
Melatonin prevents testosterone-induced suppression of immune parameters and splenocyte...
7
General and Comparative Endocrinology 141 (2005) 226–232 www.elsevier.com/locate/ygcen 0016-6480/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2005.01.005 Melatonin prevents testosterone-induced suppression of immune parameters and splenocyte proliferation in Indian tropical jungle bush quail, Perdicula asiatica Shiv Shankar Singh, Chandana Haldar ¤ Pineal Research Lab, Department of Zoology, Banaras Hindu University, Varanasi 221 005, India Received 5 June 2004; revised 22 November 2004; accepted 11 January 2005 Abstract During reproductive inactive phase (winter months) peripheral high melatonin and low testosterone was noted in male Indian tropical bird, Perdicula asiatica. During this phase exogenous testosterone (1 mg/100 g Bwt./day) suppressed the immune parameters [spleen weight, total leukocyte count (TLC), lymphocyte count (LC), and percent stimulation ratio (% SR)] and depleted splenic cel- lular density, while melatonin (25 g/100 g Bwt./day) restored the immune parameters and splenic cellular density to the level of con- trol birds. In vitro suppression of splenocyte proliferation by testosterone (3 ng/ml) was also reversed by melatonin (500 pg/ml) supplementation. Therefore, high melatonin in circulation during the reproductive inactive phase acts as immunostimulator and is of high adaptive signiWcance to this bird for survival during the adverse conditions of season and reproductive phase. 2005 Elsevier Inc. All rights reserved. Keywords: Melatonin; Testosterone; Splenocyte; Blastogenic response; In vivo; In vitro 1. Introduction The eVect of gonadal steroids on immune function was best illustrated by the fact of profound sex diVer- ences in immune function of vertebrates (Schuurs and Verheul, 1990). Males generally exhibit lower immune responses than females (Schuurs and Verheul, 1990; Zuk and Mckeam, 1996). The ability of gonadal steroids to alter the course of disease and autoimmunodisease has been suspected in the clinical setting for more than 100 years, while animal model has provided more durable evidence of the immunomodulatory eVects of the gonadal steroids (Olsen and Kovacs, 1996). The maintenance of high testosterone throughout the breeding season may have adverse eVects on physical condition and therefore longevity, which is an important component of individual Wtness (Lochmiller and Deer- enberg, 2000). Testosterone suppresses immune function and enhances female attractive characters in male dark eyed Jankus (Folstad and Karter, 1992). A negative rela- tionship between testosterone titers and parasite load was found in barn swallows (Hirundo rustica; Saino and Moller, 1994), but no such relationship was detected in red winged bird (Agelains phoeniceus; Weatherhead et al., 1993). High testosterone in circulation reduced antibody production in adult barn swallows, house spar- rows Passer domesticus, and chickens (Evans et al., 2000; Saino et al., 1995; Verhulst et al., 1999) but not in juve- nile black-headed gull (Larus ridibundus; Ros et al., 1997) or adult red winged black bird (Hasselquist et al., 1999), suggesting a species-speciWc eVect. It has been well established that neurohormone mela- tonin is an immunomodulator regulating the develop- ment, diVerentiation, and function of lymphoid tissues (Lissoni, 1999; Maestroni, 1998; Reiter et al., 2000; Skwarlo-Sonta, 1999) in mammals and birds. Indeed ¤ Corresponding author. E-mail addresses: [email protected], chaldar2001@yahoo. com (C. Haldar).
Transcript of Melatonin prevents testosterone-induced suppression of immune parameters and splenocyte...

General and Comparative Endocrinology 141 (2005) 226–232
www.elsevier.com/locate/ygcen
Melatonin prevents testosterone-induced suppression of immune parameters and splenocyte proliferation in
Indian tropical jungle bush quail, Perdicula asiatica
Shiv Shankar Singh, Chandana Haldar ¤
Pineal Research Lab, Department of Zoology, Banaras Hindu University, Varanasi 221 005, India
Received 5 June 2004; revised 22 November 2004; accepted 11 January 2005
Abstract
During reproductive inactive phase (winter months) peripheral high melatonin and low testosterone was noted in male Indiantropical bird, Perdicula asiatica. During this phase exogenous testosterone (1 mg/100 g Bwt./day) suppressed the immune parameters[spleen weight, total leukocyte count (TLC), lymphocyte count (LC), and percent stimulation ratio (% SR)] and depleted splenic cel-lular density, while melatonin (25 �g/100 g Bwt./day) restored the immune parameters and splenic cellular density to the level of con-trol birds. In vitro suppression of splenocyte proliferation by testosterone (3 ng/ml) was also reversed by melatonin (500 pg/ml)supplementation. Therefore, high melatonin in circulation during the reproductive inactive phase acts as immunostimulator and is ofhigh adaptive signiWcance to this bird for survival during the adverse conditions of season and reproductive phase. 2005 Elsevier Inc. All rights reserved.
Keywords: Melatonin; Testosterone; Splenocyte; Blastogenic response; In vivo; In vitro
1. Introduction component of individual Wtness (Lochmiller and Deer-
The eVect of gonadal steroids on immune functionwas best illustrated by the fact of profound sex diVer-ences in immune function of vertebrates (Schuurs andVerheul, 1990). Males generally exhibit lower immuneresponses than females (Schuurs and Verheul, 1990; Zukand Mckeam, 1996). The ability of gonadal steroids toalter the course of disease and autoimmunodisease hasbeen suspected in the clinical setting for more than 100years, while animal model has provided more durableevidence of the immunomodulatory eVects of thegonadal steroids (Olsen and Kovacs, 1996).
The maintenance of high testosterone throughout thebreeding season may have adverse eVects on physicalcondition and therefore longevity, which is an important
¤ Corresponding author.E-mail addresses: [email protected], chaldar2001@yahoo.
com (C. Haldar).
0016-6480/$ - see front matter 2005 Elsevier Inc. All rights reserved.doi:10.1016/j.ygcen.2005.01.005
enberg, 2000). Testosterone suppresses immune functionand enhances female attractive characters in male darkeyed Jankus (Folstad and Karter, 1992). A negative rela-tionship between testosterone titers and parasite loadwas found in barn swallows (Hirundo rustica; Saino andMoller, 1994), but no such relationship was detected inred winged bird (Agelains phoeniceus; Weatherheadet al., 1993). High testosterone in circulation reducedantibody production in adult barn swallows, house spar-rows Passer domesticus, and chickens (Evans et al., 2000;Saino et al., 1995; Verhulst et al., 1999) but not in juve-nile black-headed gull (Larus ridibundus; Ros et al.,1997) or adult red winged black bird (Hasselquist et al.,1999), suggesting a species-speciWc eVect.
It has been well established that neurohormone mela-tonin is an immunomodulator regulating the develop-ment, diVerentiation, and function of lymphoid tissues(Lissoni, 1999; Maestroni, 1998; Reiter et al., 2000;Skwarlo-Sonta, 1999) in mammals and birds. Indeed

S.S. Singh, C. Haldar / General and Comparative Endocrinology 141 (2005) 226–232 227
diurnal and seasonal changes in immune function arethought to reXect directly the changes in pineal-melato-nin production (Haldar et al., 2001; Moore and Siopes,2000; Nelson and Drazen, 1999). Further, immune sys-tem varies according to reproductive status as well andreproductive hormone plays important role in the regu-lation of immune system in mammals (Bilbo and Nelson,2001). Cell mediated immune responses are also dimor-phic. Further, neutralizing or protective antibodies ofthe IgG class and IgM level also show the most signiW-cant sexual dimorphism.
The immunocompetence handicap hypothesis sug-gested by Folstad and Karter (1992) for testosteronepredicts a trade-oV between: (i) suppressive inXuences oftestosterone on immune function and (ii) enhancinginXuences of testosterone on characters that are attrac-tive to females. To note the eVect of exogenous testoster-one on immune parameters when peripheral level oftestosterone is low in the subject, the present investiga-tion was undertaken during reproductively inactivephase for a better understanding of immunomodulatoryrole of melatonin in relation to testosterone in a tropicalseasonally breeding bird, male Indian jungle bush quail,Perdicula asiatica.
2. Materials and methods
All experiments were conducted in accordance withinstitutional practice and within the framework ofrevised Animals (ScientiWc Procedures) Act of 2002 ofGovernment of India on Animal Welfare.
Indian jungle bush quail, P. asiatica belonging to theorder Galliformes and family Phasianidae is a semi-migratory wild bird well distributed at northern regionof India. Adult male quails were collected from the vicin-ity of Varanasi (latitude 25°18�N, longitude 83°01�E)during reproductively inactive phase (winter, November;11L:13D). They were acclimatized for 10–15 days to lab-oratory conditions (in an aviary exposed to natural con-ditions; 11L:13D). The birds were fed with millet seed(Pennisetum typhoides) and water ad libitum. After accli-matization birds were randomly divided into two sets.
Experiment I: Birds of Wrst set were divided into Wvegroups containing seven healthy male birds in each. Twocontrol groups were taken, Wrst control group birds weregiven ethanolic saline (0.01% ethanol in 0.85% NaClsolution; 0.1 ml/day) and second were given olive oil(0.1 ml/day). Third group birds received testosterone(Sigma, St. Louis, USA) at a concentration of 1 mg/100 gof body mass (testosterone was dissolved in olive oil)intramuscularly (im) at 14.00–14.50 h (Chaturvedi andThapliyal, 1978). Fourth group birds received testoster-one and melatonin simultaneously. Melatonin (Sigma,St. Louis, USA) at a concentration of 25 �g/100g of bodymass (melatonin was dissolved in ethanolic saline; Sud-
hakumari, 1995) was given subcutaneously (sc) to Wfthgroup of birds at 16.30–17.00 h (Haldar and Ghosh,1988). All treatments were carried out for 15 consecutivedays. After the last day of injection the birds weresacriWced by decapitation during the dark phase (20.00–21.00 h) of light/dark cycle and blood was taken in hepa-rinized tube. Spleen was dissected out on ice, weighed onSartorius balance, and processed for culture to observethe blastogenic response and histological observations.
Experiment II: Birds of second set were sacriWced bydecapitation during dark phase (20.00–21.00 h) of light/dark cycle. Spleen were taken out and processed forsplenocyte culture as described in Section 2.2. Fourgroups of culture sets were prepared having Wve replica-tion in each. First group was taken as control. Secondsets of culture tubes were supplemented with testoster-one (3 ng/ml) (Bilbo and Nelson, 2001) in each tube (tes-tosterone was dissolved in DMSO and diluted withculture medium RPMI 1640). Third sets of culture tubeswere supplemented with testosterone and melatoninboth while fourth sets of culture tubes were supple-mented with melatonin (500 pg/ml) (Drazen et al., 2001)in each tube (melatonin was Wrst dissolved in a drop ofethyl alcohol and then diluted with culture medium forrequired concentration).
2.1. Haematological parameters
Blood Wlm was prepared on glass slides for lympho-cyte count. Total leukocyte count was done in a Neu-bauer hemocytometer. Lymphocyte count was donefollowing Leishman’s staining.
2.2. Reagent and culture medium for blastogenic response
Tissue culture medium RPMI-1640 and all otherchemicals were purchased from Sigma Chemicals, St.Louis, USA. The culture medium was supplementedwith 100 U/ml penicillin, 100 �g/ml streptomycin, and10% fetal calf serum. Spleen was processed for prepara-tion of single cell suspensions. The number of cells wasadjusted to 1 £ 106 cells/ml in complete medium. Twomilliliters of cell suspension was placed in duplicate cul-ture tubes and kept at 37 °C in a 5% CO2 incubator(Hera Cell, Heraus, Germany) for 72 h. Blastogenicresponse was measured in terms of [3H]thymidine (spe-ciWc activity 8.9 Ci/mM) uptake against stimulation byCon A of the splenocytes (Pauly and Sokal, 1972).
2.3. Histological parameters
The spleen was dissected out and Wxed in Bouin’sXuid. Sections of 5 �m were cut and stained with hema-toxylin and eosin. Photographs of each group of spleenwere taken with the help of Leitz MPV3 microscopeunder 40£ magniWcation.

228 S.S. Singh, C. Haldar / General and Comparative Endocrinology 141 (2005) 226–232
2.4. Statistical analysis
Statistical analysis of the data was performed withone-way ANOVA followed by Student Newman–Keulsmultiple range test. The diVerences were consideredsigniWcant when P < 0.05. Since, there was no signiW-
cance diVerence in both the control group (ethanolicsaline and olive treated) data therefore, only one con-trol data (ethanolic saline treated) was presented in allhistogram.
3. Result
3.1. Spleen weight
SigniWcant (P < 0.01) decrease in spleen weight wasnoted in testosterone treated bird when compared withcontrol birds. Melatonin treatment along with testoster-one restored the spleen weight up to control level. How-ever, melatonin (alone) treatment to the birds showedsigniWcant (P < 0.01) increase of spleen weight whencompared with control group (Fig. 1).
3.2. Total leukocyte count and lymphocyte count
SigniWcant (P < 0.01) decrease of total leukocytecount and lymphocyte count (TLC and LC) wasnoted in testosterone treated bird when comparedwith control bird. Melatonin treatment along withtestosterone antagonized the testosterone-inducedsuppression of TLC and LC and restored up to con-trol level. Whereas, melatonin treatment signiWcantly(P < 0.01) increased the total leukocyte and lym-phocyte count when compared with control group(Figs. 2A and B).
Fig. 1. EVect of testosterone and melatonin administration on spleenweight of Indian jungle bush quail, P. asiatica during reproductivelyinactive phase (November–January). Histograms representmeans § SE, n D 5 for each group within this experiment. Con, con-trol; Mel, melatonin; and T, testosterone. **P < 0.01 Con vs T, Con vsMel, Con vs Mel + T, and T vs Mel + T.
3.3. Blastogenic response of splenocytes and percent stimulation ratio
3.3.1. Experiment ITestosterone treatment signiWcantly (P <0.01)
decreased the basal and mitogen Con A-induced blasto-genic response of splenocytes as well as percent stimula-tion ratio (% SR). Restoration of these parameters up tocontrol level was noted in melatonin along with testoster-one treated birds. However, melatonin treatment signiW-
cantly (P < 0.01) increased the basal as well as mitogenCon A-induced blastogenic response of splenocytes and %SR when compared with control group. (Figs. 3A and B).
3.3.2. Experiment IITestosterone supplementation in culture signiWcantly
(P < 0.01) suppressed the basal as well as mitogen ConA-induced blastogenic response and % SR when com-pared with control culture. Melatonin supplementationalong with testosterone restored the basal as well asmitogen Con A-induced blastogenic response and per-cent stimulation ratio as was observed in control culture.Melatonin supplementation to the splenocyte culturesigniWcantly (P < 0.01) increased basal and mitogen Con
Fig. 2. EVect of testosterone and melatonin administration on (A) totalleukocyte count and (B) lymphocyte count of Indian jungle bush quail,P. asiatica during reproductively inactive phase (November–January).Histograms represent means § SE, n D 5 for each group within thisexperiment. Con, control; Mel, melatonin; and T, testosterone.**P < 0.01 Con vs T, Con vs Mel, Con vs Mel + T, and T vs Mel + T.
S.S. Singh, C. Haldar / General and Compar
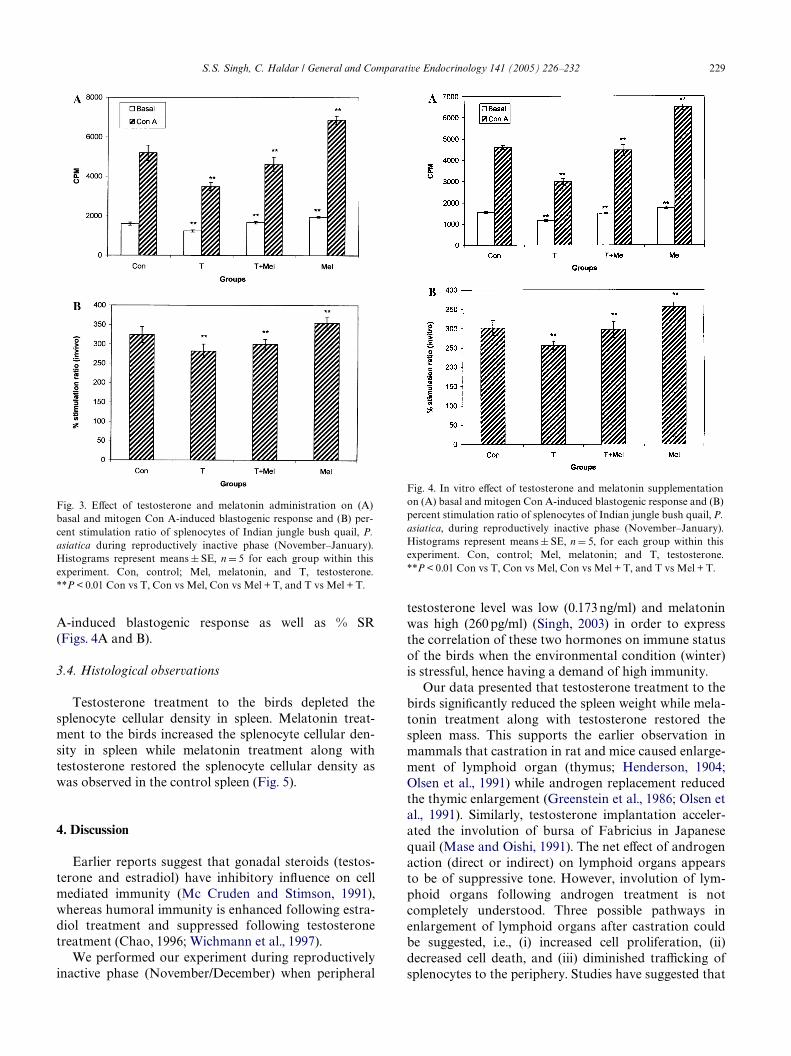
A-induced blastogenic response as well as % SR(Figs. 4A and B).
3.4. Histological observations
Testosterone treatment to the birds depleted thesplenocyte cellular density in spleen. Melatonin treat-ment to the birds increased the splenocyte cellular den-sity in spleen while melatonin treatment along withtestosterone restored the splenocyte cellular density aswas observed in the control spleen (Fig. 5).
4. Discussion
Earlier reports suggest that gonadal steroids (testos-terone and estradiol) have inhibitory inXuence on cellmediated immunity (Mc Cruden and Stimson, 1991),whereas humoral immunity is enhanced following estra-diol treatment and suppressed following testosteronetreatment (Chao, 1996; Wichmann et al., 1997).
We performed our experiment during reproductivelyinactive phase (November/December) when peripheral
Fig. 3. EVect of testosterone and melatonin administration on (A)basal and mitogen Con A-induced blastogenic response and (B) per-cent stimulation ratio of splenocytes of Indian jungle bush quail, P.asiatica during reproductively inactive phase (November–January).Histograms represent means § SE, n D 5 for each group within thisexperiment. Con, control; Mel, melatonin, and T, testosterone.**P < 0.01 Con vs T, Con vs Mel, Con vs Mel + T, and T vs Mel + T.
testosterone level was low (0.173 ng/ml) and melatoninwas high (260 pg/ml) (Singh, 2003) in order to expressthe correlation of these two hormones on immune statusof the birds when the environmental condition (winter)is stressful, hence having a demand of high immunity.
Our data presented that testosterone treatment to thebirds signiWcantly reduced the spleen weight while mela-tonin treatment along with testosterone restored thespleen mass. This supports the earlier observation inmammals that castration in rat and mice caused enlarge-ment of lymphoid organ (thymus; Henderson, 1904;Olsen et al., 1991) while androgen replacement reducedthe thymic enlargement (Greenstein et al., 1986; Olsen etal., 1991). Similarly, testosterone implantation acceler-ated the involution of bursa of Fabricius in Japanesequail (Mase and Oishi, 1991). The net eVect of androgenaction (direct or indirect) on lymphoid organs appearsto be of suppressive tone. However, involution of lym-phoid organs following androgen treatment is notcompletely understood. Three possible pathways inenlargement of lymphoid organs after castration couldbe suggested, i.e., (i) increased cell proliferation, (ii)decreased cell death, and (iii) diminished traYcking ofsplenocytes to the periphery. Studies have suggested that
Fig. 4. In vitro eVect of testosterone and melatonin supplementationon (A) basal and mitogen Con A-induced blastogenic response and (B)percent stimulation ratio of splenocytes of Indian jungle bush quail, P.asiatica, during reproductively inactive phase (November–January).Histograms represent means § SE, n D 5, for each group within thisexperiment. Con, control; Mel, melatonin; and T, testosterone.**P < 0.01 Con vs T, Con vs Mel, Con vs Mel + T, and T vs Mel + T.
ative Endocrinology 141 (2005) 226–232 229

230 S.S. Singh, C. Haldar / General and Comparative Endocrinology 141 (2005) 226–232
out of those three possibilities, Wrst one occurred aftercastration in normal male mice (Olsen et al., 1994). Spe-ciWc receptors for androgen have been reported on thelymphoid organs (Grossman et al., 1983) suggesting alsohow gonadal steroid can modulate immune responsive-ness and might have inXuenced the development of thecells that express cytokines. Whether gonadal steroidsdirectly regulate the expression of speciWc cytokine genein the diVerentiated cells of the immune system, suchexplorations have only begun.
Our data on total leukocyte and lymphocyte countshowed a signiWcant reduction in testosterone treatedbirds. Melatonin treatment along with testosteronecounteracted the testosterone-induced suppression oftotal leukocyte and lymphocyte count in circulation.Similarly a negative correlation was reported betweentestosterone and lymphocyte number in circulation inred jungle fowl Gallus gallus (Zuk et al., 1995) but not inbarn swallows (Saino et al., 1995).
Further, both in vivo and in vitro testosterone supple-mentation signiWcantly decreased the basal as well asmitogen-induced splenocyte proliferation and percentstimulation ratio (Figs. 3A and B and 4A and B). Mela-tonin administration along with testosterone antago-nized the testosterone-induced suppression and restoredthe basal as well as mitogen-induced blastogenicresponse and percent stimulation ratio in both in vivo
and in vitro experiment. This supports the earlier reportthat testosterone administration decreased the prolifera-tion of splenocytes and IgG production (Yao et al.,2002) in mammals. Further, chronic administration ofmelatonin increased the splenocyte proliferation whenstimulated with T-cell mitogen concanavalin A in maleSyrian hamster (Champney et al., 1997). This result isevident from even avian spleen as noted by us becausethe spleen becomes the principal lymphatic tissue of theadult birds.
The physiological mechanism by which steroid hor-mones, like testosterone, suppress immune function isunclear. However, androgen receptors have been identi-Wed in thymic tissues particularly in the epithelial andlymphatic portion of the thymus (Mc Cruden and Stim-son, 1991; Sasson and Mayer, 1981) of mammals. Since,no androgen receptors have been identiWed on circulat-ing lymphocyte, androgenic eVects on lymphocyte couldbe a direct one or acting through aromatization ofandrogen to estrogen (Mc Cruden and Stimson, 1991).As melatonin receptors have been identiWed on circulat-ing lymphocyte (Calvo et al., 1995; Guerrero and Reiter,1992; Liu and Pang, 1993; Maestroni, 1993; Pang andPang, 1992; Pang et al., 1993) as well as on thymocytesand splenocytes (Lopez-Gonzalez et al., 1993; Martin-Cacao et al., 1993; RaWi-El-Idrissi et al., 1995) of birds,the lymphoproliferation action of melatonin, hence
Fig. 5. (1) Histology of spleen of control bird showing splenic lymphocytes (L) in periarterial region during reproductively inactive phase (Novem-ber–December) 920£. (2) Histology of spleen of testosterone treated bird, showing depletion of splenic lymphocytes (L) in periarterial region duringreproductively inactive phase (November–December) 920£. (3) Histology of spleen of testosterone and melatonin treated bird, showing restorationof splenic lymphocytes (L) in periarterial region during reproductively inactive phase (November–December) 920£. (4) Histology of spleen of mela-tonin treated bird showing more dense splenic lymphocytes (L) in periarterial region during reproductively inactive phase (November–December)920£.

S.S. Singh, C. Haldar / General and Comparative Endocrinology 141 (2005) 226–232 231
could be a direct eVect. Further, evidences of melatoninsynthesis and its presence in bone marrow of mouse andhuman strongly support the immunomodulatory actionof melatonin on immune cells (Carrillo-Vico et al., 2004;Conti et al., 2000; Tan et al., 1999).
The inverse relationship between gonadal steroid tes-tosterone and pineal hormone melatonin suggests thatvariation in peripheral melatonin level provides environ-mental signals to this seasonally breeding avian speciesP. asiatica. Similarly it might be playing an importantrole in the maintenance of the immune status by keepingthem healthy during non-breeding phase (Folstad andKarter, 1992).
Recently, it has been reported that levels of cortico-sterone as well as testosterone were higher in testoster-one treated male Juncos (Casto et al., 2001), raisingdoubt on the fact that whether the immune suppressionobserved by us is due to testosterone’s direct aVect onimmunity or testosterone inXuenced glucocorticoid pro-duction. Therefore, further experiments are needed todissociate the eVects of corticosterone and testosteroneon the avian immune system.
Acknowledgments
Financial assistance from Council of Industrial and Sci-entiWc Research and University Grant Commission, NewDelhi, India, and Instrument gift of Alexander von Hum-boldt Foundation, Germany, are gratefully acknowledged.
References
Bilbo, S., Nelson, R.J., 2001. Sex steroid hormones enhance immunefunction in male and female Siberian hamsters. Am. J. Physiol. 280,R207–R213.
Calvo, J.R., RaWi-El-Idrissi, M., Pozo, D., Guerrero, G.M., 1995.Immunomodulatory role of melatonin: SpeciWc binding sites inhuman and rodent lymphoid cells. J. Pineal Res. 18, 119–126.
Carrillo-Vico, A., Calvo, J.R., Abreu, P., Lardone, P.J., Garcia-Mau-rino, S., Reiter, R.J., Guerrero, J.M., 2004. Evidence of melatoninsynthesis by human lymphocytes and its physiological signiWcance:Possible role as intracrine, autocrine, and/or paracrine substance.FASEB J. 18, 537–539.
Casto, J.M., Nolan Jr., V., Ketterson, E.D., 2001. Steroid hormones andimmune function: Experimental studies in wild and captive dark-eyed Juncos (Junco hyemalis). Am. Nat. 157, 408–420.
Champney, T.H., Prado, J., Youngblood, T., Appel, K., McMurray,D.N., 1997. Immune responsiveness of splenocytes after chronicdaily melatonin administration in male Syrian hamsters. Immunol.Lett. 58, 95–100.
Chaturvedi, C.M., Thapliyal, J.P., 1978. EVects of orchidectomy andmale hormone administration on adrenal of common myna (Acri-dotheres tristis). Ann. Endocrinol. 39, 417–420.
Conti, A., Conconi, S., Hertens, E., Skwarlo-Sonta, K., Markowska,M., Maestroni, J.M., 2000. Evidence for melatonin synthesis inmouse and human bone marrow cells. J. Pineal Res. 28, 193–202.
Drazen, D.L., Bilu, D., Bilbo, S.D., Nelson, R.J., 2001. Melatoninenhancement of splenocyte proliferation is attenuated by luzindole,
a melatonin receptors antagonist. Am. J. Physiol. 280, R1476–R1482.
Evans, M.R., Gold smith, A.R., Norris, S.R.A., 2000. The eVect of tes-tosterone on antibody production and plumage coloration in malehouse sparrow (Passer domesticus). Behav. Ecol. Sociobiol. 47, 156–163.
Folstad, I., Karter, A.J., 1992. Parasites, bright males, and the immuno-competence handicap. Am. Nat. 139, 603–622.
Greenstein, B.D., Fitzpatric, F.T., Adcock, I.M., Kendal, M.D.,Wheeler, M.J., 1986. Reappearance of the thymus in old rats afterorchidectomy; inhibition of regeneration by testosterone. J. Endo-crinol. 110, 417–422.
Grossman, C.J., Sholiton, J., Helmsworth, J.A., 1983. Characteristics ofthe cytoplasmic and nuclear dihydrotestosterone receptors ofhuman thymic tissue. Steroids 42, 11–22.
Haldar, C., Ghosh, M., 1988. EVect of 5-methoxy indoles on the testic-ular function of of the Indian jungle bush quail, Perdicula asiatica.Arch. Anat. Hist. Embriol. Norm. et exp. 71, 97–107.
Haldar, C., Singh, R., Guchhait, P., 2001. Relationship between theannual rhythm in melatonin and immune system in the tropicalpalm squirrel F. pennanti. Chronobiol. Int. 18, 61–69.
Hasselquist, D., Marsh, J.A., Sherman, P.W., WingWeld, J.C., 1999. Inavian humoral immunocompetence suppressed by testosterone.Behav. Ecol. Sociobiol. 45, 167–175.
Henderson, J., 1904. On the relationship of thymus to the sexual organsI. The inXuence of castration on the thymus. J. Physiol. 31, 222–229.
Liu, Z.M., Pang, S.F., 1993. [125I]Iodomelatonin binding sites in the bursaof fabricius of birds: Binding characteristics, sub-cellular distribution,diurnal variations and age studies. J. Endocrinol. 138, 51–57.
Lochmiller, R.L., Deerenberg, C., 2000. Trade-oVs in evolutionaryimmunology. Just what is cost of immunity?. Oikois 88, 87–98.
Lopez-Gonzalez, M.A., Calvo, J.R., Osuna, C., Secura, J.J., Guerrero,J.M., 1993. Characterization of melatonin sites in human peripheralblood neutrophils. Biotechnol. Ther. 4, 253–262.
Maestroni, G.J.M., 1998. The photoperiod transducer melatonin andthe immune-hematopoietic system. J. Photochem. Photobiol. 43,186–192.
Mase, Y., Oishi, T., 1991. EVect of castration and testosterone treatmenton the development and involution of the bursa of Fabricius andthymus in Japanese quail. Gen. Comp. Endocrinol. 84, 426–433.
Mc Cruden, A.B., Stimson, W.H., 1991. Sex steroids and immune func-tion. In: Ader, R., Cohen, J. (Eds.), Psychoneuroimmunology. Aca-demic Press, New York, pp. 475–493.
Moore, C.B., Siopes, T.D., 2000. EVect of lighting conditions and mela-tonin supplementation of the cellular and humoral immuneresponses in Japanese quail, Coturnix coturnix japonica. Gen.Comp. Endocrinol. 119, 95–104.
Nelson, R.J., Drazen, D.L., 1999. Melatonin mediates seasonal adjust-ments in immune function. Reprod. Nutr. Dev. 39, 383–398.
Olsen, N.J., Kovacs, W.J., 1996. Gonadal steroids and immunity. Endo-crine Rev. 17, 369–384.
Olsen, N.J., Viselli, S.M., Shults, K., Stelzer, G., Kovacs, W.J., 1994.Induction of immature thymocytes proliferation after castration ofnormal male mice. Endocrinology 134, 107–113.
Olsen, N.J., Watson, M.B., Henderson, G.S., Kavacs, W.S., 1991.Androgen deprivation induces phenotypic and functionalchanges in the thymus of male adult mice. Endocrinology 129,2471–2476.
Pang, C.S., Pang, S.F., 1992. High aYnity speciWc binding of2[125I]iodomelatonin by spleen membrane preparations of chicken.J. Pineal Res. 12, 167–173.
Pauly, J.L., Sokal, J.E., 1972. A simpliWed technique for in vitro studiesof lymphocytes reactivity. Proc. Soc. Exp. Biol. Med. 140, 40–44.
RaWi-El-Idrissi, M., Calvo, J.R., Pozo, D., Harmouch, A., Guerrero,J.M., 1995. SpeciWc binding of 2[125I] iodomelatonin by rat spleeno-cyte: Characterization and its role on regulation of cyclic AMPproduction. J. Neuroimmunol. 57, 171–178.

232 S.S. Singh, C. Haldar / General and Comparative Endocrinology 141 (2005) 226–232
Reiter, R.J., Calvo, J.R., Karbownik, M., Qi, W., Tan, D.X., 2000. Mela-tonin and its relation to the immune system and inXammation.Ann. NY Acad. Sci. 917, 376–386.
Ros, A.F.H., Groothius, T.T.G., Apanius, V., 1997. The relationshipamong gonadal steroids, immunocompetence body mass andbehaviour in young black-headed gulls (Larus ridibudus). Am.Nutr. 150, 201–219.
Saino, N., Moller, A.P., 1994. Secondary sexual characters, parasitesand testosterone in the barn swallows (Hirundo rustica). Anim.Behav. 48, 1325–1333.
Saino, N., Moller, A.P., Bolzern, A.M., 1995. Testosterone eVect onimmune system and parasitic infestations in the barn swallows(Hirundo rustica): An experimental test for the immunocompetencehypothesis. Behav. Ecol. 6, 397–404.
Sasson, S., Mayer, M., 1981. Antiglucocorticoid activity of androgensin rat thymus lymphocytes. Endocrinology 108, 760–766.
Schuurs, A.H., Verheul, H.A.M., 1990. EVects of gender and sex ste-roids on the immune response. J. Steroid Biochem. 35, 157–172.
Singh, S.S., 2003. Avian pineal organ: Role in maintenance of Immu-nity in Indian jungle bush quail, Perdicula asiatica. PhD Thesis,Banaras Hindu University, India.
Skwarlo-Sonta, K., 1999. Reciprocal interdependence between pinealgland and avian immune system. Neuroendocrinol. Lett. 20, 151–156.
Sudhakumari, C.C., 1995. Comparative study of pineal–adrenal inter-relationship in a nocturnal (Athene brama) and diurnal (Perdiculaasiatica) bird of tropical Indian origin. PhD Thesis, Banaras HinduUniversity, Varanasi, India.
Tan, D.X., Manchester, L.C., Reiter, R.J., Qi, W.B., Zhang, M., Wein-traub, S.T., Cabrera, J., Sainz, R.M., Mayo, J.C., 1999. IdentiWcationof highly elevated levels of melatonin in bone marrow: Its originand signiWcance. Biochim. Biophys. Acta 1472, 206–214.
Verhulst, S., Dieleman, S.J., Parmentier, H.K., 1999. A trade oV
between immunocompetence and sexual ornamentation in domes-tic fowl. Proc. Natl. Acad. Sci. USA 96, 4478–4481.
Weatherhead, P.J., Metz, K.J., Bennett, G.E., Irwin, R.E., 1993. Parasitefaunas, testosterone and secondary sexual traits in male red wingedblack birds. Behav. Ecol. Sociobiol. 33, 12–23.
Wichmann, M.W., Ayala, A., Chaudry, I.H., 1997. Male sex steroids areresponsible for depressing macrophage immune function aftertrauma hemorrhage. Am. J. Physiol. 273, C1335–C1340.
Yao, G., Hou, Y., Warg, Y., 2002. EVect of testosterone on prolifera-tion and IgG production of splenic lymphocytes in vitro. Wei.Sheng Yan. Jiu. 31, 333–335.
Zuk, M., Johnsen, T.S., MacLarty, T., 1995. Endocrine immune interac-tions, ornaments and mate choice in red jungle fowl. Proc. R. Soc.Lond. B 260, 205–210.


















![James Clavell [Seria asiatica 1] Changi - 101books.club](https://static.fdokumen.com/doc/165x107/633d1b8c5f621d25220311a1/james-clavell-seria-asiatica-1-changi-101booksclub.jpg)

