
memoire.pdf - Dspace
139
قسملبيوكيمياء ا أطروحة مقدمة من طرف جرمونيريم م على شهادةحصولل ل دكتـوراه عل ـ وم ف ـ رع: بي ـ ول ـ وجي ـ ا تخص ـ ص: بيوكيمياء الموضوعكسدة للمضاداسة التأثير ا در نبتتيخلصات لمستحرمل الPeganum harmala و الجعدةSantolina chamaecyparissus بت نوقشت اريخ/.........../.......... 4102 لمناقشـةم لجنة ا أما الرئ ـ يسة دحامنة صليح أستاذ ة جامعة سطيف1 ش ا ـ رفان بغيا عبدالر أستاذ جامعة سطيف1 متحن ا ـ ون لروي صا أستاذ جامعة باتنةي مولود أستاذ جامعة باتنة بشرة ختال أستاذ ة ناضرة( أ) جامعة بايةدعو ا عارسي عر نمي أستاذ جامعة سطيف1 مخبرلبيوكيمياء ا التطبيقيةUniversité Ferhat Abbas Sétif 1 Faculté des Sciences de la Nature et de la Vie الجمهورية الجزائرية الديمقراطية الشعبيةعلملي و البحث اللعاتعليم ا وزارة ال ي، سطيفة فرحات عباس جامع1 كلية علوملحياة الطبيعة و اN°………………………………………….…………..…….……/SNV/2014
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of memoire.pdf - Dspace

البيوكيمياء قسم أطروحة
مقدمة من طرف
مريم جرموني
للحصول على شهادة
ومـدكتـوراه عل
اـوجيـولـبي: رعـف بيوكيمياء: صـتخص
الموضوع
Peganum harmala الحرمل لمستخلصات نبتتي دراسة التأثير المضاد لألكسدة
Santolina chamaecyparissus الجعدةو
4102........../.........../ اريخنوقشت بت
أمام لجنة المناقشـة
1 جامعة سطيف ةأستاذ صليحة دحامنة يس ـالرئ 1 جامعة سطيف أستاذ عبدالرمحان بغياين رف ـاملش
باتنةجامعة أستاذ صاحل لروي ون ـاملمتحن باتنةجامعة أستاذ مولود حيي جبايةجامعة ( أ)ة حماضرة أستاذ بشرة ختال
1 جامعة سطيف أستاذ خلميسي عرعار املدعو
التطبيقية البيوكيمياء مخبر
Université Ferhat Abbas Sétif 1
Faculté des Sciences de la
Nature et de la Vie
الجزائرية الديمقراطية الشعبيةالجمهورية
يوزارة التعليم العالي و البحث العلم
1 جامعة فرحات عباس، سطيف الطبيعة و الحياةعلوم كلية
N°………………………………………….…………..…….……/SNV/2014

ـــــــــــــراتتشكـــــ
أمحد اهلل سبحانه جل شأنه وأشكره جزيل الشكر على إعانته الكبرية يل يف كل ميادين .حيايت ويف إجناز وإمتام حبثي هذا
أتوجه خبالص تشكرايت لألستاذ الفاضل بغياين عبد الرمحان على إشرافه وتوجيهه ومساعدته .ونصائحه خالل مراحل إجناز هذا البحث
األستاذة دحامنة صليحة أستاذ جبامعة : جبزيل الشكر إىل أعضاء جلنة املناقشةكما أتقدم أن املناقشة و األستاذ لروي صاحل أستاذ جبامعة باتنة لقبوله جلنة قبوهلا ترأس على 1سطيف
يف عضوا يكون أن على قبوله باتنةاملناقشة واألستاذ مولود حيي جبامعة جلنة يف يكون عضوافلهم واألستاذة ختال بشرة جبامعة جباية على قبوهلا كعضو يف جلنة املناقشة، ،املناقشة جلنة
.مين كل التقدير واالحرتام
أتقدم بتشكرايت اخلالصة إىل األستاذ عرعار خلميسي على مساعدته ودعمه خالل مراحل .إجناز هذا البحث
مساعدهتا على بومرفق صباح لألستاذةكما أتقدم بالشكر اجلزيل .ائحها الدائمةونص
لألستاذ خنوف الصديق على استقبايل يف خمربه اجلزيل بالشكر أتقدم أن يفوتين ال كما .مبخربه تربصي فرتة أثناءبكل رحب وسعة
كما أتقدم بشكري إىل كل من األستاذ لعور حسني على التصنيف العلمي للنبتتني و
.ة ومساحتهااألستاذة بوريش محامة على نصائحها ومعاملتها الطيب

اإلهداء
ي العزيزيناإىل والد
ويتخأإىل
نييإىل زوجي وولدي الغال
إىل كل العائلة
األصدقاء مجيعإىل
ز هذا العملإىل كل من ساعدين يف إجنا

المنشورات العلمية
M. Djarmounj, S. Boumerfeg, A. Baghiani, N. Boussoualim, F. Zerargui, H. Trabsa, F.
Belkhiri, S. Khennouf and L. Arrar (2012). Evaluation of antioxidant and antibacterial
properties of Peaganum harmala seed extracts. Research Journal of Pharmaceutical,
Biological and Chemical Sciences. Vol.3(4); p: 1109-1119. www.rjpbcs.com
A. Baghiani*, M. Djarmouni, S. Boumerfeg, H. Trabsa, N. Charef, S. Khennouf and L. Arrar.
(2012) Xanthine Oxidase Inhibition and Antioxidant Effects of Peaganum harmala Seed
Extracts. European Journal of Medicinal Plants. 2(1): 42-56.
الملتقيات العلمية
THE 2nd International Symposium on Medicinal Plants, their cultivation and aspects of uses.
petra marriot hotel petra– jordan 3-4 november, 2010. Abderrahmane Baghiani, Meriem
Djermouni, Djemila Ameni, Hayet Trabsa, Noureddine Charef, Sabah Boumerfeg, Seddik
Khennouf, Daoud Harzallah, Lekhmici Arrar. In vitro antioxidant effects and xanthine
oxidase inhibitor properties of paganum harmala seed extracts.
3eme Séminaire International Biologie Animale. Univesité Mentouri Constantine (09-11/
2011). Djarmouni M, Baghiani A, Boumerfeg S, Adjadj M, Ameni D, Khennouf S, Arrar L.
Studies of antioxidant potential of methanol extract/Fractions of Peganum harmala.
2nd African Congress on Biology and Health(2nd ACBH 10-12 November 2012), Faculty of
Nature and Life Sciences, University Farhat Abbas Sétif. DJARMOUNI Meriem, Baghiani
Abderrahmane, Boumerfeg Sabah, Boussoualim Naouel, Belkhiri Farida and Arrar Lekhmici,
Studies of anti bacterial and antioxidant potential of methanol extracts /fractions of Peganum
harmala.

ملـــخص Peganum harmala احلرمل ملستخلصات نبتيت تقدير النشاطية املضادة لألكسدة إىلهذه الدراسة هتدف
الكمي تقدير ال ت نتائجبين. عمالن يف الطب الشعيب باجلزائرتسامل Santolina chamaecyparisus اجلعدةو أعلى كمية حيتوي على ((EAE-Sc عدةمستخلص إثيل أسيتات لنبات اجل الفينول والفالفونيدات أنلعديدات
غ من الوزن اجلاف/محض الغاليك ئمكاف مغ373.83 ± 0.23 قدرت بـ من املركبات الفينولية والفالفونيداتالنشاطية املضادة ريقدمت ت .على التوايل ،غ من الوزن اجلاف للنبتة/مكافئ كرستنيمغ ± 61.51 7.86و
مستخلص ظهرحيث أ ABTSو DPPHختبار إزاحة اجلذور احلرة تعمال إ، باس(in vitro)خمربيا لألكسدةEAE-Sc باإلضافة إىل اختبار تثبيط فوق أكسدة الليبيدات ،ختبارينأعلى نشاطية إزاحية للجذور احلرة يف اإل (EAE- Ph) احلرملمستخلص إثيل األسيتات لنبات أظهرت النتائج أن ، حيثFTCو β-caroteneباستعمال
فقد ظهرت FTCختبار إأما فيما خيص .β –caroteneبالنسبة إلختبار (% 94.21±0.003 ) قدرةأعلى ميلكلفوق األكسدة امؤشر ك MDAاستعمل تقدير . (CE-Sc) باملستخلص اخلام لنبات اجلعدةأعلى نسبة تثبيط
Ferrozine باستعمالاستخالب املعادن اختبار بني. أقل امتصاصية EAE-Phوقد سجل مستخلص ،الليبيدية أعلى قدرةميلك CE-Ph املستخلص اخلام لنبات احلرمل أن(IC50 =0.32 ± 0.009 املؤكسد للكزانثني نزماإلبينت نتائج إختبار قدرة املستخلصات على تثبيط .)مل/مغ(XO) وإزاحة جذرO2
ات لنبات للمستخلص الكلوروفورمي وإثيل األسيت أعلى نشاطية كانت أن الناتج عنه -•O2أعلى قدرة إزاحية جلذر سجلت يف حني .(EAE-Scو CHE-Sc)اجلعدة
لمستخلص الكلوروفورمي ل -• يف الكائن احلي دراسة النشاطية املضادة لألكسدةمتت (.CHE-Ph) لنبات احلرمل
(in vivo) كل منل ثانويليمستخلص امللل P. harmala ((MOHE-Ph وS. chamaecyparissus
(MOHE-Sc) النشاطية الكلية للبالزما عن طريق (APC : antioxidant plasma capacity) باستعمال إختبارDPPH لمستخلصماعدا القدرة اإلرجاعية ل. القدرة اإلرجاعيةو MOHE-Sc، امعنوي تأثريامل تبد املستخلصات.
نسيج الكبد يف GSHو MDA ياتومستو CAT)) الكتاالزأظهرت نتائج تقدير نشاطية إنزم من جهة أخرى، بنسبة MOHE-Phو MOHE-Scعند املعاملة بكل من CATنشاطية معنويا يف اعافتر ا .على التوايل ،% 50,44و % 59,21 نسبةب GSHكما رفعت من مستوى . على التوايل ،% 35 و % 59,16
قدرة املستخلصات دراسة وبينت .على التوايل ،% 63.48و % 79.56 نسبةب MDAخفضت من مستوى كما منقريبا ميلك تأثريا تثبيطيا MOHE-Phأن ، t-BHيف وجود املؤكسد in vivo ويالدم االحناللعلى تثبيط النتائج احملصل بينت .(على التوايل، دقيقة 4,95 ± 72,05و (MOHE-Sc 78,51 ± 11,32 = HT50 املستخلص
كبدنسيج مطحون القدرة على محاية ميلكان S. chamaecyparissusو P. harmala نبات أن كل منعليها اإلستعمال يفسرهذا ما و إىل ميزاهتما املضادة لألكسدة، أساسا من اإلجهاد التأكسدي وهذا التأثري يعود الفئران
.ماهلالواسع الطيب الشعيب
xanthine، عديدات الفينول، Peganum harmala، Santolina chamaecyparisus :الكلمات المفاتيح
oxidase ،استخالب املعادن، اإلحنالل الدموي ،مضادات األكسدة.

Abstract
The aim of this study is to evaluate the antioxidant activity of the extracts from Peganum
harmala and Santolina chamaecyparisus which are largely used in traditional medicine in
Algeria. The estimation of polyphenols and flavonoids showed that Ethyl Acetate extract of
S. chamaecyparisus (EAE-Sc) contains a high amount of phenolic compounds and flavonoids:
373.83 ± 0.23 mg gallic acid equivalent and 7.86 ± 61.51 mg quercetin equivalent/g of dried
weigh, respectively. The DPPH and ABTS tests were used to evaluate the antioxidant activity
in vitro. The EAE-Sc showed the highest scavenging activity of free radicals in both tests. The
inhibition of lipid peroxydation test using FTC and β-carotene showed that the ethyl acetate
extract of P. harmala (EAE-Ph) had the highest rate (94,21 ± 0.003 %) in β-carotene test, but
for the FTC test the highest inhibition rate was noticed with crud extract of S.
chamaecyparisus (CE-Sc). The MDA evaluation was used as indicator of lipid peroxydation.
The low absorption was found in EAE-Ph level. The capacity of the extracts to chelate the
mineral ions was tested using Ferrozine. The crud extract of P. harmala (CE-Ph) showed a
high capacity to chelate ferrous ions (IC50: 0.32 ± 0.009 mg/ml). The extracts were studied
for their ability of inhibition effect on XO and O2•- scavenging activity. The results showed
that the chloroform extract of S. chamaecyparisus (CHE-Sc) and EAE-Sc have the highest
inhibiting activity of XO, but the chloroform extract of P. harmala (CHE-Ph) showed the
greatest scavenging effect on O2•- with an IC50: 0.037 ± 0.0053 mg/ml. The antioxidant
activity of methanol extract of P. harmala (MOHE-Ph) and S. chamaecyparisus (MOHE-Sc)
in vivo was estimated using the antioxidant plasma capacity (APC), DPPH and reducing
power tests. Except for the reducing power of MOHE-Sc, all extracts did not show any
significant change. The activity of catalase (CAT), the level of MDA and GSH were
evaluated in mouse liver homogenate. The results showed a high significant increase in CAT
activity with MOHE-Sc and MOHE-Ph, by 59.16 % and 35 %, respectively, and olso in the
GSH levels by 59.21 % and 50.44 %, respectively. However they decreased the level of MDA
by 79.56 % and 63.48 %, respectively. The Evaluation of the plant extracts capacity to inhibit
the hemolysis of blood in vivo in the presence of the oxidant (tBH), showed that MOHE-Ph
has an inhibition effect comparable with MOHE-Sc (78.51 ± 11.32 min and 72.05 ± 4.95 min,
respectively). The present study showed that the P. harmala and S. chamaecyparisus have the
capacity to protect liver tissue from oxidative stress and this effect is probably due to their
antioxidant capacity, which explain the large use of these plants in traditional medicine.
Key words: Peganum harmala, Santolina chamaecyparisus, polyphenols, xanthine oxidase,
antioxidant, chelating.

Resumé
L’objectif de la présente étude est d’estimer l’activité antioxydante des extraits de deux
plantes, Peganum harmala et Santolina chamaecyparisus largement utilisées en médecine
traditionnelle en Algérie. L’extrait acétate d’éthyle de S. chamaecyparisus (EAE-Sc)
renferme une grande quantité de composés phénoliques et de flavonoïdes avec des teneurs de
373.83 ± 0.23 mg d’équivalent acide gallique et 61.51 ± 7.86 mg d’équivalent quercetin/g de
poids sec de la plante, respectivement. L’activité antioxydante a été évaluée in vitro par
l’utilisation des tests de DPPH et ABTS. L’extrait EAE-Sc a montré l’effet le plus élevé dans
les deux tests. L’examen de l’activité anti-péroxidation lipidique évaluée par les tests β-
carotène et FTC, a montré que l’extrait acétate d’éthyle de P. harmala (EAE-Ph) présente
l’effet inhibiteur le plus puissant dans le test β-carotène, par contre dans le test FTC, l’effet
de l’extrait brut de S. chamaecyparisus (CE-Sc) est le plus élevé. L’évaluation du MDA
utilisée comme indicateur de la peroxidation lipidique a montré la faible absorbance a été
enregistrée au niveau de EAE-Ph, La capacité des extraits de capter les ions minéraux testée
par l’utisation de ferrozine a indiqué que l’extrait brut de P. harmala (CE-Ph) présente
l’activité chélatrice la plus puissante (IC50: 0.32±0.009 mg/ml). D’autre part, l’évaluation de
la capacité des extraits d’inhiber la xanthine oxydase (XO) et d’éliminer le radical O2•- qu’elle
produit, a montré que l’extrait chloroformique du S. chamaecyparisus (CHE-Sc) et EAE-Sc
sont plus actifs. Cependant, la plus grande capacité d’éliminer le radical O2•- est observée avec
l’extrait chloroformique de P. harmala (CHE-Ph). L’activité antioxydante, in vitro, estimée
par la capacité antioxidante plasmatique (CAP), le test DPPH et le test du pouvoir réducteur a
montré que seul le MOHE-Sc possède un effet réducteur. L’activité de la catalase (CAT) et
les taux du MDA et du GSH ont été évalués dans l’homogénat du foie de souris. Les résultats
montrent une augmentation significative dans l’activité de la CAT par MOHE-Sc et MOHE-
Ph (59.16 % et 35 %, respectivement), en plus d’une augmentation du taux du GSH de 59.21
et 50.44%, respectivement et d’une diminution du taux du MDA de 79.56 % et
63.48 %, respectivement. L’évaluation de la capacité des extraits d’inhiber l’hémolyse des
globules rouges in vivo en présence d’un oxydant tBH, a montré que le MOHE-Ph possède un
effet inhibiteur de l’hémolyse similaire à celui du MOHE-Sc (78,51 ± 11,32 et 72,05 ± 4,95
min, respectivement). La présente étude montre que les plantes Peganum harmala et
Santolina chamaecyparisus possèdent une capacité de protéger le foie contre le stress oxydatif
et que cet effet est probablement dû à leurs capacités antioxydantes. Ceci explique leur grande
utilisation en médecine traditionnelle.
Mots-clés: Peganum harmala, Santolina. chamaecyparisus, polyphenols, xanthine oxidase,
antioxidant, chelation.

قائمــــــــة المختصــــــرات
AAPH
AqE
2,2′-azobis (2-methylpropionamidine) dihydrochloride
Aqueous extract
BHT Butylated hydroxytoluene
BXOR Bovine xanthine oxidoreductase
CAT Catalase
CHE Chloroform extract
COX Cyclooxygenase
CrE Crude extract
Cyt-c Cytochrome c
DPPH 2,2'-diphenyl-1-picrylhydrazyl
DTNB 5,5'- dithio-bis- [2- nitrobenzoic acid ]
EAE Ethyl acetate extract
FTC Ferric thiocyanate
GPx Glutathion peroxidase
GSH Gluthathion
GSSG Oxidized glutathione
HT50 half-hemolysis Time
HxE Hexane extract
INF Interferon
IL Interleukin
LOX Lipooxygenase
MDA Malondialdehyde
Mo Molybdene
NO• Nitric oxide
NOS Nitric oxide synthase
O2•- Superoxide anion
OH• Hydroxyl radical
ONOO- Peroxynitrite
PAF Platelet activating factor
PFR Protein/Flavin ratio
Ph Peganum harmala
RNS Reactive nitrogen species
ROS Reactive oxygen species
Sc Santolina chamaecyparissus
SOD Superoxide dismutase
SDS-PAGE Sodium dodecyl sulfate- polyacrylamid gel electrophoresis
TBA Thiobarbituric acid
t-BHP tert-butyl hydroperoxide
TCA Trichloroacetic acid
TEMED N,N,N'N' Tetramethylenediamine
XDH Xanthine dehydrogenase
XO Xanthine oxidase

الفهرس بالعربية امللخص
امللخص باالجنليزية قائمة األشكال قائمة اجلداول
قائمة املختصرات التشكرات
0..............................................................................................................................ةمـــمقد 3.........................................................................................................يجهاد التأكسداإل .0 3.............................................................................................اجلذور احلرة واألنواع النشطة .0. 0 5.........................................................................ائية لألنواع األكسيجينية النشطةالبنية الكيمي .4. 0 7..........................................................................................املصادر الرئيسية للجذور احلرة .3. 0
8.........................................................................................جي للجذور احلرةالدور الفيزيولو .5. 0 9.................................................................................األمراض املرتبطة باإلجهاد التأكسدي .6. 0
9.................................................................................... جهاد التأكسديالنتائج اجلزيئية لإل .7. 0
Xanthine oxydoreductase.....................................................................................00إنزم . 2
XOR..........................................................................................................00بنية إنزم . 2.1
XOR..............................................................................................04و توزع إنزم تواجد. 2.2
03...............................................................................................الدور الفيزيولوجي لإلنزم. 2.3
XOR............................................................................................03الدور املمرض إلنزم . 2.4
XOR....................................................................................................15مثبطات إنزم .2.5 XOR...........................................................................................07اآللية التحفيزية إلنزم .2.6
08.................................................................................................األنظمة املضادة لألكسدة .3
44.............................................................................................................املركبات الفينولية .4
23.............................. .....................................................األقسام الرئيسية للمركبات الفينولية .1. 4
27 ...............................................................................النشاطية البيولوجية لعديدات الفينول .4 .4
Peganum harmala .................................................................................. 31 نبات احلرمل .5
33..............................................................................................اإلستعمال التقليدي للنبتة .5.0
34............................................................................................التأثري السمي لنبات احلرمل .5.4

Santolina chamaecyparisus .......................................................................34 نبات اجلعدة. 6
35..............................................................................................اإلستعمال التقليدي للنبتة. 6.0
اجلزء العملي 37................................................................................................................الطرق املواد و .0 37...........................................................................................................املواد املستعملة. 1.1 38..................................................................................................................طرق العمل .2 38...........................................................................................نوليةياستخالص املركبات الف .0. 4 40.........................................................................................تقدير عديدات الفينول الكلية .4. 4 40..........................................................................................التقدير الكمي للفالفونيدات .3. 4 41....................................................................................من حليب البقر XORتنقية إنزم .2. 4
in vitro................................................................................44النشاطية املضادة لألكسدة .2. 5 XO.............................................................................................44تثبيط نشاطية إنزم . 2.5.1
O2التأثري اإلزاحي جلذر . 2.5.2 XO.......................................................................44الناتج عن -•
DPPH.........................................................................................45ختبار ازاحة جذر إ. 2.5.3 ABTS.........................................................................................45ختبار ازاحة جذر إ. 2.5.2 β –carotene/Linoleic acid............................................................................46إختبار . 2.5.5 47...........................................................................................إختبار استخالب املعادن. 2.5.6
Ferric thiocyanate.............................................................................47 (FTC)إختبار . 2.5.7
TBA (Thiobarbituric acid) .........................................................................48إختبار . 2.5.8
in vitro..................................................... ....48 لتحلل كريات الدم احلمراءملضادة النشاطية ا.2.5.9 in vivo..........................................................................49دراسة النشاطية املضادة لألكسدة .2.6 50..........................................................................لدم احلمراءلتحلل كريات االقدرة املضادة . 2.6.1
50............................................................ (DPPH) النشاطية املضادة لألكسدة الكلية للبالزما. 2.6.2
51.....................................................................................إختبار القدرة اإلرجاعية للبالزما. 2.6.3 51........................................................تقدير املؤشرات التأكسدية غري اإلنزميية يف نسيج الكبد .2.6.4 MDA...............................................................................................51تقدير كمية . 2.6.4.1 52 .....................................................................................الكبدي GSHتقدير كمية . 2.6.4.2 52.............................................................تقدير املؤشرات اإلنزميية على مستوى نسيج الكبد .5 .2.6 Catalase...................................................................................52تقدير نشاط إنزم . 5.1 .2.6

53.............................................................................................تقدير الربوتني الكلي. 5.2 .2.6 54.................................................................................................التحاليل اإلحصائية املستعملة
املناقشة النتائج و 55............................................................................إستخالص عديدات الفينول والفالفونيدات. 1
56..........................................................................تقدير عديدات الفينول الكلية والفالفونيدات. 2
in vitro....................................................................................58النشاطية املضادة لألكسدة . 3 58..............................................................................................التأثري املزيح للجذور احلرة. 3.1 DPPH.......................................................................................................58إختبار . 3.1.1
ABTS.......................................................................................................60إختبار . 3.1.2
62..............................................................................................إستخالب املعادنإختبار . 2 .3
65..................................................................................إختبار تثبيط فوق أكسدة الليبيدات. 3.3
β –caroténe................................................................................................66 إختبار. 3.3.1
68...................................................................................................... (FTC)ختبار إ. 2 .3.3
…70................................................................................................... (TBA)إختبار . 3.3.3
73....................................................................وتقدير نشاطيته XORتنقية ومراقبة نقاوة إنزم .3.4
XO.........................................................................................73التأثري التثبيطي على إنزم .3.5
O2ذرجلزاحي ثري اإلالتأ .3.6• XO............................................................................76الناتج عن -
AAPH......................................................78باستعمال in vitroالنشاطية املضادة لتحلل الدم .7 .3
80....................................................................يف الكائن احليدة دراسة النشاطية املضادة لألكس .4
80...........................................................................اتقدير النشاطية املضادة لألكسدة للبالزم .4.1 DPPH............................................................................81جذر إزاحةقدرة البالزما على .4.1.1
.…82..............................................................................رجاعية للبالزماختبار القدرة اإلإ. 4.1.2
84.........................................................................لتحلل كريات الدم احلمراءاملضادة النشاطية . 4.2
86........................................................................لكبداعلى مستوى MDA مستويات تركيز .3 .4 88.........................................................................................الكبدي CATتقدير نشاط .4 .4 89........................................................................................السيتوزويل للكبد GSHمعايرة . 4.5
92................................................................................................................................املناقشة العامة
98............................................................................................................................املراجع

قائمة األشكال ROS))............................................................. 5إنتاج خمتلف األنواع األكسجينية النشطة .1شكل
Oxypurinol.......................................16ومستقلباته Allopurinolبواسطة XORتثبيط إنزم .2شكل 17...............................................اإلنتقال اإللكرتوين بني مراكزهو XORاآللية التحفيزية إلنزم .3شكل 18....................تنظيم إنتاج األنواع األكسجينيية النشطة بواسطة األنظمة الدفاعية املضادة لألكسدة.4شكل 23...............................................................................البنية الكيميائية لألمحاض الفينولية .5شكل
24........................................................................................البنية العامة للفالفونيدات. 6 شكل
25...................................................................................األقسام املختلفة للفالفونيدات . 7 شكل
26............(الدباغ املكثفة) proanthocyanidinsو ( الدباغ املميهة) casuarictin لالبنية الكيميائية . 8 شكل 28.................................................................مواقع ارتباط األيونات املعدنية بالفالفونيدات .9شكل 28........................................................................ ذور احلرةتفاعل الفالفونيدات مع اجل. 10 شكل
P. harmala..................................................32 نبات احلرمل واألجزاء املختلفة املكونة له .11شكل Santolina chamaecyparissus L. ................................35الشكل املورفولوجي لنبات اجلعدة .12شكل 39....................اجلعدة طرق استخالص الفالفونيدات من حبوب نبات احلرمل واجلزء اهلوائي لنبتة . 13شكل
40................................................................................. منحىن العيارية حلمض الغاليك .14شكل rutin..........................................................................41و quercetin لمنحىن العيارية .15شكل DPPH ..........59على جذر S .chamaecyparisusو P. harmalaالتأثري اإلزاحي ملستخلصات .16شكل
DPPH..... 59من جذر %51املزحية لـ S. chamaecyparisusو P .harmalaمستخلصات تركيز .17شكل
على جذر S .chamaecyparisus و P. harmala ملستخلصاتالتأثري اإلزاحي .18شكلABTS...........................................................................................................................61
من جذر %51املزحية لـ S .chamaecyparissusو P. harmalaتركيز مستخلصات .19 شكلABTS...........................................................................................................................61
Feنسبة تثبيط مركب .20 شكل+2
-Ferrozine مبستخلصات P. harmalaو S. chamaecyparisu...........63
من 50%على إستخالب S. chamaecyparisusو P.harmalaتركيز مستخلصات . 21 شكل 64............................................................................................................................املعادنيف ساعة 24خالل S. chamaecyparisus و P. harmala النشاطية املضادة لألكسدة ملستخلصات . 22شكل
..............66.........................................................................محض اللينولييك/β-caroteneإختبار
ساعة 42بعد مرور )املاء امليثانول و و (BHT تخلصات والشواهدالنشاطية املضادة لألكسدة للمس. 23 شكل 67....................................................................................محض اللينولييك/β-caroteneيف إختبار

P. harmalaحركية فوق أكسدة محض اللينولييك يف وجود وغياب مستخلصات .24 شكل S .chamaecyparisus.....................................................................................................69و
Sو P.harmala (Ph)الناتج أثناء أكسدة الليبيدات ملستخلصات MDAإمتصاصية مركب .25شكل
.chamaecyparisus (Sc) نيةبواسطة تق TBA...........................................................................71 S. chamaecyparisusو P .harmala مقارنة بني النشاطية املضادة لألكسدة ملستخلصات نبتيت .26شكل
TBA..................................................................................................72و FTC باستعمال تقنييت XO......74على نشاطية إنزم S. chamaecyparisus و P. harmalaالتأثري التثبيطي ملستخلصات .27شكل من نشاطية إنزم %05املثبطة لـ S. chamaecyparisusو P. harmalaتركيز مستخلصات .28شكل XOR............................................................................................................................74 O2 على جذر S. chamaecyparisus و P. harmala ملستخلصات زاحيالتأثري اإل .29 شكل
XO الناتج عن -• Cyt c................................................................................................................76 بإستعمال
O2و النشاطية املزحية جلذر XO من نشاطية %51لـ التأثري التثبيطي .30شكل •-...................................77
يف وجود S. chamaecyparissusو P. harmala ستخلصاتملنسبة حتلل كريات الدم احلمراء .31شكل (AAPH).......................................................................................................................79
بمقارنة Phو Scملستخلصات DPPHالنشاطية املضادة لألكسدة للبالزما اجتاه جذر .32شكل
contrôle........................................................................................................................81 83.........................................................................................القدرة اإلرجاعية للبالزما .33شكل
85................ ............... ........................................اميع املدروسةملختلف اجمل HT50زمن . 34 شكل
MDAعلى معدل MOHE-Scو MOHE-Ph تأثري معاملة الفئران باملستخلص .35 شكل 87..........................................................................................................................................لكبدا
CAT ........................88على نشاط إنزم MOHE-Scو MOHE-Phتأثري املعاملة مبستخلص .36شكل GSHعلى معدالت MOHE-Scو MOHE-Phتأثري املعاجلة مبستخلص .37شكل
90.............................................................................................................................لكبدا

قائمة الجداول
4......................................................تصنيف اجلذور احلرة وتأثريهاهتا املختلفة .1 جدول
وحمتواها من عديدات الفينول P. harmalaمردود مستخلصات حبوب نبات احلرمل .2جدول
57....................................................................................لفالفونويداتا و
وحمتواها من عديدات الفينول S .chamaecyparisusمردود مستخلصات اجلزء اهلوائي لنبتة اجلعدة .3 جدول 57....................................................................................الفالفونويدات و

مقدمـة
تعمل مضادات األكسدة على خفض األضرار التأكسدية على مستوى اخلاليا واجلزيئات احليوية اليت تسببها
ة النشطةوالنرتوجينيreactive oxygene species (ROS) األنواع األكسجينية
RNS) )reactive nitrogene species .وجد أن فقدROS وRNS ةببسمتساهم يف التخريب التأكسدي
اإللتهاب املزمنو داء السكريو العديد من األمراض عند اإلنسان مثل إلتهاب املفاصلأو استمرار ظهور بذلك
،وهلذا السبب(. 2007وآخرون، Valko)وبعض أنواع السرطان Alzheimerمرض مثل اإلختالالت العصبية و
العديد من النباتات ونظرا لكون. حضيت التأثريات الوقائية ملضادات األكسدة باهتمام كبري يف السنوات األخرية
اتمضاد أو/لألكسدة و اتمضادحيث يعمل الكثري منها ك، ذات نشاطية حيويةبإنتاجها ملركبات معروفة
نذكر عديدات ،ومن بني مضادات األكسدة هذه. ات على إختبار هذه الفعاليةركزت الكثري من الدراس للبكترييا
تعمل املركبات الفينولية . هي مستقلبات نباتية ثانوية متثل أكثر مضادات األكسدة الطبيعية انتشاراو الفينول
آخرون، و Fattouch)كعوامل مرجعة وماحنة للهيدروجني ومزحية للجذور احلرة وملتقطة لألكسجني املهيج
تستعمل العضوية مضادات أكسدة داخلية وخارجية املنشأ للدفاع ضد األضرار اليت تسببها (.2007
ROS/RNS، مثل إنزمييةأنظمة تقسم إىل و superoxide dismutase (SOD) وcatalase (CAT)
فيتامينات وال reduced glutathione (GSH)وأنظمة غري إنزميية مثل glutathione peroxidase (GPx)و
ا ممغري قادرة على احلماية من األضرار التأكسدية الداخلية يف بعض األحيان مضادات األكسدة تصبح. عادنوامل
، Ibrahimو (Habibعلى الوقاية من اإلجهاد التأكسدي هايعمل استهالكواليت يتطلب تدخل مركبات خارجية
2011 .)
Santolinaونبات اجلعدة Peganum harmalaاحلرمل نبات مها تنيطبي تنينبتعلى هذه الدراسة جرت
chamaecyparissusبذور تستعمل فعلى سبيل املثال ،، املعروفة بامتالكهما للعديد من النشاطيات البيولوجية

ومضادة للسرطان ( 2010وآخرون، Moghadam)كمضادة للبكترييا والفطريات P. harmalaنبات احلرمل
Baghiani؛ 2007وآخرون، Moura)ومضادة لألكسدة (2005وآخرون، Jahaniani)والطفرات الوراثية
وآخرون، De Logu)نشاطية مضادة لإللتهاب S. chamaecyparissusيف حني متلك (. 2012وآخرون،
Dhanabal)الكبد واقيةكما متتلك نشاطية ،Phospholipase A2 إنزم وتعمل على تثبيط نشاطية ،(2000
(.2012وآخرون،
على إزاحة اجلذور احلرة S. chamaecyparissusو P. Harmalaمت يف هذه الدراسة إختبار قدرة مستخلصات
اختبار التأثري اإلستخاليب للمعادن، كما مشلت كذا وتثبيط فوق أكسدة الليبيدات باستعمال عدة تقنيات و
O2وإزاحة جذر XOالدراسة اختبار قدرة املستخلصات على تثبيط إنزم يف حني تقصت الدراسة . الناتج عنه -•
على على التأثري S. chamaecyparissusو P. harmala كل منقدرة املستخلص امليثانويل ليف الكائن احلي
MDAحيث مت تقدير مستويات ،فوق األكسدة الليبيدية والنظام املضاد للتأكسد على مستوى نسيج الكبد
هتمت الدراسة باختبار التأثري املضاد لالحنالل الدموي للمستخلصات كما ا.Catalase إنزمونشاطية GSHو
. يف وجود املؤكسدات احملفزة لالحنالل الدموي

اإلجهاد التأكسدي. 1
هذا التوازن قد خيتل حلساب غري أن اجلذور احلرة، للعلى مستوى اخلاليا السليمة بني إنتاج و حتتوازنا يوجد
مسببا هذه احلالة باإلجهاد التأكسدي تدعى احلرة أو نقص يف كمية مضادات األكسدة، إنتاج مفرط للجذور
عرف (. 2012وآخرون، Shinde)أضرارا خلوية خطرية يف حالة ما إذا كان اإلجهاد قويا و ذو فاصل زمين طويل
د هجوم األنواع مصطلح اإلجهاد التأكسدي بأنه عدم قدرة اجلسم على الدفاع ض ،1990سنة يف ،Sieالعامل
لة قدرة األنظمة املضادة أو ق ROS، و هذا راجع لالختالل الناتج عن فرط إنتاج ROSاألكسيجينية النشطة
(.Pincemail ،2007و (Defraignet لألكسدة
الجذور الحرة و األنواع النشطة .1.1
،جية على مستوى مدارها اخلارجيزو متلك واحدا أو عدة إلكرتونات غريأو ذرة كل جزئية تعترب اجلذور احلرة
وبذلك تصبح هذه اجلزيئات غري مستقرة، إذ حتاول أخذ إلكرتون من اجلزيئات اجملاورة هلا وهبذا تؤدي إىل
تنتج اجلذور احلرة عرب حيث ، (0جدول ( )من امللي ثانية إىل نانو ثانية) تتميز مبدة حياة قصرية جدا. أكسدهتا
عنه طاقة عالية، عند تفاعلها مع جذور أو جزيئات أخرى تتحول هذه األخرية انتقال إلكرتون هذا ما ينتج
جند األنواع األكسيجينية النشطة ،من بني اجلذور احلرة (.2006وآخرون، (Finaud بدورها إىل جذور جديدة
(ROS ) و اليت تشتق من جزيء األكسيجني، باإلضافة إىل األنواع النرتوجينية النشطة(RNS ) يتية النشطة والكرب
(RSS( ) 50جدول).

(.2006وآخرون، (Finaudتصنيف اجلذور احلرة وتأثريهاهتا املختلفة . 1جدول
Free radical Contraction Half-life Main effects
Reactive oxygen species ROS
Superoxide ion O2•-
10-5
sec
Lipid oxidation and
peroxidation
protein oxidation
DNA domage
Ozone O3 Stable
Singlet oxygen 1O2 1µsec
Hydroxyl radical OH•
10-9
sec
Hydrogen peroxide H2O2 Stable
Hypochlorous acid HOCI Stable
Alkoxyl radical RO•
10-6
sec
Peroxyl radical ROO•
7 sec
Hydroperoxyl radical ROOH•
Reactive nitrogen species RNS
Nitric oxide NO•
Lipid peroxidatio
DNA domage
Proteins oxidation
Nitric dioxide NO2•
1-10 SEC
Peroxynitrite ONOO•-
0.05-1
SEC
Reactive sulphur species RSS
Thyil radical RS•
Proteins oxidation
DNA domage
ROS production

البنية الكيميائية لألنواع األكسيجينية النشطة. 2. 1
O2تشمل األنواع األكسيجينية النشطة كل من اجلذور احلرة مثل أيون فوق األكسيد OH، جذر اهليدروكسيل -•
•
(.1شكل) H2O2واألنواع غري اجلذرية واليت تعترب ذات مسية عالية مثل بريوكسيد اهليدروجني
(.ROS) ) Lucy Chen)، 2012إنتاج خمتلف األنواع األكسجينية النشطة . 1 شكل
O2) يعترب أيون فوق األكسيد. وينتج عرب اإلرجاع األحادي جلزيء األكسيجني ،اجلذر األكثر إنتشارا يف اخللية( -•
O2يتكون جذر . ستطيع اخرتاق األغشية البيولوجيةيال إذ يتميز هذا اجلذر بنشاطية ضعيفةتلقائيا على -•
مستوى الغشاء الداخلي للمتوكوندري خالل السلسلة التنفسية، أو عن طريق عدة إنزميات مثل
xanthine oxydase إنزميي ،cyclooxygenase وlipoxygenaseباإلضافة إىل إنزم ، NADPH oxydases
(. Laitonjam ، 2012 ؛Genestra ،2007؛ 2003وآخرون، Gardès-Albert) لغشاء اخلاليا البلعمية
O2+ e- O2
•-

هنا اإلنتشار والتفاعل مع اجلزيئات على بعد جزيئة مستقرة متلك نصف عمر طويل نسبيا، بإمكا H2O2يعترب
. جذرا حرا لكن بإمكانه إنتاج جذور عالية النشاط H2O2ال يعترب . Mc Cord)، (1993مسافات من تشكلها
هو جذر جذرا عايل النشاط Fentonعرب تفاعل H2O2، ينتج (احلديد والنحاس)جود األيونات املعدنية ففي و
OHاهليدروكسيل• Wardman) وCandeias ،1996؛Laitonjam ، 2012 ) كما يساهم يف تشكيل جذر ،
، Genestra؛ myeloperoxidase (Sorg ،2004عرب إنزم hypochlorous acid (HOCl)خطري آخر هو
(. Laitonjam، 2012 ؛2007
H2O2+ O2•- HO
• + O2 + OH
- OHميلك جذر اهليدروكسيل
إذ ميثل اجلذر األكثر نشاطا و يتميز بعدم استقرارية . نصف عمر يقدر بالنانو ثانية •
حملدود هلذا اجلذر بالتفاعل مع العديد من يسمح اإلنتشار ا. عالية من بني كل األنواع األكسيجينية النشطة
هذا ما يسبب تشكل العديد من األضرار ... ( .، ADNبروتينات ، ليبيدات ، )اجلزيئات اليت تكون قريبة منه
.Laitonjam ، (2012 ؛Halliwell ،1993و Guteridge)اخللوية
O2يعترب (. 2005وآخرون، (Delattreشاطه العايل نوعا مهيجا لألكسجني وهو يشبه اجلذر احلر نظرا لن 1
ينتج اجلسم جماميعا أخرى مشتقة من اآلزوت وتعرف ،باإلضافة إىل كل هذه اجملاميع األكسجينية النشطة
وفوق أكسيد النرتيت NOوتضم كل من جذر أحادي أكسيد اآلزوت RNSالنرتوجينية النشطة باجملاميع
.ONOO NOS(nitric oxide إنزم بواسطة citrullineإىل argenine محض تفاعل إستقالبمن NOينتج •
synthases) ،Xia) ،لك، مي(1999وآخرون NO جذور ) احلرةذور اجلالتفاعل مع القدرة علىperoxyl
عندما تنتج .( Laitonjam، 2012)تثبيط فوق أكسدة الليبيدات ويعمل على تها،نشاطي خمفضا من( alkylو
O2ة عالية من اخللية كميONOO)فوق أكسيد النرتيت شكاليلهذين األخريين يتحد، NOباملوازاة مع -•
•) ،
، Laitonjam ؛Grigorov) ،2012و حتفيز فوق أكسدة الليبيدات ADNجذرا مؤكسدا يقوم بتجزئة ويعترب
2012 ). NO•+ O2
• ONOO
-

المصادر الرئيسية للجذور الحرة. 1.3
الداخلية ROSمصادر . 1 .1.3
NADPH األكسدة الذاتية للجزيئات الصغرية، نذكر منها ROSمن مصادر يوجد داخل العضوية العديد
oxydase ، xanthine oxydasوامليتوكندري اليت تعترب من أهم املصادر ، البريوكسيزومات.
ROSلـ هاما مصدرا hydroquinoneو dopamine ،adrenaline ، flavineة الذاتية للجزيئات األكسدمتثل
(Freeman وGrapo ،1981 ) هو جذرعموما ذه األكسدة هناتج وO2، باإلضافة إىل ذلك فإن األكسدة -•
خاصة متثل إحدى أسباب املوت املربمج الذي يظهر خالل األمراض العصبية dopamineالذاتية جلزيئات
.(Fanburg ،2000 و (Parkinson Thannickalمرض
خالل هذا التفاعل يعمل كالكزانثني إىل محض اليوري والكزانثني إىل هدم اهليبوكزانثني xanthie oxydaseخيفز
O2األكسجني اجلزيئي كمستقبل لإللكرتونات منتجا بذلك جذر Kelley ؛1986ن، وآخرو (H2O2 Parksو -•
عرب إرجاع النرتيت غري العضوي، ويف وجود NOأيضا على إنتاج جذر XORيعمل كما ، (2010 وآخرون،
O2األكسجني يتحد NOمع -•
ONOOإلنتاج •• Harrison) ،2006.)
Xanthine+ 2O2 +H2O xanthine oxydase
acide urique +2O2•-
+ 2H+
. عية وبصفة خاصة يف املكافحة ضد العضيات الدقيقةدورا أساسيا يف االستجابة املنا NADPH oxydaseيلعب
الغشاء البالزمي للخاليا البالعة مما حيفز إرجاع إال أنه خالل عملية البلعمة، يتواجد هذا االنزم على مستوى
O2األكسجني إلنتاج جذر •- (Babior ،1999 ؛Sardina 2012، وآخرون).
NADPH+ 2O2 NADPH Oxydese NADP
+ + 2O2
•- + H
+
اخللوي ROSمن %90وذلك عرب السلسلة التنفسية، واليت تنتج ROS ـلمتثل امليتوكوندري املصدر الرئيسي
(Balaban ،2005)وآخرون .

انتقال اإللكرتونات عرب السلسلة التنفسية إىل غاية املستقبل النهائي للمتكوندري علىالغشاء الداخلي مليع
ويتم ،احلركة اإللكرتونية يصاحبها إنتقال نشط للربوتونات من احلشوة إىل الفراغ بني الغشائني هذه(. األكسجني)
؛ ATP (Mitchell ،1961شكل فتنتج بذلك طاقة على VІو ІІІ و Іذلك على مستوى النواقل
Labuschagne وBrenkman ،2013 .)ماء، يعمل األكسجني كمستقبل هنائي لإللكرتونات لريجع إىل جزيئة
O2خالل سلسلة نقل اإللكرتونات يتسرب جزء من اإللكرتونات ليتحد مع األكسجني وينتج جذر •-
(Labuschagne وBrenkman ،2013) .ةدمست يتم O2 (Mn-SOD)املتوكوندري SODبواسطة H2O2إىل -•
.(Schacht ،2013 و Böttger) ((Cu-Zn-SODأو السيتوزويل
يةالخارج ROSمصادر . 2 .1.3
يتعرض اجلسم للعديد من العوامل اخلارجية اليت تتسبب يف تشكيل اجلذور احلرة، نذكر على سبيل املثال األشعة
O2فوق البنفسجية اليت تعمل على إنتاج OH و -•
O2و •1 (Pavlou ،2009وآخرون .) كما تعمل األشعةx
اهلواءو اجلذور احلرة من تلوث املاء تنتج (. Beani ،1995)إىل جذرين حرين H2Oعلى جتزئة جزيئة γو
، Cyclosporine)ومن بعض األدوية ( Cd ،Hg ،Pb،Fe ،As)الكحول واملعادن الثقيلة و التدخنيو
Tacrolimus وBleomycine ) اللحم املقدد، استعمال الزيوت، )وبعض املذيبات الصناعية ومن جراء الطبخ
ركبات اخلارجية بعد دخوهلا إىل اجلسم عرب العديد من تتفكك امل (.2008وآخرون، Pham-Huy( )الدهون
.املسالك وهتدم إىل جذور حرة
الدور الفيزيولوجي للجذور الحرة. 1.5
كذلك يف تساهممصطلح اإلجهاد التأكسدي يف العموم باألضرار واإلصابات اخلطرية، إال أن اجلذور احلرة يرتبط
كما تلعب . حيث تؤثر بالتحديد ضد املستضدات أثناء عملية البلعمة العديد من الوظائف الفيزيولوجية كاملناعة،
بإمكاهنا أنو (. دور مراسيل خلويةب تقوم)دورا هاما يف نقل اإلشارات اخللوية، ويف البناء احليوي للخاليا

بتدخلها يف تنشيط اإلنزميات، وتلعب دورا هاما يف أيضا ROSتعرف . تعدل من حالة األكسدة اإلرجاعية
(.2006 وآخرون، Finaud)لتقلص العضلي ا
NOميلك جذر ،فعلى سبيل املثالالعديد من الوظائف الفيزيولوجية على مستوى اجلهاز العصيب، فهو يؤثر •
أما على مستوى . ويلعب دورا يف املرونة الشبكية ويف الذاكرة طويلة املدى neuromodulateurكمنظم عصيب
ويهدم بعض الكائنات احلية الدقيقة الدموية يثبط تكتل الصفائح و غط الدمويالوعائي، فهو ينظم الض اجلهاز
(Sorg ،2004.)
األمراض المرتبطة باإلجهاد التأكسدي. 6. 1
.وبعض اجلزيئات الكربى إىل ظهور العديد من األمراض الربوتيناتو ADNتؤدي األضرار التأكسدية اليت تصيب
يعترب اإلجهاد التأكسدي أيضا السبب الرئيسي يف ظهور .(Halliwell ،1994)خاصة أمراض القلب والسرطان
مراضاألو إلتهاب كبيبات الكلىو إلتهاب املفاصلو ابية مثل إلتهاب األوعية الدمويةعديد من األمراض اإللتهال
مرض و مرض نقص املناعة املكتسبو (ءاألمعا إقفارالسكتة الدماغية و و أمراض القلب) النامجة عن اإلقفار
، Alzheimer)ألمراض العصبية وا ارتفاع الضغط الدمويو القرحة املعديةو emphysemaانتفاخ النسيج اخللوي
parkinson) وغريها بالتدخني الناجتة عناألمراض و (Lobo ،2010، وآخرون.)
النتائج الجزيئية لإلجهاد التأكسدي. 1.7
يئات البيولوجية وبذلك يؤثر على خمتلف الوظائف اخللوية، أضرارا على مستوى اجلز ROSاإلنتاج املفرط ل يسبب
Kohen) األكثر عرضة هلجوم اجلذور احلرة ADNوتعترب األغشية البيولوجية واإلنزميات الربوتينية و
(.Nyska ،2002و

فوق األكسدة الليبيدية. 1.7.1
ROS (OHم و جهلغري املشبعة األهداف الرئيسية متثل األمحاض الدهنية، واليت تتواجد على مستوى (خصوصا •
(. Moussard ،2006)الفوسفوليبيدات والكولسرتول، ومتثل الليبيدات األساسية لألغشية اخللوية والليبوبروتينات
OHيقوم جذر حيث مرحلة البدأ: مراحل ثالثةتتم فوق األكسدة الليبيدية وفق مبهامجة جمموعة ميثيالن •
Rمزدوجتني حلمض دهين غري مشبع منتجا جذرا كربونيا متواجدة بني رابطتنيOHينزع ) •
ذرة هيدروجني ل •
CH2- كما يتحول ، (مث حيدث إعادة ترتيب الروابط املزدوجة RRO2إىل •
مرحلة تليها ،يف وجود األكسجني •
RO2ينزع جذر أين التضاعفRذرة هيدروجني من محض دهين جديد قريب منها، لينتج بدوره جذر •
مث •
RO2جذرalcoxyl (ROيتفاعل جذر حيث فيبدأ بذلك تفاعل سلسلة ،•
مع محض دهين جديد من أجل ( •
توقيف تفاعالت لبني جذرين حرين تفاعل رحلة النهاية حيدث أما يف امل ،احملافظة على مرحلة اإلنتشار الليبيدي
: أكسدة الليبيدات يف تتمثل املركبات الرئيسية الناجتة من فوق (.Moussard ،2006)السلسلة
MDA(malondialdehyde ) 4وHDA (4hydroxyalkenal ) 4وHDN (4hydroxynonenal .) هذه تعترب
(.Beeber ،2008و Borg)التفاعل مع بعض األمحاض األمينية هلا القدرة على املركبات مؤكسدات
أكسدة البروتينات. 1.7.2
مشكال نواجتا (R)سواءا على مستوى السلسلة اجلانبية ROSأهدافا ل متثل األمحاض األمينية املكونة للربوتينات
إن معظم األمحاض األمينية تقريبا تكون . لسلسلةامؤكسدة أو على مستوى الرابطة البيبتيدية، مؤديا إىل جتزء
هي ( Trpو Tyr)والعطرية ( Metو Cys)، إال أن األمحاض األمينية الكربيتية ROSعرضة للتأكسد بواسطة
كربونيلية على مستوى تؤدي أكسدة األمحاض األمينية إىل إنتاج جماميع هيدروكسيلية و . األكثر حساسية للهجوم
كإحداث جسور كتشكيل شبكات داخل أو بني خلوية إحداث تغريات بنيوية هامة كذلك بإمكاهناو الربوتينات
(. Beeber، 2008و Borg )وظائفها البيولوجية بذلك على ةؤثر مليت تغري يف البنية الربوتينية ثنائية الكربيت ا

يتم التخلص منها، فإما توجه للهدم اأحيانا فاقدة لنشاطيتها كليا لذعموما مصابة و الربوتينات املتأكسدة ظهرت
(.Beeber، 2008و Borg ؛Moussard ،2006)بواسطة الربوتينات احلالة، أو تشكل تكتالت صلبة
ADNأكسدة . 1.7.3
بإمكاهنا التفاعل معها ROSعبارة عن جزيئات مستقرة وحممية بصورة عالية إال أن ADNبالرغم من أن جزيئات
سلسليت ، كسور بسيطة ألحد أو كالADNمنها تغيريات على مستوى قواعد ،إلحداث العديد من األضرار
ADN فقدان ألجزاء بيورينية ،( مواقعapurinique)كر الريبوز منقوص األكسجني، ، إصابات على مستوى س
(.Priyadarsini ،2011و Kunwar) ADNوإصابة نظام تصليح ( تصالب)مع بروتني ADNإحتاد
مؤكسدة بعض القواعد اآلزوتية فتتشكل بذلك جذور هيدروكسيلية ADNتؤثر جذور اهليدروكسيل مباشرة على
OH)مضافة • adducts ) أهم القواعد اآلزوتية املعدلة نذكرمن . وتتغري طبيعتهابدورها تتأكسد :
8-hydroxydeoxyguanosine (8-OHdG ) 8و (or4- ,5-)-Hydroxyadenine وThymine peroxide
يسبب كسورا وتشوهات كروموزومية، ROSإن هجوم .uracyl (Hydroxymethyl)-5و Thymine glycolو
(.Priyadarsini ،2011و Kunwar)النسيجية خة واليت تعترب عوامل حمفزة لظهور السرطان والشيخو
Xanthine oxydoreductaseإنزيم . 2
بنيته. 2.1
ميلك . من حليب البقر عن معقد فالفوبروتني معدين، مت عزله أول مرةعبارة Xanthine oxydase (XOR )نزم إ
حتتوي كل حتت .ن بعضهما، ويقسم إىل حتت وحدتني متماثلتني ومستقلتني حتفيزيا عXOR (290 kDa) نزمإ
، واجملال املركزي حيتوي مركز Fe2S2 (20 kDa)الذي ميلك مركزين N-termإرجاعية، اجملال مراكز 4وحدة على
FAD (40 kDa ) واجملال C-term الذي حيتوي عامل مساعد للموليبدان(MO-CO( )85 kDa )Nishino)
(.2009وآخرون، Hadizadeh ؛2005، وآخرون

وهو الشكل Xanthine deshydragenase (XDH, EC1.1.1.204 )الشكل املرجع ،اإلنزم على شكلنييظهر
إن حتول . السائد يف السوائل Xanthine oxydase (XO, EC1.1.3.22)املؤكسد ، والشكل in vivoالسائد
ة أو غري عكسي اجملاميع الكربيتيقد حيدث عكسيا عرب أكسدة XOإىل الشكل املؤكسد XDHالشكل املرجع
(.2008وآخرون، Nishino؛ Harisson ،2006)عرب القطع الربوتيين لألمحاض األمينية النهائية
مسؤول عن ميتابوليزم البيورينات، الذي حيفز أكسدة الكزانثني و اهليبوكزانثني إىل محض XORيعترب إنزم
O2بإنتاج اليوريك، مرفقاتقع مورثة إنزم (. 2009وآخرون، Hadizadeh ؛ 2007وآخرون، Ferrari) H2O2و -•
XOR الشريط على مستوى البشريةp22 36تتكون من ،2للكروموزوم exones 35و intron مكونة بذلك
1333البشري من XORيتكون التسلسل الربوتيين إلنزم . النيكليوتيدية زوج من القواعد 3999اجملموع الكلي
للفئران واجلرذان XORمن التشابه البنيوي مع %91هذا التتايل ميثل . محض أميين
(Berry وHare ،2004.)
XORإنزيمتواجد و توزع . 2.2
فقد (.Granger ،1986و Parks)بشكل واسع ابتداء من البيكترييا إىل غاية الثديات XORيتواجد إنزم
نسجة مثل على مستوى بعض األ ، تواجد اإلنزم على شكلني نشط وغري نشطأظهرت تقنيات التواجد املناعي
تتوزع نشاطية. (2004 وآخرون، Martin ؛Bulkley، 2002و Meneshian)نسان قلب اإلو والدماغ الكبد
Pacher)اإلنزم بشكل واسع على مستوى أنسجة الثديات، وتظهر مبستويات مرتفعة يف الكبد واألمعاء
بطانة، و الت امللساء خاليا العض)من األنسجة واخلاليا ديف العدي XORحتديد إنزم مت كما (.2006وآخرون،
وعلى ( Hellesten-Westing ،1993( )والبالعم البدنيةالت اهليكلية، اخلاليا للقلب، العضالدموية ية وعاأل
يف حني أظهرت دراسات (. 1991وآخرون، Stevens( )الروماتيزمية)مستوى البطانة املفصلية السليمة وامللتهبة
على مستوى همتركز مستوى املناطق احمليطة بالنواة، كما أظهرت السيتوبالزمي على XORمتركز إنزم أخرى

بينما وظيفته هناك غري معروفة، ميكن أن تكون مسؤولة عن . لإلنسانالطالئية للخاليا احلويصالت داخل خلوية
الربوتني األساسي املكون لغشاء XOR إنزمباإلضافة إىل ذلك، يعترب (.Harrison ،2002)ختزين اإلنزم
. (Mather ،2000)القطريات الدهنية للحليب الطازج الكريات الدهنية للحليب، اليت تغلف
نزيمالدور الفيزيولوجي لإل. 2.3
منذ عشرات السنني، كان يعتقد أن الدور الوحيد لإلنزم يتمثل يف هدم البيورينات، إال أنه نظرا لتعقيداته وتوسع
لى مستوى فيزيولوجية العضوية، فقد ع أكثر مشوليةنشاطيته وتوزعه النوعي يف األعضاء فإن وظائفه أصبحت جمال
وآخرون، Martin)مت إظهار دورا هاما لإلنزم يف امتصاص احلديد على مستوى األمعاء الدقيقة وحركتها يف الكبد
2004.)
NO)العضوي إىل أكسيد النرتيك إرجاع النرتيت غري XORعند نقص األكسجني، حيفز إنزم ، واليت متثل (•
O2أما يف حالة وجود األكسجني، فهو يرجع بذلك مكونا . جزيئة اإلشارة اليت متلك خصائص قاتلة للبيكترييا• ،
NOالذي يتفاعل بسرعة مع ONOOإلنتاج •
وآخرون، Martin)لبيكترييا ل، والذي ميثل عامال فعاال قاتال•
ختفيض خطر ويعمل على (in vivoداخل الكائن احلي ) XORضع حليب األم الغين بإنزم يستقبل الر . (2004
(. 2000وآخرون، Stevens)ظهور إلتهابات املعدة واألمعاء
XORالدور الممرض إلنزيم . 2.4
لتهابفي اإل XORدور . 2.4.1
ثل ظاهرة طبيعية حتدث داخل العضوية متثل اإلستجابة اإللتهابية عنصرا هاما جدا يف املناعة الطبيعية، فهي مت
، بتحفيز من XORو cytokinesتنتج هذه التفاعالت العديد من جزيئات . أو عدوى حو بسبب اإلصابة جبر
( IL3و IL1)و (TNF-α) ، وعامل النخر الورميInterferon (αINF-)و Interferon (γIFN- )جزيئات
(Vorbach ،2003وآخرون.) يشارك إنزمXOR عن تهابية، كما يشارك يف توسعهااإلل انتشار التفاعالتيف

اإللتهاب على مستوى السائل XORيف حالة إلتهاب املفاصل الرثوي، يضخم إنزم . ROS ه لطريق إنتاج
كما يؤدي إىل ظهور مضاعفات مثل اإللتهابات ،املصلي يف املفاصل مؤديا إىل تآكل العظم وانتشار املرض
بإمكاهنا التفاعل مع مكونات الغشاء XORاملنتجة بواسطة ROSإن (.1997، خرونوآ Blake)الوعائية
مما يساهم يف انتشارإنتاج ليبيدات جاذبة كيميائيا للخاليا املتعادلة وتؤدي إىلاخللوي كحمض األراشيدونيك،
ية مسئولة عن جزيئات سطحإنتاج حتفيز ROS كما بإمكان(. 1990 ،وآخرون Perez)اإلستجابة اإللتهابية
و Conner)إلتصاق اخلاليا البيضاء على خاليا البطانة الوعائية وهجرهتا عرب البطانة حنو موقع اإللتهاب
Grisham ،1996.)
حتقانفي اإلقفار وإعادة اإل XORدور إنزيم . 2.4.2
يف ATPهدم تسبب إصابات إقفارية أثناء XORإنزم الناجتة عن ROS، أن (1981) وآخرون Granger وجد
، ATPعند اخنفاض مستويات . ىل زيادة مستويات اهليبوكزانثنيحالة نقص مستويات األكسجني هذا ما يؤدي إ
Caىل دخول بالزمي مؤديا إحيدث اختالل يف التدرج األيوين على مستوى الغشاء الإن ارتفاع .داخل اخللية +2
Caاملعتمدة على protéases الرتاكيز البالزمية للكالسيوم يؤدي إىل تنشيط إنزميات2+ (Granger ،وآخرون
XO (Kooij، إىل in vivo، السائد XDHاليت حتفز التحول غري العكسي ل (McCord ،1985؛ 1981
.1992)وآخرون،
O2دم اهليبوكزانثني املرتاكم ويرجع هب XOإنزم يقوم وبوجود األكسجني،أثناء إعادة اإلحتقان الذي يتبع اإلقفار
O2إىلOHينتج عن التفاعل مع هذين اجلذرين جذر . H2O2و -•
الذي ميلك قدرة عالية للهدم مما يؤدي إىل •
عن إنتاج العوامل الضارة لوحده، وإمنا بإمكان غري مسؤول XOإىل XDHإن حتول .إصابات نسيجية هامة
XDH أن يرجع ،O2 إىلROS (Harisson ،2002.) األمراض، بإمكان بعضإىل جانب XO أن يلعب دورا
، فقدان وظائف (Hare ،2004و Berry)هاما يف ظهور حاالت مرضية أخرى كأمراض القلب واألوعية

Al-Mouhtaseb)وإلتهاب املفاصل الرثوي وإلتهابات مفصلية أخرى ( 2007وآخرون، Hane)األعصاب
(.2011وآخرون،
XORنزيم مثبطات إ.2.5
األحباث يته ودوره ال تزال مستمرة، فت السنني، إال أن األحباث حول بنمنذ عشرا XORاكتشاف إنزم رغم
يف ظهور رته على املسامهةفهم بنيته وآلية تأثريه، وقدإىل أمهية بالغة وهتدف املتجهة حنو مثبطات اإلنزم متلك
XORإن مثبطات .آليات التثبيط وتتعلق بطبيعة املثبطتتنوع طبيعة متغرية، كما XORمتلك مثبطات . اضمر األ
كال . أو ملواد ليس هلا أي عالقة بنيوية مع املواد البيولوجية لإلنزم puriniqueقد تكون مشاهبة ملادة التفاعل
(. Massey ،1981و Hille) (Mo)القسمني من املثبطات تتفاعل على مستوى موقع املوليبدان
المثبطات المشابهة لمواد التفاعل.2.5.1
املثبط األكثر Allopurinolميثل العديد من املركبات ذات احللقات العطرية غري املتجانسة، يشمل هذا القسم
يستعمل املصلية والبولية حلمض اليوريك، كما لنسببذلك اخمفضا XOميلك نشاطية تثبيطية إلنزم فهوأمهية
.(2002وآخرون، Borges؛ Koij ،1994)كدواء لعالج النقرس
Oxypurinol (isoester de laإىل XORبواسطة Allopurinol (isoester de l’hypoxanthine)يتأكسد
xanthine) ( 2شكل) . هذا األخري ميلك شراهية عالية لإلنزم املرجع(Moıv
قوي شكلب معهيرتبط و (
Massey) وEdmondson ،1970 .)عند الرتاكيز املرتفعة ملركب Allopurinolنافسي ، نه يعمل كمثبط ال ت، فإ
(.2006وآخرون، Pacher)ولكن عند الرتاكيز املنخفضة، فانه يثبط اإلنزم تنافسيا
XORأن يؤثران كمزحيني للجذور احلرة املنتجة من طرف (Oxypurinolو Allopurinol)بإمكان كال املثبطني
(Augustin ،1994وآخرون)، من أجل ذلك ال نستطيع التأكيد هلAllopurinol وOxypurinol ميلكان
(.1987وآخرون، Moorhouse)زاحي للجذور احلرة تأثريا تثبيطيا لإلنزم أو إ

N
N
H
O
NH
N
N
H
O
NH
NO N
H
N
H
O
NH
NO N
H
O
H
N
N
H
N
N
OH
N
N
H
N
N
OH
OH
XanthineHypoxanthine Acide urique
Allopurinol Oxypurinol
XO XO
(.2002وآخرون، Borges) Oxypurinolومستقلباته Allopurinolبواسطة XORتثبيط إنزم . 2شكل
نزيممثبطات أخرى لإل.2.5.2
يت الضروري للعملية التحفيزية الذي يعمل على حترير الكرب Cyanureمثل XORتوجد مثبطات أخرى إلنزم
، Masseyو desulfo (Hilleغري نشط منزوع الكربيت منتجا بذلك شكال thiocyanateعلى شكل Moملركز
،لإلنزم املرجعMo للتثبيط وذلك عرب تفاعله مع الكربيت الضروري للمركز منوذجا مماثال NOميثل (.1981
(.1999وآخرون، Ichimori)ا بذلك الشكل غري النشط منزوع الكربيت منتج
حنو املراكز Moوذلك بتوقيف حركة اإللكرتونات ملركز Moبتفاعله مع XORبدوره إنزم Arséniteكما يثبط
Fe/S وFAD (Stewart ،1984وآخرون.) يعتربTEI-67204-methyl-5-thiozole-carboxyique)(3-
cyano-4-isobutoxyphenyl) مثبطا اصطناعيا إلنزمXOR تثبيطيا من نوع خمتلط، فهو يتثبت ا، وميلك تأثري
وآخرون، (Okamoto بشكل قوي مع الشكلني النشطني لإلنزم والشكل منزوع الكربيت غري النشط لإلنزم
2003 .)

XORاآللية التحفيزية إلنزيم .2.6
ويرافقه كبيورينات وذلك بتحفيزه أكسدة الكزانثني إىل محض اليوريلا هدميف XOR الدور الفيزيولوجي ل مثليت
NAD رجاعإ+ (Olson 1974، وآخرون.) الكزانثني يتم على و أظهرت الدراسات أن أكسدة اهليبوكزانثني
FADبدورها إىل مركز تتحول ، اليت (1999وآخرون، Xia) إلكرتونني لإلنزم، وذلك بانتقال Moمستوى املركز
، وآخرون Olson)لإللكرتونات ازنواليت تعترب كمخ (Anderson ،1991و Hille) Fe2/S2املراكز املتوسطة عرب
NADيرجع . (1974 إىلميكن أن يرجع بشكل ثنائي O2، أما NADH إىل دائما على شكل ثنائي التكافؤ +
H2O2 إىلأو بشكل أحاديO2•- (Nishino وNishino، 1997). ثبت تيNAD
، XDHل FADكز على مر +
NADإىل إلكرتونني نتقليوبذلك (.3الشكل ( )Massey ،1997و Harris) NADHلريجع بصورة ثنائية إىل +
(. 2006وآخرون، (Brondino اإلنتقال اإللكرتوين بني مراكزه و XORاآللية التحفيزية إلنزم . 3شكل
ل غري النشط منزوع املوليبدان أين تكون ذرة الشك( 1: )يف الطبيعة على شكلني غري نشطني XORيتواجد
( S)الشكل غري النشط منزوع الكربيت أين تكون ذرة الكربيت ( 2)غائبة، و ptériniqueاملوليبدان للعامل
XORيوجد شكال غري نشط آخر إلنزم (. Edmondson ،1970و (Massey(O) كسجنيمستبدلة بذرة األ

أكثر من . ولكنه ال يوجد يف احلالة الطبيعية in vitroميكن احلصول عليه ، والذي deflavoيتمثل يف الشكل
؛ 1992وآخرون، Abadeh)املتواجد يف حليب اإلنسان يكون يف شكل غري نشط XORمن إنزم 90%
Godber ،1997وآخرون.)
األنظمة المضادة لألكسدة . 3
املركبات فعالة هذه تعترب و قارنة باملادة املؤكسدة، تعرف مضادات األكسدة أهنا كل مادة تتواجد برتاكيز أقل م
وآخرون، Lobo؛ Nyska ،2002و Kohen)إلكرتوناهتا إىل اجلذور احلرة وبذلك تعدل آثارها اجلانبية متنحألهنا
يتم ختليقها glutathion ،ubiquinol ، uric acidثل تنقسم إىل مضادات أكسدة غري انزميية مكما ؛(2010
عليها من األغذية وال يستطيع اجلسم ختليقها، احلصولوأخرى يتم يتابوليزم الطبيعي يف اجلسم، أثناء امل
، B-caroteneو vitamine E (α-tocopherol) ،vitamine C (ascorbic acid)كالفيتامينات خصوصا
(2010وآخرون، (GPX Loboو CATو SODمضادات أكسدة إنزميية مثل باإلضافة إىل نوع ثالث هي
.(4شكل)
(Pincemailتنظيم إنتاج األنواع األكسجينيية النشطة بواسطة األنظمة الدفاعية املضادة لألكسدة . 4 شكل (.2002وآخرون،

اإلنزيميةالمضادة لألكسدة األنظمة . 3.1
Superoxide dismutase (SOD)انزيم. 3.1.1
الدفاعي الرئيسي ضد جذور فوق األكسيد وميثل خط الدفاع األول ضد اإلجهاد النظامSOD نزم إ يعترب
O2حتويل جذر SODحيفز إنزم و ،يتواجد يف معظم العضيات ويف السوائل خارج خلوية حيث التأكسدي،إىل -•
H2O2 وأكسجني (Das وLewis-Molock ،1997.)
2O2•-
+ 2H+ SOD
H2O2 + O2 ميلك أربع حتت وحدات امليتوكوندرييف Mn-SOD (SOD2)على ثالث أشكال يف اجلسم، SODتواجد إنزم ي
ميلك حتت وحدتني والعوامل املساعدة له يف السيتوزول Cu-Zn-SOD(SOD1)والعامل املساعد له هو املنغنيز و
وحداتميلك أربع حتت والذي ((Cu-Zn-SOD SOD3النحاس والزنك، باإلضافة إىل شكل خارج خلوي هي
(Lobo ،؛ 2010وآخرونGrigorov ،2012).
Catalase (CAT) إنزيم .3.1.2
يتواجد يف خاليا الثديات خاصة على مستوى و .من اإلنزميات اليت حتتوي على جمموعة اهليم CATيعترب
.(Laitonjam ،2012) إىل ماء وأكسجني بريوكسيد اهليدروجني CATحيول إنزم البريوكسيزومات،
2H2O2 CAT
2H2O + O2
Feحتت وحدات، حتتوي كل حتت وحدة يف مركزها النشط على أيون احلديد ربعأمن CATيتكون إنزم 3+
OHيف قدرته على خفض مسية جذر CATيتمثل أحد أدوار إنزم و . NADPHوجزيئة كما . H2O2الناتج عن •
وآخرون، Delattre)ترفع من فعاليته التثبيط و محايته من على تعمل اليت NADPH مبجموعة CATيرتبط
(.Laitonjam ،2012 ؛2005

Glutathion peroxydase (GPX) نزيمإ. 3.1.3
السيلينيوم على معتمد غري على شكلني، أحدمها GPXيتواجد
selenium-independent (glutathione-Stransferase, GST) بينما يعتمد اآلخر على السيلينيوم ،
selenium-dependent (GPx) . قدرهتا على إرجاع يف تتمثل النشاطية املضادة لألكسدة هلذه اإلنزمياتH2O2
(.Grigorov ،2012) إىل ماء أو كحول ويصاحبها أكسدة جزيئات اجلليتاتيون ((peroxide ROOHو
+ 2GSH GPX
GSSG +2H2O H2O2 CATبينما ROSأكثر فعالية مع الرتاكيز العالية ل GPX، إال أن H2O2نفس التأثري على CATو GPX ميلك
(.Grigorov ،2012؛2002وآخرون، (H2O2 Antunes له تأثري عايل مع الرتاكيز املنخفضة ل
Peroxiredoxinsإنزيمات.3.1.4
، وهي من اإلنزميات املكتشفة حديثا وهلا القدرة على thioredoxin peroxidaseبإسم Peroxiredoxins تعرف
تلعب . alkyl hyderoxides(Laitonjam ،2012) مثل بريوكسيد اهليدروجني و اإلرجاع املباشر للبريوكسيدات
Peroxiredoxins من الربوتينات احلرة %1.8-1.0ما يف التخلص من اهليدروبريوكسيدات، إذ متثل ادورا ه
(.2011وآخرون، (GPx Alvarezو CAT انزمياتضعيفة مقارنة ب نشاطيتها ، إال أناخللوية
غير اإلنزيمية المضادة لألكسدة األنظمة. 3.2
ال يتم تركيبها من طرف العضوية ويتم غري اإلنزميية معظم املركباتفإن على عكس اإلنزميات املضادة لألكسدة،
(GSH)من الغليتاتيون املرجع كالجند ضمن قائمة مضادات األكسدة حيث .األغذية عن طريقصول عليها احل
.والكاروتنويدات و عديدات الفينول وغريها Vit Eو Vit Cو

3.2.1 .Vitamine E(Tocopherol)
كسدة متغرية نشاطية مضادة لأل ، ذات δ و γ و β و α عدة أشكالب يف الطبيعة Vit Eيتواجد
(Riicciarelli ،2001.) يعترب الشكل وα أكثر نشاطية، فهو ذو طبيعة ليبيدية ويتثبت على الغشاء اخللوي
peroxyl (LOOويثبط سلسلة تفاعالت فوق األكسدة الليبيدية وذلك بإحتاده مع جذر فيتحول بدوره إىل ( •
LOO)جذر لكنه أقل خطورة من جذر (.Evans ،2000)جزيئة مضادة لألكسدة مع تحد يف ما بعد لي ،(•
له القدرة على إرجاع جذور ذيإما عن طريق محض األسكوربيك وال بطريقتني، -tocopherol αيتجدد
tocophéryle ومن جهة أخرى يتجدد عرب إنزم خاص يعتمد على ،glutathion نزمهو إ
tocophéryle réductase الذي يعمل على إرجاع جذورtocophéryle إىلα- tocophérol يصاحبه حتول ،
(.2005وآخرون، (Delattre (GSSG)إىل احلالة املؤكسدة (GSH)الغليتاتيون من احلالة املرجعة
3.2.2. Vitamine C
. ميثل مضاد األكسدة األكثر أمهية على مستوى السوائل اخللوية ومن املركبات الذائبة يف املاء Vit Cيعترب
يعمل شكله املرجع و . الطور املائي قبل بدأ عملية فوق أكسدة الليبيدات يفاملتواجدة ROSل تعديقوم بيحيث
AH2 كمزيح للجذور وذلك باحتاده مع جذر حرRإىل Vit C ةأكسديصاحبه مستقر و RHيرجع إىل الذي •
AH (.Moussard ،2006)مقارنة باجلذر احلر األويل ضعيفة نشاطية ،حيث ميلك هذا األخري •
O2على اإلزاحة املباشرة جلذور Vit Cيعمل ONOO و •
OHو •• (Evans ،2000) ،له القدرة على جتديد و
–tocopherol α على مستوى األغشية البالزمية والليبوبروتينات وحيافظ على مستوياتGSH داخل اخللية مما
.(Grigorov ،2012)األكسدة للربوتينات من عملية thiolجيعله يلعب دورا هاما يف محاية جماميع
AH2 + R• AH
• + RH

الكاروتنويدات. 3.2.3
محراء وبرتقالية، تتواجد يف معظم اخلضر و هي صبغات صفراءو ( xanthophyllsأو carotens)الكاروتنويدات
ونظرا .(β–cryptoxanthinو β -carotene ،γ-carotene) VitA فتامنيل امعظمها طليع ويعترب والفواكه
كما أن اخنفاض .طة مزدوجة فهي تعمل على إزاحة اجلذور احلرة و إلتقاط األكسجني املهيجإلحتواءها على راب
(.Podsȩdek ،2007)يف املصل يعترب مؤشرا ألمراض القلب واألوعية والسرطان β –caroteneمعدالت
Glutathionببتيد .3.2.4
-Glycineويسمى حبمض thiolحيتوي على جمموعة . هو عبارة عن بيبتيد ثالثي ذو وزن جزيئي ضعيف
Cystein-glutamique شكله املرجع يف (GSH ) وGSSG يف شكله املؤكسد(Halliwell وGutteridge ،
ويعترب مضاد األكسدة األكثر إنتشارا يف هذه ،النواة وامليتوكوندري و يتواجد على مستوى السيتوزول(. 1999
يف اخللية، GSSGيرتاكم الغليتاتيون املؤكسد (. Grigorov ،2012؛ 2005وآخرون، (Masellaاألجزاء اخللوية
(.Grigorov ،2012؛ Droge ،2002)إلجهاد التأكسدي اا عن مؤشرا هاما GSSG /GSH بني ومتثل النسبة
للسمية ويساهم يف نقل املزيلة يتمثل الدور الرئيسي للغليتاتيون يف مشاركته كعامل مساعد للعديد من اإلنزميات
OHر و اض األمينية عرب الغشاء البالزمي وإزاحة جذاألمحO2و •
، وآخرون VitE (Masellaو VitCوجتديد 1
(.Grigorov ،2012؛ 2005
GSH + OH• GS
•+ H2O
GSH + R• GS
•+ RH
المركبات الفينولية. 4
وية نباتية، تتميز بنيتها األساسية بوجود حلقة عطرية أو أكثر مرتبطة املركبات الفينولية عبارة عن مستقلبات ثان
فقد تتواجد املركبات الفينولية على شكل حر أو مرتبطة (. 2004وآخرون، Manach)بعدة جماميع هيدروكسيل
مع سكريات أو أسرتات أو مبلمرة، و بإمكاهنا اإلرتباط مع مكونات اجلدار اخللوي كعديدات السكريات
(.Edeas ،2007)لربوتينات وا

األقسام الرئيسية للمركبات الفينولية. 1 .4
تهاحسب بني املركبات الفينولية ميكن تقسيمو . مركب فينويل خمتلف معروف ببنيات متعددة 8000يوجد أكثر من
isocoumarinو coumarinو ةالفينوالت البسيطة، األمحاض الفينولي: األساسية إىل عدة أقسام
من و .tanninو lignaneو flavonoidesو antraquinone و stilbene و xanthone و naphtoquinone و
؛ 2002وآخرون، Debeer)بني هذه األقسام، متثل األمحاض الفينولية والفالفونيدات األقسام األكثر انتشارا
Dykes وRooney ،2007.)
األحماض الفينولية .1.1 .4
cinnamicمشتقة من محض فينولية أمحاض : ، تقسم إىل قسمني رئيسني(5شكل)ولية عبارة عن مركبات فينوهي
، (C3-C6)ذات هيكل كربوين benzoicمشتقة من محض فينولية و أمحاض ( C1-C6)ذات هيكل كربوين
مع تظهر األمحاض الفينولية على شكل مرتبط كما .القشوريف البذور و تتواجد يف اخلضر والفواكه خاصة و
، وميكن أن تتحول إىل شكل حر (Shahidi ،2010 ،Gotti ،2011و Chandrasekara)سكريات أو أسرتات
.األمحاض أو القواعد أو اإلنزمياتبواسطة بإماهتها
(.Tsao) ،2010البنية الكيميائية لألمحاض الفينولية . 5 شكل

الفالفونيدات .4.1.2
وزن وهي ذات . مركب 4000يف عائلة عديدات الفينول وتتكون من أكثر من جمموعة أهم متثل الفالفونيدات
( Bو Aحلقة )على حلقتني عطريتني C6–C3–C6 يف شكلذرة كربون متوزعة 15 توي علىجزيئي ضعيف، حت
(.6 الشكل) (2006، وآخرون (C Balasundramحللقة تسمى اذرات كربون 3 منمتصلتني بسلسلة
(2008وآخرون، (Cazarolli لعام الفالفونويدات و أهم املواقع املتدخلة يف تأثرياهتا احليويةالشكل ا .6 شكل
يف C3و C2حسب عدد املبادالت اهليدروكسيلية وتواجد الرابطة املزدوجة بني عائلة 13تقسم الفالفونيدات إىل
Isoflavonesو Flavanones و Flavanoles و Flavonoles وFlavones أمهها و ؛C احللقة
شكل مشتقات ميثيلية أو كربيتية بالفالفونيدات ظهرقد ت (.7شكل ) anthocyanidinsأو Anthocyanins و
األمحاض الكربوكسيلية و األمينات و الليبيداتأو مع أو متحدة مع سكريات أحادية أو ثنائية أو متعددة،
(.Reis Giada) ،2013واألمحاض العضوية

(.2006وآخرون، (Bahorunاألقسام املختلفة للفالفونيدات . 7 شكل
الدباغ .4.1.3
الربوتينات برتباط على اإل دالتون، تتميز بقدرهتا العالية 3000إىل 500هي مركبات فينولية ذات وزن جزيئي عايل
ميكن متييز . (2005وآخرون، Macheix) اخلضرو العديد من الفواكه يف مسؤولة عن الذوق وهي ،وتقوم برتسيبها
.الدباغ املميهة والدباغ املكثفةوهي من الدباغ واليت ختتلف عن بعضها بنشاطيتها وتركيبها الكيميائي تنيعمو جم
4z.1.3.1. الدباغ المميهة
ق عن طري فكك بفعل اإلنزميات أوتت ، وويف بعض األشجار يف النباتات ثنائية الفلقة هذه املركبات تتواجد
محض ، أوgallic (gallotannin)محض أمحاضا فينولية من نوعبذلك حمررة و القاعدية،امضية أماهة احلاإل
ellagic (ellagitannin) ( Macheix ،2005وآخرون) ) 8)شكل.

الدباغ المكثفة .4.1.3.2
تتميز بعدم . -(+) catechineاملشتقة من Flavane-3olsزيئات جبلمرة طريق عن تتشكل الدباغ املكثفة
ىل إتحول ت حيث حتت درجة حرارة عاليةو قويةالكيميائية التفاعالت ال إال يف ،للتحلل تهامقاومجتانسها و
املركبات الثنائية واملتعددة القابض يف اخلضر وتدعىالذوق ومتثل املركبات الفينولية املسؤولة عن . صبغات محراء
(.8شكل )( Reis Giada ،2013؛ 2005خرون، وآ (Macheix( proanthocyanidines)بـ
(الدباغ املكثفة) proanthocyanidinsو ( الدباغ املميهة) casuarictin لالبنية الكيميائية . 8 شكل
Reis Giada) ،2013.)

النشاطية البيولوجية لعديدات الفينول. 4.2
النشاطية المضادة لألكسدة للفالفونيدات .1 .2 .4
استخالبنزميات و تثبيط اإلو ROSلـ باشرةبآليات خمتلفة مثل اإلزاحة امل ةلألكسد كمضاداتفونيدات لفالتؤثر ا
.(Halliwell، 1994) املعدنية أو محاية األنظمة الدفاعية املضادة لألكسدة األيونات
نزيماتتثبيط اإل. 1.1 .2 .4
ت دراسة قام هباأظهر حيث xanthine oxydase (Hanasaki ،1994)نزم إلفالفونيدات القدرة على تثبيط ل
Cos (1998)، نزم ثبطة إلالعالقة بني البنية الكيميائية للفالفونيدات ونشاطيتها املXOR، أن كل من حيث وجد
Flavones وFlavonoles ا على الرابطة املزدوجة بني حتواءمهنزم إلا القدرة على تثبيط اإلمهلC2-C3 وغياب
أقل من الفالفونيدات غري XOR الفالفونيدات السكرية متلك نشاطية تثبيطية لن إ. C3على OHجمموعة
أظهرت دراسات أخرى أن و .Quercetinمرات من 10أقل ية نشاط Rutin ميلك ، فعلى سبيل املثالالسكرية
و Cyclooxygenaseنتاج اجلذور احلرة مثل إنزميات أخرى مسؤولة عن إالفالفونيدات هلا القدرة على تثبيط
Lipooxygenase (Landolfi ،1984.) كما بينت النتائج املتحصل عليها من طرفSandhar وآخرون
هي مثبطات apigenineمثل Flavoneو galangine و kaempherol وquercetin كل من أن( 2011)
. Cytochrome P450نزم إل قوية
المعدنية استخالب األيونات .1.2 .2 .4
Feستخالبية لأليونات املعدنية مثل إلفالفونيدات مركبات تعترب اجذور اهليدروكسيل إنتاجمما خيفض من ،+2
(Pietta ،2000 ؛Perron وBrumaghi ،2009؛) . فقد قامVan Acker ظهار آلية إب (1996) وآخرون
يف catecholنواة ب هيستخالاالاملواقع الفعالة املسؤولة عن كانت بعض الفالفونيدات، وقدلاستخالب احلديد

Cو A للحلقتني hydroxyl-5و oxo-4جماميع و Cيف احللقة oxo-4و hydroxyl-3جمموعة و Bاحللقة
.(9 شكل)
(.Pietta) ،2000مواقع ارتباط األيونات املعدنية بالفالفونيدات .9شكل
زاحة الجذور الحرةإ. 1.3 .2 .4
O2: كسجينية مثلذور األرجاع اجلإتعمل الفالفونيدات على •- ،OH
• ،LOLOOو •
وذلك مبنحها هيدروجينا •
، Pietta)مستقرة quinoneعطاء بنية لكرتونا، يف حني تتفاعل اجلذور الفالفونيدية الناجتة مع بعضها إلإأو
. (10شكل) (2000
OH
OH
O
OH
O
OH
O
O
..
R RH R RH
quinoneFlav-OH Flav-O
. ..
.(Pietta ،2000)تفاعل الفالفونيدات مع اجلذور احلرة . 10شكل
واليت تعترب ضرورية Cللحلقة ortho-diphenoliqueالبنية ن النشاطية املزحية للجذور احلرة تتطلب وجود كل منإ
لكرتونات انتقال اإل لىع يساعد oxo-4مع الوظيفة C2-C3الرابطة املزدوجة بني ، وجود لنشاطية الفالفونيدات

C4مع وظيفة الكربونيل يف 5و 3يف الوضع OHجماميع د ، كما أن احتاaroxylمما يعمل على استقرار جذر
يغري من مبجاميع ميثيلية B، أما استبدال جماميع اهليدروكسيل يف احللقة ة للجذور احلرةيزحامل يرفع من النشاطية
.(Reis Giada ،2013 ؛ 2006وآخرون، Balasundram؛ Pietta ،2000) النشاطية اإلزاحية للجذور احلرة
لتهابالنشاطية المضادة لإل.2 .4.2
Galati؛ 1994، وآخرون Da Silva)لتهاب ظهار دور الفالفونيدات املضاد لإلإقامت العديد من الدراسات ب
هلا chrysineو quercetin ،myricetine ،apigenineفالفالفونيدات مثل (. Read ،1995؛ 1994وآخرون،
وآخرون، lypooxygenase (Brunetton ،1993 Iwalewaو cyclooxygenaseنزم إالقدرة على تثبيط
إلتهاب وآثارها على فينوليةالركبات على بعض امل( 2011) وآخرون Kostyukأسفرت الدراسة اليت قام (. 2007
املتسبب يف تصلب ( oxidased LDL)املؤكسد LDLختفض مستويات resveratolو quercetinأن ة،األوعي
O2وإنتاج جذر ل اإللتهايب،األوعية خالل التفاعو resveratol ، كما يعملNOوخفض مستويات -•
verbascoside كيموكيناتالللخاليا الطالئية خمفضا بذلك من التعبري املفرط ل على تعديل اإلستجابة اإللتهابية
.(oxLDL)وجلزيئات اإللتصاق احملفزة ب
التأثير المضاد للسرطان. 3 .4.2
. ديثا حول عزل املركبات الطبيعية كاملركبات الفينولية واختبار قدرهتا كعوامل مضادة للسرطاناجتهت األحباث ح
، myrecetin، ومشتقاها السكرية باإلضافة إىل quercetin ،rutin ،luteolinفعلى سبيل املثال يعمل كل من
rosmarinic acid وcatechin على محايةADN من أضرارROS كما ميثل ،resveratrol مثبطا قويا
مثار العنب وبرتاكيز عالية يف اخلمر األمحر وتقوم بنشاطيتها املضادة للسرطان تتواجد هذه املركبات يف. للسرطان
يف الشاي catechineكما أنه من املعروف أن مركبات . عن طريق حتفيز املوت اخللوي للخاليا الورمية

(epigallocatechin، catechin وepicatechin، وغريها )متلك قدرة عالية مضادة للسرطان
de Mello Andrade) وFasolo ،2014.)
Neurodegenerativeالتأثير المضاد لألمراض العصبية .4 .4.2
وجد أن اإلجهاد التأكسدي يسبب أضرارا عصبية . مت تقدير دور املركبات الفينولية على اجلهاز العصيب املركزي
قام (.Fasolo ،2014و de Mello Andrade)املوت عند املرضى املصابني باحتقان دموي يف الدماغ ويسبب
Itoh بدراسة تأثري ( 2012) وآخرون(-) -epigallocatechin gallate (EGCG) يف الشاي ةواملركبات الفينولي
ذه الدراسة مت معاجلة جرذان مبحلول يف ه. األخضر على تكاثر اخلاليا العصبية اجلذعية يف املكان العصيب املصاب
(EGCG) أسابيع وأظهرت الدراسة النسيجية عند املعاجلة بـ 10مدة (EGCG) (w/v) %0.1مائي حيتوي على
O2)أو امتصاص اجلذور احلرة مثل /أنه يقوم بالتخلص و• ،OH
يتم تثبيط اجلذور احلرة . الناجتة أثناء اإلصابة( •
ربمج للخاليا العصبية اجلذعية هذا ما يسمح حبمايتها يف املراحل األوىل من حدوث احملفزة للموت اخللوي امل
.اإلصابة
الدموية النشاطية المضادة ألمراض القلب واألوعية .5 .4.2
من %68وجد أن األغذية الغنية بالفالفونيدات ختفض 0993وآخرون عام hertogإنطالقا من حبث قام به
(.2008وآخرون، Zabri)وعية مقارنة بغذاء فقري هبا خطر أمراض القلب و األ
تعمل املركبات الفينولية على محاية القلب واألوعية من خالل تثبيط تشكل أوعية دموية جديدة وهجرة اخلاليا
األوعية يعود إىل احلماية من حدوث قفارالتأثري الفعال للمركبات الفينولية على ا يعتقد أن. والتضاعف يف األوعية
ختثر الدم من خالل تثبيط نشاط الصفائح الدموية، أو من خالل خفض تعبري اجلزيئات املولدة للتخثر أو املولدة
(.Fasolo ،2014و khoo، de Mello Andrade 2000و Ong (لتصلب األوعية الدموية

Peganum harmalaنبات الحرمل .5
African، يف إيران Espandيف مشال افريقيا و harmelبإسم .Peganum harmala Lيعرف نبات احلرمل
rue وMexican rue و Turkish rue يف الواليات املتحدة(Mahmoudian ،2002وآخرون .) ينتمي نبات
يف املناطق شبه الصحراوية cm 100-30، فهي عشبة معمرة، قد تصل إىل Zygophylaceaاحلرمل إىل عائلة
يتوزع نبات احلرمل بشكل واسع يف وسط آسيا و (.2002وآخرون، Mahmoudian)والسهوب واألتربة الرملية
(.2002وآخرون، Mahmoudian)مشال إفريقيا والشرق األوسط، كما مت جلبه إىل أمريكا وأسرتاليا
، Unverzagt)يتكاثر نبات احلرمل أساسا عن طريق البذور، كما له القدرة على التكاثر عرب اجلذور املصابة
يتميز بسيقان مستقيمة كثرية التفرع، حتمل أوراقا متناوبة مقسمة إىل أشرطة ضيقة، وأزهار أحادية تبلغ (. 2013
أما البذور فهي صغرية كثرية العدد تتميز باحتواءها على . ذات لون أبيض مصفر وعروق خضراء mm 30-25من
تكون فرتة إزهار نبات احلرمل من شهر (.Iserine ،2001)زوايا ثالثية الشكل ذات لون بين قامت وذوق مر
(. 11شكل( )2011وآخرون، Dube) مارس إىل أكتوبر

بذور ال: Dالثمار، : Cاألزهار، : Bالنبتة بأكملها، : A. نبات احلرمل واألجزاء املختلفة املكونة له. 11 شكلAsgarpanah) وRamezanloo ،2010.)

اإلستعمال التقليدي للنبتة. 5.1
فبذوره تستعمل كمدرة للحيض يستعمل نبات احلرمل منذ زمن قدم يف العالج عند العرب يف الطب الشعيب،
Lamchouri)ستعمل كمضادة للعقم يف مشال إفريقيا والشرق األوسط ، كما تستعمل كمخفضة للحمى و ت
مدر للحليب، كما تستعمل البذور يف املغرب كطاردة للديدان و (Winston ،2000و Kuhn ؛1999وآخرون،
ومتلك النبتة تأثريات أخرى ضد ( Bellakhdar ،1997؛ 1995وآخرون، Munir)ومضادة للتشنجات وللتقيؤ
؛ 1998وآخرون، Kholmatov)الزكام ونزالت الربد، أما العشبة ككل فهي تستعمل للتبخري والتهابات األعصاب
Sezik ،مسببة للهذيان ومنومة لكنها و مدرة للبولعمل كمساعدة للهضم و أما الثمار فهي تست(. 2004وآخرون
(Kartal ،؛ 2003وآخرونGoel ،2009وآخرون .)(البطن أمل )املغص الربو و ملعاجلة أما األوراق فتستعمل ،
Ezer؛ Basu ،1995و Kirtikar)والروماتيزم احلادةاآلالم العصبية اهلسترييا و احلازوقة و و انقطاع احليض و
(. Iserine ،2001)ملعاجلة مرض باركنسن Harmineكما يستعمل (. Mumcu ،2006و
Asparganah)واليت تتواجد أساسا يف اجلذور و البذور %5إىل 2 بني احمتواها من القلويدات ميرتاوح
، Harmine ،Harmalineمثل B-carbolinesالقلويدات يف مركبات ، وتتمثل(Ramenzanloo ،2012و
Harmalol وHarmane (Mahmoudian ،2002وآخرون )وHarmalidine ،Ruine
Vascine (Peganine)مثل Quizolineومركبات Tetrahydroharmine (Khan ،1990 .)و
Mahmoudian)الناضجة بذور غري الناضجة يكون أقل من البذور توى القلويدات يف الإن حم. Vasicinoneو
Orientineو Vitexineحيتوي اجلزء اهلوائي لنبات احلرمل على الفالفونيدات وتشمل (. 2002، وآخرون
ر يف البذو Quercetinو Kaempherol، يف حني يتواجد (2012، وآخرون Khan) Luteolineو Hypersideو
(Pulpati ،2008وآخرون) باإلضافة إىل تواجد فالفونيدات سكرية يف اجلزء اهلوائي وهي ،acacetin-7-O-
rhamnoside وO-glycosyl-2’’-O-(3’’’-acetylrhamnosyl) glucoside ،7-O-(2’’’-O-rhamnosyl-

2’’’-O-glucosylglucoside وglucoflavone-2’’’-O- rhamnosyl-2’’-O-glucosyl-cytoside
(Sharef ،1997.)
Moghadam؛ Jhon ،1999و Prashanth)متلك بذور نبات احلرمل نشاطية مضادة للبكترييا والفطريات
وآخرون، Jahaniani؛ 2000وآخرون، Lamrouchi)، ومضادة للسرطان (Nennah ،2010و 2010وآخرون،
وآخرون، Baghiani؛ 2012وآخرون، Djarmouni؛ 2007وآخرون، Moura)، ومضادة لألكسدة ( 2005
Berraouagui)ونشاطية موسعة لألوعية ( 2009وآخرون، Im)ومضادة لتكتل الصفائح الدموية (. 2012
(.2008وآخرون، Astulla؛ 2006وآخرون،
التأثير السمي لنبات الحرمل. 5.2
أن بعض الدراسات أظهرت اجلانب السمي بالرغم من خمتلف النشاطيات البيولوجية اليت ميلكها نبات احلرمل، إال
B-carboline Alcaloides (Harmine ،Harmalineفهضم النبتة ونظرا الحتواءها على مركبات . هلذا النبات
حتفز اجلهاز العصيب B-carbolineيسبب تأثريات مسية خمتلفة، كما أن مركبات ( Tetrahydroharmineو
وآخرون، Frison)ة األمينية، أو التثبت مباشرة على املستقبالت النوعية املركزي بتثبيط أيض النواقل العصبي
(.2008وآخرون، Bukhari)على خفض ضغط الدم Harmineكما يعمل (. 2008
Santolina chamaecyparissusنبات الجعدة . 6
Cyprèأو Aurone femelleأو Santolina incanaأو Santolina petit cyprèتدعى نبتة اجلعدة بالفرنسية
de jardin أما باإلجنليزية فتسمى ،Lavender cotton و باإليطاليةSantolina.
متوضعة عموديا تنتهي مبجموع زهري ، قد تصل اخشبية ذات رائحة قوية، متلك فروععبارة عن نبتة نبتة اجلعدة
6إىل 2وراقها حلمية ضيقة جدا وال تتعدى أ. سم، بينما املزروعة تبلغ مستويات معتربة 50النبتة يف الرباري إىل
(.12شكل ( )Fournier ،1948)و األراضي احلصوية مم طوال ذات لون أبيض خمضر، جندها يف التالل اجلافة

. ، مظهر عام لنبتة اجلعدةSantolina chamaecyparissus :Aالشكل املورفولوجي لنبات اجلعدة . 12شكل
Bاألوراق ، .C، األزهار.
اإلستعمال التقليدي للنبتة. 6.1
والدورة الشهرية، متلك األوراق العديد من اخلصائص فهي حمفزة ومنشطة وتستعمل أحيانا ضد اضطرابات املعدة
مضادة للهسترييا، و احتقان الكبد ، هلا تأثريات(Debuigne ،1974)كما أهنا مثرية للشهوة و طاردة للديدان
هي األكثر نشاطا الزيوت و بذور إال أن ال ،مضادة للديدانمتلك النبتة خصائص . املتناوبةمى والطحال وضد احل
(Fournier ،1984.)

spathulenol (82 - 45 %)من تتميز نبتة اجلعدة بغناها بالزيوت األساسية، فهي حتتوي على نسبة عالية
camphor (7,1 %) ،L-cadinol (5,8 %) ، Methoxy-1,2-dihydrobenzen(2,83 % ) و
كما حتتوي مستخلصات النبتة على الفالفونيدات cis-chrysanthenol (2,52 %( )El Rafari ،2008 .)و
، glucoronides)وكذلك مشتقاهتا السكرية luteolineو apigenineمثل Flavoneخاصة
rhamnoglucoside ) ومركباتFlavone methoxylées مثلpectolinaringinine ،hispiduline،nepetine
،jaceosidine ومركباتFlavonol methoxylées 7يف الوضع-glycosyl (Giner Pons وRios
Canavate ،2000) باإلضافة إىل مركبات ،sesquiterpene من نوعfarnesane ،germacrane ،eudesmane
(Sanz ،؛ 1991وآخرونBarrero 1999ن، وآخرو ) و على جماميعacétylénique (Christensen ،1992 ) و
Coumarin (Ferrari ،2005وآخرون.)
Giner)تأثريها املسكن لألمل وآخرون Gingerمتلك نبتة اجلعدة العديد من التأثريات البيولوجية، فقد أظهر
القدرة على تثبيط التقلصات ، كما أن ملستخلصات نبتة اجلعدة(2000وآخرون، De logu؛ 1988وآخرون،
De؛ 1988وآخرون، Giner)العضلية احملفزة باهلستامني والسريوتونني، باإلضافة إىل نشاطيتها املضادة لإللتهاب
logu ،يف دراسة أخرى أجريت من طرف (. 2000وآخرونSala وجد أن نبتة اجلعدة متلك تأثريا مثبطا وآخرون ،
، كما أن مستخلصاهتا اهليدروكحولية هلا تأثري واقي للكبد (in vivo و in vitro) Phospholipase A2لنشاطية
(Dhanabal ،2012وآخرون .) كما تستعمل النبتة كذلك ملعاجلة العديد من اإللتهابات اجللدية(Da Silva ،
(.1997وآخرون، Suresh)، ومتلك زيوهتا األساسية تأثريا مضادا للفطريات (2004

يالجزء العمل
المواد و الطرق. 1
المواد المستعملة. 1.1
المواد البيولوجية .1.1.1
من منطقة 2012يف جوان جلبت نبتة اجلعدةو من منطقة نقاوس والية باتنة، 2010يف مارس جلبت نبتة احلرمل
ت، كلية النباالبيئة وبيولوجيا قسم ،سنيح من طرف األستاذ الدكتور لعور نيفهماصمت ت، و عني آرنات بسطيف
.حليب البقر الطازج من مزرعة بضواحي سطيفمت جلب .سطيفامعة علوم الطبيعة واحلياة جب
مت احلصول عليها ،غ 30ىل إ 25، يرتاوح وزهنا بني Swiss albinosعلى فئران ذكور in vivoأجريت الدراسة
. الغذاء املصنع اخلاص هبا دملفرتة تأقلم مدة أسبوع، مع تقأخضعت الفئران . من معهد باستور باجلزائر
واألجهزة المواد الكيميائية .1.1.2
من .Flukaو Sigma-Aldrichمن هامت شراءواليت واحملاليل العضوية استعملت العديد من املركبات الكيميائية
-UV/VISوجهاز التحليل الطيفي ( BUchi461)جهاز التبخري ،بني األجهزة اليت استعملت أثناء البحث
8500TechomP ومحام مائيMemmert وجهاز الطرد املركزيSigma 1-15K.
طرق العمل. 2
لفينولية استخالص المركبات ا.2.1
Markham (1982.)اإلستخالص العديد من املذيبات القطبية وغري القطبية حسب طريقة استعمل يف طرق
يعرض (. % 85) يثانولمة، يضاف إليها غ من الوزن اجلاف للنبت100تعتمد طريقة اإلستخالص على أخذ
ن زجاجي، مث حتفظ الرشاحة قطبعدها يرشح احمللول عرب، °م 4اخلليط لعملية رج مدة ليلة كاملة يف درجة حرارة
ميثانولويعاد استخالص الراسب مرة ثانية بنفس الطريقة ولكن باستعمال °م 4األوىل املتحصل عليها يف

احتني مث تعرض لعملية تبخري باستعمال جهاز التبخري لنحصل يف النهاية على املستخلص متزج الرش(. % 50)
للحصول على مستخلص عدة مرات ل اهلكسانب يبيدات من هذا األخري بالغسلنزع الليتم (. CrE)اخلام
معطيا يةبالكلوروفورم إلستخالص الفالفونيدات غري السكر ائي املتبقي اجلزء امليتم غسل . (HxE)اهلكسان
إيثيل األسيتات إلستخالص الفالفونيدات ف له ، أما اجلزء املائي الناتج فيضا((CHEمستخلص الكلوروفورم
حيتوي املستخلص املائي النهائي . ((EAEللحصول على مستخلص إثيل األسيتات أحادية وثنائية السكر
(AqE) إىل حني °م -20فظ املستخلصات حتت حت. على الفالفونيدات األكثر قطبية ثنائية وثالثية السكر
.(13 شكل)استعماهلا

(.(Markham 1982 نبات احلرمل واجلزء اهلوائي لنبتة اجلعدة حسببذور من طرق استخالص الفالفونيدات. 13شكل
اإلستخالص باهلكسان
85%اإلستخالص باملثانول -
ساعة 24الرج مدة -
ح بالقطن الرتشي -
50%اإلستخالص باملثانول -
ساعات 4الرج مدة -
الرتشيح بالقطن -
40 °م تبخري يف درجة حرارة
(فالفونويدات سكرية)جزء عضوي
اجلزء املائي
اإلستخالص باإلثيل أسيتات
اإلستخالص بالكلوروفورم
(فالفونويدات سكرية)جزء عضوي
اجلزء املائي
الرتشيحالرتشيح بورق -
م °40تبخري يف درجة حرارة -
(خام)مستخلص امليثانول
اجلزء املائي
2+1 2راسب 2رشاحة رشاحة
اجلزء اهلوائي لنبات اجلعدة / نبات احلرملبذور غ من 100
1 راسب 1رشاحة
(لبيدات)جزء عضوي

تقدير عديدات الفينول الكلية .2.2
حيث تعتمد على (. 2007 ،وآخرون (Folin-Ciocalteu Lieباستعمال طريقة مت تقدير عديدات الفينول الكلية
ة اجملاميع اهليدروكسيلية الفينولية، مشكلة ناجتا أزرقا يف وسط بواسط Folin-Ciocalteuإرجاع مكونات كاشف
ج تر . مرات 10املخفف Folin-Ciocalteuمن حملول 1mlمن املستخلصات يضاف هلا µl 200نأخذ .قاعدي
من حملول كربونات الصوديوم µl 800ضاف يدقائق، بعدها 4األنابيب وحتفظ يف درجة حرارة املخرب ملدة
(g/l 75)765تقرأ اإلمتصاصية عند . ، وحتفظ األنابيب يف درجة حرارة املخرب ملدة ساعتني nm . ويستعمل محض
يتم التعبري عن النتائج مبكافئ محض الغاليك (. 14 شكل)لتحديد منحىن العيارية ( µg/ml 160-20)الغاليك
.املستخلص لكل غرام من وزن
حلمض الغاليك منحىن العيارية. 14شكل
لكمي للفالفونيداتالتقدير ا .2.3
من 1ml، يؤخذ (1996)وآخرون Bahorunحسب AlCl3مت التقدير الكمي للفالفونيدات بطريقة
دقائق من احلضن تقرأ 10، بعد % 2برتكيز AlCl3من حملول ml 1 املستخلصات املستعملة ويضاف هلا
لكل من العيياري نيدات انطالقا من املنحىنمت حساب تركيز الفالفو . nm 430تصاصية عند طول موجة اإلم

quercetin وrutin برتكيز يرتاوح بني(40-1 µg/ml.) يعرب عن النتائج بعدد امللغرامات املكافئة لكل من
quercetin وrutin (15 شكل)لكل غ من املستحلص.
.rutinو quercetin ل منحىن العيارية .15شكل
حليب البقر من XORتنقية إنزيم .2.4
تعتمد هذه الطريقة على .(2003 ،وآخرون (Baghianiمن حليب البقر بطريقة XORمت فصل وتنقية إنزم
تسرتجع بعدها ،(°م 4دقيقة حتت / دورة 5000)استخالص القشدة من حليب البقر بعد عملية طرد مركزي
من mM 1حيتوي على ، ((K2HPO4 0,2 M جم من حملول فوسفات البوتاسيومويضاف هلا ضعفي احل القشدة
EDTA 10و mM منDTT وmM 1,5 منSalycylate و mM 0,1 منPMSF يرتك املزيج حتت الرج ،
طايف دقيقة، يرشح اجلزء ال 30دقيقة ملدة /دورة 6000، بعد ذلك جترى له عملية طرد مركزي مدة ساعتني
تدرجييا (حجم/حجم % 15)األمونيوم يتاتكرب نضيف للمحلول املرشح و املتحصل عليه عرب قطن زجاجي
دقيقة /دورة 10000عملية طرد مركزي ، مث يعرض لساعةيرج احمللول ملدة (. حجم/حجم % 15)البيتانول البارد و
يرسب اإلنزم من مث ساعةمع الرج ملدة تدرجييا (حجم/وزن % 20)كربيتات األمونيوم يضافدقيقة، 20ملدة

مدة ( دقيقة/دورة 11000)يعرض احمللول اإلنزميي املرشح للطرد املركزي ،رب القطن الزجاجياجلزء الطايف للمرشح ع
/ Na2HPO4)احمللول املنظم اهليباريين دقيقة و يتم اسرتجاع الطبقة البنية احلاوية على اإلنزم مث يتم إذابتها يف 30
NaH2PO4 )(25 mM)1 ، حيتويmM منEDTA ذوpH 6,2 نزميي الناتج لعملية ميز ول اإل، ويعرض احملل
18000)وائب غري الذائبة بالطرد املركزي يتم التخلص من الش. ضد نفس احمللول املنظم ملدة ليلة كاملة
، والناتج النهائي املتحصل عليه يدعى µm 0,45، مث يرشح احمللول عرب غشاء بقطر ساعةملدة ( دقيقة/دورة
.XORباملستخلص اخلام إلنزم
مبعدل تدفق NaCl (ml 30)من M 1 حيتوي املنظم اهليباريين احمللولأغاروز باحمللول -اهليبارينمود يغسل ع
ويتم التخلص من الربوتينات ضعيفة اإلرتباط سا،/مل 10سا، مث حيقن اإلنزم اخلام يف العمود بتدفق /مل 10-15
املنظم اهليباريين، ويتم اسرتجاع اإلنزم باحمللول NaClمن M 0,02املنظم اهليباريين حيتوي بغسل العمود باحمللول
Bicine (mM 05pH= 8,3 ; )بعدها يعرض اإلنزم املنقى لعملية ميز ضد احمللول املنظم ، NaClمن M 0,1 به
.°م 4مجيع خطوات التجربة متت يف درجة حرارة . إىل حني استعماله °م 20-مث حيضن يف ملدة ليلة كاملة،
BXORراسة درجة نقاوة د. 2.4.1
فوق بنفسجي -تم إجراء مسح طيفي مرئيياملتحصل عليه من كروماتوغرافيا الشراهة BXORللتأكد من نقاوة
لProtein Flavin Ratio (PFR )، حيث نسبة الربوتني إىل الفالفني 700إىل nm 250لإلنزم من طول موجة
(A450 / A280)، درجة عالية من نقاوة اإلنزم ل متث 5أو القريبة من املساوية(Bray ،1975وآخرون .)
SDSالفصل الكهربائي على هالم األكريالميد المتعدد في وجود . 2.4.2
وفق SDSبعملية فصل كهربائي على هالم األكريالميد املتعدد يف وجود نا، قمنقاوة اإلنزم املتحصل عليه ملعرفة
Tris-HCl (Mيف حملول ( T ، 2,74 = % C % = 10)الفصل مت حتضري هالم . Laemmli (1970)طريقة
0,125 pH=6,8 ; )مضاف إليه SDS (% 0,1 حجم/وزن ) وTEMED (0,042 % حجم/ حجم ) وفوق

Tris- HCl (M 0,125 pH=6,8 ; ) هالم الرتكيز حمضر يف حملول(. حجم/وزن % 0,1)كربيتات األمونيوم
% 0,1) وفوق كربيتات األمونيوم( حجم/حجم % 0,1) TEMEDو ( حجم/وزن % 0,1) SDSمضاف له
5ملدة °م 100بالتسخني يف ( مل/مغ 1تقريبا )وبروتينات العيارية ( مل/مغ 1تقريبا )حتضر العينات (.حجم/وزن
Tris- HCl (mM 45,6 pH=6,8 ;) ،SDS، املكون من حملول (حجم/حجم)دقائق بعد إذابتها يف حملول العينة
تعامل (. حجم/حجم % 0,005)وأزرق الربوموفينول ( حجم/حجم % 20)غليسريول ،(حجم/وزن % 2)
Tris-HClتتم اهلجرة الكهربائية يف حملول منظم . بنفس الطريقة (MW-SDS-200 Kit, Sigma) الشواهد
(mM 25 ) و غليسني(mM 091) حاوي علىSDS (0,1 % حجم/وزن ) حتت شدة تيارmA 10 حىت
تثبت وتلون العينات املفصولة ملدة ساعة على األقل (. mA) 37-45 الم الفصل، مث ترفع إىلنات إىل هدخول العي
و أواملثانول ( حجم/حجم % 10)مذاب يف محض اخلل ( حجم/وزن % 0,2) يف حملول التلوين أزرق كوماسي
% 10-5)أما إزالة الصبغة الزائدة فتكون بوضع اهلالم يف حملول اخلل (. حجم/حجم % 45)اإلثانول
(.حجم/حجم 5 %)يف وجود أو غياب اإليثانول ( حجم/حجم
نزيمنشاطية اإل تقدير. 2.4.3
بقياس تركيز تراكم إنتاج محض اليوريك باستعمال معامل nm 295 قدرت النشاطية الكلية لإلنزم عند طول موجة
ε = 9600 M)اإلمتصاصية 1-
Cm1-
NADمن µM 500ل و تفاع من الكزانثني كمادة µM 100يف وجود و ،( +
أما نشاطية النمط املؤكسد فتم . pH 8,3ذو Bicineكمادة مستقبلة لإللكرتونات مذابة يف احمللول املنظم
NADتقديرها بنفس الطريقة لكن يف غياب + (Avis ،1956وآخرون).
نشاطية النمط املرجع+نشاطية النمط املؤكسد = حيث النشاطية الكلية لإلنزم
من رامغلبري عن النشاطية اخلاصة لإلنزم بعدد النانوموالت من محض اليوريك الناجتة يف كل دقيقة لكل ممت التع
n mol.min)اإلنزم 1-
.mg-1
)

S. chamaecyparissusو P. harmalaلمستخلصات in vitro النشاطية المضادة لألكسدة.2.5
XOتثبيط نشاطية . 2.5.1
وذلك بقياس XO لـستخلصات نبتيت احلرمل واجلعدة على نشاطية النمط املؤكسد مت دراسة التأثري التثبيطي مل
عند طول موجة XORمن انزم 10µlويف وجود من الكزانثني µM 100كمية محض اليوريك الناجتة عن أكسدة
295 nm (Bray ،1975) 50املذابة يف احمللول املنظم mM Na2HPO4 / NaH2PO4) ) ذوpH = 7,4 الذي و
مت يف وجود تراكيز متغرية من مستخلصات نبات احلرمل ونبات اجلعدة، و EDTAمن mM 0,1 على حيتوي
شاطية يف غياب النعتربت. XOم لـكشاهد للمقارنة باعتباره املثبط املتخصص Allopurinolل ااستعم
باتباع القاعدة وقد مت حساب نسبة التثبيط % 100هي النشاطية الكلية (Allopurinolمستخلصات أو )
:التالية
% Inhibition = (A1 - A2) / A1 × 100
A1 : نشاطيةXO يف غياب املثبطات.
A2 : نشاطيةXO يف وجود املثبطات.
O2 لجذرالتأثير اإلزاحي . 2.5.2 XOالناتج عن -•
O2يعمل جذر Cyt-cعلى إرجاع XOالناتج عن -•
الذي ميلك امتصاصية عظمى عند طول موجة ) 3+
550 nm ) إىلCyt- cCyt- cوجود مركبات مزحية هلذا اجلذر ميكن أن متنع إرجاع ف عليه ، و +2
حيتوي وسط . +3
Cyt-c من µM 25من الكزانثني و µM 100 التفاعل علىتراكيز متغرية من مع XORمن انزم 10µl و 3+
، pH=7,4 وmM 50تركيز ذو( (Na2HPO4 / NaH2PO4مستخلصات كال النبتتني املذابة يف احمللول املنظم
% Inhibition = Ac - As / Ac × 100 :كاآليتCyt C (I % ) ومت حساب نسبة تثبيط إرجاع
Ac :إمتصاصية الشاهد
As :اإلمتصاصية يف وجود املستخلصات

Cyt c (IC50 )إرجاع من % 50 ل مت التعبري عن النتائج بالرتكيز املثبط
DPPHجذر زاحة إإختبار . 2.5.3
DPPH، حيث يتم إرجاع جذر DPPHإزاحة اختبار بجلذور احلرة زاحة االنشاطية املضادة إلمت تقدير
(2,2’ diphenyl-1-picrylhydrazyl) ركبات مزحية للجذور احلرة، إىلذو اللون البنفسجي الداكن مب
2,2’-diphenyl-1-picrylhydrazine معطيا اللون األصفر (Cuendet ،؛ 1997وآخرونBurits وBucar ،
.نانومرت 517عند طول موجة ويقرأ اللون ،(2000
DPPHمن احمللول امليثانويل ملركب µl 1250مع اتيةالنبلمستخلصات من خمتلف الرتاكيز احملضرة ل µl 50 نأخذ
النتائج احملصل عليها يتم مقارنةحيث . nm 517دقيقة، تقرأ اإلمتصاصية عند 30بعد و 0,004 % الرتكيز ذو
%) (Iللمستخلصات DPPHجذر إزاحة تقاس نسبة. الذي استعمل كشاهد BHTلكل مستخلص مع مركب
:كالتايل
% I = ( (AC - AE) / Ac) /100
AC : (الشاهد)اإلمتصاصية يف غياب املثبط
AE : (العينة)اإلمتصاصية يف وجود املثبط
لكل مستخلص من منحىن نسب التثبيط DPPH (IC50 )ن نشاطية جذر م % 50 ـالرتكيز املثبط ليتم حساب
(% I) ضد تركيز املستخلصات.
ABTS إزاحة جذر إختبار. 2.5.4
ABTS على قياس قدرة املركبات املضادة لألكسدة على إزاحة جذر ABTSيعتمد اختبار •+
2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonate)( ) Re ،ينتج جذر (.1999وآخرونABTS•+
املاء يف 2,45 potassium persulfate (K2SO5)) (mM و 7 ABTS) (mMكل من تفاعل بعد الكاتيوين

ABTSبعدها يتم إذابة حملول . سا 12درجة حرارة الغرفة مدة و املقطر، يرتك املزيج يف الظالمول إىل نيف امليثا +°
.nm 734عند 0,70غاية الوصول إىل إمتصاصية
ABTSمن حملول ml 1يبدأ التفاعل مبزج من املستخلصات النباتية احملضرة وتقرأ اإلمتصاصية بعد µl 50مع +°
Trolox (6-Hydroxy-2,5,7,8-tetramethylchroman-2-carboxylicيستعمل .دقيقة ضد املثانول 30
acid) تقاس نسبة التثبيطو ،كشاهد للمقارنة I) (% يلللمستخلصات كالتا:
% I = (AC - AE) /Ac) / 100
AC : (الشاهد)اإلمتصاصية يف غياب املثبط
AE : (العينة)اإلمتصاصية يف وجود املثبط
β –carotene/linoleic acid تبييض إختبار. 2.5.5
أكسدة محض اللينولييك الناجتة من البريوكسيدات س قدرة املستخلصات على تثبيطيعتمد هذا اإلختبار على قيا
(Aslan ،2006 ؛Barriere ،2001وآخرون.) ة يتم إذابة بداي mg 0,5 منβ -carotene يفml 1 من
يتم تبخري الكلوروفورم عند . Tween 40من mg 200من محض اللينولييك و µl 25الكلوروفورم، مث يضاف هلا
حيقن بغاز األكسجني ) من املاء املقطر املشبع باألكسجني ml 100يضاف بعدها . عرب جهاز التبخري °م 40
من اخلليط السابق ويوضع يف أنابيب ml 2,5يؤخذ . مع حتريك اخلليط بشدة (ساعة قبل االستعمالدة مل
BHTونستعمل mg/ml 2من كل مستخلص حمضر لكال النبتتني برتكيز µl 350اإلختبار، ويضاف له
(mg/ml 2 )490أ اإلمتصاصية عند تقر . انسالب انكشاهد موجب واملثانول واملاء كشاهد nm خالل أزمنة ،
للمستخلصات النشاطية املضادة لألكسدةويتم حساب . سا 24سا، 6سا، 4سا، 2سا، 1سا، 0متتالية
:املعادلة التاليةب( % AA)النباتية
AA = AE / ABHT × 100 %
AE :إمتصاصية العينة. ABHT : إمتصاصيةBHT.

نختبار استخالب المعادإ. 2.5.6
Fe(П)-Ferrozineبقياس تثبيط تشكل معقد ف املستخلصات النباتيةاستخالب املعادن من طر قدرت نشاطية
من µl 250ميزج .(2007) وآخرون Leاملعدلة من طرف Welch (1990 )و Deckerحسب طريقة
µl 50يضاف دقائق 5ميثانول بعد µl 450 وFeCl2 (mM 0,6 )من حملول µl 50مع املستخلصات النباتية
اصية عندصدقائق من احلضن يف الظالم تقرأ اإلمت 10، ترج األنابيب وبعد Ferrozine (mM 5)من حملول
nm 562 ،ستعمل مركب يEDTA (0-40 µg/ml ) ب نسبة استخالب أيونات احلديد حتسكشاهد للمقارنة و
Chelation = [1- (AC - AE) × 100] % :كما يلي
Ac :إمتصاصية الشاهد
AE :اإلمتصاصية يف وجود املستخلصات
Ferric thiocyanate (FTC)إختبار . 2.5.7
حسب طريقة بيداتيعتمد هذا اإلختبار على قياس كمية البريوكسيدات الناجتة يف املرحلة األولية لفوق أكسدة اللي
(Chang ،2002وآخرون). حيضر حملول acid linoleic من بإضافة كلacid linoleic (155 µl) tween 20 +
(158 µl) + املستخلصات(2 mg/ml )يف ml 50 من احمللول املنظمml) (pH=7, 0,02M , 50 .املزيج ضنحي
من احمللول السابق ويضاف ml 0,1يتم تقدير مستويات فوق أكسدة اللبيدات بأخذ يف الظالم °م 40 درجة عند
FeCl2 (M ,0,02 من ml 0,1 ، مع إضافةNH4CN6 (05 % ) من ml 0,1 و 4,7ml (75 % ) له اإليثانول
3,5 % HCl .) دقائق وتقرأ اإلمتصاصية عند 3بعد مزج اخلليط، حيضن ملدةnm 500 سا 24وذلك بعد كل
تعطى قيمة نسبة تثبيط فوق أكسدة محض اللينولييك . إىل غاية وصول قيمة إمتصاصية الشاهد إىل أقصاها
Inhibition % = (1-AbsS / AbsC) × 100 :لعالقة التاليةبا
AbsS :إمتصاصية العينة AbsC :إمتصاصية الشاهد

TBAإختبار . 2.5.8
حسب الناجتة يف آخر مرحلة من مراحل فوق أكسدة الليبيدات MDAتعتمد هذه التقنية على تقدير كمية
حملول محض اللينولييك مع ) FTCلتقنية السابق اخلليطمن نفس 1ml فتضا(. Tunalier ،2007)طريقة
TCA (Trichloroacetic acid )من ml 2مع أيام 5مت حضنه ، الذي(FeCl2 و NH4CN6اإليثانول و
جلليدبعد التربيد يف او دقائق، 10مدة ( °م 80)يوضع املزيج يف محام مائي .ml 2 TBA و( حجم/وزن % 20)
كلما .nm 532دقيقة، وتقرأ امتصاصية اجلزء الطايف عند 20دة دقيقة مل/ دورة 3000رد مركزي عند نقوم بط
.خفض القدرة التثبيطية للمستخلصدلت على كانت اإلمتصاصية عالية
:وتقاس نسبة التثبيط باستعمال العالقة التالية
Inhibition % = (0- AbsExt /Abs C+)×100
AbsExt:لعينةإمتصاصية ا
AbsC+ :إمتصاصية الشاهد
AAPH باستعمال in vitro لتحلل كريات الدم الحمراءالنشاطية المضادة .2.5.9
عني فئرانوريد دم من ال قمنا بنزع .(2006 وآخرون، Girard)تبعا لطريقة مت قياس النشاطية املضادة لتحلل الدم
للتخلص من البالزما، بعدها (دقيقة 10دقيقة، /دورة 6000) املركزي تعريضه للطرديتم ،EDTA يف أنبوب حيتوي
Tampon phosphate 10mM)تغسل كريات الدم احلمراء ثالث مرات بواسطة حملول نظامي
NaHPO4/pH7,4/ 125mM NaCl). مركب مت استعمالAAPH ( جزيئاتAZO) ، الذي ينتج عن جتزئه يف
ذور احلرة لكريات الدم ل حتفيز سلسلة أكسدة اجلمن أج ،وسط مائي ويف وجود األكسجني جذور البريوكسيل
يف Tampon phosphate (80µl )من املعلق اخللوي لكريات الدم احلمراء مع حملول %2يتم حضن .احلمراء
S. chamaecyparissusو P. harmalaنبتيت لمن املستخلص املثانويل ل( µl) 20مع إضافة Elisaصفائح
، مث حتضن mg/ml 0,1 مستخلص الكلوروفورم وإثيل األسيتات للنبتتني برتكيزو اخلام للنبتتنيواملستخلص

°م 37يف ) mM) 300 AAPHمن مركب ( µl) 136املستخلصات يف وجود أوغياب ساعات ونصف 4ملدة
حتسب . nm 620متصاصية عند تايل لكريات الدم احلمراء بتتبع إخنفاض اإلخنفاض املتويتم مالحظة اإلتقريبا
:حتلل كريات الدم احلمراء بالعالقة نسبة
I% = (Abs t0- Abs tX) × 100 / (Abs t0- Abs tfinale)
Abs t0 : اإلمتصاصية يف الزمنt = 0 mn
Abs tX : اإلمتصاصية يف الزمنt = x
Abs tfinale :يف الزمن اإلمتصاصيةt = 270 mn
.Sو P. harmalaلمثانولي لنبتتي للمستخلص ا in vivoدراسة النشاطية المضادة لألكسدة .2.6
chamaecyparissus جمموعات حتتوي 4قسمت إىل حيث غ ، 30-20يرتاوح وزهنا بني اليت ، Swiss albinosران ئفاستعمال ذكور مت
:كمايلي يف صفاق البطناملستخلص املثانويل بيوما 21دة حقنها يوميا ملفئران، مت 9-7كل جمموعة من
.S. chamaecyparissusمن املستخلص املثانويل لنبتة mg/kg 100دمت هلا ق Scاجملموعة -
.P. harmalaمن املستخلص املثانويل لنبتة mg/kg 10قدمت هلا Phاجملموعة -
NaCl (0,9 %.)احمللول امللحي قدمت هلا Controleجمموعة -
.Vit cمن mg/kg 50قدمت هلا Vit cجمموعة -
للقيام باختبار من الدم µl 100يؤخذ بعد هناية مدة املعاجلة،ت الفئران قتلعني الفئران مث يد من ور دم ال نزعمت
أما كمية الدم املتبقية (. NaCl 125mM ،10mM NaPOH4 ،pH7,4) من ml 2,4 حتلل الدم ويضاف له
إىل حني °م 20-يف ، السرتجاع البالزما ويتم حفضها(°م 4دقائق يف 5مدة g 1500) طرد مركزي فيجرى هلا
Feوقدرهتا اإلرجاعية أليون DPPHهبدف قياس القدرة اإلزاحية للبالزما جلذر استعماهلا بعد .+3
ورق بواسطة جيفف بعدها ، (% ,NaCl (0,9 ويغسل بواسطة احمللول الفيزيولوجيالكبد خذؤ ي رانئالف تشريح

يتم طحن احمللول و من الوزن الكلي للكبد، g 0,5 لـ (C° KCl 1,15%, 4 ( من ml 4,5 يضاف. الرتشيح
دقيقة مدة /دورة 4000، يعرض الناتج لعملية طرد مركزي sonicator مث ميرر عرب جهاز Dounce millبواسطة
mn 10 لتقدير مؤشرات اإلجهاد التأكسدي ليسرتجع اجلزء الطايف وحيفظ من أجل استعمالهMDA ،GSH و
Catalase ربوتني الكليوكذلك ال.
in vivo لتحلل كريات الدم الحمراءالقدرة المضادة .2.6.1
مع بعض ( 2010) وآخرون Takebayashi طريقة حسب لتحلل كريات الدم احلمراءمت قياس النشاطية املضادة
من حملول الدم السابق ELISA :µl 80باختصار نضع على صفائح (. 2002وآخرون، Manna)التعديالت
، ويتم قياس tBH (µM 166)من حملول µl 136يضاف له ،phosphate Tampon (1/25)ب يف حملول املذا
ويعرب عن مقاومة. nm 620وذلك بتتبع تغري امتصاصية العينات عند °م 37إمكانية حتلل كريات الدم احلمراء يف
احلمراء الدم كريات من % 50 لتحلل روريـــــالض بالزمن اجلذري للهجوم احلمراء الدم كريات
(half-hemolysis Time; HT50).
DPPHللبالزما على جذر تقدير النشاطية المضادة لألكسدة الكلية .2.6.2
وآخرون Brand-Williamsحسب طريقة DPPHة جذر ـــــبار على قدرة البالزما على إزاحــــيعتمد هذا اإلخت
حيفظ و ،% DPPH 0,004من حملول µl 625من البالزما µl 25 لـ فيضا. مع بعض التعديالت (1995)
يتم .nm 517دقيقة بعدها نقوم بطرد مركزي وتقرأ اإلمتصاصية للجزء الطايف عند 30املزيج يف الظالم مدة
:حساب النشاطية املضادة لألكسدة للبالزما بالعالقة التالية
% scavenging activity = [(Abs C –Abs ech) / Abs c] × 100
Abs C: الشاهد امتصاصية
Abs ech : (البالزما)امتصاصية العينة

ختبار القدرة اإلرجاعية للبالزما إ.2.6.3
Feالنشاطية املضادة لألكسدة للبالزما بقياس قدرهتا على إرجاع تقديرمت Feإىل +3
وآخرون، Chung)حسب +2
من ml 0,1و NaHPO4 (0,2M / pH6,6)املنظم لول احمل من ml 0,1 مع من البالزما ml 0,1 منزج (.2005
له ، يضافدقيقة 20مدة °م 50 يفحيضن املزيج . potassium fericyanide (K3Fe(CN)6) (1 %)حملول
ميرر املزيج على جهاز الطرد املركزيtrichloroacetic acid (:TCA 1 % .)من ml 0,25بعدها
ماء مقطر و ml 0,25من اجلزء الطايف وميزج مع ml 0,25مث يسرتجع مقدار ،(دقائق 10ة مدة دقيق/دورة (3000
0,5 ml FeCl3) ) مث تقرأ اإلمتصاصية عندnm 700 ، زيادة اإلمتصاصية تعكس زيادة القدرة اإلرجاعية.
في نسيج الكبد تقدير المؤشرات التأكسدية غير اإلنزيمية. 2.6.4
MDAتقدير كمية .2.6.4.1
الناجتة، وذلك حسب طريقة MDAميكن قياس فوق أكسدة الليبيدات املتشكلة على مستوى الكبد بتقدير كمية
Ohkawa) ،يعترب (. 1979وآخرونMDA يف وجود ركبات النهائية املتشكلة من فوق أكسدة الليبيداتأحد امل
لتشكيل معقد وردي اللون TBAمع جزيئيت MDAجزيئة يعتمد هذا اإلختبار على اإلرتباط بني . اجلذور احلرة
.nm 530ميتص عند
% TCA) (20 من حملول ml 0,25يضاف له (homogenat)من العينة املراد اختبارها ml 0,25 باختصار نأخذ
جدقيقة، يتم تربيد املزي 15مدة °م 100 خيلط املزيج ويسخن يف درجة. TBA) % (0,67من حملول ml 0,5 و
يتم (دقيقة15 دقيقة، مدة /دورة 3000)بعد القيام بعملية طرد مركزي n-butanolمن ml 2مث يضاف له
nmol/gramme ـيف العينة ب MDAيعرب عن كمية . nm 532اسرتجاع اجلزء الطايف وتقرأ كثافته الضوئية عند
tissus نسيج الكبد من.

الكبدي GSHتقدير كمية . 2.6.4.2
وذلك dithio-bis- [2- nitrobenzoic acid ] (DTNB -'5,5)قمنا باستخدام كاشف أملان GSHية لقياس كم
( SH)بواسطة جمموعات الثيول DTNBيعتمد أساس التفاعل على إرجاع Ellman (1959.)حسب طريقة
:كما هو موضح يف التفاعل التايل nitro-5-mercaptobenzoic acid-2للجليتاتيون فيتشكل محض
من حملول ml 0,5 يضاف لـ .nm 412عند ميتصبلون شديد الصفرة Nitromercaptobenzoicيتميز محض
10 % TCA مقدار ml 0,5 من العينة املراد اختبارها(homogenat ) ونقوم برجها رجا خفيفا على فرتات
من ml 0,2 نأخذ. (دقائق 5/ورةد 2000)ىل عملية طرد مركزي د بعدها يعرض املزيج إ 15 - 10متقطعة ملدة
من µl 100مع إضافة phosphate buffer (pH8) لول املنظماحمل من ml 1,7 اجلزء الطايف ونضيف إليها
ـيف العينة ب GSHيعرب عن تراكيز .nm 412دقائق عند طول موجة 5كاشف أملان، مث تقرأ اإلمتصاصية بعد
µmol/g tissus ا من منحىن العيارية ملركب ويتم احلصول عليها إنطالقGSH يف نفس الشروط السابقة.
على مستوى نسيج الكبد اإلنزيمية المؤشراتتقدير . 5 .2.6
Catalaseتقدير نشاط .5.1 .2.6
وهذا يعود إىل حتلل nm 240على اخنفاض اإلمتصاص الضوئي عند طول موجة CATيعتمد تقدير نشاط
H2O2 بفعل CAT (Clairborne ،1985).

احملضر يف H2O2 (19 mM )ومتزج مع حملول (homogenat) ارهاختباملراد امن العينة µl 50نأخذ ،ختصاربا
، يعرب عن نشاط ( دقيقتني)ثانية 120ملدة نقيس تغري اإلمتصاصية . M) NaHPO4 pH 7.4 (0.1 حملول نظامي
CAT حسب العالقة التالية:
K= 2.303 / T * log A1 / A2 :حيث أن
K :سرعة التفاعل
T :الفرق الزمين بالدقائق
A1 : اإلمتصاصية يف الزمنt0
A2 : اإلمتصاصية يف الزمنt1
أو U/mg protein دة ــــوحـــبال طىـــــــتع و K/n ب CAT ل ية ـــــالنوع ية ــــــاإلنزمي يةـــــالنشاط بـــــحتس
μmol H2O2 / mg protein / min .املتواجدة يف احلجم املستعمل mg ـثل كمية الربوتني بمي nحيث
تقدير البروتين الكلي. 5.2 .2.6
تكون شدة اللون املتشكل متناسبة . تعطي الربوتينات معقدا بنفسجيا مزرقا مع أمالح النحاس يف وسط قاعدي
وآخرون، (nm 540 Biuret ىمع تركيز الربوتني الكلي يف العينة والذي ميكن قياسه بواسطة املطياف الضوئي عل
، Kit total protein Biuret (ml Ref :1001290, 2×250 ) قمنا بقياس الربوتني الكلي باستعمال. (1999
عندها . °م 37يف محام مائي دقائق 5من العينة، مث ترتك ملدة µl 25من كواشف بيوريت مع 1mlوذلك مبزج
.mg/dl ب، تقدر كمية الربوتني يف العينة nm 540تقاس الكثافة الضوئية عند طول موجة

حصائية المستعملةالتحاليل اإل
االحنراف ±احلسايب باملتوسط، أو mean ± (SD)االحنراف املعياري ± احلسايب مت التعبري عن القيم باملتوسط
.mean ±(SEM) املعياري املتوسط
ANOVA عن طريق إختبار « logiciel « Graphpad Prism باستعمالحللت النتائج إحصائيا
(one-way ANOVA) متبوع باختبارTukey بالنسبة للمقارنات بني خمتلف املستخلصات والشواهد و بني
التحديد معامل أن أي % 5 الداللة عند معىن ذو إحصائيا الفرق اعترب حيث ،املستخلصات فيما بينها
(P < 0.05).

ئج والمناقشةالنتا
ص عديدات الفينول والفالفونيداتإستخال. 1
، واليت تعتمد Markham (1982)مت حتضري خمتلف املستخلصات املكونة لنبتيت احلرمل واجلعدة حسب طريقة
تعتمد طريقة الفصل على ذوبانية املركبات الفينولية يف هذه .القطبية متزايدة احملاليلعلى استعمال سلسلة من
( 85 %)ماء /يعرض مسحوق النبتتني اجملفف إىل اإلستخالص بواسطة خليط من حملول ميثانولحيث يل، احملال
هذا األخري يتم تعريضه لسلسلة من اإلستخالصات مبحاليل متزايدة . للحصول ابتداء على املستخلص اخلام
: مستخلصات مخسةول على يسمح هذا اإلستخالص باحلص(. اهلكسان، الكلوروفورم مث إثيل األسيتات)القطبية
، مستخلص إثيل (CHE)ستخلص الكلوروفورم م، ((HxE، مستخلص اهلكسان (CrE)املستخلص اخلام
(. AqE)و املستخلص املائي ( EAE)األسيتات
النبتتني، وقد لكال غ من الوزن اجلاف100مت تقدير مردودية مستخلصات نبتيت احلرمل واجلعدة إنطالقا من
بالنسبة لنبات اجلعدة CrE، يف حني قدرت أعلى مردودية يف EAEمردودية يف نبات احلرمل مع ظهرت أعلى
(.3، 2جدول )
شروط و املستعمل يف اإلستخالص و اجلزء يتإن املردود الناتج من عملية اإلستخالص خيتلف حسب النوع النبا
.عمل يف اإلستخالص املذيب املست ةوقطبي نوعو حمتوى كل نوع من املستقلبات و التجفيف
الفالفونيدات والدباغ من أهم املركبات املسؤولة على النشاطية و تعترب املركبات الفينولية كاألمحاض الفينولية
لوجية، كما متلك هذه املركبات العديد من النشاطيات البيو (. 2007وآخرون، Li)املضادة لألكسدة يف النباتات
مضادة للتخثر الدموي و مضادة للحساسية و مضادة للفريوسات و كترييامضادة للب و مضادة لإللتهاب منها
(.2010وآخرون، Gulcin) كل هذه النشاطيات تعود أساسا إىل النشاطية املضادة لألكسدة. وموسعة لألوعية
.اجلعدةنبات احلرمل واجلزء اهلوائي لنبتة بذور لكل من و الفالفونيدات ول الكليةتقدير عديدات الفينهلذا مت

تقدير عديدات الفينول الكلية والفالفونيدات. 2
نبات بذور ل( CrE ،CHE ،EAE) ةمت تقدير عديدات الفينول الكلية والفالفونيدات للمستخلصات الثالث
AlCl3 (Bahorumوطريقة ( 2007وآخرون، Li) Folin-ciocalteauاحلرمل واجلزء اهلوائي لنبتة اجلعدة بطريقة
.على الرتتيب (1996، وآخرون
مل يتضح أن حمتوى عديدات الفينول ملستخلصات نبات احلر ،(3،4)النتائج املسجلة يف اجلدول إنطالقا من
حيث (. املستخلص من g/محض الغاليك املكافئ mg( )183.95 ± 1.66 ) و (137.98 ± 6.9 ) بنييرتاوح
كمية عديدات الفينول يف مستخلصات نبتة ، يف حني تراوحتCrE-Phظهرت أعلى كمية لعديدات الفينول يف
وقد ظهرت . املستخلص من g/ ئمحض الغاليك املكاف mg 373.83 ± 0.23و 143.08 ±0.47 اجلعدة بني
. CHEمث CrEيليه EAEأعلى كمية لعديدات الفينول يف نبات اجلعدة على مستوى
، أن (1996) وآخرون Bahorumحسب AlCl3أظهرت النتائج حول تقدير الفالفونيدات باستعمال تقنية
quercetinمغ 61.51 ± 7.86و 7.39 ±0.68 تني يرتاوح بني االنب ىحمتوى الفالفونيدات يف مستخلصات كل
فقد ظهرت أعلى كمية من .نبات احلرمل واجلزء اهلوائي لنبتة اجلعدة على التوايلبذور غ من ملستخلصات /مكافئ
يليه EAE-Scوظهرت كمية عالية من الفالفونيدات يف CrE-Ph مثEAE-Ph يه ، يلCHE-Phالفالفونيدات يف
CrE-Sc مثCHE-Sc.

وحمتواها من عديدات الفينول و P. harmalaنبات احلرمل بذور مردود مستخلصات .2 الجدول
.الفالفونويدات
وحمتواها من عديدات الفينول S .chamaecyparissusمردود مستخلصات اجلزء اهلوائي لنبتة اجلعدة .3 الجدول
و الفالفونويدات
الفالفونويدات rutinمكافئ مغ
لكل غ املستخلص
الفالفونويدات مكافئ لكل quercetinمغ
غ من املستخلص
عديدات الفينول الكلية مكافئ مغ محض الغاليك
لكل غ من املستخلص %املردود
نوع املستخلص
اخلام 0.20 ± 9.71 0.35 ± 196.08 1.96 ± 16.94 1.23 ± 31.93 الكلوروفورم 0.22 ± 1.487 0.47 ± 143.08 0.98 ± 8.71 0.64 ± 6.46 1 إثيل األسيتات 0.34 ± 1.352 0.23 ± 373.83 7.86 ± 61.51 4.5 ± 115.12
يتضح من خالل النتائج املتحصل عليها أن مستخلصات نبات اجلعدة غنية باملركبات الفينولية والفالفونيدات
Asteraceaفقد أظهرت دراسات سابقة أن كمية عديدات الفينول يف عائلة . مقارنة مبستخلصات نبات احلرمل
.(Kapoor ،2002و (Kaurمع باقي العائالت تكون أعلى مقارنة
الفالفونويدات rutinمكافئ مغ
لكل غ املستخلص
الفالفونويدات مكافئ لكل quercetinمغ
غ من املستخلص
عديدات الفينول الكلية مكافئ مغ محض الغاليك
لكل غ من املستخلص نوع املستخلص %املردود
اخلام 0.27 ± 1.65 1.66 183.95± 0.68 ± 7.39 2.01 ± 21.38 الكلوروفورم 0.16 ± 2.19 6.97 ± 137.98 0.13 ± 13.88 0.39 ± 40.08 اثيل األسيتات 0.31 ± 2.41 1.66 153.12± 0.08 ± 12.1 0.24 ± 35.09

لمستخلصات نبات الحرمل والجعدة in vitro النشاطية المضادة لألكسدة. 3
ثير المزيح للجذور الحرةالتأ. 3.1
حيث تعمل كعوامل مرجعة ةاإلرجاعي-تعود النشاطية املضادة لألكسدة للمركبات الفينولية إىل نشاطيتها األكسدة
O2 مزحية لهيدروجني أو للماحنة أو (.1995وآخرون، Rice Evans)، كما هلا القدرة على استخالب املعادن 1
وتتمثل ،ملختلف املستخلصات باستعمال العديد من اإلختبارات in vitroمت تقدير النشاطية املضادة لألكسدة
وإزاحة XORو تثبيط استخالب املعادنو FTC و β –carotene و تبييض ABTS و DPPHازاحة اختبار يف
O2اختباري باستعمالعلى انتقال اإللكرتونات اليت تعتمددراسة القدرة املضادة لألكسدة حيث متت . -•
DPPH) وABTS ) وانتقال اهليدروجني عرب اختباريB-carotene) و(FTC (Li ،2009وآخرون ) أما اختبار
Ferrozine يونات احلديد الثائي، باإلضافة إىل اختبار تثبيط أل ستخالب املستخلصاتإفاستعمل لتقدير خاصية
O2وإزاحة جذر XORإنزم .الناتج عنه -•
DPPH إختبار إزاحة جذر. 3.1.1
البنفسجي إىل األصفر، اللون ذوعرب إرجاع هذا اجلذر DPPHجذر على إزاحةملستخلصات اقدرت نشاطية
على ملستخلصات املدروسة قدرة اائج احملصل عليها أن يتضح من خالل النت .517nmاإلمتصاصية عند قيست و
.(16 شكل)تناسب طرديا مع الزيادة يف الرتكيز ت DPPH ذرج ةزاحإ

Sc
0.0 0.1 0.2 0.3 0.4 0.50
20
40
60
80
100CrE
CHE
EAE
Concentration (mg/ml)
Inh
ibitio
n (%
)
Ph
0.0 0.5 1.0 1.5 2.00
20
40
60
80
100CrECHEEAE
Concentration (mg/ml)
Inh
ibitio
n (%
)
على جذر S .chamaecyparissus (Sc)و P.harmala (Ph)التأثري اإلزاحي ملستخلصات . 16الشكل DPPH.
DPPH (IC50من جذر 50 %تلف املستخلصات والشواهد املستعملة اليت تزيح خمتراكيز 17الشكل يوضح
mg/ml .) أظهرت النتائج أن لـEAE-Sc أعلى تأثري إزاحي على جذرDPPH ((0.01± 0.0001 mg/ml يفوق
تأثريا أقل EAE-Scو CHE-Phيف حني أبدت املستخلصات. مرة 2.8ب BHTالتأثري اإلزاحي لـ
، أبدت gallic acidو quercetin ،rutinأما الشواهد ( .، على التوايل 0.711± 0.029و 0.744 0.019± )
.> gallic acid) quercetin < (rutinاملوايل على الرتتيب BHTتأثريا إزاحيا أعلى من
CrECHE
EAE
QuercetinRutin
Gallic acid
BHT
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Ph
Sc
ns
nsns nsns
IC50
(m
g/m
l)
من جذر %51املزحية لـ S. chamaecyparissusو P .harmala تركيز مستخلصات كل من .17 الشكلDPPH .. (***: p ≤ 0.001, **: p ≤ 0.01, ns: p > 0.05 )

يتضح من خالل النتائج املتحصل عليها وجود عالقة طردية بني حمتوى املركبات الفينولية ملستخلصات نبات
إليه العديد من الباحثني، ما يتوافق مع ما توصل DPPHاحلرمل واجلعدة والتأثري اإلزاحي هلا على جذر
Agbor) ،؛ 2007 وآخرونOzsoy ،؛2008وآخرون Hossain وShah ،2011.)
Tsimogiannis)ببنيتها الكيميائية DPPHاملضادة لألكسدة وجذر آلية التفاعل بني املركبات تتعلق
على إزاحة جذور باإلضافة إىل أن قدرة هذه املركبات(. 2007وآخرون، Kouri؛ Oreopoulou ،2006و
DPPH ترتبط بعدد اجملاميع اهليدروكسيلية اليت حتتوي عليها(Boudet ،1997وآخرون .) ،بني فعلى سبيل املثال
Williams 4, 3وجود اجملموعة أن (4112( وآخرون-dihydroxylated يف احللقةB مع 3-4 والرابطة املزدوجة بني
يرفع من النشاطية املزحية C5و C3إضافية على كربون OH جماميعو وجود Cيف احللقة oxo-4وجود الوظيفة
. fisetinو myricetinو quercetinمثل للجذور احلرة
ABTSإختبار . 3.1.2
فعاال اجتاه معظم املركبات املضادة لألكسدة، كما أنه يذوب يف كل من املذيبات العضوية ABTSيعترب جذر
قدير نشاطية كل من مضادات األكسدة اليت تذوب يف املاء والدهون يف أطوار واملائية، لذلك ميكن استعماله لت
(.2010و آخرون، çelik) خمتلفة
(.18شكل )تزداد بزيادة الرتكيز ABTSبينت النتائج أن ملستخلصات النبتتني قدرة إزاحة معتربة جلذر

Sc
0.00 0.02 0.04 0.06 0.080
20
40
60
80
100
CrE
CHE
EAE
Concentration (mg/ml)
Inh
ibitio
n (%
)
Ph
0.0 0.1 0.2 0.3 0.40
20
40
60
80
100CrE
CHE
EAE
Concentration (mg/ml)
Inh
ibitio
n (%
) .ABTSعلى جذر S.chamaecyparissus و P. harmala ملستخلصاتالتأثري اإلزاحي .18الشكل
جذر على ميلك أعلى تأثري إزاحي EAE-Scملستخلصات نبات اجلعدة أن IC50(mg/ml ) بينت مقارنة قيم
ABTS (105-
mg/ml.9.40 ± 0.0085 )(19شكل )، وهو أقل من تأثريTrolox هذه توافقت .اتمر 6 حوايلبـ
أهنا ا، كمEAE-Scظهر أعلى تأثري إزاحي على مستوى أين ،DPPHاإلزاحي جلذر مع نتائج التأثري النتائج
الغنية لنباتات خمتلفة ستخلص إثيل األسيتاتتتوافق مع ما توصل إليه العديد من الباحثني حول القدرة العالية مل
وآخرون، ABTS (Fanأو DPPH ةإزاح على إزاحة اجلذور احلرة سواء عرب اختباربالفالفونيدات السكرية
(.2013وآخرون، Coqueiro؛ 2011وآخرون، Sudha؛ 2010
CrE
CHE
EAE
Trolo
x
0.00
0.05
0.10
0.15
0.20Ph
Sc
IC5
0 (
mg
/ml)
من جذر %05املزحية لـ S .chamaecyparissusو P. harmala تركيز مستخلصات كل من .19 الشكل
ABTS . (***: p ≤ 0.001, ns: p > 0.05 )

قد يعود الحتواءها على أكرب كمية من عديدات الفينول وباخلصوص EAE-Scكما أن التأثري اإلزاحي العايل لـ
إختالفا بني قيم التأثري اإلزاحي سواء باستعمال اختبار احلرمل يف حني أبدت مستخلصات نبات. الفالفونيدات
DPPH أوABTS،أعطى حيثCHE-Ph أعلى تأثري إزاحي جلذرABTS يليهCrE-Ph وبنسبة أقلEAE-
Phإىل نوع املركبات الفينولية املتواجدة يف يرجع هذاقد . يتناسب مع كمية عديدات الفينول املتواجدة هبا، وهذا ال
. مستخلصات نبات احلرمل أو لتدخل مركبات طبيعية نشطة أخرى
فينولية هذا القسم بنية حيمل . تصنع النباتات املركبات الفالفونيدية من أجل الوقاية من اإلصابة باجلذور احلرة
منح إلكرتونات مما يؤدي إىل استقرار ىمتلك العديد من الروابط املزدوجة واجملاميع اهليدروكسيلية اليت هلا القدرة عل
(.Pietta ،2000)اجلذور احلرة
تخفيض من خطر اإلصابة بأمراض الكسدي، و ىل احلماية من اإلجهاد التأإلفالفونيدات لالقدرة اإلزاحية تؤدي
(Hollman يةاالحناللمراض األي من السرطان، كما ختفض من ظهور الشيخوخة املسؤولة عن ظهور القلب، وتق
(.Harvsteen ،2002؛ Katan ،1997و
إختبار إستخالب المعادن . 3.2
Feأيونات تعترب(. 2010وآخرون، (Gulcinكسدة األكثر قوة، وهذا راجع لنشاطيتها العالية األمولدات من +2
فوق أكسدة الليبيدات ، مما يوسع Peroxyl و Alkoxylفكك اهليدروبريوكسيدات إىل جذور فهي حتفز ت
(Pincemail 1999، وآخرون .) كما بإمكاهنا أن تساهم يف تفاعالتFenton املنتجة جلذرOHلذلك . •
ذلك عرب استخالهبا بإمكاهنا أن تؤمن احلماية من األضرار التأكسدية و فاملركبات الفعالة املزحية أليونات احلديد
(.2010وآخرون، (Gulcinهلذه األيونات

أيونات إستخالب اختبارباستعمال على استخالب املعادنمستخلصات نبتيت احلرمل واجلعدة مت تقدير قدرة
FeFeمعقدا مع أيونات Ferrozineيشكل ، حيث+2
احلرة، يرافقه إخنفاضا يف إمتصاصية املعقد +2
–Ferrozine) (Fe . يعمل املركب املضاد لألكسدة على منع تشكل هذا املعقد. nm 562ند ع +2
تزداد قدرهتا على تثبيط تشكل ((EDTAالشاهد و نبتيت احلرمل واجلعدة مستخلصات أظهرت النتائج أن مجيع
Ferrozine) (Fe–املركب (. 20شكل)كيز مع الزيادة يف الرت +2
Sc
0 1 2 3 4 50
20
40
60
80
100CrE
CHE
EAE
Concentration (mg/ml)
Inh
ibitio
n (%
)
Ph
0 1 2 3 4 50
20
40
60
80
100CrE
CHE
EAE
Concentration (mg/ml)
Inh
ibitio
n (%
)
Feسبة تثبيط مركب ن . 20الشكل +2
-Ferrozine بواسطة مستخلصات P. harmalaو S.
chamaecyparissus.
هذا ما يعكس قدرته اإلستخالبية العالية، هذه ( 5.8 ± (0.0831بـ EDTAملركب IC50 (µg/ml )قيمة قدرت
.µg/ml 5.6 واليت كانت مساوية ل( 2007) وآخرون Leاليت توصل إليها IC50القيمة قريبة جدا لقيمة
اأعطى أعلى تأثري CrE-PHأن تبني ملستخلصات نبات احلرمل واجلعدة IC50 (mg/ml)قيم من خالل
مرة، 15 حوايلـب EDTAمقارنة مع باقي املستخلصات وبقيمة أقل من مركب (0.088 ±0.0008 ) ااستخالبي
،مرة على التوايل 209.31و 56 ـب EDTA، واليت أعطت قيم نشاطية أقل من مركب CHE-Scو CrE-Scيليه
، على الرتتيب( 2.631 ± 0.081و 2.256 ± 0.039)تأثريا متقاربا EAE-Scو EAE-Phيف حني أظهرت
(. 21شكل) CHE-Phأقل نشاطية يف وسجلت

CrE
CHE
EAE
EDTA
0
1
2
3
4
5
6 Ph
Sc
IC5
0 (
mg
/ml)
املعادن يف من %50على إستخالب S. chamaecyparissusو P.harmala تركيز مستخلصات .21 الشكل .p ≤ 0.001 :***) كشاهد EDTAوجود
احملتوى مع من خالل النتائج املتحصل عليها يظهر أن التأثري اإلستخاليب ملستخلصات نبات احلرمل يتناسب
يف حني . الفينويل للنبتة، لذلك قد يعود هذا التأثري اإلستخاليب إىل املركبات الفينولية املتواجدة هبذه املستخلصات
الفينولية يف هذه الظاهرةركبات نوع امليظهر هذا التناسب يف مستخلصات نبات اجلعدة، مما يعتقد تدخل مل
.طبيعية أخرى خبالف املركبات الفينولية يف استخالب أيونات احلديد الثنائيأو تدخل مركبات
-Amessis ،2009 وآخرون، Shyu)تعترب املركبات الفينولية املركبات املسؤولة عن استخالب األيونات املعدنية
Ouchemoukh ،من طرفيف حني أجريت دراسة سابقة .(2014، وآخرون Zhao عدم ظحو ل، (2008) وآخرون
.واحملتوى الفينويل، هذا ما تؤكده نتائج مستخلصات نبتة اجلعدة تناسب بني القدرة اإلستخالبية
لضرورة املركبات اإلستخالبية املتواجدة يف املستخلصات الفينولية ليست هي با املركباتهذه النتائج تشري إىل أن
، (phosphoric acid ،citric acid ،ascorbic acid)العديد من األمحاض العضوية مثل تتدخلالنباتية، وإمنا
carnosine مثل ، البيبتيدات والربوتيناتمينيةاألمحاض األبعض وtransferring و ovotransferrin يف
(.2011وآخرون، (Cornago ونات املعدنيةستخالب األيإ

(VanAckerتتطلب وجود عدد معني من اجملاميع الوظيفية املتوضعة بشكل حمدد فالقدرة اإلزاحية ملركب فينويل
إذن فالعينات الغنية باملركبات الفينولية ليس بإمكاهنا استخالب املعادن الناقلة إذا مل تكن (.1996وآخرون،
ترتبط بقوة مع احلديد على Catécholفمثال جماميع . اميع الوظيفية الضرورية للنشاطية اإلستخالبيةحتتوي على اجمل
(Wong من الفعالية اإلستخالبية نقصكذلك احتاد مركب فينويل مع جزء سكري ي. phénolعكس جماميع
النبتتني ميلك قدرة يظهر من خالل النتائج املتحصل عليها أن املستخلص اخلام لكال (. 2006وآخرون،
العالية للمركبات القطبية على استخالب املعادن القدرة استخالبية أعلى من باقي املستخلصات، هذا ما يظهر
، أن املستخلصات النباتية متلك (2010) وآخرون Sahreenوقد بينت دراسة . مقارنة باملركبات غري القطبية
عبارة mucilageفعلى سبيل املثال مركبات . املذيبات العضويةتأثريات استخالبية تتناسب طردا مع قطبيتها يف
، (Watanabeعن عديدات سكرية تذوب يف املاء وال تذوب يف الكحول تتميز خبصائص إستخالبية عالية
2008).
(.2011وآخرون، (Ozenستخالب املعادن إخاصية منكما أن اجملاميع القطبية اليت متلك نواة حمبة للماء تزيد
إختبار تثبيط فوق أكسدة الليبيدات. 3 .3
تسبب فوق أكسدة اللبيدات العديد من األمراض كأمراض القلب واألوعية والشيخوخة والسرطان
Ramarathnam) ،خاصة فوق أكسدة اللبيدات للذلك فمن الضروري البحث عن مثبطات (. 1995وآخرون
(.2009ون، وآخر Bougatef)املواد الغذائية منها املتواجدة يف
تتميز ظاهرة فوق أكسدة الليبيدت بتحول األمحاض الدهنية إىل ليبيدات بريوكسيدية يف وجود األيونات املعدنية
HNE وMDA (malonedialdéhyde )مثل تتجزأ إىل جمموعة من املركبات كاأللدهيدات و اهليدروكربوناتو
(4-hydroxynonénal)أكسدة الليبيدات عند املرضى فوق عملية، واليت تستعمل ككواشف ل(Pincemail

- β تبييضإىل إجراء اختبارين لفوق أكسدة الليبيدات، اختبار ةدراسهذه اللذلك ارتأينا يف ( .1999، وآخرون
caroténe واختبار فوق أكسدة محض اللينولييكFTC لفوق أكسدة الليبيداتقياس النشاطية التثبيطية ل.
β –caroténe تبييض إختبار. 3.3.1
عبارة عن آلية تعتمد على إزاحة اجلذور احلرة من جهة وتثبيط فوق أكسدة β -caroténeإن ابيضاض جزيئات
.الليبيدات الناجتة عن أكسدة محض اللينولييك من جهة أخرى
على أكسدة محض اللينولييك مما ينتج عنه تشكل جذور حرة ذات طبيعة β -caroténe تبييض يعتمد اختبار
كسدة على تعديل اجلذور احلرة األيعمل مضاد ،β –caroténeجزيئات ةهامجمتعمل على دروبريوكسيدية هي
حركية ، وقد مت تتبع β –caroteneاملشتقة من محض اللينولييك وبذلك حيمي من أكسدة وابييضاض مركب
كشاهد BHTواستعمال ساعة 24ستخلصات خالل امليف وجود وغياب β -caroténeإبيضاض مركبات
(. 22لالشك)
0 5 10 15 20 250
20
40
60
80
100CrE-Sc
CHE-Sc
EAE-Sc
CrE-Ph
CHE-Ph
EAE-Ph
MOH
H2O
BHT
Time (h)
Inh
ibit
ion
%
.S و P. harmalaملستخلصات β –caroténe / linoleic acidالنشاطية املضادة لألكسدة . 22الشكل
chamaecyparissus و الشواهدBHT) ساعة 24 خالل )و امليثانول و املاء.

. p < 0.001بصورة جد معتربة β -caroténeدة جزيئات النتائج أن مستخلصات نبات احلرمل تثبط أكستظهر
واليت كانت % 94.21 ± 0.003ب EAE-Phل ساعة 24فقد قدرت أعلى نشاطية مضادة لألكسدة عند
والذي ميلك بدوره نشاطية مضادة لألكسدة CrE-Phمتبوعة بـ % BHT 0.55 ± 99.66مقاربة لنشاطية مركب
35 .77 %± 0.032 أقل نشاطية بنسبة CHE-Ph، يف حني أظهر 86.24 % ±0.027 عالية واليت قدرت ب
(. 23شكل)
بعد )املاء امليثانول و و (BHT لمستخلصات والشواهدل β -caroténeالنشاطية املضادة لألكسدة . 23الشكل .(p ≤ 0.001, **: p ≤ 0.01, ns: p > 0.05 :***).محض اللينولييك/β-carotene يف إختبار ساعة 42مرور
CrE-Scفقد أظهر . ت مستخلصات نبتة اجلعدة نسبة تثبيطية منخفضة مقارنة مبستخلصات نبات احلرملهرأظ
.58 % حبوايل قريبة جدا من بعضها البعض تثبيطية نسب CHE-Scو EAE-Scو
- βض جزيئات يعتقد أن النشاطية املضادة لألكسدة للمركبات املتواجدة باملستخلصات تؤخر من معدل إبيضا
caroténe أو تثبيط أكسدة الليبيدات /عن طريق إزاحة اجلذور احلرة و(Jayaprakasha ،2001وآخرون.) تشري
العديد من الدراسات إىل أن النشاطية املضادة ألكسدة الليبيدات للمستخلصات النباتية تعود أساسا الحتوائها
إال أن نتائج هذه الدراسة مل (. Sarikurkcu ،2008 ؛2008 ،وآخرون Ozsoy)الفينول على مركبات عديدات
هلذا . تظهر التناسب الطردي بني النشاطية املضادة لألكسدة هلذه املستخلصات وحمتواها من عديدات الفينول
CrECHE
EAEM
OH
H2OBHT
0
20
40
60
80
100
Ph
Sc
***
***
***
***
***
***
***
***
Ant
ioxi
dant
act
ivity
%

ـميكن تفسري النشاطية العالية املضادة لألكسدة ل .فقد يعود هذا التأثري إىل نوعية املركبات الفينولية املتواجدة هبا
CrE وEAE إىل احتوائهما على املركبات األكثر قطبية على عكسCHE هذه النتائج تتوافق مع ما توصل اليه ،
Baghiani يف دراسة أجريت على جذور نبات ( 2010) وآخرون Carthamus caeruleus حيث أظهرت ،
% 77.13و 79.75 %قدرت ب نشاطية مضادة لألكسدة عالية واليت AqEو EAEاملستخلصات القطبية
Wuكما تتماشى هذه النتائج مع ما توصل إليه . على الرتتيب، رغم احتوائها الضئيل على املركبات الفينولية
Frankelإال أن (. CrE ،AqE)، حيث ظهرت أعلى نسبة تثبيطية مع املستخلصات القطبية (2010) وآخرون
دة غري القطبية متلك مميزات تثبيطية عالية جدا يف إقرتحوا فرضية تنص على أن مضادات األكس( 1994)، وآخرون
مبعىن أن املستخلصات القطبية اليت هلا . ماء مشكلة عازال حيمي الليبيدات من عملية األكسدة -املعلق زيت
خصائص ذوبانية يف الطور املائي متلك نشاطية أقل يف محاية اللبيدات من األكسدة مقارنة باملستخلصات األقل
وآخرون، Koleva) هلا القدرة على الذوبان يف الطور الدهين، وهذا ما يسمى بظاهرة القطبية العكسية قطبية اليت
(.2008وآخرون، Dastamalchi ؛ 2002
Ferric thiocyanate method (FTC)إختبار .3.2 .3
هر األكسدة، خصوصا محض يعترب الغشاء البالزمي غين باألمحاض الدهنية اليت تعترب املركبات األكثر عرضة لظوا
متثل ظاهرة (.Halliwell ،1994) ذان يعتربان هدفا رئيسيا لفوق أكسدة الليبيداتلاللينولييك واألراشيدونيك ال
أكسدة الليبدات أحد أسباب ظهور أمراض القلب واألوعية والسرطان، وتتدخل يف ظهور العديد من فوق
ومتنع الليبيدية، peroxylللهيدروجني على التفاعل مع جذور ملاحنةتعمل مضادات األكسدة ا .األضرار البيولوجية
لقياس كمية البريوكسيدات الناجتة يف بداية ظاهرة FTCتستعمل تقنية . سلسلة إنتاج جذور حرة جديدة استمرار
Feلتشكيل أيونات FeCl2مع peroxideفوق أكسدة الليبدات، حيث يتفاعل تتحد فيما بعد أيونات ،+3
.ذو الصبغة احلمراء Ammonium thiocyanateديد الثالثية مع احل

يعتقد أن املستخلصات اليت متلك نسب تثبيطية عالية، حتتوي على مركبات أكسدة أولية اليت بإمكاهنا التفاعل
OHاحلرة خصوصا جذر بقوة مع اجلذور ، وتعمل على إيقاف سلسلة تفاعالت اجلذور احلرة وبالتايل تأخري •
النشاطية املضادة لألكسدة بزيادة إمتصاصية املعقد تقاس. (2013 وآخرون، (Abbasi اهليدروبريوكسيداتتشكل
ferric thiocyacanate األكسدة الكلية حلمض اللينولييك سا، إىل غاية 24كل (Elmastas ،2006وآخرون).
قدرت . معلق محض اللينولييك متلك مستخلصات نبتيت احلرمل واجلعدة نشاطية مضادة لألكسدة عالية يف
على تثبيط فوق أكسدة اللبيدات ملعلق محض 2mg/mlنشاطية مستخلصات نبتيت احلرمل واجلعدة برتكيز
. 24اللينولييك والنتائج موضحة يف الشكل
0 24 48 72 96
0.0
0.2
0.4
0.6
0.8CrE-Ph
CHE-Ph
EAE-Ph
CrE-Sc
CHE-Sc
EAE-Sc
BHT
VITC
MOH
Time (h)
Ab
so
rba
nc
e
.Sو P. harmalaحركية فوق أكسدة محض اللينولييك يف وجود وغياب مستخلصات . 24شكلال
chamaecyparissus باستعمالBHT وVit C كشواهد.
VitC (0.007 .)وهي أقل من EAE-Ph (0.001)يف إمتصاصية كانت يف بداية التجربة أن أقلأظهرت النتائج
تعكس أقل قيمة لإلمتصاصية أعلى نشاطية مضادة لألكسدة؛ هذا ما يدل على القدرة التثبيطية العالية
. مث تزداد اإلمتصاصية مبرور الزمن، Scيف CHEكما ُسجلت أقل امتصاصية للمستخلص . EAE-Phملستخلص

إذ أن كل العينات (. t= 72h)مقارنة بآخرها ( t= 0h)فالنشاطية املضادة لألكسدة تكون عالية يف بداية التجربة
فقد أظهر(. t= 0h)ليوم األول مقارنة با( t= 72h)املستعملة تقريبا أظهرت إمتصاصية عالية يف اليوم األخري
CE-Sc يليها (0.00066)يف اإلمتصاصية إخنفاضا ملحوظا ،VitC (0.0863) مث ،BHT (0.0877 .) ومبقارنة
أقل إمتصاصية ) CrEعلى عكس مستخلص Phلكال النبتتني، فيالحظ أن أقل إمتصاصية يف CHEاملستخلص
فقد سجلت أعلى نشاطية . نشاطية عالية مضادة لألكسدة أظهرت النتائج أن مجيع املسخلصات متلك (.Scيف
BHT (% 87.87)و VitCتقريبا، واليت فاقت نشاطية كل من % 100بنسبة CrE-Scتثبيطية على مستوى
تعكس أعلى نسبة تثبيط لفوق أكسدة اللبيدات أعلى نشاطية مضادة لألكسدة . على التوايل( 87.68 %)و
ية أو ضاد لألكسدة ملستخلصات نبات احلرمل قد يعود الحتوائها على املركبات الفينولإن التأثري امل .26)شكل (
، واليت هلا القدرة على تسريع توقيف سلسلة تفاعالت فوق األكسدة اللبيدية مركبات مزحية للجذور احلرة
وآخرون، (Abbasiأخرىاللبيدات وجزيئات بيولوجية والتخلص من األنواع األكسجينية النشطة بتثبيط أكسدة
، يف حني مل يظهر التناسب الطردي بني احملتوى الفينويل والنشاطية املضادة لألكسدة ملستخلصات نبتة (2013
وآخرون، Djanna)اجلعدة، لذلك قد يعود هذا التأثري لتدخل مركبات غري فينولية أخرى مسؤولة عن هذا التأثري
2011 .)
Thiobarbituric acid (TBA) إختبار .3.3.3
لقياس كمية البريوكسيدات الناجتة يف املرحلة األولية من ظاهرة أكسدة الليبيدات، بينما FTC إختبار يستعمل
أثناء عملية األكسدة تتفكك البريوكسيدات تدرجييا . لقياس املركبات الثانوية الناجتة عن األكسدة TBAيستعمل
والذي يتم malonaldehyde (MDA )كبات جند مركب إىل مركبات ذات وزن جزيئي ضعيف، من بني هذه املر
.(2012 وآخرون،(TBA Emynur Shafekh قياسه بواسطة إختبار

S. chamaecyparissusو P. harmalaالنشاطيىة املضادة لألكسدة ملختلف مستخلصات 25يظهر الشكل
TBAباستعمال تقنية
Ebr
Ech
EAE
Vit C
BHT
MOH
0.00
0.04
0.08
0.12
0.16
0.20P. harmala
S. chamaecyparisus
Ab
so
rba
nc
e (
53
2n
m)
بواسطة S.chamaecyparissusو P.harmalaستخلصات املضادة لألكسدة يف وجود مالنشاطية .25 شكل .الناتج أثناء أكسدة الليبيدات MDAإمتصاصية قياسوذلك TBA تقنية
واليت كانت أقل ((EAE-Ph 0.026ظهرت بالنسبة لـ MDAيتضح من خالل النتائج أن أقل إمتصاصية ملركب
املتشكلة أثناء املرحلة األخرية من MDAيعكس إخنفاض كمية هذا ما .BHT )p> 0,05 (0.033متصاصية إمن
ىل اخنفاض تشكل مما أدى إلبيدات من عملية األكسدة قوية للحبماية EAE-Phأكسدة اللبيدات، وبالتايل قام
MDA . يف حني ظهرت أعلى امتصاصية على مستوىCHE-Sc مما يعكس قدرته الضعيفة على محاية الليبيدات
مبقارنة .BHT مث Vit Cيليه يف امليثانول أعلى امتصاصيةأما فيما خيص الشواهد، لوحظت .من عملية األكسدة
فنالحظ أهنا كانت متساوية فقط مع ( 26 الشكل) TBAو FTCباستعمال نسب تثبيط أكسدة اللبيدات
CHE-Sc (% 64 )رهتا على تثبيط أكسدة الشواهد يف قداقي املستخلصات و با بني يف حني يوجد اختالف
(.TBA)والنهائية ( (FTCالليبيدات يف مرحلتها األولية

CrE
-Ph
CHE-P
h
EAE- P
h
CrE
-Sc
CHE-S
c
EAE-S
c
Vit C
BHT
0
20
40
60
80
100
120FTC
MDAns
nsns
ns
ns
nsns
In
hib
itio
n %
S. chamaecyparissusو P .harmalaمقارنة بني النشاطية املضادة لألكسدة ملستخلصات نبتيت . 26شكل
.كشواهد BHTو VitCيف وجود TBAو FTCباستعمال تقنييت
التفاعل اليت يعتمد عليها كال ختالف آلية إىل عدة عوامل من بينها بني اإلختبارين إ ختالفاإلعزى قد ي
النشاطية املضادة لألكسدة اليت تقوم هبا هذه و ختبارين، بنية املركبات الفينولية املختلفة يف كل مستخلصاإل
فقد تعود النشاطية . (2003وآخرون، (Zainolاملركبات واحتمال وجود تعاون بينها يف القيام هبذه النشاطية
نواع األكسجينية النشطة كقدرهتا على إزاحة األ هذه املركبات إىل عدة آليات املضادة لألكسدة اليت متلكها
(quercetin وcatechine) ، فوق أكسدة الليبيدات سلسلةاجلذور احلرة و هدم بيط انتاجتثو
(p-coumaric acids)، ت املعدنيةاأليونا واستخالب Zainol) ،هذه ما يدل على أن اآللية .(2003وآخرون
.املعتمدة من طرف املركبات الفينولية للحماية من عملية األكسدة تتميز بنوعية عالية لكل مركب فينويل

XOR وتقدير نشاطيةتنقية ومراقبة نقاوة .4 .3
عرب سلسلة من عمليات الطرد (4113) آخرونو Baghiani طريقة من حليب البقر حسب XORمت تنقية
املركزي و الرتسيب بواسطة كربيتات األمونيوم إلستخالص اإلنزم اخلام، متبوع بالفصل على عمود كروماتوغرافيا
اليت (mg/ml 20.32)، هذه القيمة قريبة من القيمة mg/ml 43.40 قدرت مرودية اإلنزم ب .(هيبارين-أغاروز)
املنقى XORفوق البنفسجي إلنزم -أعطى املسح الطيفي املرئي (.4113)آخرون و Baghianiتوصل إليها
، وهي تدل على 5.15بـ (PFR)و قدرت نسبة الربوتني إىل الفالفني nm ،325 ،450 280ثالث معاول أساسية
ربائي على قمنا بإجراء فصل كه، XORوللتأكد من درجة نقاوة . (1996، وآخرون (Bray درجة عالية من النقاوة
هذا . كيلودالتون051، أظهر الفصل الكهربائي شريطا رئيسيا يف حدود (SDS-PAGE)هالم األكريالميد املتعدد
(.2004) وآخرون Atmani و (4113)و آخرون Baghianiما يتوافق مع نتائج
NAD ولكرتونات املنقى من حليب البقر يف وجود الكزانثني كمادة ماحنة لال XOR لقدرت النشاطية الكلية +
كمادة O2 يف وجودمغ بروتني و النشاطية املؤكسدة / دقيقة / نانومول 4155.15 ـب كمادة مستقبلة لالكرتونات
% 90وقد كانت نسبة النمط املؤكسد تفوق مغ بروتني، / دقيقة / نانومول 0988.55 ـب مستقبلة لالكرتونات
(.4113وآخرون، Baghiani)هذه النتائج تتماشى مع ما توصل إليه
XOدراسة التأثير التثبيطي على إنزيم .3.5
باستعمال ،XOعلى نشاطية إنزم S.chamaecyparissusو P. harmalaقمنا بدراسة تأثري مستخلصات
أظهرت .نانومرت مع الزمن 495و تتبع إنتاج محض اليوريك عند µM 011تركيز ثابت من مادة التفاعل الكزانثني
بصورة معتربة، بشكل يتناسب طرديا مع الزيادة يف تركيز XOئج أن كل املستخلصات تثبط إنزم النتا
.(27الشكل )املستخلصات

Sc
0.00 0.05 0.10 0.15 0.200
20
40
60
80
100CrE
CHE
EAE
Concentration (mg/ml)
Inh
ibitio
n (%
)
Ph
0.0 0.2 0.4 0.6 0.80
20
40
60
80
100CrE
CHE
EAE
Concentration (mg/ml)
Inh
ibitio
n (%
)
.XOعلى نشاطية إنزم S. chamaecyparissus و P harmalaالتأثري التثبيطي ملستخلصات . 27 شكل
مساوية IC 50 (mg/ml)بقيمة EAE-Scو CHE-Scمع XOم إلنز أعلى نشاطية تثبيطية سجلت
(0.055 ±0.005 ) بقيمة EAE-Phو CHE-Phيليها ،(0.052 ± (0.0003 و (0.051 ± (0.002
(. 28شكل)على التوايل (0.057 ±0.014 )و
CrE
CHE
EAE
Allo
purinol
0.0
0.1
0.2
0.3Ph
Sc
IC50
(m
g/m
l)
من نشاطية إنزم %05املثبطة لـ S. chamaecyparissus و P.harmalaتركيز مستخلصات . 28الشكل XOR (***: p ≤ 0.001).
مستخلصاتو ((EAE-Sc P > 0.05 و CHE-Sc مستخلصاتطة بواس XOمبقارنة نسبة تثبيط إنزم
CHE-Ph و EAE-Ph (P > 0.05) مع التأثري التثبيطي ملركبAllopurinol الذي يعترب مثبطا صناعيا نوعيا
.مرة على التوايل 6.7و 6.3 ـفقد كانت أقل ب XOإلنزم

XOR (Gawlik-Dzikiإنزم أجريت العديد من الدراسات حول قدرة املستخلصات النباتية على تثبيط
إذ يعتقد أن املركبات اليت تعمل على تثبيط إنزم (. 2009وآخرون، Ummamaheswari؛ 2012وآخرون،
XOR ركز النشط وترتبط هذه املركبات بدورها على مستوى املراكز تقوم مبنع تثبت مادة التفاعل باملMO
بني احملتوى الفينويل عدم وجود ارتباطيتضح من خالل النتائج .(1997وآخرون، Sanders ) لإلنزم FADو
يف حني أشار العديد من ،XORوقدرهتا التثبيطية إلنزم S. chamaecyparissus و P. harmalaلكال النبتتني
Lespade) الفالفونيداتخصوصا يعود إىل تدخل املركبات الفينولية XORالباحثني إىل أن تثبيط إنزم
(.2013 وآخرون، Bakasso ؛ 2012 وآخرون، Sowndhararajan ؛Bercion ،2010 و
ذ إ، ثبيطفإن نوعية الفالفونويدات املوجودة يف كل مستخلص ميكن أن حتدد قوة الت ،إضافة إىل العامل الكمي
تأثريا متلك CHEوالفالفونيدات غري السكرية املتواجدة يف EAEيعتقد أن الفالفونيدات السكرية املتواجدة يف
يف دراسة أجريت ( 2012؛ 2010وآخرون، (Baghianiهذه النتائج تتوافق مع ما توصل إليه . CrEأقوى من
أعلى تأثريا EAEو CHE فيها أظهر ،التوايلعلى Carthamus caeruleus و Capparis spinosaعلى نبات
.CrEمقارنة مع XORتثبيطيا إلنزم
، أن مركب (2002وآخرون، Van Hoorn ؛2002وآخرون، Lin)دراسات سابقة أجريت من طرف أظهرت
Flavone (Myrecetin ) ميلك تأثريا تثبيطيا عاليا إلنزمXOR بينما مل يظهر ،بشكل تنافسي
Catechine و Epicatechine أي تأثري تثبيطي (Cos ،؛ 1998وآخرونHsieh وChang ،2010) . كما أن
حيث أن وجود الرابطة املزدوجة بني، بنيتها الكميائيةيعود إىل قد XOR الفعل التثبيطي للفالفونويدات على
C2-C3 و غياب جمموعةOH يفC3 يرفع من القدرة التثبيطية للفالفونويدBorges) 2002 ون،وآخر).

O2جذرل زاحياإلالتأثير .3.6 XOالناتج عن -•
O2بإنتاج جذر XORنزم إيقوم O2يقوم جذر ،إرجاع األكسجني اجلزيئيوذلك عرب يف وجود الكزانثني -•
•-
Cyt-cبإرجاع Cyt-cإىل 3+
.S و P.hamala لدراسة التأثري اإلزاحي ملستخلصات نبتيت. 2+
chamaecyparissus جذر على O2O2بواسطة Cyt cقمنا بتتبع إرجاع XOالناتج عن-•
551عند طول موجة -•
Cyt cتثبط إرجاع S. chamaecyparissus و P .hamala أظهرت النتائج أن كل مستخلصات نبتيت .نانومرت
.(29 شكل)بشكل يتناسب طرديا مع الزيادة يف الرتكيز
Sc
0.00 0.05 0.10 0.15 0.200
20
40
60
80
100CrE
CHE
EAE
Concentration (mg/ml)
Inh
ibitio
n (%
)
Ph
0.0 0.2 0.4 0.6 0.80
20
40
60
80
100CrE
CHE
EAE
Concentration (mg/ml)
Inh
ibitio
n (%
)
O2 على جذر S. chamaecyparissus و P. harmala ملستخلصات زاحيلتأثري اإلا. 29 الشكل عن لناتجا -•
XO بإستعمالCyt c.
فلوحظ أعلى تأثري يف CrEميلك أعلى تأثري تثبيطي، أما بالنسبة ملستخلص CHE-Phأظهرت النتائج أن كما
Sc على عكس مستخلصEAE ظهر أعلى تأثري يفPh . النتائج يتضح عدم تناسب بني التأثري مبالحظة هذه
لكل من التأثري IC50مقارنة قيم مت. هلذه املستخلصات مع حمتواها من املركبات الفينولية Cytcاملثبط إلرجاع
O2و التأثري اإلزاحي جلذر XOاملثبط إلنزم . 30والنتائج موضحة يف الشكل ،-•

CrE
-Ph
CHE-P
h
EAE-P
h
CrE
-Sc
CHE-S
c
EAE-S
c
0.0
0.1
0.2
0.3XOR Inhibition
O2°- scavengerIC
50 (
mg
/ml)
O2و النشاطية املزحية جلذر XO من نشاطية %51ري التثبيطي لـ التأث. 30 الشكل•-.
مساويا للتأثري XORميلك تأثريا مثبطا إلنزم CrE-Scأن 30 يتضح من خالل النتائج املوضحة يف الشكل
O2اإلزاحي جلذر -CrEمبعىن أن ( Bالقسم )ضمن ( 1998) وآخرون Cosحسب CrE-Sc، هذا ما يصنف -•
Sc ثريا تثبيطيا إلنزم ميلك تأXO دون تأثري إضايف مزحيا لO2 Costantino، هذا ما يتوافق مع ما توصل إليه -•
O2للتأثري اإلزاحي جلذر IC50ميلكان EAE-Scو CHE-Scيف حني (.1992) وآخرون IC50أعلى من -•
O2ىن مستوى تشكل ، مبع(Dالقسم )على التوايل، هذا ما يصنفهما ضمن محض اليوريك إلنتاج املثبطةتكون -•
لتأثري اإلزاحي جلذر ا P. harmala IC50مستخلصات نبات متلك بينما . أعلى من مستوى تشكل محض اليوريك
O2O2وتأثريا إضافيا مزحيا جلذر XOمتلك تأثريا تثبيطيا إلنزم امبعىن أهن XOنزم إل ةطثبملا IC50أقل من -•
هذا -•
.(Cالقسم )ضمن ما جيعلها
.Sو P. harmalaيتضح عدم تناسب طردي بني كل من احملتوى الفينويل لنبتة ،ة النتائجظمن خالل مالح
chamaecyparissus والنشاطية املزحية جلذرO2لذلك يعتقد أن البنية الكيميائية للمركبات املتواجدة هبذه .-•
تأثريا epigallocatechineلك مركب مي ،املستخلصات هي اليت تتحكم يف هذه النشاطية، فعلى سبيل املثال
O2إزاحيا قويا جلذر O2أي تأثري إزاحي جلذر ال ميلك naringenineبينما مركب -•
•- Cos) ،1998آخرون.)

AAPHباستعمال in vitroالنشاطية المضادة لتحلل كريات الدم الحمراء .3.7
نولية على محاية كريات الدم احلمراء من عملية أجريت العديد من الدراسات مؤخرا حول تأثري املركبات الفي
حتفز جزيئات .(2011وآخرون، Valente؛ 2011وآخرون، Tulipani ؛ 2011وآخرون، Mendes)األكسدة
AAPH ( مركباتAZO )تتفاعل املركبات الناجتة بسرعة مع ،إنتاج اجلذور احلرة وذلك بعد جتزئتها يف وسط مائي
.مهامجة غشاء كريات الدم احلمراء لبريوكسيل واليت تعمل بدورها علىاألكسجني إلنتاج جذور ا
تعترب كريات الدم احلمراء من أكثر اخلاليا املتواجدة يف اجلسم، كما أهنا األكثر عرضة لألكسدة وذلك نظرا
عدنية إلحتواءها على كمية عالية من الليبيدات وغناها باألكسجني باإلضافة إىل احتواءها على األيونات امل
إىل حدوث فوق اإلجهاد التأكسدي يف العموم يؤدي(. 2012وآخرون، (Alvarez-Suarez كاحلديد والنحاس
ألغشية كريات الدم احلمراء مما يسبب إختالل وظائفها من خالل التأثري على ميوعتها وعلى األكسدة الليبيدية
د أن هذه اإلختالالت هي املسببة يف ظهور حتلل إذ يعتق ،وظائف املستقبالت واإلنزميات املدجمة يف األغشية
.كريات الدم احلمراء مما يؤدي إىل إصابة اخلاليا أو موهتا
باإلضافة إىل مستخلص الكلوروفورمي (MOHEو CrE) %100و %85قمنا بدراسة تأثري مستخلصي امليثانول
لى محاية كريات الدم احلمراء من ع S. chamaecyparissusو P. harmalaالنبتتني وإثيل األسيتات لكال
وتأثري AAPHحتلل كريات الدم احلمراء بواسطة 31 يظهر الشكل. AAPHالتحلل احملفز بواسطة املؤكسد
إىل معلق كريات الدم احلمراء، نالحظ زيادة يف نسبة AAPHعند إضافة . املستخلصات املدروسة على حتلل الدم
نالحظ أن عملية ،300mM AAPH ـبعد تعريض كريات الدم احلمراء ل .نحتلل كريات الدم احلمراء مبرور الزم
يالحظ من خالل النتائج أن كل من .ساعة ونصف 4دقيقة لتستقر بعد حوايل 60تبدأ بعد الدموي لالحناإل
قامت حبماية EAE-Scو EAE-Ph مستخلص إثيل األسيتاتو CrE-Phو MOHE-Phاملستخلص املثانويل
-CrE، إذ ظهرت أعلى نسبة محاية على مستوى P < 0,001بتأثري معنوي جدا من التحللاحلمراء كريات الدم

Ph وMOHE-Ph يف حني كل من املستخلص املثانويل .على التوايل %50.0و %56.65بMOHE-Sc و
CrE-Sc الكلوروفورمي املستخلصوCHE-Ph وCHE-Sc بتأثري مراءمتلك تأثريا حمفزا لتحلل كريات الدم احل
و %67.88 ـب CHE-Phو MOHE-Scإذ ظهرت أعلى نسبة حتلل يف كل من ، P < 0.001معنوي جدا
.(31شكل ) على التوايل 65.92%
Contr
oleCrE
CHE
EAE
MOHE
0
20
40
60
80
100
Ph
Sc
******
***
***
***
***
***
***
He
mo
lys
e %
S. chamaecyparissusو P. harmalaنسبة حتلل كريات الدم احلمراء يف وجود مستخلصات نبتيت .31شكل (0,1 mg/ml) 300بواسطة µM من حملولAAPH لإلحنالل الدموياحملفز (n=3. )
يف النشاطية املضادة اإختالففيها د وجفقد ،(2008) آخرونو de Freitasنتائج املتحصل عليها النتائج ؤكدت
ة متلك نشاطية حملل Solidago microglossaو Bryophyllum spذ أن كل من احلمراء إالدم كريات لتحلل
ويل الدمالحنإللقد يعود التأثري احملفز . ويل الدمالإلحنتأثريا مضادا لباقي النباتات متلك للدم يف حني
، إذ تعمل هذه املركبات على تكوين معقدات مع Saponinesعلى مركبات هاإلحتواءللمستخلصات
يف حني يعتقد أن تأخري ، الزميالستريوالت و الربوتينات والفوسفولبيدات مغرية بذلك من نفاذية الغشاء الب
لباقي املستخلصات قد يعود إىل وجود الفالفونيدات اليت هلا القدرة على اإلندماج ضمن أغشية وي الدم لالاإلحن

.(2007وآخرون، (Chaudhuri كريات الدم احلمراء وتعمل على محايتها من عملية األكسدة
يف وجوداخللوية اليت تصيب غشاء كريات الدم احلمراء حول السمية in vitroجريت العديد من الدراساتأ
عديدات الفينول فعلى سبيل املثال،. مستخلصات العديد من النباتات ويعزى ذلك إىل املركبات اليت حتتويها
املستخرجة من epicactechin isomers، (1997وآخرون، Grinberg)املستخرجة من الشاي األخضر واألسود
Jasmine green tea (Zhang ،1997وآخرون ) وsteryl glycosides من جذور نبات املستخرجة
Momordica cochinchinensis Ng) ،و (1986 وآخرونtriterpenoid saponins كل من املستخرجة من
Solidago viraurea ،Heteropappus altaicus، H. biennis ،Helianthus annuus وجد هلذه املركبات
.(1996وآخرون، (Baderدة بتحلل كريات الدم احلمراء عالقة وطي
إلختالف طبيعة أيضا يعود قد بني املستخلصات املدروسة حنالل الدمويإن االختالف يف النشاطية املضادة لإل
املركبات املتواجدة هبا، لذا من الضروري جتزئة هذه املستخلصات ملعرفة نوعية وآلية املركبات املسؤولة عن هذا
.أثريالت
في الكائن الحيدراسة النشاطية المضادة لألكسدة .4
( (APC تقدير النشاطية المضادة لألكسدة للبالزما .4.1
، albumine)،bilirubineحتتوي البالزما على جمموعة من املركبات املضادة لألكسدة داخلية املنشأ
reduced glutathion، (uric acid ميكن هلذه املركبات أن تعمل بشكل . من األغذيةوأخرى خارجية املنشأ تأيت
.العضوية تعاوين على سلسلة اجلذور احلرة من أجل القيام حبماية
للجذور احلرة وذلك على مستوى النظام املزيحمهية وهذا يعود لدورها األبالغة النشاطية املضادة للجذور احلرة إن

قدرة البالزما على إزاحة :متكاملني مهاستعمال اختبارين مسح ا. (Gulçin) ،2010الغذائي و األنظمة البيولوجية
Feوإرجاع احلديد الثنائي DPPHجذر .يف البالزما لمستخلصات النباتيةلألكسدة لاملضاد دور اللدراسة +2
DPPHجذر إزاحةقدرة البالزما على . 4.1.1
نشاطية تقاس حيث الفعل االزاحي للجذور احلرةمن اإلختبارات املستعملة بكثرة لتقدير DPPHيعترب اختبار
البنفسجي يف وجود البالزما مقارنة باحمللول املثانويل DPPHاملضادة لألكسدة بدرجة اختفاء لون حملول البالزما
.لوحده DPPHالذي حيتوي على
لص املثانويل للمستخ mg / kg 10و mg / kg 100جرعة إعطاء أن 33النتائج املوضحة يف الشكل تظهر أ
النشاطية املضادة لألكسدة للبالزما ب مل يؤثر علىعلى التوايل P .harmala و S. chamaecyparissusلنبتيت
.(32شكل ( )4.03 ± 32.11 %) الشاهدعلى التوايل مقارنة 4,45 ± 32.70 %و % 2.40 ± 59 .33
Controle VIT C MOHE-Ph MOHE-Sc
0
10
20
30
40nsns ns
Inh
ibit
ion
(%
)
الشاهدبمقارنة Phو Scملستخلصات DPPHكسدة للبالزما اجتاه جذر النشاطية املضادة لأل. 32الشكل
( (Scل mg/kg 100، يالحظ أن اجلرعة (HT50)والنشاطية الكلية للدم ( (APCمبقارنة النشاطية الكلية للبالزما
اجلرعة يف حني و النشاطية الكلية للدميف النشاطية الكلية للبالزما (p > 0.05) أظهرت تأثريا غري معنوي
10mg/kg لPh) ) أظهرت تأثريا جد معنويا(p < 0.001) يعود حيتمل أن ،فقط النشاطية الكلية للدميف

، ascorbate)اإلختالف بني النشاطية الكلية للبالزما والكلية للدم لتنوع واختالف األنظمة املضادة لألكسدة
urate ،tocopherole إن الدراسات اليت أجريت (.املوجودة يف الدمة مضادات األكسدو املوجودة يف البالزما
تقدير النشاطية املضادة لألكسدة الكلية للبالزما أو املصل قليلة وهذا يعود لصعوبة قياس كل مركب مضاد ل
يعتمد قياس النشاطية املضادة . (1999وآخرون، Re) املركبات دة على حدى والتفاعل احلاصل بنيلألكس
املستعمل يف القياس، ( املادة املؤكسدة)ريقة املستعملة ونوع املركب املنتج للجذور احلرة لألكسدة للعينة على الط
اختيار إختبارين خمتلفني مت على تسهيل الفهم وتفسري النتائج، هلذا قارنة بني التقنيات يساعد أكثر كما أن امل
(DPPH وreducing power). يدل إزاحة اجلذور احلرة يف حني األول على قدرة البالزما علىاالختبار يعتمد
مبستويات حمددة للمركبات املضادة لألكسدة يف نيتين التقنيتتعلق هذ. الثاين على القدرة اإلرجاعية للبالزما
القدرة اختبارمبستويات الغليتاتيون الداخلية يف حني يتعلق DPPHالبالزما فعلى سبيل املثال يتعلق اختبار
.البالزمية -tocopherol αو كاألسكوربيك ومحض اليوري مبستويات محضاإلرجاعية
رجاعية للبالزما ختبار القدرة اإلإ. 2 .4.1
له لقياس النشاطية املضادة غالت األكسدة اإلرجاعية ميكن استإلكرتونات يف تفاعال منحإن قدرة العينة على
Feملركب تحولياس القدرة اإلرجاعية للبالزما بمت ق. لألكسدة3+
(CN-Feشكل ل 6(
2+ (CN
-)6 (Le ،وآخرون
لبالزما اقدرة على شدة هذا اللون تعربيتغري اللون األصفر للوسط التفاعلي إىل اللون األخضر و (.2007
. 2004)وآخرون، Zou)اإلرجاعية
الشاهدبمقارنة رجاعية للبالزما بشكل معنوي رفع من القدرة اإل MOHE-Scأن 33يتضح من خالل الشكل
(p < 0.001) ارتفاع طفيف وغري معنوي يف كل من ، يليه0.298 ± 1.237 ببإمتصاصية قدرت MOHE-Ph
.(p > 0.05)على التوايل 0.279 ± 0.867 و0.915 ± 0.217 ب Vit C و

Controle VIT C MOHE-PH MOHE-GH0.0
0.5
1.0
1.5
2.0
***
**A
bs
orb
an
ce
(7
00
nm
)
.S. chamaecyparissusو P. harmalaللمستخلص املثانويل لكل من القدرة اإلرجاعية للبالزما. 33 شكل .(غري معنوي= ns ؛ (***mean ± SEM (n=8-9) p < 0.001 القيم متثل
،يوجد اختالف يف نتائج الباحثني حول استجابة مضادات األكسدة بعد استهالك عديدات الفينول ملدة قصرية
Pinnus maritimaمن مستخلص بعد استهالك كل النشاطية املضادة لألكسدةعدم تغري يف أبدتمن هافمن
(Silliman ،2003وآخرون ) واستهالكanthocyanins من نباتMyrciaria jaboticaba Leite) ،وآخرون
يف حني أجريت دراسة أخرى وجد فيها أن مستخلص التوت يرفع النشاطية املضادة لألكسدة للبالزما ،( 2011
(Hassimotto ،2011وآخرون). أجريت من طرف اسة سابقة ر يف د(da Silva ،2013وآخرون ) أظهرت أن
مل يظهر تغريا يف النشاطية املضادة لألكسدة للبالزما Passiflora edulisمستخلص اهليدروكحويل لنبات
اليت مل يظهر فيها هذا ( يوما 15)لقصر مدة التجربة أرجع السببوقد FRAPو ORACختبار إباستعمال
ال استهالك الشاي األسود مل يظهر تغريا يف النشاطية املضادة لألكسدة للبالزما باستعمال فعلى سبيل املث أثري،الت
على تدل النتائج احملصل عليها ، (Widlansky ،2005)إال بعد إطالة زمن التجربة FRAPو ORACاختبار
متثل الفالفونيدات املركبات إذ . قدرة هذه العينات على منح إلكرتونات، وبالتايل قدرهتا على تعديل اجلذور احلرة
كما أظهرت دراسات أخرى أن املركبات الفينولية هي ،(2007وآخرون، Le)لإللكرتونات احنةاألساسية امل
إال أن التناسب بني . (2011وآخرون، (Krishnamoorthy املسؤولة عن القدرة اإلرجاعية للمستخلصات النباتية

آخرونو Ozenففي دراسة أجريت من طرف ،ال حيدث يف معظم احلاالتاحملتوى الفينويل والقدرة اإلرجاعية
ميلك أعلى قدرة إرجاعية مقارنة بباقي Thymus praecoxتة وجد أن املستخلص املثانويل لنب( 2011)
وقد يعزى هذا لنوعية املركبات املستخلصات املدروسة بالرغم من احتوائه على أقل كمية من املركبات الفينولية
.تواة يف املستخلصاحمل
نا ظبالقدرة اإلرجاعية، فيما خيص النتائج احملصل عليها الح رتبطت لجذور احلرة الاإلزاحية لحظ أن النشاطية و ل
Priorو Caoنتائج عنالنتائج هذه ختتلف ،reducing powerو DPPHبني قيم كل من إختبار توافقوجود
مل يالحظ وجود هذا حيث باستعمال طرق متعددة دة ملصل اإلنساندراسة النشاطية املضادة لألكسيف ( (1998
مطحون أنسجة )نوعية العينة املستعملة وطريقة حتضريها النشاطية املضادة لألكسدة حسب ف تلخت .التناسب
يف املصل، urateو ascorbateتواجد )العينات يف هذهواختالف مضادات األكسدة ( مصل إنسان أوحيوان
(. كسدة اخللوية يف األنسجةومضادات األ
لتحلل كريات الدم الحمراء النشاطية المضادة .4.2
حتلل يف زمن اليت تدل على إرتفاع HT50نتائج قيم .كريات الدم احلمراءعلى حتفيز حتلل tBHPيعمل مركب
. 34مبينة يف الشكل MOHE-Scو MOHE-Phلكل من نصف كريات الدم احلمراء
HT50يف اإلحنالل الدموي بقيمة p < 0.001تأخرا جد معنوي Phحفز مستخلص
مقارنة p < 0. 05( min = HT50 4.95 ± 72.05)بقيمة Scمستخلص ، يليه min 11.32 ± 78.51مساوية ل
.(min 9.32 ± 59.88)مع جمموعة الشاهد

Controle Vit C MOHE-Ph MOHE-Sc0
20
40
60
80
100
ns
**
HT
50
(min
)
ns
و S. chamaecyparissusو P. harmalaاملثانويل لكل من بعد إضافة املستخلص HT50زمن ال .34شكل
Vit C القيم متثلmean ± SEM (n=8-9)، *** : p≤0.001) ؛ns =غري معنوي.)
على كريات الدم احلمراء يكون من خالل اإلرتباط مع الربوتينات الغشائية مما tBHPيعتقد أن آلية تأثري مركب
إذ ،(1992وآخرون، Deuticke)الغشائية الليبيدية وتكوين ثغور بالغشاء اخللوي يؤدي إىل تغيري يف هيئة الطبقة
كما أن اخنفاض ،(1996وآخرون، Haraguchi)للربوتينات أكثر عرضة لإلجهاد التأكسدي SH–تعترب اجملاميع
يف اختالل ؤديا إىلم(. 1994وآخرون، (SH Grinberg–يؤدي إىل اخنفاض يف جماميع يونثاجلليتامستويات
. (1996وآخرون، (Kitagawaاستقرار األغشية
على مستوى كريات الدم احلمراء MDAإنتاج مركب هفوق أكسدة الليبيدات من خالل حتفيز على tBHP يؤثر
ومن أجل محاية األغشية من هجوم .(1987وآخرون، Deuticke)مما يؤدي إىل اختالل النفاذية الغشائية
بح استعمال مضادات األكسدة من مصادر طبيعية حمل اهتمام كبرياجلذور احلرة، أص
Machlin) وBendich ،1987) .من أهم مضادات األكسدة املتواجدة على مستوى كريات الدم احلمراء جند

من أهم Vit Eإذ يعترب ،Vit Eوأنظمة غري انزميية مثل اجلليتاتيون و GPXو SOD ،CATنزميية مثل إأنظمة
وآخرون، Dalle-Donne؛ Gutteridge ،1992و (Halliwellغشية اخللوية دات األكسدة الدفاعية لألمضا
، فهو يعمل على استقرار in vitroجهاد التأكسدي إللد اضامل quercetinأظهرت بعض الدراسات دور (.2006
(.2010خرون، وآ Hapner)لسهولة نفاذه إىل داخل اخللية ويرجع هذاغشاء كريات الدم احلمراء
درجة ( 2) ،نوع املركبات الفينولية( 1: )يعود تأثري محاية املستخلصات النباتية من أكسدة كريات الدم احلمراء عرب
إال أن سلوك هذه املركبات ،(2007وآخرون، (Chaudhuriنفاذية الفالفونيدات بداخل كريات الدم احلمراء
كريات الدم احلمراء متثل حشوة خلوية معقدة، إذ تؤثر لكون نظرا اجتاه اجلذور احلرة يبقى عموما صعب التفسري
(.2007وآخرون، (Djeridaneمتنوعة بتفاعالت املركبات الصيدالنية على الغشاء البالزمي
يلكبدا MDA مستويات تركيز .3 .4
أكسدة فوق تثبيطيستخدم ،اخللوي املوت من مشرتكة وحالة ذاتية حتليلية عملية الليبيدات أكسدة فوق تعد
وآخرون، (MDA Mazunderلألكسدة والذي يظهر من خالل تقدير املضاد للنشاط مهم كمؤشر الليبيدات
2005 .)
MOHE-Scو MOHE-Phران املعاجلة مبستخلص ئالف لدىالكبدي MDAتغري تركيز نسب 35 يظهر الشكل
و MOHE-Phستخلصاملران املعاجلة بئالفعند MDAيف تركيز خنفاظ جد معنوياالحظ ي .مقارنة بالشاهد
MOHE-Sc وVit C (P < 0.001) بالنسبة % 63.48قدر هذا اإلخنفاض ب . الشاهدمقارنة معPh
(14.47 nmol /g tissus ± 41.77 ) بالنسبة % 79.56وبSc (23.37 ± 6.73 nmol/g tissus ) 84.58و %
. (nmol / g tissus 64.78 ±7.14) الشاهدبمقارنة Vit C (nmol / g tissus 17.63 ± 6.06 )بالنسبة ل

Controle VIT C MOHE-Ph MOHE-Sc0
20
40
60
80
TB
A (
n m
ol/ g
wet
tissu
e)
السيتوزويل MDAعلى معدل VitCو MOHE-Scو MOHE-Phتأثري معاملة الفئران باملستخلص .35شكل n=6-9) ). *** : p≤0.001 للكبد
،مؤشر فوق األكسدة الليبيدية مستوىعلى خفض S. chamaecyparissus و P. harmala يقدرة مستخلصب
وذلك لغىن املستخلصني بعديدات الفينول اليت TBARS تعمل على وقاية الكبد من خالل خفض مؤشر فإهنا
.تزيح اجلذور احلرة
اليت املميزات و باخلصائص تتمتع S. chamaecyparissusو P. harmala يتنبت نالحظ سبق ما خالل من
بشكل معنوي MDA معدالت اخنفاض خالل من جليا ذلك يبدو و لألكسدة املضادة فهاوظائ بأداء هلا تسمح
p≤0.001 أجريت العديد من الدراسات على .امليثانويل مسبقا باملستخلص املعاملتني للمجموعتني بالنسبة
كبد، ال)سواء يف الدم أو األعضاء MDAوالذي يظهر فيها اخنفاض مستويات TBARSتقدير ومت احليوانات
تؤكده ا ماهذ(. 2013وآخرون، da Silva، 2011وآخرون، Chen ؛2008 وآخرون، Decorde( )الكلى، املخ
عند % 63.48و % 79.56 بنسبةيف الكبد MDAيف مستويات اخنفاضإ النتائج احملصل عليها، إذ أثبتت
دراسة أجريت من طرف يف. على التوايل MOHE-Phو MOHE-Scمبستخلصي ران املعاجلةئالف اتعو جمام
Khennouf يفحول تأثري املركبات الفينولية و الدباغ النقية على تثبيط فوق أكسدة الليبيدات ( 2010) آخرون
األرانب، أظهرت النتائج أن األمحاض الفينولية والدباغ النقية تثبط فوق أكسدة الليبيدات يف املخ عند مجيع

لوحظ حيث( 2010 آخرون، (Rop ةدراس أيضا كدتؤ ، هذه النتائج (µg/mL 50 ,25 ,10)الرتاكيز املدروسة
blackب ةعاجلاملكبد اجلرذان يف % 19.81 و % 12.57 بني نسبةتثبيط أكسدة الليبيدات ب يف خنفاضا
chokeberries.
الكبدي CATتقدير نشاط .4 .4
على بشكل غري مباشر ا يعمل كم أكسجنيو ماءإىل H2O2هيموبروتني يعمل على إرجاع هو CATإنزم
.(Subramanian ،2005و (Sathishsekarمحاية األنسجة من جذور اهليدروكسيل عالية النشاط
على نشاط إنزم S. chamaecyparissusو P. harmalaنتائج تأثري املعاجلة مبستخلصي 36الشكل يبني
CAT ارتفاعا معنويا ظحو ل .على مستوى الكبدP < 0.01 اطية يف نشCAT ران املعاجلة ئسواءا عند الف
ه ذقدرت ه .ران الشاهدئمقارنة مع جمموعة ف S. chamaecyparissusو مستخلص أ P. harmalaستخلص مب
P % 35 و Sc (0.382 ± 0.068 U/mg protein)ران ئبالنسبة جملموعة الف ((P < 0.001 % 59.16الزيادة ب
بالشاهد مقارنة Ph (0.021 U / mg protein ± (0.24بالنسبة جملموعة الفئران( (0.01 >
(0.033 U / mg protein ± 0.156.)
Controle VIT C CrE-Ph CrE-Sc0.0
0.1
0.2
0.3
0.4
0.5
ns
Ca
tala
se
ac
tiv
ity
(U
/mg
pro
tein
)
CATعلى نشاط إنزم MOHE-Scو MOHE-Phتأثري املعاملة مبستخلص .36شكل
n=6-9) *** : p≤0.001, ** : p≤0.01))

من األضرار اليت تسببها يحيمالذي CATات على الرفع من نشاطية املركبات الفعالة املتواجدة هبذه النباتتعمل
املنتجة لإلنزم mRNAزيادة التعبري عن جزيئات ناجتا عن هذا اإلرتفاع يكون أن وميكنالكبد، يفاجلذور احلرة
Zhong) ،النشاطية املضادة لألكسدة لإلنزميات أجريت دراسات أخرى حول تقدير .(1999وآخرونCAT ،
SOD ،GSH-PX عند جرذان معاجلة مبستخلصات نبتةDouchi ،ففي الكبد على مستوى الكبد والكلى
نشاطية يف وغري معنوي طفيفوارتفاع ، (p < 0.05)مقارنة بالشاهد SODو CATنشاطية إنزميي ارتفعت
-SOD (p < 0.05) GSHظهرت زيادة معنوية يف نشاط فأما على مستوى الكلى . GSH-PX (p > 0.05)إنزم
PX يف نشاطية و زيادة غري معنويةCAT (p > 0.05) مقارنة بالشاهد Wang) ،يف حني .(2008وآخرون
فلم نبات املشمش بأشهر 5 مبعاجلة جرذان مدة (2009) وآخرون Ozturkأجريت دراسات سابقة من طرف
املتحصل إن النتائج .(CAT ،SOD ،GSH-PX)كسدة يالحظ أي تغيري يف نشاطية اإلنزميات املضادة لأل
مما يبني جليا MDAواخنفاض يف معدالت GSHوزيادة يف تراكيز CATنشاطية إنزم عليها أظهرت إرتفاع يف
ورفع النظام متلك قدرة فعالة على إلتقاط اجلذور احلرة S. chamaecyparissusو P. harmalaأن مستخلصات
.املضاد لألكسدة
يلكبدا GSHمعايرة . 4.5
.Sو P. harmalaمبستخلصي فئرانيف نسيج الكبد بعد معاجلة ال GSHالنتائج اليت تظهر تركيز
chamaecyparissus يف تركيز حظ ارتفاع معنويو ل .37 يوما موضحة يف الشكل 21ملدةGSH فئرانعند ال
MOHE-Scلصوتلك املعاجلة مبستخ( (MOHE-Ph (6.74 ± 1.89 µmol / g) P < 0.05املعاجلة مبستخلص
(8.19 ± 2.98 µmol/g )P < 0.01)) 3.34 1.12الشاهدبمقارنة على التوايل ±.
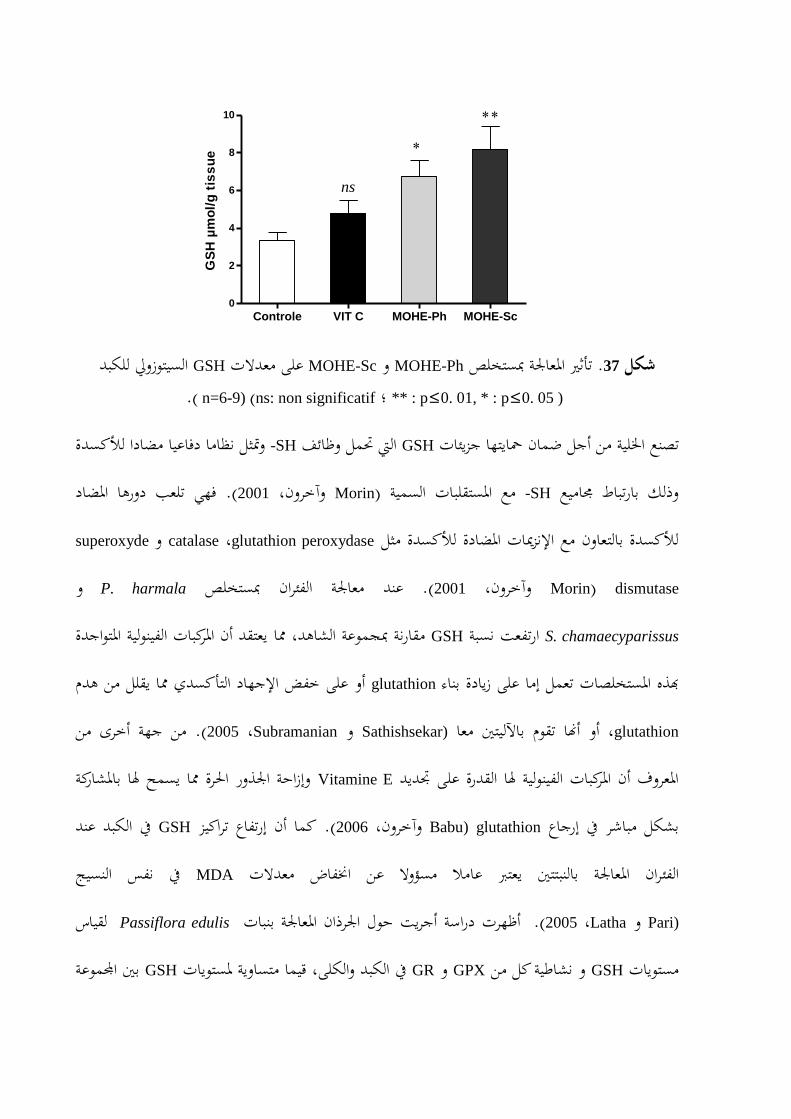
Controle VIT C MOHE-Ph MOHE-Sc0
2
4
6
8
10
ns
GS
H µ
mo
l/g
tis
su
e
السيتوزويل للكبد GSHعلى معدالت MOHE-Scو MOHE-Phتأثري املعاجلة مبستخلص . 37 شكل
** : p≤0. 01, * : p≤0. 05 ) ؛ns: non significatif) n=6-9) ).
مضادا لألكسدة دفاعيا ومتثل نظاما SH-اليت حتمل وظائف GSHاخللية من أجل ضمان محايتها جزيئات صنع ت
فهي تلعب دورها املضاد (. 2001وآخرون، Morin)مع املستقلبات السمية SH-وذلك بارتباط جماميع
superoxydeو glutathion peroxydase، catalase لألكسدة بالتعاون مع اإلنزميات املضادة لألكسدة مثل
dismutase (Morin ،2001وآخرون .)مبستخلص فئرانعند معاجلة ال P. harmalaو
S. chamaecyparissus نسبة تفعتر اGSH يعتقد أن املركبات الفينولية املتواجدة مما ،مقارنة مبجموعة الشاهد
ض اإلجهاد التأكسدي مما يقلل من هدم خف أو على glutathionستخلصات تعمل إما على زيادة بناءهبذه امل
glutathion أو أهنا تقوم باآلليتني معا ،Sathishsekar) وSubramanian ،2005) . من جهة أخرى من
وإزاحة اجلذور احلرة مما يسمح هلا باملشاركة Vitamine Eاملعروف أن املركبات الفينولية هلا القدرة على جتديد
يف الكبد عند GSHكما أن إرتفاع تراكيز (. 2006وآخرون، (glutathion Babuبشكل مباشر يف إرجاع
يف نفس النسيج MDAران املعاجلة بالنبتتني يعترب عامال مسؤوال عن اخنفاض معدالت ئالف
Pari) وLatha ،2005) . دراسة أجريت حول اجلرذان املعاجلة بنبات أظهرت Passiflora edulis لقياس
بني اجملموعة GSHقيما متساوية ملستويات ،الكبد والكلى يف GRو GPXطية كل من و نشا GSHمستويات

، باإلضافة إىل ارتفاع يف الكلىيف GSH (% 40)يات يف مستو وارتفاع الكبد، يف جمموعة الشاهد املعاجلة و
.(2013 آخرون، da Silva)يف الكبد والكلى GPXو GRنشاطية

قشة العامةالمنا
تظهر أ اليت ثل موضوع العديد من دراسات علم األوبئة متأصبحت مضادات األكسدة يف العقدين األخريين،
تحيث تركز ،خنفاض معدل اإلجهاد التأكسدي املسبب لألمراضوااستهالك مضادات األكسدة العالقة بني
، Ngو Liu)سني صحة اإلنسان عمال مضادات األكسدة خاصة الطبيعية منها لتحتاالهتمامات حول اس
Peganum harmalaقمنا يف هذه الدراسة باستعمال نبتتني طبيتني .(Wang ،2001و Zheng؛ 2000
كمصدر ملضادات األكسدة الطبيعية وذلك باختبار نشاطيتهما املضادة Santolina chamaecyparissusو
. (in vivo)فئران الوجتريبها على (in vitro) خمربيالألكسدة
يثانول املستخلص متقدير عديدات الفينول الكلية والفالفونيدات املتواجدة يف مت قبل التطرق إىل هذه اإلختبارات
أن كمية ةأظهرت الدراس. S. chamaecyparissusو P. harmalaلكل من ثيلاإلأسيتات والكلوروفورم و
373.83 ± 0.23 و 137.98 ± 6.97 رتاوح بنيتنبتتني واجدة يف مستخلصات كال العديدات الفينول الكلية املت
على P. harmala ظهرت أعلى كمية لعديدات الفينول لنباتو املستخلصغ من / محض غاليكمكافئ مغ
أما .S. chaaecyparisusبالنسبة ل EAE-Sc أعلى نسبة يف مستخلصكانت يف حني CrE-Phمستوى
غ من /كرستنيمكافئ مغ 61.51 ±7.86 إىل 7,39 ± 0.68بني تهاسبة للفالفونيدات فقد تراوحت كميبالن
كمية عالية من املركبات الفينولية والفالفونيدية على احتواءها S. chamaecyparissusأبدت نبتة .املستخلص
أن كمية عديدات بينت دراسات سابقة كديؤ هذا ما ،P. harmala يف مجيع املستخلصات مقارنة مع نبات
يتغري تركيز (.Kapoor ،2002و (Kaurتكون أعلى مقارنة مع باقي العائالت Asteraceaول يف عائلة الفين
.وئية واحلرارةعدة عوامل منها املوقع اجلغرايف، املرحلة الضتبعا لاملركبات احليوية يف النباتات وكذا تأثرياهتا البيولوجية
الثانوية كالفالفوندات هذا ما يغري نشاطيتها املضادة لألكسدة هذه العوامل على البناء احليوي للمستقلبات تؤثر
(Kumazawa ،؛ 2004وآخرونJaakola وHohtola ،2010).

مقارنة ABTSو DPPHجلذر ميلك أعلى تأثري إزاحي EAE-Scأن ABTSو DPPHإختباري أظهرت نتائج
إال أنه ، ن املركبات الفينولية والفالفونيداتهذا الحتواءه على كمية عالية مميكن إعزاء باقي املستخلصات و ب
Zhouهذا ما يتوافق مع ما توصل إليه ،اختالف بني نتائج اإلختبارين بالنسبة لباقي املستخلصاتيوجود
يعتقد أن الفالفونيدات السكرية متلك قدرة إزاحية عالية للجذور احلرة مقارنة مع باقي هذا و . ((2013 وآخرون
. املستخلصات
تتم وفق سلسلة من و . ل ظاهرة فوق أكسدة الليبدات أحد أسباب ظهور أمراض القلب واألوعية والسرطانمتث
أظهرت . (2013 وآخرون، (Abbasi من األضرار البيولوجيةعدد تفاعالت اجلذور احلرة، كما أهنا تتدخل يف
بينما ،EAE-Phبواسطة كاني أن أعلى تأثري تثبيط / linoleic acid B-carotene ختباراالنتائج اخلاصة ب
تقودنا هذه النتائج إىل .60 %حوايل تأثريا متوسطا بنسب S. chamaecyparissusأظهرت مستخلصات نبات
لنبات احلرمل ترجع إىل قدرهتا اإلزاحية للجذور B-caroteneاستنتاج أن النشاطية املضادة ألكسدة جزيئات
،تخلصات متلك نشاطية مضادة لألكسدة عاليةسالنتائج أن مجيع امل أظهرتف FTCأما فيما خيص اختبار .احلرة
تقريبا، واليت فاقت نشاطية كل من % 100 بنسبة CrE-Scفقد سجلت أعلى نشاطية تثبيطية على مستوى
VitC وBHT . تشري العديد من الدراسات أن النشاطية املضادة ألكسدة الليبيدات للمستخلصات النباتية تعود
إال أنه فيما ،(Sarikurkcu ،2008 ؛2008وآخرون، Ozsoy)حتوائها على مركبات عديدات الفينول أساسا ال
مع إال الطردي مل يظهر هذا التناسب ، TBAو FTCو B-caroteneيف اختبار يهالاحملصل عنتائج الخيص
وجدههذا ما و لفينوليةري إىل نوعية املركبات اقد يعود هذا التأث، FTCمستخلصات نبات احلرمل يف تقنية
Tuberoso وجد أن كل من بنية و ذوبانية عديدات الفينول يف الدهون متثل العوامل األساسية .(2009) وآخرون
م يف النشاطية املضادة لألكسدة وهي املسؤولة عن اإلندماج العميق هلذه املركبات ضمن الطبقة اليبيدية كاليت تتح
(Djeridane ،2010وآخرون.) ت دراسة سابقة من طرف أجربHayet وجد أن و (2010) ،وآخرون

يف ظهرت أعلى نشاطية وقد FTC استعمالمتلك نشاطية مضادة لألكسدة ب P. harmalaمستخلصات أوراق
.املستخلص املثانويل
يونات تعترب النشاطية اإلستخالبية لأليونات املعدنية ذات أمهية بالغة وهذا يرجع لقدرهتا على خفض تراكيز األ
تفاعالت سلسلة األكسدة على مستوى النظام انطالقاجلذور احلرة األولية و حتفيز نتاج إ ناملعدنية املسؤولة ع
Liu)اهليدروبريوكسيدات وتفكيك Fentonكما تعمل املركبات اإلستخالبية على تثبيط تفاعالت ،البيولوجي
قد يعود التأثري ،CrE-Ph على مستوى IC50 ةقيمى من خالل النتائج احملصل عليها كانت أعل .(2010وآخرون،
يف حني مل يظهر هذا التناسب يف ا،إىل املركبات الفينولية املتواجدة هبملستخلصات نبات احلرمل اإلستخاليب
مثل ، يف استخالب أيونات احلديد الثنائي غري فالفونيديةمستخلصات نبات اجلعدة، مما يعتقد تدخل مركبات
النتائج اإلمكانية العالية للمركبات تظهر أ. (2011، وآخرون Cornago)ينية واألمحاض العضوية األمحاض األم
(.2010) وآخرون Sahreen يتوافق معهذا ما ،القطبية على استخالب املعادن مقارنة باملركبات غري القطبية
والتأثري S. chamaecyparissusو P. harmalaبواسطة مستخلصات XOنسبة تثبيط إنزم نتائج أظهرت
O2اإلزاحي هلا على جذر يليها EAE-Sc و CHE-Scكان على مستوى أن أعلى تأثري تثبيطي ،-•
CHE-Ph و EAE-Ph . وقدرهتا احملتوى الفينويل لكال النبتتني عالقة بنيال غيابيتضح من خالل النتائج
قد يعود إىل تدخل العديد من املركبات XORثبيط إنزم الباحثني أن ت بعضيف حني أشار ،XOالتثبيطية إلنزم
(.2013 وآخرون، Bakasso ؛2012 وآخرون، (Sowndhararajanخصوصا الفالفونيدات النشطة
رضة لألكسدة وذلك نظرا تعترب كريات الدم احلمراء من أكثر اخلاليا املتواجدة يف اجلسم، كما أهنا األكثر ع
ها على األيونات املعدنية ئها باألكسجني باإلضافة إىل احتوامن الليبيدات وغناها على كمية عالية ئإلحتوا
P. harmalaإختبار قدرة مستخلصات مت (. 2012وآخرون، (Alvarez-Suarezكاحلديد والنحاس
اإلحناللمن t-BHP و AAPHكل من باحملفزة على محاية كريات الدم احلمراء S. chamaecyparissusو

in vitro وin vivo نتائج أظهرت .على التوايلin vitro ستخلصاتأن مMOHE-Ph وCrE-Ph
CHE-Ph و CrE-Scو MOHE-Scبينما متلك ،ويل الدمالإلحنل مثبطاتأثريا متلك EAE-Scو EAE-Phو
ات املتواجدة هبذه إىل بنية املركب أثريهذا االختالف يف التيعود أن ميكن ،ويل الدمالإلحنلتأثريا حمفزا CHE-Scو
اليت هلا القدرة إىل الفالفونيداتللمستخلصات وي ل الدمالاإلحن ثبيطفعلى سبيل املثال قد يعود ت ،املستخلصات
وآخرون، (Chaudhuri على اإلندماج ضمن أغشية كريات الدم احلمراء وتعمل على محايتها من عملية األكسدة
لى تكوين تعمل ع اليت ،Saponineاقي املستخلصات إىل مركبات لب ويل الدمالاإلحن، بينما قد يرجع (2007
de Freitas)نفاذية الغشاء البالزمي يف ريغيمسببة تالربوتينات والفوسفولبيدات معقدات مع الستريوالت و
حتلل كريات الدم EAEالفالفونيدات السكرية املتواجدة مبستخلص تثبط أن ميكنكما ،(2008 آخرون،و
حتلل كريات الدم على حتفيز CHEحني تعمل الفالفونيدات غري السكرية املتواجدة مبستخلص احلمراء، يف
قويا مثبطا لإلحنالل تأثريا MOHE-Phأبدى فقد in vivoنتائج اإلحنالل الدموي أما فيما خيص .احلمراء
اية كريات الدم احلمراء حبم املستخلصاتتقوم .in vitro للنتائج احملصل عليها ثالامم MOHE-Scالدموي يليه
Chaudhuri)داخل الكريات احلمراء تهادرجة نفاذياملتواجدة هبا و من األكسدة حسب نوع املركبات الفينولية
(.2007وآخرون،
in vivoقمنا مبواصلة الدراسة in vitroجلذور احلرة على اذين النبتتني التأثري املثبط هلوعليه وبعد التأكد من
كبد مصل و شاطية املضادة لألكسدة على مستوى ندراسة تأثري املستخلصات امليثانولية على ال وذلك من خالل
.الفئران
P. harmala كل من للمستخلص املثانويل ل( CPA)قدرت النشاطية املضادة لألكسدة الكلية للبالزما
CPAيف أثريتأي د مل يالحظ وجو .reducing powerو DPPHباستعمال اختبار S. chamaecyparissusو
تغريا معنويا CrE-Scيف حني أبدى مستخلص ،بالنسبة لكلى املستخلصني DPPHفيما خيص اختبار

ختتلف قدرة املستخلصات النباتية على . reducing powerاختبار يف CPAيف تغريا غري معنوي CrE-Phو
مل يظهر تغريا يف ( (2013وآخرون da Silvaأجريت دراسة سابقة من طرف فعلى سبيل املثال ،CPAتغيري
CPA باستعمال اختبارORAC وFRAP، يف حني مل ،(يوما 15)لقصر مدة التجربة اسناد هذاإذ يعتقد
.(Widlansky ،2005)إال بعد إطالة زمن التجربة FRAPو ORACباستعمال اختبار CPAيظهر تغري يف
P. harmalaملة باملستخلص املثانويل لكل من عند اجملاميع املعا MDAأن قيمة نتائج البينت
ا يدل على أن املستخلصني قاما حبماية نسيج الكبد من فوق مم كانت منخفضة S. chamaecyparissusو
يف كبد الفئران املعاجلة باملستخلص GSHهذه الدراسة ارتفاعا معنويا يف مستوى تسجل. أكسدة الليبيدات
يعود هذا االرتفاع إىل قدرة املستخلصات علىقد S. chamaecyparissusو P. harmalaاملثانويل لكل من
أو اإلثنني معا GSHخفض اإلجهاد التأكسدي املؤدي إىل التقليل من هدم أو GSH لزيادة البناء احليوي
(Rajasekaran ،2005وآخرون). جلنا خالل هذه الدراسة إرتفاعا معنويا يف نشاطية سCAT عند الفئران
% 35بنسبة قدرت ب S. chmaecyparissusو P. harmalaملعاجلة باملستخلص املثانويل لكل من ا
قدرة هذه املستخلصات على حتسني نشاط اإلنزميات املضادة للتأكسد يعود إىلهذا ،على التوايل % 59,16و
(Finley ،؛ 2011وآخرونSilva ،2012وآخرون)، ة التعبري عن املورثات املستخلصات على زياد أو إىل عمل
.SOD ،CAT ،GPXاملشفرة لإلنزميات املضادة لألكسدة مثل
قياس القدرة املضادة لألكسدة للنباتات، إال أنه يف بعض لبكثرة in vitroتستعمل اإلختبارات املضادة لألكسدة
قلة إىل ئيسي لذلك يعود السبب الر و in vivo و in vitroاألحيان حيث تداخل بني النشاطية املضادة لألكسدة
، باإلضافة إىل قلة نصف عمر املركبات الفينولية يف البالزما anthocyaninsإمتصاصية بعض املركبات خصوصا
والذي يعد بالساعات، كل هذه العوامل حتد من استعمال املركبات الفينولية لتقدير النشاطية املضادة لألكسدة
in vivo، جيب تكثيف الدراسات لذلكin vivo لتحديد الدور احلقيقي للمركبات املضادة لألكسدة

( Lotito و Fre،2006.)
متلك نشاطية S. chamaecyparissusو P. harmalaيتضح من خالل النتائج احملصل عليها أن كل من نبتة
األحباث سيع جيدر تو عالية مضادة لألكسدة خمربيا ويف الكائن احلي ونشاطية مضادة لإلحنالل الدموي، لذلك
وتنقية املركبات النشطة ودراسة تأثرياهتا املضادة لألكسدة، دراسة آلية عمل كل مركب على خالل عزل وجتزئة من
أمراض إلتهابية، مرض )مشاهبة ملا عند اإلنسان باستعمال حيوانات مصابة بأمراض in vivoوتنويع الدراسة حدة
ا بأمان مدراسة اجلانب السمي للنبتتني للتأكد من استعماهلو ...( ها،إلتهاب املفاصل الرثوي، فرط احلساسية وغري
.يف اجلانب الصيدالين

عــــــــــــة المراجــــــــــقائم
Abadeh, S., Killacky, J., Benboubetra, M., Harrison, R. (1992). Purification and partial
characterization of xanthine oxidase from human milk. Biochimica et Biophysica Acta 1117, 25-32.
Abbasi M A, Saleem H, Rehman A, Riaz T and Ajaib M. (2013). Determination of Antioxidant
Activity and Phytoconstituent Screening of Euphorbia heterophylla Linn. British Journal of
Pharmaceutical Research. 3(2): 202-216.
Agbor, G.A., Kuate, D., Oben, J.E. (2007). Medicinal plants can be good source of antioxidants: case
study in Cameroon. Pakistan Journal of Biological Sciences 10, 537-544.
Al-Muhtaseb, N., Al-Kaissi, E., Thawaimi, A.J., Muhi Eldeen, Z., Al-Muhtaseb, S., Al-Saleh, B.
(2012). The role of human xanthine oxidoreductase (HXOR), anti-HXOR antibodies, and
microorganisms in synovial fluid of patients with joint inflammation. Rheumatology International
32(08), 2355-2362.
Alvarez, M. A., Debattista, N. B., Pappano, N. B. (2008). Antimicrobial activity and synergism of
some substituted flavonoids. Folia Microbiologica (Praha) 53, 23-28.
Amessis-Ouchemoukh, N., Madani, K., Falé, P.L.V. Serralheiro, M. L., Araújo, M.E.M.
(2014). Antioxidant capacity and phenolic contents of some Mediterranean medicinal
plants and their potential role in the inhibition of cyclooxygenase-1 and
acetylcholinesterase activities. Industrial Crops and Products 53, 6-15.
Amzad Hossain, M., Shah, M.D. (2011). A study on the total phenols content and antioxidant activity
of essential oil and different solvent extracts of endemic plant Merremia borneensis. Arabian
Journal of Chemistry 08, 66-71.
Antonella, S., Mario, S., Maria, L., Daniela, M., Francesco, B., Francesco, C. (1995). Flavonoids as
antioxidant agents: importance of their interaction with biomembrane. Free Radical Biology and
Medicine 19, 481-486.
Antunes, F., Derick, H., Cadenas, E. (2002) Relative contributions of heart mitochondria glutathione
peroxidase and catalase to H2O2 detoxification in in vivo conditions. Free Radical Biology and
Medecine 33 (9), 1260-1267.
Aqil, F., Ahmad, I., Mehmood, Z. (2006). Antioxidant and free radical scavenging properties of
twelve traditionally used Indian medicinal plants. Turkish Journal of Biology 30, 177-183.

Asgarpanah, J., Ramezanloo, F. (2012). Chemisty, pharmacology and medicinal properties of
Peganum harmala L.. African Journal Pharmacology Pharmacologica 22, 1573-1580.
Aslan, A., Güllüce, M., Sökmen, M., Adigüzel, A., Sahin, F., Özkan, H. (2006). Antioxidant and
antimicrobial properties of lichens Cladonia foliacea, Dermatocarpon miniatum, Everinia
divaricata, Everinia prunastri and Neofuscella pulla. Pharmacology Biology 44, 247-252.
Astulla, A., Zaima, K., Matsuno, Y., Hirasawa, Y., Ekasari, W., Widyawaruyanti, A., et al. (2008).
Alkaloids from the seeds of Peganum harmala showing antiplasmodial and vasorelaxant activities.
Journal of Natural Medicines 62, 470-472.
Atmani, D., Benboubetra, M., Harrison, R. (2004) Goat’s milk xanthine oxidoreductase is grossly
deficient in molybdenum. Journal of Dairy Research 71, 7-13.
Augustin, A.J., Boker, T., Blumenroder, S.H., Lutz, J., Spitznas, M. (1994). Free radical scavenging
and antioxidant activity of allopurinol and oxypurinol in experimental lensinduced uveitis.
Investigation Ophthalmology Visual Science 35, 3897-3904.
Avis, P.G., Bergel, F., Bray, R.C. (1956). Cellular constituents. The chemistry of xanthine oxidase.
Part III. Estimations of the cofactors and the catalytic activities of enzyme fractions from cow’s
milk. Journal of the Chemistry Society 1219-1226.
Babior, B.M. (1999). NADPH oxidase: an update. Blood 93, 1464-1476.
Babu, P.V.A., Sabitha, K.S., Shyamaladevi, C.S. (2006). Therapeutic effect of green tea extract on
oxidative stress in aorta and heart of streptozotocin diabetic rats. Chemico-Biological Interactions
162, 114-120.
Bader, G., Plohmann, B., Hiller, K., Franz, G. (1996). Cytotoxicity of triterpenoid saponins. Part 1:
activities against tumor cells in vitro and hemolytical index. Pharmazie 51 (6), 414-417.
Baghiani, A., Boumerfeg, S., Belkhiri, F., Khennouf, S., Charef, N., Harzallah, D., Arrar L., Abdel-
Wahhab, M. (2010). Antioxidant and radical scavenging properties of Carthamus caeruleus
L.extracts grow wild in Algeria flora. Comunicata Scientiae 1, 128-136.
Baghiani, A., Djarmouni, M., Boumerfeg, S., Trabsa, H., Charef, N., Khennouf, S., Arrar, L. (2012).
Xanthine Oxidase Inhibition and Antioxidant Effects of Peaganum harmala Seed Extracts.
European Journal of Medicinal Plants 2(1), 42-56.
Baghiani, A., Harrison, R., Benboubetra, M. (2003). Purification and partial characterisation of camel
milk xanthine oxidoreductase. Archives of Physiology and Biochemistry 111, 407-414.

Bahorun, T., Gressier, B., Trotin, F., Brunete, C., Dine, T., Vasseur, J., Gazin, J.C., Pinkas, M.,
Luycky, M., Gazin, M. (1996). Oxigen species scavenging activity of phenolic extract from
howthorn fresh plant organs and pharmaceutical preparation. Arzneimittel Forsh / Drug Research
1-6.
Bahorun, T., Soobrattee, M.A., Luximon-Ramma, V., Aruoma, O.I. (2006). Free radicals and
antioxidants in cardiovascular health and disease. Internet Journal Medicine Update 1, 25-41.
Bakasso, S., Lamien-meda, A., Lamien, C.E., Kiendrebeogo, M., Coulibaly, A.Y., Compaoré, M.,
Meda, N.R., Nacoulma, O.G. (2013). In vitro Inhibition of Acetyl Cholinesterase, Lipoxygenase,
Xanthine Oxidase and Antibacterial Activities of Five Indigofera (Fabaceae) Aqueous Acetone
Extracts from Burkina Faso. Research Journal of Biological Sciences 5(3), 115-122.
Balaban, R.S., Nemoto, S., Finkel, T. (2005). Mitochondria, oxidants, and aging. Cell 120, 483-495.
Balasundram, N., Sundram, K., Samman, S. (2006). Phenolic compounds in plants and agri-industrial.
by-products: Antioxidant activity, occurrence, and potential uses. Food Chemistry 99, 191-203.
Barrero, A.F., Mar Herrador, M., Quilez, J.F., Alvarez- Manzaneda, R., Portal, D., Gavin, J.A,
Gravalos, D.G., Simmonds, M.S.J., Blaney, W.M. (1999). Bioactive sesquiterpenes from Santolina
rosmarinifolia subsp. Canescens. A conformational analysis of the germacrane ring.
Phytochemistry 51, 529-541.
Barriere, C., Centeno, D., Lebert, A., Leroy-Setrin, S., Berdague, J.L., Talon, R. (2001). Roles of
superoxide dismutase and catalase of Staphylococcus xylosus in the inhibition of linoleic acid
oxidation. FEMS Microbiology Letters 201, 181-185.
Beani, J. C. (1995). Actions biologiques du rayonnement solaire sur la peau. Review Internationnal
Pédiatric 259, 2-7.
Bellakhdar, J.L. (1997). A pharmacopee marocaine traditionnelle. Medecine arabe ancienne et savoirs
populaires. Paris: Ibis Press.
Ben Abid, Z., Feki, M., Hédhili, A., Hédi Hamdaoui, M. (2007). Artemisia herba-alba Asso
(Asteraceae) Has Equivalent Effects to Green and Black Tea Decoctions on Antioxidant Processes
and Some Metabolic Parameters in Rats. Annals of Nutrition and Metabolism 51, 216-222.
Benzie, I.F., Strain, J.J. (1996). The ferric reducing ability of plasma (FRAP) as a measure of
bantioxidant powerQ the FRAP assay. Analytical Biochemistry 239, 70-76.

Berrougui, H., Martin-Cordero, C., Khalil, A., Hmamouchi, M., Ettaib, A., Marhuenda, E., et al.
(2006). Vasorelaxant effects of harmine and harmaline extracted from Peganum harmala L. seeds
in isolated rat aorta. Pharmacological Research 54, 150-157.
Berry, C.E., Hare, J.M. (2004). Xanthine oxidoreductase and cardiovascular disease: molecular
mechanisms and pathophysiological implications. The Journal of Physiology 555, 589-606.
Bieber, J. D., Terkeltaub, R. A. (2004). Gout: on the brink of novel therapeutic options for an ancient
disease. Arthritis and Rheumatology 50, 2400-2414.
Blake, D. R., Stevens, C. R., Sahinoglu, T. M., Ellis, G., Gaffney, K., Edmonds, S., Benboubetra, M.,
Harrison, R., Jawed , S., Kanezler, J., Millar, T. M., Winyord, P. G., Zhang, Z. (1997). Xanthine
oxidase; four roles for the enzyme in rheuatoid pathology. Biochemical Society Transactions 25,
812-816.
Bondet, V., Williams, W.B., Berset, C. (1997). Kinetic and mechanism of antioxidant activity using
the DPPH free radical method. Lebensmittel -Wissenschaft und Technologie 30, 609-615.
Borg, J., Beeber, A. (2008). Le stress oxydant. In: Biochimie métabolique, édité par: Ellipses: France,
2eme
édition, pp. 257-273.
Borges, F., Fernandes, E., Rolivera, F. (2002). Progress towards the discovery of xanthine oxidase
inhibitors. Current Medicinal Chemistry 9, 195-217.
Böttger, E. C., Schacht J. (2013). The mitochondrion: A perpetrator of acquired hearing loss. Hearing
Research 303, 12-19.
Bougatef, A., Hajji, M., Balti, R., Lassoued, I., Triki-Ellouz , Y., Nasri, M. (2009). Antioxidant and
free radical-scavenging activities of smooth hound (Mustelus mustelus) muscle protein
hydrolysates obtained by gastrointestinal proteases. Food Chemistry 114, 1198-1205.
Brand-Williams, W., Cuvelier, M.E., Berset, C. (1995). “Use of free radical method to evaluate
antioxidant activity”. Lebensmittel-Wissenschaft und –Technologie 28, 25-30.
Bray, R.C. (1975). Molybdenum iron–sulfur flavin hydroxylases and related enzymes. In Boyer P.D.
The enzyme. New York: Academic Press 299-419.
Brondino, C. D., Romao, M. J., Moura, I., and Moura, J. J. (2006). Molybdenum and tungsten
enzymes: the xanthine oxidase family. Current Opinion in Chemical Biology. 10, 109-114.
Bruneton, J. (1993). Pharmacognosie, phytochimie, plantes médicinales. Technique et documentation,
Lavoisier, 2eme
ed., Paris 915.

Bukhari, N., Choi, J. H., Jeon, C. W., Park, H. W., Kim, W. H., Khan, M. A., et al. (2008).
Phytochemical studies of the alkaloids from Peganum harmala. Applied Chemistry 12, 101-104.
Burits, M., Bucar, F. (2000). Antioxidant activity of Nigella sativa essential oil. Phytotheraphy
Research 14, 323-328.
Cao, G., Prior, R.L. (1998). Comparison of different analytical methods for assessing total antioxidant
capacity of human serum. Clinical Chemistry 44, 1309-1315.
Casetta, I., Govoni, V., Granieri, E. (2005). Oxidative stress, antioxidants and neurodegenerative
diseases. Current Pharmaceutical Design 11(16), 2033-2052.
Çelik, S.E., Ozyürek, M.,Güçlü , K., Apak, R. (2010). Solvent effect s on the antioxida nt capac ity of
lipophilic and hy drophilic antioxidants measured by CUPRA C, ABTS/ persulp hate and FRAP
methods. Talanta 81, 1300–1309.
Chandrasekara, A., Shahidi, F. (2010). Content of insoluble bound phenolics in millets and their
contribution to antioxidant capacity. Journal of Agricultural and Food Chemistry 58, 6706–6714.
Chang, C., Yang, M., Wen, H., Chern, J. (2002). Estimation of total flavonoid content in propolis by
two complementary colorimetric methods. Journal Food Drug Analaysis 10, 178-182.
Chapple, I.L., Mason, G.I., Garner, I., Matthews, J.B., Thorpe, G.H., Maxwell, S.R., Whitehead, T.P.
(1997). Enhanced chemiluminescent assay for measuring the total antioxidant capacity of serum,
saliva and crevicular fluid. Annals of Clinical Biochemistry 34(4), 412- 421.
Chaudhuri, S., Banerjee, A., Basu, K., Sengupta, B., Sengupta, P.K. (2007). Interaction of flavonoids
with red blood cell membrane lipids and proteins: antioxidant and antihemolytic effects.
International Journal of Biological Macromolecules 41, 42-48.
Chen Y, Huang B, He J, Han L, Zhan Y, Wang Y. (2011). In vitro and in vivo antioxidant effects of
the ethanolic extract of Swertia chirayita. Journal of Ethnopharmacology 136, 309–315.
Christensen, L.P.(1992). Acetylenes and related compounds in Anthemideae. Phytochemistry 31, 7-
49.
Cazarolli L. H., Zanatta L., Alberton E .H., Figueiredo M. S., Folador P., Damazio R. G., Pizzolatti M.
G. and Silva F. R.(2008). Flavonoids: prospective drug candidates. Mini Rev Med Chem. 8: 1429-
1440
Chung, Y.C., Chen, S.J., Hsu, C.K., Chang, C.T., Chou, S.T. (2005). Studies on the antioxidative
activity of Graptopetalum paraguayense E. Walther. Food Chemistry 91, 419-424.
Clairborne, A. (1985). Catalase activity. In: CRC Handbook of methods for oxygen radical research.
Ed. Greenwald. R.A. CRC Press. Boca. Raton 283-284.

Conner, E. M., Grisham, M. B. (1996). Inflammation, free radicals, and antioxidants. Nutrition 12,
274-277.
Coqueiro, A., Regasini, L.O., Gutkoski, S.C.S., Marçal, M., Queiroz, F., Dulce, H., iqueira, S.,
Vanderlan da, S.B. (2013). Free Radical Scavenging Activity of Kielmeyera variabilis
(Clusiaceae). Molecules 18, 2376-2385.
Cos, P., Ying, L., Calomme, M., Hu, J.P., Cimanga, K., Van Poel, B., Pieters, L., Vlietinck, A.J.,
Vanden Berghe, D. (1998). Structure activity relationship and classification of flavonoids as
inhibitors of xanthine oxidase and superoxide scavengers. Journal of Natural Products 61, 71-76.
Costantino, L., Albasini , A., Rastelli, G., and Benvenuti, S. (1992). Activity of polyphenolic crude
extracts as scavengers of superoxide radicals and inhibitors of xanthine oxidase. Planta Medica 58,
342-345.
Cuendet, M., Hostettmann, K., Potterat, O. (1997). Iridoid glucosides with free radical scavenging
properties from Fagraea blumei. Helvetica Chimica Acta 80, 1144-1152.
da Silva, J K., Cazarin, C B B., Colomeu, T. C., Batista, Â. G., Meletti ,L.M.M., Paschoal, J. A. R.,
Júnior , S .B., Furlan, M. F., Reyes, F. G. R., Augusto, F., Júnior, M. R. M., Zollner, R. de Lima.
(2013). Antioxidant activity of aqueous extract of passion fruit (Passiflora edulis) leaves: In vitro
and in vivo study. Food Research International 53, 882-890.
da Silva, E.J.A., Oliveira, A. B., Lapa, A.J. (1994). Pharmacological evaluation of the anti-
inflammatory activity of a citrus bioflavonoid, hesperidin, and the isoflavonoids, duartin and
claussequinone, in rats and mice. Journal of Pharmacy and Pharmacology 46(2), 118-122.
da Silva, J.A.T. (2004). Mining the essential oils of the Anthemidea. African Journal of Biotechnology
3(12), 706-720.
Dalle-Donne, I., Rossi, R., Colombo, R., Giustarini, D., Milzani, A. (2006). Biomarkers of oxidative
damage in human disease. Clinical Chemistry 52, 4601–4623.
Das, K.C., Lewis-Molock, Y., White, C.W. (1997). Elevation of manganese superoxide dismutase
gene expression by thioredoxin. American Journal of Respiratory Cell and Molecular Biology 17,
713-726.
Dastmalchia, K., Dormana, H.J.D., Oinonen, P.P., Darwisd, Y., Laaksoa, I., Hiltunena, R. (2008).
Chemical composition and in vitro antioxidative activity of a lemon balm (Melissa officinalis L.)
extract. Food Science and Technology 41, 391-400.

De beer, D., Joubert, E., Gelderblom, W. C. A., Manley, M. (2002). Phenolic compounds: a review of
their possible role as in vivo antioxidants of wine. South African Journal of Enology and Viticulture
20, 48-61.
de Freitas, M. V., Netto, M. de Cássia. R., da Costa Huss, J.C., de Souza, T.M.T., Costa, J. O.,
Firmino, C. B., Penha-Silva, N. (2008). Influence of aqueous crude extracts of medicinal plants on
the osmotic stability of human erythrocytes. Toxicology in Vitro 22, 219–224.
de Logu, A., Loy, G., Pellerano, M.L., Bonsignore, L., Schivo , M.L. (2000). In a ctivation of HSV-1
and HSV-2 and prevention of cell-to-cell virus spread by Santolina insularis essential oil. Antiviral
Research 48, 177-185.
de Lourdes Reis Giada, M. (2013). Food Phenolic Compounds: Main Classes, Sources and Their
Antioxidant Power. Chapter 4. licensee In Tech. 88-112.
de Mello Andrade, J. M., Fasolo, D. (2014). Polyphenol Antioxidants from Natural Sources and
Contribution to Health Promotion Chapter 20. Polyphenols in Human Health and Disease.
Debuigne, G. (1974). Larousse des plantes qui guérissent. Larousse paris, pp. 254.
Decker, E. A., Welch, B. (1990). Role of Ferritin as lipid oxidation catalyst in muscle food. Journal of
Agricultural and Food Chemistry 38, 674-677.
Decorde, K., Teissedre, P.L., Auqer, C., Cristol, J.P., Rouanet, J.M. (2008). Phenolics from purple
grape, apple, purple grape juice and apple juice prevent early atherosclerosis induced by an
atherogenic diet in hamsters. Molecular Nutrition & Food Research 2, 400-407.
Defraigne, J.O., Pincemail, J. (2007). Stress oxydant et antioxydants: mythes et réalités. Revue
Médicale de Liège 62, 4.
Delattre, J., Beaudeux, J.L., Bonnefont-Rousselot. (2005). Radicaux libres et stress bxydant: aspects
biologiques et pathologiques. Lavoisier edition TEC & DOC editions medicales internationales
Paris 1-405.
Delmas-Beauvieux, M.C., Combe, C., Peuchant, E., Carbonneau, M.A., Dubourg, L., de Precigout, V.,
Aparicio, M., Clerc, M. (1995). Evaluation of red blood cell lipoperoxidation in hemodialysed
patients during erythropoietin therapy supplemented or not with iron. Nephron 69, 404-410.
Deuticke, B., Heller, K.B., Haest, W.M. (1987). Progressive oxidative membrane damage in
erythrocytes after pulse treatment with t-butylhydroperoxide. Biochimica et Biophysica Acta 899,
113-124.

Deuticke, B., Klonk, S., Haest, C.W.M. (1992). Erythrocyte membrane barrier formation: role of
bilayer polarity and skeletal proteins. Biochemical Society Transactions 20, 769-773.
Dhanabal, S.P., Rahul, J., Priyanka Dwarampudi, L., Muruganantham, N., Raghu, P.S. (2012).
Hepatoprotective activity of Santolina chamaecyparissus Linn against D-Galactosamine Induced
Hepatotoxicity in Rats. Pharmacognosy Communications. 2.
Djanna, F., Cornago, Rowena Grace, O., Rumbaoa, Inacrist, M., Geronimo. (2011). Philippine Yam
(Dioscorea spp.) Tubers Phenolic Content and Antioxidant Capacity. Philippine Journal of Science
140(2), 145-152.
Djarmouni M., Boumerfeg S., Baghiani A., Boussoualim N., Zerargui F., Trabsa H., Belkhiri F.,
Khennouf S., Arrar L. (2012). Evaluation of Antioxidant and Antibacterial Properties of Peaganum
Harmala. Seed Extracts. Research Journal of Pharmaceutical, Biological and Chemical Sciences.
3(4), 1109-1119.
Djeridane, A., Yousfi, M., Brunel, J. M., Stocker, P. (2010). Isolation and characterization of a new
steroid derivative as a powerful antioxidant from Cleome arabica in screening the in vitro
antioxidant capacity of 18 Algerian medicinal plants. Food and Chemical Toxicology 48, 2599-
2606.
Djeridane, A., Yousfi, M., Nadjemi, B., Vidal , N., Lesgards, JF., Stocker, P. (2007). Screening of
some Algerian medicinal plants for the phenolic compounds and their antioxidant activity.
European Food Research and Technology 224, 801-809.
Droge, W. (2002). Free radicals in the physiological control of cell function. Physiological Reviews
82, 47-95.
Dube, A., Misra, P., Khaliq, T., Tiwari, S., Kumar, N., Narender, T. (2011). Therapeutic potential of
Harmala (Peganum harmala L.) seeds with an array of pharmacological activities. In: Nuts &
Seeds in Health and Disease Prevention, edited by: Victor R. Preedy, Ronald Ross Watson and
Vinood B. Patel, ELSEVIER: UK, USA 601-609.
Dykes, L., Rooney, L. W. (2007). Phenolic compounds in cereal grains and health benefits. Cereal
Foods World 50, 105-117.
Edeas, M. (2007). Les polyphénols et les polyphénols de thé. Phytothérapie 5, 264-270.
EL Rhaffari, L. (2008). Catalogue des plantes potentielles pour la conception de tisanes.
Errachidia.14p.
Ellman, G.L. (1959). Tissue sulfhydryl groups. Archives of Biochemistry and Biophysics 82, 70-77.

Elmastas, M., Gulcin, I., Isildak, O., Kufrevioglu, O.I., Ibaoglu, K., Aboul-Enein, H.Y. (2006).
Radical scavenging activity and antioxidant capacity of bay leaf extracts. Journal of Iranian
chemical society 03(03), 258-266.
Erben-Russ, M., Bors, W., Saran, M. (1987). Reactions of linoleic acid peroxyl Radicals with phenolic
antioxidants: a pulse Radiolysis study. International Journal of Radiation Biology 52(3), 393-412.
Evans, W.J. (2000). Vitamin E, vitamin C, and exercise. The American Journal of Clinical Nutrition
72, 647-652.
Ezer, N., Mumcu, A.O. (2006). Folk medicines in Merzifon (Amasya, Turkey). Turkish Journal of
Botany 30, 223-230.
Fan, H., Yang, G.Z, Zheng, T., Mei, Z. N. X.M., Liu Chen, Y., Chen, S. (2010). Chemical
Constituents with Free-Radical-Scavenging Activities from the Stem of Microcos paniculata.
Molecules 15, 5547.
Fang, Y.Z. (2002). Free radicals and nutrition. In: Fang YZ, Zheng RL, eds. Theory and application of
free radical biology. Beijing: Scientific Press 647.
Fattouch, S., Caboni, P., Coroneo, V., Tuberoso, C.I.G., Angioni, A., Dessi, S., Marzouki, N., Cabras,
P. (2007). Antimicrobial activity of tunisian quince (Cydonia oblonga Miller) pulp and peel
polyphenolic extracts. Journal of Agricultural and Food Chemistry 55, 963-969.
Ferrari, A.M., Sgobba, M., Gamberina, M.C., Rastelli, G. (2007). Relationship between quantum-
chemical descriptors of proton dissociation and experimental acidity constants of various
hydroxylated coumarins. Identification of the biologically active species for xanthine oxidase
inhibition. European Journal of Medicinal Chemistry 42, 1028-1031..
Finaud, J., Lac G., Filaire, E. (2006). Oxidative Stress Relationship with Exercise and Training.
Sports Medicine 36(4), 327-358.
Finley, J. W., Kong, A. N., Hintze, K. J., Jeffrey, E. H., Ji, L. L., Lei, X. G. (2011). flavonoids and
risk of coronary heart disease: the Zutphen Elderly Study. Lancet 342(8878), 1007-1011.
Fournier, P. (1948). Livre des plantes medicinales et venéneuses de France. Tome Ш. Lechevalier-
Paris, pp. 415 .
Frankel, E.N., Huang, S.W. Kanner, J., German, J.B. (1994). Interfacial phenomena in the evaluation
of antioxidants: Bulk oils vs. emulsions. Journal of Agricultural and Food Chemistry 42, 1054-
1059.

Freeman, B.A., Crapo, J.D. (1981). Hyperoxia increases oxygen radical production in rat lungs and
lung mitochondria. Journal of Biological Chemistry 256, 10986-10992.
Frison, G., Favretto, D., Zancanaro F., Fazzin, G., Ferrara, S. D. (2008). A case of beta-carboline
alkaloid intoxication following ingestion of Peganum harmala seed extract. Forensic Science
International 179, 37-43.
Galati, E. M., Monforte, M. T., Kirjavainen, S., Forestieri, A. M., Trovato, A., Tripodo, M.M. (1994).
Biological effects of hesperidin, a citrus flavonoid. (Note I): antiinflammatory and analgesic
activity. Farmaco 40(11), 709-712.
Gardès-Albert, M., Bonnefont-Rousselot, D., Abedinzadeh, Z. (2003). Comment l'oxygène peut-il
devenir toxique?. L'actualité chimique 14, 91-96.
Gawlik-Dziki, U. (2012). Dietary spices as a natural effectors of lipoxygenase, xanthine oxidase,
peroxidase and antioxidant agents. Food Science and Technology 47, 138-146.
Genestra, M .(2007). Oxyl radicals, redox-sensitive signalling cascades and antioxidants. Cellular
Signalling 19, 1807-1819.
Ghiselli, A., Serafini, M., Maiani, G., Azzini, E., Ferro-Luzzi, A. (1995). A fluorescence-based
method for measuring total plasma antioxidant capability. Free Radical Biology and Medicine 18,
29- 36.
Giner Pons, R. M., Rios Canavate, J. L. (2000). Santolina chamaecyparissus: Especie mediterranea
con potenciales aplicaciones terapeuticas en procesos inflamatorios y transtornos digestivos.
Revista de Fitoterapia 1, 27-34.
Giner, R.M., Rios, J.L., Villar, A. (1988). C N S depressant effects, anti-inflammatory activity and
anti-cholinergic actions of Santolina chamaecyparissus extracts. Phytotherapy Research 12, 37-41.
Girard, A., Madani, S., Boukortt, F., Cherkaoui-Malki, M., Belleville, J., Prost, J. (2006). Fructose
enriched diet modifies antioxidant status and lipid metabolism in spontaneously hypertensive rats.
Nutrition 22, 758-766.
Godber, B., Sanders, S., Harrison, R., Eisenthal, R., Bray, R.C. (1997) of xanthine oxidase in human
milk is present as the demolybdo form, lacking molybdopterin. Biochemical Society Transactions
25, 519 .
Goel, N., Singh, N., Saini, R. (2009). Efficient in vitro multiplication of Syrian Rue (Peganum
harmala) using 6-benzylaminopurine preconditioned seedling explants. Nature Sciences 7, 129-
134.

Gotti, R. (2011). Capillary electrophoresis of phytochemical substances in herbal drugs and medicinal
plants. Journal of Pharmaceutical and Biomedical Analysis 55, 775-801.
Gramza, A., Korczak, J., Amarowicz, R. (2005). Tea polyphenols – their antioxidant properties
and biological activity - a review. Polish Journal of Food And Nutrition Sciences 3, 219-235.
Granger, D.N., Rutili, G., and McCord, J.M. (1981). Role of superoxide radical in feline intestinal
ischemia. Gastroenterology 81, 22-29.
Grigorov, B. (2012). Reactive oxygen species and their relation to carcinogenesis. Trakia Journal of
Sciences 10 (3), 83-92.
Grinberg, L.N., Newmark, H., Kitrossk, N., Rahamim, E., Chevion, M., Rachmilewitz, E.A. (1997).
Protective effects of tea polyphenols against oxidative damage to red blood cells. Biochemical
Pharmacology 54 (9), 973-978.
Grinberg, L.N., Rachmilewitz, E.A., Newmark, H. (1994). Protective effects of rutin against
hemoglobin oxidation. Biochemical Pharmacology 48, 643-649.
Gulcin, I., Huyut, Z., Elmastas, M., Aboul-Enein, H. Y. (2010). Radical scavenging and antioxidant
activity of tannic acid. Arabian Journal of Chemistry 3, 43-53.
Habib, H.M., Ibrahim, W. H. (2011). Effect of date seeds on oxidative damage and antioxidant status
in vivo. Journal of the Science of Food and Agriculture 91(9), 1674-1679.
Hadizadeh, M., Keyhani, E., Keyhani, J., Khodadadi, C. (2009). Functional and structural alterations
induced by copper in xanthine oxidase. Acta Biochimica et Biophysica Sinica 41 (7), 603–617.
Halliwell B (1995). Antioxidant characterization. Methodology and mechanism. Biochemical
Pharmacology 49(10), 1341-1348.
Halliwell, B. (1994). Free radicals and antioxidants : a personal view. Nutrition Reviews 52, 253-265.
Halliwell, B. (1994). Free Radicals, Antioxidants, and Human Disease: Curiosity, Cause, or
Consequence?. Lancet 344, 721-724.
Halliwell, B., Gutteridge, J. M. C. (1986). Oxygen free radicals and iron in relation to biology and
medicine: some problems and concepts. Archives of Biochemistry and Biophysics 246(2), 501-514.
Halliwell, B., Gutteridge, J.M.C. (Eds.) (1999). In: Free Radical in Biology and Medicine, Oxford
University Press, UK 617.

Halliwell, B., Gutteridge, J.M.C., Cross , C. E. (1992). Free radicals, antioxidants, and human disease:
where are we now?. Journal of Laboratory and Clinical Medicine 119(6), 598-620.
Han, S.T., Bhopale, V.M., Om, S.R. (2007). Xanthine oxidoreductase and neurological sequelae of
carbon monoxide poisoning. Toxicology Letters 170, 111-115.
Hanasaki, Y., Ogawa, S., Fukui, S. (1994). The correlation between active oxygens scavenging and
antioxidative effects of flavonoids. Free Radical Biology and Medicine 16, 845-850.
Hapner, C.D., Deuster, P., Chen, Y. (2010). Inhibition of oxidative hemolysis by quercetin, but not
other antioxidants. Journal Of Chemical And Biological Interfaces186(3), 275-279.
Haraguchi, H., Saito, T., Ishikawa, H., et al. (1996). Antiperoxidative components in Thymus vulgaris.
Planta Medica 62, 217-221.
Harborne, J.B., and Williams, C.A. (2000). Advances in flavonoid research since 1992.
Phytochemistry 55, 481-504.
Harris, C.M., Massey, V. (1997). The oxidative half-reaction of xanthine dehydrogenase with NAD;
reaction kinetics and steady-state mechanism. Journal of Biological Chemistry 272, 28335-28341.
Harrison, R. (2002). Structure and function of xanthine oxidoreductase: Where are we now?. Free
Radical Biology and Medicine 33, 774-797.
Harrison, R. (2006). Milk xanthine oxidase: properties and physiological roles. International Dairy
Journal 16, 546-554.
Harvsteen, B.M. (2002). The biochemistry and medical significance of the flavonoids. Pharmacology
and Therapeutics 96, 67-202.
Hassimotto, N. M. A., Pinto, M. D., & Lajolo, F. M. (2008). Antioxidant status in humans after
consumption of blackberry (Rubus fruticosus L.) juices with and without defatted milk. Journal of
Agricultural and Food Chemistry 56(24), 11727-11733.
Hayet, E., Maha, M., Mata, M., Mighri, Z., Laurant, G., Mahjoub, A. (2010). Biological activities of
Peganum harmala leaves. African Journal of Biotechnology 9(48), 8199-8205.
Hellsten-Westing, Y. (1993). Immunohistochemical localization of xanthine oxidase in human cardiac
and skeletal muscle. Histochemistry 100, 215-222.
Hille, R., and Massey, V. (1981). Studies on the oxidative half-reaction of xanthine oxidase. Journal
of Biological Chemistry 256, 9090-9095.

Hille, R., Anderson, R.F. (1991). Electron transfer in milk xanthine oxidase as studied by pulse
radiolysis. Journal of Biological Chemistry 266, 5608-5615.
Holley, A.E. and Cheeseman, K.H. (1993). Mesuring free radical reactions in vivo. British Medical
Bulletin 49, 494-505.
Hollman, P.C.H., Katan, M.B. (1997). Absorpition, metabolism and health effects of dietary
flavonoids in man. Biomedicine and Pharmacotherapy 51, 305-310.
Hsieh, C.Y., Chang, S.T. (2010). Antioxidant activities and xanthine oxidase inhibitory effects of
phenolic phytochemicals from Acacia confusa twigs and branches. Journal of Agricultural and
Food Chemistry 58, 1578-1583.
Huang, T.C., Hsu, H.K., Fu , H.Y., Ho , C.T. (2005). Effect of lactic acid fermentation on quercetin
composition and antioxida tiv e properties of Toona sin ensis lea ves. In F.Shahidi &C.T.Ho (Eds.),
Phenolic compoun ds in foods and nat ur al health products (pp.46–56).Washingt on, DC:ACS
Division of Agricultural and Food Chemistry , Inc..
Ichimori, K., Fukahori, M., Nakazawa, H., Okamoto, K., and Nishino, T. (1999). Inhibition of
xanthine oxidase and xanthine dehydrogenase by nitric oxide. Nitric oxide converts reduced
xanthine-oxidizing enzymes into the desulfo-type inactive form. Journal of Biological Chemistry
274, 7763-7768.
Im, J. H., Jin, Y. R., Lee, J. J., Yu, J. Y., Han, X. H., Im, S. H., et al. (2009). Antiplatelet activity of
beta-carboline alkaloids from Peganum harmala: a possible mechanism through inhibiting PLC
gamma2 phosphorylation. Vascular Pharmacology 50, 147-152.
Iserin, P. (2001). Encyclopédie Encyclopedia of Médicinal Plants. La Rousse. 2nd Edition, pp. 244-
245.
Ismail, H.I., Chan, K.W., Mariod, A.A., Ismail, M. (2010). Phenolic content and antioxidant activity
of cantaloupe (cucumis melo) methanolic extracts. Food Chemistry 119, 643-647.
Itoh, T., Imano, M., Nishida, S. (2012). (2)-Epigallocatechin-3-gallate increases the number of neural
stem cells around the damaged area after rat traumatic brain injury. Journal of Neural Transmission
- Springer 119(8), 877-890.
Iwalewa, E.O., McGaw, L.J., Naidoo, V., Eloff, J.N. (2007). Inflammation: the foundation of diseases
and disorders. A review of phytomedicines of South African origin used to treat pain and
inflammatory conditions. African Journal of Biotechnology 6, 2868-2885.

Jaakola, L., Hohtola, A. (2010). Effect of latitude on flavonoid biosynthesis in plants. Plant, Cell &
Environment 33(8), 1239-1247.
Jahaniani, F., Ebrahimi, S. A., Rahbar-Roshandel, N., Mahmoudian, M. (2005). Xanthomicrol is the
main cytotoxic component of Dracocephalum kotschyii and a potential anti-cancer agent.
Phytochemistry 66, 1581-1592.
Jayaprakasha, G. K., Singh, R. P., Sakariah, K. K. (2001). Antioxidant activity of grape seed (Vitis
vinifera) extracts on peroxidation models in vitro. Food Chemistry 73, 285-290.
Kang, H.M., Saltveit, M.E. (2002). Antioxidant capacity of lettuce leaf tissue increases after
wounding. Journal of Agricultural and Food Chemistry 50, 7536-7541.
Kartal, M., Altun, M.L., Kurucu, S. (2003). HPLC method for the analysis of harmol, harmalol,
harmine and harmaline in the seeds of Peganum harmala. Journal of Pharmaceutical and
Biomedical Analysis 31, 263-269.
Kaur, C., Kapoor, H.C. (2002). Antioxidant activity and total phenolic content of some Asian
vegetables. International Journal of Food Science & Technology 37, 53-161.
Kelley, E. E., Kelley, N. K. H., Hundley, N. J., Malik, U. Z., Freeman, B. A., Tarpey, M. M. (2010).
Hydrogen peroxide is the major oxidant product of xanthine oxidase. Free Radical Biology and
Medicine 48, 493-498.
Khan, A. M., Qureshi, R. A., Ullah, F., Shinwari, Z.K., khan, J. (2012). Flavonoids distribution in
selected medicinal plants of margalla hills and surroundings. Pakistan Journal of Botany 44(4),
1241-1245.
Khan, O. Y. (1990). Studies in the chemical constituents of Peganum harmala L. PhD Thesis, H.E.J.
Research Institute of Chemistry University of Karachi, Paki
Khennouf, S., Amira, S., Arrar, L., Baghiani, A. (2010). Effect of Some Phenolic Compounds and
Quercus Tannins on Lipid Peroxidation. World Applied Sciences Journal 8 (9), 1144-1149.
Kholmatov, Kh., Kharlamov, I.A., Mavlankulova, Z.I. (1998). Medicinal plants of Central Asia (In
Russian). Med Lit Publ Ibn Sino. Tashkent 296.
Kirtikar, K.R., & Basu, B.D. (1995). Indian Medicinal Plants. Dehradune, India: International Book
Distributors.

Kitagawa, S., Sugiyama, Y., Sakuma, T. (1996). Inhibitory effects of catechol derivatives on
hydrophilic free radical initiator-induced hemolysis and their interaction with hemoglobin.
Chemical and Pharmaceutical Bulletin (Tokyo) 44, 881-4.
Klinenberg, J.R., Goldfinger, S.E., Seegmiller, J.E. (1965). The effectiveness of xanthine oxidase
inhibitor allopurinol in the treatment of gout. Annals of internal medicine 62, 639-647.
Kohen, R., Nyska, A. (2002). Oxidation of biological systems: oxida-tive stress phenomena,
antioxidants, redox reactions, and methods for their quantification. Toxicologic Pathology 30, 620-
630.
Koleva, I.I., van Beek, T.A., Linssen, J.P.H., de Groot, A., Evstatieva, L.N. (2002). Screening of plant
extracts for antioxidant activity: A comparative study on three testing methods. Phytochemical
Analysis 13(1), 8-17.
Kooij, A. (1994). A re-evaluation of the tissue distribution and physiology of xanthine oxidoreductase.
Histochemical Journal 26, 889-915.
Kooij, A., Bosch, K.S., Frederiks, W.M., and Van Noorden, C.J.F. (1992). High levels of xanthine
oxidoreductase in endothelial, epithelial and connective tissue cells in rat tissues. A relation
between localization and function. Virchows Archiv [B] Cell Pathology 62, 143-150.
Kostyuk, V.A., Potapovich, A.I., Suhan, T.O., De Luca, C., Korkina, L.G. (2011). Antioxidant and
signal modulation properties of plant polyphenols in controlling vascular inflammation. European
Journal of Pharmacology 658(2_3), 248-256.
Kouri, G., Tsimogiannis, D., Bardouki, H., Oreopoulou, V. (2007). Extraction and analysis of
antioxidant components from Origanum dictamnus. Innovative Food Science and Emerging
Technologies 8, 155-162.
Krishnamoorthy, M., Sasikumar, J.M., Shamna, R., Pandiarajan, C., Sofia, P., Nagarajan, B. (2011).
Antioxidant activities of bark extract from mangroves, Bruguiera cylindrica (L.) Blume and
Ceriops decandra Perr. Indian Journal of Pharmacology 43(5), 557-562.
Kuhn, M.A., Winston, D. (2000). Herbal therapy and supplement, a scientific and traditional approach.
lippincot, New York, pp. 347-350.
Kumazawa, S., Hamasaka, T., Nakayama, T. (2004). Antioxidant activity of propolis of various
geographic origins. Food Chemistry 84(3), 329–339.
Kunwar, A., Priyadarsini, K.I. (2011). Free radicals, oxidative stress and importance of antioxidants in
human health. Journal of Medical and Allied Sciences 2, 53-60.

Labuschagne, C. F., Brenkman, A. B. (2013). Current methods in quantifying ROS and oxidative
damage in Caenorhabditis elegans and other model organism of aging. Ageing Research Reviews
12, 918-930.
Laemmli, U.K. (1970). Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature 227, 680-685.
Laitonjam W. S. (2012). Natural Antioxidants (NAO) of Plants Acting as Scavengers of Free Radicals,
Chapter 9. In Studies in Natural Products Chemistry Vol. 37. (review)
Lamchouri, F., Settaf, A., Cherrah, Y., Zemzami, M., Lyoussi, B., Zaid, A., et al. (1999). Antitumour
principles from Peganum harmala seeds. Therapie 54, 753-758.
Landolfi, R., Mower, R.L., Steiner, M. (1984). Modification of platelet function and arachidonic acid
metabolism by bioflavonoids. Structure-activity relations. Biochemical Pharmacology 33, 1525-
1530.
Le, K., Chiu, F., Ng, K. (2007). Identification and quantification of antioxidants in Fructus lycii. Food
Chemistry 105, 353-363.
Leite, A. V., Malta, L. G., Riccio, M. F., Eberlin, M. N., Pastore, G. M., Marostica, M. R. (2011).
Antioxidant potential of rat plasma by administration of freeze-dried Jaboticaba peel (Myrciaria
jaboticaba Vell Berg). Journal of Agricultural and Food Chemistry 59(6), 2277-2283.
Lespade, L., Bercion, S. (2010). Theoretical study of the mechanism of inhibition of xanthine oxydase
by flavonoids and gallic acid derivatives. Journal of Physical Chemistry 114, 921-928.
Li, W., Hydamaka, A.W., Lowry, L., Beta, T. (2009). Comparison of antioxidant capacity and
phenolic compounds of berries, chokecherry and sea buckthorn. Central European Journal of
Biology
Li, H. B., Cheng, K. W., Wong, C. C., Fan, K. W., Chen, F. D., and Jiang, Y. S. (2007). Evaluation of
antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food
Chemistry 102, 771-776.
Lin, C.M., Chen, C.S., Chen, C.T., Liang, Y.C., and Lin, J.K. (2002). Molecular modeling of
flavonoids that inhibits xanthine oxidase. Biochemical and Biophysical Research Communications
294, 167-172.
Liu, F., Ng, T.B. (2000). Antioxidative and free radical scavenging activities of selected medicinal
herbs. Life Science 66(8), 725-735.

Liu, W., Wang, H., Yao, W., Gao, X., Yu, L. (2010). Effects of sulfation on the physicochemical and
function properties of a water-insoluble polysaccharide preparation from Ganoderma lucidum.
Journal of Agricultural and Food Chemistry 58, 3336-3341.
Lobo, V., Patil, A., Phatak, A., Chandra, N. (2010). Free radicals, antioxidants and functional foods:
Impact on human health. Pharmacognosy reviews 4(8), 118-126.
Lotito, S.B., Frei, B. (2006). Consumption of flavonoid-rich foods and increased plasma antioxidant
capacity in humans: cause, consequence, or epiphenomenon?. Free Radical Biology and Medicine
41, 1727-1746.
Lucy Chen, B.A., Judy, Y., Hu, M.D., Steven, Q., Wang, M.D. (2012). The role of antioxidants in
photoprotection: A critical review. Journal of the American Academy of Dermatology 67, 1013-
1024.
Macheix, J.J., Fleuriet, A., Jay-Allemand, C. (2005). Nature et diversité des composés phénoliques des
végétaux. In : Les composes phénoliques des végétaux, Presses polytechnichniques et universitaires
Fomandes: Italie, 1er
édition 1-33.
Machlin, L.J., Bendich, A. (1987). Free radical tissue damage: protective role of antioxidant nutrients.
The Journal of American Societies for Experimental Biology 1, 441-445.
Mahmoudian, M., Jalilpour, H., Salehian, P. (2002). Toxicity of Peganum harmala. Iranian Journal
of Pharmacology & Therapeutics 1, 1-4.
Manach, C., Scalbert, A., Morand, C., Remesy, C., Jimenez, L. (2004). Polyphenols: food sources and
bioavailability The American Journal of Clinical Nutrition 79, 727-747.
Manna, C., D’angelo, S., Migliardi, V., Loffredi, E., Mazzoni, O., Morrica, P., Galletti, P., Zappia, V.
(2002). Protective effect of the phenolic fraction from virgin olive oils against oxidative stress in
human cells. Journal of agricultural and food chemistry 50, 6521-6526.
Markham, K.R. (1982). Techniques of flavonoid identification. Academic press, London, Chap. 1 and
2, 1-113.
Martin, H.M., Moore, K.P., Bosmans, E., Davies, S., Burroughs, A.K., Dhillon, A.P., Tosh, D.,
Harrison, R. (2004). Xanthine oxidoreductase is present in bile ducts of normal and cirrhotic liver.
Free Radical Biology and Medicine 37, 1214-1223.
Masella, R., Di Benedetto, R., Vari, R., et al. (2005). Novel mechanisms of natural antioxidant
compounds in biological systems: involvement of glutathione and glutathione-related enzymes. The
Journal of Nutritional Biochemistry 16, 577-586.

Massey, V., Edmondson, D. (1970). On the mechanism of inactivation of xanthine oxidase by cyanide.
Journal of Biological Chemistry 245, 6595-6598.
Mather, I.H. (2000). A review and proposed nomenclature for major proteins of the milk-fat globule
membrane. Journal of Dairy Science 83, 203-247.
Matsumura, F., Yamaguchi, Y., Goto, M., Ichiguchi, O., Akizuki, E., Matsuda, T., Okabe, K., Liang,
J., Ohshiro, H., Iwamoto, T., Yamada, S., Mori, K., and Ogawa, M. (1998). Xanthine oxidase
inhibition attenuates kupffer cell production of neutrophil chemoattractant following ischemia-
reperfusion in rat liver. Hepatology 28, 1578.
Mazunder, U.K., Gupta, M., Rajeshwar, Y. (2005). Antihyperglycemic effect and antioxidant potential
of phyllanthus niruri (euphorbiaceae) in streptozotocin induced diabetic rats. European Bulletin of
Drug Research 13(1), 13-23.
Mc Cord, J. (1993). Human disease, free radicals and the oxidant/antioxidant balance. Clinical
Biochemistry 26-32.
Mc Cord, J.M. (1985). Oxygen-derived free radicals in post ischemic tissue injury. New England
Journal of Medicine 312, 159-163.
McCord, J. M., and Fridovich, I. (1969). Superoxide dismutase: An enzymic function for
erythrocuprein (hemocaprein). Journal of Biological Chemistry 244, 6049-6055.
Mendes, L., de Freitas, V., Baptista, P., Carvalho, M. (2011). Comparative antihemolytic and radical
scavenging activities of strawberry tree (Arbutus unedo L.) leaf and fruit. Food and Chemical
Toxicology doi:10.1016/j.fct.2011.06.028.
Meneshian, A.V.E.D., Bulkley, G.B. (2002). The physiology of endothelial xanthine oxidase: from
urate catabolism to reperfusion injury to inflammatory signal transduction. Microcirculation 9,
161-175.
Milde, J., Elsttner, E.F., Grabmann, J. (2004). Synergistic inhibition of Low-density lipoprotein
oxidation by rutin, terpinene, and ascorbic acid. Phytomedicine 11, 105-113.
Mitchell, P. (1961). Coupling of phosphorylation to electron and hydrogen transfer by a chemi-
osmotic type of mechanism. Nature 191, 144-148.
Moghadam, M. S., Maleki, S., Darabpour, E., Motamedi, H., Nejad, S. M. S. (2010). Antibacterial
activity of eight Iranian plant extracts against methicillin and cefixime restistant Staphylococcous
aureus strains. Asian Pacific Journal of Tropical Medicine 3, 262-265.

Moorhouse, P.C., Grootvelt, M., Halliwell, B., Quinlan, J.G., and Gutteridge, M.C.J. (1987).
Allopurinol and oxypurinol are hydroxyl radical scavengers. European Journal of Biochemistry
Letters 213, 23-28.
Morin D., Thiery H., Spedding M., Tillement J., P. (2001). Mitochondria as target for antiischemic
drugs. Journal of Physiology and Pharmacology 13, 22-67.
Moura, D. J., Richter, M. F., Boeira, J. M., Pegas Henriques, J. A., & Saffi, J. (2007). Antioxidant
properties of betacarboline alkaloids are related to their antimutagenic and antigenotoxic activities.
Mutagenesis 22, 293-302.
Moure, A., Cruz, J. M., Franco, D., Dominguez, J. M., Sineiro, J., Dominguez, H., Nunez, M. J.,
Parajo, J. C.(2001). Natural antioxidants from residual sources. Food Chemistry 72, 145-171.
Moussard, C. (2006). Radicaux libres oxygénés et antioxydants. In: Biochimie structural et
métabolique, De boeck: Bruxelles, 3eme
édition, pp. 321-335.
Munir, C., Zaidi, M.I., Ahmad, N., Attaur, R. (1995). An Easy Rapid Metal Mediated Method of
Isolation of Harmine and Harmaline from Peganum harmala. Fitoterapia 1,73.
Nakamura, M., Yamazaki, I. (1982). Preparation of bovine milk xanthine oxidase as dehydrogenase
form. The Journal of Biochemistry 92, 1279-1286.
Nenaah, G. (2010). Antibacterial and antifungal activities of (beta)-carboline alkaloids of Peganum
harmala (L) seeds and their combination effects. Fitoterapia 81(7), 779-782.
Ng, T.B., Li, W.W., Yeung, H.W. (1986). A steryl glycoside fraction with hemolytic activity from
tubers of Momordica cochinchinensis. Journal of Ethnopharmacology 18 (1), 55-61.
Nishino, T., Nishino, T. (1997). The conversion from the dehydrogenase type to the oxidase type of rat
liver xanthine dehydrogenase by modification of cysteine residues with fluorodinitrobenzene. The
Journal of Biological Chemistry 272, 29859-29864.
Nishino, T., Okamoto, K., Eger, B.T., Pai, E.F., Nishino, T. (2008). Mammalian xanthine
oxidoreductase mechanism of transition from xanthine dehydrogenase to xanthine oxidase.
Federation of the Societies of Biochemistry and Molecular Biology Journal 275, 3278-3289.
Nishino, T., Okamoto, K., Kawaguchi, Y., Hori, H., Matsumura, T., Eger, B.T., Pai, E.F., Nishino, T.
(2005). Mechanism of the conversion of xanthine dehydrogenase to xanthine oxidase: identification
of the two cysteine disulfide bonds and crystal structure of a non-convertibal rat liver xanthine
dehydrogenase mutant. The Journal of Biological Chemistry 280, 24888-24894.

Ohkawa, H., Ohishi, N., Yagi, K. (1979). Assay of lipid peroxides in animal tissue by thiobarbituric
reaction. Analytical Biochemistry 95(2), 351-358.
Okamoto, K., Eger, B. T., Nishino, T., Kondo, S., Pai, E. F., Nishino, T. (2003). An extremely potent
inhibitor of xanthine oxidoreductase. Journal of Biological Chemistry 278, 1848-1855.
Olson, J. S., Ballou, D. P., Palmer, G., Massey, V. (1974). The mechanism of action of xanthine
oxidase. Journal of Biological Chemistry 249, 4363-4382.
Ong, K.C., Khoo, H.E. (2000). Effects of myricetin on glycemia and glycogen metabolism in diabetic
rats. Life Science 67, 1695-1705.
Ozen, T., Demirtas, I., Aksit, H. (2011). Determination of antioxidant activities of various extracts and
essential oil compositions of Thymus praecox subsp. skorpilii var. skorpilii. Food Chemistry 124,
58-64.
Ozsoy, N., Can, A., Yanardag, R., Akev, N. (2008). Antioxidant activity of Smilax excelsa L. leaf
extracts. Food Chemistry 110, 571-583.
Ozturk, F., Gul, M., Ates, B., Ozturk, I.C., Cetin, A., Vardi, N., Otlu, A., 2009. Protective effect of
apricot (Prunus armeniaca L.) on hepatic steatosis and damage induced by carbon tetrachloride in
Wistar rats. British Journal of Nutrition 102, 1767-1775.
Pacher, P., Nivorozhkin, A., Szabó, C. (2006). Therapeutic effects of xanthine oxidase inhibitors:
renaissance half a century after the discovery of Allopurinol. Pharmacological Reviews 58, 87-
114.
Pari ,L., Latha, M.. (2005). Antidiabetic effect of Scoparia dulcis: effect on lipid peroxidation
peroxidation in streptozotocin diabetes. General Physiology and Biophysics 24, 13-26.
Parks, D. A., Granger, D. N. (1986). Xanthine oxidase: Biochemistry, distribution and physiology.
Acta Physiologica Scandinavica 548, 87-99.
Parks, D.A., Williams, T.K., Beckman, J.S. (1988). Conversion of xanthine dehydrogenase to oxidase
in ischemic rat intestine: a reevaluation. American Journal of Physiology 254, 768-774.
Pavlou, P., Rallis, M., Deliconstantinos, G., Papaioannou, G., Grando, S. A. (2009). In-vivo data on
the influence of tobacco smoke and UV light on murine skin. Toxicology and Industrial Health 25,
231-239.
Perez, H. D., Weksler, B. B., Goldstein, I. M. (1990). Generation of chemotactic lipid from
arachidonic acid by exposure to a superoxide generation system. Inflammation 4, 313-328.

Perron, N. R., Brumaghim, J. L. (2009). A review of the antioxidant mechanisms of polyphenol
compounds related to iron binding. Cell Biochemistry and Biophysics 53, 75-100.
Pham-Huy, Lien, Ai., He, Hua., Pham-Huy, Chuong. (2008). Free Radicals, Antioxidants in Disease
and Health. International Journal of Biomedical Science 4(2), 89-96.
Pietta, P.G. (2000). Flavonoids as antioxidants. Journal of Natural Products 63, 1035-42.
Pincemail, J, Meurisse, M, Limet, R., Defraigne, J. O. (1999). L’évaluation du stress oxydatif d’un
individu: une réalité pour le médecin. Vaisseaux, Coeur, Poumons 4 (5). (review)
Pincemail, J., Bonjean, K., Cayeux, K., Defraigne, J.O. (2002). Physiological action of antioxidant
defences. Nutrition Clinique et Métabolisme 3 (16), 233-239.
Podsȩdek, A. (2007). Natural antioxidants and antioxidant capacity of Brassica vegetables. A review.
Food Science and Technology 40, 1-11.
Prashanth, D., John, S. (1999). Antibacterial activity of Peganum harmala. Fitoterapia 70, 438-439.
Pulpati, H., Biradar, Y. S., Rajani, M. (2008). High-performance thin-layer chromatography
densitometric method for the quantification of harmine, harmaline, vasicine, and vasicinone in
Peganum harmala. Journal of AOAC International 91(5), 1179-1185.
Rajasekaran, S., Sivagnanam, K ., Subramanian, S. (2005). Antioxidant effect of Aloe vera gel extract
instreptozotocin-induced diabetes in rats Pharmacological. Reports 57, 90-96.
Ramarathnam, N., Osawa, T., Ochi, H., Kawakishi, S. (1995). The contribution of plant food
antioxidants to human health. Trends in Food Sciences and Technology 6, 75.
Rastelli, G., Costantino, L., and Albasini, A. (1997). A model of the interaction of substrates and
inhibitors with xanthine oxidase. Journal of the American Chemical Society 119, 3007-3016.
Re, R., Pellegrini, N., Proteggente, A., Annala, A., Yang, M., Rice-Evans, C. (1999). Antioxidant
activity applying an improved ABTS radical cation decolourisation assay. Free Radical Biology
and Medicine 26, 1231-1237.
Read G., Randat R. (1988). Structure and properties of thiobarbituric acid malonaldehyde chromogen.
Journal of The Chemical Society 1103-1105.
Read, M. A. (1995). Flavonoids: naturally occurring anti-inflammatory agents Vascular. American
Journal of Pathology 147(2), 235-237.

Record, I. R., Dreosti, I. E., McInerney, J. K. (2001). Changes in plasma antioxidant status following
consumption of diets high or low in fruit and vegetables or following dietary supplementation with
an antioxidant mixture. British Journal of Nutrition 85(4), 459-464.
Ricciarelli, R., Zingg, J.M., Azizi, A. (2001). Vitamin E: protective role of a Janus molecule. FASEB.
Journal 15, 2314-2325.
Rice-Evans, C.A., Miller, N.J., Bolwell, P.G., Bramley, P.M., Pridham, J.B. (1995). The relative
antioxidant activities of plant derived polyphenolic flavonoids. Free Radical Research. 22, 375-
383.
Rop, O., Mlcek, J., Jurikova, T., Valsikova, M., Sochor, J., Reznicek, V., Kramarova, D. (2010).
Phenolic content, antioxidant capacity, radical oxygen species scavenging and lipid peroxidation
inhibiting activities of extracts of five black chokeberry (Aronia melanocarpa (Michx.) Elliot)
cultivars. Journal of Medicinal Plants Research 4(22), 2431–7.
Rott, K. T., Agudelo, C. A. (2003). Gout. Journal of the American Medical Association 289, 2857-
2860.
Rundles, R.W., Metz, E.N., Sliberman, H.R. (1966). Allopurinol in the treatment of gout. Annals of
Internal Medicine 64, 229-258.
Sahreen, S., Khan, M. R., Khan, R. A. (2010). Evaluation of antioxidant activities of various solvent
extracts of Carissa opaca fruits. Food Chemistry 122, 1205-1211.
Sala, A., Recio, M.C., Giner, R.M., Mane, S., Rios, J.L. (2000). Anti-phospholipase A2 and anti-
inflammatory activity of Santolina chamaecyparissus. Life Science 66, 35-40.
Sanchez-Moreno, C. (2002). Review: methods used to evaluate the free radical scavenging activity in
foods and biological systems. Food Science and Technology International 8, 121-137.
Sanders, S., Eisenthal, R.S., and Harrison, R. (1997). NADH oxidase activity of human xanthine
oxidoreductase – reneration of superoxyde anion. European Journal of Biochemistry 245, 541-
548.
Sandhar, H. K., Kumar, B., Prasher S., Tiwari, P., Salhan, M., Sharma, P. (2011). A Review of
Phytochemistry and Pharmacology of Flavonoids. Internationale Pharmaceutica Sciencia 1(1), 25-
41.
Sanz, J. F., Garcia-Sarrion, A., Marco, J. A. (1991). Germacrane derivatives from Santolina
chamaecyparissus. Phytochemistry 30, 3339-3342.

Sardina, J. L., López-Ruano, G., Sánchez-Sánchez, B., Lianillo, L., Hernández-Hernández, A. (2012).
Reactive oxygen species: Are they important for haematopoiesis?. Critical Reviews in
Oncology/Hematology 81, 257–274.
Sarikurkcu, C. Tepe, B. Daferera, D. Polissiou, M. Harmandar, M. (2008). Studies on the antioxidant
activity of the essential oil and methanol extract of Marrubium globosum subsp. Globosum
(lamiaceae) by three different chemical assays. Bioresource Technology 99, 4239-4246.
Sathishsekar, D., Subramanian, S. (2005). Antioxidant properties of Momordica Charantia (bitter
gourd) seeds on Streptozotocin induced diabetic rats. Asia Pacific Journal of Clinical Nutrition
14(2), 153-158.
Sezik, E., Yesilada, E., Shadidoyatov, H., Kuliev, Z., Nigmatullaev, A.M., Aripov, H.N. (2004). Folk
Medicine in Uzbekistan. I. Toshkent, Djizzah and Samarqand provinces. Journal of
Ethnopharmacol 92, 197-207.
Sharaf, M., el-Ansari, M.A., Matlin, S.A., Saleh, N.A. (1997). Four flavonoid glycosides from
Peganum harmala. Phytochemistry 44, 533-536.
Shinde, A., Ganu, J., Naik, P. (2012). Effect of free radicals & antioxidants on oxidative stress: A
review. Journal of Dentistry and Allied Science 2, 63-66.
Shyu, Y.S., Lin, J.T., Chang, Y.T., Chiang, C.J., Yang, D.J. (2009). Evaluation of antioxidant ability
of ethanolic extract from dill (Anethum graveolens L.) flower. Food Chemistry 115, 515–521.
Sies, H. Oxidative stress: from basic research to clinical application. (1991). American Journal of
Medicine 91, 31-38.
Silliman, K., Parry, J., Kirk, L. L., & Prior, R. L. (2003). Pycnogenol does not impact the antioxidant
or vitamin C status of healthy young adults. Journal of the American Dietetic Association 103(1),
67-72.
Silva, A. M. O., Vidal-Novoa, A., Batista-Gonzlez, A. E., Pinto, J. R., Mancini, D. A. P., Reina-
Urquijo, W., et al. (2012). In vivo and in vitro antioxidant activity and hepatoprotective properties
of polyphenols from Halimeda opuntia (Linnaeus) Lamouroux. Redox Report 17, 47-53.
Simoes, C. M.O., Amoros, M., and Girre, L. 1999. Mechanism of antiviral activity of triterpenoid
saponins. Phytotherapy Research 13, 323-328.
Sorg, O. (2004). Oxidative stress: a theoretical model or a biological reality. Comptes Rendus
Biologies 327, 649-662.

Sowndhararajan, K., Joseph, J.M., Rajendrakumaran D. (2012). In vitro xanthine oxidase inhibitory
activity of methanol extracts of Erythrina indica Lam. leaves and stem bark. Asian Pacific Journal
of Tropical Biomedicine S1415-S1417.
Stevens, C.R., Miller, T.M., Clinch, J.G., Kanczler, J.M., Bodamyali, T., Blake, D.R. (2000).
Antibacterial properties of xanthine oxidase in human milk. Lancet 356, 829-830.
Stewart, R. C., Hille, R., Massey, V. (1984) Characterization of arsenite-complexed xanthine oxidase
at room temperature. Spectral properties and pH-dependant redox behaviour of the molybdenum-
arsenite center. Journal of Biological Chemistry 259, 14426-14436.
Sudha, G., sangeetha priya, M., indhu shree, R., vadivukkarasi, S. (2011). in vitro free radical
scavenging activity of raw pepino fruit (solanum muricatum aiton). International Journal of
Current Pharmaceutical Research 3(2), 0975-7066.
Suresh, B., Srirama, S., Dhanaraja, S.A., Elangoa, K., Chinnaswamy, K. (1997). "Anticandidal activity
of Santolina chamaecyparissus volatile oil." Journal of Ethnopharmacology 55(2), 151-159.
Takebayashi, J., Chen, J., Tai, A. (2010). A Method for Evaluation of Antioxidant Activity Based on
Inhibition of Free Radical-Induced Erythrocyte Hemolysis. D. Armstrong (ed.), Advanced
Protocols in Oxidative Stress II, Methods in Molecular Biology, vol 594, 287-296. (review)
Thannickal, V.J., Fanburg, B.L. (2000). Reactive oxygen species in cell signaling. The American
Journal of Physiology - Lung Cellular and Molecular Physiology 279, 1005-1028.
Tsao, R. Chemistry and biochemistry of dietary polyphenols (2010). Nutrients 2, 1231-1246.
Tsimogiannis, D.I., Oreopoulou, V. (2006). The contribution of flavonoid C-ring on the DPPH free
radical scavenging efficiency. A kinetic approach for the 3’,4’-hydroxy substituted members.
Innovative Food Science and Emerging Technologies 7, 140-146.
Tuberoso C I G, Montoro P, PiacenteS, Corona G, Deiana M, Dessi M A, Pizza C and Cabras P
(2009). Flavonoid characterization and antioxidant activity of hydroalcoholic extracts from
Achillea ligustica All. Journal of Pharmaceutical and Biomedical Analysis 50, 440-448.
Tulipani, S., Armeni, T., Giampieri, F., Alvarez-Suarez, J.M., Gonzalez-Paramas, A.M., Santos-Buelga, C.,
Busco, F., Principato, G., Bompadre, S., Quiles, J.L., Mezzetti, B., Battino, M. (2014). Strawberry
intake increases blood fluid, erythrocyte and mononuclear cell defenses against oxidative
challenge. Food Chemistry 156, 87-93.

Tunalier, Z., Koşar, M., Küpeli, E., Caliş, I., Başer, K.H. (2006). Antioxidant, anti-inflammatory,
anti-nociceptive activities and composition of Lythrum salicaria L. extracts. Journal of
Ethnopharmacology 4;110(3), 539-547.
Tung, Y.-T., Wu, J.-H., Huang, C.-Y., Kuo, Y.-H., and Chang, S.-T. (2009). Antioxidant activities and
phytochemical characteristics of extracts from Acacia confusa bark. Bioresource Technology 100,
509-514.
Umamaheswari, M., Asokkumar, K., Sivashanmungam, A. T., Remyarayu, A., Subhadredevi,
V., Ravi, T. K. (2009). In vitro xanthine oxidase inhibitory activity of the fraction of
Erythrina stricta Roxb. Journal of Ethnopharmacology 124, 646-648.
Unverzagt, J. (2013). African rue. In: Field guide wyoming weed watchlist, edited by: Steven L.
miller, university of wyoming extension: USA 47-49.
Urban, M.K., Zainol, A., Abd-Hamid, S., Yusof, R. Muse. (2003). Antioxidative activity and total
phenolic compounds of leaf, root and petiole of four accessions of Centella asiatica (L.). Food
Chemistry 81, 575-581.
Valente, M.J., Baltazar, A.F., Henrique, R., Estevinho, L., Carvalho, M., (2011). Biological
activities of Portuguese propolis: protection against free radical-induced erythrocyte
damage and inhibition of human renal cancer cell growth in vitro. Food and Chemical
Toxicology 49, 86–92.
Valko, M., Dieter, L., Jan, M., Mark, T.D., Cronin, Milan, Mazur, Joshua Telser. (2007). Free radicals
and antioxidants in normal physiological functions and human disease. The International Journal
of Biochemistry & Cell Biology 39, 44-84.
Valko, M., Rhodes, C.J., Moncol, J., Izakovic, M., Mazur, M. (2006). Free radicals, metals and
antioxidants in oxidative stress-induced cancer. Chemico-Biological Interactions 160(1), 1-40.
Van Acker, S. A. B. E., van Balen, G. P., van den Berg, D. J., Bast, A., van der Vijgh, S. A. B. E.
(1998). Influence of iron chelation on the antioxidant activity of flavonoids. Biochemical
Pharmacology 56(8), 935-943.
Van Acker, S.A.B.E., van den Berg, D.J., Tromp, M.N.J.L., Griffioen, D.H., van Bennekom, W.P.,
van der Vijgh, W.J.F., Bast, A. (1996). Structural aspect of antioxidant activity of flavonoids. Free
Radical Biology and Medicine 20, 331-342.
Van Hoorn, D.E.C., Nijveldt, R.J., Van Leeuwen, P.A.M., Hofman, Z., M’Rabet, L., De Bont,
D.B.A., and Van Norren, K. (2002). European Journal of Pharmaceutical 451, 111-118.
http://www.ncbi.nlm.nih.gov/pubmed?term=Ko%C5%9Far%20M%5BAuthor%5D&cauthor=true&cauthor_uid=17125946

Vorbach, C., Harrison, R., Capecchi, M. (2003). Xanthine oxidoreductase is central to the evolution
and function of the innate immune system. Trends in Immunology 24, 512-517.
Wang, D., Wang, L., Zhu, F., Zhu, J., Chen, X. D. L. Z., Saito, M., Li, L. (2008). In vitro and in vivo
studies on the antioxidant activities of the aqueous extracts of Douchi (a traditional Chinese salt-
fermented soybean food). Food Chemistry 107, 1421-1428
Wardman, P., Candeias Fenton, L.P. (1996). Chemistry: an introduction. Radiation Res 145, 523-531.
Watanabe, T., Misawa, S., Hiradate, S., Osaki, M. (2008). Characterization of root mucilage from
Melastoma malabathricum, with emphasis on its roles in aluminum accumulation. New Phytologist
178, 581-589.
Widlansky, M. E., Duffy, S. J., Hamburg, N. M., Gokce, N., Warden, B. A., Wiseman, S., et al.
(2005). Effects of black tea consumption on plasma catechins and markers of oxidative stress and
inflammation in patients with coronary artery disease. Free Radical Biology & Medicine 38(4),
499-506.
Williams, R.J., Spencer, J.P.E., Rice Evans, C. (2004). Flavonoids: antioxidants or signaling
molecules?. Free Radical Biology and Medicine 7, 838–849.
Wong, S.P., Leong, L.P., William Koh, J.H. (2006). Antioxidant activities of aqueous extracts of
selected plants. Food Chemistry 99, 775-783.
Wu, N., Zu, Y., Fu, Y., Kong, Y., Zhao, J., Li, X., Li, J., Wink, M., Efferth, T.(2010). Antioxidant
activities and xanthine oxidase inhibitory effects of extracts and main polyphenolic compounds
obtained from Geranium sibiricum L. Journal of Agricultural and Food Chemistry 58, 4737-4743.
Xia, M., Dempski, R., and Hille, R. (1999). The reductive half-reaction of xanthine
oxidasereactionwith aldehyde substrats and identification of the catalytically labile oxygen. Journal
of Biological Chemistry 274, 3323-3330.
Yalçın, D., Bayraktar, O. (2009). Screening of Some Plant-Derived Alkaloids and Phenolics for
Catechol-OMethyltransferase Inhibition. International Journal of Natural and Engineering
Sciences 3 (2), 23-27.
Young, J.L., Boswell, R.B., Nies, A.S. (1974). Severe allopurinol hypersensitivity. association with
Thiazides and Prior Renal Compromise. Archives of internal medicine 134, 553-559.
Zabri, H., Kodjo, C., Beni, A., Marnyrbekova, B., Ko, J., Bekro, Y.A. (2008). Phytochemical
screening and determination of flavonoids in Secamone afzeli (Asclepiadaceae) extracts. African
journal of Pure and Applied Chemistry 2(8), 80-82.

Zhang, A., Zhu, Q.Y., Luk, Y.S., Ho, K.Y., Fung, K.P., Chen, Z.Y. (1997). Inhibitory effects of
jasmine green tea epicatechin isomers free radical induced lysis of red blood cells. Life Science
61(4), 383-394.
Zhao, H., Fan W., Dong J., Lu J., Chen J., Shan L., Lin Y., Kong W. (2008). Evaluation of antioxidant
activities and total phenolic contents of typical malting barley varieties. Food Chemistry 107, 296-
304.
Zheng, W., Wang, S.Y. (2001). Antioxidant activity and phenolic compounds in selected. herbs.
Journal of Agricultural and Food Chemistry 49(11), 5165-5170.
Zhong, W., Yan, T., Lim, R., Oberley, L.W., 1999. Expression of superoxide dismutases, catalase, and
glutathione peroxidase in glioma cells. Free Radical Biology & Medicine 27, 1334-1345.
Zhou, F. G., Chen, Y., Liu, S., Yao, X., Wang, Y. (2013). In vitro and in vivo hepatoprotective and
antioxidant activity of ethanolic extract from Meconopsis integrifolia (Maxim.) Franch. Journal of
Ethnopharmacology 148, 664-670.
Zhu, J.X., Wang, Y., Kong, L.D., Yang, C., and Zhang, X. (2004). Effects of Biota orientalis extract
and its flavonoid constituents, quercetin and rutin on serum uric acid levels in oxonate-induced
mice and xanthine dehydrogenase and xanthine oxidase activities in mouse liver. Journal of
Ethnopharmacology 93, 133-140.
Zou, Y.P., Lu, Y.H., Wei, D.Z. (2004). Antioxidant activity of a flavonoid-rich extract of Hypericum
perforatum L. in vitro. Journal of Agricultural and Food Chemistry 52, 5032-5039.

املستعمالن يف Santolina chamaecyparisusواجلعدة Peganum harmalaملستخلصات نبتيت احلرمل هتدف هذه الدراسة إىل تقدير النشاطية املضادة لألكسدة: ملـــخصحيتوي على أعلى كمية من املركبات ((EAE-Scتقدير الكمي لعديدات الفينول والفالفونيدات أن مستخلص إثيل أسيتات لنبات اجلعدة بينت نتائج ال. الطب الشعيب باجلزائر
مت .ة، على التوايلغ من الوزن اجلاف للنبت/مكافئ كرستنيمغ ± 61.51 7.86غ من الوزن اجلاف و/محض الغاليك ئمغ مكاف373.83 ± 0.23 الفينولية والفالفونيدات قدرت بـأعلى نشاطية إزاحية للجذور احلرة EAE-Scحيث أظهر مستخلص ABTSو DPPH، باستعمال إختبار إزاحة اجلذور احلرة (in vitro)خمربيا تقدير النشاطية املضادة لألكسدة
(EAE- Ph)، حيث أظهرت النتائج أن مستخلص إثيل األسيتات لنبات احلرمل FTCو β-caroteneيف اإلختبارين، باإلضافة إىل اختبار تثبيط فوق أكسدة الليبيدات باستعمال . (CE-Sc)فقد ظهرت أعلى نسبة تثبيط باملستخلص اخلام لنبات اجلعدة FTCأما فيما خيص إختبار .β –caroteneبالنسبة إلختبار (% 94.21±0.003 )أعلى قدرة ميلك
أن املستخلص Ferrozineاختبار استخالب املعادن باستعمال بني. أقل امتصاصية EAE-Phليبيدية، وقد سجل مستخلص كمؤشرا لفوق األكسدة ال MDAاستعمل تقدير O2ر وإزاحة جذ (XO)بينت نتائج إختبار قدرة املستخلصات على تثبيط اإلنزم املؤكسد للكزانثني .)مل/مغ IC50 =0.32 ± 0.009)ميلك أعلى قدرة CE-Phاخلام لنبات احلرمل
•- O2يف حني سجلت أعلى قدرة إزاحية جلذر (.EAE-Scو CHE-Sc)الناتج عنه أن أعلى نشاطية كانت للمستخلص الكلوروفورمي وإثيل األسيتات لنبات اجلعدة
للمستخلص -• .Sو P. harmala ((MOHE-Phخلص امليثانويل لكل من للمست (in vivo)متت دراسة النشاطية املضادة لألكسدة يف الكائن احلي (.CHE-Ph)الكلوروفورمي لنبات احلرمل
chamaecyparisus (MOHE-Sc) النشاطية الكلية للبالزما عن طريق (APC : antioxidant plasma capacity) باستعمال إختبارDPPH ماعدا القدرة . القدرة اإلرجاعيةويف نسيج GSHو MDAومستويات CAT))الكتاالزمن جهة أخرى، أظهرت نتائج تقدير نشاطية إنزم .، مل تبد املستخلصات تأثريا معنوياMOHE-Scاإلرجاعية للمستخلص
59,21بنسبة GSHكما رفعت من مستوى . ، على التوايل% 35و % 59,16بنسبة MOHE-Phو MOHE-Scعند املعاملة بكل من CATالكبد ارتفاعا معنويا يف نشاطية inدراسة قدرة املستخلصات على تثبيط االحنالل الدموي وبينت .، على التوايل% 63.48و % 79.56بنسبة MDAخفضت من مستوى كما . على التوايل، % 50,44و %
vivo يف وجود املؤكسدt-BH أن ،MOHE-Ph قريبا من املستخلص ميلك تأثريا تثبيطياMOHE-Sc 78,51 ± 11,32 = HT50) ينت ب .(على التوايل، دقيقة 4,95 ± 72,05وميلكان القدرة على محاية نسيج الكبد من اإلجهاد التأكسدي وهذا التأثري يعود أساسا إىل ميزاهتما S. chamaecyparisusو P. harmala أن كل من نبات النتائج احملصل عليها
. اإلستعمال الطيب الشعيب الواسع هلما يفسراملضادة لألكسدة، وهذا ما
Abstract :The aim of this study is to evaluate the antioxidant activity of the extracts from Peganum harmala and Santolina chamaecyparisus
which are largely used in traditional medicine in Algeria. The estimation of polyphenols and flavonoids showed that the Ethyl Acetate extract
of S. chamaecyparisus (EAE-Sc) contains a high amount of phenolic compounds and flavonoids: 373.83 ± 0.23 mg gallic acid equivalent and 7.86 ± 61.51 mg quercetin equivalent/g of dried weigh, respectively. The DPPH and ABTS tests were used to evaluate the antioxidant
activity in vitro. The EAE-Sc showed the highest scavenging activity of free radicals in both tests. The inhibition of lipid peroxydation test
using FTC and β-carotene showed that the ethyl acetate extract of P .harmala (EAE-Ph) had the highest rate (94,21 ± 0.003 %) in β-carotene test, but for the FTC test the highest inhibition rate was noticed with crud extract of S. chamaecyparisus (CE-Sc). The MDA evaluation was
used as indicator of lipid peroxydation. The low absorption was found in EAE-Ph level. The capacity of the extracts to chelate the mineral
ions was tested using Ferrozine. The crud extract of P. harmala (CE-Ph) showed a high capacity to chelate ferrous ions (IC50: 0.32 ± 0.009 mg/ml). The extracts were studied for their ability of inhibition effect on XO and O2
•- scavenging activity. The results showed that the
chloroform extract of S. chamaecyparisus (CHE-Sc) and EAE-Sc have the highest inhibiting activity of XO, but the chloroform extract of P. harmala (CHE-Ph) showed the greatest scavenging effect on O2
•- with an IC50: 0.037 ± 0.0053 mg/ml. The antioxidant activity of methanol
extract of P. harmala (MOHE-Ph) and S. chamaecyparisus (MOHE-Sc) in vivo was estimated using the antioxidant plasma capacity (APC),
DPPH and reducing power tests. Except for the reducing power of MOHE-Sc, all extracts did not show any significant change. The activity of catalase (CAT), the level of MDA and GSH were evaluated in mouse liver homogenate. The results showed a high significant increase in
CAT activity with MOHE-Sc and MOHE-Ph, by 59.16 % and 35 %, respectively, and olso in the GSH levels by 59.21 % and 50.44 %,
respectively. However they decreased the level of MDA by 79.56 % and 63.48 %, respectively. The Evaluation of the plant extracts capacity to inhibit the hemolysis of blood in vivo in the presence of the oxidant (tBH), showed that MOHE-Ph has an inhibition effect comparable
with MOHE-Sc (78.51 ± 11.32 min and 72.05 ± 4.95 min, respectively). The present study showed that the P. harmala and S.
chamaecyparisus have the capacity to protect liver tissue from oxidative stress and this effect is probably due to their antioxidant capacity, which explain the large use of these plants in traditional medicine.
Resumé :L’objectif de la présente étude est d’estimer l’activité antioxydante des extraits de deux plantes, Peganum harmala et Santolina chamaecyparisus largement utilisées en médecine traditionnelle en Algérie. L’extrait acétate d’éthyle de S. chamaecyparisus (EAE-Sc)
renferme une grande quantité de composés phénoliques et de flavonoïdes avec des teneurs de 373.83 ± 0.23 mg d’équivalent acide gallique
et 61.51 ± 7.86 mg d’équivalent quercetin/g de poids sec de la plante, respectivement. L’activité antioxydante a été évaluée in vitro par l’utilisation des tests de DPPH et ABTS. L’extrait EAE-Sc a montré l’effet le plus élevé dans les deux tests. L’examen de l’activité anti-
péroxidation lipidique évaluée par les tests β-carotène et FTC, a montré que l’extrait acétate d’éthyle de P. harmala (EAE-Ph) présente
l’effet inhibiteur le plus puissant dans le test β-carotène, par contre dans le test FTC, l’effet de l’extrait brut de S. chamaecyparisus (CE-Sc) est le plus élevé. L’évaluation du MDA utilisée comme indicateur de la peroxidation lipidique a montré la faible absorbance a été enregistrée
au niveau de EAE-Ph, La capacité des extraits de capter les ions minéraux testée par l’utisation de ferrozine a indiqué que l’extrait brut de P.
harmala (CE-Ph) présente l’activité chélatrice la plus puissante (IC50: 0.32±0.009 mg/ml). D’autre part, l’évaluation de la capacité des extraits d’inhiber la xanthine oxydase (XO) et d’éliminer le radical O2
•- qu’elle produit, a montré que l’extrait chloroformique du S.
chamaecyparisus (CHE-Sc) et EAE-Sc sont plus actifs. Cependant, la plus grande capacité d’éliminer le radical O2•- est observée avec
l’extrait chloroformique de P. harmala (CHE-Ph). L’activité antioxydante, in vitro, estimée par la capacité antioxidante plasmatique (CAP), le test DPPH et le test du pouvoir réducteur a montré que seul le MOHE-Sc possède un effet réducteur. L’activité de la catalase (CAT) et les
taux du MDA et du GSH ont été évalués dans l’homogénat du foie de souris. Les résultats montrent une augmentation significative dans
l’activité de la CAT par MOHE-Sc et MOHE-Ph (59.16 % et 35 %, respectivement), en plus d’une augmentation du taux du GSH de 59.21 et 50.44%, respectivement et d’une diminution du taux du MDA de 79.56 % et63.48 %, respectivement. L’évaluation de la capacité des extraits
d’inhiber l’hémolyse des globules rouges in vivo en présence d’un oxydant tBH, a montré que le MOHE-Ph possède un effet inhibiteur de
l’hémolyse similaire à celui du MOHE-Sc (78,51 ± 11,32 et 72,05 ± 4,95 min, respectivement). La présente étude montre que les plantes Peganum harmala et Santolina chamaecyparisus possèdent une capacité de protéger le foie contre le stress oxydatif et que cet effet est
probablement dû à leurs capacités antioxydantes. Ceci explique leur grande utilisation en médecine traditionnelle.




















