
Contextual responsiveness: an enduring challenge for educational assessment in Africa
Upload
technology-iraqCategory
view
2download
0
Mice lacking the schizophrenia-associated proteinFEZ1 manifest hyperactivity and enhancedresponsiveness to psychostimulants
Nobutaka Sakae1,2,3, Nobuyuki Yamasaki2,4, Kiyoyuki Kitaichi5, Takaichi Fukuda6,
Mitsunori Yamada7, Hiroo Yoshikawa8, Takato Hiranita5,{, Yoshiki Tatsumi8, Jun-ichi Kira3,
Tsuneyuki Yamamoto5, Tsuyoshi Miyakawa2,4,9 and Keiichi I. Nakayama1,2,�
1Department of Molecular and Cellular Biology, Medical Institute of Bioregulation, Kyushu University, 3-2-2 Maidashi,
Higashi-ku, Fukuoka 812-8582, Japan, 2CREST, Japan Science and Technology Agency (JST), Kawaguchi, Saitama
332-0012, Japan, 3Department of Neurology, Graduate School of Medical Sciences, Neurological Institute, Kyushu
University, Higashi-ku, Fukuoka 812-8582, Japan, 4Genetic Engineering and Functional Genomics Group, Frontier
Technology Center, Kyoto University Graduate School of Medicine, Yoshida-Konoe-cho, Sakyo-ku, Kyoto 606-8501,
Japan, 5Department of Pharmacology, Nagasaki International University, Sasebo, Nagasaki 859-3298, Japan,6Department of Anatomy and Neurobiology, Graduate School of Medical Sciences, Kyushu University, Higashi-ku,
Fukuoka 812-8582, Japan, 7Department of Pathology, Brain Research Institute, Niigata University, 1-757 Asahi-
machi, Niigata, Niigata 951-8585, Japan, 8Division of Neurology and Stroke Care Unit, Department of Internal
Medicine, Hyogo College of Medicine, Nishinomiya, Hyogo 663-8501, Japan and 9Division of Systems Medicine,
Institute for Comprehensive Medical Science, Fujita Health University, 1-98 Dengakugakubo, Kutsukake-cho,
Toyoake 470-1192, Japan
Received April 23, 2008; Revised July 1, 2008; Accepted July 18, 2008
FEZ1 (fasciculation and elongation protein zeta 1), a mammalian ortholog of Caenorhabditis elegans UNC-76,interacts with DISC1 (disrupted in schizophrenia 1), a schizophrenia susceptibility gene product, and poly-morphisms of human FEZ1 have been associated with schizophrenia. We have now investigated the role ofFEZ1 in brain development and the pathogenesis of schizophrenia by generating mice that lack Fez1.Immunofluorescence staining revealed FEZ1 to be located predominantly in g-aminobutyric acid-containinginterneurons. The Fez12/2 mice showed marked hyperactivity in a variety of behavioral tests as well as enhancedbehavioral responses to the psychostimulants MK-801 and methamphetamine. In vivo microdialysis revealedthat the methamphetamine-induced release of dopamine in the nucleus accumbens was exaggerated in themutant mice, suggesting that enhanced mesolimbic dopaminergic transmission contributes to their hyperactivityphenotype. These observations implicate impairment of FEZ1 function in the pathogenesis of schizophrenia.
INTRODUCTION
FEZ1 (fasciculation and elongation protein zeta 1) is a mam-malian ortholog of the Caenorhabditis elegans UNC-76protein (1). Worms harboring mutations in unc-76 manifestsevere abnormalities in movement and in the elongation ofaxons along other axonal surfaces (but not in that along non-
neuronal surfaces) during development, and these abnormal-ities are rescued by introduction of human FEZ1 (1). Thismovement of axons along other axonal surfaces, which isknown as fasciculation, allows axons to associate in specificbundles and plays an important role in the determination ofneural structures. In Drosophila, Unc-76 associates withkinesin heavy chain and its loss of function results in defects
†Psychobiology Section, Medications Discovery Research Branch, Intramural Research Program, National Institution on Drug Abuse, 251 BayviewBoulevard, Suite 200, Baltimore, MD 21224, USA.
�To whom correspondence should be addressed. Tel: þ81 92 642 6815; Fax: þ81 92 642 6819; Email: [email protected]
# The Author 2008. Published by Oxford University Press. All rights reserved.For Permissions, please email: [email protected]
Human Molecular Genetics, 2008, Vol. 17, No. 20 3191–3203doi:10.1093/hmg/ddn215Advance Access published on July 22, 2008
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from

in locomotion and axonal transport reminiscent of thoseinduced by kinesin mutation, suggesting that Unc-76 isrequired for axonal transport (2). In mammals, FEZ1 mRNAis present during all developmental stages of the mousebrain (3,4) and is abundant in the adult rat brain (5). FEZ1is phosphorylated by protein kinase Cz in response to nervegrowth factor, and this phosphorylation of FEZ1 triggersaxonal elongation. Coexpresssion of FEZ1 and a constitutivelyactive mutant of protein kinase Cz in PC12 cells enhancedneurite outgrowth in the absence of nerve growth factor (5).We previously showed that FEZ1 interacts with E4B (alsoknown as UFD2a), a U box-type ubiquitin–protein isopeptideligase, in the yeast two-hybrid system. Furthermore, theassociation between FEZ1 and E4B was enhanced by coex-pression of a constitutively active form of protein kinase Cz,and phosphorylation of FEZ1 by this kinase and its subsequentubiquitylation by E4B resulted in neurite extension in PC12cells (6). FEZ1 also contributes to the polarization of hippo-campal neurons by controlling mitochondrial motility (7).
Two-hybrid analysis has shown that FEZ1 interacts withvarious proteins that function in transcriptional regulation,neuronal cell development, intracellular transport, apoptosisor neurotransmitter release (8). These proteins includenecdin, which is inactivated in Prader–Willi syndrome (9),and disrupted in schizophrenia 1 (DISC1) (10). DISC1 is a sus-ceptibility gene for schizophrenia that was originally identifiedin a large Scottish family with severe mental illness includingschizophrenia (11,12). Subsequent genetic and clinical evi-dence has shown that DISC1 is potentially one of the mostimportant risk genes for schizophrenia and other mental dis-orders (13). DISC1 forms complexes with a large number ofproteins in addition to FEZ1 (10,14–17), suggesting that itmay function as a hub in a multidimensional risk pathwayfor major mental illness and that it plays roles in neurodeve-lopment, cytoskeletal function, cAMP signaling (18) andadult neurogenesis (19). Two of the most studied proteinsthat interact with DISC1 are FEZ1 and nuclear distributiongene E homolog-like 1 (NDEL1), and expression of thegenes for these two proteins has been found to be altered inpostmortem brain specimens from individuals with schizo-phrenia (20). Studies in primates and rodents have shownthat FEZ1 and DISC1 have overlapping spatial and temporalpatterns of expression (4,21,22). Moreover, disruption of theDISC1–FEZ1 interaction inhibits the stimulation of neuriteoutgrowth in PC12 cells by DISC1 (10). Together, theseobservations are consistent with a possible role for FEZ1 inthe pathogenesis of schizophrenia. Whereas associationbetween single nucleotide polymorphisms of FEZ1 andschizophrenia was demonstrated in a Japanese cohort (23),two other studies of FEZ1 polymorphisms and schizophreniafailed to demonstrate an association (24,25).
To explore the physiological role of FEZ1 as well as itsrelation to schizophrenia, we have generated mice with a dis-rupted FEZ1 gene by homologous recombination in embryo-nic stem (ES) cells. Whereas no anatomic abnormality wasevident in the mutant mice, several behavioral tests revealeda phenotype characterized by hyperactivity under novel,social or stressful conditions as well as enhanced responsive-ness to psychostimulants. These defects were associated withan abnormal increase in dopamine release. Our observations
suggest that loss of FEZ1 function may contribute to thepathogenesis of schizophrenia.
RESULTS
Expression of fez1 in mouse brain
In situ hybridization and northern blot analysis have shownthat FEZ1 mRNA is abundant in the adult rat brain (5) aswell as throughout brain development in mouse embryos (4).We examined the expression pattern of FEZ1 protein inmice. Immunoblot analysis revealed that FEZ1 is expressedexclusively in the central nervous system (Fig. 1A). The low-intensity bands apparent in other organs such as the stomachseem to be attributable to non-specific cross-reaction withthe antibodies to FEZ1, given that these bands were alsodetected in FEZ1-deficient mice (Supplementary Material,Fig. S1). FEZ1 was found to be present in a variety of brainregions (Fig. 1B–D). It was detected from the prenatalperiod to adulthood; expression peaked at postnatal day 10and gradually declined thereafter, but was maintained untilat least 5 months of age (Supplementary Material, Fig. S1).
We examined the localization of FEZ1 in the adult brain bymultilabel immunohistofluorescence analysis. FEZ1 immunor-eactivity was detected in a subset of (not all) neurons invarious brain regions including the hippocampus (Fig. 1E),neocortex (Fig. 1F), caudate-putamen (Fig. 1G) and nucleusaccumbens (Fig. 1H). In all these regions, FEZ1-positiveneurons were also labeled with antibodies to glutamic aciddecarboxylase 67 (GAD67), the enzyme responsible for syn-thesis of the inhibitory neurotransmitter g-aminobutyric acid(GABA), indicating that FEZ1 is expressed mainly in GABA-ergic inhibitory neurons. One notable exception to this patternof distribution of FEZ1 was apparent in the dentate gyrus,where granule cells were distinctly labeled with antibodiesto FEZ1 along their axon bundles that form the mossy fiberpathway (Fig. 1C and D). Although dentate granule cells aretypical excitatory neurons with glutamate as their neurotrans-mitter, they also express GAD67 along the mossy fiberpathway (26), resulting in merged signals for colocalizationof GAD67 and FEZ1 therein (Fig. 1C, D). Tyrosinehydroxylase-positive dopaminergic neurons in the midbraindid not manifest FEZ1 immunoreactivity (Fig. 1I).
Generation of fez1-deficient mice
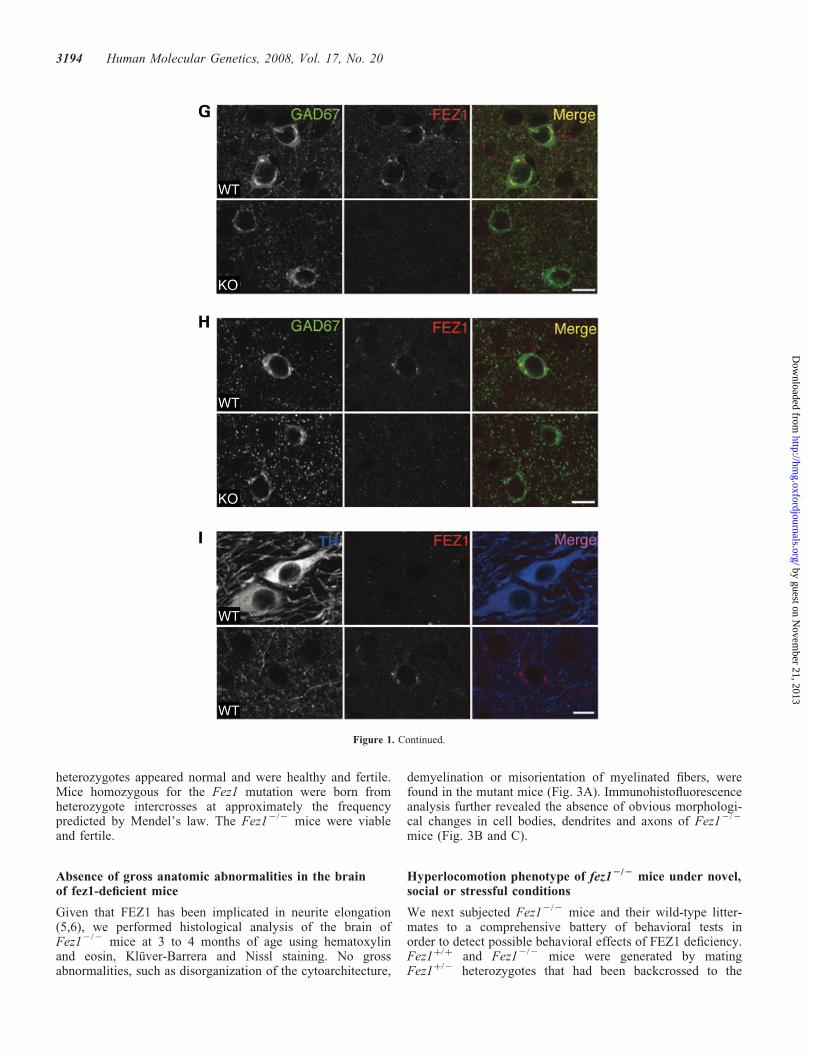
To probe the physiological function of FEZ1, we generatedmice deficient in this protein. Fez1 was disrupted in mouseES cells by replacement of exon 2, which encodes the N-terminus of FEZ1, with a PGK-lox-neo-poly(A) cassette(Fig. 2A). We verified homologous recombination by Southernblot analysis (Fig. 2B) and polymerase chain reaction (PCR)analysis of genomic DNA (Fig. 2C) from the resultingmutant mice. FEZ1 mRNA was not detected in the homozy-gous mutant animals by reverse transcription (RT) and PCRanalysis (Fig. 2D). Immunoblot (Fig. 2E) and immunohisto-fluorescence (Fig. 1C–H) analyses also revealed the absenceof detectable FEZ1 protein in homozygous mutant mice.Although the abundance of FEZ1 in heterozygous micewas about half of that in wild-type animals (Fig. 2E), the
3192 Human Molecular Genetics, 2008, Vol. 17, No. 20
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from
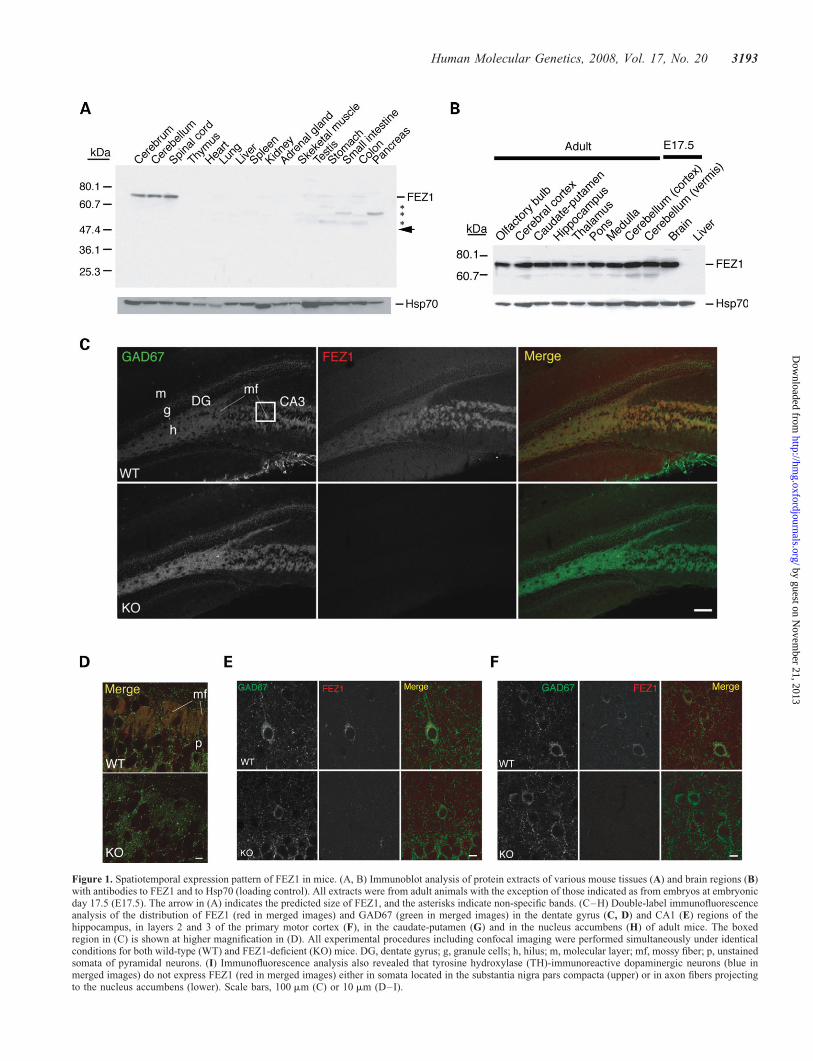
Figure 1. Spatiotemporal expression pattern of FEZ1 in mice. (A, B) Immunoblot analysis of protein extracts of various mouse tissues (A) and brain regions (B)with antibodies to FEZ1 and to Hsp70 (loading control). All extracts were from adult animals with the exception of those indicated as from embryos at embryonicday 17.5 (E17.5). The arrow in (A) indicates the predicted size of FEZ1, and the asterisks indicate non-specific bands. (C–H) Double-label immunofluorescenceanalysis of the distribution of FEZ1 (red in merged images) and GAD67 (green in merged images) in the dentate gyrus (C, D) and CA1 (E) regions of thehippocampus, in layers 2 and 3 of the primary motor cortex (F), in the caudate-putamen (G) and in the nucleus accumbens (H) of adult mice. The boxedregion in (C) is shown at higher magnification in (D). All experimental procedures including confocal imaging were performed simultaneously under identicalconditions for both wild-type (WT) and FEZ1-deficient (KO) mice. DG, dentate gyrus; g, granule cells; h, hilus; m, molecular layer; mf, mossy fiber; p, unstainedsomata of pyramidal neurons. (I) Immunofluorescence analysis also revealed that tyrosine hydroxylase (TH)-immunoreactive dopaminergic neurons (blue inmerged images) do not express FEZ1 (red in merged images) either in somata located in the substantia nigra pars compacta (upper) or in axon fibers projectingto the nucleus accumbens (lower). Scale bars, 100 mm (C) or 10 mm (D–I).
Human Molecular Genetics, 2008, Vol. 17, No. 20 3193
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from
heterozygotes appeared normal and were healthy and fertile.Mice homozygous for the Fez1 mutation were born fromheterozygote intercrosses at approximately the frequencypredicted by Mendel’s law. The Fez12/2 mice were viableand fertile.
Absence of gross anatomic abnormalities in the brainof fez1-deficient mice
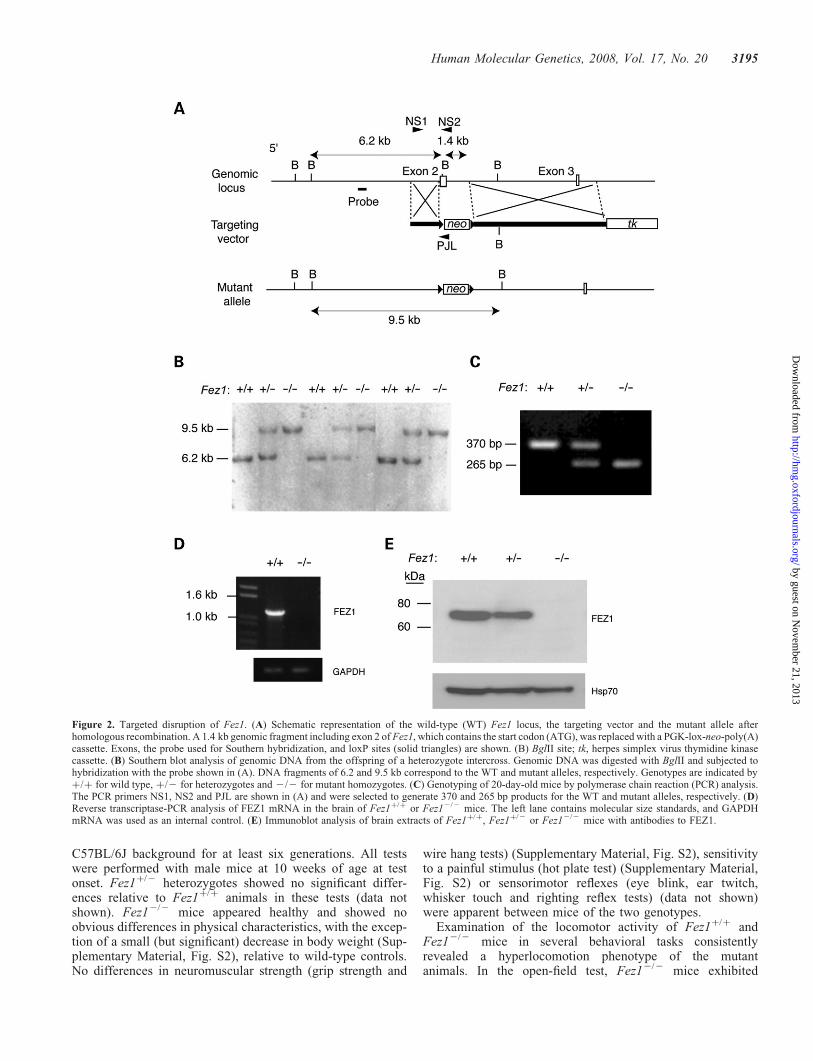
Given that FEZ1 has been implicated in neurite elongation(5,6), we performed histological analysis of the brain ofFez12/2 mice at 3 to 4 months of age using hematoxylinand eosin, Kluver-Barrera and Nissl staining. No grossabnormalities, such as disorganization of the cytoarchitecture,
demyelination or misorientation of myelinated fibers, werefound in the mutant mice (Fig. 3A). Immunohistofluorescenceanalysis further revealed the absence of obvious morphologi-cal changes in cell bodies, dendrites and axons of Fez12/2
mice (Fig. 3B and C).
Hyperlocomotion phenotype of fez12/2 mice under novel,social or stressful conditions
We next subjected Fez12/2 mice and their wild-type litter-mates to a comprehensive battery of behavioral tests inorder to detect possible behavioral effects of FEZ1 deficiency.Fez1þ/þ and Fez12/2 mice were generated by matingFez1þ/2 heterozygotes that had been backcrossed to the
Figure 1. Continued.
3194 Human Molecular Genetics, 2008, Vol. 17, No. 20
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from
C57BL/6J background for at least six generations. All testswere performed with male mice at 10 weeks of age at testonset. Fez1þ/2 heterozygotes showed no significant differ-ences relative to Fez1þ/þ animals in these tests (data notshown). Fez12/2 mice appeared healthy and showed noobvious differences in physical characteristics, with the excep-tion of a small (but significant) decrease in body weight (Sup-plementary Material, Fig. S2), relative to wild-type controls.No differences in neuromuscular strength (grip strength and
wire hang tests) (Supplementary Material, Fig. S2), sensitivityto a painful stimulus (hot plate test) (Supplementary Material,Fig. S2) or sensorimotor reflexes (eye blink, ear twitch,whisker touch and righting reflex tests) (data not shown)were apparent between mice of the two genotypes.
Examination of the locomotor activity of Fez1þ/þ andFez12/2 mice in several behavioral tasks consistentlyrevealed a hyperlocomotion phenotype of the mutantanimals. In the open-field test, Fez12/2 mice exhibited
Figure 2. Targeted disruption of Fez1. (A) Schematic representation of the wild-type (WT) Fez1 locus, the targeting vector and the mutant allele afterhomologous recombination. A 1.4 kb genomic fragment including exon 2 of Fez1, which contains the start codon (ATG), was replaced with a PGK-lox-neo-poly(A)cassette. Exons, the probe used for Southern hybridization, and loxP sites (solid triangles) are shown. (B) BglII site; tk, herpes simplex virus thymidine kinasecassette. (B) Southern blot analysis of genomic DNA from the offspring of a heterozygote intercross. Genomic DNA was digested with BglII and subjected tohybridization with the probe shown in (A). DNA fragments of 6.2 and 9.5 kb correspond to the WT and mutant alleles, respectively. Genotypes are indicated byþ/þ for wild type, þ/2 for heterozygotes and 2/2 for mutant homozygotes. (C) Genotyping of 20-day-old mice by polymerase chain reaction (PCR) analysis.The PCR primers NS1, NS2 and PJL are shown in (A) and were selected to generate 370 and 265 bp products for the WT and mutant alleles, respectively. (D)Reverse transcriptase-PCR analysis of FEZ1 mRNA in the brain of Fez1þ/þ or Fez12/2 mice. The left lane contains molecular size standards, and GAPDHmRNA was used as an internal control. (E) Immunoblot analysis of brain extracts of Fez1þ/þ, Fez1þ/2 or Fez12/2 mice with antibodies to FEZ1.
Human Molecular Genetics, 2008, Vol. 17, No. 20 3195
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from
increased locomotor activity compared with wild-type litter-mates, as revealed by significant increases in distance traveled(Fig. 4A), vertical activity (Fig. 4B) and stereotypic activity(Fig. 4D). The hyperlocomotion phenotype of Fez12/2 micewas also apparent in specialized environments, as revealedby total distance traveled in the elevated plus-maze test(Fig. 5A), in the light–dark transition test (distance traveledin the dark chamber) (Fig. 5B), and in the social interactiontest in a novel environment (Fig. 5C). We also measured loco-motor activity under a stressful condition by the Porsolt forcedswim test. Fez12/2 mice showed a significant decrease inimmobility time compared with wild-type animals (Fig. 5D);this difference reflects a hyperactive tendency rather thanreduced depression-related behavior. In contrast, measures ofanxiety-related behavior, such as the time spent in thecentral part of the open field (Fig. 4C), in the light box ofthe light–dark transition test (Supplementary Material,Fig. S3) and in the open arms in the elevated plus-maze test(Supplementary Material, Fig. S3) as well as a measure ofsocial behavior in the social interaction test in a novel environ-ment (Supplementary Material, Fig. S3) did not differ sig-nificantly between Fez1þ/þ and Fez12/2 mice. Theseobservations indicated that FEZ1 deficiency did not affectemotional behavior.
Effects of fez1 deficiency on learning and memory
To assess whether the loss of FEZ1 was associated with cogni-tive abnormalities, we performed various memory and learningtests with Fez12/2 mice. In a contextual and cued fear con-ditioning test, Fez12/2 mice showed a significantly reducedfrequency of freezing during conditioning compared with wild-type controls (Fig. 6A). One day after conditioning, a contextualtest also revealed a significant decrease in freezing behavior forthe mutant mice (Fig. 6B). It was possible, however, that thedifference in performance in this test between mice of the twogenotypes was attributable to the hyperactive phenotype ofFez12/2 mice. To examine the possible effect of FEZ1deficiency on spatial working memory, we subjected mice tothe Barnes maze test and eight-arm radial maze test. Neitherof these tests revealed a significant difference in performancebetween genotypes (Supplementary Material, Fig. S4),suggesting that Fez12/2 mice are not cognitively impaired.Moreover, in the T-maze test, Fez12/2 mice showed a signifi-cantly higher percentage of correct choices than did their wild-type littermates (Fig. 6C). These data were thus also indicativeof the lack of a cognitive abnormality in Fez12/2 mice,although they were also suggestive of enhanced food rewardand reinforcement in the mutant animals.
Figure 3. Absence of gross structural abnormalities in the brain of FEZ1-deficient mice. (A) Coronal sections through the striatum and hippocampus of wild-type(WT) or Fez12/2 (KO) mice at 3 months of age were stained by the Kluver-Barrera method. No marked differences in cytoarchitecture revealed by staining ofcell bodies (dark purple) or fiber tracts (light blue) were apparent between the two genotypes. (B, C) Confocal immunofluorescence analysis of the morphology ofcells in the hippocampus of WT or Fez12/2 mice at 3 months of age. Immunostaining was performed with antibodies to calbindin (B) or to parvalbumin (C). cc,corpus callosum; ic, internal capsule; fmb, fimbria hippocampi; g, granule cells of the dentate gyrus; m, molecular layer; mf, mossy fiber. Scale bars, 1 mm (A) or100 mm (B, C).
3196 Human Molecular Genetics, 2008, Vol. 17, No. 20
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from
Lack of sensorimotor gating defect in fez12/2 mice
Fez12/2 mice were tested for sensorimotor gating deficits inthe prepulse inhibition test. The amplitudes of the startleresponse were similar in wild-type and Fez12/2 mice beforetesting (Supplementary Material, Fig. S5). Furthermore, thepercentage prepulse inhibition did not differ significantlybetween the two genotypes.
Enhanced responsiveness of fez12/2 mice topsychostimulants
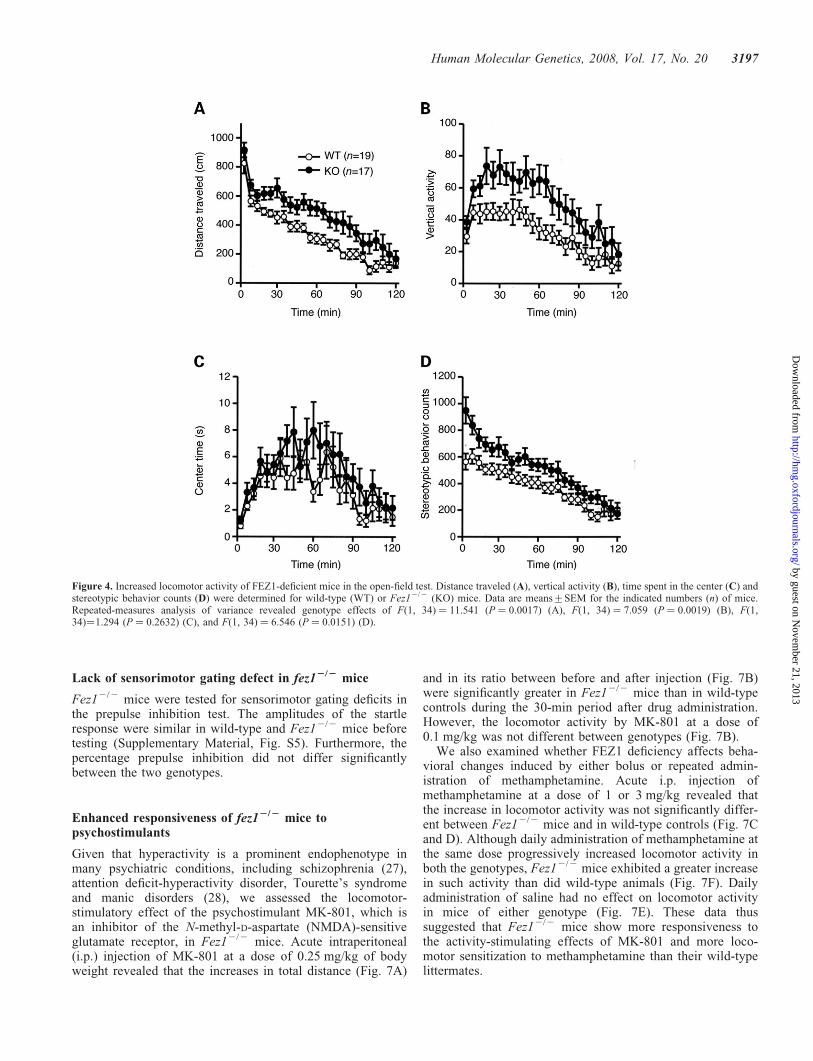
Given that hyperactivity is a prominent endophenotype inmany psychiatric conditions, including schizophrenia (27),attention deficit-hyperactivity disorder, Tourette’s syndromeand manic disorders (28), we assessed the locomotor-stimulatory effect of the psychostimulant MK-801, which isan inhibitor of the N-methyl-D-aspartate (NMDA)-sensitiveglutamate receptor, in Fez12/2 mice. Acute intraperitoneal(i.p.) injection of MK-801 at a dose of 0.25 mg/kg of bodyweight revealed that the increases in total distance (Fig. 7A)
and in its ratio between before and after injection (Fig. 7B)were significantly greater in Fez12/2 mice than in wild-typecontrols during the 30-min period after drug administration.However, the locomotor activity by MK-801 at a dose of0.1 mg/kg was not different between genotypes (Fig. 7B).
We also examined whether FEZ1 deficiency affects beha-vioral changes induced by either bolus or repeated admin-istration of methamphetamine. Acute i.p. injection ofmethamphetamine at a dose of 1 or 3 mg/kg revealed thatthe increase in locomotor activity was not significantly differ-ent between Fez12/2 mice and in wild-type controls (Fig. 7Cand D). Although daily administration of methamphetamine atthe same dose progressively increased locomotor activity inboth the genotypes, Fez12/2 mice exhibited a greater increasein such activity than did wild-type animals (Fig. 7F). Dailyadministration of saline had no effect on locomotor activityin mice of either genotype (Fig. 7E). These data thussuggested that Fez12/2 mice show more responsiveness tothe activity-stimulating effects of MK-801 and more loco-motor sensitization to methamphetamine than their wild-typelittermates.
Figure 4. Increased locomotor activity of FEZ1-deficient mice in the open-field test. Distance traveled (A), vertical activity (B), time spent in the center (C) andstereotypic behavior counts (D) were determined for wild-type (WT) or Fez12/2 (KO) mice. Data are means+SEM for the indicated numbers (n) of mice.Repeated-measures analysis of variance revealed genotype effects of F(1, 34) ¼ 11.541 (P ¼ 0.0017) (A), F(1, 34) ¼ 7.059 (P ¼ 0.0019) (B), F(1,34)¼1.294 (P ¼ 0.2632) (C), and F(1, 34) ¼ 6.546 (P ¼ 0.0151) (D).
Human Molecular Genetics, 2008, Vol. 17, No. 20 3197
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from
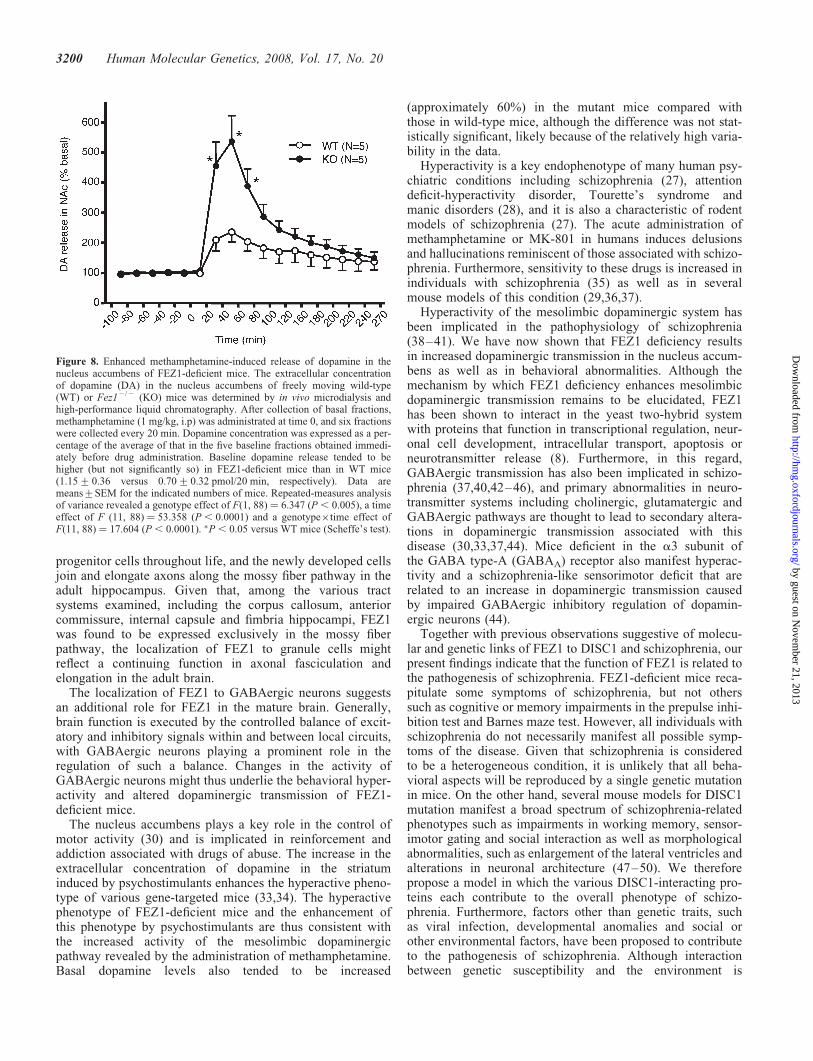
Increased methamphetamine-induced release of dopaminein fez12/2 mice
Locomotor hyperactivity is typically associated with increaseddopaminergic transmission in major motor areas of the brainsuch as the striatum (29). The caudate-putamen and nucleusaccumbens play an important role in the control of locomotoractivity (30). Given that Fez12/2 mice manifest both hyperac-tivity and enhanced responsiveness to the locomotor-stimulatory effects of methamphetamine and MK-801, weperformed in vivo microdialysis to examine whether FEZ1deficiency affects the extracellular level of dopamine in thenucleus accumbens after methamphetamine administration.Samples were collected from the nucleus accumbens offreely moving mice, and the extracellular concentration ofdopamine was determined by high-performance liquid chrom-atography both before and after the injection of methamphet-amine (1 mg/kg, i.p.). The basal levels of dopamine before
drug administration were similar in Fez1þ/þ and Fez12/2
mice, although those in Fez12/2 mice tended to be higherthan those in Fez1þ/þ mice (the difference was not statisti-cally significant possibly because of the substantial level ofvariability in the data). Whereas methamphetamine adminis-tration increased the extracellular level of dopamine in miceof both genotypes, the drug-induced increase in dopamineconcentration was significantly greater in Fez12/2 micethan in wild-type controls (Fig. 8). These data thus suggestthat FEZ1 deficiency results in an increase inmethamphetamine-induced dopaminergic neurotransmissionin the nucleus accumbens.
DISCUSSION
FEZ1 has been implicated in processes that underlie neuronalmorphology, including axon fasciculation, axonal transport
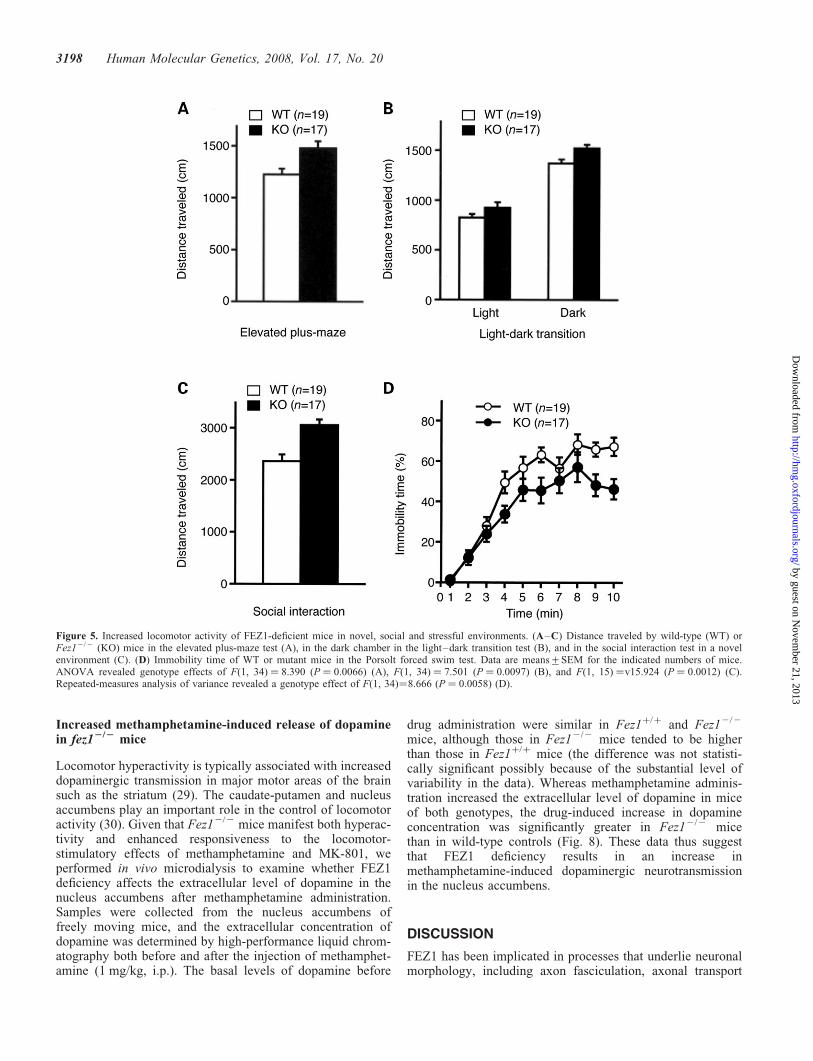
Figure 5. Increased locomotor activity of FEZ1-deficient mice in novel, social and stressful environments. (A–C) Distance traveled by wild-type (WT) orFez12/2 (KO) mice in the elevated plus-maze test (A), in the dark chamber in the light–dark transition test (B), and in the social interaction test in a novelenvironment (C). (D) Immobility time of WT or mutant mice in the Porsolt forced swim test. Data are means+SEM for the indicated numbers of mice.ANOVA revealed genotype effects of F(1, 34) ¼ 8.390 (P ¼ 0.0066) (A), F(1, 34) ¼ 7.501 (P ¼ 0.0097) (B), and F(1, 15) ¼v15.924 (P ¼ 0.0012) (C).Repeated-measures analysis of variance revealed a genotype effect of F(1, 34)¼8.666 (P ¼ 0.0058) (D).
3198 Human Molecular Genetics, 2008, Vol. 17, No. 20
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from
and neurite extension, as well as in aspects of viral infectionsuch as postentry block of retroviral infection (31) and associ-ation with the agnoprotein of the human polyomavirus JCvirus (32). FEZ1 may also play a role both in control of tran-scription, as evidenced by its association with transcriptionalfactors (SAP30L, DRAP1, BAF60a) (8), as well as in mito-chondrial motility (7). Given the predicted function of FEZ1in the determination of neuronal morphology, our findingthat mice deficient in this protein do not exhibit abnormalbrain architecture was unexpected. The FEZ1-deficient micedid, however, manifest behavior abnormalities including ahyperlocomotion phenotype and enhanced responsiveness topsychostimulants. These abnormalities are likely attributableto changes in brain-regulatory systems, particularly in thealtered control of dopamine release in the mesolimbicpathway.
Our immunohistofluorescence analysis revealed that FEZ1is expressed in GABAergic inhibitory neurons of variousbrain regions as well as in granule cells of the dentate gyrusin adult mice. Dentate granule cells are generated from
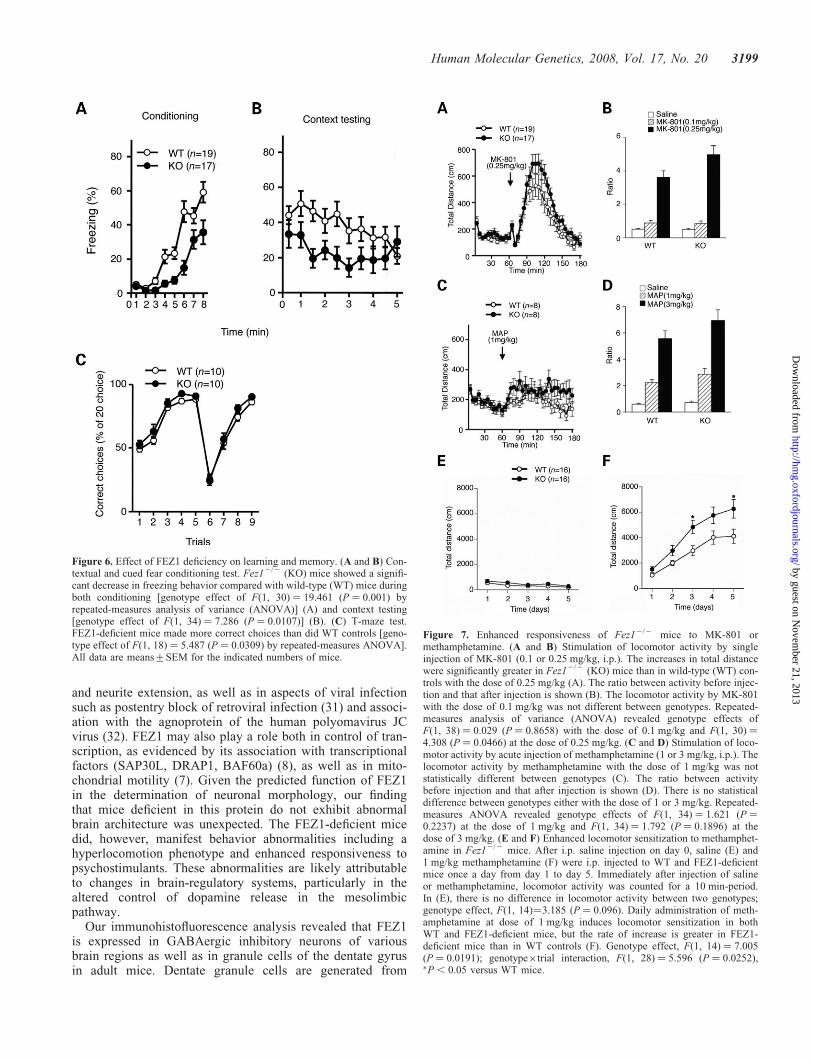
Figure 6. Effect of FEZ1 deficiency on learning and memory. (A and B) Con-textual and cued fear conditioning test. Fez12/2 (KO) mice showed a signifi-cant decrease in freezing behavior compared with wild-type (WT) mice duringboth conditioning [genotype effect of F(1, 30) ¼ 19.461 (P ¼ 0.001) byrepeated-measures analysis of variance (ANOVA)] (A) and context testing[genotype effect of F(1, 34) ¼ 7.286 (P ¼ 0.0107)] (B). (C) T-maze test.FEZ1-deficient mice made more correct choices than did WT controls [geno-type effect of F(1, 18) ¼ 5.487 (P ¼ 0.0309) by repeated-measures ANOVA].All data are means+SEM for the indicated numbers of mice.
Figure 7. Enhanced responsiveness of Fez12/2 mice to MK-801 ormethamphetamine. (A and B) Stimulation of locomotor activity by singleinjection of MK-801 (0.1 or 0.25 mg/kg, i.p.). The increases in total distancewere significantly greater in Fez12/2 (KO) mice than in wild-type (WT) con-trols with the dose of 0.25 mg/kg (A). The ratio between activity before injec-tion and that after injection is shown (B). The locomotor activity by MK-801with the dose of 0.1 mg/kg was not different between genotypes. Repeated-measures analysis of variance (ANOVA) revealed genotype effects ofF(1, 38) ¼ 0.029 (P ¼ 0.8658) with the dose of 0.1 mg/kg and F(1, 30) ¼4.308 (P ¼ 0.0466) at the dose of 0.25 mg/kg. (C and D) Stimulation of loco-motor activity by acute injection of methamphetamine (1 or 3 mg/kg, i.p.). Thelocomotor activity by methamphetamine with the dose of 1 mg/kg was notstatistically different between genotypes (C). The ratio between activitybefore injection and that after injection is shown (D). There is no statisticaldifference between genotypes either with the dose of 1 or 3 mg/kg. Repeated-measures ANOVA revealed genotype effects of F(1, 34) ¼ 1.621 (P ¼0.2237) at the dose of 1 mg/kg and F(1, 34) ¼ 1.792 (P ¼ 0.1896) at thedose of 3 mg/kg. (E and F) Enhanced locomotor sensitization to methamphet-amine in Fez12/2 mice. After i.p. saline injection on day 0, saline (E) and1 mg/kg methamphetamine (F) were i.p. injected to WT and FEZ1-deficientmice once a day from day 1 to day 5. Immediately after injection of salineor methamphetamine, locomotor activity was counted for a 10 min-period.In (E), there is no difference in locomotor activity between two genotypes;genotype effect, F(1, 14)¼3.185 (P ¼ 0.096). Daily administration of meth-amphetamine at dose of 1 mg/kg induces locomotor sensitization in bothWT and FEZ1-deficient mice, but the rate of increase is greater in FEZ1-deficient mice than in WT controls (F). Genotype effect, F(1, 14) ¼ 7.005(P ¼ 0.0191); genotype�trial interaction, F(1, 28) ¼ 5.596 (P ¼ 0.0252),�P , 0.05 versus WT mice.
Human Molecular Genetics, 2008, Vol. 17, No. 20 3199
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from
progenitor cells throughout life, and the newly developed cellsjoin and elongate axons along the mossy fiber pathway in theadult hippocampus. Given that, among the various tractsystems examined, including the corpus callosum, anteriorcommissure, internal capsule and fimbria hippocampi, FEZ1was found to be expressed exclusively in the mossy fiberpathway, the localization of FEZ1 to granule cells mightreflect a continuing function in axonal fasciculation andelongation in the adult brain.
The localization of FEZ1 to GABAergic neurons suggestsan additional role for FEZ1 in the mature brain. Generally,brain function is executed by the controlled balance of excit-atory and inhibitory signals within and between local circuits,with GABAergic neurons playing a prominent role in theregulation of such a balance. Changes in the activity ofGABAergic neurons might thus underlie the behavioral hyper-activity and altered dopaminergic transmission of FEZ1-deficient mice.
The nucleus accumbens plays a key role in the control ofmotor activity (30) and is implicated in reinforcement andaddiction associated with drugs of abuse. The increase in theextracellular concentration of dopamine in the striatuminduced by psychostimulants enhances the hyperactive pheno-type of various gene-targeted mice (33,34). The hyperactivephenotype of FEZ1-deficient mice and the enhancement ofthis phenotype by psychostimulants are thus consistent withthe increased activity of the mesolimbic dopaminergicpathway revealed by the administration of methamphetamine.Basal dopamine levels also tended to be increased
(approximately 60%) in the mutant mice compared withthose in wild-type mice, although the difference was not stat-istically significant, likely because of the relatively high varia-bility in the data.
Hyperactivity is a key endophenotype of many human psy-chiatric conditions including schizophrenia (27), attentiondeficit-hyperactivity disorder, Tourette’s syndrome andmanic disorders (28), and it is also a characteristic of rodentmodels of schizophrenia (27). The acute administration ofmethamphetamine or MK-801 in humans induces delusionsand hallucinations reminiscent of those associated with schizo-phrenia. Furthermore, sensitivity to these drugs is increased inindividuals with schizophrenia (35) as well as in severalmouse models of this condition (29,36,37).
Hyperactivity of the mesolimbic dopaminergic system hasbeen implicated in the pathophysiology of schizophrenia(38–41). We have now shown that FEZ1 deficiency resultsin increased dopaminergic transmission in the nucleus accum-bens as well as in behavioral abnormalities. Although themechanism by which FEZ1 deficiency enhances mesolimbicdopaminergic transmission remains to be elucidated, FEZ1has been shown to interact in the yeast two-hybrid systemwith proteins that function in transcriptional regulation, neur-onal cell development, intracellular transport, apoptosis orneurotransmitter release (8). Furthermore, in this regard,GABAergic transmission has also been implicated in schizo-phrenia (37,40,42–46), and primary abnormalities in neuro-transmitter systems including cholinergic, glutamatergic andGABAergic pathways are thought to lead to secondary altera-tions in dopaminergic transmission associated with thisdisease (30,33,37,44). Mice deficient in the a3 subunit ofthe GABA type-A (GABAA) receptor also manifest hyperac-tivity and a schizophrenia-like sensorimotor deficit that arerelated to an increase in dopaminergic transmission causedby impaired GABAergic inhibitory regulation of dopamin-ergic neurons (44).
Together with previous observations suggestive of molecu-lar and genetic links of FEZ1 to DISC1 and schizophrenia, ourpresent findings indicate that the function of FEZ1 is related tothe pathogenesis of schizophrenia. FEZ1-deficient mice reca-pitulate some symptoms of schizophrenia, but not otherssuch as cognitive or memory impairments in the prepulse inhi-bition test and Barnes maze test. However, all individuals withschizophrenia do not necessarily manifest all possible symp-toms of the disease. Given that schizophrenia is consideredto be a heterogeneous condition, it is unlikely that all beha-vioral aspects will be reproduced by a single genetic mutationin mice. On the other hand, several mouse models for DISC1mutation manifest a broad spectrum of schizophrenia-relatedphenotypes such as impairments in working memory, sensor-imotor gating and social interaction as well as morphologicalabnormalities, such as enlargement of the lateral ventricles andalterations in neuronal architecture (47–50). We thereforepropose a model in which the various DISC1-interacting pro-teins each contribute to the overall phenotype of schizo-phrenia. Furthermore, factors other than genetic traits, suchas viral infection, developmental anomalies and social orother environmental factors, have been proposed to contributeto the pathogenesis of schizophrenia. Although interactionbetween genetic susceptibility and the environment is
Figure 8. Enhanced methamphetamine-induced release of dopamine in thenucleus accumbens of FEZ1-deficient mice. The extracellular concentrationof dopamine (DA) in the nucleus accumbens of freely moving wild-type(WT) or Fez12/2 (KO) mice was determined by in vivo microdialysis andhigh-performance liquid chromatography. After collection of basal fractions,methamphetamine (1 mg/kg, i.p) was administrated at time 0, and six fractionswere collected every 20 min. Dopamine concentration was expressed as a per-centage of the average of that in the five baseline fractions obtained immedi-ately before drug administration. Baseline dopamine release tended to behigher (but not significantly so) in FEZ1-deficient mice than in WT mice(1.15+0.36 versus 0.70+0.32 pmol/20 min, respectively). Data aremeans+SEM for the indicated numbers of mice. Repeated-measures analysisof variance revealed a genotype effect of F(1, 88) ¼ 6.347 (P , 0.005), a timeeffect of F (11, 88) ¼ 53.358 (P , 0.0001) and a genotype�time effect ofF(11, 88) ¼ 17.604 (P , 0.0001). �P , 0.05 versus WT mice (Scheffe’s test).
3200 Human Molecular Genetics, 2008, Vol. 17, No. 20
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from

thought to play a prominent role in schizophrenia, the preciseetiology of this disorder remains unclear. Our results suggestthat characterization of the function of FEZ1 may provideinsight into the pathophysiology of schizophrenia, and thatFez12/2 mice may thus prove to be a useful animal modelfor studies of human schizophrenia as well as for the develop-ment of new drugs for this disorder.
MATERIALS AND METHODS
Generation of fez1-deficient mice
The Fez1 locus was amplified by PCR from the genome ofE14 (embryonic day 14) mouse ES cells with the use ofLA-Taq polymerase (TaKaRa). The targeting vector was con-structed by the replacement of a 1.4-kb fragment of genomicDNA containing exon 2 of Fez1 with a PGK-lox-neo-poly(A)cassette. The vector thus contained 1.6- and 6.5-kb regions ofhomology located at 50 and 30, respectively, relative to the neo-mycin resistance gene (neo). A PGK-tk-poly(A) cassette wasligated at the 30 end of the targeting construct. The mainten-ance, transfection and selection of ES cells were performedas described previously (51). The recombination event wasconfirmed by Southern blot analysis with a 0.4-kb fragmentof genomic DNA that flanked the 50 homology region(Fig. 2A). The expected sizes of hybridizing fragments afterdigestion of genomic DNA with BglII were 6.2 and 9.5 kbfor the wild-type and mutant Fez1 alleles, respectively.Mutant ES cells were microinjected into C57BL/6 blastocysts,and the resulting male chimeras were mated with femaleC57BL/6 mice. The germline transmission of the mutantallele was confirmed by Southern blot analysis. Heterozygousoffsprings were intercrossed to produce homozygous mutantanimals. For genotyping of mice, DNA was extracted fromthe tail and analyzed by PCR with the primers NS1(50-TTTCCAAGGGACATAGAGTAA-30), NS2 (50-AAAGCAGACATTGAGCTTCTC-30) and PJL (50-TGCTAAAGCGCATGCTCCAGACTG-30). Procedures related to the generationof the knockout mice were approved by the animal ethics com-mittee of Kyushu University.
Immunoblot analysis
Immunoblot analysis was performed with antibodies to Hsp70(1 mg/ml) (clone7, Transduction Laboratories, Lexington, KY)and to FEZ1 (1 mg/ml). The rabbit polyclonal antibodies tohuman FEZ1 were generated by standard procedures using arecombinant full-length protein expressed in and purifiedfrom Sf21 cells as described previously (6). Mouse tissueswere homogenized in homogenization buffer [50 mM Tris–HCl (pH 7.6), 150 mM NaCl, 0.5% Triton X-100, aprotinin(10 mg/ml), leupeptin (10 mg/ml), 10 mM iodoacetamide,1 mM phenylmethylsulfonyl fluoride, 0.4 mM ethylenediammi-netetraacetic acid, 10 mM NaF, 10 mM sodium pyrophosphate].Proteins were separated by sodium dodecylsulfate–polyacryl-amide gel electrophoresis and transferred to a Hybond P mem-brane (Amersham Biosciences), which was then exposed for1 h at room temperature to Tris-buffered saline containing 5%dried skim milk before consecutive incubations for 1 h withprimary antibodies and for 30 min with horseradish peroxidase-
conjugated secondary antibodies. Immune complexes weredetected using Signal West Pico chemiluminescence reagents(Pierce, Rockford, IL).
Histological analysis
Male mice (12–15 weeks of age) were anesthetized with pen-tobarbital (100 mg/kg, i.p.) and perfused through the ascend-ing aorta first briefly with phosphate-buffered saline (PBS)and then with 50 ml of 4% paraformaldehyde in 0.1 M phos-phate buffer (pH 7.4). For immunostaining, the brain wasexcised and cut serially into 40 mm coronal or saggital sec-tions using a vibrating microtome (VT1000, Leica, Heidel-berg, Germany). For hematoxylin and eosin, Nissl orKluver-Barrera staining, portions of the brain were embeddedin paraffin and cut coronally into 5 mm serial sections. Aftercryoprotection with 30% (w/v) sucrose in 0.1 M phosphatebuffer (pH 7.4), sections were rapidly subjected to freezingin liquid nitrogen and thawing before processing for multilabelimmunostaining as described previously (52), but with slightmodifications. In brief, sections were incubated for 7 days atroom temperature with the following combinations ofprimary antibodies in PBS containing 1% bovine serumalbumin and 0.3% Triton X-100: (i) rabbit polyclonal anti-bodies to FEZ1 (1:1000 dilution) (6), mouse monoclonal anti-bodies to GAD67 (1:5000) (Chemicon, Temecula, CA) andgoat polyclonal antibodies to tyrosine hydroxylase (1:5000)(Chemicon); or (ii) rabbit polyclonal antibodies to parvalbu-min (1:5000) (kindly provided by C.W. Heizmann) andmouse monoclonal antibodies to calbindin (1:10 000)(SWANT). After washing the sections thoroughly in PBS,they were incubated overnight with biotinylated donkey anti-bodies to goat immunoglobulin G (Invitrogen, Carlsbad, CA)and then overnight with a mixture of Alexa 488-conjugateddonkey antibodies to mouse immunoglobulin G, rhodamine-conjugated donkey antibodies to rabbit immunoglobulin G(Invitrogen) and streptavidin-conjugated Cy5 (Invitrogen).The sections were mounted in Vectashield (Vector) andexamined with a laser-scanning confocal microscope (C1plus, Nikon). Sections from Fez12/2 mice were used as con-trols for FEZ1 immunostaining. Specimens from Fez1þ/þ andFez12/2 mice were thus prepared and processed at each stepsimultaneously and were examined under identical microscopesettings. The images are presented as digital figures withminimal changes in brightness or contrast.
Behavioral analysis
Fez1þ/þ and Fez12/2 mice were generated by matingFez1þ/2 heterozygotes that had been backcrossed to theC57BL/6J background for at least six generations. All beha-vioral tests were carried out with male mice that were10-weeks-old at the start of the testing. Female mice wereexcluded because their behavior is influenced by the menstrualcycle. Mice were housed four per cage in a room with a12 h-light and 12 h-dark cycle (lights on at 0700 h), and theyhad free access to food and water. Behavioral testing wasperformed between 0900 and 1800 h in a manner similar tothat described previously (29,34,53). All behavioral testingprocedures were approved by the Animal Care and Use
Human Molecular Genetics, 2008, Vol. 17, No. 20 3201
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from

Committee of Kyoto University Graduate School of Medicine.Detailed descriptions of the behavioral tests are available inSupplementary Methods.
In vivo microdialysis
In vivo microdialysis measurements of extracellular dopaminewere performed in freely moving mice as described previously(54) (see Supplementary methods for details).
Statistical analysis
Data are presented as means+SEM and were analyzed by thetwo-tailed Student’s t-test, Scheffe’s test, analysis of variance(ANOVA) or repeated-measures ANOVA using StatViewsoftware (SAS Institute, Cary, NC).
SUPPLEMENTARY MATERIAL
Supplementary Material is available at HMG Online.
ACKNOWLEDGEMENTS
We thank Y. Yamada for generating FEZ1 knock-out mice;K. Nakanishi and H. Ougino for technical assistance withbehavioral analysis; and A. Ohta for help in preparing themanuscript. This work was supported in part by Grants-in-Aidfrom the Ministry of Education, Culture, Sports, Science andTechnology of Japan; by a research grant from the HumanFrontier Science Program; by Grants-in-Aid for ScientificResearch from the Japan Society for the Promotion ofScience; by the BIRD program of the Japan Science and Tech-nology Agency; and by the National Institute of BiomedicalInnovation of Japan.
Conflict of Interest statement. The authors declare nocompeting financial interests.
FUNDING
This work was supported by Takeda Science Foundation.
REFERENCES
1. Bloom, L. and Horvitz, H.R. (1997) The Caenorhabditis elegans geneunc-76 and its human homologs define a new gene family involved inaxonal outgrowth and fasciculation. Proc. Natl Acad. Sci. USA, 94,3414–3419.
2. Gindhart, J.G., Chen, J., Faulkner, M., Gandhi, R., Doerner, K.,Wisniewski, T. and Nandlestadt, A. (2003) The kinesin-associated proteinUNC-76 is required for axonal transport in the Drosophila nervoussystem. Mol. Biol. Cell., 14, 3356–3365.
3. Fujita, T., Ikuta, J., Hamada, J., Okajima, T., Tatematsu, K., Tanizawa, K.and Kuroda, S. (2004) Identification of a tissue-non-specific homologue ofaxonal fasciculation and elongation protein z-1. Biochem. Biophys. Res.Commun., 313, 738–744.
4. Honda, A., Miyoshi, K., Baba, K., Taniguchi, M., Koyama, Y., Kuroda,S., Katayama, T. and Tohyama, M. (2004) Expression of fasciculation andelongation protein z-1 (FEZ1) in the developing rat brain. Brain Res. Mol.Brain Res., 122, 89–92.
5. Kuroda, S., Nakagawa, N., Tokunaga, C., Tatematsu, K. and Tanizawa, K.(1999) Mammalian homologue of the Caenorhabditis elegans UNC-76
protein involved in axonal outgrowth is a protein kinase C z-interactingprotein. J. Cell Biol., 144, 403–411.
6. Okumura, F., Hatakeyama, S., Matsumoto, M., Kamura, T. andNakayama, K.I. (2004) Functional regulation of FEZ1 by the U-box-typeubiquitin ligase E4B contributes to neuritogenesis. J. Biol. Chem., 279,53533–53543.
7. Ikuta, J., Maturana, A., Fujita, T., Okajima, T., Tatematsu, K., Tanizawa,K. and Kuroda, S. (2007) Fasciculation and elongation protein z-1 (FEZ1)participates in the polarization of hippocampal neuron by controlling themitochondrial motility. Biochem. Biophys. Res. Commun., 353, 127–132.
8. Assmann, E.M., Alborghetti, M.R., Camargo, M.E. and Kobarg, J. (2006)FEZ1 dimerization and interaction with transcription regulatory proteinsinvolves its coiled-coil region. J. Biol. Chem., 281, 9869–9881.
9. Lee, S., Walker, C.L., Karten, B., Kuny, S.L., Tennese, A.A., O’Neill,M.A. and Wevrick, R. (2005) Essential role for the Prader-Willi syndromeprotein necdin in axonal outgrowth. Hum. Mol. Genet., 14, 627–637.
10. Miyoshi, K., Honda, A., Baba, K., Taniguchi, M., Oono, K., Fujita, T.,Kuroda, S., Katayama, T. and Tohyama, M. (2003) Disrupted-in-schizophrenia 1, a candidate gene for schizophrenia, participates in neuriteoutgrowth. Mol. Psychiatry, 8, 685–694.
11. Millar, J.K., Wilson-Annan, J.C., Anderson, S., Christie, S., Taylor, M.S.,Semple, C.A., Devon, R.S., Clair, D.M., Muir, W.J., Blackwood, D.H.et al. (2000) Disruption of two novel genes by a translocationco-segregating with schizophrenia. Hum. Mol. Genet., 9, 1415–1423.
12. Blackwood, D.H., Fordyce, A., Walker, M.T., St Clair, D.M., Porteous,D.J. and Muir, W.J. (2001) Schizophrenia and affective disorders—cosegregation with a translocation at chromosome 1q42 that directlydisrupts brain-expressed genes: clinical and P300 findings in a family.Am. J. Hum. Genet., 69, 428–433.
13. Wang, Q., Jaaro-Peled, H., Sawa, A. and Brandon, N.J. (2008) How hasDISC1 enabled drug discovery? Mol. Cell. Neurosci., 37, 187–195.
14. Morris, J.A., Kandpal, G., Ma, L. and Austin, C.P. (2003) DISC1(Disrupted-In-Schizophrenia 1) is a centrosome-associated protein thatinteracts with MAP1A, MIPT3, ATF4/5 and NUDEL: regulation and lossof interaction with mutation. Hum. Mol. Genet., 12, 1591–1608.
15. Ozeki, Y., Tomoda, T., Kleiderlein, J., Kamiya, A., Bord, L., Fujii, K.,Okawa, M., Yamada, N., Hatten, M.E., Snyder, S.H. et al. (2003)Disrupted-in-Schizophrenia-1 (DISC-1): mutant truncation preventsbinding to NudE-like (NUDEL) and inhibits neurite outgrowth. Proc. Natl
Acad. Sci. USA, 100, 289–294.
16. Brandon, N.J., Handford, E.J., Schurov, I., Rain, J.C., Pelling, M.,Duran-Jimeniz, B., Camargo, L.M., Oliver, K.R., Beher, D., Shearman,M.S. et al. (2004) Disrupted in Schizophrenia 1 and Nudel form aneurodevelopmentally regulated protein complex: implications forschizophrenia and other major neurological disorders. Mol. Cell.
Neurosci., 25, 42–55.17. Millar, J.K., Pickard, B.S., Mackie, S., James, R., Christie, S., Buchanan,
S.R., Malloy, M.P., Chubb, J.E., Huston, E., Baillie, G.S. et al. (2005)DISC1 and PDE4B are interacting genetic factors in schizophrenia thatregulate cAMP signaling. Science, 310, 1187–1191.
18. Chubb, J.E., Bradshaw, N.J., Soares, D.C., Porteous, D.J. and Millar, J.K.(2008) The DISC locus in psychiatric illness. Mol. Psychiatry, 13, 36–64.
19. Duan, X., Chang, J.H., Ge, S., Faulkner, R.L., Kim, J.Y., Kitabatake, Y.,Liu, X.B., Yang, C.H., Jordan, J.D., Ma, D.K. et al. (2007)Disrupted-In-Schizophrenia 1 regulates integration of newly generatedneurons in the adult brain. Cell, 130, 1146–1158.
20. Lipska, B.K., Peters, T., Hyde, T.M., Halim, N., Horowitz, C., Mitkus, S.,Weickert, C.S., Matsumoto, M., Sawa, A., Straub, R.E. et al. (2006)Expression of DISC1 binding partners is reduced in schizophrenia andassociated with DISC1 SNPs. Hum. Mol. Genet., 15, 1245–1258.
21. Austin, C.P., Ky, B., Ma, L., Morris, J.A. and Shughrue, P.J. (2004)Expression of Disrupted-In-Schizophrenia-1, a schizophrenia-associatedgene, is prominent in the mouse hippocampus throughout braindevelopment. Neuroscience, 124, 3–10.
22. Schurov, I.L., Handford, E.J., Brandon, N.J. and Whiting, P.J. (2004)Expression of disrupted in schizophrenia 1 (DISC1) protein in the adultand developing mouse brain indicates its role in neurodevelopment. Mol.
Psychiatry, 9, 1100–1110.
23. Yamada, K., Nakamura, K., Minabe, Y., Iwayama-Shigeno, Y., Takao, H.,Toyota, T., Hattori, E., Takei, N., Sekine, Y., Suzuki, K. et al. (2004)Association analysis of FEZ1 variants with schizophrenia in Japanesecohorts. Biol. Psychiatry, 56, 683–690.
3202 Human Molecular Genetics, 2008, Vol. 17, No. 20
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from

24. Koga, M., Ishiguro, H., Horiuchi, Y., Albalushi, T., Inada, T., Iwata, N.,Ozaki, N., Ujike, H., Muratake, T., Someya, T. et al. (2007) Failure toconfirm the association between the FEZ1 gene and schizophrenia in aJapanese population. Neurosci. Lett., 417, 326–329.
25. Hodgkinson, C.A., Goldman, D., Ducci, F., DeRosse, P., Caycedo, D.A.,Newman, E.R., Kane, J.M., Roy, A. and Malhotra, A.K. (2007) The FEZ1gene shows no association to schizophrenia in Caucasian or AfricanAmerican populations. Neuropsychopharmacology, 32, 190–196.
26. Sloviter, R.S., Dichter, M.A., Rachinsky, T.L., Dean, E., Goodman, J.H.,Sollas, A.L. and Martin, D.L. (1996) Basal expression and induction ofglutamate decarboxylase and GABA in excitatory granule cells of the ratand monkey hippocampal dentate gyrus. J. Comp. Neurol., 373, 593–618.
27. Gainetdinov, R.R., Mohn, A.R. and Caron, M.G. (2001) Genetic animalmodels: focus on schizophrenia. Trends Neurosci., 24, 527–533.
28. Paule, M.G., Rowland, A.S., Ferguson, S.A., Chelonis, J.J., Tannock, R.,Swanson, J.M. and Castellanos, F.X. (2000) Attention deficit/hyperactivity disorder: characteristics, interventions and models.Neurotoxicol. Teratol., 22, 631–651.
29. Miyakawa, T., Leiter, L.M., Gerber, D.J., Gainetdinov, R.R., Sotnikova,T.D., Zeng, H., Caron, M.G. and Tonegawa, S. (2003) Conditionalcalcineurin knockout mice exhibit multiple abnormal behaviors related toschizophrenia. Proc. Natl Acad. Sci. USA, 100, 8987–8992.
30. Di Chiara, G., Morelli, M. and Consolo, S. (1994) Modulatory functionsof neurotransmitters in the striatum: ACh/dopamine/NMDA interactions.Trends Neurosci., 17, 228–233.
31. Naghavi, M.H., Hatziioannou, T., Gao, G. and Goff, S.P. (2005)Overexpression of fasciculation and elongation protein z-1 (FEZ1)induces a post-entry block to retroviruses in cultured cells. Genes Dev.,19, 1105–1115.
32. Suzuki, T., Okada, Y., Semba, S., Orba, Y., Yamanouchi, S., Endo, S.,Tanaka, S., Fujita, T., Kuroda, S., Nagashima, K. et al. (2005)Identification of FEZ1 as a protein that interacts with JC virus agnoproteinand microtubules: role of agnoprotein-induced dissociation of FEZ1 frommicrotubules in viral propagation. J. Biol. Chem., 280, 24948–24956.
33. Gerber, D.J., Sotnikova, T.D., Gainetdinov, R.R., Huang, S.Y., Caron,M.G. and Tonegawa, S. (2001) Hyperactivity, elevated dopaminergictransmission, and response to amphetamine in M1 muscarinicacetylcholine receptor-deficient mice. Proc. Natl Acad. Sci. USA, 98,15312–15317.
34. Morishima, Y., Miyakawa, T., Furuyashiki, T., Tanaka, Y., Mizuma, H.and Nakanishi, S. (2005) Enhanced cocaine responsiveness and impairedmotor coordination in metabotropic glutamate receptor subtype 2knockout mice. Proc. Natl Acad. Sci. USA, 102, 4170–4175.
35. Lieberman, J.A., Kane, J.M. and Alvir, J. (1987) Provocative tests withpsychostimulant drugs in schizophrenia. Psychopharmacology (Berl), 91,415–433.
36. Paterlini, M., Zakharenko, S.S., Lai, W.S., Qin, J., Zhang, H., Mukai, J.,Westphal, K.G., Olivier, B., Sulzer, D., Pavlidis, P. et al. (2005)Transcriptional and behavioral interaction between 22q11.2 orthologsmodulates schizophrenia-related phenotypes in mice. Nat. Neurosci., 8,1586–1594.
37. Pillai-Nair, N., Panicker, A.K., Rodriguiz, R.M., Gilmore, K.L.,Demyanenko, G.P., Huang, J.Z., Wetsel, W.C. and Maness, P.F. (2005)Neural cell adhesion molecule-secreting transgenic mice displayabnormalities in GABAergic interneurons and alterations in behavior.J. Neurosci., 25, 4659–4671.
38. Weinberger, D.R. (1987) Implications of normal brain development forthe pathogenesis of schizophrenia. Arch. Gen. Psychiatry, 44, 660–669.
39. Breier, A., Su, T.P., Saunders, R., Carson, R.E., Kolachana, B.S., deBartolomeis, A., Weinberger, D.R., Weisenfeld, N., Malhotra, A.K.,Eckelman, W.C. et al. (1997) Schizophrenia is associated with elevatedamphetamine-induced synaptic dopamine concentrations: evidence from a
novel positron emission tomography method. Proc. Natl Acad. Sci. USA,94, 2569–2574.
40. Carlsson, A., Waters, N., Holm-Waters, S., Tedroff, J., Nilsson, M. andCarlsson, M.L. (2001) Interactions between monoamines, glutamate, andGABA in schizophrenia: new evidence. Annu. Rev. Pharmacol. Toxicol.,41, 237–260.
41. Abi-Dargham, A., Rodenhiser, J., Printz, D., Zea-Ponce, Y., Gil, R.,Kegeles, L.S., Weiss, R., Cooper, T.B., Mann, J.J., Van Heertum, R.L.et al. (2000) Increased baseline occupancy of D2 receptors by dopaminein schizophrenia. Proc. Natl Acad. Sci. USA, 97, 8104–8109.
42. Blum, B.P. and Mann, J.J. (2002) The GABAergic system inschizophrenia. Int. J. Neuropsychopharmacol., 5, 159–179.
43. Lewis, D.A. and Gonzalez-Burgos, G. (2006) Pathophysiologically basedtreatment interventions in schizophrenia. Nat. Med., 12, 1016–1022.
44. Yee, B.K., Keist, R., von Boehmer, L., Studer, R., Benke, D., Hagenbuch,N., Dong, Y., Malenka, R.C., Fritschy, J.M., Bluethmann, H. et al. (2005)A schizophrenia-related sensorimotor deficit links a3-containing GABAA
receptors to a dopamine hyperfunction. Proc. Natl Acad. Sci. USA, 102,17154–17159.
45. Erbel-Sieler, C., Dudley, C., Zhou, Y., Wu, X., Estill, S.J., Han, T.,Diaz-Arrastia, R., Brunskill, E.W., Potter, S.S. and McKnight, S.L. (2004)Behavioral and regulatory abnormalities in mice deficient in the NPAS1and NPAS3 transcription factors. Proc. Natl Acad. Sci. USA, 101, 13648–13653.
46. Winterer, G. and Weinberger, D.R. (2004) Genes, dopamine and corticalsignal-to-noise ratio in schizophrenia. Trends Neurosci., 27, 683–690.
47. Hikida, T., Jaaro-Peled, H., Seshadri, S., Oishi, K., Hookway, C., Kong,S., Wu, D., Xue, R., Andrade, M., Tankou, S. et al. (2007) Dominant-negative DISC1 transgenic mice display schizophrenia-associatedphenotypes detected by measures translatable to humans. Proc. Natl Acad.Sci. USA, 104, 14501–14506.
48. Kvajo, M., McKellar, H., Arguello, P.A., Drew, L.J., Moore, H.,MacDermott, A.B., Karayiorgou, M. and Gogos, J.A. (2008) A mutationin mouse Disc1 that models a schizophrenia risk allele leads to specificalterations in neuronal architecture and cognition. Proc. Natl Acad. Sci.USA, 105, 7076–7081.
49. Li, W., Zhou, Y., Jentsch, J.D., Brown, R.A., Tian, X., Ehninger, D.,Hennah, W., Peltonen, L., Lonnqvist, J., Huttunen, M.O. et al. (2007)Specific developmental disruption of disrupted-in-schizophrenia-1function results in schizophrenia-related phenotypes in mice. Proc. Natl.Acad. Sci. USA, 104, 18280–18285.
50. Pletnikov, M.V., Ayhan, Y., Xu, Y., Nikolskaia, O., Ovanesov, M.,Huang, H., Mori, S., Moran, T.H. and Ross, C.A. (2008) Enlargement ofthe lateral ventricles in mutant DISC1 transgenic mice. Mol. Psychiatry,13, 115.
51. Nakayama, K., Ishida, N., Shirane, M., Inomata, A., Inoue, T., Shishido,N., Horii, I., Loh, D.Y. and Nakayama, K.I. (1996) Mice lacking p27Kip1
display increased body size, multiple organ hyperplasia, retinal dysplasia,and pituitary tumors. Cell, 85, 707–720.
52. Fukuda, T., Kosaka, T., Singer, W. and Galuske, R.A. (2006) Gapjunctions among dendrites of cortical GABAergic neurons establish adense and widespread intercolumnar network. J. Neurosci., 26, 3434–3443.
53. Ihara, M., Yamasaki, N., Hagiwara, A., Tanigaki, A., Kitano, A., Hikawa,R., Tomimoto, H., Noda, M., Takanashi, M., Mori, H. et al. (2007) Sept4,a component of presynaptic scaffold and Lewy bodies, is Required for thesuppression of alpha-synuclein neurotoxicity. Neuron, 53, 519–533.
54. Niwa, M., Nitta, A., Yamada, Y., Nakajima, A., Saito, K., Seishima, M.,Shen, L., Noda, Y., Furukawa, S. and Nabeshima, T. (2007) An inducerfor glial cell line-derived neurotrophic factor and tumor necrosis factor-aprotects against methamphetamine-induced rewarding effects andsensitization. Biol. Psychiatry, 61, 890–901.
Human Molecular Genetics, 2008, Vol. 17, No. 20 3203
by guest on Novem
ber 21, 2013http://hm
g.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















