
Antitumor properties of two traditional aromatic rice genotypes ...
Upload
independentCategory
view
0download
0
ORIGINAL ARTICLE
Modulation by Melatonin of the Cardiotoxicand Antitumor Activities of Adriamycin
Chunghui Kim,* Nari Kim,* Hyun Joo,* Jae Boum Youm,* Won Sun Park,* Dang Van Cuong,*
Young Shik Park,* Euiyong Kim,* Churl-Ki Min,† and Jin Han*
Abstract: In this study, we investigated the effects of melatonin on
adriamycin-induced cardiotoxicity both in vivo in rats and in vitro,
and on the antitumor activities of adriamycin on MDA-231 and NCI
breast cancer cells. Rats that received a single intraperitoneal in-
jection of 25 mg/kg adriamycin showed a mortality rate of 86%,
which was reduced to 20% by melatonin treatment (10 mg/kg, SC for
6 days). Melatonin attenuated adriamycin-induced body-weight loss,
hemodynamic dysfunction, and the morphologic and biochemical
alterations caused by adriamycin. Melatonin also reduced adriamycin-
induced nuclear DNA fragmentation, as assessed by the comet
assay. In addition, the antitumor activity of adriamycin could be main-
tained using lower doses of this drug in combination with melatonin.
Melatonin treatment in the concentration range of 0.1–2.5 mM in-
hibited the growth of human breast cancer cells. In terms of oncolytic
activity, the combination of adriamycin and melatonin improved the
antitumor activity of adriamycin, as indicated by an increase in the
number of long-term survivors as well as decreases in body-weight
losses resulting from adriamycin treatment. These results indicate that
melatonin not only protects against adriamycin-induced cardiotox-
icity but also enhances its antitumor activity. This combination of
melatonin and adriamycin represents a potentially useful regimen for
the treatment of human neoplasms because it allows the use of lower
doses of adriamycin, thereby avoiding the toxic side effects asso-
ciated with this drug.
Key Words: melatonin, adriamycin, cardiotoxicity, nuclear DNA
fragmentation, antitumor activity
(J Cardiovasc PharmacolTM 2005;46:200–210)
Adriamycin, which is an anthracyclin antibiotic, is effectiveagainst a wide range of human neoplasms, such as acute
leukemia, lymphoma, and a number of solid tumors.1 How-ever, clinical applications of adriamycin have been restrictedby its toxicity, especially its cardiotoxic side effects, whichlead to cardiomyopathy and heart failure.2
Although the pathogenesis of adriamycin-induced car-diotoxicity remains unclear, oxygen free radical productionand lipid peroxidation are considered to be the 2 most im-portant factors.1,3 The underlying mechanism involves 1-electronreduction of adriamycin for the generation of the semiquinoneradical, which reduces oxygen, thereby producing superoxideand regenerating adriamycin. The net result of this process isthat adriamycin catalyzes the reduction of oxygen by NADPHto form a superoxide radical, which is subsequently reducedto hydrogen peroxide (H2O2) by the antioxidant enzymesuperoxide dismutase. In the presence of Fe2+, H2O2 is furtherreduced to the extremely reactive hydroxyl (�OH) radical,which can react with polyunsaturated fatty acids to yield lipidhydroperoxide. Biologic membranes contain large amounts ofpolyunsaturated fatty acids, which are susceptible to perox-idative attack by oxygen free radicals, resulting in lipid per-oxidation. Of the various organs in which peroxidative tissuedamage has been documented, the heart is known to be themost vulnerable to attack by free radicals produced followingadriamycin administration.4
Assuming that oxygen radical–induced lipid peroxida-tion of membrane lipids is indeed a major cause of adriamycin-induced cardiotoxicity, it seems reasonable to speculate thatantioxidants would prevent this toxicity. Indeed, several anti-oxidants have been reported to reduce or delay adriamycin-induced cardiotoxicity in experimental animals.5–7
Melatonin is produced and secreted by the pineal glandin almost all species of animal and is best known for its abilityto modulate circadian rhythms.8 There are numerous reportsdealing with the effects of melatonin, such as the inhibitionof cancer growth,9 stimulation of the immune system,10 andoxygen free radical scavenging.11 The antioxidant activityof melatonin has been ascribed to 2 different mechanisms:(1) melatonin reduces oxidative stress by stimulating antioxi-dant enzymes12,13; and (2) melatonin directly scavenges �OHradicals.14 It has been demonstrated that melatonin protectscells, tissues, and organs from oxidative damage that is in-duced by a variety of free radical–generating agents andprocesses.13,15 Melatonin acts effectively as an antioxidant inprotecting membrane lipids, nuclear DNA, and proteins fromoxidative damage both in vivo and in vitro.16,17
Received for publication December 7, 2004; accepted April 12, 2005.From the *Mitochondrial Signaling Laboratory, Department of Physiology
and Biophysics, College of Medicine, Cardiovascular & Metabolic DiseaseCenter, Biohealth Products Research Center, Inje University, Busan,Korea; and †College of Natural Science, Ajou University, Suwon, Korea.
This work was supported by Grants R05-2003-000-00413-0 andR05-2004-000-00905-0, and by the BioHealth Products Research Centerof the Korea Science and Engineering Foundation.
The first two authors contributed equally to this work.Chunghui Kim’s present address is Department of Animal Science & Bio-
technology, Jinju National University, Jinju, 660-758, Korea.Reprints: Jin Han, MD, PhD, Mitochondrial Signaling Laboratory, Depart-
ment of Physiology and Biophysics, College of Medicine, BiohealthProducts Research Center, 2020 Cardiovascular Institute, Inje University633-165 Gaegeum-Dong, Busanjin-Ku, Busan 614-735, Korea (e-mail:[email protected]).
Copyright � 2005 by Lippincott Williams & Wilkins
200 J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005

In this study, we investigated whether melatonin couldattenuate adriamycin-induced cardiotoxicity in rats both in vivoand in vitro. We examined adriamycin-induced cardiotoxicityusing various criteria, such as mortality, body-weight loss,hemodynamic dysfunction, morphologic alterations, nuclearDNA fragmentation as a measure of DNA damage, the level ofmalondialdehyde (MDA) as an index of lipid peroxidation,and lactate dehydrogenase (LDH) release and the level ofserum creatine kinase as indices of irreversible cell damage.We also investigated whether the protective effects of mel-atonin are mediated by alterations in the above criteria. Inaddition, it was of interest to examine the effects of melatoninon the antitumor effects of adriamycin. In this respect, weconsidered the direct antitumor effects of melatonin, whichmight influence the effects of adriamycin in antagonistic,additive, or synergistic ways. Thus, we determined the anti-tumor effects of melatonin on human breast cancer cells andexamined whether the antitumor efficacy of adriamycin couldbe maintained at lower doses when used in combination withmelatonin.
MATERIALS AND METHODS
AnimalsMale Sprague-Dawley rats (175–240 g) were used in
these experiments. The rats were fed on standard rat chow andhoused in plastic cages. Food and water were provided adlibitum. All of the experiments in this study were performedin accordance with the Guidelines for Animal Research of theNational Institutes of Health and were approved by the localCommittee on Animal Research.
ChemicalsAdriamycin was dissolved in normal saline immediately
before use in each experiment. Melatonin was dissolved indimethylsulfoxide (DMSO) and diluted as required in normalsaline. The DMSO concentration never exceeded 1%. Thechemicals were freshly prepared immediately before use. Allthe other chemicals used in this study, unless specified other-wise, were purchased from Sigma Chemical Co (St Louis, MO).
Dose–Response Kinetics ofAdriamycin-Induced Mortality
Four groups of rats were used to evaluate the dose–response kinetics of adriamycin toxicity. The percentage ofanimals that survived in each group was plotted against time(days) after adriamycin injection. Each animal receiveda single IP injection of adriamycin. Group 1 was the control(1% DMSO) (n = 25); group 2 received 5 mg/kg adriamycin(n = 25); group 3 received 15 mg/kg adriamycin (n = 26);and group 4 received 25 mg/kg adriamycin (n = 23). To assessthe effect of melatonin on adriamycin-induced cardiac dam-age, the rat hearts were used in the assay described below.
Modulation of Adriamycin-Induced LethalityThe mortality rate for adriamycin was studied after the
administration of melatonin. One group of rats (n = 33) servedas the control group (1% DMSO). The second group of rats(n = 28) received adriamycin alone (25 mg/kg IP). The third
group (n = 31) was injected with melatonin (10 mg/kg SC)1 hour before receiving adriamycin (25 mg/kg IP) and every12 hours thereafter for 6 days. The fourth group (n = 24)received only melatonin (10 mg/kg SC) every 12 hours for6 days. To assess the effect of melatonin on adriamycin-induced cardiac damage, rat hearts were used in the assay de-scribed below.
Modulation of Adriamycin-InducedBody-Weight Changes
The following groups of rats were used to evaluate thebody-weight changes and cardiotoxicity of adriamycin: control(n = 12); adriamycin alone (n = 11); melatonin alone (n = 8);and adriamycin plus melatonin (n = 9). In the adriamycingroup, 15 mg/kg adriamycin was injected IP as a single dose,whereas in the adriamycin plus melatonin group, the rats weretreated with melatonin (10 mg/kg SC) for 6 consecutive days,starting 2 days before adriamycin injection. To observe theeffects of melatonin on normal rats, melatonin (10 mg/kg SC)was administered daily to the melatonin-alone group for5 consecutive days. Body-weight measurements were performedevery other day. The numbers of deaths were recorded daily.Finally, the effect of melatonin on adriamycin-induced cardiacdamage was assayed as described below.
Hemodynamics, Heart Weights, and Levels ofCholesterol, Glucose, Insulin, andCreatine Kinase
Using a 2F micromanometer-tipped catheter (ModelSPR-407; Millar Instrument, Houston, TX) that was insertedthrough the right carotid artery, we measured the mean LVsystolic and diastolic pressures of 5 consecutive pressure cy-cles. The maximal rates of left ventricular pressure rise(+dP/dt) and decrease (2dP/dt) were measured. The heart wasexcised rapidly and subjected to retrograde perfusion witha Langendorff apparatus. At the completion of the heart rate(HR) measurements, the heart was rapidly divided into theright and left atria, and right and left ventricles, and each tissuewas weighed individually. To evaluate pulmonary edema, thelungs were also weighed. To determine the confounding rolesof cholesterol, glucose, insulin, and creatine kinase, we as-sayed blood samples from the aorta. The plasma cholesterollevels were determined enzymatically using kits from Hitachi7150 (Hitachi, Japan), and expressed as milligram per deciliterof serum. The plasma insulin concentrations were measuredby collecting 4-mL blood samples in test tubes that contained2% ethylenediaminetetraacetic acid (80 mL/ml of blood). Theblood samples were immediately centrifuged at 3000 3 g for10 minutes, and the plasma samples were stored at 270�Cuntil further analysis. Insulin was measured by the ultrasen-sitive rat enzyme immunoassay (Mercodia, Uppsala, Sweden).Creatine kinase was determined using kits purchased fromCal-Tech Diagnostics (Chino, CA).
Electron MicroscopyThe following groups of rats were used to evaluate the
ultrastructural alterations to the heart that resulted fromadriamycin treatment: control (n = 4); adriamycin alone (n = 5);adriamycin plus melatonin (n = 5); and melatonin alone (n = 4).
q 2005 Lippincott Williams & Wilkins 201
J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005 Melatonin Effects on Adriamycin

In the adriamycin group, a single dose of 15 mg/kg adriamycinwas injected IP, whereas in the adriamycin plus melatoningroup, the rats were treated with melatonin (10 mg/kg SC)for 6 consecutive days, starting 2 days before adriamycininjection. Six days after adriamycin injection, the hearts wereisolated, as described above, and fixed by perfusion with 0.1 Mphosphate buffer that contained 1.7% glutaraldehyde and1.6% paraformaldehyde through the aorta for 10 minutes ata pressure of 120 mm Hg. The hearts were then excised andstored in a fixative. Random samples (n = 11) of heart tissueswere postfixed in Epon 812. Ultrathin sections (50 to 70 nmthick) were stained with uranyl acetate and lead nitrate. Theultrastructure of the cardiac muscles was examined at mag-nification powers of 35000 and 315,000 in the electron mi-croscope (Jeol 1200 EX II; Jeol Ltd, Tokyo, Japan) andanalyzed using Image Pro-plus software (Media CyberneticsInc, Silver Spring, MD).
Preparation of Single Ventricular Myocytesand Heart Slices
Single ventricular myocytes were isolated from rat heartsby an enzymatic dissociation procedure, as described pre-viously.18 The isolated ventricular cells were stored in KBsolution at 4�C and used for measurements of cell length,width, and area as well as for the comet assay described below.
Thin (0.4- to 0.5-mm-thick) slices of rat heart were pre-pared as described previously,3 and the sections were storedin ice-cold modified Cross-Taggart medium that contained130 mM NaCl, 10 mM KCl, 1.5 mM CaCl2, 5 mM glucose,and 20 mM Tris-HCl (adjusted to pH 7.4 with NaOH). Theywere used for measurements of LDH release and lipid perox-idation described below.
Measurement of LDH Releaseand Lipid Peroxidation
For the measurement of LDH release, the heart sliceswere centrifuged at 1000 rpm for 5 minutes. The pellets werediscarded, and the supernatants were saved. LDH activity wasdetermined in the supernatants and incubation medium usingthe LDH Assay Kit (Asan Pharmaceutical Co, Ltd, Seoul,Korea). The final values are expressed as percentages of thecontrol value.
Lipid peroxidation was estimated by measuring the con-tent of malondialdehyde (MDA) according to the method ofOkawa et al.19 The final values are expressed as percentages ofthe control value.
Comet AssayThe comet assay was performed as described by Sestili
and Cantoni,20 with slight modifications. The assay works onthe principle that strand breakage of the supercoiled duplexDNA leads to the reduction of the size of the large molecule,and these strands can be stretched out by electrophoresis. Also,under highly alkaline conditions there is denaturation, un-winding of the duplex DNA, and expression of alkali-labilesites as single strand breaks. Comets form as the broken endsof the negatively charged DNA molecule become free tomigrate in the electric field toward the anode. Two principlesin the formation of the comet are that (1) DNA migration is
a function of both size and the number of broken ends of theDNA, and that (2) tail length increases with damage initiallyand then reaches a maximum that is dependent on the elec-trophoretic conditions, not the size of fragments. In eachexperiment, the DNA migration distance was calculated for atleast 50 randomly selected cells. For each nucleus in the cell,the total length of the image (including the nucleus and tail)and the diameter of the nucleus were measured in micrometers.The extent of DNA damage was assessed from the extent ofDNA migration (derived by subtracting the diameter of thenucleus from the total length of the image).
Colony-Forming AssayA clonogenic assay was used to measure the ability of
cells to form colonies. MDA-231 and NCI cells were culturedin 5% CO2 at 37�C in DMEM that was supplemented with10% FBS. The cells (n = 500) were seeded on 100-mm2 tissueculture dishes, following treatment with melatonin and/oradriamycin. The controls consisted of untreated cells. The cellswere exposed continuously for 7 days to the indicated con-centrations of melatonin and/or adriamycin; colonies were thenstained with crystal violet, and colonies that contained .50cells were counted. Each point represents the mean value 6SEM for 6 plates. The dose–response relationships for MDA-231 or NCI cell growth inhibition by adriamycin or melatoninwere fitted to the Hill equation using the least-squares method:
Relative cell growth ¼ 1=f1 þ ð½Drug�=KiÞng
where [Drug] is the concentration of adriamycin or melatonin,Ki is the concentration that gave half-maximal inhibition byadriamycin or melatonin, and n is the Hill coefficient.
Statistical AnalysisAll the values are expressed as mean 6 SEM. The dif-
ferences between the mean values were analyzed by the Studentt test, Dunnett procedure after analysis of variance or x2
analysis, ANOVA, and Kaplan-Meier (Mantel-Cox) methods.A P, 0.05 value was considered to be statistically significant.
RESULTS
Survival StudySingle-dose adriamycin treatment was dose-dependently
lethal for rats (Fig. 1A). The following survival rates werenoted: 100% for group 1 (control) and group 2 (5 mg/kgadriamycin); 85% for group 3 (15 mg/kg adriamycin); and14% for group 4 (25 mg/kg adriamycin). Figure 1B showshow the survival rate from IP administration of 25 mg/kgadriamycin was modified by SC injection of 10 mg/kg mel-atonin for 7 days. Melatonin treatment significantly attenuatedadriamycin-induced mortality from 86% to 20% (P , 0.05).On the other hand, the administration of melatonin aloneaccording to this regimen did not kill any of the rats.
Body Weight ChangesAdriamycin injection significantly decreased (216% of
the initial weight) the rat body weights by day 7 (Fig. 1C; P,0.05 versus control). In contrast, the body weights of the rats
202 q 2005 Lippincott Williams & Wilkins
Kim et al J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005
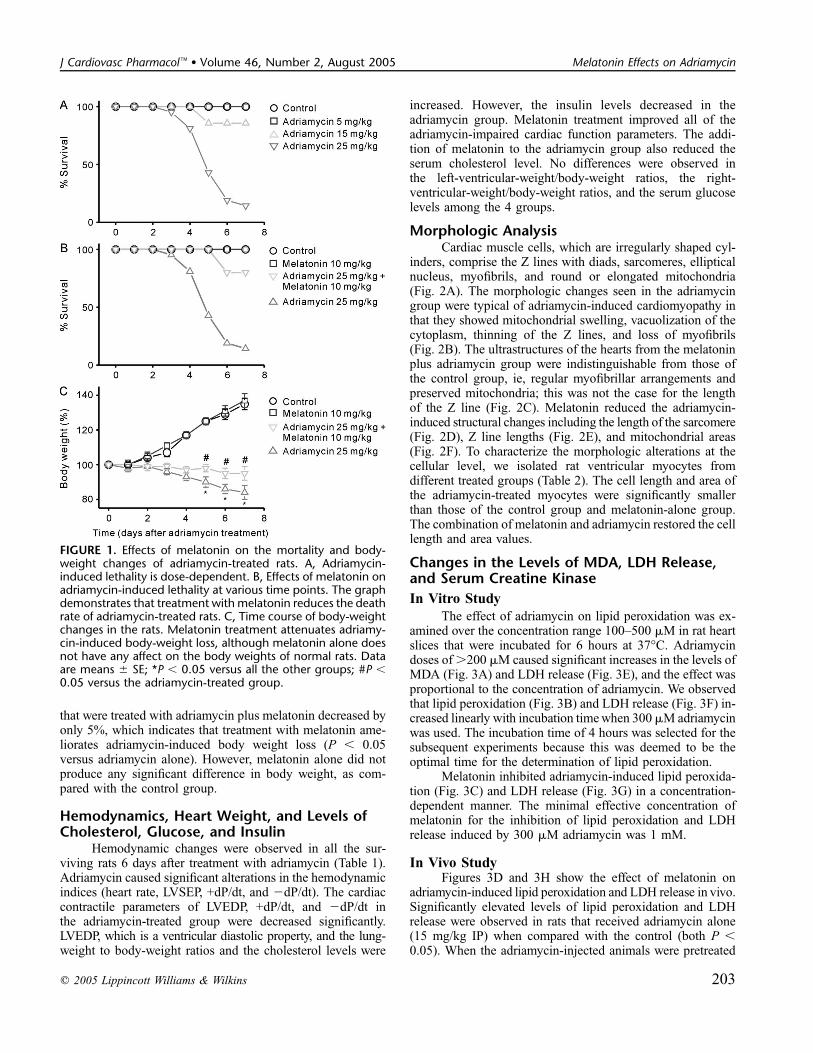
that were treated with adriamycin plus melatonin decreased byonly 5%, which indicates that treatment with melatonin ame-liorates adriamycin-induced body weight loss (P , 0.05versus adriamycin alone). However, melatonin alone did notproduce any significant difference in body weight, as com-pared with the control group.
Hemodynamics, Heart Weight, and Levels ofCholesterol, Glucose, and Insulin
Hemodynamic changes were observed in all the sur-viving rats 6 days after treatment with adriamycin (Table 1).Adriamycin caused significant alterations in the hemodynamicindices (heart rate, LVSEP, +dP/dt, and 2dP/dt). The cardiaccontractile parameters of LVEDP, +dP/dt, and 2dP/dt inthe adriamycin-treated group were decreased significantly.LVEDP, which is a ventricular diastolic property, and the lung-weight to body-weight ratios and the cholesterol levels were
increased. However, the insulin levels decreased in theadriamycin group. Melatonin treatment improved all of theadriamycin-impaired cardiac function parameters. The addi-tion of melatonin to the adriamycin group also reduced theserum cholesterol level. No differences were observed inthe left-ventricular-weight/body-weight ratios, the right-ventricular-weight/body-weight ratios, and the serum glucoselevels among the 4 groups.
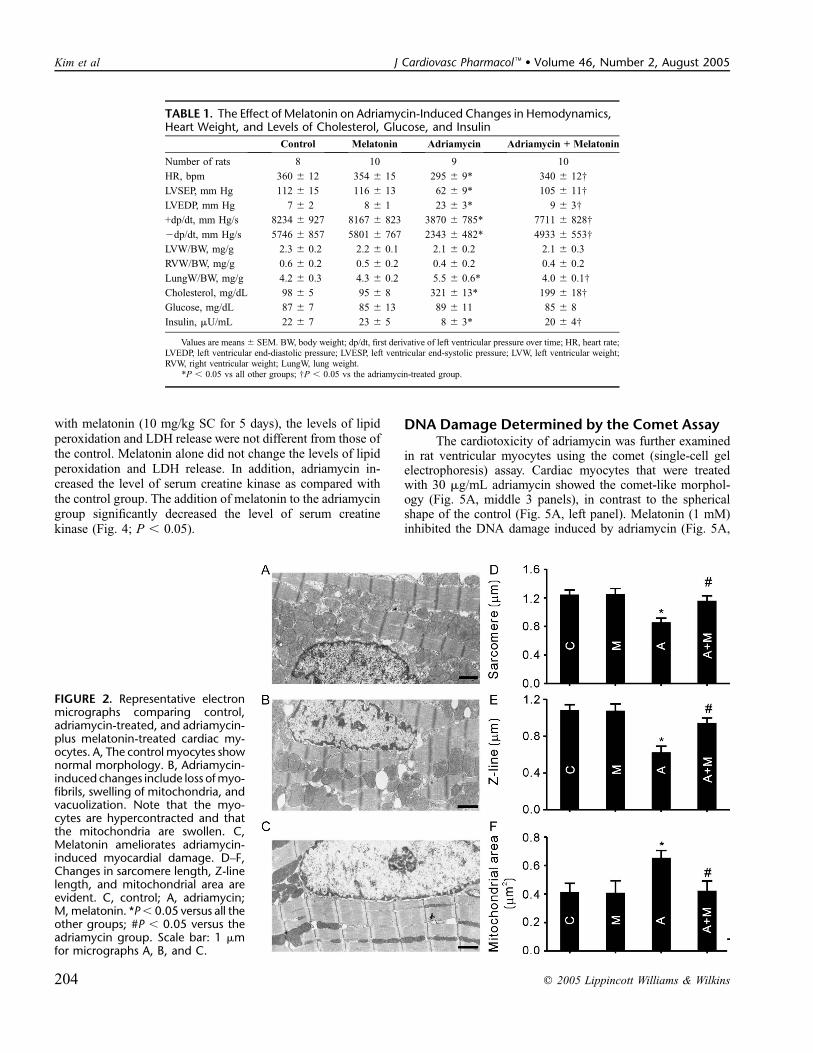
Morphologic AnalysisCardiac muscle cells, which are irregularly shaped cyl-
inders, comprise the Z lines with diads, sarcomeres, ellipticalnucleus, myofibrils, and round or elongated mitochondria(Fig. 2A). The morphologic changes seen in the adriamycingroup were typical of adriamycin-induced cardiomyopathy inthat they showed mitochondrial swelling, vacuolization of thecytoplasm, thinning of the Z lines, and loss of myofibrils(Fig. 2B). The ultrastructures of the hearts from the melatoninplus adriamycin group were indistinguishable from those ofthe control group, ie, regular myofibrillar arrangements andpreserved mitochondria; this was not the case for the lengthof the Z line (Fig. 2C). Melatonin reduced the adriamycin-induced structural changes including the length of the sarcomere(Fig. 2D), Z line lengths (Fig. 2E), and mitochondrial areas(Fig. 2F). To characterize the morphologic alterations at thecellular level, we isolated rat ventricular myocytes fromdifferent treated groups (Table 2). The cell length and area ofthe adriamycin-treated myocytes were significantly smallerthan those of the control group and melatonin-alone group.The combination of melatonin and adriamycin restored the celllength and area values.
Changes in the Levels of MDA, LDH Release,and Serum Creatine Kinase
In Vitro Study
The effect of adriamycin on lipid peroxidation was ex-amined over the concentration range 100–500 mM in rat heartslices that were incubated for 6 hours at 37�C. Adriamycindoses of .200 mM caused significant increases in the levels ofMDA (Fig. 3A) and LDH release (Fig. 3E), and the effect wasproportional to the concentration of adriamycin. We observedthat lipid peroxidation (Fig. 3B) and LDH release (Fig. 3F) in-creased linearly with incubation time when 300 mM adriamycinwas used. The incubation time of 4 hours was selected for thesubsequent experiments because this was deemed to be theoptimal time for the determination of lipid peroxidation.
Melatonin inhibited adriamycin-induced lipid peroxida-tion (Fig. 3C) and LDH release (Fig. 3G) in a concentration-dependent manner. The minimal effective concentration ofmelatonin for the inhibition of lipid peroxidation and LDHrelease induced by 300 mM adriamycin was 1 mM.
In Vivo StudyFigures 3D and 3H show the effect of melatonin on
adriamycin-induced lipid peroxidation and LDH release in vivo.Significantly elevated levels of lipid peroxidation and LDHrelease were observed in rats that received adriamycin alone(15 mg/kg IP) when compared with the control (both P ,0.05). When the adriamycin-injected animals were pretreated
FIGURE 1. Effects of melatonin on the mortality and body-weight changes of adriamycin-treated rats. A, Adriamycin-induced lethality is dose-dependent. B, Effects of melatonin onadriamycin-induced lethality at various time points. The graphdemonstrates that treatment with melatonin reduces the deathrate of adriamycin-treated rats. C, Time course of body-weightchanges in the rats. Melatonin treatment attenuates adriamy-cin-induced body-weight loss, although melatonin alone doesnot have any affect on the body weights of normal rats. Dataare means 6 SE; *P , 0.05 versus all the other groups; #P ,0.05 versus the adriamycin-treated group.
q 2005 Lippincott Williams & Wilkins 203
J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005 Melatonin Effects on Adriamycin
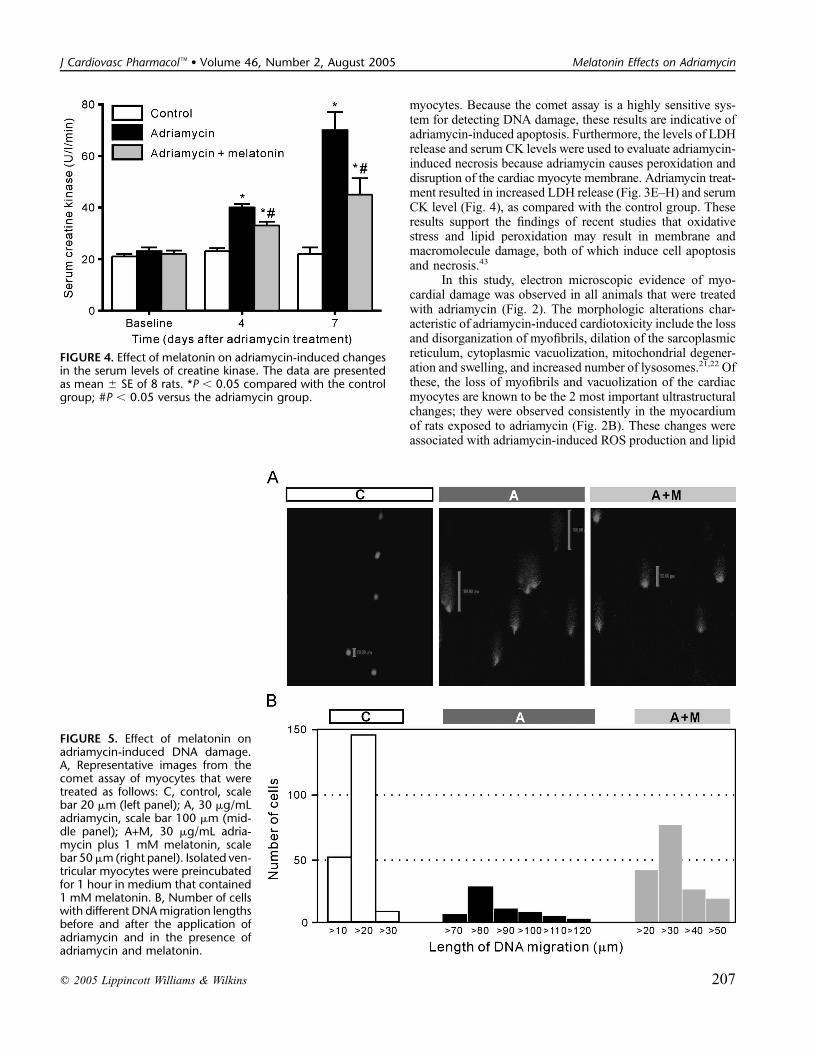
with melatonin (10 mg/kg SC for 5 days), the levels of lipidperoxidation and LDH release were not different from those ofthe control. Melatonin alone did not change the levels of lipidperoxidation and LDH release. In addition, adriamycin in-creased the level of serum creatine kinase as compared withthe control group. The addition of melatonin to the adriamycingroup significantly decreased the level of serum creatinekinase (Fig. 4; P , 0.05).
DNA Damage Determined by the Comet AssayThe cardiotoxicity of adriamycin was further examined
in rat ventricular myocytes using the comet (single-cell gelelectrophoresis) assay. Cardiac myocytes that were treatedwith 30 mg/mL adriamycin showed the comet-like morphol-ogy (Fig. 5A, middle 3 panels), in contrast to the sphericalshape of the control (Fig. 5A, left panel). Melatonin (1 mM)inhibited the DNA damage induced by adriamycin (Fig. 5A,
TABLE 1. The Effect of Melatonin on Adriamycin-Induced Changes in Hemodynamics,Heart Weight, and Levels of Cholesterol, Glucose, and Insulin
Control Melatonin Adriamycin Adriamycin + Melatonin
Number of rats 8 10 9 10
HR, bpm 360 6 12 354 6 15 295 6 9* 340 6 12†
LVSEP, mm Hg 112 6 15 116 6 13 62 6 9* 105 6 11†
LVEDP, mm Hg 7 6 2 8 6 1 23 6 3* 9 6 3†
+dp/dt, mm Hg/s 8234 6 927 8167 6 823 3870 6 785* 7711 6 828†
2dp/dt, mm Hg/s 5746 6 857 5801 6 767 2343 6 482* 4933 6 553†
LVW/BW, mg/g 2.3 6 0.2 2.2 6 0.1 2.1 6 0.2 2.1 6 0.3
RVW/BW, mg/g 0.6 6 0.2 0.5 6 0.2 0.4 6 0.2 0.4 6 0.2
LungW/BW, mg/g 4.2 6 0.3 4.3 6 0.2 5.5 6 0.6* 4.0 6 0.1†
Cholesterol, mg/dL 98 6 5 95 6 8 321 6 13* 199 6 18†
Glucose, mg/dL 87 6 7 85 6 13 89 6 11 85 6 8
Insulin, mU/mL 22 6 7 23 6 5 8 6 3* 20 6 4†
Values are means 6 SEM. BW, body weight; dp/dt, first derivative of left ventricular pressure over time; HR, heart rate;LVEDP, left ventricular end-diastolic pressure; LVESP, left ventricular end-systolic pressure; LVW, left ventricular weight;RVW, right ventricular weight; LungW, lung weight.
*P , 0.05 vs all other groups; †P , 0.05 vs the adriamycin-treated group.
FIGURE 2. Representative electronmicrographs comparing control,adriamycin-treated, and adriamycin-plus melatonin-treated cardiac my-ocytes. A, The control myocytes shownormal morphology. B, Adriamycin-induced changes include loss ofmyo-fibrils, swelling of mitochondria, andvacuolization. Note that the myo-cytes are hypercontracted and thatthe mitochondria are swollen. C,Melatonin ameliorates adriamycin-induced myocardial damage. D–F,Changes in sarcomere length, Z-linelength, and mitochondrial area areevident. C, control; A, adriamycin;M,melatonin. *P, 0.05 versus all theother groups; #P , 0.05 versus theadriamycin group. Scale bar: 1 mmfor micrographs A, B, and C.
204 q 2005 Lippincott Williams & Wilkins
Kim et al J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005

right panel). The control, adriamycin-treated, and adriamycin-plus melatonin-treated cells exhibited various DNA migrationdistances in several different experiments; the data were pooledand are expressed graphically in Figure 5B. The distribution ofthe control cells with respect to DNA migration distance wassimilar in all the experimental series. The DNA migrationdistance increased markedly in adriamycin-treated cells, andthis effect was prevented by melatonin treatment.
The Effect of Melatonin on the AntitumorActivity of Adriamycin
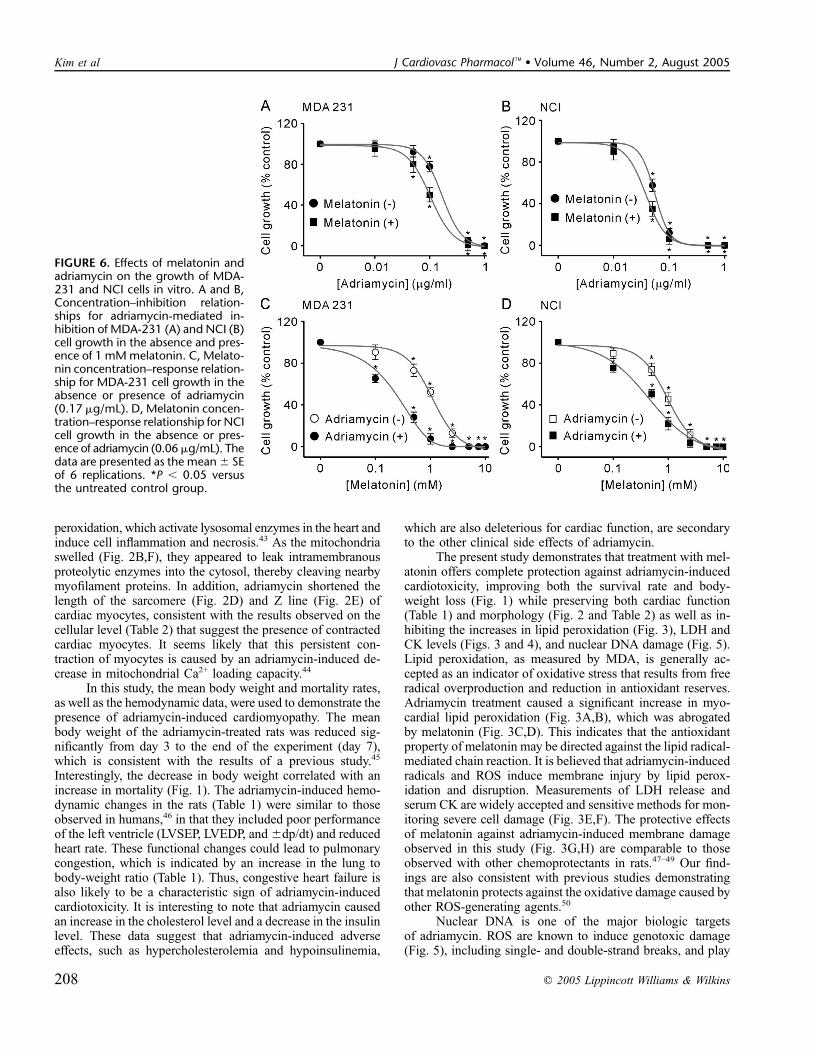
An initial series of experiments was performed todetermine the effect of different concentrations of adriamycin,within the range of 0.01 to 1 mg/mL, on in vitro culturedhuman breast cancer cell lines MDA-231 and NCI. Adriamy-cin inhibited the growth of both cell lines. Figure 6A shows thedose–response relationships for MDA-231 cell growth in-hibition by adriamycin in the absence and presence of 1 mMmelatonin. The adriamycin concentration at half-maximal inhi-bition (Ki) and the Hill coefficient were 0.17 6 0.03 mg/mLand 0.99 6 0.18, respectively, in the absence of melatonin and0.10 6 0.01 mg/mL and 0.99 6 0.11, respectively, in thepresence of melatonin (P, 0.05). For the NCI cells (Fig. 6B),treatment of cells with 1 mM melatonin changed the Ki ofadriamycin (0.06 6 0.01 mg/mL in the absence of melatoninand 0.03 6 0.01 mg/mL in the presence of melatonin; P ,0.05). No significant changes in the Hill coefficient wereobserved in the absence (0.98 6 0.16) and presence (0.98 60.14) of melatonin. These data indicate that melatonin in-creases the efficacy of adriamycin.
At concentrations of 0.5–10 mM, melatonin inducedsignificant reductions in cell growth. The melatonin concen-tration at half-maximal inhibition and the Hill coefficientwere 1.07 6 0.06 mM and 0.99 6 0.13, respectively, forthe MDA-231 cells (Fig. 6C) and 0.95 6 0.12 mM and 0.99 60.16, respectively, for the NCI cells (Fig. 6D). The mela-tonin concentration at half-maximal inhibition in the presenceof adriamycin was significantly less than that in the absence ofadriamycin (0.18 6 0.02 mM in the presence of 0.17 mg/mLadriamycin for the MDA-231 cells, Fig. 6C; and 0.486 0.09 mMin the presence of 0.06 mg/mL adriamycin for the NCI cells,Fig. 6D). No significant changes in the Hill coefficient wereobserved in the absence or presence of adriamycin (0.99 60.15 in the presence of 0.17 mg/mL adriamycin for the MDA-231 cells and 0.98 6 0.14 in the presence of 0.06 mg/mLadriamycin for the NCI cells).
DISCUSSIONPrevious studies have shown that adriamycin induces car-
diac dysfunction and causes severe cell damage.1–3 Adriamycincardiotoxicity is mainly the result of the formation of reactiveoxygen species (ROS).4 Although several ROS scavengershave been tested, they do not completely eliminate adriamycincardiotoxicity because the problems of high-density lipopro-tein lowering, unchanged mortality rate and weight loss,and potentiation of hematotoxicity persist, as well as the factthat these scavengers interfere with the antitumor properties ofadriamycin.21,22 Recently, it has been reported that melatoninis a potent antioxidant and ROS scavenger.11–14 On the basisof previous findings, it seems reasonable to assume that mela-tonin represents a valid candidate for reducing adriamycin-induced cardiotoxicity. Melatonin, besides being an efficientROS scavenger, is a lipophilic compound that readily crossescell membranes and distributes itself within the subcellularcompartments, where the antioxidant protection is required.Furthermore, it is well tolerated and safe after oral adminis-tration.23 In this study, melatonin was effective in attenuatingadriamycin-induced cardiotoxicity in rats. It is interesting tonote that melatonin itself inhibited the growth of human cancercells. In addition, the concomitant administration of melatoninimproved the antitumor action of adriamycin. The functionallyrefractory nature of adriamycin-induced cardiomyopathy andthe heart failure observed in humans have also been observedin rats.24,25 It seems likely that rats simulate most of theadriamycin-induced clinical and hemodynamic changes. Thus,in this study, we chose the rat as the animal model of adriamycin-induced cardiomyopathy.
Since the first report on adriamycin-induced cardiomy-opathy,26 several mechanisms have been proposed to explainthe development of this disease.4,27–41 It is generally acceptedthat mechanisms involving ROS are the main contributorsto adriamycin-mediated cardiotoxicity.35,36 In this study, in-creased ROS formation caused by adriamycin was evidencedby an increase in tissue MDA formation, which is a breakdownproduct of lipid peroxidation (Fig. 3A–D). The measurementof thiobarbituric acid–reactive substances (TBARs) is a widelyaccepted and sensitive method for monitoring oxidative stressin tissues. These results are consistent with those of previousstudies conducted by other investigators that suggest thatadriamycin-induced cardiotoxicity is associated with ROSgeneration and lipid peroxidation.4,38,42 The results of thecomet assay reported in the present study (Fig. 5) clearlydemonstrate adriamycin-induced DNA damage of cardiac
TABLE 2. Effects of Adriamycin and/or Melatonin on the Characteristics of RatVentricular Myocytes
Control Melatonin Adriamycin Adriamycin + Melatonin
Number of rats 6 6 6 6
Number of cells measured 95 90 160 110
Myocyte length, mm 137 6 17 134 6 13 72 6 15* 125 6 10†
Myocyte width, mm 23 6 3 22 6 3 20 6 2 22 6 2
Measured myocytes areas, mm2 2896 6 427 2872 6 387 1903 6 405* 2760 6 211†
Values are means 6 SEM. *P , 0.05 vs all other groups; †P , 0.05 vs the adriamycin-treated group.
q 2005 Lippincott Williams & Wilkins 205
J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005 Melatonin Effects on Adriamycin
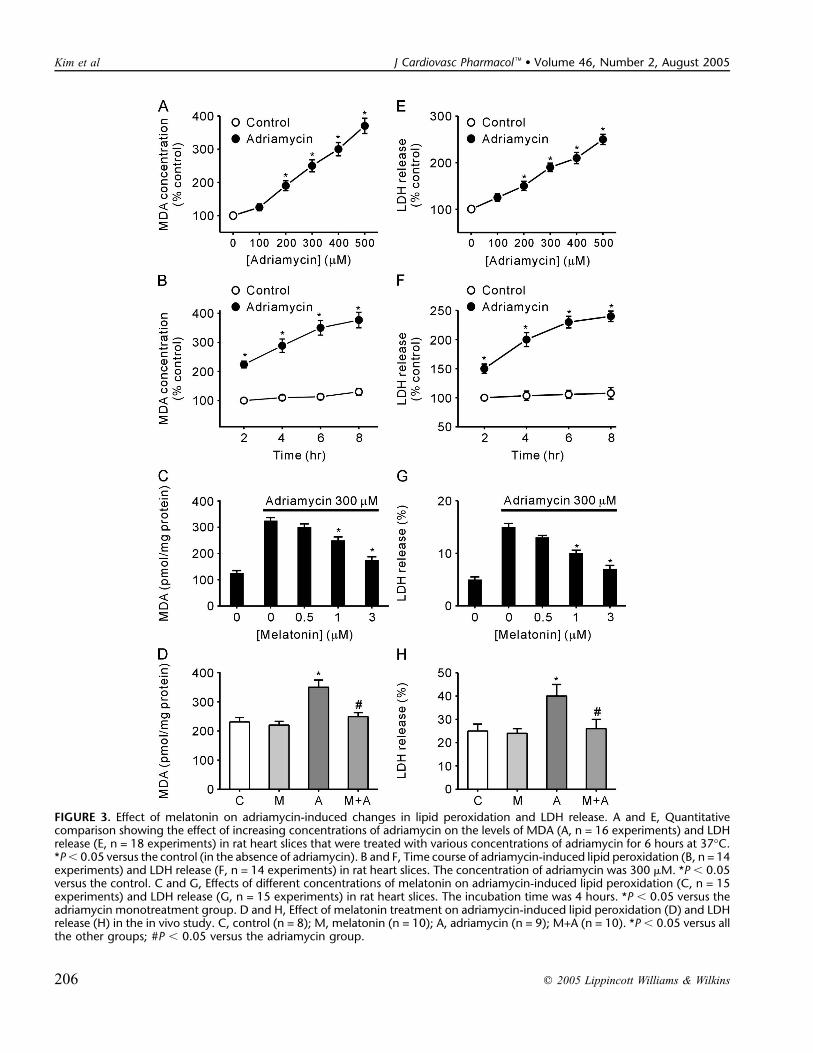
FIGURE 3. Effect of melatonin on adriamycin-induced changes in lipid peroxidation and LDH release. A and E, Quantitativecomparison showing the effect of increasing concentrations of adriamycin on the levels of MDA (A, n = 16 experiments) and LDHrelease (E, n = 18 experiments) in rat heart slices that were treated with various concentrations of adriamycin for 6 hours at 37�C.*P, 0.05 versus the control (in the absence of adriamycin). B and F, Time course of adriamycin-induced lipid peroxidation (B, n = 14experiments) and LDH release (F, n = 14 experiments) in rat heart slices. The concentration of adriamycin was 300 mM. *P , 0.05versus the control. C and G, Effects of different concentrations of melatonin on adriamycin-induced lipid peroxidation (C, n = 15experiments) and LDH release (G, n = 15 experiments) in rat heart slices. The incubation time was 4 hours. *P , 0.05 versus theadriamycin monotreatment group. D and H, Effect of melatonin treatment on adriamycin-induced lipid peroxidation (D) and LDHrelease (H) in the in vivo study. C, control (n = 8); M, melatonin (n = 10); A, adriamycin (n = 9); M+A (n = 10). *P , 0.05 versus allthe other groups; #P , 0.05 versus the adriamycin group.
206 q 2005 Lippincott Williams & Wilkins
Kim et al J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005
myocytes. Because the comet assay is a highly sensitive sys-tem for detecting DNA damage, these results are indicative ofadriamycin-induced apoptosis. Furthermore, the levels of LDHrelease and serum CK levels were used to evaluate adriamycin-induced necrosis because adriamycin causes peroxidation anddisruption of the cardiac myocyte membrane. Adriamycin treat-ment resulted in increased LDH release (Fig. 3E–H) and serumCK level (Fig. 4), as compared with the control group. Theseresults support the findings of recent studies that oxidativestress and lipid peroxidation may result in membrane andmacromolecule damage, both of which induce cell apoptosisand necrosis.43
In this study, electron microscopic evidence of myo-cardial damage was observed in all animals that were treatedwith adriamycin (Fig. 2). The morphologic alterations char-acteristic of adriamycin-induced cardiotoxicity include the lossand disorganization of myofibrils, dilation of the sarcoplasmicreticulum, cytoplasmic vacuolization, mitochondrial degener-ation and swelling, and increased number of lysosomes.21,22 Ofthese, the loss of myofibrils and vacuolization of the cardiacmyocytes are known to be the 2 most important ultrastructuralchanges; they were observed consistently in the myocardiumof rats exposed to adriamycin (Fig. 2B). These changes wereassociated with adriamycin-induced ROS production and lipid
FIGURE 4. Effect of melatonin on adriamycin-induced changesin the serum levels of creatine kinase. The data are presentedas mean 6 SE of 8 rats. *P , 0.05 compared with the controlgroup; #P , 0.05 versus the adriamycin group.
FIGURE 5. Effect of melatonin onadriamycin-induced DNA damage.A, Representative images from thecomet assay of myocytes that weretreated as follows: C, control, scalebar 20 mm (left panel); A, 30 mg/mLadriamycin, scale bar 100 mm (mid-dle panel); A+M, 30 mg/mL adria-mycin plus 1 mM melatonin, scalebar 50mm(right panel). Isolated ven-tricular myocytes were preincubatedfor 1 hour in medium that contained1 mM melatonin. B, Number of cellswith different DNAmigration lengthsbefore and after the application ofadriamycin and in the presence ofadriamycin and melatonin.
q 2005 Lippincott Williams & Wilkins 207
J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005 Melatonin Effects on Adriamycin
peroxidation, which activate lysosomal enzymes in the heart andinduce cell inflammation and necrosis.43 As the mitochondriaswelled (Fig. 2B,F), they appeared to leak intramembranousproteolytic enzymes into the cytosol, thereby cleaving nearbymyofilament proteins. In addition, adriamycin shortened thelength of the sarcomere (Fig. 2D) and Z line (Fig. 2E) ofcardiac myocytes, consistent with the results observed on thecellular level (Table 2) that suggest the presence of contractedcardiac myocytes. It seems likely that this persistent con-traction of myocytes is caused by an adriamycin-induced de-crease in mitochondrial Ca2+ loading capacity.44
In this study, the mean body weight and mortality rates,as well as the hemodynamic data, were used to demonstrate thepresence of adriamycin-induced cardiomyopathy. The meanbody weight of the adriamycin-treated rats was reduced sig-nificantly from day 3 to the end of the experiment (day 7),which is consistent with the results of a previous study.45
Interestingly, the decrease in body weight correlated with anincrease in mortality (Fig. 1). The adriamycin-induced hemo-dynamic changes in the rats (Table 1) were similar to thoseobserved in humans,46 in that they included poor performanceof the left ventricle (LVSEP, LVEDP, and 6dp/dt) and reducedheart rate. These functional changes could lead to pulmonarycongestion, which is indicated by an increase in the lung tobody-weight ratio (Table 1). Thus, congestive heart failure isalso likely to be a characteristic sign of adriamycin-inducedcardiotoxicity. It is interesting to note that adriamycin causedan increase in the cholesterol level and a decrease in the insulinlevel. These data suggest that adriamycin-induced adverseeffects, such as hypercholesterolemia and hypoinsulinemia,
which are also deleterious for cardiac function, are secondaryto the other clinical side effects of adriamycin.
The present study demonstrates that treatment with mel-atonin offers complete protection against adriamycin-inducedcardiotoxicity, improving both the survival rate and body-weight loss (Fig. 1) while preserving both cardiac function(Table 1) and morphology (Fig. 2 and Table 2) as well as in-hibiting the increases in lipid peroxidation (Fig. 3), LDH andCK levels (Figs. 3 and 4), and nuclear DNA damage (Fig. 5).Lipid peroxidation, as measured by MDA, is generally ac-cepted as an indicator of oxidative stress that results from freeradical overproduction and reduction in antioxidant reserves.Adriamycin treatment caused a significant increase in myo-cardial lipid peroxidation (Fig. 3A,B), which was abrogatedby melatonin (Fig. 3C,D). This indicates that the antioxidantproperty of melatonin may be directed against the lipid radical-mediated chain reaction. It is believed that adriamycin-inducedradicals and ROS induce membrane injury by lipid perox-idation and disruption. Measurements of LDH release andserum CK are widely accepted and sensitive methods for mon-itoring severe cell damage (Fig. 3E,F). The protective effectsof melatonin against adriamycin-induced membrane damageobserved in this study (Fig. 3G,H) are comparable to thoseobserved with other chemoprotectants in rats.47–49 Our find-ings are also consistent with previous studies demonstratingthat melatonin protects against the oxidative damage caused byother ROS-generating agents.50
Nuclear DNA is one of the major biologic targetsof adriamycin. ROS are known to induce genotoxic damage(Fig. 5), including single- and double-strand breaks, and play
FIGURE 6. Effects of melatonin andadriamycin on the growth of MDA-231 and NCI cells in vitro. A and B,Concentration–inhibition relation-ships for adriamycin-mediated in-hibition of MDA-231 (A) and NCI (B)cell growth in the absence and pres-ence of 1 mMmelatonin. C, Melato-nin concentration–response relation-ship for MDA-231 cell growth in theabsence or presence of adriamycin(0.17 mg/mL). D, Melatonin concen-tration–response relationship for NCIcell growth in the absence or pres-ence of adriamycin (0.06mg/mL). Thedata are presented as the mean6 SEof 6 replications. *P , 0.05 versusthe untreated control group.
208 q 2005 Lippincott Williams & Wilkins
Kim et al J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005

an important role in the pathogenesis of cytotoxicity. Adriamycin-induced free radicals can also induce DNA damage andcardiotoxicity. In the present study, we used the single-cell gelelectrophoresis (SCGE) assay, also known as the comet assay,to test the ability of melatonin to prevent DNA breaks inducedby adriamycin. The comet assay is a very sensitive method forthe detection of DNA damage. The administration of mela-tonin (1) decreased the extent of DNA damage (DNA migra-tion distance) and (2) increased the number of cells withdecreased levels of DNA damage (Fig. 5). These results are ofinterest in view of the proposed chemopreventive activity ofmelatonin.
Our results suggest that melatonin may be a useful ad-juvant to prevent adriamycin toxicity in cancer chemother-apies. The anticancer activity of adriamycin and lipid peroxidationare reportedly independent properties of adriamycin; andantioxidants, such as vitamin E and vitamin C, have been re-ported to reduce the lipid peroxidation and cardiotoxic activ-ities of adriamycin.6,38 In this context, it seems unlikely thatthe ameliorating effect of melatonin on adriamycin toxicityhas any affect on the antitumor activity of adriamycin. In thisstudy, we also examined whether the antioxidant effect ofmelatonin interfered with the antineoplastic effects of adriamycin.Surprisingly, melatonin itself had antiproliferative effects onhuman breast cancer cells. Furthermore, the direct antitumoreffects of melatonin were additive or synergistic with those ofadriamycin, which suggests the potential of this drug combi-nation for improving the efficacy of chemotherapy in terms ofboth survival and quality of life. These results are consistentwith previous reports that melatonin does not attenuate theantitumor activity of adriamycin under various in vitro andin vivo conditions.51 Indeed, in a large group of advanced cancerpatients who were treated with chemotherapy, melatonin in-creased the 1-year and 5-year survival rates as well as theoverall rate of tumor regression.52
In conclusion, the results of the present study dem-onstrate that melatonin protects against adriamycin-inducedlethality, body-weight loss, hemodynamic changes, lipid per-oxidation, severe cell damage, morphologic changes, andDNA damage in rat heart and that it also inhibits the growth ofthe human cancer cells. These findings support the results ofclinical trials that suggest the use of melatonin to preventadriamycin-induced cardiotoxicity and to enhance the antitu-mor activity of adriamycin.
REFERENCES1. Singal PK, Siveski-Iliskovic N, Hill M, et al. Combination therapy with
probucol prevents adriamycin-induced cardiomyopathy. J Mol CellCardiol. 1995;27:1055–1063.
2. Booser DJ, Hortobagyi GN. Anthracycline antibiotics in cancer therapy.Focus on drug resistance. Drugs. 1994;47:223–258.
3. Han J, Kim C, Kim N, et al. The protective effect of melatonin admin-istration against adriamycin-induced in rats. Kor J Physiol Pharmacol.2001;5:333–342.
4. Singal PK, Deally CM, Weinberg LE. Subcellular effects of adriamycin inthe heart: a concise review. J Mol Cell Cardiol. 1987;19:817–828.
5. Wang YM, Madanat FF, Kimball JC, et al. Effect of vitamin E againstadriamycin-induced toxicity in rabbits. Cancer Res. 1980;40:1022–1027.
6. Fujita K, Shinpo K, Yamada K, et al. Reduction of adriamycin toxicity byascorbate in mice and guinea pigs. Cancer Res. 1982;42:309–316.
7. Tesoriere L, Ciaccio M, Valenza M, et al. Effect of vitamin A admin-istration on resistance of rat heart against doxorubicin-induced cardio-toxicity and lethality. J Pharmacol Exp Ther. 1994;269:430–436.
8. Brzezinski A. Melatonin in humans. N Engl J Med. 1997;336:186–195.9. Blask D. Melatonin in oncology. In Yu HS, Reiter RJ (eds): Melatonin.
Boca Raton, FL: CRC Press, 1993:447–475.10. Maestroni GJ. The immunoneuroendocrine role of melatonin. J Pineal
Res. 1993;14:1–10.11. Marshall KA, Reiter RJ, Poeggeler B, et al. Evaluation of the antioxidant
activity of melatonin in vitro. Free Radic Biol Med. 1996;21:307–315.12. Antolin RL, Rodriquez C, Saints RM, et al. Neurohormone melatonine
prevents cell damage: effects of gene expression for antioxidant enzymes.FASEB J. 1996;10:882–890.
13. Reiter RJ, Tang L, Garcia JJ, et al. Pharmacological actions of melatoninin oxygen radical pathophysiology. Life Sci. 1997;60:2255–2271.
14. Poeggeler B, Saarela S, Reiter RJ, et al. Melatonin—a highly potentendogenous radical scavenger and electron donor: new aspects of theoxidation chemistry of this indole accessed in vitro. Ann N Y Acad Sci.1994;738:419–420.
15. Reiter RJ. Oxidative damage in the central nervous system: protection bymelatonin. Prog Neurobiol. 1998;56:339–384.
16. Reiter RJ, Tan DX, Kim SJ, et al. Melatonin as a pharmacological agentagainst oxidative damage to lipids and DNA. Proc West Pharmacol Soc.1998;41:229–236.
17. Reiter RJ, Tan DX, Qi W. A brief survey of observations and mechanismsrelated to suppression of oxygen toxicity by melatonin. Acta PharmacolSin. 1998;19:575–581.
18. Han J, Kim N, Joo H, et al. ATP-sensitive K+ channel activation by nitricoxide and protein kinase G in rabbit ventricular myocytes. Am J PhysiolHeart Circ Physiol. 2002;283:H1545–H1554.
19. Okawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animals andtissues by thiobarbituric aid reaction. Anal Biochem. 1979;95:351–358.
20. Sestili P, Cantoni O. Osmotically driven radical diffusion of single-stranded DNA fragments on an agarose bed as a convenient measure ofDNA strand scission. Free Radicals Biol Med. 1999;26:1019–1026.
21. Herman EH, Zhang J, Ferrans VJ. Comparison of the protective effects ofdesferrioxamine and ICRF-187 against doxorubicin-induced toxicity inspontaneously hypertensive rats. Cancer Chemother Pharmacol. 1994;35:93–100.
22. Koning J, Palmer P, Franks CR, et al. Cardioxane–ICRF-187 towardsanticancer drug specificity through selective toxicity reduction. CancerTreat Rev. 1991;18:1–19.
23. Ivanenko A, Crabtree VM, Tauman R, et al. Melatonin in children andadolescents with insomnia: a retrospective study. Clin Pediatr (Phila).2003;42:51–58.
24. Weinberg LE, Singal PK. Refractory heart failure and age-related dif-ferences in adriamycin-induced myocardial changes in rats. Can J PhysiolPharmacol. 1987;65:1957–1965.
25. Tong J, Ganguly PK, Singal PK. Myocardial adrenergic changes at twostages of heart failure due to adriamycin treatment in rats. Am J Physiol.1991;260:H909–H916.
26. Bonadonna G, Monfardini S, De Lena M, et al. Phase I and preliminaryphase II evaluation of adriamycin (NSC 123127). Cancer Res. 1970;30:2572–2582.
27. Buja LM, Ferrans VJ, Mayer RJ, et al. Cardiac ultrastructural changesinduced by daunorubicin therapy. Cancer. 1973;32:771–788.
28. Arena E, D’Alessandre N, Dusonchet L, et al. DNA, RNA and proteinsynthesis in heart, liver and brain of mice treated with daunorubicin andadriamycin. Int Res Commun Systemic Med Sci. 1974;2:1053–1061.
29. Bristow MR, Sageman WS, Scott RH, et al. Acute and chronic car-diovascular effects of doxorubicin in the dog: the cardiovascular phar-macology of drug-induced histamine release. J Cardiovasc Pharmacol.1980;2:487–515.
30. Gosalvez M, van Rossum GD, Blanco MF. Inhibition of sodium-potassium-activated adenosine 5#-triphosphatase and ion transport byadriamycin. Cancer Res. 1979;39:257–261.
31. Singal PK, Segstro RJ, Singh RP, et al. Changes in lysosomal morphologyand enzyme activities during the development of adriamycin-inducedcardiomyopathy. Can J Cardiol. 1985;1:139–147.
32. Singal PK, Pierce GN. Adriamycin stimulates low-affinity Ca2+ bindingand lipid peroxidation but depresses myocardial function. Am J Physiol.1986;250:H419–H425.
q 2005 Lippincott Williams & Wilkins 209
J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005 Melatonin Effects on Adriamycin

33. Singal PK, Panagia V. Direct effects of adriamycin on the rat heart sar-colemma. Res Commun Chem Pathol Pharmacol. 1984;43:67–77.
34. Olson RD, MacDonald JS, vanBoxtel CJ, et al. Regulatory role of glu-tathione and soluble sulfhydryl groups in the toxicity of adriamycin.J Pharmacol Exp Ther. 1980;215:450–454.
35. Kalyanaraman B, Perez-Reyes E, Mason RP. Spin-trapping and directelectron spin resonance investigations of the redox metabolism of quinoneanticancer drugs. Biochim Biophys Acta. 1980;630:119–130.
36. Doroshow JH. Anthracycline antibiotic-stimulated superoxide, hydrogenperoxide, and hydroxyl radical production by NADH dehydrogenase.Cancer Res. 1983;43:4543–4551.
37. Siveski-Iliskovic N, Kaul N, Singal PK. Probucol promotes endogenousantioxidants and provides protection against adriamycin-induced cardio-myopathy in rats. Circulation. 1994;89:2829–2835.
38. Myers CE, McGuire WP, Liss RH, et al. Adriamycin: the role of lipidperoxidation in cardiac toxicity and tumor response. Science. 1977;197:165–167.
39. Odom AL, Hatwig CA, Stanley JS, et al. Biochemical determinations ofadriamycin toxicity in mouse liver, heart and intestine. Biochem Pharmacol.1992;43:831–836.
40. Kumar D, Kirshenbaum L, Li T, et al. Apoptosis in isolated adult car-diomyocytes exposed to adriamycin. Ann N Y Acad Sci. 1999;874:156–168.
41. Kaul N, Siveski-Iliskovic N, Hill M, et al. Free radicals and the heart.J Pharmacol Toxicol Methods. 1993;30:55–67.
42. Iliskovic N, Hasinoff BB, Malisza KL, et al. Mechanisms of beneficialeffects of probucol in adriamycin cardiomyopathy. Mol Cell Biochem.1999;196:43–49.
43. Kumar D, Jugdutt BI. Apoptosis and oxidants in the heart. J Lab ClinMed. 2003;142:288–297.
44. Zhou S, Starkov A, Froberg MK, et al. Cumulative and irreversible cardiacmitochondrial dysfunction induced by doxorubicin. Cancer Res. 2001;61:771–777.
45. Xu MF, Ho S, Qian ZM, et al. Melatonin protects against cardiac toxicityof doxorubicin in rat. J Pineal Res. 2001;31:301–307.
46. de Graaf H, Dolsma WV, Willemse PH, et al. Cardiotoxicity fromintensive chemotherapy combined with radiotherapy in breast cancer. Br JCancer. 1997;76:943–945.
47. Mohamed HE, El-Swefy SE, Hagar HH. The protective effect of glu-tathione administration on adriamycin-induced acute cardiac toxicity inrats. Pharmacol Res. 2000;42:115–121.
48. Mostafa MG, Mima T, Ohnishi ST, et al. S-Allylcysteine amelioratesdoxorubicin toxicity in the heart and liver in mice. Planta Med. 2000;66:148–151.
49. Nagi MN, Mansour MA. Protective effect of thymoquinone againstdoxorubicin-induced cardiotoxicity in rats: a possible mechanism ofprotection. Pharmacol Res. 2000;41:283–289.
50. Qi W, Reiter RJ, Tan DX, et al. Inhibitory effects of melatonin on ferricnitrilotriacetate-induced lipid peroxidation and oxidative DNA damage inthe rat kidney. Toxicology. 1999;139:81–91.
51. Granzotto M, Rapozzi V, Decorti G, et al. Effects of melatonin ondoxorubicin cytotoxicity in sensitive and pleiotropically resistant tumorcells. J Pineal Res. 2001;31:206–213.
52. Lissoni P, Chilelli M, Villa S, et al. Five years survival in metastatic non–small cell lung cancer patients treated with chemotherapy alone or che-motherapy and melatonin: a randomized trial. J Pineal Res. 2003;35:12–15.
210 q 2005 Lippincott Williams & Wilkins
Kim et al J Cardiovasc Pharmacol� � Volume 46, Number 2, August 2005
Copyright © 2022 FDOKUMEN




















