
Modulation of corticospinal excitability by paired associative stimulation: Reproducibility of...
8
Modulation of corticospinal excitability by paired associative stimulation: Reproducibility of effects and intraindividual reliability Fabiana Fratello a , Domenica Veniero a,b , Giuseppe Curcio a , Michele Ferrara c , Cristina Marzano a , Fabio Moroni a , Maria Concetta Pellicciari a,d , Mario Bertini a , Paolo Maria Rossini d,e,f , Luigi De Gennaro a,d,g, * a Department of Psychology, University of Rome ‘‘La Sapienza’’, Rome, Italy b Department of Biomedical Science and Biotechnology, University of Brescia, Italy c Department of Internal Medicine and Public Health, University of L’Aquila, Italy d IRCCS ‘‘Centro S. Giovanni di Dio-FBF-’’, Brescia, Italy e AFaR, Department of Neuroscience, Hosp. ‘‘Fatebenefratelli’’, Isola Tiberina, Rome, Italy f Department of Neurology, University ‘‘Campus Biomedico’’, Rome, Italy g ‘‘Universita ` Europea di Roma’’, Rome, Italy Accepted 28 July 2006 Available online 2 October 2006 Abstract Objective: Electrical stimulation of the median nerve followed by a magnetic pulse on the primary motor cortex (M1) is effective to cause an increase in the amplitude of motor evoked potential (MEP) registered in the target muscle with the interstimulus interval (ISI) at 25 ms (paired associative stimulation, PAS). The aim of this study is to evaluate the reproducibility of PAS with ISI 25 (PAS25), assessed in two separate sessions. Intraindividual reliability of TMS measures was also evaluated. Methods: Motor threshold of abductor pollicis brevis (APB), assessed at rest, and MEP amplitude of APB and abductor digiti minimi (ADM) were assessed before and after PAS25 in 18 healthy volunteers (nine males and nine females). Results: Data showed a significant increase of MEP amplitude in the target muscle (APB) after PAS25 and a reproducibility of group effect in the two sessions, as assessed by ANOVA, but a lack of intraindividual reliability, as assessed by intraclass correlation coefficients (ICC). Conclusions: The results underline the reproducibility of mean effects and the need to be careful when comparing the same subject on different days. Significance: Electrical stimulation of the median nerve followed by a magnetic pulse delivered on M1 after 25 ms causes a reproducible increase in MEP amplitude, without showing an acceptable intraindividual reliability. Ó 2006 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved. Keywords: PAS; TMS; Reproducibility; Reliability; Cortical plasticity; Peripheral stimulation 1. Introduction Ever since Barker et al. (1985) applied the Faraday rules to develop a non-invasive painless technique to assess cor- ticospinal excitability, the use of transcranial magnetic stimulation (TMS) in human studies and in clinical settings has been growing. In the last decade, TMS has been used to measure plastic cortical changes after different experimen- tal procedures (Rossini and Pauri, 2000; Siebner and Roth- well, 2003). TMS can be employed for this in two ways. First, it can be used to evaluate variations in cortical 1388-2457/$32.00 Ó 2006 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.clinph.2006.07.315 * Corresponding author. Tel.: +39 06 49917647; fax: +39 06 4451667. E-mail address: [email protected] (L. De Gennaro). www.elsevier.com/locate/clinph Clinical Neurophysiology 117 (2006) 2667–2674
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Modulation of corticospinal excitability by paired associative stimulation: Reproducibility of...

www.elsevier.com/locate/clinph
Clinical Neurophysiology 117 (2006) 2667–2674
Modulation of corticospinal excitability by pairedassociative stimulation: Reproducibility of effects
and intraindividual reliability
Fabiana Fratello a, Domenica Veniero a,b, Giuseppe Curcio a, Michele Ferrara c,Cristina Marzano a, Fabio Moroni a, Maria Concetta Pellicciari a,d, Mario Bertini a,
Paolo Maria Rossini d,e,f, Luigi De Gennaro a,d,g,*
a Department of Psychology, University of Rome ‘‘La Sapienza’’, Rome, Italyb Department of Biomedical Science and Biotechnology, University of Brescia, Italyc Department of Internal Medicine and Public Health, University of L’Aquila, Italy
d IRCCS ‘‘Centro S. Giovanni di Dio-FBF-’’, Brescia, Italye AFaR, Department of Neuroscience, Hosp. ‘‘Fatebenefratelli’’, Isola Tiberina, Rome, Italy
f Department of Neurology, University ‘‘Campus Biomedico’’, Rome, Italyg ‘‘Universita Europea di Roma’’, Rome, Italy
Accepted 28 July 2006Available online 2 October 2006
Abstract
Objective: Electrical stimulation of the median nerve followed by a magnetic pulse on the primary motor cortex (M1) is effective to causean increase in the amplitude of motor evoked potential (MEP) registered in the target muscle with the interstimulus interval (ISI) at25 ms (paired associative stimulation, PAS). The aim of this study is to evaluate the reproducibility of PAS with ISI 25 (PAS25), assessedin two separate sessions. Intraindividual reliability of TMS measures was also evaluated.Methods: Motor threshold of abductor pollicis brevis (APB), assessed at rest, and MEP amplitude of APB and abductor digiti minimi
(ADM) were assessed before and after PAS25 in 18 healthy volunteers (nine males and nine females).Results: Data showed a significant increase of MEP amplitude in the target muscle (APB) after PAS25 and a reproducibility of groupeffect in the two sessions, as assessed by ANOVA, but a lack of intraindividual reliability, as assessed by intraclass correlation coefficients(ICC).Conclusions: The results underline the reproducibility of mean effects and the need to be careful when comparing the same subject ondifferent days.Significance: Electrical stimulation of the median nerve followed by a magnetic pulse delivered on M1 after 25 ms causes a reproducibleincrease in MEP amplitude, without showing an acceptable intraindividual reliability.� 2006 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved.
Keywords: PAS; TMS; Reproducibility; Reliability; Cortical plasticity; Peripheral stimulation
1. Introduction
Ever since Barker et al. (1985) applied the Faraday rulesto develop a non-invasive painless technique to assess cor-
1388-2457/$32.00 � 2006 International Federation of Clinical Neurophysiolo
doi:10.1016/j.clinph.2006.07.315
* Corresponding author. Tel.: +39 06 49917647; fax: +39 06 4451667.E-mail address: [email protected] (L. De Gennaro).
ticospinal excitability, the use of transcranial magneticstimulation (TMS) in human studies and in clinical settingshas been growing. In the last decade, TMS has been used tomeasure plastic cortical changes after different experimen-tal procedures (Rossini and Pauri, 2000; Siebner and Roth-well, 2003). TMS can be employed for this in two ways.First, it can be used to evaluate variations in cortical
gy. Published by Elsevier Ireland Ltd. All rights reserved.

2668 F. Fratello et al. / Clinical Neurophysiology 117 (2006) 2667–2674
connectivity induced by different experimental paradigms,such as by prolonged peripheral nerve stimulation (Rid-ding et al., 2000) or muscle electric stimulation (Riddingand Uy, 2003; Pyndt and Ridding, 2004). The presence ofplastic changes was shown by an increase in MEP ampli-tude after the application of these stimulations. Pioneeringstudies had previously shown that conditioning stimuli onperipheral nerve-fibers, followed by a test stimulation onthe primary motor cortex via transcranial electrical or mag-netic stimuli, depict a clearcut inhibitory/facilitatory influ-ence (Mariorenzi et al., 1991; Kasai et al., 1992). Namely,when the interstimulus interval between the peripheralnerve and the cortical stimulation is appropriate for senso-ry input arrival to the controlateral sensorimotor cortex,this cortex is facilitated at a level that triggers largerresponses to individual TMS (Deuschl et al., 1991; Mario-renzi et al., 1991).
One of the limitations of these paradigms concerns theirlength: depending on the specific protocol, one (Riddingand Uy, 2003; Pyndt and Ridding, 2004) or even two hours(Ridding et al., 2000) of continuative stimulation are neces-sary to obtain changes in cortical excitability. This limita-tion can be avoided by using TMS in the second way, i.e.to directly induce plastic cortical changes. It has beenshown, for example, that by pairing a train of peripheralstimulation with TMS, the time and number of stimula-tions required to produce cortical excitability changes canbe shortened to 30 min (Ridding and Taylor, 2001; Charl-ton et al., 2003). Accordingly, the coupling of a single elec-tric stimulus delivered on a peripheral nerve with a singleTMS pulse causes changes in the excitability of the motor(Stefan et al., 2000, 2002; Wolters et al., 2003) and sensorycortex (Wolters et al., 2005) with a shorter (about 20 min)and more comfortable stimulation. These plastic changesdepend on a temporally asymmetric Hebbian rule (Hebb,1949); hence, a shorter Inter Stimulus Interval(ISI = 10 ms) produces a decrease in the motor evokedpotential (MEP), measured after delivering a TMS singlepulse, and a longer interval (ISI = 25 ms) induces anincrease in MEP amplitude (Wolters et al., 2003). Theseinterventions were, respectively, termed PAS10 (PairedAssociative Stimulation with ISI = 10 ms) and PAS25(Paired Associative Stimulation with ISI = 25 ms) (Classenet al., 2004). The PAS-related excitability change evolvesrapidly (within 30 min), is long lasting (at least 60 min),reversible and cortically generated (Stefan et al., 2000).The changes in MEP amplitude were blocked by dextro-methorphan, showing a PAS dependence on N-methyl-D-aspartate (NMDA) glutamatergic receptors. Consideringthese properties, a LTP-like mechanism mediating thePAS-related effects was proposed (Stefan et al., 2000; Wol-ters et al., 2003).
Recently, PAS was employed to investigate cerebralplasticity in different groups of patients suffering fromParkinson’s disease (Bagnato et al., 2005) and writer’scramp (Quartarone et al., 2003) compared to healthy sub-jects. For example, Bagnato and co-workers (2005)
showed an abnormal sensorimotor cortical plasticity inParkinson’s disease, as shown by a more pronouncedincrease in MEP amplitude after PAS in off-medicationpatients compared to healthy controls. The use of PASalso in clinical settings and the comparison of patientsand healthy controls requires a more complete assessmentof the normative data on healthy subjects and on thereproducibility of PAS effects, with particular attentionto inter-individual and intra-individual differences. Withrespect to these issues, indeed, the effect of PAS has beennot evaluated.
In view of this, the present study examined the stabilityof TMS measures in cortical plasticity studies in healthyhumans. Hence, it aimed to assess the magnitude of thePAS effect and its reproducibility over time, within subjectsand in the group as a whole.
2. Methods
2.1. Subjects
Eighteen right-handed healthy experimental subjects(nine males and nine females, mean age = 26.11,SE = 0.62) were selected from a university student popula-tion. In a clinical interview, subjects reported the absenceof epilepsy or other neurological disease in themselvesand in their family history. All subjects gave their writteninformed consent and the study was approved by the localInstitutional Ethics Committee and was conducted inaccordance with the Declaration of Helsinki.
2.2. Materials
2.2.1. Transcranial magnetic brain stimulation and electrical
nerve stimulation
The study used a Magstim 200 Mono Pulse connected toa Bistim module and a figure-of-eight coil with an externaldiameter of 9 cm (Magstim Company Limited, UK). Thepeak magnetic field produced by such a coil is 2.0 T. Elec-trical stimulation was delivered using a bipolar electrode(cathode proximal) placed on the right median nerve atthe level of the wrist, connected to an electromyograph(Myto, EBNeuro, Italy).
2.2.2. Electromyographic recording
Motor evoked potentials (MEPs) were obtained fromthe right APB by positioning two Ag–AgCl surface cupelectrodes of 9 mm diameter with the active electrode onthe muscle belly and the reference electrode on the metacar-pophalangeal joint of the thumb. The same montage wasused for the abductor digiti minimi (ADM) with the refer-ence electrode on the metacarpophalangeal joint of the lit-tle finger. In both cases the skin-electrode conductance waskept to <10 kX. The electromyographic signals were band-pass filtered between 1 and 1 kHz, recorded using an elec-tromyogram (EMG)-dedicated software (Myto, EBNeuro,Italy), and stored in a computer for off-line analysis.

F. Fratello et al. / Clinical Neurophysiology 117 (2006) 2667–2674 2669
2.3. Experimental procedure
Subjects were seated on a comfortable chair in a sound-proof room and tested in two identical sessions (test andretest) a week apart. Test and retest sessions were carriedout at the same time of day to avoid circadian effects.The coil was held tangentially to the skull, 45� from themidsagittal axis of the subject’s head to stimulate fiberstravelling horizontally with respect to the cortical surface(Rothwell, 1997). The optimal site of stimulation (‘‘hot-spot’’) for eliciting MEPs in the right abductor pollicis bre-
vis (APB) was chosen by positioning the coil approximatelyover the central sulcus and moving it on the scalp in 0.5 cmsteps on the primary motor area (M1) of the left cortex.The hotspot was marked on the scalp with a soft-tip pen.
Motor threshold (MT) was assessed at rest over theAPB-hot spot and corresponded to the lower intensity ofstimulation able to produce at least 100 lV in the relaxedmuscle in at least five of ten consecutive stimulations (Ros-sini et al., 1994). Stimulation started at suprathresholdintensity (about 60% of stimulator output) and wasdecreased in steps of 1–2% of the stimulator output (Roth-well et al., 1999). Then the intensity of stimulation neces-sary to elicit reliable MEPs in 90% of a cascade of 10stimuli was assessed. This intensity was 132%(SEM = 0.03) of the individual motor threshold in the testsession (intensity output = 55.55 ± 1.45) and 132%(SEM = 0.02) in the retest session (intensity out-put = 56.8 ± 1.47), and it was kept constant throughoutthe session.
PAS consisted of 140 pairs of stimuli delivered during23 min. The magnetic stimulus, delivered on the optimalsite for APB on the left cortex, was preceded by an electri-cal stimulus delivered to the right median nerve at wristlevel, with an ISI of 25 ms (Stefan et al., 2000, 2002). Theelectrical stimulus lasted 200 ls and had an intensity ofthree times the perceptual threshold (averaged intensityof electric stimulation = 5.0 mA, SE = 0.2 in the test ses-sion; 5.0 mA, SE = 0.2 in the retest session). The repetitionrate of paired stimulation was 0.1 Hz (Stefan et al., 2006).During PAS subjects were asked to watch their own righthand, since this condition has been demonstrated to givethe maximal PAS-induced plasticity (Stefan et al., 2004).The experimental session lasted about 90 min. Before andafter PAS, MT of the APB was assessed and twenty MEPselicited by single magnetic pulses were collected with a rateof stimulation of 0.20 Hz.
Subjects were required to keep their right arm and handcompletely relaxed. Muscle relaxation was monitored bygiving subjects visual feedback of their EMG, and trialscontaminated with voluntary EMG activity were discardedfrom further analysis.
2.4. Data analysis
The dependent variables considered in the study wereMT of APB, before (pre-PAS) and after PAS (post-PAS),
and MEP amplitude recorded in the target muscle (APB)and in a control muscle (ADM), in test and retest sessions.
The data were log-transformed before any statisticalprocedure, in order to obtain a better approximation togaussianity and a higher homoschedasticity.
2.4.1. Motor thresholds
Resting motor thresholds were analyzed by a within-subjects ANOVA design Treatment (pre-PAS vs. post-PAS) · Session (test vs. retest). This analysis aimed toevaluate differences in MTs after the intervention and thestability between sessions. The MT was expressed as themagnetic stimulator output ± SEM.
2.4.2. Treatment effect and reproducibility of the effect
MEP amplitude was measured peak-to-peak. To testchanges in APB–MEP amplitude after PAS and to testthe reproducibility of the effect, a within-subjects ANOVATreatment (pre-PAS vs. post-PAS) · Session (test vs. ret-est) was performed. The same analysis was carried out onADM–MEPs to assess the specificity of the treatmenteffect. The significance of main effects and interactionswas set at 0.05.
2.4.3. Intraindividual reliability
Intraindividual reliability of MTs was assessed by intra-class correlation coefficients (ICC) between the two ses-sions. The same correlation was carried out on changesin MEP amplitude as a result of treatment, expressed asthe ratio between pre-PAS responses divided by responsesobtained after PAS.
3. Results
3.1. Motor thresholds
In the test session, the mean MT was 42.4% ± 1.5% ofthe maximum stimulator output before PAS and41.9% ± 1.51% after PAS. In the retest session, the meanMT was 43.3% ± 1.4% before PAS and 43.3% ± 1.4% afterPAS. As a result of these small changes, the Treatment ·Session ANOVA on log-transformed MT showed no sig-nificant main effect for Treatment (F1,17 = 0.51;p = 0.48), no significant main effect for Session (F1,17 =2.72; p = 0.12) and no significant interaction (F1,17 =1.12; p = 0.30).
3.2. Treatment effect
Paired associative stimulation was effective in increasingAPB–MEP amplitude in the muscle affected by PAS (APB)both in the test and retest session (Fig. 1a). MEP amplitudeincreased by 36.4% ± 10.3% in the test session and by31.7% ± 7.6% in the retest session.
The Treatment · Session ANOVA on log transformedresting MEPs in APB showed a significant effect for Treat-ment (F1,17 = 29.77; p = 0.00004), while the main effect for
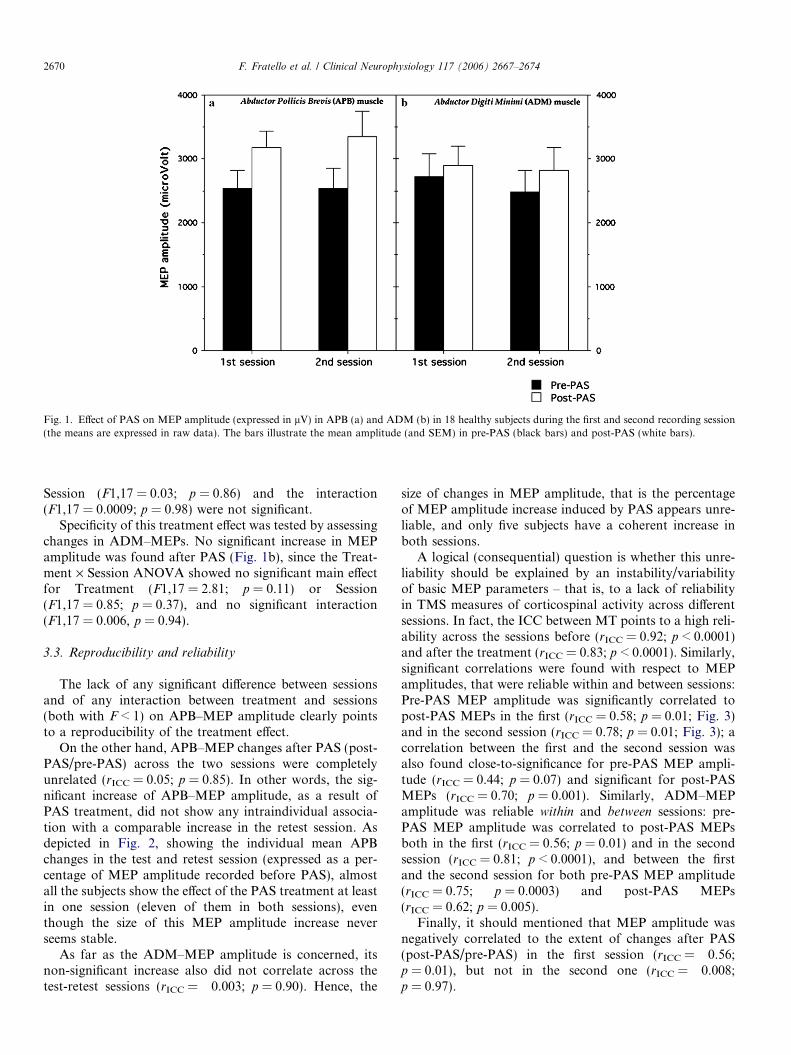
Fig. 1. Effect of PAS on MEP amplitude (expressed in lV) in APB (a) and ADM (b) in 18 healthy subjects during the first and second recording session(the means are expressed in raw data). The bars illustrate the mean amplitude (and SEM) in pre-PAS (black bars) and post-PAS (white bars).
2670 F. Fratello et al. / Clinical Neurophysiology 117 (2006) 2667–2674
Session (F1,17 = 0.03; p = 0.86) and the interaction(F1,17 = 0.0009; p = 0.98) were not significant.
Specificity of this treatment effect was tested by assessingchanges in ADM–MEPs. No significant increase in MEPamplitude was found after PAS (Fig. 1b), since the Treat-ment · Session ANOVA showed no significant main effectfor Treatment (F1,17 = 2.81; p = 0.11) or Session(F1,17 = 0.85; p = 0.37), and no significant interaction(F1,17 = 0.006, p = 0.94).
3.3. Reproducibility and reliability
The lack of any significant difference between sessionsand of any interaction between treatment and sessions(both with F < 1) on APB–MEP amplitude clearly pointsto a reproducibility of the treatment effect.
On the other hand, APB–MEP changes after PAS (post-PAS/pre-PAS) across the two sessions were completelyunrelated (rICC = 0.05; p = 0.85). In other words, the sig-nificant increase of APB–MEP amplitude, as a result ofPAS treatment, did not show any intraindividual associa-tion with a comparable increase in the retest session. Asdepicted in Fig. 2, showing the individual mean APBchanges in the test and retest session (expressed as a per-centage of MEP amplitude recorded before PAS), almostall the subjects show the effect of the PAS treatment at leastin one session (eleven of them in both sessions), eventhough the size of this MEP amplitude increase neverseems stable.
As far as the ADM–MEP amplitude is concerned, itsnon-significant increase also did not correlate across thetest-retest sessions (rICC = �0.003; p = 0.90). Hence, the
size of changes in MEP amplitude, that is the percentageof MEP amplitude increase induced by PAS appears unre-liable, and only five subjects have a coherent increase inboth sessions.
A logical (consequential) question is whether this unre-liability should be explained by an instability/variabilityof basic MEP parameters – that is, to a lack of reliabilityin TMS measures of corticospinal activity across differentsessions. In fact, the ICC between MT points to a high reli-ability across the sessions before (rICC = 0.92; p < 0.0001)and after the treatment (rICC = 0.83; p < 0.0001). Similarly,significant correlations were found with respect to MEPamplitudes, that were reliable within and between sessions:Pre-PAS MEP amplitude was significantly correlated topost-PAS MEPs in the first (rICC = 0.58; p = 0.01; Fig. 3)and in the second session (rICC = 0.78; p = 0.01; Fig. 3); acorrelation between the first and the second session wasalso found close-to-significance for pre-PAS MEP ampli-tude (rICC = 0.44; p = 0.07) and significant for post-PASMEPs (rICC = 0.70; p = 0.001). Similarly, ADM–MEPamplitude was reliable within and between sessions: pre-PAS MEP amplitude was correlated to post-PAS MEPsboth in the first (rICC = 0.56; p = 0.01) and in the secondsession (rICC = 0.81; p < 0.0001), and between the firstand the second session for both pre-PAS MEP amplitude(rICC = 0.75; p = 0.0003) and post-PAS MEPs(rICC = 0.62; p = 0.005).
Finally, it should mentioned that MEP amplitude wasnegatively correlated to the extent of changes after PAS(post-PAS/pre-PAS) in the first session (rICC = �0.56;p = 0.01), but not in the second one (rICC = �0.008;p = 0.97).

Fig. 2. Effect of PAS on MEP amplitude recorded in APB, expressed as a percentage (post-PAS/pre-PAS) for individual subjects in the first (black bars)and second session (white bars) in APB. Zero corresponds to the pre-PAS MEP amplitude.
F. Fratello et al. / Clinical Neurophysiology 117 (2006) 2667–2674 2671
4. Discussion
Paired nerve-brain stimulation is known to influence theexcitability of the sensorimotor cortex, when conditioningstimuli on peripheral nerves governing a given body districtare properly timed with respect to transcranial stimulationof the motor cortex to muscles of the same body regions(Deuschl et al., 1991; Mariorenzi et al., 1991; Kasaiet al., 1992). Within this theoretical framework, prolongednerve-brain paired stimuli – namely in accordance with thePAS protocol – can modify excitability of the related motor
Fig. 3. Correlations between MEP amplitude obtained pre-PAS and post-PAS in the APB muscle, assessed by intraclass correlations. The values areexpressed in lV and refer to both sessions (test and retest).
output for a relatively long time (Stefan et al., 2000; Wol-ters et al., 2003).
This study confirms the efficacy of PAS in causing plas-tic changes in motor cortex excitability and shows thereproducibility of this effect when the averaged data areconsidered. On the other hand, this effect does not showan intraindividual stability. This lack of intraindividual sta-bility in the efficacy of PAS changes does not seem attrib-utable to the unreliability of the basic parameters ofcorticospinal activity, that is, motor thresholds and uncon-ditioned MEP amplitudes.
4.1. Treatment effect
4.1.1. MEP amplitude
As shown by Wolters and co-workers (2003), an intervalof 25 ms between the peripheral electric stimulus and thecentral magnetic pulse is able to induce an enhancementof the excitability in the human motor cortex. The specific-ity of this effect is demonstrated by the non-significant var-iation of MEP amplitude in ADM after PAS.
The present results show an increase of MEP amplitudeafter PAS ranging between +32% and +36%. Previousstudies had shown different percentage increases rangingbetween 40% (Stefan et al., 2002) and 79% (Quartaroneet al., 2003). It must be considered that the effect is notpresent in all the subjects undergoing the study, as shownby our data (14 of the 18 subjects in the test session and15 of the 18 subjects in the retest session) and previous data(11 out of 13 subjects in Wolters et al. (2003); 13 out of 17subjects in Stefan et al. (2004); 10 out of 12 subjects in Ste-fan et al. (2006)). For this reason, we hypothesize that theintersubject differences may play a determinant role in themagnitude of the effects.

2672 F. Fratello et al. / Clinical Neurophysiology 117 (2006) 2667–2674
4.2. Reproducibility of the considered measures over time
4.2.1. Treatment effect
The present results show a large degree of variability inthe interindividual magnitude of PAS-related effects and alow intraindividual reliability of the effect on different days(see Fig. 2). Nevertheless, the group effects show that thePAS protocol is effective in modulating cortical excitability,and this effect is reproducible, as shown by the lack of anydifferences in the mean effect between sessions evaluated byANOVA. This result is in agreement with previous obser-vations that showed no significant differences between thePAS-effect obtained in two different recording sessions(Ziemann et al., 2004; Stefan et al., 2005). This findingpoints to a good reproducibility of the PAS-effect. On theother hand, as regards the reliability of the effect, pertain-ing to a within-subjects correlation between the dependentvariables considered at different times (De Gennaro et al.,2003), this is not guaranteed. Reliability of the PAS-effectis based on the intraindividual correlation between theratio of change (MEPs obtained post-PAS/pre-PAS) in dif-ferent sessions. When this analysis is performed, large dif-ferences emerge, as demonstrated by the very lowcorrelations reported in this work. Furthermore, largerpre-PAS amplitudes were associated with a smaller PAS-ef-fect, since MEP amplitude was negatively correlated to theextent of changes after PAS, but only in the first session.Hence, specific aspects of the PAS effect again do not seemto be reliable. This lack of reliability does not pertain exclu-sively to PAS effects, but it is common to different TMStreatments and it involves both single-pulse TMS andrepetitive TMS (rTMS) (e.g., Boroojerdi et al., 2000).
In a previous study that used a different TMS design(interhemispheric paired pulses), we reported no significantcorrelation between the ratio of changes in MEP amplitudein two different sessions (De Gennaro et al., 2003),although the mean effect obtained in the two sessions didnot significantly differ. In the same way, McDonnell andcolleagues (2004) compared two blocks recorded in thesame session and showed no significant differences inMEP amplitude assessed by ANOVA, but a moderate reli-ability of MEP amplitude in the muscles concerned, asassessed by intraclass correlation analysis (ICC = 0.46–0.55). It has been repeatedly shown that also the reproduc-ibility of cortical inhibition and facilitation in differentsessions is problematic (Boroojerdi et al., 2000; Maedaet al., 2002), as well the reproducibility of effects of rTMSon cortical plasticity (Maeda et al., 2000) and the evalua-tion of MEP amplitude size (Humm et al., 2004).
Carroll and colleagues (2001) speculated that the largervariability of TMS measures with respect to TES (transcra-nial electrical stimulation) is probably due to the non-com-plete overlapping of stimulation sites between sessions.This issue should also be considered in the present study.Moreover, fluctuations in subjects’ attention within andbetween sessions have to be considered (Ellaway et al.,1998; Funase et al., 1999). The TMS session is well tolerated
but considered boring by our experimental subjects. Thisholds especially true for the retest session, characterizedby more familiarity with TMS, so it is very difficult to guar-antee the same level of attention. This issue seems relevantalso because it has been recently shown that asking subjectsto focus attention on the stimulated hand (Stefan et al.,2004), or to focus attention on the stimulated hand andcounting peripheral electrical stimuli (Rosenkranz andRothwell, 2006), increases magnitude of PAS-inducedchanges. In the current study, we have chosen the experi-mental procedure showing the maximal PAS-induced plas-ticity, that is asking the subjects to watch their target handduring peripheral stimulation (Stefan et al., 2004). Howev-er, we had no direct measure of attention, and the possibil-ity that poor intrasubject reproducibility was due todifferent intraindividual levels of attention cannot be com-pletely discarded.
Finally, prestimulus muscle contraction should also beconsidered, since this parameter affects the variability ofMEP amplitudes in healthy participants, with a decreasedvariability with increased prestimulus muscle contraction(Kiers et al., 1993). Although muscle relaxation was con-trolled by visual inspection of EMG, one cannot rule outthat small changes actually affected the recordings ofTMS measures. However, there is no reason to accept thisinterpretation for the changes induced by the PAS effect,while it does not affect MTs and MEP amplitudes.
It should be remembered that the lack of reliabilitybetween different sessions is not limited to TMS. Eventhe reproducibility of other techniques, such as fMRI(functional magnetic resonance imaging), is controversial.For example, some studies stressed that fMRI is betterfor defining brain activation qualitatively than quantita-tively (Liu et al., 2004; Gustard et al., 2001; Yetkin et al.,1996; Yoo et al., 2005; Willem et al., 2000); in other words,fMRI can quite reliably reproduce the locations of activa-tion, while their sizes fluctuate substantially. In our opin-ion, the TMS has similar problems of reliability as thefMRI, and, similarly, the reproducibility of its measuresdepends on the data treatment method. Hence, currentresults suggest taking great care when evaluating differentTMS parameters in repeated measure studies, and whencomparing healthy subjects and patients. An improvementof TMS reliability may be obtained with more careful coilpositioning with the help of neuronavigation that can guidethe correct repositioning of the coil in the same area in dif-ferent sessions, and by using an adaptation session toenhance the familiarity of experimental subjects withTMS, conforming the level of attention between sessionsas much as possible.
4.2.2. Motor threshold
In contrast with MEP amplitude, MTs remainunchanged over time, showing a high reliability with highcorrelations between sessions (test–retest). This result isin agreement with previous findings (Traversa et al.,1998; Maeda et al., 2002; Corneal et al., 2005; Mortifee

F. Fratello et al. / Clinical Neurophysiology 117 (2006) 2667–2674 2673
et al., 1994; Wolf et al., 2004; Kimiskidis et al., 2004). Kim-iskidis and co-workers (2004), for example, tested 84 sub-jects in six sessions and showed a satisfactory level ofreliability in the three threshold measurements considered(lower threshold, upper threshold, standard threshold),despite wide inter-subject variations. These data makeMT the most stable TMS parameter.
Although of different magnitude, the reliability of theother measure of corticospinal activity, i.e. unconditionedMEP amplitude, also seems acceptable.
5. Conclusions
In conclusion, our results confirmed that the pairedstimulation proposed by Stefan and co-workers (2000) iseffective in causing topographically specific excitabilitychanges in the human motor cortex, as indexed by anincrease of MEP amplitude ranging between +32% and+36%. This effect is reproducible in different sessions, asshown by the lack of any differences in the group measureobtained in test and retest sessions. The large inter- andintra-subject variability can explain the lack of correlationbetween sessions when we consider the individual values.
References
Bagnato S, Agostino R, Modugno N, Quartarone A, Berardelli A.Plasticity of the motor cortex in Parkinson’s disease patients on and offtherapy. Mov Disord 2005, Epub ahead of print.
Barker AT, Freeston IL, Jalinous R, Jarratt JA. Non-invasive stimulationof motor pathways within the brain using time-varying magnetic fields.Electroenceph Clin Neurophysiol 1985;61:S245.
Boroojerdi B, Kopylev L, Battaglia F, Facchini S, Ziemann U,Muellbacher W, et al. Reproducibility of intracortical inhibitionand facilitation using the paired-pulse paradigm. Muscle Nerve2000;23:1594–7.
Carroll TJ, Riek S, Carson RG. Reliability of the input-output propertiesof the cortico-spinal pathway obtained from transcranial magnetic andelectrical stimulation. J Neurosci Methods 2001;112:193–202.
Charlton CS, Ridding MC, Thompson PD, Miles TS. Prolongedperipheral nerve stimulation induces persistent changes in excitabilityof human motor cortex. J Neurol Sci 2003;208:79–85.
Classen J, Wolters A, Stefan K, Wycislo M, Sandbrink F, Schmidt A,et al. Paired associative stimulation. Suppl Clin Neurophysiol2004;57:563–9.
Corneal SF, Butler AJ, Wolf SL. Intra- and intersubject reliability ofabductor pollicis brevis muscle motor map characteristics withtranscranial magnetic stimulation. Arch Phys Med Rehabil2005;86:1670–5.
De Gennaro L, Ferrara M, Bertini M, Pauri F, Cristiani R, Curcio G,et al. Reproducibility of callosal effects of transcranial magneticstimulation (TMS) with interhemispheric paired pulses. Neurosci Res2003;46:219–27.
Deuschl G, Michels R, Berardelli A, Schenck E, Inghilleri M, LuckingCH. Effects of electric and magnetic transcranial stimulation on longlatency reflexes. Exp Brain Res 1991;83:403–10.
Ellaway PH, Davey NJ, Maskill DW, Rawlinson SR, Lewis HS,Anissimova NP. Variability in the amplitude of skeletal muscleresponses to magnetic stimulation of the motor cortex in man.Electroencephal Clin Neurophysiol 1998;109:104–13.
Funase K, Miles TS, Gooden BR. Trial-to-trial fluctuations in H-reflexesand motor evoked potentials in human wrist flexor. Neurosci Lett1999;271:25–8.
Gustard S, Fadili J, Williams EJ, Hall LD, Carpenter TA, Brett M, et al.Effect of slice orientation on reproducibility of fMRI motor activationat 3 Tesla. Magn Reson Imaging 2001;19:1323–31.
Hebb DO. The organization of behavior: a neuropsychological theo-ry. New York: Wiley; 1949.
Humm AM, Z’Graggen WJ, von Hornstein NE, Magistris MR,Rosler KM. Assessment of central motor conduction tointrinsic hand muscles using the triple stimulation technique:normal values and repeatability. Clin Neurophysiol 2004;115:2558–66.
Kasai T, Hayes KC, Wolfe DL, Allatt RD. Afferent conditioning of motorevoked potentials following transcranial magnetic stimulation ofmotor cortex in normal subjects. Electroenceph Clin Neurophysiol1992;85:95–101.
Kiers L, Cros D, Chiappa KH, Fang J. Variability of motor potentialsevoked by transcranial magnetic stimulation. Electroenceph ClinNeurophysiol 1993;89:415–23.
Kimiskidis VK, Papagiannopoulos S, Sotirakoglou K, Kazis DA,Dimopoulos G, Kazis A, et al. The repeatability of corticomotorthreshold measurements. Neurophysiol Clin 2004;34:259–66.
Liu JZ, Zhang L, Brown RW, Yue GH. Reproducibility of fMRI at1.5 T in a strictly controlled motor task. Magn Reson Med2004;52:751–60.
Maeda F, Keenan JP, Tormos JM, Topka H, Pascual-Leone A.Modulation of corticospinal excitability by repetitive transcranialmagnetic stimulation. Clin Neurophysiol 2000;111:800–5.
Maeda F, Gangitano M, Thall M, Pascual-Leone A. Inter- and intra-individual variability of paired-pulse curves with transcranial magneticstimulation (TMS). Clin Neurophysiol 2002;113:376–82.
Mariorenzi R, Zarola F, Caramia MD, Paradiso C, Rossini PM. Non-invasive evaluation of central motor tract excitability changes follow-ing peripheral nerve stimulation in healthy humans. ElectroencephClin Neurophysiol 1991;81:90–101.
McDonnell MN, Ridding MC, Miles TS. Do alternate methods ofanalysing motor evoked potentials give comparable results? J NeurosciMethods 2004;136:63–7.
Mortifee P, Stewart H, Schulzer M, Eisen A. Reliability of transcranialmagnetic stimulation for mapping the human motor cortex. Electro-enceph Clin Neurophysiol 1994;93:131–7.
Pyndt HS, Ridding MC. Modification of the human motor cortex byassociative stimulation. Exp Brain Res 2004;159:123–8.
Quartarone A, Bagnato S, Rizzo V, Siebner HR, Dattola V, Scalfari A,et al. Abnormal associative plasticity of the human motor cortex inwriter’s cramp. Brain. 2003;126:2586–96.
Ridding MC, Brouwer B, Miles TS, Pitcher JB, Thompson PD. Changesin muscle responses to stimulation of the motor cortex induced byperipheral nerve stimulation in human subjects. Exp Brain Res2000;131:135–43.
Ridding MC, Taylor JL. Mechanisms of motor-evoked potential facili-tation following prolonged dual peripheral and central stimulation inhumans. J Physiol 2001;537:623–31.
Ridding MC, Uy J. Changes in motor cortical excitability inducedby paired associative stimulation. Clin Neurophysiol 2003;114:1437–44.
Rosenkranz K, Rothwell JC. Differences between the effects of threeplasticity inducing protocols on the organization of the human motorcortex. Eur J Neurosci 2006;23:822–9.
Rossini PM, Barker AT, Berardelli A, Caramia MD, Caruso G, CraccoRQ, et al. Non-invasive electrical and magnetic stimulation of thebrain, spinal cord and roots: basic principles and procedures forroutine clinical application. Report of an IFCN committee. Electro-enceph Clin Neurophysiol 1994;91:79–92.
Rossini PM, Pauri F. Neuromagnetic integrated methods tracking humanbrain mechanisms of sensorimotor areas ‘plastic’ reorganisation. BrainRes Rev 2000;33:131–54.
Rothwell JC. Techniques and mechanism of action of transcranialmagnetic stimulation of the human motor cortex. J Neurosci Methods1997;74:113–22.

2674 F. Fratello et al. / Clinical Neurophysiology 117 (2006) 2667–2674
Rothwell JC, Hallett M, Berardelli A, Eisen A, Rossini P, Paulus W.Magnetic stimulation: motor evoked potentials. The InternationalFederation of Clinical Neurophysiology. Electroencephalogr ClinNeurophysiol Suppl 1999;52:97–103.
Siebner HR, Rothwell J. Transcranial magnetic stimulation: new insightsinto representational cortical plasticity. Exp Brain Res 2003;148:1–16.
Stefan K, Kunesch E, Cohen LG, Benecke R, Classen J. Induction ofplasticity in the human motor cortex by paired associative stimulation.Brain. 2000;123:572–84.
Stefan K, Kunesch E, Benecke R, Cohen LG, Classen J. Mechanismsof enhancement of human motor cortex excitability induced byinterventional paired associative stimulation. J Physiol2002;543:699–708.
Stefan K, Wycislo M, Classen J. Modulation of associative human motorcortical plasticity by attention. J Neurophysiol 2004;92:66–72.
Stefan K, Wycislo M, Gentner R, Schramm A, Naumann M, Reiners K,Clossen J. Temporary occlusion of associative motor cortical plasticityby prior dynamic motor training. Cereb Cortex 2006;16:376–85.
Traversa R, Cicinelli P, Pasqualetti P, Filippi M, Rossini PM. Follow-upof interhemispheric differences of motor evoked potentials from the‘affected’ and ‘unaffected’ hemispheres in human stroke. Brain Res1998;803:1–8.
Willem CM, Machielsen, Serge ARB, Rombouts, Barkhof F, Scheltens P,et al. fMRI of visual encoding: reproducibility of activation. HumanBrain Mapping 2000;9:156–164.
Wolf SL, Butler AJ, Campana GI, Parris TA, Struys DM, Weinstein SR,et al. Intra-subject reliability of parameters contributing to mapsgenerated by transcranial magnetic stimulation in able-bodied adults.Clin Neurophysiol 2004;115:1740–7.
Wolters A, Sandbrink F, Schlottmann A, Kunesch E, Stefan K, CohenLG, et al. A temporally asymmetric Hebbian rule governing plasticityin the human motor cortex. J Neurophysiol 2003;89:2339–45.
Wolters A, Schmidt A, Schramm A, Zeller D, Naumann M, Kunesch E,et al. Timing-dependent plasticity in human primary somatosensorycortex. J Physiol 2005;565:1039–52.
Yetkin FZ, McAuliffe TL, Cox R, Haughton VM. Test–retest precision offunctional MR in sensory and motor task activation. Am J Neuro-radiol 1996;17:95–8.
Yoo Seung-schik, Wei Xingchang. Long-term reproducibility analysis offMRI using hand motor task. Intern J Neurosc 2005;115:55–77.
Ziemann U, Ilic TV, Pauli C, Meintzschel F, Ruge D. Learning modifiessubsequent induction of long-term potentiation-like and long-termdepression-like plasticity in human motor cortex. J Neurosci2004;24:1666–72.




















