
Kinase-likeness and Kinase-Privileged Fragments: Toward Virtual Polypharmacology
Upload
independentCategory
view
0download
0
Metapopulation viability of the marsupial Micoureus demeraraein small Atlantic forest fragments in south-eastern Brazil
INTRODUCTIONHabitat loss and fragmentation are a serious threat tobiological diversity, and a primary cause of the presentextinction crisis (Wilcox & Murphy, 1985; Krebs, 1994).This world-wide process is probably the main factorendangering the rich Atlantic Forest fauna of Brazil. Afew centuries ago, Atlantic Forest was found all alongthe Brazilian eastern coast, covering over a millionsquare kilometers. Nowadays it is one of the most endan-gered ecosystems of the world (Fonseca, 1985): only 5%of its original cover is left and, with the exception ofsome biological reserves and parks, most of the rem-nants are small fragments, ranging from tens to hundredsof hectares.
Populations of different species of Atlantic Forestmarsupials and rodents show a diverse array of responsesto the changed landscape: some are largely unaffected,some become completely isolated, and others are foundas metapopulations (Fernandez et al., 1998; Pires &Fernandez, 1999). Much of the habitat loss occurredwithin the last few decades, and some of the effects of
isolation of populations may be felt only over a longertime span; therefore an important conservation issue isto evaluate the perspectives for long-term survival ofthese isolated populations and metapopulations.Although most of the species concerned are not globallyendangered, frequent losses of populations found todayin the small fragments may lead to ecological extinc-tions (sensu Redford, 1992), causing considerablechanges in the structure and functioning of the localcommunities preserved in the remnants.
Since the 1980s, the understanding of the demo-graphic, environmental and genetic processes that maybring small populations to extinction has increased con-siderably; such ideas merged in what the late GraemeCaughley (1994) called the ‘small population paradigm’.Building up from this conceptual basis, PopulationViability Analysis (PVA) established itself as a usefuland increasingly popular tool, not only for evaluating theperspectives for long-term survival of populations, butalso for providing useful guidelines for choosing amongmanagement options (Possingham, Lindenmayer &Norton, 1993). It has proved its usefulness as a tool in conservation and management, being applied world-wide by wildlife researchers and managers(Lindenmayer et al., 1993a). The growing list of species
Animal Conservation (2000) 3, 201–209 © 2000 The Zoological Society of London Printed in the United Kingdom
Daniel Brito and Fernando A. S. FernandezLaboratório de Ecologia e Conservação de Populações, Departamento de Ecologia, CCS, Universidade Federal do Rio de Janeiro, CxP68020, Ilha do Fundão, Rio de Janeiro, RJ, 21941-590, Brazil
(Received 30 June 1999; accepted 21 December 1999)
AbstractA population viability analysis (PVA) was carried out for populations of the arboreal didelphidMicoureus demerarae in eight small (1.5–15.0 ha) forest fragments in south-eastern Brazil. Analysiswas based on field data obtained through demographic studies carried out since 1995. Populations aresmall, but connected by dispersing individuals, thus forming a metapopulation. Frequency of cata-strophic fires was estimated from the Reserve’s historical records. We used the computer packageVORTEX for all analyses. All populations and the metapopulation were found to be endangered within100 years (extinction probability > 0.98). A sensitivity analysis was run varying six parameters: threedemographic (sex ratio, migration and mortality rates), two environmental (K, fire frequency) and onegenetic (level of inbreeding depression). Genetics, K, mortality rates and sex ratio seemed to playmajor roles to population persistence, whereas catastrophes and migration rates had a secondary role.Among demographic factors, extinction rate was least sensitive to migration rate. Micoureus demer-arae can be used as a model species, thus improving our knowledge of how extinction-prone popu-lations of neotropical arboreal marsupials in forest fragments might be, and which management actionscould decrease such risks.
All correspondence to: D. Brito. Tel: 55-21-2903308 ext. 31; Fax:55-21-2903308 ext. 34; E-mail: [email protected].

for which PVAs have already been carried out includesseveral arboreal marsupials such as the yellow-belliedglider (Goldingay & Possingham, 1995), theLeadbeater’s possum (Lindenmayer & Possingham,1995a,b, 1996a), the mountain brushtail possum(Lindenmayer & Lacy, 1995a), and the greater glider(Possingham et al., 1994).
PVAs are particularly useful when associated withsensitivity analysis (McCarthy, Burgman & Ferson,1995). In sensitivity analysis, as well as simulationsusing the parameters actually estimated, additional sim-ulations are run varying those parameters across speci-fied ranges, and observing the effects of this procedureon the PVA results, that is, on the estimated probabili-ties of population survival. This method brings twoadvantages. First, it increases the reliability of PVA byallowing for imperfections in parameter estimation.Second, it shows if the probabilities of survival of thepopulation are especially sensitive to just a few factors,which are the ones that affect most the persistence ornot of these populations, and therefore the ones thatshould be the main target of conservation and manage-ment strategies (Possingham et al., 1993).
Neotropical marsupials are a taxon that is particularlythreatened by habitat loss and the fragmentation processdue to the arboreal habits observed in the great major-ity of species (Nowak, 1991; Emmons & Feer, 1997).In this study, we had the objective of evaluating the rel-ative roles of several factors that may threaten popula-tions of a marsupial, the long-furred wooly mouseopossum, Micoureus demerarae, in small fragments ofAtlantic Forest, both a taxon and an area largelyneglected so far by published PVAs. The approach usedwas a PVA based mostly on field data obtained from the
populations under study, followed by sensitivity analy-sis in order to compare the importance of the factorsunder a range of plausible conditions.
METHODS
Study area and trapping designPoço das Antas Biological Reserve (22°30!–22°33! S;42°15!–42°19! W) covers a total area of 5500 ha, overhalf of which is covered with lowland Atlantic Forest(Pádua & Coimbra Filho, 1989). Data collection was car-ried out in two patches that are part of a group of eightsmall fragments of Atlantic Forest, with areas rangingfrom 1.5–15 ha. The set of fragments, known as ‘Ilhasdos Barbados’ (‘howler monkeys’ islands’), are sur-rounded by an open area composed mostly of grassesand bracken, hereafter called matrix. The nearest largeAtlantic Forest block is the reserve’s main forest patch,of about 3000 ha and located approximately 2 km fromthe fragments (Fig. 1).
The fragments named A (7.0 ha) and D (8.8 ha) weresampled every other month, A from March1995–September 1998, and D from April 1996–June1998. Each trapping session comprised five consecutivenights. We marked trapping paths transversal to thegreatest length of each fragment, separated by 50 m.Within each path, trapping points were separated by20 m. The grids covered the whole area of each frag-ment. There were two live traps at each trapping point,one on the ground and the other on a tree at breast height,each baited with a mixture of banana, peanut butter,bacon, oat grain and manioc. We used Sherman andTomahawk live traps. Animals were individually marked
202 D. BRITO & F. A. S. FERNANDEZ
Fig. 1. The study area: the group of eight Atlantic Forest fragments known as ‘Ilhas dos Barbados’ (‘Howler Monkeys’ islands’),within Poço das Antas Biological Reserve, south-eastern Brazil.

with two coded ear-tags (Le Boulengé-Nguyen & LeBoulengé, 1986). They were weighed, sized (ear and taillength), aged, sexed and their reproductive condition wasdetermined. After recording the data, each individualwas released at the same point that it had been captured.
Life history attributes of M. demeraraeMicoureus demerarae is a small (≈ 130 g) solitary didel-phid marsupial. The species is widespread in SouthAmerica, from Colombia to northern Argentina,although recent taxonomic work indicates that theAtlantic coast populations actually belong to a separatespecies in relation to the Amazonian forms (Vidigal,1997). Micoureus demerarae is found in mature andsecondary evergreen rainforest; it is nocturnal and hasmostly arboreal habits, although it often descends to theground. These marsupials favour dense viney vegetationwith many palm trees, but are also found in open highforest. They feed on insects, small animals, fruits andnectar (Emmons & Feer, 1997). Micoureus demeraraeis polygynous, with male home ranges overlapping thoseof females (Pires, Fernandez & Freitas, 1999). Adultfemales first breed successfully at 6 months of age witha maximum of 11 young per litter. The longest life spanrecorded in our field study was about 24 months (Rocha,2000).
The two populations studied at Poço das Antas arepart of a metapopulation (Pires & Fernandez, 1999).Both populations are very small, since population sizesestimated using the ‘jack-knife’ method (Burnham &Overton, 1979) seldom exceeded 20 individuals foreither fragment during the study (Quental et al., inpress). Micoureus demerarae inhabits all eight of theforest fragments, and movements have already beenrecorded among at least four of the fragment popula-tions.
PVA package usedThe computer simulation package VORTEX version8.21 (Miller & Lacy, 1999) was used. VORTEX is aMonte Carlo simulation of the effects of deterministicforces as well as demographic, environmental andgenetic stochasticity on the dynamics of wildlife popu-lations (Miller & Lacy, 1999). This package is one ofthe most often used programs for PVAs that focus on endangered populations, including in workshops withofficers from conservation and land management agen-cies (Lindenmayer et al., 1995).
Data sourcesThe demographic parameters used to parameterize thePVA model were all based on the results obtained dur-ing our field study, some of them published elsewhere(Fernandez et al., 1998; Pires & Fernandez, 1999; Pireset al., 1999). Parameters estimated for populations A and D were averaged and set for all eight patches. Thecarrying capacities (K) of the populations (A, 23; B, 5;
C, 35; D, 29; E, 37; F, 41; G, 32; H, 50) were estimatedusing the areas of the fragments and number of homeranges each fragment would support, based on home-range sizes and patterns of overlap for M. demeraraewithin the study area, as studied by Pires & Fernandez(1999). The initial population size for all iterations washalf the K of each patch. Forest fires are probably themost dangerous and destructive catastrophes in Poço dasAntas; fire frequency was estimated from the reserve’shistorical records. The most recent fire took place dur-ing the study, on August 18th, 1997. The only parame-ters for which we could not use actual field informationwere the genetic ones; there is an ongoing genetic studyon M. demerarae at the Howler Monkeys’ islands, butit has not yet provided conclusive results on the geneticstructure of these populations. Therefore, the geneticparameters are based on VORTEX 8.21 default valuesof 3.14 lethal equivalents with 50% of that due to lethalalleles (Miller & Lacy, 1999). The PVA input data usedare listed in Table 1.
Data analysis and output dataData derived from the computer simulation analysisusing VORTEX were: (1) population growth rate (± 1SD), (2) probability of population and metapopulationextinction, (3) median time to extinction, (4) mean pop-ulation size (± 1 SD) and (5) the decline in genetic vari-ability (expressed as the expected heterozygosity or genediversity). The probability of extinction is the numberof simulations that went extinct during a certain periodof time divided by the total number of simulations(Ballou, 1992). Extinct populations were not included incalculations of mean population growth, population size,or levels of expected heterozygosity.
The program was run first using the actual estimated
203PVA for the neotropical marsupial M. demerarae
Table 1. Life history parameters of Micoureus demerarae used asinput to the computer program VORTEX for Population ViabilityAnalysis
Parameter ValueInbreeding depression (number of lethal equivalents) 3.14Breeding system PolygynousAdult males in breeding pool (%) 100Breeding age of males / females (months) 6 / 6Maximum number of young/year 11Adult females producing:
6 young (%) 8.57 young (%) –8 young (%) 49 young (%) 4
10 young (%) –11 young (%) 83.5
Adult mortality (annual) (%) 53Maximum longevity (months) 24Adult sex ratio 1:1Home range of males/females (in ha) 0.8 / 0.4Probability of catastrophe (fire) (%) 8.33
Effect on reproduction 0.715Effect on survival 0.730
Migration rate (%) 0.65
Parameters were averaged from patches A and D, for which detailedfield data are available, and the averages used for all eight patches.
values of the parameters, as described in the previoussection and shown in Table 1 (baseline, BL). For sensi-tivity analysis, we selected six factors: three demo-graphic (migration rates, mortality rates and sex ratio),two environmental (carrying capacity and frequency ofcatastrophe) and one genetic (level of inbreeding depres-sion). These factors were assumed to be potentially ofgreat importance for determining the likelihood of per-sistence of the populations. In the case of sex ratio, thisfactor was regarded as worth analyzing because it var-ied between the populations studied: adult sex ratio wasfemale-biased in the population in fragment A (17 malesand 41 females, χ2 with Yates correction = 4.359, P =0.0368), whereas in fragment D sex ratio did not differsignificantly from 1:1 (18 males and 13 females, χ2 withYates correction = 0.146, P = 0.7023). In our model, sexratio (measured as male percentage in the population)was simulated through differential adult mortalitybetween the sexes resulting in the ratios observed in eachpopulation.
For each of the demographic and environmental fac-tors described above, taken separately, the program wasrun a second time using values set at 50%, 75%, 125%and 150% of the baseline estimates in Table 1, thus char-acterizing 24 different scenarios. We labelled the simu-lations in sensitivity analysis as highly optimistic (HO),optimistic (OP), pessimistic (PM) and highly pessimistic(HP) scenarios. Such labels were used because chang-ing a baseline value by a given percentage can haveeither a positive or a negative effect on population per-sistence, according to the factor analysed (e.g. 50% ofmortality has a positive effect, whereas 50% of K has anegative impact on the population). Optimistic scenar-ios were built either by increasing the values of a fac-tor with positive effects or by decreasing the values ofa factor with negative effects, whereas the inverse pro-cedure was used to build pessimistic scenarios. As malesare the dispersing sex, local extinctions of this sex canbe reversed by recolonization by dispersing males, butthe same is not true for females; therefore we consid-ered a female-biased population to be a better scenario
for population persistence. Sex ratio had populationswith male percentages set at: 25% (HO), 37.5% (OP),50% (BL), 62.5% (PM) and 75% (HP). Regarding thegenetic factor, as well as the baseline scenario (with 3.14lethal equivalents), we ran a simulation with one lethalequivalent to characterize the optimistic scenario,whereas in the highly optimistic scenario we assumedzero lethal equivalents affecting the populations; no sce-nario worse than the baseline was used for genetics.
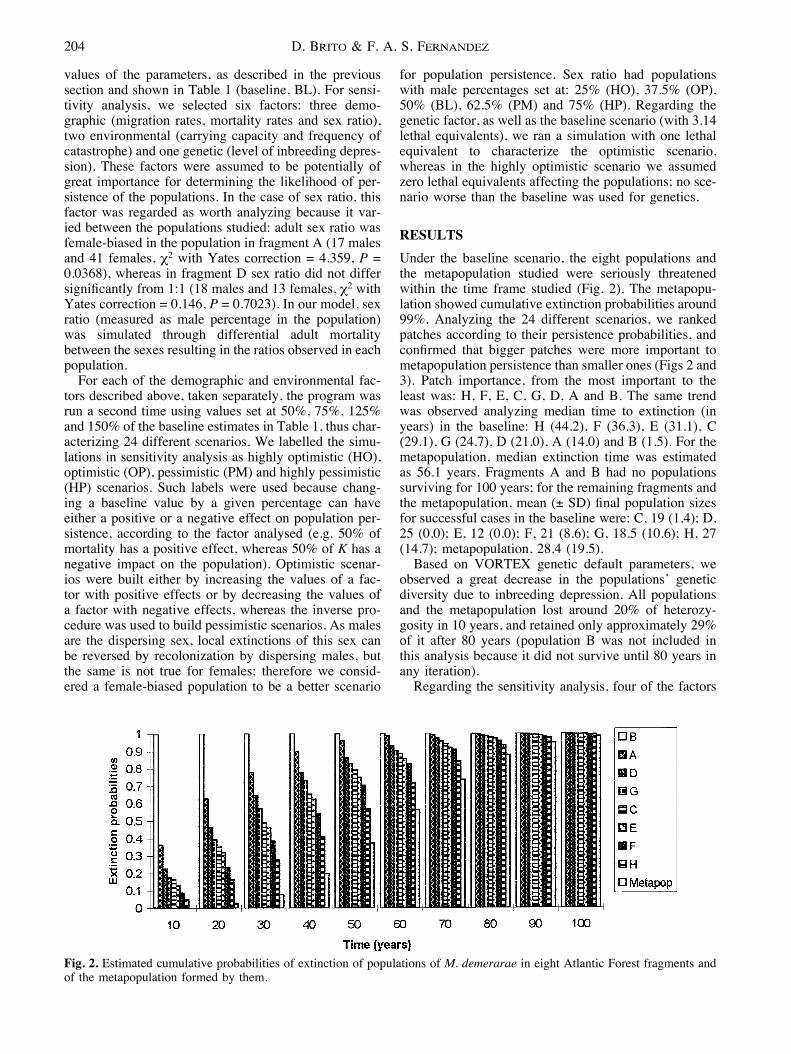
RESULTSUnder the baseline scenario, the eight populations andthe metapopulation studied were seriously threatenedwithin the time frame studied (Fig. 2). The metapopu-lation showed cumulative extinction probabilities around99%. Analyzing the 24 different scenarios, we rankedpatches according to their persistence probabilities, andconfirmed that bigger patches were more important tometapopulation persistence than smaller ones (Figs 2 and3). Patch importance, from the most important to theleast was: H, F, E, C, G, D, A and B. The same trendwas observed analyzing median time to extinction (inyears) in the baseline: H (44.2), F (36.3), E (31.1), C(29.1), G (24.7), D (21.0), A (14.0) and B (1.5). For themetapopulation, median extinction time was estimatedas 56.1 years. Fragments A and B had no populationssurviving for 100 years; for the remaining fragments andthe metapopulation, mean (± SD) final population sizesfor successful cases in the baseline were: C, 19 (1.4); D,25 (0.0); E, 12 (0.0); F, 21 (8.6); G, 18.5 (10.6); H, 27(14.7); metapopulation, 28.4 (19.5).
Based on VORTEX genetic default parameters, weobserved a great decrease in the populations’ geneticdiversity due to inbreeding depression. All populationsand the metapopulation lost around 20% of heterozy-gosity in 10 years, and retained only approximately 29%of it after 80 years (population B was not included inthis analysis because it did not survive until 80 years inany iteration).
Regarding the sensitivity analysis, four of the factors
204 D. BRITO & F. A. S. FERNANDEZ
Fig. 2. Estimated cumulative probabilities of extinction of populations of M. demerarae in eight Atlantic Forest fragments andof the metapopulation formed by them.

studied were important, with the probabilities of persis-tence of the populations and the metapopulation beingvery sensitive to changes in their values. They weregenetics, carrying capacity, mortality rates and sex ratio(Fig. 3). However, migration rates and frequency of cat-astrophes were found to be of lesser importance in thesimulations: manipulating their values had little effecton the probabilities of persistence of either the popula-tions or the metapopulation (Fig. 3).
DISCUSSIONBoyce (1992) suggested that performing PVAs withspecies in no danger of extinction could be a great con-tribution, because the populations could be manipulatedexperimentally to rigorously test the predictions of themodel. Besides, even species that are not currentlyendangered and are presently common may be vulnera-
ble to the long-term effects of fragmentation if they arereduced to small populations (Lindenmayer & Lacy,1995a). Micoureus demerarae is roughly representativeof neotropical marsupials in many important character-istics: its arboreal habits together with its capacity forusing the ground occasionally, its typical population den-sities (and consequently its K in small fragments), itssurvival rates and its reproductive output. Therefore,evaluating the risks suffered by its populations providesa starting point for understanding the vulnerability ofmarsupials to local extinction in fragmented neotropicalforests. Such information coupled with decision analy-sis (Maguire, Seal & Brussard, 1987) may suggestmanagement options that could be used not only forM. demerarae, but hopefully for other neotropical arbo-real marsupials as well.
A point of concern is that fluctuations of the two pop-ulations studied in detail (A and D) were synchronous;
205PVA for the neotropical marsupial M. demerarae
Fig. 3. Sensitivity analysis: the probabilities of extinction of each population and the metapopulation in a 100 year time frame,varying each of the six factors studied, under five different scenarios, which are described in the text. Scenarios are: HO, highlyoptimistic scenario ( ); OP, optimistic scenario ( ); BL, baseline scenario (■■); PM, pessimistic scenario ( ); HP, highly pes-simistic scenario ( ).

if the same applies to all fragments, the vulnerability ofthe metapopulation may be higher than estimated.Asynchrony can average out fluctuations caused bydemographic or environmental stochasticity and mayensure species survival in the face of catastrophes(Gilpin, 1987), whereas correlated fluctuations amongpopulations can drastically reduce survival. Also,M. demerarae forms an atypical metapopulation whereonly the males migrate, and therefore local extinctionscannot be replaced by recolonization, since males alonecannot found new populations (Pires & Fernandez,1999). As noticed by Lindenmayer & Lacy (1995b), ifsex-biased migration occurs, the instability of metapop-ulations composed of ensembles of small populations isprobably greater than predicted by the analysis.
Persistence times and probabilities were very sensi-tive to variation in some demographic factors (mortalityrates and sex ratio). This is an expected result, since atsmall population sizes demography plays an importantrole in population persistence (Goodman, 1987; Nunney
& Campbell, 1993; Lindenmayer & Lacy, 1995a).Catastrophes, on the other hand, were found not to beso important in the iterations. Low intensity fires had aminimal effect on populations of arboreal marsupials inAustralia (Inions, 1985 cited in Lindenmayer et al.,1993b), but for Gymnobelideus leadbeateri, large firesincreased mortality rates because of elevated predationrates and/or a shortage of food and hollows(Lindenmayer & Possingham, 1996b). However, thegreater glider, Petauroides volans, was found to beseverely threatened by fire (Possingham et al., 1994). Inour case, M. demerarae populations were severely hitby fire in August 1997 but recovered quickly. With suchsmall population sizes, the immediate dangers of thegenetic and demographic vortexes are so overwhelmingthat the effects of fire may be of lesser importance.
Lindenmayer & Lacy (1995b) stated that whereasmigration decreased the chances of extinction for largepopulations, it could be detrimental to the persistence ofsmall populations of the Australian marsupials
206 D. BRITO & F. A. S. FERNANDEZ
Fig. 3. continued

Trichosurus caninus and G. leadbeateri. In these cases,median time to extinction was predicted to be longer ina single isolated population than in metapopulations.Increased emigration could actually reduce thepersistence of very small populations, desestabilizingmetapopulation dynamics; this pattern could be the resultof negative feedback processes creating a metapopula-tion extinction vortex (Lindenmayer & Lacy, 1995b,c)similar to the extinction vortexes described by Gilpin &Soulé (1986). However, in this same study, they foundthat higher rates of migration had no negative impact insmall populations of P. volans, another marsupial. Theeffects of migration are therefore complex and can dif-fer according to population size and species studied,even if they are from closely related taxa. In the presentstudy, we observed a small, but positive effect of migra-tion to the persistence of a small metapopulation, sug-geting that improved migration rates would bring onlya limited benefit as a management action.
There is a common view within the small population
paradigm that demographic effects are likely to be feltbefore genetic ones (Soulé, 1987; Nunney & Campbell,1993; Caughley & Gunn, 1996). However, genetics, andespecially inbreeding depression, can play a major rolein extinction vulnerability, its effects being as severe andas fast as the demographic ones. We should be carefulabout putting too much emphasis on our conclusionssince they are based on default parameters rather thanon actual field data. A natural step for our work is theongoing evaluation of the genetic impoverishment of thereal populations, which would allow this information tobe included in future analyses. While studying the arbo-real marsupial G. leadbeateri in western Australia,Lindenmayer et al. (1993b) found that populations of100 or more were demographically stable, and popula-tions of 500 or more would suffer no significant loss ingenetic variation over 100 years, but small populationsof 25 and 50 individuals would not be viable. In ourstudy we had very small population sizes (around 20individuals) and, using default parameters, we observed
207PVA for the neotropical marsupial M. demerarae
Fig. 3. continued

a great loss of genetic variability (around 20%) in thefirst 10 years of simulation, suggesting that for these populations, the short-term genetic depauperationcould be a severe threat. As heterozygosity and fitnessare directly related in natural populations of animals,depressing the first should lead to a reduction of thesecond (Allendorf & Leary, 1986). Genetic problems area real threat to the persistence of wild populations of several mammalian taxa, acting through their effectson demographic and ecological processes (Lacy, 1997);gradual depletion of genetic variability can be well onits way before there is any evidence of a marked declinein population size (Lindenmayer et al., 1993b).
The factors chosen for our sensitivity analysis werethose that could be manipulated experimentally, increas-ing our knowledge of the reliability of PVA. These fac-tors can also be the target for different potentialmanagement options. Our results suggest several man-agement priorities, affecting the four factors pointed outby sensitivity analysis. Genetic problems could bereduced by the periodic translocation of individuals fromthe nearby larger population of the reserve’s main for-est block to the metapopulation, in order to preventexcessive inbreeding. Carrying capacities (K values) arelinked to the sizes of the fragments, which are decreasedby fire and edge effects, and could be managed by firecontrol and/or by reforestation programmes. A decreasein mortality rates could possibly be achieved by addi-tional food supply during periods of resource scarcity.Also, since the results suggested that a female-biased sexratio is beneficial to population persistence, periodictranslocation of females to the fragments could be anespecially useful management option. More generally,we hope that this study, by pointing out some importantfactors, will help to focus attention on them for furtherstudy and potentially for the management of very smallpopulations of marsupials in fragments, which areincreasingly common in the Neotropics.
AcknowledgementsWe thank IBAMA – Brazilian Institute of theEnvironment and Renewable Natural Resources – espe-cially through D. Pessamilio, W. Junior and I. C. Astor,for allowing us to work at Poço das Antas and for pro-viding facilities there. D. Rambaldi of the Golden LionTamarin Association provided helpful administrativeand practical support. We also thank the many col-leagues who helped in field work and contributed withfield data, especially A. S. Pires, T. B. Quental, F. S.Rocha and L. C. Oliveira. Useful comments and sug-gestions were provided by A. S. Pires, T. B. Quental,M. Passamani, J. H. Brown and D. Simberloff. We alsothank Barry W. Brook and an anonymous referee, fortheir suggestions which greatly improved the article. Thework was supported by Fundação O Boticário deProteção à Natureza, The MacArthur Foundation,FAPERJ, and Fundação Universitária José Bonifácio,and by Brazil’s National Biodiversity Program (PRO-BIO / PRONABIO-MMA, funded by BIRD / GEF).
REFERENCESAllendorf, F. W. & Leary, R. F. (1986). Heterozygosity and fit-
ness in natural populations of animals. In Conservation Biology:the Science of Scarcity and Diversity: 57–76. Soulé, M. E. (Ed.).Sunderland, MA: Sinauer Associates.
Ballou, J. (1992). Small population overview. In VORTEX: UsersManual. A Stochastic Simulation of the Extinction Process:40–49. Lacy, R. & Kreeger, T. (Eds). Apple Valley: CaptiveBreeding Specialist Group.
Boyce, M. S. (1992). Population viability analysis. Annu. Rev.Ecol. Syst. 23: 481–506.
Burnham, K. P. & Overton, W. S. (1979). Robust estimation ofpopulation size when capture probabilities vary among animals.Ecology 60: 927–936.
Caughley, G. (1994). Directions in conservation biology. J. Anim.Ecol. 63: 215–244.
Caughley, D. & Gunn, A. (1996). Conservation Biology in Theoryand Practice. Oxford: Blackwell.
Emmons, L. H. & Feer, F. (1997). Neotropical RainforestMammals: A Field Guide (2nd edn). Chicago: The Universityof Chicago Press.
Fernandez, F. A. S., Pires, A. S., Freitas, D., Rocha, F. S. &Quental, T. B. (1998). Respostas de pequenos mamíferos, à frag-mentação de habitats em remanescentes de Mata Atlântica. InAnais do IV Simpósio de Ecossistemas Brasileiros – volume 5:184–189. Águas de Lindóia, SP: Academia de Ciências doEstado de Sâo Paolo.
Fonseca, G. A. B. (1985). The vanishing Brazilian Atlantic Forest.Biol. Conserv. 34: 17–34.
Gilpin, M. E. & Soulé, M. E. (1986). Minimum viable popula-tions: processes of extinction. In Conservation Biology: theScience of Scarcity and Diversity: 19–34. Soulé, M. E. (Ed.).Sunderland, MA: Sinauer Associates.
Gilpin, M. E. (1987). Spatial structure and population viability.In Viable Populations for Conservation:: 125–139. Soulé, M. E.(Ed.). Cambridge: Cambridge University Press.
Goldingay, R. & Possingham, H. P. (1995). Area requirementsfor viable populations of the Australian gliding marsupialPetaurus australis. Biol. Conserv. 73: 161–167.
Goodman, D. (1987). The demography of chance extinction. InViable Populations for Conservation: 11–34. Soulé, M. E. (Ed.).Cambridge: Cambridge University Press.
Krebs, C. J. (1994). Ecology: the Experimental Analysis ofDistribution and Abundance (4th edn). New York: HarperCollins College Publishers.
Lacy, R. C. (1997). Importance of genetic variation to the via-bility of mammalian populations. J. Mammal. 78: 320–335.
Le Boulengé-Nguyen, P. & Le Boulengé, E. (1986). A new ear-tag for small mammals. J. Zool. Lond. 209: 302–304.
Lindenmayer, D. B. & Lacy, R. C. (1995a). A simulation studyof the impacts of population subdivision on the mountain brush-tail possum Trichosurus caninus Ogilby (Phalangeridae :Marsupialia) in south-eastern Australia. I. Demographic stabil-ity and population persistence. Biol. Conserv. 73: 119–129.
Lindenmayer, D. B. & Lacy, R. C. (1995b). Metapopulation via-bility of arboreal marsupials in fragmented old-growth forests:comparison among species. Ecol. Applic. 5: 183–199.
Lindenmayer, D. B. & Lacy, R. C. (1995c). Metapopulation ofthe Leadbeater’s possum Gymnobelideus leadbeateri, in frag-mented old-growth forests. Ecol. Applic. 5: 164–182.
Lindenmayer, D. B. & Possingham, H. P. (1995a). The conser-vation of arboreal marsupials in the montane ash forests of thecentral highlands of Victoria, south-eastern Australia – VII. Model-ling the persistence of Leadbeater’s possum in response to mod-ified timber harvesting practices. Biol. Conserv. 73: 239–257.
Lindenmayer, D. B. & Possingham, H. P. (1995b). Modelling theviability of metapopulations of the endangered Leadbeater’s pos-sum in south-eastern Australia. Biodiv. Conserv. 4: 984–1018.
208 D. BRITO & F. A. S. FERNANDEZ

Lindenmayer, D. B. & Possingham, H. P. (1996a). Ranking con-servation and timber management options for Leadbeater’s pos-sum in southeastern Australia using population viabilityanalysis. Conserv. Biol. 10: 235–251.
Lindenmayer, D. B. & Possingham, H. P. (1996b). Modelling theinter-relationships between habitat patchiness, dispersal capa-bility and metapopulation persistence of the endangered species,Leadbeater’s possum, in south-eastern Australia. Landsc. Ecol.11: 79–105.
Lindenmayer, D. B., Clark, T. W., Lacy, R. C. & Thomas, V. C.(1993a). Population viability analysis as a tool in wildlife con-servation policy: with reference to Australia. Environ. Mgmt. 17:745–758.
Lindenmayer, D. B., Lacy, R. C., Thomas, V. C. & Clark, T. W.(1993b). Predictions of the impacts of changes in population sizeand environmental variability on Leadbeater’s possumGymnobelideus leadbeateri McCoy (Marsupialia : Petauridae)using population viability analysis: an application of the com-puter program VORTEX. Wildl. Res. 20: 67–86.
Lindenmayer, D. B., Burgman, M. A., Akçakaya, H. R., Lacy, R.C. & Possingham, H. P. (1995). A review of the generic com-puter programs ALEX, RAMAS/space and VORTEX for mod-elling the viability of wildlife metapopulations. Ecol. Mod. 82:161–174.
Maguire, L. A., Seal, U. S. & Brussard, P. F. (1987). Managingcritically endangered species: the Sumatran rhino as a case study.In Viable Populations for Conservation: 141–158. Soulé, M. E.(Ed.). Cambridge: Cambridge University Press.
McCarthy, M. A., Burgman, M. A. & Ferson, S. (1995).Sensitivity analysis for models of population viability analysis.Biol. Conserv. 73: 93–100.
Miller, P. S. & Lacy, R. C. (1999). VORTEX: A StochasticSimulation of the Extinction Process. Version 8 User’s Manual.Apple Valley: Conservation Breeding Specialist Group(SSC/IUCN).
Nowak, R. M. (1991). Walker’s Mammals of the World (5th edn).London: The Johns Hopkins University Press.
Nunney, L. & Campbell, K. A. (1993). Assessing minimum viable
population size: demography meets population genetics. TrendsEcol. Evol. 8: 234–239.
Pádua, M. T. J. & Coimbra Filho, A. F. (1989). Os ParquesNacionais do Brasil. São Paulo: José Olympio.
Pires, A. S. & Fernandez, F. A. S. (1999). Use of space by themarsupial Micoureus demerarae in small Atlantic Forest frag-ments in southeastern Brazil. J. Trop. Ecol. 15: 279–290.
Pires, A. S., Fernandez, F. A. S. & Freitas, D. (1999). Patterns ofspace use by Micoureus demerarae (Marsupialia : Didelphidae)in a fragment of Atlantic Forest in Brazil. Mastozool. Neotrop.6: 39–45.
Possingham, H. P., Lindenmayer, D. B. & Norton, T. W. (1993).A framework for the improved management of threatenedspecies based on population viability analysis (PVA). PacificConserv. Biol. 1: 39–45.
Possingham, H. P., Lindenmayer, D. B., Norton, T. W. & Davies,I. (1994). Metapopulation viability analysis of the greater gliderPetauroides volans in a wood production area. Biol. Conserv.70: 227–236.
Redford, K. H. (1992). The empty forest. Bioscience 42: 412–421.Rocha, F. S. (2000). Ecologia Reprodutiva de Pequenos
Mamíferos (com Ênfase no Marsupial Micoureus demerarae)em Fragmentos de Mata Atlântica no Sudeste do Brasil. MScthesis; Universidade Federal do Rio de Janeiro, Rio de Janeiro.
Quental, T. B., Fernandez, F. A. S., Dias, A. T. C. & Rocha, F.S. (2000). Population dynamics of the marsupial Micoureusdemerarae in small fragments of Atlantic Coastal Forest inBrazil. J. Trop. Ecol. (in press).
Soulé, M. E. (Ed.) (1987). Viable Populations for Conservation.Cambridge: Cambridge University Press.
Vidigal, V. C. S. (1997). Determination of Dental Age Classesand Estimation of Seasonal Population Dynamics andReproductive Patterns in Micoureus demerarae and Micoureusregina (Marsupialia : Didelphidae) from Rio Jurua, Brasil. BScHonor Thesis; University of California, Berkeley.
Wilcox, B. A. & Murphy, D. D. (1985). Conservation strategy:the effects of fragmentation on extinction. Am. Nat. 125:879–887.
209PVA for the neotropical marsupial M. demerarae

Copyright © 2022 FDOKUMEN




















