and 20 to 40% decrease after seed treatments with MET (Fig. 4).
Field TrialsAnalyses of variance indicated diff erences between the Niger and Mali locations for Striga emergence and popula-tion. Striga infection was very heavy in Niger, and signifi -cant herbicide seed treatment eff ects were found for days to Striga emergence and Striga population at 60 and 90 DAP (Table 3). At this location, IMI and MET treatments delayed Striga emergence at all rates (Table 4). Striga emergence was delayed by as much as 17 d with the IMI treatments and 21 d with the MET treatments. Because of the early infestation of plots in Niger, large diff erences in Striga population were observed among herbicide treatments at 60 DAP (Table 3). The untreated control had the largest numbers of Striga at 60 DAP, with nearly 20 plants m–2 compared with 2 or fewer plants m–2 observed in plots with the highest treatment rates of IMI and MET (Table 4). Diff erences in Striga population among herbicide treatments persisted through 90 DAP; sig-nifi cant diff erences were observed in Niger and Mali (Table 3). In Niger, the untreated control had the greatest num-ber of Striga, with more than 46 plants m–2 compared with approximately 14 plants m–2 in plots with the highest rates of IMI and MET (Table 4). Similar trends were observed in Mali, where the untreated control had more than 10 plants m–2 compared with fewer than 1 plant m–2 in plots with the highest rate of IMI and fewer than 5 plants m–2 in plots with the highest rate of MET (Table 4).
DISCUSSIONResults from greenhouse and fi eld experiments indicated that IMI and MET herbicide seed treatments delayed and reduced Striga attachment in sorghum. This is a positive outcome given that timing of attachment has been shown to have a tremendous eff ect on host-plant and parasite performance. Cechin and Press (1993) indicated that growth stage at which a plant becomes infected by Striga hermonthica is an important determinant of the extent to which the parasite can aff ect host productivity. van Ast and Bastiaans (2006) evaluated sorghum grown under controlled conditions and infected with Striga hermonthica at diff erent times. They concluded that early infection had a more severe eff ect on photosynthesis and biomass accumulation than did delayed infection. Delayed attach-ment resulted in smaller reductions in sorghum stem and panicle dry weight and signifi cant reductions in Striga dry weight. Gurney et al. (1999) evaluated the eff ects of infection time of Striga in sorghum and found that the greatest biomass accumulation of Striga hermonthica was observed with early infection and lowest with late infec-tion. Similar observations have been reported in maize; delayed Striga emergence, indicating delayed infection,
Figure 3. Effects of (A) imazapyr and (B) metsulfuron herbicide
seed treatments on grain yield, dry matter production, and height
of sorghum plants produced in Striga-infested soils. Each trait is
expressed as a percentage of the value observed in untreated
control plants without Striga. Standard errors of the means are
shown with T-bars and the LSD values for grain yield, dry matter,
and height are 7.0, 8.3, and 13.5, respectively.
Figure 4. Effects of (A) imazapyr and (B) metsulfuron herbicide
seed treatments on grain yield, dry matter production, and height
of sorghum plants produced in soils without Striga. Each trait is
expressed as a percentage of the value observed in untreated
control plants without Striga. Standard errors of the means are
shown with T-bars and the LSD values for grain yield, dry matter,
and height are 24.2, 9.9, and 5.4, respectively.

928 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, MAY–JUNE 2009
substantially increased host yield (Abayo et al., 1998; Kanampiu et al., 2003).
The MET and IMI seed treatments used in these experiments signifi cantly reduced Striga emergence in fi eld and greenhouse studies. Kanampiu et al. (2001) demonstrated similar results in maize and showed a lin-ear decrease in Striga emergence with increasing rates of imazapyr. In a subsequent study, Kanampiu et al. (2003) showed that Striga emergence was suppressed by imazapyr and pyrithiobac herbicide seed treatments at 8 and 12 wk after planting. Both herbicides delayed Striga asiatica emer-gence, and higher rates provided seasonlong control, pro-moting greater grain yield than the untreated control. de Groote et al. (2007) found similar results in on-farm tri-als; seed treatments contributed to reduced Striga infection and increased yield. Controlling Striga populations during crop development should reduce Striga seed production and improve management of the seed bank.
This was a proof-of-concept study to determine whether imazapyr or metsulfuron-methyl seed treatments could con-trol Striga hermonthica infection of sorghum. Although seed treatments resulted in reduced Striga growth and improved sorghum productivity, some herbicide damage was observed in young plants with reductions in grain yield and dry matter compared with untreated control plants under noninfested
conditions. This indicates that the level of herbicide tolerance in the heterozygous genotype evaluated in this study was not suffi cient to fully protect sorghum at the treatment rates used. Recent studies by Tuinstra and Al-Khatib (2007) have shown that ALS herbicide tolerance in sorghum is inherited as an incompletely dominant trait in F
1 hybrids with two to three modifi er
genes. Ongoing research eff orts are focused on incorporating ALS herbicide tolerance as a homozygous trait into locally adapted varieties with appropriate modifi ers to pro-vide higher levels of tolerance. These vari-
eties will then be used to deploy this trait and method for managing Striga infestations in sorghum.
Improving crop yield by delaying or preventing parasit-ism provides farmers with more resources and incentives to control reproduction of Striga (Abayo et al., 1998). Although no single technology is completely eff ective at controlling Striga or containing its spread, eff orts to integrate control options developed in diff erent disciplines have been shown to be highly eff ective at reducing damage by the parasite. Development of herbicide-tolerant varieties and use of herbicide seed treatments in integrated Striga management systems has shown promise for witchweed management in maize (Kanampiu et al., 2003; de Groote et al., 2007). These programs and successes are slowly expanding in dif-ferent parts of Africa. Eff orts in sorghum crop improvement are focused on combining ALS herbicide tolerance with various Striga resistance traits including low germination stimulant (Hess and Ejeta, 1992; Rodenburg et al., 2005), low haustorial factor (Rich et al., 2004), and hypersensitive response (Mohamed et al., 2003) to maximize opportuni-ties for integrated crop protection strategies against Striga infection. Successes in managing Striga achieved to date suggest that herbicide seed treatments may play a signifi cant role in developing and implementing of a successful Striga management program for sorghum in Africa.
Table 3. Mean squares from the ANOVA comparing herbicide seed treatment
effects on time to Striga emergence and Striga population at 60 and 90 d after
sorghum planting in fi eld trials in West Africa.
Country SourceStriga
EmergenceStriga
at 60 DAPStriga
at 90 DAP†
Niger Herbicide treatment 264*** 0.529*** 0.217**
Error 32.9 0.080 0.048
Mali Herbicide Treatment 158 NS‡ 0.048 NS 0.422*
Error 81.0 0.024 0.142
*Signifi cance at p ≤ 0.05.
**Signifi cance at p 0.01.
***Signifi cance at p 0.001.†DAP, days after planting.‡NS, nonsignifi cant.
Table 4. Herbicide treatment effects on time to Striga emergence and Striga number at 60 and 90 d after planting in fi eld trials.
TreatmentCinzana, Mali Konni, Niger
Rate Emergence Striga at 60 d Striga at 90 d Emergence Striga at 60 d Striga at 90 d
μg a.i. seed–1 d —————— Striga m–2 —————— d —————— Striga m–2 ——————
Control 0 55.3 1.1 10.2 31.2 19.7 46.1
Imazapyr 19 56.8 0.4 8.5 42.8 5.5 30.9
38 66.2 0.1 2.9 47.5 4.0 23.2
75 72.7 0 0.1 48.3 1.5 14.5
Metsulfuron 3 62.3 0 5.4 47.0 4.1 23.1
6 58.3 0.5 7.5 51.2 3.2 20.5
12 56.0 0.4 4.5 48.4 2.0 13.5
Mean 59.8 0.4 5.7 45.0 5.7 24.6
LSD0.05
NS† NS 7.6 6.8 6.4 14.2
†NS, nonsignifi cant.

CROP SCIENCE, VOL. 49, MAY–JUNE 2009 WWW.CROPS.ORG 929
CONCLUSIONSThese studies demonstrated that IMI and MET seed treat-ments contribute to improved Striga management in ALS herbicide–tolerant sorghum genotypes. Striga emerged more slowly and in fewer numbers in plots planted with herbicide treated seeds compared with untreated seeds. This is the fi rst report of successful use of ALS herbicide seed treat-ments on sorghum for controlling Striga hermonthica.
AcknowledgmentsThis research was funded in part by grants from the Interna-
tional Sorghum and Millet Collaborative Research Support
Program (INTSORMIL) and the C.T. de Wit Graduate School
for Production Ecology & Resource Conservation at Wagenin-
gen University, Wageningen, the Netherlands.
ReferencesAbayo, G.O., J.K. Ransom, J. Gressel, and G.D. Odhianbo. 1996.
Striga hermonthica control with acetolactate synthase inhibiting
herbicides seed-dressing to corn with target site resistance.
p. 762–768. In M.T. Moreno et al. (ed.) Advances in parasitic
weed research. Junta de Andalucia, Cordoba, Spain.
Abayo, G.O., T. English, R.E. Eplee, F.K. Kanampiu, J.K. Ran-
son, and J. Gressel. 1998. Control of parasitic witchweeds
(Striga spp.) on corn (Zea mays) resistant to acetolate synthase
inhibitors. Weed Sci. 46:459–466.
Berner, D.K., A.E. Awad, and E.I. Aigbokhan. 1994. Potential of
imazaquin seed treatment for control of Striga gesnerioides and
Alectra vogelii in cowpea (Vigna unguiculata). Plant Dis. 78:18–23.
Berner, D.K., B.B. Ikie, and B.B. Green. 1997. ALS-inhibiting
herbicide seed treatment control Striga hermonthica in ALS–
modifi ed corn (Zea mays L.). Weed Technol. 11:704–707.
Cechin, E.T., and M.C. Press. 1993. Nitrogen relations of the sor-
ghum-Striga hermonthica host-parasite association: Growth and
photosynthesis. Plant Cell Environ. 16:237–247.
Dembele, B., D. Dembele, and J.H. Westwood. 2005. Herbicide
seed treatments for control of purple witchweed (Striga her-
monthica) in sorghum and millet. Weed Technol. 19:629–635.
Diaz-Sanchez, J., N. Jurado-Exosito, F. Lopez-Granados, M.
Castejon-Munoz, and L. Garcia-Torres. 2003. Pronamide
applied to sunfl ower seeds for Orobanche cumana control. Weed
Technol. 17:314–319.
Diaz-Sanchez, J., N. Lopez-Martinez, F. Lopez-Granados, R. De
Prado, and L. Garcia-Torres. 2002. Absorption, translocation,
and fate of herbicides in Orobanche cumana–sunfl ower system.
Pest Biochem. Physiol. 74:9–15.
Ejeta, G. 2005. Integrating biotechnology, breeding, and agronomy
in the control of Striga in sorghum. p. 239–251. In R. Tuberosa et
al. (ed.) In the wake of the double helix: From the green revolu-
tion to the gene revolution. Avenue Media Press, Bologna, Italy.
Ejeta, G. 2007. The Striga scourge in Africa: A growing pandemic.
p. 3–16. In G. Ejeta and J. Gressel (ed.) Integrating new tech-
nologies for Striga control: Towards ending the witch-hunt.
World Scientifi c, Singapore.
Ejeta, G., and L.G. Butler. 1993. Host-parasite interactions
throughout the Striga life cycle and their contributions to
Striga resistance. Afr. Crop Sci. 1:75–80.
Greaves, J.A., G.K. Rufener, M.T. Chang, and P.H. Koehler. 1993.
Development of resistance to pursuit TM herbicide in corn.
p. 104–118. In D. Wilkinson (ed.) The IT Gene: Proc. of the
48th Annu. Corn and Sorghum Industry Research Conf. Am.
Seed Trade Assoc., Washington, DC.
de Groote, H. 2007. Striga economics. p. 265–280. In G. Ejeta and
J. Gressel (ed.) Integrating new technologies for Striga control:
Towards ending the witch-hunt. World Scientifi c, Singapore.
de Groote, H., L. Wangare, and F. Kanampiu. 2007. Evaluating
the use of herbicide-coated imidazolinone-resistant (IR)
maize seeds to control Striga in farmers’ fi elds in Kenya. Crop
Prot. 26:1496–1506.
Gurney, A.L., M.C. Press, and J.D. Scholes. 1999. Infection time
and density infl uence the response of sorghum to the parasitic
angiosperm Striga hermonthica. New Phytol. 143:573–580.
Hess, D.E., and G. Ejeta. 1992. Inheritance of resistance to Striga
in sorghum genotype SRN39. Plant Breed. 109:233–241.
Jurado-Esposito, M., M. Castejon-Munoz, and L. Garcia-Torres.
1996. Broomrape (Orobanche crenata) control with Imazethapyr
applied to pea (Pisum sativum) seed. Weed Technol. 10:774–780.
Jurado-Exposito, M., N. Castejon-Munoz, and L. Garcia-Torres.
1999. Uptake and translocation of Imazethapyr in peas as
aff ected by parasitism of Orobanche crenata and herbicide appli-
cation methods. Weed Res. 39:129–136.
Jurado-Exposito, M., L. Garcia, and M. Castejon-Munoz. 1997.
Broad bean and lentil seed treatment with imidazolinones
for the control of broomrape (Orobanche crenata). J. Agric. Sci.
129:307–314.
Kanampiu, F.K., V. Kabambe, C. Massawe, L. Jasi, D. Friesen, J.K.
Ransom, and J. Gressel. 2003. Multi-site, multi-season fi eld
tests demonstrate that herbicide seed-coating herbicide resis-
tance maize controls Striga spp. and increases yields in several
African countries. Crop Prot. 22:697–706.
Kanampiu, F.K., J.K. Ransom, D. Friesen, and J. Gressel. 2002.
Imazapyr and pyrithiobac movement in soil and from maize
seed coats to control Striga in legume intercropping. Crop
Prot. 21:611–619.
Kanampiu, F.K., J.K. Ransom, and J. Gressel. 2001. Imazapyr seed
dressings for Striga control on acetolactate synthase target-site
resistant maize. Crop Prot. 20:885–895.
Mohamed, A., A. Ellicott, T.L. Housley, and G. Ejeta. 2003.
Hypersensitive response to Striga infection in sorghum. Crop
Sci. 43:1320–1324.
Rich, P.J., C. Grenier, and G. Ejeta. 2004. Striga resistance in the
wild relatives of sorghum. Crop Sci. 44:2221–2229.
Rodenburg, J., L. Bastiaans, M.J. Kropff , and A. van Ast. 2006.
Eff ects of host plant genotype and seed bank density on Striga
reproduction. Weed Res. 46:251–263.
Rodenburg, J., L. Bastiaans, E. Weltzien, and D.E. Hess. 2005.
How can fi eld selection for Striga resistance and tolerance in
sorghum be improved? Field Crops Res. 93:34–50.
Tuinstra, M.R., and K. Al-Khatib. 2007. New herbicide tolerance
traits in sorghum. In Proc. of the 2007 Corn, Sorghum, and
Soybean Seed Research Conf. and Seed Expo, Chicago, IL.
5–7 Dec. 2007. Am. Seed Trade Assoc., Alexandria, VA.
van Ast, A., and L. Bastiaans. 2006. The role of infection time in
the diff erential response of Sorghum cultivars to Striga her-
monthica infection. Weed Res. 46:71–89.
van Ast, A., L. Bastiaans, and S. Katile. 2005. Cultural control mea-
sures to diminish sorghum yield loss and parasite success under
Striga hermonthica infestation. Crop Prot. 24:1023–1034.
Wright, T.R., and D. Penner. 1998. Corn (Zea mays) acetolac-
tate synthase sensitivity to four of ALS-inhibiting herbicides.
Weed Sci. 46:8–12.




















