
Melatonin prevents the development of hyperplastic urothelium induced by repeated doses of...
10
ORIGINAL ARTICLE Melatonin prevents the development of hyperplastic urothelium induced by repeated doses of cyclophosphamide Daša Zupančič & Gaj Vidmar & Kristijan Jezernik Received: 16 October 2008 / Revised: 14 February 2009 / Accepted: 19 March 2009 / Published online: 21 April 2009 # Springer-Verlag 2009 Abstract Repeated cyclophosphamide (CP) chemotherapy increases the risk of developing bladder cancer, which could be due to the extremely rapid proliferation of urothelial cells observed in hyperplastic urothelium induced by CP treatment. We investigated the effect of melatonin on the development of urothelial hyperplasia induced by repeated CP treatment. Male ICR mice were injected with CP (150 mg/kg) or melatonin (10 mg/kg) with CP once a week for 3, 4 and 5 weeks. Transmission and scanning electron microscopy, immunohistochemistry and Western blot analysis were used to study the ultrastructure, apoptosis, proliferation and differentiation of urothelial cells. Repeated doses of CP caused the development of hyperplastic urothelium with up to ten cell layers and increased proliferation and apoptotic indices regarding Ki-67 and active caspase-3 immunohistochemistry, respec- tively. Scanning electron microscopy observations, cytoker- atin and asymmetrical unit membrane immunohistochemistry and Western blot analysis showed a lower differentiation state of superficial urothelial cells. Melatonin co-treatment prevented the development of hyperplastic urothelium, statistically significantly decreased proliferation and apo- ptotic indices after four and five doses of CP and caused higher differentiation state of superficial urothelial cells. Keywords Urothelium . Cyclophosphamide . Hyperplasia . Melatonin Introduction The three-layered mammalian urinary bladder urothelium is a normally very stable and highly differentiated epithelium [1]. Nevertheless, it shows a great capacity for regeneration after a mechanically or chemically induced damage [1–3]. Urothelial regeneration usually gives rise to hyperplasia and can contribute to a wide range of neoplasms [2]. Hicks and Chowaniec [2] and our unpublished data showed that repeated intraperitoneal doses of cyclophosphamide (CP) cause persistent urothelial hyperplasia. Cohen et al. have developed a model of carcinogenesis which incorporates several basic principles: cancer results from genetic errors within normal cells; carcinogenesis is a multi-stage process, requiring more than one genetic event; and a chance of a genetic event occurring per cell division is a finite quantity and, therefore, that increased cell division increases the chance for a genetic event [4]. Based on this model, they then postulate that changes which lead to increased cell proliferation in the urothelium should be carefully evaluat- ed as a possible contributing factor to carcinogenesis [5–7]. CP has long been known to produce severe hyperplasia following cytotoxic damage to the urothelium [8, 9]. However, it is also a weak complete carcinogen and after prolonged application will produce cancer of the urothe- lium, but its hyperplastic action is probably associated with its late-stage carcinogenic activity rather than with its first- stage initiating activity [10]. In a lifelong experiment of the carcinogen action of CP, low-dose oral application of CP led to the induction of bladder cancer in 30% to 43% of rats [11]. However, in the last few months of the experiments, Virchows Arch (2009) 454:657–666 DOI 10.1007/s00428-009-0765-3 D. Zupančič (*) : K. Jezernik Institute of Cell Biology, Faculty of Medicine, Lipičeva 2, 1000 Ljubljana, Slovenia e-mail: [email protected] G. Vidmar Institute for Rehabilitation, Linhartova cesta 51, 1000 Ljubljana, Slovenia
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Melatonin prevents the development of hyperplastic urothelium induced by repeated doses of...

ORIGINAL ARTICLE
Melatonin prevents the development of hyperplasticurothelium induced by repeated doses of cyclophosphamide
Daša Zupančič & Gaj Vidmar & Kristijan Jezernik
Received: 16 October 2008 /Revised: 14 February 2009 /Accepted: 19 March 2009 /Published online: 21 April 2009# Springer-Verlag 2009
Abstract Repeated cyclophosphamide (CP) chemotherapyincreases the risk of developing bladder cancer, whichcould be due to the extremely rapid proliferation ofurothelial cells observed in hyperplastic urothelium inducedby CP treatment. We investigated the effect of melatonin onthe development of urothelial hyperplasia induced byrepeated CP treatment. Male ICR mice were injected withCP (150 mg/kg) or melatonin (10 mg/kg) with CP once aweek for 3, 4 and 5 weeks. Transmission and scanningelectron microscopy, immunohistochemistry and Westernblot analysis were used to study the ultrastructure,apoptosis, proliferation and differentiation of urothelialcells. Repeated doses of CP caused the development ofhyperplastic urothelium with up to ten cell layers andincreased proliferation and apoptotic indices regardingKi-67 and active caspase-3 immunohistochemistry, respec-tively. Scanning electron microscopy observations, cytoker-atin and asymmetrical unit membrane immunohistochemistryand Western blot analysis showed a lower differentiationstate of superficial urothelial cells. Melatonin co-treatmentprevented the development of hyperplastic urothelium,statistically significantly decreased proliferation and apo-ptotic indices after four and five doses of CP and causedhigher differentiation state of superficial urothelial cells.
Keywords Urothelium . Cyclophosphamide . Hyperplasia .
Melatonin
Introduction
The three-layered mammalian urinary bladder urothelium isa normally very stable and highly differentiated epithelium[1]. Nevertheless, it shows a great capacity for regenerationafter a mechanically or chemically induced damage [1–3].Urothelial regeneration usually gives rise to hyperplasia andcan contribute to a wide range of neoplasms [2]. Hicks andChowaniec [2] and our unpublished data showed thatrepeated intraperitoneal doses of cyclophosphamide (CP)cause persistent urothelial hyperplasia. Cohen et al. havedeveloped a model of carcinogenesis which incorporatesseveral basic principles: cancer results from genetic errorswithin normal cells; carcinogenesis is a multi-stage process,requiring more than one genetic event; and a chance of agenetic event occurring per cell division is a finite quantityand, therefore, that increased cell division increases thechance for a genetic event [4]. Based on this model, theythen postulate that changes which lead to increased cellproliferation in the urothelium should be carefully evaluat-ed as a possible contributing factor to carcinogenesis [5–7].CP has long been known to produce severe hyperplasiafollowing cytotoxic damage to the urothelium [8, 9].However, it is also a weak complete carcinogen and afterprolonged application will produce cancer of the urothe-lium, but its hyperplastic action is probably associated withits late-stage carcinogenic activity rather than with its first-stage initiating activity [10]. In a lifelong experiment of thecarcinogen action of CP, low-dose oral application of CPled to the induction of bladder cancer in 30% to 43% of rats[11]. However, in the last few months of the experiments,
Virchows Arch (2009) 454:657–666DOI 10.1007/s00428-009-0765-3
D. Zupančič (*) :K. JezernikInstitute of Cell Biology, Faculty of Medicine,Lipičeva 2,1000 Ljubljana, Sloveniae-mail: [email protected]
G. VidmarInstitute for Rehabilitation,Linhartova cesta 51,1000 Ljubljana, Slovenia

repeated intraperitoneal injections of CP were given to ratsand mice and no bladder tumours were observed [2, 12].Nevertheless, it is known that patients treated withcyclophosphamide have up to a ninefold increased risk ofdeveloping bladder cancer [13, 14].
Urothelial carcinogenesis in the rat and mouse proceedsthrough a sequence of morphologic changes beginning assimple hyperplasia [15]. In the rat, it then progresses tonodular and papillary hyperplasia, while in the mouse nodularhyperplasia is considerably more common than papillaryproliferations [4]. The rat model then progress to papillomasand strongly resembles low-grade papillary urothelial neopla-sia in humans, whereas the mouse model resembles the high-grade, highly malignant disease of humans [15].
A single-dose CP treatment causes a destruction of thebladder urothelium [8, 9, 16, 17], followed by rapidproliferation of urothelial cells resulting in reversible hyper-plasia and gradual restoration of normal three-layeredurothelium [6, 18–20]. It is documented that reversiblehyperplasia is accompanied not only by an increasedproliferation activity of urothelial cells but also by anincreased number of apoptotic cells [20] and lower differen-tiation state of superficial urothelial cells [16, 21, 22]. In thehyperplastic urothelium, most of the urothelial cells undergoapoptosis within the superficial layer and are exfoliated intothe lumen of the urinary bladder [20] and subsequentlyremoved from the body with urine and lost for detection.Thus, it is important to identify apoptotic cells before classicalfeatures of apoptosis are present, which can be achieved byimmunohistochemical detection of active caspase-3 [19, 23,24]. Specific antisera allow a reliable detection of apoptoticcells without overlap with necrotic cell population [25, 26].Caspase-3 is an effector caspase and when activated byinitiator caspases, such as caspase-8, it cleaves varioussubstrates, leading to the apoptotic cell death [24, 27].
The terminally differentiated superficial urothelial cells–umbrella cells have two unique characteristics: a subapicaltrajectorial network of cytokeratins [22] and membrane–protein plaques, which cover the apical surface of theumbrella cells, forming a rigid-looking asymmetrical unitmembrane (AUM) [28–31]. Immunostaining against AUMproteins may serve as a well-defined marker of celldifferentiation, since it shows no labelling signal in theundifferentiated cells with microvilli, whereas stronglabelling in the umbrella cells with rigid-looking micro-ridges can be observed [21, 22]. Anti-AUM antibody ismade against highly purified bovine AUM and reactsstrongly with uroplakin III (47 kDa) [32]. Differentiationof urothelial cells can also be characterised by changes inthe cytokeratin (CK) expression patterns. For example, CK7 is expressed in differentiated and undifferentiated cells,whereas CK 20 is present exclusively in terminallydifferentiated umbrella cells [33–37].
In the last decade, research has specifically addressed therole of melatonin, the main secretory product of the pinealgland, in controlling the proliferation of neoplastic cells[38]. Both in vitro and on different animal models oftumorigenesis, melatonin has been found to inhibit cellproliferation and to delay tumour progression [39, 40].Most studies exploring the relationship between melatoninand oncogenesis have focused on melanoma and breastcancer, but more recently melatonin has been shown toinfluence neoplastic growth in many other cancers includ-ing carcinomas of the urinary bladder [41]. Furthermore,increasing evidence indicates that melatonin may exert itsanticancer effects not only via inhibition of cell prolifera-tion but also by stimulation of differentiation and apoptosis[39]. Moreover, it is documented that melatonin has aprotective effect against single-dose CP-induced urothelialdamage [42, 43] and a favourable impact on regenerationand restoration of normal urothelium, since it reduces thenumber of apoptotic and proliferating urothelial cells andresults in their earlier differentiation [43].
Repeated doses of CP cause the development of urothelialhyperplasia, which could lead to bladder cancer development[6]. In different studies, melatonin has been found to inhibitcell proliferation and also influence differentiation andapoptosis. The aim of this study was therefore to examinethe effect of melatonin on the development of hyperplasticurothelium and on the proliferation, apoptosis and differen-tiation of urothelial cells after repeated doses of CP.
Materials and methods
Animals and treatment
The animal experiments were approved by the Slovenianveterinary administration of the Ministry of Agriculture,Forestry and Food according to the Animal HealthProtection Act and Introductions for Granting Permits forAnimal Experimentation for Scientific Purposes. Fortyalbino ICR (CD-1) adult male mice were divided into tengroups by a simple random sampling method. They werehoused in plastic cages at 23±2°C and 50% to 60% relativehumidity. Basal diet and water were available ad libitum.
Group 1 (control) animals were injected with saline oncea week for 3 weeks and they served as controls, whereasgroup 2 (Mel) received 15 mg/kg of body weight ofmelatonin once a week for 3 weeks and served as controlfor melatonin administration only. All the animals fromgroups 3 to 10 were injected with CP (Sigma) at a dose of150 mg/kg of body weight once a week for 3, 4 and5 weeks and half of them (groups 7 to 10) also received15 mg/kg of body weight of melatonin with CP, before andthe day after CP injection. All the drugs were administered
658 Virchows Arch (2009) 454:657–666

intraperitoneally (i.p.). Animals were sacrificed after three(3×), four (4×) and five (5×) doses of CP or melatonin andCP co-treatment (Mel+CP) and 1 month after five doses(1 m) of CP or melatonin and CP co-treatment (Mel+CP).
From each group, the bladders were processed forelectron microscopy, for active caspase-3, Ki-67, CK 7,CK 20 and AUM immunohistochemistry and for Westernblot analysis.
Electron microscopy
The tissue was fixed in 4.5% paraformaldehyde and 2%glutaraldehyde for 3 h. For transmission electron micros-copy, the samples were then postfixed in osmium tetroxide,dehydrated in ethanol and embedded in Epon. Ultrathinsections were stained with lead citrate and uranyl acetateand examined with a Jeol 100 CX electron microscope(Jeol Ltd., Tokyo, Japan). For scanning electron microsco-py, the samples were postfixed in osmium tetroxide andcritical-point dried. After sputter-coating with gold, theywere examined at 15 kV with a Jeol JSM 840 A scanningelectron microscope (Jeol Ltd., Tokyo, Japan).
Immunohistochemical detection of active caspase-3, Ki-67and AUM
The tissue was fixed in 4% paraformaldehyde overnightand embedded in paraffin. After deparaffinisation, theendogenous peroxidase activity was blocked by 3% H2O2
in methanol. For detection of Ki-67, sections weremicrowave-heated. Non-specific labelling was blocked by2.5% bovine serum albumin (Sigma, Taufkirchen, Ger-many) for Ki-67 or by 20% normal swine serum for activecaspase-3 and AUM. Sections were incubated with primarymonoclonal anti-Ki-67 (clone MIB-5, Dako, Glostrup,Denmark; 1:50 dil.), polyclonal anti-active-caspase-3(R&D Systems, Wiesbaden, Germany; 1:1,500 dil.) orpolyclonal rabbit antibodies raised against highly purifiedbovine AUM (a kind gift from Prof. T.-T. Sun, Departmentof Dermatology, New York University Medical School,New York, NY 10016; 1:10,000 dil.) overnight at 4°C. Fornegative controls, the incubation with primary antibody wasomitted or the specific primary antibody was replaced by anon-relevant antibody. For secondary antibodies, biotiny-lated rabbit anti-mouse (Dako, Glostrup, Denmark; 1:100dil.) or swine anti-rabbit immunoglobulins (Dako, Glostrup,Denmark; 1:400 dil.) were applied for 1 h at roomtemperature, followed by ABC/HRP complex (Dako,Glostrup, Denmark) incubation. After the standard DAB(Sigma, Taufkirchen, Germany) development procedure,sections were counterstained with haematoxylin and exam-ined with a Nikon Eclipse TE300 fluorescence microscope(Nikon Corporation, Tokyo, Japan).
Immunohistochemical detection of CK 7 and CK 20
The tissue was frozen and cut into 7-µm-thick cryosectionsand fixed in absolute ethanol (−20°C) for 20 min. Afterblocking with 2% BSA in PBS, incubation with primarymonoclonal anti-CK 7 (clone OV/TL 12/30, Dako, Glostrup,Denmark; 1:20 dil.) and anti-CK 20 (clone KS20.8, Dako,Glostrup, Denmark; 1:100 dil.) was performed overnight at4°C. For negative controls, the incubation with primaryantibody was omitted or the specific primary antibody wasreplaced by a non-relevant antibody. Adequate secondaryantibodies conjugated with TRITC (Sigma, Taufkirchen,Germany) were incubated for 1 h at 37°C. After washing,the sections were mounted in mounting medium Vectashieldwith DAPI (Vector Laboratories, Burlingame, CA, USA) andexamined with a Nikon Eclipse TE300 fluorescence micro-scope (Nikon Corporation, Tokyo, Japan).
Western blot analysis
Urothelial cells were scraped from the wall of the urinarybladder and homogenised in ice-cold buffer (0.8 M Tris–HCl,7.5% SDS, 1 mM phenylmethylsulfonyl fluoride). The lysatewas centrifuged and the protein concentration in the superna-tant was determined by using a BCA™ protein assay kit(Pierce, Rockford, IL, USA). Sample proteins (30 µg/lane)were size-fractionated on 12% SDS–polyacrylamide gels andthen transferred to Hybond ECL nitrocellulose membranes(Amersham Biosciences, Little Chalfont, UK) by electro-blotting. After blocking overnight at 4°C in 5% skim milk inPBS-Tween, membranes were incubated for 2 h at roomtemperature with primary polyclonal rabbit antibodies raisedagainst highly purified bovine AUM (a kind gift fromProf. T.-T. Sun, Department of Dermatology, New YorkUniversity Medical School, New York, NY 10016, USA;1:10,000 dil.) or monoclonal anti-CK 7 (Santa Cruz Biotech-nology, Inc., Santa Cruz, CA, USA; 1:200 dil.). After washingin PBS-Tween, membranes were incubated for 1 h withadequate secondary antibodies horseradish peroxidase-conjugated goat anti-rabbit (Sigma, Taufkirchen, Germany),diluted 1:1,000, or anti-mouse (Dako, Glostrup, Denmark),diluted 1:1,000. Membranes were finally probed with enhancedchemoluminescence reagent (Amersham Biosciences, LittleChalfont, UK) and exposed to X-ray films. Rabbit anti-actinantibodies, diluted 1:1,000 (Sigma, Taufkirchen, Germany),were used to confirm an equal loading of protein in each lane.
Methods and counting for quantitative analysis
Four animals of each group were used for immunohisto-chemical detection of active caspase-3 and detection ofKi-67 on paraffin sections. Urinary bladders were cut intoserial tissue sections and three sections of each urinary
Virchows Arch (2009) 454:657–666 659
bladder were examined. Cells with a positive immunohis-tochemical reaction against active caspase-3 and Ki-67were accepted as labelled and used for scoring of apoptoticand proliferation indices, respectively. All urothelial cells pertissue section were counted and the indices for each tissuesection were defined as a percentage ratio of labelledurothelial cells to the total number of counted cells.
Statistical analysis
For each animal, the mean of the three slices was used forfurther analysis of apoptotic index and of proliferationindex. First, the distributions of both indices were plotted
for each group using boxplots. Because of departures fromnormality, and because of non-homogeneous variances, thedata were analysed using two-way analysis of variance onranks instead of on original observations, which is a robustyet efficient solution [44].
Since the effect of different doses of CP was found to behighly statistically significant, post-hoc comparisons wereperformed between different doses of CP using the Scheffeprocedure on the ranked data (it should be noted that othermethods, such as Sidak's, or Dunnet's method for plannedcomparisons with baseline, lead to equivalent conclusions).Since the effect of group was also highly statisticallysignificant, while the (non-)significance of the interaction
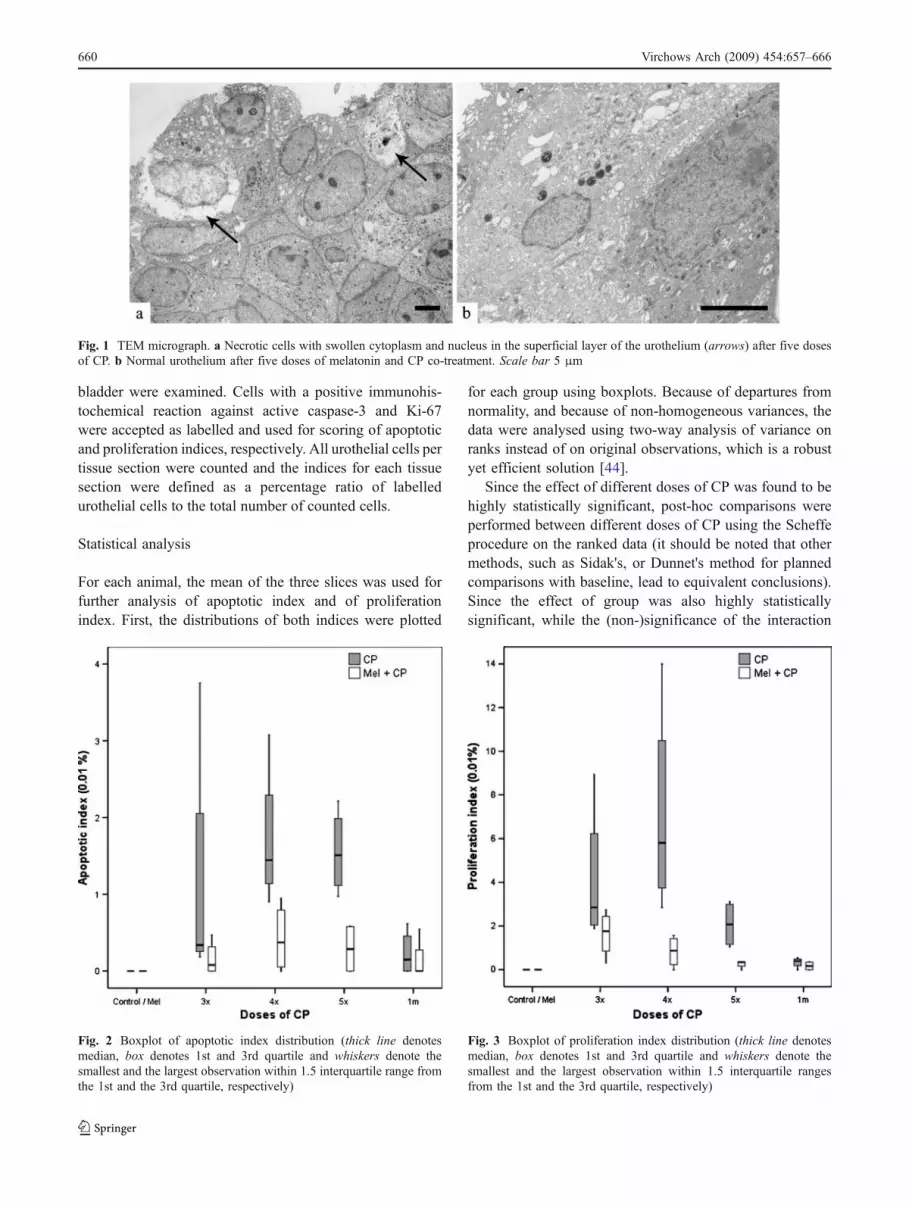
Fig. 1 TEM micrograph. a Necrotic cells with swollen cytoplasm and nucleus in the superficial layer of the urothelium (arrows) after five dosesof CP. b Normal urothelium after five doses of melatonin and CP co-treatment. Scale bar 5 μm
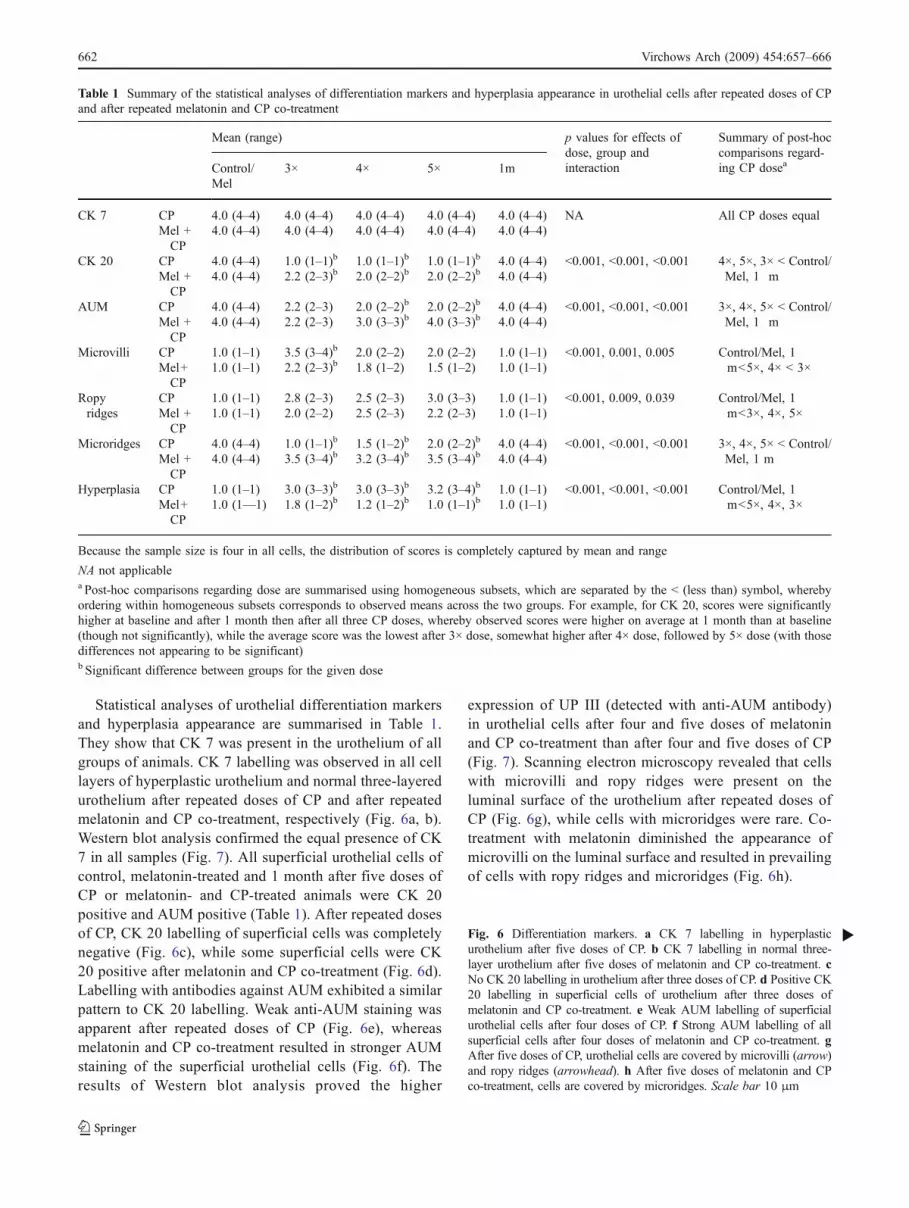
Fig. 2 Boxplot of apoptotic index distribution (thick line denotesmedian, box denotes 1st and 3rd quartile and whiskers denote thesmallest and the largest observation within 1.5 interquartile range fromthe 1st and the 3rd quartile, respectively)
Fig. 3 Boxplot of proliferation index distribution (thick line denotesmedian, box denotes 1st and 3rd quartile and whiskers denote thesmallest and the largest observation within 1.5 interquartile rangesfrom the 1st and the 3rd quartile, respectively)
660 Virchows Arch (2009) 454:657–666

effect should be treated with caution in ANOVA on rankeddata [45], the groups with or without melatonin co-treatment were compared after different doses of CP usingthe exact Mann–Whitney test.
For morphological data (urothelial differentiationmarkers and hyperplasia appearance), which were in theform of ratings of four-point scale (from 1 = least present to4 = maximum presence), the median value of the threesections was calculated and analysed in the same way asthat for apoptotic and proliferation indices.
Results
In control and melatonin-treated animals, the urotheliumwas composed of three cell layers and was ultrastructurallynormal. Repeated CP treatment at 3, 4 and 5 weeks causedthe development of hyperplastic urothelium with up to tencell layers. After three, four and five doses of CP,prominent dilatations in the intercellular spaces were
observed and some necrotic cells were perceived in thesuperficial layer of the urothelium (Fig. 1a). Melatonin co-treatment resulted in the normal appearance of theurothelium (Fig. 1b). One month after five doses of CPand 1 month after five doses of melatonin and CP co-treatment, the urothelium was composed of three cell layersand was ultrastructurally similar to normal urothelium.
Active caspase-3-positive and Ki-67-positive cells incontrol and melatonin-treated animals were extremely rareand the mean for all indices was zero (Figs. 2 and 3). Afterrepeated doses of CP (3×, 4× and 5×), substantial elevationof apoptotic and proliferation index was detected, whilemelatonin co-treatment resulted in much less apoptotic andproliferation index increase (Figs. 2 and 3). An activecaspase-3 and Ki-67 reaction positive signal was observedmainly in the cytoplasm of the superficial urothelial cells andin the nuclei of the basal cell layer, respectively (Figs. 4 and 5).One month after five doses of CP and 1 month after fivedoses of melatonin and CP co-treatment, the apoptotic andproliferation indices returned to near the initial zero level.
Fig. 4 Active caspase-3 immunohistochemistry. a Active caspase-3-positive cell (brown) in the urothelium after four doses of CP and b afterfour doses of melatonin and CP co-treatment. Scale bar 10 μm
Fig. 5 Ki-67 immunohistochemistry. a Ki-67-positive cells (brown nuclei) are most abundant in the basal layer of the urothelium after four dosesof CP and b after four doses of melatonin and CP co-treatment. Scale bar 10 μm
Virchows Arch (2009) 454:657–666 661
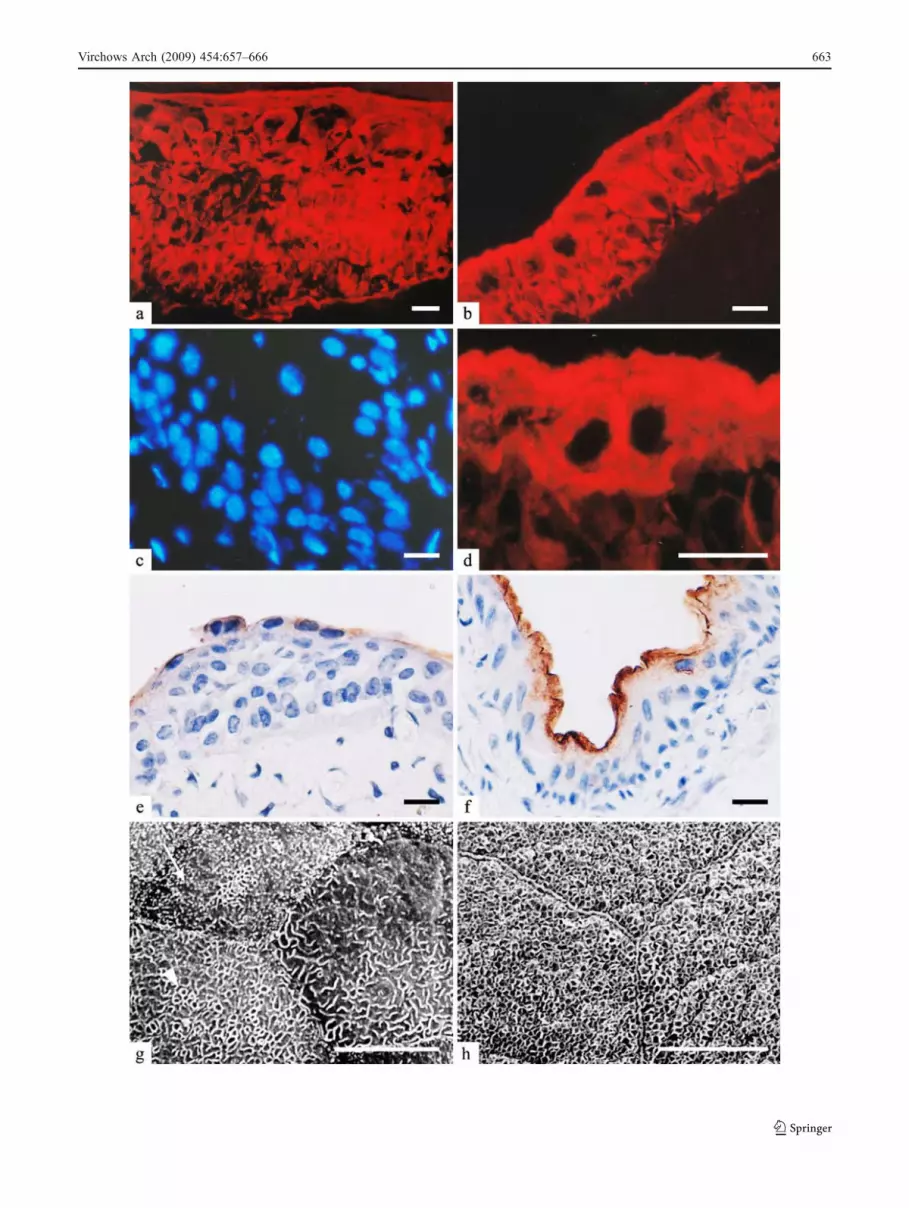
Statistical analyses of urothelial differentiation markersand hyperplasia appearance are summarised in Table 1.They show that CK 7 was present in the urothelium of allgroups of animals. CK 7 labelling was observed in all celllayers of hyperplastic urothelium and normal three-layeredurothelium after repeated doses of CP and after repeatedmelatonin and CP co-treatment, respectively (Fig. 6a, b).Western blot analysis confirmed the equal presence of CK7 in all samples (Fig. 7). All superficial urothelial cells ofcontrol, melatonin-treated and 1 month after five doses ofCP or melatonin- and CP-treated animals were CK 20positive and AUM positive (Table 1). After repeated dosesof CP, CK 20 labelling of superficial cells was completelynegative (Fig. 6c), while some superficial cells were CK20 positive after melatonin and CP co-treatment (Fig. 6d).Labelling with antibodies against AUM exhibited a similarpattern to CK 20 labelling. Weak anti-AUM staining wasapparent after repeated doses of CP (Fig. 6e), whereasmelatonin and CP co-treatment resulted in stronger AUMstaining of the superficial urothelial cells (Fig. 6f). Theresults of Western blot analysis proved the higher
expression of UP III (detected with anti-AUM antibody)in urothelial cells after four and five doses of melatoninand CP co-treatment than after four and five doses of CP(Fig. 7). Scanning electron microscopy revealed that cellswith microvilli and ropy ridges were present on theluminal surface of the urothelium after repeated doses ofCP (Fig. 6g), while cells with microridges were rare. Co-treatment with melatonin diminished the appearance ofmicrovilli on the luminal surface and resulted in prevailingof cells with ropy ridges and microridges (Fig. 6h).
Table 1 Summary of the statistical analyses of differentiation markers and hyperplasia appearance in urothelial cells after repeated doses of CPand after repeated melatonin and CP co-treatment
Mean (range) p values for effects ofdose, group andinteraction
Summary of post-hoccomparisons regard-ing CP doseaControl/
Mel3× 4× 5× 1m
CK 7 CP 4.0 (4–4) 4.0 (4–4) 4.0 (4–4) 4.0 (4–4) 4.0 (4–4) NA All CP doses equalMel +CP
4.0 (4–4) 4.0 (4–4) 4.0 (4–4) 4.0 (4–4) 4.0 (4–4)
CK 20 CP 4.0 (4–4) 1.0 (1–1)b 1.0 (1–1)b 1.0 (1–1)b 4.0 (4–4) <0.001, <0.001, <0.001 4×, 5×, 3× < Control/Mel, 1 mMel +
CP4.0 (4–4) 2.2 (2–3)b 2.0 (2–2)b 2.0 (2–2)b 4.0 (4–4)
AUM CP 4.0 (4–4) 2.2 (2–3) 2.0 (2–2)b 2.0 (2–2)b 4.0 (4–4) <0.001, <0.001, <0.001 3×, 4×, 5× < Control/Mel, 1 mMel +
CP4.0 (4–4) 2.2 (2–3) 3.0 (3–3)b 4.0 (3–3)b 4.0 (4–4)
Microvilli CP 1.0 (1–1) 3.5 (3–4)b 2.0 (2–2) 2.0 (2–2) 1.0 (1–1) <0.001, 0.001, 0.005 Control/Mel, 1m<5×, 4× < 3×Mel+
CP1.0 (1–1) 2.2 (2–3)b 1.8 (1–2) 1.5 (1–2) 1.0 (1–1)
Ropyridges
CP 1.0 (1–1) 2.8 (2–3) 2.5 (2–3) 3.0 (3–3) 1.0 (1–1) <0.001, 0.009, 0.039 Control/Mel, 1m<3×, 4×, 5×Mel +
CP1.0 (1–1) 2.0 (2–2) 2.5 (2–3) 2.2 (2–3) 1.0 (1–1)
Microridges CP 4.0 (4–4) 1.0 (1–1)b 1.5 (1–2)b 2.0 (2–2)b 4.0 (4–4) <0.001, <0.001, <0.001 3×, 4×, 5× < Control/Mel, 1 mMel +
CP4.0 (4–4) 3.5 (3–4)b 3.2 (3–4)b 3.5 (3–4)b 4.0 (4–4)
Hyperplasia CP 1.0 (1–1) 3.0 (3–3)b 3.0 (3–3)b 3.2 (3–4)b 1.0 (1–1) <0.001, <0.001, <0.001 Control/Mel, 1m<5×, 4×, 3×Mel+
CP1.0 (1––1) 1.8 (1–2)b 1.2 (1–2)b 1.0 (1–1)b 1.0 (1–1)
Because the sample size is four in all cells, the distribution of scores is completely captured by mean and range
NA not applicablea Post-hoc comparisons regarding dose are summarised using homogeneous subsets, which are separated by the < (less than) symbol, wherebyordering within homogeneous subsets corresponds to observed means across the two groups. For example, for CK 20, scores were significantlyhigher at baseline and after 1 month then after all three CP doses, whereby observed scores were higher on average at 1 month than at baseline(though not significantly), while the average score was the lowest after 3× dose, somewhat higher after 4× dose, followed by 5× dose (with thosedifferences not appearing to be significant)b Significant difference between groups for the given dose
Fig. 6 Differentiation markers. a CK 7 labelling in hyperplasticurothelium after five doses of CP. b CK 7 labelling in normal three-layer urothelium after five doses of melatonin and CP co-treatment. cNo CK 20 labelling in urothelium after three doses of CP. d Positive CK20 labelling in superficial cells of urothelium after three doses ofmelatonin and CP co-treatment. e Weak AUM labelling of superficialurothelial cells after four doses of CP. f Strong AUM labelling of allsuperficial cells after four doses of melatonin and CP co-treatment. gAfter five doses of CP, urothelial cells are covered by microvilli (arrow)and ropy ridges (arrowhead). h After five doses of melatonin and CPco-treatment, cells are covered by microridges. Scale bar 10 μm
b
662 Virchows Arch (2009) 454:657–666
Virchows Arch (2009) 454:657–666 663

The distributions of apoptotic index and proliferationindex are depicted in Figs. 2 and 3, respectively.
For apoptotic index and for proliferation index, the maineffects of different doses of CP and of CP only or melatoninand CP co-treatment were statistically significant (p<0.001from two-way ANOVA on ranked data), while theinteraction effect was not (p=0.161 and p=0.053 forapoptotic and proliferation indices, respectively).
Post-hoc comparisons between different doses of CPindicate that, for both apoptotic and proliferation index, theaverage does not differ statistically significantly betweencontrol/Mel and 1-m groups (p=0.605 and p=0.175 forapoptosis and proliferation index, respectively), while thatlevel differs significantly from the 3×, 4× and 5× doses ofCP (p<0.05), which, in turn, do not differ significantlyamong themselves (p>0.05).
Comparisons between the CP and Mel+CP groups afterdifferent doses showed that the groups differed mostnotably at 4× and 5× (p=0.029 for apoptosis andproliferation index at both doses). They differed less at 3×without reaching statistical significance (p=0.029 forapoptosis and proliferation index), they were practicallyequal at 1 m (p=0.686 and p=0.200 for apoptosis andproliferation index, respectively), and they were identical atthe onset of the study as expected (p=1.000 for bothindices). It should be noted that the p values forcomparisons between groups after different doses of CPmight have been adjusted for multiple tests, but they areconservative enough because of data aggregation (and thusa reduced sample size, which is very small).
Discussion
Hicks and Chowaniec [2] and our previous unpublishedexperiments showed that repeated doses of CP causeurothelial hyperplasia. In the present study, we demonstrat-ed that melatonin prevents the development of hyperplasticurothelium induced by repeated CP treatment.
Cohen and co-workers [4–7, 46, 47] proposed a modelof carcinogenesis which takes into account the proliferativeeffects of agents in carcinogenesis. In the current experi-ment, we demonstrated that urothelial hyperplasia inducedby repeated-dose CP is accompanied by increased prolifer-ation activity of urothelial cells. Cohen et al. assumed that,every time a cell undergoes a cell division, there is a lowprobability, but not zero, of a genetic error occurring [4, 6].It is, in general, accepted that at least two irreversiblegenetic events must occur for tumour formation. Thus, theextremely rapid proliferation of urothelial cells seenfollowing repeated CP treatment greatly increased thelikelihood of at least one genetic error occurring (initiation).If the increased cell proliferation would be continued for along period of time, the likelihood of a second promotingevent would increase sufficiently that a significant inci-dence of tumours would occur. Persistent high-proliferationindices observed after repeated CP doses could increase theprobability of bladder carcinogenesis and therefore explainthe fact that patients treated with repeated CP chemotherapyhave up to ninefold increased risk of developing bladdercancer [13, 14].
In numerous experiments in vitro and in different animalmodels of tumorigenesis, melatonin has been found toinhibit proliferation and neoplastic growth [38, 40, 48].Some studies also documented that melatonin reduced thecell proliferation in neoplastically unchanged cells [49–51].It is therefore not surprising that, in the present experiment,melatonin significantly decreased the proliferation activityof urothelial cells and therefore prevented hyperplasia fromdeveloping. Since the rapid proliferation of urothelial cellsseen after repeated doses of CP could increase thelikelihood of bladder carcinogenesis [4–7], melatonin mightprevent the development of bladder tumours by decreasingthe proliferation activity of urothelial cells.
Experimentally induced urothelial hyperplasia is accom-panied by an increased number of apoptotic cells [20, 52–54]. It happened similarly in the present experiment, sincenot only increased proliferation but also increased apoptotic
Fig. 7 Western blot analysis of CK 7 and AUM. CK 7 (54 kDa) isequally present in urothelial cells after repeated doses of CP and afterrepeated doses of melatonin and CP co-treatment. After four and fivedoses of melatonin and CP co-treatment, the expression of UP III
(47 kDa; detected with anti-AUM antibody) is higher than after fourand five doses of CP. The immunoblot experiments were performedtwice with similar results. MW molecular weight standards
664 Virchows Arch (2009) 454:657–666

indices were found in the hyperplastic urothelium afterrepeated doses of CP. Melatonin co-treatment significantlydecreased not only the proliferation but also the apoptosisof urothelial cells. Numerous reports documented the roleof melatonin in inhibiting apoptosis in different cells afterischemia [55–57], hyperoxia [58] or oxidative stress [59].Moreover, melatonin also decreased the apoptosis ofurothelial cells after a single-dose CP treatment [43]. Allthese experiments investigated the acute injury in whichmelatonin diminished cell death, while in the present studythe anti-apoptotic effect of melatonin was observed inchronic hyperplastic state. It is possible that since melatonininhibited the proliferation activity of urothelial cells andprevented the development of hyperplastic urothelium,decreased apoptosis of urothelial cells appeared per se andnot as a consequence of direct melatonin action.
The scanning electron microscopy observations andexpression of urothelial key differentiation markers, the CK7, CK 20 and AUM, showed that the differentiation state ofsuperficial urothelial cells of hyperplastic urothelium, inducedby repeated doses of CP, was lower than in control andresembled the differentiation of superficial urothelial cellsduring reversible hyperplasia following the destruction of theurothelium after a single dose of CP [16, 21, 22, 60].Melatonin co-treatment resulted in the higher differentiationstate of superficial urothelial cells as indicated by theappearance of urothelial apical surface and confirmed bythe expression of analysed differentiation markers. Theseresults are similar to a variety of in vivo and in vitroexperimental models of neoplasia, in which evidence wasshown that melatonin influences the differentiation of tumourcells [39, 61]. Although it is documented that melatonin iscapable of promoting differentiation of neoplastically un-changed osteoblasts and ovarian cells [49, 62], we do notknow whether melatonin influences urothelial cell differen-tiation directly or through its preventive effect against CP-induced hyperplasia. Further studies will be needed to gain acomplete insight into this question.
In conclusion, our results document that repeated dosesof CP cause the development of hyperplastic urotheliumwith rapid proliferation of urothelial cells, which could leadto an increased risk of bladder cancer. Melatonin co-treatment prevents the development of hyperplastic urothe-lium, statistically significantly decreases proliferation andapoptotic indices and causes higher differentiation state ofsuperficial urothelial cells. Despite the lack of clinicalstudies testing its clinical relevance, the present resultssuggest that supplementing patients treated with repeatedCP chemotherapy with adjuvant melatonin therapy maydecrease their risk of developing bladder cancer.
Acknowledgement The authors would like to thank Prof. T.-T. Sunfor providing the anti-AUM antibody used in this study.
Conflict of interest statement We declare that we have no conflictof interest.
References
1. Hicks RM (1975) The mammalian urinary bladder: an accommo-dating organ. Biol Rev 50:215–246
2. Hicks RM, Chowaniec J (1978) Experimental induction, histolo-gy, and ultrastructure of hyperplasia and neoplasia of the urinarybladder epithelium. Int Rev Exp Pathol 18:199–280
3. Liang DS (1962) Bladder regeneration following subtotalcystec-tomy. J Urol 88:503–505
4. Cohen SM, Ellwein LB (1991) Genetic errors, cell proliferation,and carcinogenesis. Cancer Res 51:6493–6505
5. Cohen SM (1983) Promotion in urinary bladder carcinogenesis.Environ Health Perspect 50:51–59
6. Cohen SM (1989) Toxic and nontoxic changes induced in theurothelium by xenobiotics. Toxicol Appl Pharmacol 101:484–498
7. Cohen SM, Greenfield RE, Ellwein LB (1983) Multistagecarcinogenesis in the urinary bladder. Environ Health Perspect49:209–215
8. Koss LG (1967) A light and electron microscopic study of theeffects of a single dose of cyclophosphamide on various organs inthe rat. Lab Invest 16:44–65
9. Locher GW, Cooper EH (1970) Repair of rat urinary bladderepithelium following injury by cyclophosphamide. Investig Urol8:116–123
10. Hicks RM (1983) Effect of promoters on incidence of bladdercancer in experimental animal models. Environ Health Perspect50:37–49
11. Schmähl D, Habs M (1979) Carcinogenic action of low-dosecyclophosphamide given orally to Sprague–Dawley rats in alifetime experiment. Int J Cancer 23:706–712
12. Tokuoka S (1965) Induction of tumor in mice with N,N-bis(2-chloroethyl)-N′,O-propylenephosphoric acid ester diamide (cyclo-phosphamide). Gann 56:537–541
13. Jung I, Messing E (2000) Molecular mechanisms and pathways inbladder cancer development and progression. Cancer Control7:325–334
14. Tuttle TM, Williams GM, Marshall FF (1988) Evidence forcyclophospamide-induced transitional cell carcinoma in a renaltransplant patient. J Urol 140:1009–1011
15. Cohen SM (2002) Comparative pathology of proliferative lesionsof the urinary bladder. Toxicol Pathol 30:663–671
16. Fukushima S, Arai M, Cohen SM et al (1981) Scanning electronmicroscopy of cyclophosphamide-induced hyperplasia of the raturinary bladder. Lab Invest 44:89–96
17. Philips FS, Sternberg SS, Cronin AP et al (1961) Cyclophospha-mide and urinary bladder toxicity. Cancer Res 21:1577–1589
18. Farsund T (1976) Cell kinetics of mouse urinary bladderepithelium. II. Changes in proliferation and nuclear DNA contentduring necrosis regeneration, and hyperplasia caused by a singledose of cyclophosphamide. Virchows Arch 21:279–298
19. Jezernik K, Romih R, Mannherz HG et al (2003) Immunohisto-chemical detection of apoptosis, proliferation and inducible nitricoxide synthase in rat urothelium damaged by cyclophosphamidetreatment. Cell Biol Int 27:863–869
20. Romih R, Koprivec D, Stiblar Martincic D et al (2001)Restoration of the rat urothelium after cyclophosphamide treat-ment. Cell Biol Int 25:531–537
21. Romih R, Jezernik K (1996) Reorganization of the urothelialluminal plasma membrane in the cyclophosphamide treated rats.Pflugers Arch Suppl 431:R241–R242
Virchows Arch (2009) 454:657–666 665

22. Romih R, Veranic P, Jezernik K (2002) Appraisal of differentia-tion markers in urothelial cells. Appl Immunohisto M M 10:339–343
23. Hoshi T, Sasano H, Kato K et al (1998) Immunohistochemistry ofcaspase3/CPP32 in human stomach and its correlation with cellproliferation and apoptosis. Anticancer Res 18:4347–4353
24. Stadelmann C, Lassmann H (2000) Detection of apoptosis intissue sections. Cell Tissue Res 301:19–31
25. Armstrong RC, Aja TJ, Hoang KD et al (1997) Activation of theCED3/ICE-related protease CPP32 in cerebellar granule neuronsundergoing apoptosis but not necrosis. J Neurosci 17:553–562
26. Sirinivasan A, Roth KA, Sayers RO et al (1998) In situimmunodetection of activated caspase-3 in apoptotic neurons inthe developing nervous system. Cell Death Differ 5:1004–1016
27. Bratton SB, MacFarlane M, Cain K et al (2000) Protein complexactivate distinct caspase cascades in death receptor and stress-induced apoptosis. Exp Cell Res 256:27–33
28. Hicks RM (1965) The fine structure of the transitional epitheliumof rat ureter. J Cell Biol 26:215–246
29. Kachar B, Liang F, Lin SU et al (1999) Three-dimensionalanalysis of the 16 nm urothelial plaque particle: luminal surfaceexposure, preferential head-to-head interaction, and hinge forma-tion. J Mol Biol 285:595–608
30. Koss LR (1969) The asymmetric unit membrane of the epitheliumof the urinary bladder of the rat. An electron microscopy study ofa mechanism of epithelial maturation and function. Lab Invest21:154–168
31. Staehelin LA, Chlapowski FJ, Bonneville MA (1972) Lumenalplasma membrane of the urinary bladder: 1. Three-dimensionalreconstruction from freeze-etch images. J Cell Biol 53:73–91
32. Wu XR, Manabe M, Yu J et al (1990) Large scale purification andimmunolocalization of bovine uroplakins I, II and III. Molecularmarkers of urothelial differentiation. J Biol Chem 265:19170–19179
33. Harnden P, Allam A, Joyce AD et al (1995) Cytokeratin 20expression by non-invasive transitional cell carcinomas: potentialfor distinguishing recurrent from non-recurrent disease. Histopa-thology 27:169–174
34. Moll R, Achtstätter T, Brecht E et al (1988) Cytokeratins innormal and malignant transitional epithelium. Maintenance ofexpression of urothelial differentiation features in transitional cellcarcinomas and bladder carcinoma cell culture lines. Am J Pathol132:123–144
35. Moll R, Schiller DL, Franke WW (1990) Identification of proteinIT of the intestinal cytoskeleton as a novel type I cytokeratin withunusual properties and expression patterns. J Cell Biol 77:685–697
36. Romih R, Jezernik K, Mašera A (1998) Uroplakins andcytokeratins in the regenerating rat urothelium after sodiumsaccharin treatment. Histochem Cell Biol 109:263–269
37. Schaafsma HE, Ramaeker FCS, Vanmujien GNP et al (1989)Distribution of cytokeratin polypeptides in epithelial of the adulthuman urinary tract. Histochemistry 91:151–191
38. Zachary I, Woll PJ, Rozengurt A (1987) A role for neuropeptidesin the control of cell proliferation. Dev Biol 124:295–308
39. Blask DE, Dauchy RT, Sauer LA (2005) Putting cancer to sleep atnight: the neuroendocrine/circadian melatonin signal. Endocr27:179–188
40. Hill SM, Blask DE (1988) Effects of the pineal hormone onthe proliferation and morphological characteristics of humanbreast cancer (MCF-7) in culture. Cancer Res 48:6121–6126
41. Blask DE (1993) Melatonin in oncology. In: Yi HS, Reiter RJ(eds) Melatonin. CRC, Boca Raton, pp 447–476
42. Topal T, Oztas Y, Korkmaz A et al (2005) Melatonin amelioratesbladder damage induced by cyclophosphamide in rats. J PinealRes 38:272–277
43. Zupančič D, Jezernik K, Vidmar G (2008) Effect of melatoninon apoptosis, proliferation and differentiation of urothelialcells after cyclophosphamide treatment. J Pineal Res 44:299–306
44. Hora SC, Conover WJ (1984) The F-statistic in the two-waylayout with rank-score transformed data. J Am Stat Assoc 79:668–673
45. Conover WJ (1999) Practical nonparametric statistics, 3rd edn.Wiley, New York
46. Ellwein LB, Cohen SM (1988) A cellular dynamics model ofexperimental bladder cancer: analysis of the effect of sodiumsaccharin in the rat. Risk Anal 8:215–221
47. Greenfield RE, Ellwein LB, Cohen SM (1984) A generalprobabilistic model of carcinogenesis: analysis of experimentalurinary bladder cancer. Carcinogenesis 5:437–445
48. Cini G, Coronnello M, Mini E et al (1998) Melatonin’s growth/inhibitory effect on hepatoma AH 130 in the rat. Cancer Lett125:51–59
49. Sainz RM, Mayo JC, Tan D-X (2003) Antioxidant activity ofmelatonin in Chinese hamster ovarian cells: changes in cellularproliferation and differentiation. Biochem Biophys Res Commun302:625–634
50. Lewinski A, Rybicka I, Wajs E et al (1991) Influence of pinealindoleamines on the mitotic activity of gastric and mucosaepithelial cells in the rat: interaction with omeprazole. J PinealRes 10:104–108
51. Yu HS, Hernandez V, Haywood M et al (1993) Melatonin inhibitsthe proliferation of retinal pigment epithelial (RPE) cells in vitro.In Vitro Cell Dev Biol Anim 29:415–418
52. Otori K, Yano Y, Takada N et al (1997) Reversibility andapoptosis in rat urinary bladder papillomatosis induced by uracil.Carcinogenesis 18:1485–1489
53. Santarosa R, Colombel MC, Kaplan S et al (1994) Hyperplasiaand apoptosis. Opposing cellular processes that regulate theresponse of the rabbit bladder to transient outlet obstruction. LabInvest 70:503–510
54. Shirai T, Shibata M-A, Takahashi S et al (1995) Differences in cellproliferation and apoptosis between reversible and irreversiblemucosal lesions associated with uracil-induced urolithiasis in N-butyl-N-(4-hydroxybutyl) nitrosamine-pretreated rats. Carcinogen-esis 16:501–505
55. Bertuglia S, Marchiafava PL, Colantuoni A (1996) Melatoninprevents ischemia reperfusion injury and hamster cheek pouchmicrocirculation. Cardiovasc Res 31:947–952
56. Kunduzova OR, Escourrou G, Seguelas M-H et al (2003)Prevention of apoptotic and necrotic cell death, caspase-3activation, and renal dysfunction by melatonin after ischemia/reperfusion. FASEB J 17:872–874
57. Osborne NN, Nash MS, Wood JP (1998) Melatonin counteractsischemia-induced apoptosis in human retinal pigment epithelialcells. Invest Ophthalmol Vis Sci 39:2374–2383
58. Shaikh AY, Xu J, Wu Y et al (1997) Melatonin protects bovinecerebral endothelial cells from hyperoxia-induced DNA damageand death. Neurosci Lett 229:193–197
59. Jou MJ, Peng TI, Reiter RJ et al (2004) Visualization of theantioxidative effects of melatonin at the mitochondrial levelduring oxidative stress-induced apoptosis of rat brain astrocytes.J Pineal Res 37:55–70
60. Veranič P, Romih R, Jezernik K (2004) What determines differen-tiation of urothelial umbrella cells? Eur J Cell Biol 83:27–34
61. Cos S, Fernandez R, Guezmes A et al (1998) Influence ofmelatonin on invasive and metastatic properties of MCF-7 humanbreast cancer cells. Cancer Res 58:4383–4390
62. Roth JA, Kim B-G, Lin W-L (1999) Melatonin promotesosteoblast differentiation and bone formation. J Biol Chem274:22041–22047
666 Virchows Arch (2009) 454:657–666




















