
Mercuric Ion Attenuates Nuclear Factor-κB Activation and DNA Binding in Normal Rat Kidney...
12
Mercuric Ion Attenuates Nuclear Factor-kB Activation and DNA Binding in Normal Rat Kidney Epithelial Cells: Implications for Mercury-Induced Nephrotoxicity Francisco J. Dieguez-Acun ˜a, Maureen E. Ellis, John Kushleika, and James S. Woods 1 Center for Ecogenetics and Environmental Health, Department of Environmental Health, University of Washington, Seattle, Washington 98105 Received February 19, 2001; accepted March 23, 2001 Mercuric Ion Attenuates Nuclear Factor-kB Activation and DNA Binding in Normal Rat Kidney Epithelial Cells: Implica- tions for Mercury-Induced Nephrotoxicity. Dieguez-Acun ˜ a, F. J., Ellis, M. E., Kushleika, J., and Woods, J. S. (2001). Toxicol. Appl. Pharmacol. 173, 176 –187. Mercuric ion (Hg 21 ), one of the strongest thiol-binding agents known, mediates the toxicity associated with elemental, inorganic, and organic mercurial compounds. Studies of cellular events as- sociated with Hg 21 toxicity have focused largely on disruption of cell membranes and impairment of mitochondrial functions. In contrast, few studies have sought to define the specific molecular mechanisms through which Hg 21 might affect toxicity via alter- ation of thiol-dependent signal transduction pathways that regu- late cell proliferation and survival. Of particular interest in this regard is the effect of Hg 21 on nuclear factor-kB (NF-kB), a pleiotropic transcriptional factor that is known to require reduced cysteine moieties at critical steps of activation and DNA binding. Here, we evaluated the effects of Hg 21 on the expression of NF-kB in normal rat kidney epithelial (NRK52E) cells, a principal target of Hg 21 toxicity. The lipopolysaccharide (LPS)-inducible form of NF-kB was readily detected in kidney cells and has been charac- terized as the p50p65 heterodimer. NF-kB–DNA binding was prevented in a dose-related manner by Hg 21 (0 –55 mM) in vitro when added to DNA binding reactions containing the nonthiol reducing agent Tris(2-carboxyethyl)phosphine hydrochloride (TCEP). Similarly, Hg 21 at the same concentrations prevented DNA binding of a human recombinant wild-type p50p50 ho- modimer in binding reactions, and this effect was attenuated using a mutant form of the p50 protein containing a cys 62 3ser 62 muta- tion. The inhibition of p50 –DNA binding by Hg 21 was reversible in a dose-related manner in vitro by competitive thiols DTT, GSH, and L-cysteine in binding reactions. In contrast, competitive thiols added to nuclear binding reactions were unable to reverse atten- uation of LPS-mediated NF-kB–DNA binding affinity when cells were pretreated in vivo with Hg 21 at concentrations as low as 2 mM prior to LPS administration. Immunoblot analyses indicted that Hg 21 pretreatment of kidney cells substantially diminished, in a dose-related manner, the concentration of p65 translocated into the nucleus following LPS administration. Additionally, Hg 21 pre- treatment impaired both the phosphorylation and degradation of IkBa, suggesting a specific effect on NF-kB activation at the level of IkBa proteolysis. Finally, Hg 21 at concentrations as low as 5 mM significantly diminished NF-kB-mediated transcriptional ac- tivity when administered to kidney cells transiently transfected with an NF-kB-driven luciferase reporter gene (pLuc-43NF-kB) prior to LPS treatment. These findings demonstrate that Hg 21 , at low cellular concentrations, attenuates NF-kB activation at sites associated with IkBa phosphorylation and degradation, nuclear translocation of the p50p65 heterodimer, and association of p50- cys 62 with the DNA kB binding site. Attenuation of NF-kB acti- vation by Hg 21 through these mechanisms may underlie apoptotic or other cytotoxic responses that are known to be associated with low level Hg 21 exposure in kidney epithelial cells. © 2001 Academic Press Key Words: mercury; mercuric ion; nuclear factor-kB; NF-kB; kidney; epithelial cells; lipopolysaccharide; LPS, DNA binding. Mercuric ion (Hg 21 ) is a potent nephrotoxicant with princi- pal effects directed toward proximal tubular epithelial cells of the S3 segment (pars recta) (Rodin and Crowson, 1962; Gratzka and Krump, 1968; Magos et al., 1984). Two principal properties of Hg 21 contribute to its distribution and toxicity: Hg 21 is among the most potent thiol binding agents known, having a first-order thiol binding constant in the range of 10 30–40 (Oram et al., 1996), many orders of magnitude greater than that of lead, cadmium, and other thiol-binding metals; and the ligand exchange rate constant of Hg 21 for thiol interactions is also among the highest known (10 9 /s) (Martin, 1986), en- suring extremely rapid partitioning of Hg 21 among thiol groups in tissues. Previous studies have demonstrated that the princi- pal effects of Hg 21 in kidney cells derive from thiol-directed modification of cellular constituents. These include structural alteration of the mitochondrial inner membrane with conse- quent dissipation of membrane potential, disruption of oxida- tive phosphorylation (Weinberg et al., 1982a,b; Southard et al., 1973, 1974; Lund et al., 1993), and alteration of intracellular calcium homeostasis (Lund et al., 1993; Smith et al., 1987, 1991; Chevez and Hoguin, 1988). Recent studies further sug- gest that modification of thiol-dependent signal transduction 1 To whom correspondence should be addressed at Department of Environ- mental Health, University of Washington, 4225 Roosevelt Way NE, Suite 100, Seattle, WA 98105. Fax: (206) 528-3550; E-mail: [email protected]. Toxicology and Applied Pharmacology 173, 176 –187 (2001) doi:10.1006/taap.2001.9195, available online at http://www.idealibrary.com on 176 0041-008X/01 $35.00 Copyright © 2001 by Academic Press All rights of reproduction in any form reserved.
-
Upload
washington -
Category
Documents
-
view
0 -
download
0
Transcript of Mercuric Ion Attenuates Nuclear Factor-κB Activation and DNA Binding in Normal Rat Kidney...

105
ioNtp
Dmat
pH
Toxicology and Applied Pharmacology173,176–187 (2001)doi:10.1006/taap.2001.9195, available online at http://www.idealibrary.com on
Mercuric Ion Attenuates Nuclear Factor-kB Activationand DNA Binding in Normal Rat Kidney Epithelial Cells:
Implications for Mercury-Induced Nephrotoxicity
Francisco J. Dieguez-Acun˜a, Maureen E. Ellis, John Kushleika, and James S. Woods1
Center for Ecogenetics and Environmental Health, Department of Environmental Health, University of Washington, Seattle, Washington 98
Received February 19, 2001; accepted March 23, 2001
ttI
p
vol
cip s oft 962;G lp y:
wn,of
terand
s srinci-d
turalnse-ida-
ar
1 sug-g tion
nvir10.ed
Mercuric Ion Attenuates Nuclear Factor-kB Activation andDNA Binding in Normal Rat Kidney Epithelial Cells: Implica-tions for Mercury-Induced Nephrotoxicity. Dieguez-Acuna, F. J.,Ellis, M. E., Kushleika, J., and Woods, J. S. (2001). Toxicol. Appl.Pharmacol. 173, 176–187.
Mercuric ion (Hg21), one of the strongest thiol-binding agentsknown, mediates the toxicity associated with elemental, inorganic,and organic mercurial compounds. Studies of cellular events as-sociated with Hg21 toxicity have focused largely on disruption ofcell membranes and impairment of mitochondrial functions. Incontrast, few studies have sought to define the specific molecularmechanisms through which Hg21 might affect toxicity via alter-ation of thiol-dependent signal transduction pathways that regu-late cell proliferation and survival. Of particular interest in thisregard is the effect of Hg21 on nuclear factor-kB (NF-kB), apleiotropic transcriptional factor that is known to require reducedcysteine moieties at critical steps of activation and DNA binding.Here, we evaluated the effects of Hg21 on the expression of NF-kBn normal rat kidney epithelial (NRK52E) cells, a principal targetf Hg21 toxicity. The lipopolysaccharide (LPS)-inducible form ofF-kB was readily detected in kidney cells and has been charac-
erized as the p50p65 heterodimer. NF-kB–DNA binding wasrevented in a dose-related manner by Hg21 (0–55 mM) in vitro
when added to DNA binding reactions containing the nonthiolreducing agent Tris(2-carboxyethyl)phosphine hydrochloride(TCEP). Similarly, Hg21 at the same concentrations prevented
NA binding of a human recombinant wild-type p50p50 ho-odimer in binding reactions, and this effect was attenuated usingmutant form of the p50 protein containing a cys623ser62 muta-
ion. The inhibition of p50–DNA binding by Hg21 was reversible ina dose-related manner in vitro by competitive thiols DTT, GSH,and L-cysteine in binding reactions. In contrast, competitive thiolsadded to nuclear binding reactions were unable to reverse atten-uation of LPS-mediated NF-kB–DNA binding affinity when cellswere pretreated in vivo with Hg21 at concentrations as low as 2 mM
rior to LPS administration. Immunoblot analyses indicted thatg21 pretreatment of kidney cells substantially diminished, in a
dose-related manner, the concentration of p65 translocated into
1 To whom correspondence should be addressed at Department of E-mental Health, University of Washington, 4225 Roosevelt Way NE, SuiteSeattle, WA 98105. Fax: (206) 528-3550; E-mail: [email protected]
1760041-008X/01 $35.00Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
he nucleus following LPS administration. Additionally, Hg21 pre-reatment impaired both the phosphorylation and degradation ofkBa, suggesting a specific effect on NF-kB activation at the level
of IkBa proteolysis. Finally, Hg21 at concentrations as low as 5mM significantly diminished NF-kB-mediated transcriptional ac-tivity when administered to kidney cells transiently transfectedwith an NF-kB-driven luciferase reporter gene (pLuc-43NF-kB)
rior to LPS treatment. These findings demonstrate that Hg21, atlow cellular concentrations, attenuates NF-kB activation at sitesassociated with IkBa phosphorylation and degradation, nucleartranslocation of the p50p65 heterodimer, and association of p50-cys62 with the DNA kB binding site. Attenuation of NF-kB acti-ation by Hg21 through these mechanisms may underlie apoptoticr other cytotoxic responses that are known to be associated withow level Hg21 exposure in kidney epithelial cells. © 2001 Academic Press
Key Words: mercury; mercuric ion; nuclear factor-kB; NF-kB;kidney; epithelial cells; lipopolysaccharide; LPS, DNA binding.
Mercuric ion (Hg21) is a potent nephrotoxicant with prin-al effects directed toward proximal tubular epithelial cell
he S3 segment (pars recta) (Rodin and Crowson, 1ratzka and Krump, 1968; Magoset al.,1984). Two principaroperties of Hg21 contribute to its distribution and toxicit
Hg21 is among the most potent thiol binding agents knohaving a first-order thiol binding constant in the range1030–40 (Oramet al., 1996), many orders of magnitude greathan that of lead, cadmium, and other thiol-binding metals;the ligand exchange rate constant of Hg21 for thiol interactionsis also among the highest known (109/s) (Martin, 1986), en-uring extremely rapid partitioning of Hg21 among thiol group
in tissues. Previous studies have demonstrated that the ppal effects of Hg21 in kidney cells derive from thiol-directemodification of cellular constituents. These include strucalteration of the mitochondrial inner membrane with coquent dissipation of membrane potential, disruption of oxtive phosphorylation (Weinberget al.,1982a,b; Southardet al.,1973, 1974; Lundet al., 1993), and alteration of intracellulcalcium homeostasis (Lundet al., 1993; Smithet al., 1987,
991; Chevez and Hoguin, 1988). Recent studies furtherest that modification of thiol-dependent signal transduc
on0,u.

s,
ndb do
aln oro
1 llsubedate
c da 9),
es-
er
aac
ea
cyetar
rip89s
y
rteitei-
eeofinal
ntringin,ing
taie
p50foreF-n toeys
b itsi r,
fF-gc-
,ide0,
2 reea ls, andb epesa anap-o uiredf iums fastfl -t hamL romG la pur-c l
A).
E k( Tris–g fromN ottingg CA).E Phar-m
werep lep owerswc edb nd).P chasedf diumd kers-v
E)c ltureC ifiedE -
erumll
m ostar,
177Hg21 IMPAIRS NF-kB ACTIVATION AND DNA BINDING IN KIDNEY CELLS
processes may be associated with Hg21-mediated toxic eventincluding apoptosis (Shumillaet al., 1998; Eckhardtet al.,1999; McCabeet al., 1999). However, the mechanisms a
iochemical consequences of these effects have not beennstrated.Nuclear factor-kB (NF-kB) is a mammalian transcription
activator known to be involved in the inducible expressioa variety of genes, particularly those involved in cellular pliferative, antiapoptotic, and tumorigenic processes (Shaet al.,
995; Kretz-Renyet al.,1996; Chenet al.,1999). In most cetypes, NF-kB exists as a heterodimer composed of twounits with molecular mass values of 50 and 65 kDa, referras p50 (NF-kB1) and p65 (Rel A), respectively. In cells thhave inducible NF-kB activity, NF-kB exists as an inactivytoplasmic form consisting of a p50p65 heterodimer bounn inhibitory protein, IkB (Baeuerle and Baltimore, 198
comprised of two principal moieties, IkBa and IkBb. Thetranscriptional activation of NF-kB-dependent genes involvdissociation of IkB from the NF-kB heterodimer in the cytoplasm and subsequent translocation of active NF-kB into thenucleus, where it binds with thek enhancer in the promotregion of various genes to effect transcription (Mu¨ller et al.,1993). Depending on the cell type, NF-kB is activated bywide range of stimuli including viruses, bacterial lipopolyscharide (LPS), cytokines, and reactive oxygen species.
NF-kB–DNA binding is thought to be regulated by nuclredox state (Staalet al., 1990; Schulze-Osthoffet al., 1995).Both p50 and p65 homodimers contain a redox-sensitiveteinyl group that effects DNA binding. In the p50p65 herodimer it is believed that the p50 subunit plays the primrole in DNA binding, and p65 is responsible for the transctional activity of the complex (Baeuerle and Baltimore, 19Fujita et al.,1992; Toledanoet al.,1993). Dimer formation ialso required for DNA binding of NF-kB subunits (Toledanoetal., 1993). In vitro oxidation (by diamide) or alkylation (bN-ethylmaleimide) of free sulfhydryls attenuated NF-kB–DNAbinding (Toledano and Leonard, 1991). Moreover, Kumaetal. (1992) found that serine substitution of a conserved cysresidue in the N-terminal domain of the v-Rel oncopro(conserved in all NF-kB/rel/Dorsal family transcription factors) eliminated redox control of DNA binding. It has also bdemonstrated that cys62 located in the N-terminal domainp50 mediates the redox effect required for p50 DNA bind(Toledanoet al., 1993). Cysteine to serine mutations onother cysteines located on p50 had no effect on redox coMitomo and colleagues (1994) showed similar results usp50 cys623ser62 mutant. Additional studies using thioredoxan endogenous disulphide redox catalyst, enhanced bindwild-type p50 but did not increase cys623ser62 mutant p50DNA binding (Matthewset al., 1992).
The purpose of the present studies was to conduct a deanalysis of the effects of Hg21 on NF-kB activation and thinteraction of the NF-kB binding with DNA, specifically with
em-
f-
-to
to
-
r
s--y-;
nen
n
glol.a
of
led
respect to the role of the critical cysteinyl moiety of thesubunit (cys62). Renal epithelial cells, a major target siteHg21 bioaccumulation and Hg21-induced tubular injury, weremployed as the experimental model for these studies. NkBwas activated using LPS, a bacterial endotoxin knowactivate NF-kB in a number of cell types, including kidnepithelial cells (Woodset al., 1999). DNA-binding studieconfirmed that the capacity of Hg21 to block NF-kB–DNA
inding was dependent largely, although not solely, onnteraction with the cys62 moiety of the p50 protein. Howeveadditional effects of Hg21 on NF-kB activation at the level oIkBa phosphorylation and degradation and subsequent NkBnuclear translocation also contribute significantly to H21-mediated attenuation of NF-kB-dependent transcriptional ativity in kidney cells.
MATERIALS AND METHODS
Materials. Lipopolysaccharide fromEscherichia coliserotype 026:B6dithiothreitol (DTT), deoxycholic acid, phenylmethylsulfonyl fluor(PMSF), mercuric chloride (HgCl2), sodium orthovanadate, Triton X-10
-mercaptoethanol (b-ME), L-cysteine (free base), glutathione (GSH) (fcid), glutathione agarose, human thrombin protease, aprotinin, alcohouffer salts were purchased from Sigma Chemical Co. (St. Louis, MO). Hnd 10% Nonidet P-40 were obtained from Boehringer Mannheim (Indilis, IN). Tris(2-carboxyethyl) phosphine hydrochloride (TCEP) was acq
rom Pierce (Rockford, IL). Poly(dI-dC)-poly(dI-dC), double stranded sodalt [poly(dI-dC)], calf thymus DNA, and CNBr-activated Sepharose 4ow were purchased from Pharmacia Biotech (Alameda, CA). Isopropylb-D-hiogalactopyranoside (IPTG) and ampicillin were obtained from Amersife Science Products (Arlington Heights, IL). LB broth was acquired fibco–BRL (Gaithersburg, MD). NF-kB (p65), IkBa, and actin polyclonantibodies and radioimmunoprecipitation assay (RIPA) lysis buffer werehased from Santa Cruz Biotechnology (Santa Cruz, CA). IkB-P polyclona
antibody was purchased from Cell Signaling Technology (Beverly, MPrecast, 10 cm2, 6% polyacrylamide gels prepared with 0.53 Tris–borate–
DTA (TBE) buffer, 53 TBE running buffer stock, 63 sample buffer stocwith bromophenyl blue and xylene cyanol dyes), PVDF paper, 4–20%lycine precast gels and Tris–glycine SDS running buffer were obtainedOVEX (San Diego, CA). Kaleidoscope prestained standards and blrade blocker were purchased from Bio-Rad Laboratories (Hercules,CL Western blotting detection reagent was acquired from Amershamacia (Piscataway, NJ).Polyacrylamide gel electrophoresis (PAGE) and Western transfers
erformed with an E19001-Xcell II Mini Cell and X Cell II Blot Moduurchased from NOVEX and driven by a model 250 electrophoresis pupply obtained from Gibco–BRL. Human recombinant p50 NF-kB1 (p50)as purchased from Promega Corporation (Madison, WI).E. coli JM101 withys623ser62 mutant p50 (p50MT)–GST fusion protein was generously providy Dr. Ronald Hay (University of Saint Andrews, Saint Andrews, Scotlarotein assays were performed using the BioRad DC protein assay pur
rom BioRad Laboratories. Molecular biology grade water and 10% soodecyl sulfate (SDS) solution were purchased from BioWhittaker (Walille, MA).
Cell culture and treatments. Normal rat kidney epithelial-type (NRK52ell stocks (ATCC CRL 1571) were acquired from the American Type Cuollection (Rockville, MD) and were propagated in Dulbecco’s modagle’s medium (DMEM) with high glucose, pyruvate, andL-glutamate (Bio
Whittaker) supplemented with 5% heat-inactivated newborn calf s(Gibco) plus 100 units/ml penicillin and 100mg/ml streptomycin (Sigma). Ce
onolayer mats were grown to confluence in 6- or 12-well plates (C

tedfl tod n
to9 rifugs sa
s-oo
wasthectioford ater
rolt cehatwew
cid,a and3 uget amw 0,00r was
aT nt p( ssa
fr as
ria
ec diup andm oola 5,0r affinc NTb al 1c h 1m erep sep tedt ubles diup ftew db cteS mnr as
seding
r
L say
TpN to
on of-type
a asm
eanrobese
( freen
hy.E ne usly
as).at
r idine,2T ing ofa l to
ubat dded
titiven
g
T ARerealyst
werehphe-ecastishedufac-
ional,
of aeam-
b athol-o oningE Y),
andbuffern to
ent kitomeser-
c MV( rter:r iationi wasi f thec 80%c wereh Lucif-e ometer
178 DIEGUEZ-ACUNA ET AL.
Corning, NY) and also in 25- and 75-cm2 cant-necked, vent-cap, uncoaasks (Becton Dickinson, Oxnard, CA). Medium was replaced every 2ays, irrespective of confluence. LPS treatment (1mg/ml of LPS 30 to 60 mi
before harvesting) to activate NF-kB was performed when cells covered 800% of flask surface. Nuclear extracts were prepared in microcentample tubes as described below from LPS-, Hg21-, and H2O-treated cell flasknd stored at280°C.
Preparation of nuclear and cytoplasmic extracts.Nuclear and cytoplamic extracts of NRK52E cells were prepared as previously described (Wet al., 1999; Dieguez-Acun˜a and Woods, 2000) except that 1 mM TCEPemployed in place of DTT as the reducing agent in buffers employed inprocedures and 1 M sodium orthovanadate was added to cytoplasmic fraFollowing centrifugation steps, a 5-ml aliquot of each extract was isolatedprotein determination. The remaining portion of each extract was removestored at280°C until used in electrophoretic mobility shift assays or Wesblot analysis.
Preparation of whole cell extracts. Medium was evacuated from contand experimentally treated NRK52E cell culture flasks. The still-adherenmats were gently and quickly washed twice in 5 ml of Dulbecco’s phospbuffered saline (PBS) (Gibco) at room temperature. All further operationsperformed on ice with chilled buffer. The washed cell mats were scraped0.6 ml RIPA lysis buffer (13 PBS, 1% Nonidet P-40, 0.5% deoxycholic a
nd 0.1% SDS) supplemented with 1 M PMSF and sodium orthovanadate0 ml of aprotinin per ml of RIPA. Cells were transferred into microcentrif
ubes and passed through a 23-gauge needle to sheer the DNA. The sere then incubated on ice for 30 to 60 min and microcentrifuged at 1
pm for 10 min at 4°C. The supernatant containing the total cell lysatetored in fresh microcentrifuge tubes at280°C after removing 5ml for BioRad
protein assay.
Preparation of recombinant p50. Human recombinant NF-kB p50 wasdiluted to 0.143 ng/ml using C/D (2:3) buffer and stored at280°C. Buffers C
nd D were prepared as described (Woodset al., 1999) except that 1 mMCEP was employed in place of DTT as the reducing agent. Recombina0.67 ng) was added directly to binding reactions prepared for gel shift a
Preparation of mutant (cys623ser62) recombinant p50. Purification oecombinant p50 cys623ser62 mutant GST fusion protein was performed
described by Matthewset al. (1992) with the following modifications:E. coliJM101 transformed with the pGEX derivatives containing the mutant vaof p50 were grown in 1 liter of L-broth base containing 50mg/ml ampicillin toan A600 of 0.7 and induced with 0.5 mM IPTG for 4 h at 25°C. Cells werollected by centrifugation, resuspended in 20 ml ice-cold 20 mM sohosphate buffer, pH 7.5, 0.5 M NaCl, 1 mM TCEP (PNT1 buffer),aintained at 4°C. Cells were disrupted by sonication, samples were pnd Triton X-100 was added to 1%. Samples were then centrifuged at 1pm for 20 min and supernatant was loaded onto a glutathione agaroseolumn (10 ml packed column) equilibrated with 10 column volumes of Puffer. After loading the sample, the column was washed with an additionolumn volumes of PNT1 buffer, and the fusion protein was eluted witM GSH, pH 7.5, in 50 mM Tris HCl and 0.5 M NaCl. Peak fractions wooled and fusion proteins were cleaved using 6 U human thrombin proteaer mg of fusion protein for 4 h at20°C. The NaCl concentration was dilu
o 0.25 M and proteins were applied to a column of calf thymus dotranded DNA Sepharose (2 ml packed volume) equilibrated in 20 mM sohosphate buffer, pH 7.0, 0.2 M NaCl, and 2 mM TCEP (PNT2 buffer). Aashing the column with PNT2 buffer, the bound p50MT protein was removey raising the NaCl concentration to 0.6 M and peak fraction was colleamples were then loaded onto a glutathione agarose affinity colu
emove uncleaved GST bound p50MT proteins. The protein concentration wthen determined using BioRad DC protein assay. Recombinant p50MT samplewere diluted to 0.144 ng/ml using “C/D” (2:3) nuclear extract buffer and storat 280°C. Recombinant p50MT (0.7 ng) was then added directly to bindeactions prepared for gel shift assays.
In vitro DNA-binding studies. Nuclear extracts prepared from H2O- orPS-treated cells provided sufficient protein for up to 4 nuclear binding as
5
e
ds
sens.
ndn
lle-reith
ples0s
50ys.
nt
m
ed,00ity
15
0
-mr
d.to
s.
hese studies evaluated the effects of either Hg21 alone or of Hg21 in theresence of competitive thiol reagents (DTT,b-ME, GSH, andL-cysteine) onF-kB-DNA binding. Hg21, b-ME, DTT, GSH, andL-cysteine were added
binding reactions at room temperature 10 to 15 min prior to the additi32P-labeled oligonucleotide. In assays involving recombinant p50 wildnd mutant proteins, samples were stored at280°C in aliquots prepared forany as 15in vitro assessments.
Oligonucleotide. An oligonucleotide containing the sensekB sequenc(59-AGT TGA GGG GAC TTT CCC AGG C-39) was purchased asannealed probe from IDT (Coralville, IA). The double-stranded oligopwas end-labeled with [g-P32]dATP (DuPont, Wilmington, DE) using T4 kinaBoehringer Mannheim). The radiolabeled probe was separated fromucleotide using a NucTrap push column (Stratagene, La Jolla, CA).
Electrophoretic mobility shift assays (EMSAs) and autoradiograpMSAs were used to demonstrate activation of NF-kB in nuclear proteixtracts from H2O- or LPS-treated cells and were performed as previo
described (Woodset al., 1999) with the exception that 1 mM TCEP wsubstituted for DTT in binding reactions (Dieguez-Acun˜a and Woods, 2000Recombinant p50WT and p50MT proteins were incubated for 30 to 40 minoom temperature in a solution containing 23.1 ng BSA, 3.6 mM sperm3 mM Hepes, 6.2 mM MgCl2, 8.6 mM KCl, 6% glycerol, 88 mM NaCl, 6 mMCEP HCl, and 0.14% Nonidet P-40. A signal-generating probe consist
32P-end-labeled NF-kB-binding double-stranded oligonucleotide (equa4 3 104 cpm/5ml) was used for all reactions. In studies requiring preinc-ion with Hg21 and/or thiol-directed agents, the radiolabeled probe was aafter 15–20 min with an additional incubation time of 15–20 min. Compethiol reagents were either added before or after Hg21 treatment depending othe study. Following electrophoresis in 0.53 TBE running buffer containin44.5 mM Tris base, 44.5 mM boric acid, and 1 mM EDTA at 4°C for 1.5W2 hat 90 V constant O/C, the gels were dried and exposed to Kodak X-OMAX-ray film with intensifying screens for up to 48 h. Autoradiograms wanalyzed using BioRad Gel Doc 1000 and the BioRad Molecular Anversion 2.1.1 software from BioRad Laboratories.
Western blot analysis. To quantitate p65, IkBa and phosphorylated IkBa
(IkB-P) protein levels, nuclear, cytoplasmic, and whole cell extractsprepared as described above. A total of 10–40mg of proteins were mixed wit53 sample buffer (312.5 mM Tris–HCl, 50% glycerol, and 0.05% bromonol blue) and resolved with SDS–PAGE using 4–20% Tris–glycine prgels and then transferred onto PVDF paper using the NOVEX publprotocol. The blots were visualized using ECL as described by the manturer. Selected blots were stripped with ChemiStrip (Chemicon InternatInc., Temecula, CA) and reblotted.
Transient transfections. Plasmids containing a construct composed43 tandem repeat of the NF-kB promoter response element inserted upstrof the coding region of a firefly (Photinus pyralis) luciferase gene in pGL2
asic were generously provided by Dr. Jean Campbell, Department of Pgy, University of Washington. The plasmids were propogated via Subclfficiency DH5a Competent Cells (Life Technologies, Grand Island, N
extracted with Endofree Plasmid Maxi Kit (Qiagen, Chatsworth, CA),evaluated with a 0.5% agarose–Tris buffer gel, eluted, and stored in TEat 4°C. NRK52E cells were seeded into 6- or 12-well plates and growapproximately 60% confluence. Using the Effectene Transfection Reagmanufacturer’s protocol (Qiagen, Valencia, CA), cotransfection liposwere formed with the NF-kB luciferase reporter plasmid and with a commially available constitutively active luciferase reference plasmid pRL-CRenilla reniformis,Promega) in a ratio of 1000:1 experimental repoeference reporter. The latter was employed to adjust for well-to-well varn cell number and transfection efficiency. The cotransfection mixturentroduced to the NKR52E cultures for 24 h followed by refreshment oulture medium. Cells were used in induction experiments at aboutonfluency 48–60 h following cotransfection. Experimental culturesarvested and assayed according to product instructions with the Dual-rase Reporter Assay System (Promega) and the Lumat LB9507 lumin
lize
waas
W nN tion
dod b
GS
prtoea),ea,edPS
dieof
of
p ct
p
thst
t yp5ss
LP
ionies
nvbuas
o . 2w 01
t.
er-
saythatd-ud-dndant.
b
in
c e (10d
gp anes4 . (B)B rcent-a as
ehiclet threealue
179Hg21 IMPAIRS NF-kB ACTIVATION AND DNA BINDING IN KIDNEY CELLS
(EG&G Berthold, Bundorra, Australia). Luminescence values were normato theRenilla reference signal.
Statistical analyses. Analysis of differences between treatment groupsdetermined using a paired, one-tailedt test. The level of significance wchosen atp , 0.05.
RESULTS
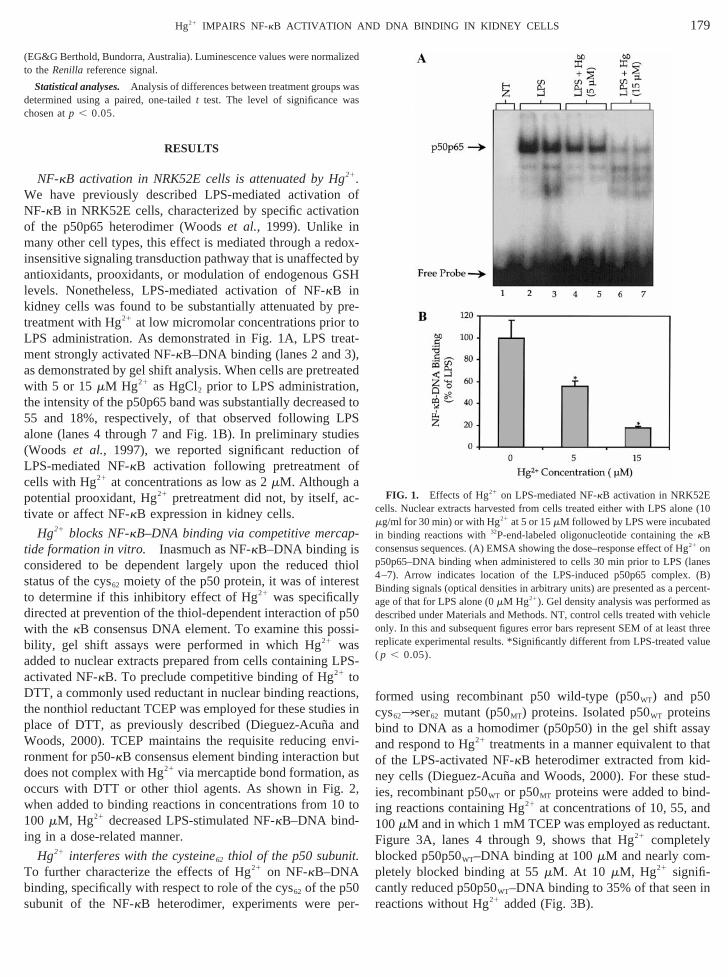
NF-kB activation in NRK52E cells is attenuated by Hg21.e have previously described LPS-mediated activatioF-kB in NRK52E cells, characterized by specific activa
of the p50p65 heterodimer (Woodset al., 1999). Unlike inmany other cell types, this effect is mediated through a reinsensitive signaling transduction pathway that is unaffecteantioxidants, prooxidants, or modulation of endogenouslevels. Nonetheless, LPS-mediated activation of NF-kB inkidney cells was found to be substantially attenuated bytreatment with Hg21 at low micromolar concentrations priorLPS administration. As demonstrated in Fig. 1A, LPS trment strongly activated NF-kB–DNA binding (lanes 2 and 3as demonstrated by gel shift analysis. When cells are pretrwith 5 or 15mM Hg21 as HgCl2 prior to LPS administrationthe intensity of the p50p65 band was substantially decreas55 and 18%, respectively, of that observed following Lalone (lanes 4 through 7 and Fig. 1B). In preliminary stu(Woods et al., 1997), we reported significant reductionLPS-mediated NF-kB activation following pretreatmentcells with Hg21 at concentrations as low as 2mM. Although a
otential prooxidant, Hg21 pretreatment did not, by itself, a-ivate or affect NF-kB expression in kidney cells.
Hg21 blocks NF-kB–DNA binding via competitive merca-tide formation in vitro. Inasmuch as NF-kB–DNA binding isconsidered to be dependent largely upon the reducedstatus of the cys62 moiety of the p50 protein, it was of intereo determine if this inhibitory effect of Hg21 was specificalldirected at prevention of the thiol-dependent interaction ofwith the kB consensus DNA element. To examine this pobility, gel shift assays were performed in which Hg21 wasadded to nuclear extracts prepared from cells containingactivated NF-kB. To preclude competitive binding of Hg21 toDTT, a commonly used reductant in nuclear binding reactthe nonthiol reductant TCEP was employed for these studplace of DTT, as previously described (Dieguez-Acun˜a andWoods, 2000). TCEP maintains the requisite reducing eronment for p50-kB consensus element binding interactiondoes not complex with Hg21 via mercaptide bond formation,
ccurs with DTT or other thiol agents. As shown in Fighen added to binding reactions in concentrations from 100 mM, Hg21 decreased LPS-stimulated NF-kB–DNA bind-
ing in a dose-related manner.
Hg21 interferes with the cysteine62 thiol of the p50 subuniTo further characterize the effects of Hg21 on NF-kB–DNAbinding, specifically with respect to role of the cys62 of the p50subunit of the NF-kB heterodimer, experiments were p
d
s
of
x-yH
e-
t-
ted
to
s
iol
0i-
S-
s,in
i-t
,to
formed using recombinant p50 wild-type (p50WT) and p50cys623ser62 mutant (p50MT) proteins. Isolated p50WT proteinsbind to DNA as a homodimer (p50p50) in the gel shift asand respond to Hg21 treatments in a manner equivalent toof the LPS-activated NF-kB heterodimer extracted from kiney cells (Dieguez-Acun˜a and Woods, 2000). For these sties, recombinant p50WT or p50MT proteins were added to bin-ing reactions containing Hg21 at concentrations of 10, 55, a100mM and in which 1 mM TCEP was employed as reductFigure 3A, lanes 4 through 9, shows that Hg21 completely
locked p50p50WT–DNA binding at 100mM and nearly com-pletely blocked binding at 55mM. At 10 mM, Hg21 signifi-cantly reduced p50p50WT–DNA binding to 35% of that seenreactions without Hg21 added (Fig. 3B).
FIG. 1. Effects of Hg21 on LPS-mediated NF-kB activation in NRK52Eells. Nuclear extracts harvested from cells treated either with LPS alon
mg/ml for 30 min) or with Hg21 at 5 or 15mM followed by LPS were incubatein binding reactions with32P-end-labeled oligonucleotide containing thekBconsensus sequences. (A) EMSA showing the dose–response effect of H21 on
50p65–DNA binding when administered to cells 30 min prior to LPS (l–7). Arrow indicates location of the LPS-induced p50p65 complexinding signals (optical densities in arbitrary units) are presented as a pege of that for LPS alone (0mM Hg21). Gel density analysis was performed
described under Materials and Methods. NT, control cells treated with vonly. In this and subsequent figures error bars represent SEM of at leasreplicate experimental results. *Significantly different from LPS-treated v( p , 0.05).
hegh
nesnti
stt.r th-
tioe
d t
d
a
Hgob-
nggts of
5 inn n inF
teins
b (1e
the-
antsas
rloca-
s intunder
gnal
win
oowsing
r LPi itrarud tho*
180 DIEGUEZ-ACUNA ET AL.
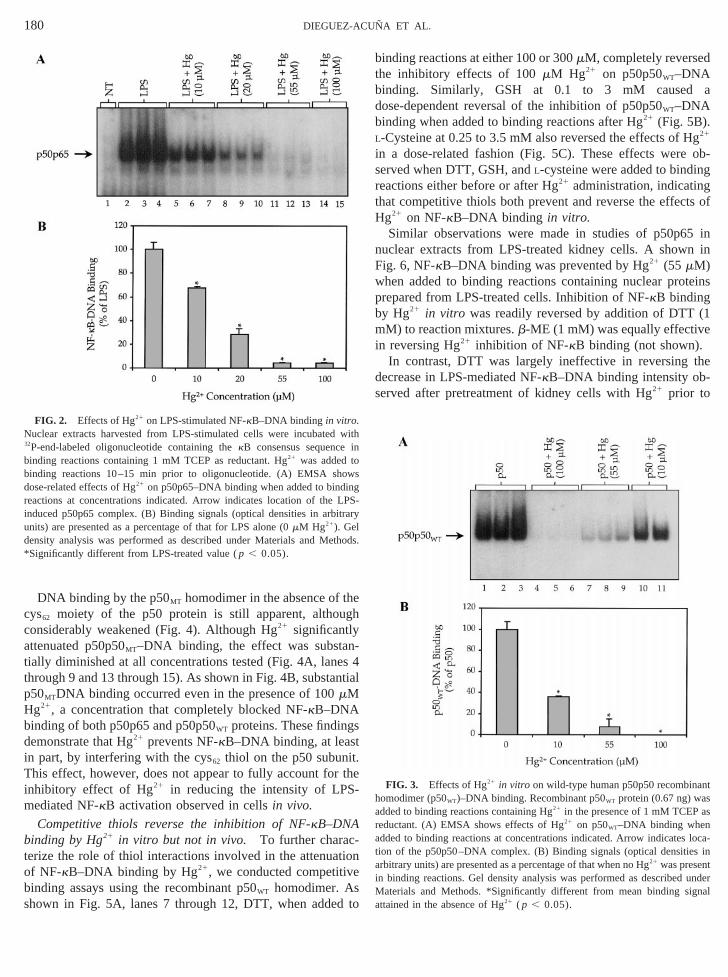
DNA binding by the p50MT homodimer in the absence of tcys62 moiety of the p50 protein is still apparent, althouconsiderably weakened (Fig. 4). Although Hg21 significantlyattenuated p50p50MT–DNA binding, the effect was substa-tially diminished at all concentrations tested (Fig. 4A, lanthrough 9 and 13 through 15). As shown in Fig. 4B, substap50MTDNA binding occurred even in the presence of 100mMHg21, a concentration that completely blocked NF-kB–DNAbinding of both p50p65 and p50p50WT proteins. These findingdemonstrate that Hg21 prevents NF-kB–DNA binding, at leasin part, by interfering with the cys62 thiol on the p50 subuniThis effect, however, does not appear to fully account foinhibitory effect of Hg21 in reducing the intensity of LPSmediated NF-kB activation observed in cellsin vivo.
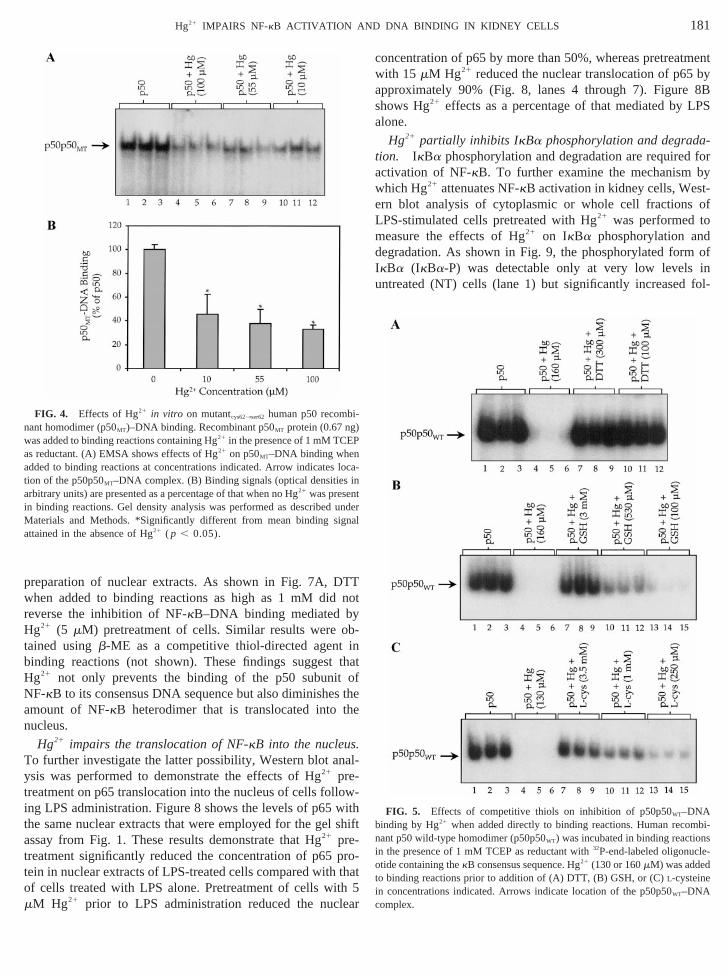
Competitive thiols reverse the inhibition of NF-kB–DNAbinding by Hg21 in vitro but not in vivo. To further charac-terize the role of thiol interactions involved in the attenuaof NF-kB–DNA binding by Hg21, we conducted competitivbinding assays using the recombinant p50WT homodimer. Asshown in Fig. 5A, lanes 7 through 12, DTT, when adde
FIG. 2. Effects of Hg21 on LPS-stimulated NF-kB–DNA binding in vitro.Nuclear extracts harvested from LPS-stimulated cells were incubated32P-end-labeled oligonucleotide containing thekB consensus sequencebinding reactions containing 1 mM TCEP as reductant. Hg21 was added tbinding reactions 10–15 min prior to oligonucleotide. (A) EMSA shdose-related effects of Hg21 on p50p65–DNA binding when added to bindeactions at concentrations indicated. Arrow indicates location of thenduced p50p65 complex. (B) Binding signals (optical densities in arbnits) are presented as a percentage of that for LPS alone (0mM Hg21). Gelensity analysis was performed as described under Materials and MeSignificantly different from LPS-treated value (p , 0.05).
4al
e
n
o
binding reactions at either 100 or 300mM, completely reversethe inhibitory effects of 100mM Hg21 on p50p50WT–DNAbinding. Similarly, GSH at 0.1 to 3 mM causeddose-dependent reversal of the inhibition of p50p50WT–DNAbinding when added to binding reactions after Hg21 (Fig. 5B).L-Cysteine at 0.25 to 3.5 mM also reversed the effects of21
in a dose-related fashion (Fig. 5C). These effects wereserved when DTT, GSH, andL-cysteine were added to bindireactions either before or after Hg21 administration, indicatinthat competitive thiols both prevent and reverse the effecHg21 on NF-kB–DNA binding in vitro.
Similar observations were made in studies of p50p6uclear extracts from LPS-treated kidney cells. A showig. 6, NF-kB–DNA binding was prevented by Hg21 (55 mM)
when added to binding reactions containing nuclear proprepared from LPS-treated cells. Inhibition of NF-kB binding
y Hg21 in vitro was readily reversed by addition of DTTmM) to reaction mixtures.b-ME (1 mM) was equally effectivin reversing Hg21 inhibition of NF-kB binding (not shown).
In contrast, DTT was largely ineffective in reversingdecrease in LPS-mediated NF-kB–DNA binding intensity observed after pretreatment of kidney cells with Hg21 prior to
FIG. 3. Effects of Hg21 in vitro on wild-type human p50p50 recombinhomodimer (p50WT)–DNA binding. Recombinant p50WT protein (0.67 ng) waadded to binding reactions containing Hg21 in the presence of 1 mM TCEPeductant. (A) EMSA shows effects of Hg21 on p50WT–DNA binding when
added to binding reactions at concentrations indicated. Arrow indicatestion of the p50p50–DNA complex. (B) Binding signals (optical densitiearbitrary units) are presented as a percentage of that when no Hg21 was presenin binding reactions. Gel density analysis was performed as describedMaterials and Methods. *Significantly different from mean binding siattained in the absence of Hg21 ( p , 0.05).
ith
S-y
ds.
TTn
yobintho
s tthe
.na
lowwitl sh
t prt tho th
lea
tmentby8B
LPS
at for
bytof
od
ofinfol-
mbin nsi e
dt
in )
P
locint
i unM gnaa
181Hg21 IMPAIRS NF-kB ACTIVATION AND DNA BINDING IN KIDNEY CELLS
preparation of nuclear extracts. As shown in Fig. 7A, Dwhen added to binding reactions as high as 1 mM didreverse the inhibition of NF-kB–DNA binding mediated bHg21 (5 mM) pretreatment of cells. Similar results were-tained usingb-ME as a competitive thiol-directed agentbinding reactions (not shown). These findings suggestHg21 not only prevents the binding of the p50 subunitNF-kB to its consensus DNA sequence but also diminisheamount of NF-kB heterodimer that is translocated intonucleus.
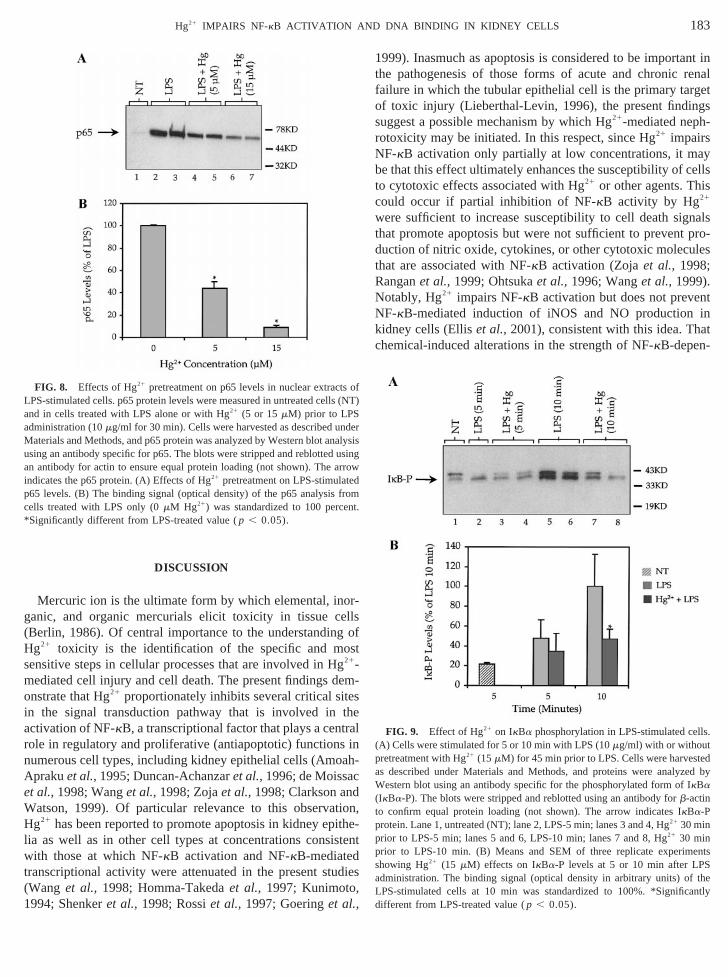
Hg21 impairs the translocation of NF-kB into the nucleusTo further investigate the latter possibility, Western blot aysis was performed to demonstrate the effects of Hg21 pre-treatment on p65 translocation into the nucleus of cells foling LPS administration. Figure 8 shows the levels of p65the same nuclear extracts that were employed for the geassay from Fig. 1. These results demonstrate that Hg21 pre-reatment significantly reduced the concentration of p65ein in nuclear extracts of LPS-treated cells compared withf cells treated with LPS alone. Pretreatment of cells wi
mM Hg21 prior to LPS administration reduced the nuc
FIG. 4. Effects of Hg21 in vitro on mutantcys623ser62 human p50 recomb-ant homodimer (p50MT)–DNA binding. Recombinant p50MT protein (0.67 ng
was added to binding reactions containing Hg21 in the presence of 1 mM TCEas reductant. (A) EMSA shows effects of Hg21 on p50MT–DNA binding whenadded to binding reactions at concentrations indicated. Arrow indicatestion of the p50p50MT–DNA complex. (B) Binding signals (optical densitiesarbitrary units) are presented as a percentage of that when no Hg21 was presenn binding reactions. Gel density analysis was performed as described
aterials and Methods. *Significantly different from mean binding sittained in the absence of Hg21 ( p , 0.05).
ot
atfhe
l-
-hift
o-at5r
concentration of p65 by more than 50%, whereas pretreawith 15 mM Hg21 reduced the nuclear translocation of p65approximately 90% (Fig. 8, lanes 4 through 7). Figureshows Hg21 effects as a percentage of that mediated byalone.
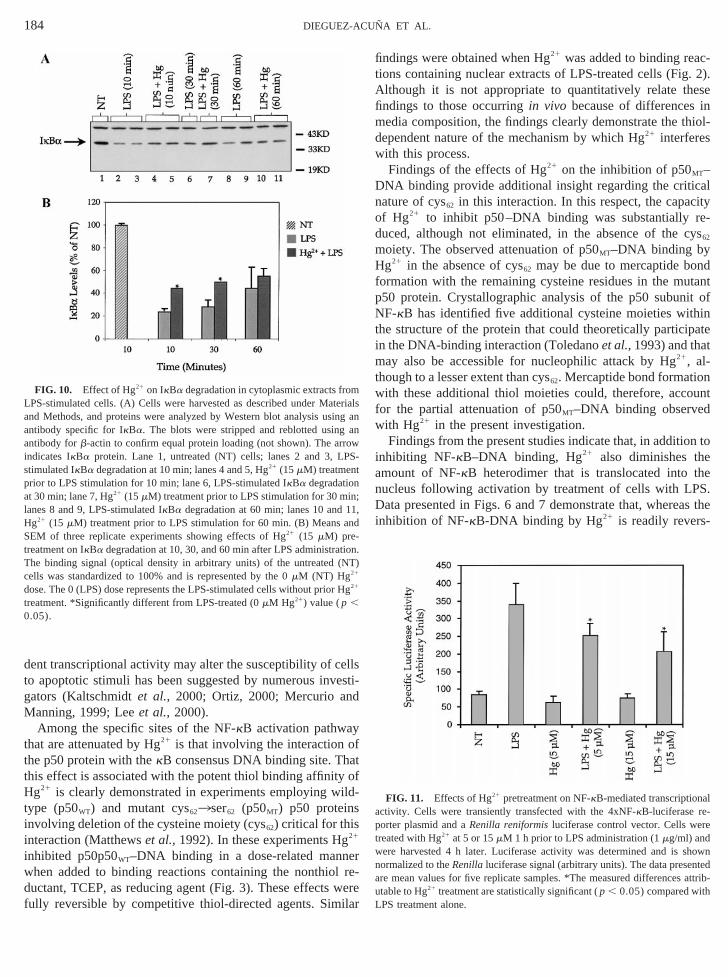
Hg21 partially inhibits IkBa phosphorylation and degrad-ion. IkBa phosphorylation and degradation are requiredactivation of NF-kB. To further examine the mechanismwhich Hg21 attenuates NF-kB activation in kidney cells, Wes-ern blot analysis of cytoplasmic or whole cell fractionsLPS-stimulated cells pretreated with Hg21 was performed tmeasure the effects of Hg21 on IkBa phosphorylation andegradation. As shown in Fig. 9, the phosphorylated formIkBa (IkBa-P) was detectable only at very low levelsuntreated (NT) cells (lane 1) but significantly increased
FIG. 5. Effects of competitive thiols on inhibition of p50p50WT–DNAbinding by Hg21 when added directly to binding reactions. Human reco-
ant p50 wild-type homodimer (p50p50WT) was incubated in binding reaction the presence of 1 mM TCEP as reductant with32P-end-labeled oligonucl-otide containing thekB consensus sequence. Hg21 (130 or 160mM) was addeo binding reactions prior to addition of (A) DTT, (B) GSH, or (C)L-cysteinein concentrations indicated. Arrows indicate location of the p50p50WT–DNAcomplex.
a-
derl
esonntrelledy
L gi-
e
m the
cmi-ith0,
ftees
ss nth
y.T
n-
n bygion
aseeat-
ithse26ity
tatehese
a ofN
ofted
f rei ss MSAs NAb .A , un-t tb –5 butp dings ted byt asd eanb
utheP
r too f tho f
s.A , unt LPs 7,
d 9.
182 DIEGUEZ-ACUNA ET AL.
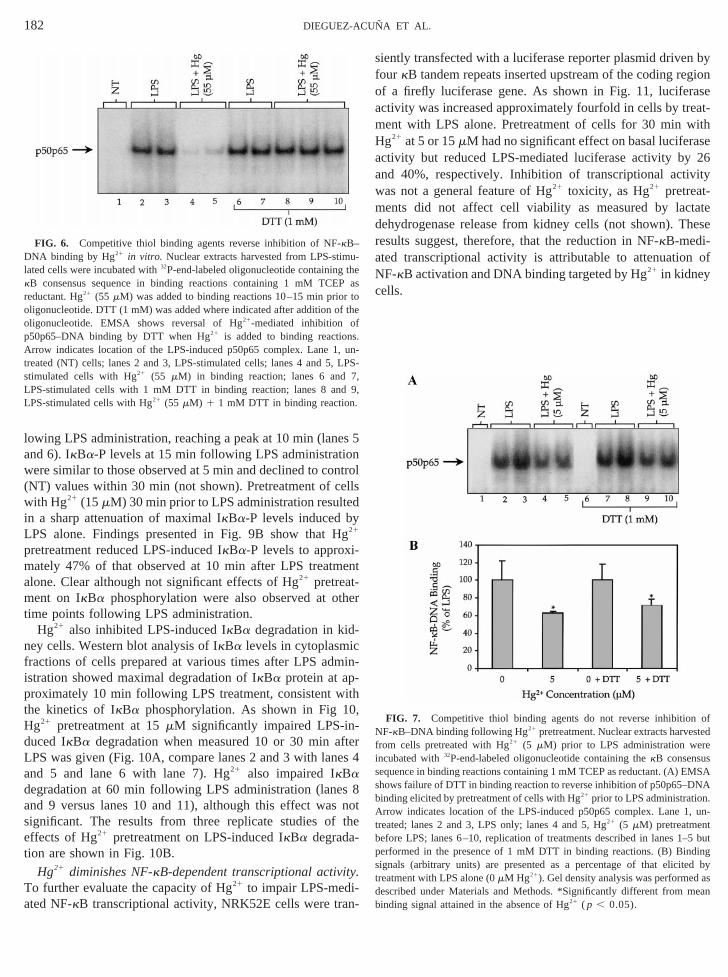
lowing LPS administration, reaching a peak at 10 min (lanand 6). IkBa-P levels at 15 min following LPS administratiwere similar to those observed at 5 min and declined to co(NT) values within 30 min (not shown). Pretreatment of cwith Hg21 (15mM) 30 min prior to LPS administration resultin a sharp attenuation of maximal IkBa-P levels induced b
PS alone. Findings presented in Fig. 9B show that H21
pretreatment reduced LPS-induced IkBa-P levels to approxmately 47% of that observed at 10 min after LPS treatmalone. Clear although not significant effects of Hg21 pretreat-
ent on IkBa phosphorylation were also observed at otime points following LPS administration.
Hg21 also inhibited LPS-induced IkBa degradation in kid-ney cells. Western blot analysis of IkBa levels in cytoplasmifractions of cells prepared at various times after LPS adistration showed maximal degradation of IkBa protein at approximately 10 min following LPS treatment, consistent wthe kinetics of IkBa phosphorylation. As shown in Fig 1Hg21 pretreatment at 15mM significantly impaired LPS-in-duced IkBa degradation when measured 10 or 30 min aLPS was given (Fig. 10A, compare lanes 2 and 3 with lanand 5 and lane 6 with lane 7). Hg21 also impaired IkBadegradation at 60 min following LPS administration (laneand 9 versus lanes 10 and 11), although this effect wasignificant. The results from three replicate studies ofeffects of Hg21 pretreatment on LPS-induced IkBa degrada-tion are shown in Fig. 10B.
Hg21 diminishes NF-kB-dependent transcriptional activito further evaluate the capacity of Hg21 to impair LPS-medi-
ated NF-kB transcriptional activity, NRK52E cells were tra
FIG. 6. Competitive thiol binding agents reverse inhibition of NF-kB–DNA binding by Hg21 in vitro. Nuclear extracts harvested from LPS-stim-lated cells were incubated with32P-end-labeled oligonucleotide containingkB consensus sequence in binding reactions containing 1 mM TCEreductant. Hg21 (55 mM) was added to binding reactions 10–15 min prio
ligonucleotide. DTT (1 mM) was added where indicated after addition oligonucleotide. EMSA shows reversal of Hg21-mediated inhibition o
p50p65–DNA binding by DTT when Hg21 is added to binding reactionrrow indicates location of the LPS-induced p50p65 complex. Lane 1
reated (NT) cells; lanes 2 and 3, LPS-stimulated cells; lanes 4 and 5,timulated cells with Hg21 (55 mM) in binding reaction; lanes 6 and
LPS-stimulated cells with 1 mM DTT in binding reaction; lanes 8 anLPS-stimulated cells with Hg21 (55 mM) 1 1 mM DTT in binding reaction
5
ols
nt
r
n-
r4
8ote
siently transfected with a luciferase reporter plasmid drivefour kB tandem repeats inserted upstream of the coding reof a firefly luciferase gene. As shown in Fig. 11, luciferactivity was increased approximately fourfold in cells by trment with LPS alone. Pretreatment of cells for 30 min wHg21 at 5 or 15mM had no significant effect on basal luciferaactivity but reduced LPS-mediated luciferase activity byand 40%, respectively. Inhibition of transcriptional activwas not a general feature of Hg21 toxicity, as Hg21 pretreat-ments did not affect cell viability as measured by lacdehydrogenase release from kidney cells (not shown). Tresults suggest, therefore, that the reduction in NF-kB-medi-
ted transcriptional activity is attributable to attenuationF-kB activation and DNA binding targeted by Hg21 in kidney
cells.
FIG. 7. Competitive thiol binding agents do not reverse inhibitionNF-kB–DNA binding following Hg21 pretreatment. Nuclear extracts harvesrom cells pretreated with Hg21 (5 mM) prior to LPS administration wencubated with32P-end-labeled oligonucleotide containing thekB consensuequence in binding reactions containing 1 mM TCEP as reductant. (A) Ehows failure of DTT in binding reaction to reverse inhibition of p50p65–Dinding elicited by pretreatment of cells with Hg21 prior to LPS administrationrrow indicates location of the LPS-induced p50p65 complex. Lane 1
reated; lanes 2 and 3, LPS only; lanes 4 and 5, Hg21 (5 mM) pretreatmenefore LPS; lanes 6–10, replication of treatments described in lanes 1erformed in the presence of 1 mM DTT in binding reactions. (B) Binignals (arbitrary units) are presented as a percentage of that elicireatment with LPS alone (0mM Hg21). Gel density analysis was performedescribed under Materials and Methods. *Significantly different from minding signal attained in the absence of Hg21 ( p , 0.05).
as
e
-S-
,
or-ellsg oostgemesthtras inahcdtioith
ste
di,
ant inrenalgetgs
aycellsis
nalspro-les
N ntinat
-
lls.(p ed
zed byI
p
entsS
a theL antlyd
s os (N
a ndealyusi
arroedfromnt.
*
183Hg21 IMPAIRS NF-kB ACTIVATION AND DNA BINDING IN KIDNEY CELLS
DISCUSSION
Mercuric ion is the ultimate form by which elemental, inganic, and organic mercurials elicit toxicity in tissue c(Berlin, 1986). Of central importance to the understandinHg21 toxicity is the identification of the specific and msensitive steps in cellular processes that are involved in H21-mediated cell injury and cell death. The present findings donstrate that Hg21 proportionately inhibits several critical sitin the signal transduction pathway that is involved inactivation of NF-kB, a transcriptional factor that plays a cenrole in regulatory and proliferative (antiapoptotic) functionnumerous cell types, including kidney epithelial cells (AmoAprakuet al.,1995; Duncan-Achanzaret al.,1996; de Moissaet al.,1998; Wanget al.,1998; Zojaet al.,1998; Clarkson anWatson, 1999). Of particular relevance to this observaHg21 has been reported to promote apoptosis in kidney ep-lia as well as in other cell types at concentrations consiwith those at which NF-kB activation and NF-kB-mediatedtranscriptional activity were attenuated in the present stu(Wang et al., 1998; Homma-Takedaet al., 1997; Kunimoto1994; Shenkeret al., 1998; Rossiet al., 1997; Goeringet al.,
FIG. 8. Effects of Hg21 pretreatment on p65 levels in nuclear extractLPS-stimulated cells. p65 protein levels were measured in untreated celland in cells treated with LPS alone or with Hg21 (5 or 15mM) prior to LPS
dministration (10mg/ml for 30 min). Cells were harvested as described uMaterials and Methods, and p65 protein was analyzed by Western blot anusing an antibody specific for p65. The blots were stripped and reblottedan antibody for actin to ensure equal protein loading (not shown). Theindicates the p65 protein. (A) Effects of Hg21 pretreatment on LPS-stimulatp65 levels. (B) The binding signal (optical density) of the p65 analysiscells treated with LPS only (0mM Hg21) was standardized to 100 perceSignificantly different from LPS-treated value (p , 0.05).
f
-
el
-
n,ent
es
1999). Inasmuch as apoptosis is considered to be importthe pathogenesis of those forms of acute and chronicfailure in which the tubular epithelial cell is the primary tarof toxic injury (Lieberthal-Levin, 1996), the present findinsuggest a possible mechanism by which Hg21-mediated neph-rotoxicity may be initiated. In this respect, since Hg21 impairsNF-kB activation only partially at low concentrations, it mbe that this effect ultimately enhances the susceptibility ofto cytotoxic effects associated with Hg21 or other agents. Thcould occur if partial inhibition of NF-kB activity by Hg21
were sufficient to increase susceptibility to cell death sigthat promote apoptosis but were not sufficient to preventduction of nitric oxide, cytokines, or other cytotoxic molecuthat are associated with NF-kB activation (Zojaet al., 1998;Ranganet al.,1999; Ohtsukaet al.,1996; Wanget al.,1999).
otably, Hg21 impairs NF-kB activation but does not preveNF-kB-mediated induction of iNOS and NO productionkidney cells (Elliset al.,2001), consistent with this idea. Thchemical-induced alterations in the strength of NF-kB-depen
FIG. 9. Effect of Hg21 on IkBa phosphorylation in LPS-stimulated ceA) Cells were stimulated for 5 or 10 min with LPS (10mg/ml) with or withoutretreatment with Hg21 (15mM) for 45 min prior to LPS. Cells were harvest
as described under Materials and Methods, and proteins were analyWestern blot using an antibody specific for the phosphorylated form ofkBa(IkBa-P). The blots were stripped and reblotted using an antibody forb-actinto confirm equal protein loading (not shown). The arrow indicates IkBa-P
rotein. Lane 1, untreated (NT); lane 2, LPS-5 min; lanes 3 and 4, Hg21 30 minprior to LPS-5 min; lanes 5 and 6, LPS-10 min; lanes 7 and 8, Hg21 30 minprior to LPS-10 min. (B) Means and SEM of three replicate experimshowing Hg21 (15 mM) effects on IkBa-P levels at 5 or 10 min after LP
dministration. The binding signal (optical density in arbitrary units) ofPS-stimulated cells at 10 min was standardized to 100%. *Significifferent from LPS-treated value (p , 0.05).
fT)
rsisngw
ellvesd
yfat
y oild
s
erl reweila
c. 2).eseinhiol-
calcitye
d cys
ndtantit ofhinpatet
nountd
n toetheS.
as the
ala -p re
w ownn nteda attrib-uL
omL teria inga an
rowPS
tnin;11,and
tionT (NTc
r Hg
184 DIEGUEZ-ACUNA ET AL.
dent transcriptional activity may alter the susceptibility of cto apoptotic stimuli has been suggested by numerous ingators (Kaltschmidtet al., 2000; Ortiz, 2000; Mercurio anManning, 1999; Leeet al., 2000).
Among the specific sites of the NF-kB activation pathwathat are attenuated by Hg21 is that involving the interaction othe p50 protein with thekB consensus DNA binding site. Ththis effect is associated with the potent thiol binding affinitHg21 is clearly demonstrated in experiments employing wtype (p50WT) and mutant cys623ser62 (p50MT) p50 proteininvolving deletion of the cysteine moiety (cys62) critical for thisinteraction (Matthewset al.,1992). In these experiments Hg21
inhibited p50p50WT–DNA binding in a dose-related mannwhen added to binding reactions containing the nonthioductant, TCEP, as reducing agent (Fig. 3). These effectsfully reversible by competitive thiol-directed agents. Sim
FIG. 10. Effect of Hg21 on IkBa degradation in cytoplasmic extracts frPS-stimulated cells. (A) Cells were harvested as described under Mand Methods, and proteins were analyzed by Western blot analysis usntibody specific for IkBa. The blots were stripped and reblotted using
antibody forb-actin to confirm equal protein loading (not shown). The arindicates IkBa protein. Lane 1, untreated (NT) cells; lanes 2 and 3, Lstimulated IkBa degradation at 10 min; lanes 4 and 5, Hg21 (15mM) treatmenprior to LPS stimulation for 10 min; lane 6, LPS-stimulated IkBa degradatioat 30 min; lane 7, Hg21 (15mM) treatment prior to LPS stimulation for 30 mlanes 8 and 9, LPS-stimulated IkBa degradation at 60 min; lanes 10 andHg21 (15 mM) treatment prior to LPS stimulation for 60 min. (B) MeansSEM of three replicate experiments showing effects of Hg21 (15 mM) pre-treatment on IkBa degradation at 10, 30, and 60 min after LPS administra
he binding signal (optical density in arbitrary units) of the untreatedells was standardized to 100% and is represented by the 0mM (NT) Hg21
dose. The 0 (LPS) dose represents the LPS-stimulated cells without prio21
treatment. *Significantly different from LPS-treated (0mM Hg21) value (p ,0.05).
sti-
f-
-re
r
findings were obtained when Hg21 was added to binding rea-tions containing nuclear extracts of LPS-treated cells (FigAlthough it is not appropriate to quantitatively relate thfindings to those occurringin vivo because of differencesmedia composition, the findings clearly demonstrate the tdependent nature of the mechanism by which Hg21 interfereswith this process.
Findings of the effects of Hg21 on the inhibition of p50MT–DNA binding provide additional insight regarding the critinature of cys62 in this interaction. In this respect, the capaof Hg21 to inhibit p50–DNA binding was substantially r-
uced, although not eliminated, in the absence of the62
moiety. The observed attenuation of p50MT–DNA binding byHg21 in the absence of cys62 may be due to mercaptide boformation with the remaining cysteine residues in the mup50 protein. Crystallographic analysis of the p50 subunNF-kB has identified five additional cysteine moieties witthe structure of the protein that could theoretically particiin the DNA-binding interaction (Toledanoet al.,1993) and thamay also be accessible for nucleophilic attack by Hg21, al-though to a lesser extent than cys62. Mercaptide bond formatiowith these additional thiol moieties could, therefore, accfor the partial attenuation of p50MT–DNA binding observewith Hg21 in the present investigation.
Findings from the present studies indicate that, in additioinhibiting NF-kB–DNA binding, Hg21 also diminishes thamount of NF-kB heterodimer that is translocated intonucleus following activation by treatment of cells with LPData presented in Figs. 6 and 7 demonstrate that, whereinhibition of NF-kB-DNA binding by Hg21 is readily revers-
FIG. 11. Effects of Hg21 pretreatment on NF-kB-mediated transcriptionctivity. Cells were transiently transfected with the 4xNF-kB-luciferase reorter plasmid and aRenilla reniformisluciferase control vector. Cells we
treated with Hg21 at 5 or 15mM 1 h prior to LPS administration (1mg/ml) andere harvested 4 h later. Luciferase activity was determined and is shormalized to theRenilla luciferase signal (arbitrary units). The data presere mean values for five replicate samples. *The measured differencestable to Hg21 treatment are statistically significant (p , 0.05)compared withPS treatment alone.
alsan
-
.)

e oPSg
p arthee
P ati. 8nt
wit
prelyratthqugsquere
hd s( athp
rouis
-
t ges of
n
ateIK
1 yby
p6med.ted). Aat
;L Td l
lysisfinedingstHg.f
theea-
Hgactiv-rved
ge-atinthe
tional
rved.F-are
elicitd
-arsf
acti-iol-
d and
-aykid-nt
rtialethe
these
entergramealthgtonfrom
185Hg21 IMPAIRS NF-kB ACTIVATION AND DNA BINDING IN KIDNEY CELLS
ible in vitro when added to binding reactions either beforafter addition of competitive thiols, the attenuation of Lmediated NF-kB–DNA binding that is observed followin
retreatment of cells with Hg21 prior to preparation of nucleextracts is not reversible. Hg21, therefore, appears to reducenuclear concentration of NF-kB available for binding to th
32-labeled consensus probe. Confirmation of this observis provided in the immunoblot analyses described in Figwhich demonstrate a dose-related reduction of p65 concetion in nuclear extracts from kidney cells that were treatedHg21 prior to LPS administration. Notably, NF-kB–DNA bind-ing activity, as measured in nuclear extracts from cellstreated with 15mM Hg21, was decreased to approximat20% of that mediated by LPS alone, whereas the concentof p65 detectable by immunoblot analysis in nuclei fromsame cells was decrased to less than 10%. A similar inewas observed at other Hg21 concentrations. These findinsuggest that step(s) involved in the activation and consenuclear translocation of the NF-kB heterodimer may be mosensitive or susceptible to impairment by Hg21 than NF-kB–DNA binding per se.
Phosphorylation of IkBa is the key point of control at whiciverse stimuli converge to activate NF-kB in many cell typee.g., Karin, 1999). Of note in the present studies was the rronounced inhibitory effect of Hg21 on maximal phosphory-
lation of IkBa induced by LPS. The effects of Hg21 on IkBaphosphorylation at treatment concentrations as low as 2mMare consistent with evidence that a metal-sensitive thiol gexists in the IkB kinase (IKK), the enzyme complex thatconsidered to be principally responsible for IkBa phosphorylation (Didonatoet al.,1997; Regnieret al.,1997). Jeonet al.(2000) have recently reported that Hg21 inhibited IKK activityin RAW 264.7 cells in vitro with an ID50 of 12.3 mM.However, the effects of Hg21 treatment on NF-kB activation inhat cell line were not reported. The present findings sugimilar inhibition kinetics of Hg21 with respect to prevention
IkBa phosphorylation in kidney cells.Hg21 was also effective in preventing IkBa degradatio
(Fig. 10). These findings imply a possible effect of Hg21 at thelevel of the ubiquitin–proteasome pathway, which mediIkBa degradation subsequent to phosphorylation by thecomplex (Chenet al.,1995; Schereret al.,1995; Alkalayet al.,
995). Since release of the activated NF-kB heterodimer marequire IkB degradation, inhibition of the latter processHg21 could account for the diminished concentration ofobserved. The specific mechanisms involved in the impairof IkBa degradation by Hg21 have not as yet been identifieDirect inhibition of the proteasome by other thiol-direcagents has been reported (Klemperer and Pickart, 1989ternatively, Hg21 impairs mitochondrial ATP productioncellular concentrations comparable to those that inhibit IkBadegradation in the present studies (Weinberget al., 1982a
undet al.,1993). Inasmuch as the proteasome activity is Aependent, it is possible that Hg21-mediated reduction of ce-
r-
on,
ra-h
e-
ioneity
nt
er
p
st
sK
5nt
l-
P
lular ATP levels could contribute to the decreased proteoof IkBa observed. While further studies are required to dethe specific mechanisms involved in this process, the finthat Hg21 substantially impairs IkBa degradation at the loweconcentrations tested may define a preferential effect of21
in the control of NF-kB activation in kidney epithelial cellsFunctional consequences of Hg21-mediated impairment o
NF-kB activation and DNA binding are strongly implied bydecrease in NF-kB-mediated transcriptional activity as msured in cells transiently transfected with akB-driven lucif-erase reporter gene construct (Fig. 11). In these studies,21
pretreatment of cells attenuated LPS-induced luciferaseity in a dose-related manner, consistent with the obsedecrease in p65 protein and in NF-kB–DNA binding affinity.While these effects may not be directly indicative of endonous transcriptional activity because of changes in chrompackaging or other effects that may be associated withtransfection process, they, nonetheless, support a funcassociation between the inhibitory effects of Hg21 on NF-kBactivation and the decrease in transcriptional activity obseNotably, moreover, the Hg21 concentrations that impaired NkB-mediated transcriptional activity in the present studieshighly consistent with those that have been shown toother early/reversible intracellular effects of Hg21-associatenephrotoxicity in situ (Weinberget al., 1982b; Lundet al.,1993).
In conclusion, the present findings demonstrate that Hg21, apotent nephrotoxicant, attenuates NF-kB activation and transactivational activity in renal epithelial cells. This effect appeto be mediated through the actions of Hg21 at multiple sites othe NF-kB activation pathway, most predominantly, IkBadegradation and subsequent nuclear translocation of thevated p50p65 heterodimer and p50-DNA binding. The thdependent nature of the latter effect is clearly demonstratelikely is related to the other steps affected by Hg21 as well. Thetoxicological consequences of attenuation of NF-kB transactivational activity by Hg21 are currently unresolved but minclude apoptosis, an important pathogenic mechanism inney cells that is elicited by Hg21 at concentrations consistewith those that are shown to impair NF-kB activation in thepresent studies. More intriguing is the possibility that paimpairment of NF-kB activation by Hg21 may alter the balancof NF-kB-mediated functional processes, enhancingsusceptibility of kidney cells to toxicity or cell death by Hg21
or other chemicals. Studies are in progress to investigatepossibilities.
ACKNOWLEDGMENTS
This research was supported in part by University of Washington CGrant P30 ES07033 and by University of Washington Superfund ProProject Grant ES04696 from the National Institute of Environmental HSciences, NIH. F.J.D.-A. was supported by the University of WashinEnvironmental Pathology/Toxicology training program (5 T32 ES07032)the NIEHS.

ene
A ngeal
B -
.s.),
s initi-
C d, Dta
.
e a
C n
d L.a-.
D , Mion
Dxy--
D airture.
E iblector
E ,NO
nde-
ide
G dof th
H 997d b
J un
, W
K
K s in
K n of
motifity
ellar
, andh
d its
cedney
saline
,
M . T.
andwth
e
M o, K.ding
lear
, M.,ted
ber of
O 1996)..
O nal
R ition
186 DIEGUEZ-ACUNA ET AL.
REFERENCES
Alkalay, I., Yaron, A., Hatsubai, A., Orian, A., Ciechanover, A., and BNeriah, Y. (1995). Stimulation-dependent IkBa phosphorylation marks th-kB inhibitor for degradation via the ubiquitin–proteasome pathway.Proc.Natl. Acad. Sci. USA92, 10599–10603.
moah-Apraku, B., Chandler, L. J., Harrison, J. K., Tang, S-S., IngelfiJ. R., and Guzman, N. J. (1995). NF-kB and transcriptional control of renepithelial-inducible nitric oxide synthase.Kidney Int.48, 674–682.
aeuerle, P. A., and Baltimore, D. (1989). A 65-kDa subunit of active NFkBis required for inhibition of NF-kB by IkB. Genes Dev.3, 1689–1698.
Berlin, M. (1986). Mercury. InHandbook on the Toxicology of Metals,2nd edVol. 2, Specific Metals.(L. Friberg, G. F. Nordberg, and C. Nordman, EdAmsterdam, Elsevier, pp. 386–445.
Chen, F., Castranova, V., Shi, X., and Demers, L. M. (1999). New insightthe role of nuclear factor-kB, a ubiquitous transcription factor in the ination of diseases.Clin. Chem.45, 7–17.
hen, Z., Hagler, J., Palombella, V. J., Meldandri, F., Scherer, D., Ballarand Maniatis, T. (1995). Signal-induced site-specific phosphorylationgets IkB to the ubiquitin–proteasome pathway.Genes Dev.9, 1586–1597
Chevez, E., and Hoguin, J. A. (1988). Mitochondrial calcium releasinduced by Hg21. J. Biol. Chem.263,3582–3587.
larkson, R. W. E., and Watson, C. J. (1999). NF-kB and apoptosis imammary epithelial cells.J. Mam. Gland Biol. Neoplasm4, 165–175.
e Moissac, D., Mustapha, S., Greenberg, A. H., and Kirshenbaum,(1998). Bcl-2 activates the transcription factor NF-kB through the degradtion of the cytoplasmic inhibitor IkBa. J. Biol. Chem.273,23946–23951
idonato, J. A., Hayakawa, M., Rothwaft, D. M., Zandi, E., and Karin(1997). A cytokine-responsive IkB kinase that activates the transcriptfactor NF-kB. Nature388,548–554.
ieguez-Acun˜a, F. J., and Woods, J. S. (2000). Inhibition of NF-kB-DNAbinding by mercuric ion: Utility of the non-thiol reductant, tris(2-carboethyl)phosphine hydrochloride (TCEP), on detection of impaired NFkB–DNA binding by thiol-directed agents.Toxicol. In Vitro14, 7–16.
uncan-Achanzar, K. B., Jones, J. T., Burke, M. F., Carter, D. E., and LH. E. (1996). Inorganic mercury chloride-induced apoptosis in the culporcine renal cell line LLC-PK1.J. Pharmacol. Exp. Ther.277,1726–1732
ckhardt, W., Bellmann, K., and Kolb, H. (1999). Regulation of inducnitric oxide synthase expression in beta cells by environmental faHeavy metals.Biochem. J.338,695–700.
llis, M. E., Corral, J., Kushleika, J., Simmonds, P. L., Dieguez-Acun˜a, F. J.and Woods, J. S. (2001). Mercuric ion induces nitric oxide synthetase (independently of NF-kB activation in rat kidney (NRK52E) cells.Toxicol-ogist 60, 211.
Fujita, T., Nolan, G. P., Ghosh, S., and Baltimore, D. (1992). Indepemodes of transcriptional activation by the p50 and p65 subunits of NFkB.Genes Dev.6, 775–787.
Goering, P. L., Rojko, J. L., and Lucas, A. D. (1999). Mercuric chlorinduced apoptosis is dependent on protein synthesis.Toxicol. Lett.105,183–195.
ratzka, T. L., and Trump, B. F. (1968). Renal tubular lesions causemercuric chloride. Electron microscopic observations: Degenerationpars recta.Am. J. Pathol.52, 1225–1277.
omma-Takeda, S., Takenaka, Y., Kumagai, Y., and Shimojo, N. (1Selective induction of apoptosis of renal proximal tubular cells causemercury in vivo.Environ. Toxicol. Pharmacol.7, 179–187.
eon, K-I., Jeong, J-Y., and Jue, D-M. (2000). Thiol-reactive metal compoinhibit NF-kB activation by blocking IkB kinase.J. Immunol.164, 5981–5989.
Kaltschmidt, B., Kaltschmidt, C., Hofmann, T. G., Hehner, S. P., Durge
-
r,
to
.,r-
s
A.
.
d,d
s:
S)
nt
-
bye
).y
ds
.,
and Schmidt, M. L. (2000). The pro- or anti-apoptotic function of NF-kB isdetermined by the nature of the apoptotic stimulus.Eur. J. Biochem.267,3828–3835.
arin, M. (1999). How NF-kB is activated: The role of the IkB kinase (IKK)complex.Oncogene18, 6867–6874.
lemperer, N. S., and Pickart, C. M. (1989). Arsenite inhibits two stepthe ubiquitin-dependent proteolytic pathway.J. Biol. Chem.264,19245–19252.
retz-Reny, C., Mehlen, P., Mirault, P., and Arrigo, A. P. (1996). InhibitioIkB-a phosphorylation and degradation and subsequent NF-kB activationby glutathione peroxidase overexpression.J. Cell Biol. 133,1083–1093.
Kumar, S., Rabson, A. B., and Gelinas, C. (1992). The RxxRxRxxCconserved in all Rel/k B proteins is essential for the DNA-binding activand redox regulation of the v-Rel oncoprotein.Mol. Cell. Biol. 12, 3094–3106.
Kunimoto, M. (1994). Methylmercury induces apoptosis of rat cerebneurons in primary culture.Biochem. Biophys. Res. Commun.204, 310–317.
Lee, E. G., Boone, D. L., Chai, S., Libby, S. L., Chien, M., Lodolce, J. P.Ma, A. (2000). Failure to regulate TNF-induced NF-kB and cell deatresponses in A20-deficient mice.Science289,2350–2354.
Lieberthal, W., and Levin, J. S. (1996). Mechanisms of apoptosis anpotential role in renal tubular epithelial cell injury.Am. J. Physiol.271,F477–F488.
Lund, B.-O., Miller, D. M., and Woods, J. S. (1993). Studies on Hg(II)-induH2O2 formation and oxidative stress in vivo and in vitro in rat kidmitochondria.Biochem. Pharmacol.45, 2017–2024.
Magos, L., Sparrow, S., and Snowden, R. T. (1984). Effect of prolongedloading on HgCl2-induced renal tubular damage.Br. J. Exp. Pathol.65,567–575.
Martin, R. B. (1986). Bioinorganic chemistry of metal ion toxicity. InMetalIons in Biological Systems Concepts in Metal Ion Toxicity(H. Sigel, Ed.)Vol. 20, pp. 21–66. Dekker, New York.
atthews, J. R., Wakasugi, N., Virelizier, J. L., Yodoi, J., and Hay, R(1992). Thioredoxin regulates the DNA binding activity of NF-kB byreduction of a disulphide bond involving cysteine 62.Nucleic Acids Res.20,3821–3830.
McCabe, M. J., Jr., Santini, R. P., and Rosenspire, A. J. (1999). Lownontoxic levels of ionic mercury interfere with the regulation of cell groin the WEHI-231 B-cell lymphoma.Scand. J. Immunol.50, 233–241.
Mercurio, F., and Manning, A. M. (1999). NF-kB as a primary regulator of thstress response.Oncogene18, 6163–6171.
itomo, K., Nakayama, K., Fujimoto, K., Sun, X., Seki, S., and Yamamot(1994). Two different cellular redox systems regulate the DNA-binactivity of the p50 subunit of NF-kB in vitro. Gene145,197–203.
Muller, J. M., Ziegler-Heitbrock, J. H. W., and Baeuerle, P. A. (1993). Nucfactor kB, a mediator of lipopolysaccharide effects.Immunolbiology187,233–256.
Ohtsuka, T., Kubota, A., Hirano, T., Watanabe, K., Yoshida, H., TsurufujiIizuka, Y., Konishi, K., and Tsurufuji, S. (1996). Glucocorticoid-mediagene suppression of rat cytokine-induced neutrophil CINC/gro, a memthe interleukin-8 family, through impairment of NF-kB activation.J. Biol.Chem.271,1651–1659.
ram, P. D., Fang, X., Fernando, Q., Letkeman, P., and Letkeman, D. (The formation of constants of mercury(II)-glutathione complexes.ChemRes. Toxicol.9, 709–712.
rtiz, A. (2000). Nephrology forum: Apoptotic regulatory proteins in reinjury. Kidney Int.58, 467–485.
angan, G. K., Wang, Y., Tay, Y.-C., and Harris, D. C. H. (1999). Inhib

cyto
R e, M
R t. 1
R M.,s.
S d, Dti-
S g bl.
etein
mernce
n onite
B. Fl
al.
S an
S on
ove-
S 990).n
T inalties
actor.
W in,nd
99).ed by
drialjury.nc-
drialjury.mer-
ventsby
W 9).is
Z M.,ction
187Hg21 IMPAIRS NF-kB ACTIVATION AND DNA BINDING IN KIDNEY CELLS
of NFkappaB activation with antioxidants is correlated with reducedkine transcription in PTC.Am. J. Physiol.277,F779–F789.
egnier, C. H., Song, H. Y., Gao, X., Goeddel, D. V., Cao, Z., and Roth(1997). Identification and characterization of an IkB kinase.Cell 90, 373–383.
odin, A. E., and Crowson, C. N. (1962). Mercury nephrotoxicity in the raFactors influencing the localization of the tubular lesions.Am. J. Pathol.41,297–313.
ossi, A. D., Vivani, B., Zhivotovsky, B., Manzo, L., Orrenius, S., Vahter,and Nicotera, P. (1997). Inorganic mercury modifies Ca21 signals, triggerapoptosis and potentiates NMDA toxicity in cerebellar granule neuronsCellDeath Diff.4, 317–324.
cherer, D. C., Brockman, A. J., Chen, Z. J., Maniatis, T., and Ballar(1995). Signal-induced degradation of IkBa requires site-specific ubiquination.Proc. Natl. Acad. Sci. USA92, 11259–11263.
chulze-Osthoff, K., Los, M., and Baeuerle, P. A. (1995). Redox signallintranscription factors NF-kB and AP-1 in lymphocytes.Biochem. Pharmaco50, 735–741.
Sha, W. C., Liou, H. C., Toumanen, E. I., and Baltimore, D. (1995). Targdisruption of the p50 subunit of NF-kB leads to multifocal defectsimmune responses.Cell 80, 321–330.
Shenker, B. J., Guo, T. L., and Shapiro, I. M. (1998). Low-level methylcury exposure causes human T-cells to undergo apoptosis: Evidemitochondrial dysfunction.Environ. Res.77, 149–159.
Shumilla, J. A., Wetterhahn, K. E., and Barchowsky, A. (1998). InhibitioNF-kB binding to DNA by chromium, cadmium, mercury, zinc, and arsein vitro: Evidence of a thiol mechanism.Arch. Biochem. Biophys.349,356–362.
Smith, M. W., Ambudkar, I. S., Phelps, P. C., Regec, A. L., and Trump,(1987). HgCl2-induced changes in cytosolic Ca21 of cultured rabbit renatubular cells.Biochim. Biophys. Acta931,130–142.
Smith, M. W., Phelps, P. C., and Trump, B. F. (1991). Cytosolic C21
deregulation and blebbing after HgCl2 injury to cultured rabbit proximatubule cells as determined by digital imaging microscopy.Proc. Natl. AcadSci. USA88, 4926–4930.
outhard, J., Nitisewojo, P., and Green, D. E. (1974). Mercurial toxicitythe perturbation of the mitochondrial control system.Fed. Proc.33, 2147–2153.
outhard, J. H., Penniston, J. T., and Green, D. E. (1973). Inducti
-
.
.
.
y
d
-of
f
.
d
of
transmembrane proton transfer by mercurials in mitochondria. I. Ion mments accompanying transmembrane proton transfer.J. Biol. Chem.248,3546–3550.
taal, F. J. T., Roederer, M., Herzenberg, L. A., and Herzenberg, L. A. (1Intracellular thiols regulate activation of nuclear factorkB and transcriptioof human immunodeficiency virus.Proc. Natl. Acad. Sci. USA87, 9943–9947.
oledano, M. B., Ghosh, D., Trinh, F., and Leonard, W. J. (1993). N-termDNA-binding domains contribute to differential DNA-binding specificiof NF-kB p50 and p65.Mol. Cell. Biol. 13, 852–860.
Toledano, M. B., and Leonard, W. J. (1991). Modulation of transcription fNF-kB binding activity by oxidation-reduction in vitro.Proc. Natl. AcadSci. USA88, 4328–4332.
ang, C. Y., Mayo, M. W., Korneluk, R. G., Goeddel, D. V., and BaldwA. S., Jr. (1998). NF-kB antiapoptosis: Induction of TRAF1 and TRAF2 ac-IAP1 and c-IAP2 to suppress caspase-8 activation.Science281, 1680–1683.
Wang, Y., Rangan, G. K., Tay, Y. C., Wang, Y., and Harris, D. C. H. (19Induction of monocyte chemoattractant protein-1 by albumin is mediatnuclear factorkB in proximal tubule cells.J. Am. Soc. Nephrol.10, 1204–1213.
Weinberg, J. M., Harding, P. G., and Humes, H. D. (1982a). Mitochonbioenergetics during the initiation of mercuric chloride-induced renal inI. Direct effects of in vitro mercuric chloride on renal mitochondrial fution. J. Biol. Chem.257,60–67.
Weinberg, J. M., Harding, P. G., and Humes, H. D. (1982b). Mitochonbioenergetics during the initiation of mercuric chloride-induced renal inII. Functional alterations of renal cortical mitochondria isolated aftercuric chloride treatment.J. Biol. Chem.257,68–74.
Woods, J. S., Dieguez, F. J., and Ellis, M. E. (1997). Mercuric ion preexpression of NF-kB in normal rat kidney epithelial (NRK52E) cellsblocking binding to DNA.Toxicologist36, 293.
oods, J. S., Ellis, M. E., Dieguez-Acun˜a, F. J., and Corral, J. (199Activation of NF-kB in normal rat kidney epithelial (NRK52E) cellsmediated via a redox-insensitive, calcium-dependent pathway.Toxicol.Appl. Pharmacol.154,219–227.
oja, C., Donaldelli, R., Colleoni, S., Figliuzzi, M., Bonazzola, S., Morigi,and Remuzzi, G. (1998). Protein overload stimulates RANTES produby proximal tubular cells depending on NF-kB activation.Kidney Int.53,1608–1615.




















