
The stability of tolerance of Sorghum spp to Striga asiatica L ...
230
i The stability of tolerance of Sorghum spp to Striga asiatica L. Kuntze under diverse conditions and existence of pre- attachment resistance Mandumbu Ronald (201317414) A Thesis Submitted to the Faculty of Science and Agriculture in Fulfillment of the Requirements of the Degree of Doctor of Philosophy Department of Agronomy Faculty of Science and Agriculture University of Fort Hare July 2017
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of The stability of tolerance of Sorghum spp to Striga asiatica L ...

i
The stability of tolerance of Sorghum spp to Striga asiatica
L. Kuntze under diverse conditions and existence of pre-
attachment resistance
Mandumbu Ronald (201317414)
A Thesis Submitted to the Faculty of Science and Agriculture in
Fulfillment of the Requirements of the Degree of Doctor of Philosophy
Department of Agronomy
Faculty of Science and Agriculture
University of Fort Hare
July 2017

ii
DECLARATION
I Ronald Mandumbu, declare that the work contained in this thesis is entirely my own work
and that all reference materials contained in this thesis have been duly acknowledged. This
thesis has not been previously submitted to this or any other University for the award of a
degree.
Signature………………………………………………
Date…………………………………………………….

iii
PUBLICATIONS
1. Mandumbu R, Mutengwa C, Mabasa S, Mwenje E (2016). Existence of different
physiological strains of Striga asiatica (L.) Kuntze on Sorghum spp [Sorghum bicolor
and Sorghum arundinaceum (Desv) Stapf) in Zimbabwe. Research on Crops 17 (3):
468 – 478. DOI10.5958/2348-7542.2016.00077.2
2. Mandumbu R, Mutengwa C, Mabasa S, Mwenje E (2017) The effect of witchweed
(Striga asiatica) infestation and moisture stress on selected morpho-physiological
traits of sorghum in Zimbabwe. Journal of Agronomy 16 (2): 65 - 75.
3. Mandumbu R, Mutengwa C, Mabasa S, Mwenje E (2017) determination of resistance
to Striga asiatica (L.) Kuntze using agar jel analysis and sand culture in Sorghum
bicolor and Sorghum arundinaceum in Zimbabwe. Accepted by the Asian Journal of
Crop Science.
4. Mandumbu R, Mutengwa C, Mabasa S, Mwenje E (2017) Response of Sorghum
bicolor (L) Moench and Sorghum arundinaceum (Desv) Stapf to Striga asiatica (L)
Kuntze infestation under mulch. Accepted by Tropical Agriculture .
5. Mandumbu R, Mutengwa C, Mabasa S and Mwenje E (2017). The Striga scourge
under changing climate in southern Africa: A perspective. Accpeted by Journal of
Biological Science.
6. Mandumbu R, Mutengwa C, Mabasa S, Mwenje E (2017). Factors affecting the
success of resistance as a management strategy in Striga management: A review.
Under review with the Asian Journal of Plant Science.

iv
PREFACE
This thesis is presented in a form of a General Introduction (Chapter 1) and Literature
Review (Chapter 2) which introduces the reader to sorghum and Striga issues in Zimbabwe
and elsewhere in the world. Chapter three deals with the simultaneous effect of reduced water
availability and Striga asiatica infestation on the morpho-physiological attributes of
sorghum. Chapter four investigates the effect of mulching as a cultural practice on the
stability of tolerance by Sorghum spp to Striga asiatica. Chapter five focuses on the variable
effects of two Striga asiatica strains sourced from two distant places in Zimbabwe on
sorghum productivity and the stability of tolerance. Chapter 6 deals with the quantification of
strigolactones in Sorghum bicolor and Sorghum arundinaceaum and determines the
relationship between strigolactone quantities and sorghum tillering. Finally, the general
discussion, conclusions and recommendations are reported in the last chapter (Chapter 7).
This thesis was written in paper format and there is therefore unavoidable repetition of some
information, including references.

v
ABSTRACT
Sorghum is the fifth most important cereal in the world and a staple food for humans. It is
also a source of food and fodder for animals. In addition to the abiotic stresses such as
drought, parasitic weeds of the genus Striga cause losses in sorghum production in sub
Saharan Africa. Striga asiatica is a parasitic weed that attacks cereals in low input
agricultural systems and is distributed throughout semi- arid regions of Africa. Most sorghum
producing farmers rely on tolerance for their harvests in Striga infested fields yet the stability
of tolerance in the face of a changing climate (recurrent droughts), new farming systems
(mulch based agriculture) and existence of various Striga strains needs further investigation.
Reduced strigolactones production was also studied as a resistance mechanism.
The first study was focused on the determination of tolerance of Striga asiatica infested
sorghum under drought in a pot study. Five sorghum lines were subjected to infestation with
Striga and some were not infested while watering was done at 50 % field capacity (FC) and
100% FC. The results showed that the five sorghum lines differed significantly in chlorophyll
content and Normalised Differential Vegetation Index (NDVI). Infection did not lower
chlorophyll content when it co-occurred with drought across all sorghum lines. Drought and
infestation had mutually exclusive effects on chlorophyll content and NDVI. Under
infestation, internode length was similar both at 100 % FC and at 50 % FC while under
uninfested conditions, 100 % FC gave the longer internode compared to 50 % FC. Both
infestation and irrigation regime reduced the sorghum head weight, illustrating that the two
effects have synergistic effects on sorghum head weight.
The second study sought to determine the effects of mulching and infestation on sorghum spp
tolerance to Striga asiatica. The experiments were carried out in the seasons 2013/14 and
2014/15 summer seasons. The results indicated that mulching increased chlorophyll content

vi
in the 2014/15 season which was a drier season compared to 2013/14. In the 2014/15 season,
mulching increased chlorophyll content in all varieties except Ruzangwaya, Mukadziusaende
and SC Sila. When the same varieties were infested under mulch and infested without mulch,
the results showed that mulching overcomes the effects of infestation in some varieties.
Mulch also negates the effect of Striga parasitism and results in yield maintenance in
sorghum varieties.
The third study sought to determine the stability of sorghum lines when exposed to two
Striga asiatica lines sourced from two places which are 500 km apart in Zimbabwe. The two
strains were termed the Chiundura and Rushinga strains, based on where they were sourced.
The experiments were conducted at Henderson Research Station (HRS) at Mazoe and at
Bindura University of Science Education (BUSE). The results showed differential virulence
for some traits while the two strains were equally virulent for some traits. The two strains
were equally virulent on all sorghum lines with respect to chlorophyll content. The different
sorghum lines responded differently to the effects of the two strains. The effects of the two
strains were generally similar for head index, root index, and leaf index at all sites. Generally
the Chiundura strain was more virulent to sorghum lines compared to Rushinga strain,
confirming the existence of physiological strains of Striga in Zimbabwe. Therefore
physiological speciation of Striga asiatica exists and this adds a further dimension to the
complexity of Striga management in the smallholder sector.
Quantification of strigolactone production by different sorghum genotypes was conducted in
the laboratory using the agar gel assay. The genotype Mukadziusaende produced
significantly the least (P<0.01) quantities of strigolactones, as inferred from the maximum
germination distance (MGD) from the sorghum root. The MGD was negatively correlated to
tiller numbers illustrating that the more the strigolactones the less the tillering capacity. Tiller
numbers and MGD can therefore be used to select for reduced strigolactones production.

vii
Key words
Sorghum bicolor, Sorghum arundinaceum, Striga asiatica, resistance, tolerance, drought,
mulch, physiological speciation, strigolactones.

viii
DEDICATION
This study is dedicated to my wife, Virginia, my children, Praise, Gladiness, Anesu and Ruth,
my parents Ndedza and Enita Mandumbu.

ix
ACKNOWLEDGEMENTS
In a doctoral study, one is indebted to so many sources of assistance that it is practically
impossible to list them all. At the risk of omission, I would like particularly to thank the
people and institutions listed below.
I would like to thank my supervisors: Professor C.S. Mutengwa, Dr Stanford Mabasa and
Professor Eddie Mwenje for their guidance throughout the course of this work. This work
was made possible by the financial assistance from the Zimbabwe Manpower Development
Fund (ZIMDEF), the Research Council of Zimbabwe (RCZ) and the Research and Post
Graduate Centre of the Bindura University of Science Education (BUSE).
I want to thank Mr Chikaka, Mr Gochera, Zengeza Tapiwa, Chitaukire Charity and Maramba
Komborero, Mr Kufa Mutsengi and Munyati Vincent for the assistance of setting the
experiments and data collection. Mr Senga and Mr Kamhapa for the assistance in laboratory
work. Mr Parwada and Mr Mafuse for assembling at our reading hub as we had the same
challenges of coming up with a thesis. The Weed Research Team at Henderson Research
station is acknowledged for their assistance with space and data collection for the
experiments which were carried out there. I also want to thank the Mafirenyika family for
hosting me in East London every time I was in South Africa. They made my stay a
memorable experience with great dinners, break fast and sight seeing. I also want to thank my
brothers and sisters who include Tongai, Pesanai, Georgina, Tawanda, Solomon, Ruvarashe
and Chiedza and their families for inspiration. I also thank Diriri Simbarashe and family for
their support. My brother Zvamaida Gumbo is acknowledged for teaching me to read, I am
sure you also did not know it was coming to this. To Effort Macheza, Noel Gumbo and
Model Macheza, the button is now in your hands and the tracklines are open.
Never should I forget the Lord Jesus Christ for His unwarranted favour, for giving me
strength and wisdom to complete this study. Last but definitely not least I want to thank
Prophet Emmanuel Makandiwa, Prophetess Ruth Makandiwa and the whole United Family
International Church for the teachings, motivation and inspiration I got which have kept me
going and grounded in the word of God. And He said I give unto you POWER.

x
Table of Contents
PREFACE ................................................................................................................................. iv
ABSTRACT ............................................................................................................................... v
DEDICATION ....................................................................................................................... viii
ACKNOWLEDGEMENTS ...................................................................................................... ix
1.1 Statement of the problem ................................................................................................. 1
1.2 Justification of the study .................................................................................................. 5
1.3 Main objective .................................................................................................................. 7
1.4 Specific objectives............................................................................................................ 7
References .............................................................................................................................. 9
CHAPTER TWO: LITERATURE REVIEW ......................................................................... 15
2.1. Introduction to literature review .................................................................................... 15
2.2 Sorghum production in Zimbabwe ................................................................................. 15
2.3 The genus Striga............................................................................................................. 16
2.3.1 Striga hermonthica ................................................................................................... 17
2.3.2 Striga asiatica ........................................................................................................... 17
2.3.3 Striga gesneroides .................................................................................................... 18
2.4 Extent of the Striga problem in sub-Saharan Africa ...................................................... 19
2.5 Striga problem in the smallholder sector of Zimbabwe ................................................. 20
2.6 History and distribution of Striga asiatica in Zimbabwe ............................................... 24
2.7 Striga asiatica research in Zimbabwe ............................................................................ 24
2.8 Striga asiatica lifecycle .................................................................................................. 27
2.8.1 Dormancy and conditioning .................................................................................... 27
2.8.2 Germination ............................................................................................................. 28
2.8.3 Haustorium development ......................................................................................... 28
2.8.5 Establishment of parasitism and flowering ............................................................. 30

xi
2.9. Strigolactones in plants ................................................................................................. 31
2.10 The parasite as a sink ................................................................................................... 33
2.11 Mechanisms of sorghum tolerance to Striga asiatica .................................................. 33
2.11.1 Host photosynthesis ............................................................................................... 33
2.11.2 Sorghum tillering as Striga tolerance mechanism ................................................. 35
2.11.3 Environmental regulation of tillering ....................................................................... 36
2.12.1 Propensity to tiller ................................................................................................. 36
2.13 Host plant resistance against Striga ............................................................................. 37
2.13.1 Reduced host stimulant production ....................................................................... 37
2.13.2 Reduced haustorium inducing factors (Low haustorial factor) ............................ 38
2.13.3 Parasite establishment resistance ........................................................................... 39
2.13.4 Post-attachment resistance ..................................................................................... 39
2.13.5 Incompatible response (IR) ................................................................................... 39
2.13.6 Hypersensitive response ........................................................................................ 40
2.14 Managing Striga asiatica in sub-Saharan Africa ......................................................... 40
2.15. Sorghum response to drought stress ............................................................................ 42
2.16 Existence of Striga physiological speciation and differential virulence ...................... 43
2.17 Mulch effects on Striga asiatica incidence .................................................................. 44
2.18 Photosynthesis in a Striga-infested plant ..................................................................... 46
2.18.1 Stomatal conductance ............................................................................................ 47
Chapter Three: ......................................................................................................................... 68
The effect of witchweed (Striga asiatica L. Kuntze) and moisture stress on selected morpho-
physiological traits which impart tolerance to Sorghum spp ................................................... 68
3.1 Abstract .......................................................................................................................... 68
3.2 Introduction .................................................................................................................... 69
3.3 Methodology .................................................................................................................. 73
3.3.1 Experimental site ..................................................................................................... 73

xii
3.3.3 Experimental design and treatments ........................................................................ 73
3.3.5 Irrigation .................................................................................................................. 74
3.3.6 Data collection ......................................................................................................... 74
3.4 Results ............................................................................................................................ 75
3.4.1 Chlorophyll concentration and NDVI ......................................................................... 75
3.4.2 Dry matter traits ....................................................................................................... 81
3.5 Discussion ...................................................................................................................... 89
References ............................................................................................................................ 94
CHAPTER FOUR:................................................................................................................. 103
The response of tolerance traits of Sorghum bicolor (L) Moench and Sorghum arundinaceum
(Desv) Stapf to Striga asiatica (L) Kuntze infestation under mulch ..................................... 103
4.1 Abstract ........................................................................................................................ 103
4.3 Materials and methods ................................................................................................. 106
4.3.1 Experimental site ................................................................................................... 106
4.3.2 Experimental design and pot layout ...................................................................... 106
4.3.4 Source of Seeds ..................................................................................................... 107
4.4 Results .......................................................................................................................... 108
4.4.4 Plant height ............................................................................................................ 114
4.4.5 Grain yield ............................................................................................................. 119
4.5 Discussion .................................................................................................................... 121
6.0 References .................................................................................................................... 126
The existence of different physiological ‘strains’ of Striga asiatica (L.) kuntze on Sorghum
bicolor (L.) Moench and Sorghum arundinaceum (desv) Stapf in Zimbabwe ...................... 131
5.1 Abstract ........................................................................................................................ 131
5.2 Introduction .................................................................................................................. 132
5.3 Materials and Methods ................................................................................................. 134
5.3.1 Experimental sites.................................................................................................. 134
5.3.4 Experimental details .............................................................................................. 135

xiii
5.3.5 Data Collection ...................................................................................................... 136
5.4 Results .......................................................................................................................... 136
5.4.1 Sorghum plant height............................................................................................. 136
5.4.3 Sorghum tillering ................................................................................................... 143
5.4.4 Sorghum dry matter traits ...................................................................................... 145
5.4.5 Sorghum head weight ............................................................................................ 148
5.4.6 Total dry weight..................................................................................................... 150
5.5 Discussion .................................................................................................................... 152
5.7 References .................................................................................................................... 156
CHAPTER SIX ...................................................................................................................... 161
Sorghum response to Striga asiatica based on maximum germination distance, Striga counts
and sorghum tillering in Zimbabwe ....................................................................................... 161
6.1 Abstract ........................................................................................................................ 161
6.2 Introduction .................................................................................................................. 162
6.3 Materials and methods ................................................................................................. 165
6.3.1 Experiment 1: Agar jel assays ................................................................................... 165
6.3.2 Sorghum germplasm and Striga asiatica seed sources ............................................. 165
6.3.3 Experimental design .................................................................................................. 165
6.3.4 Surface Sterilisation and sorghum seed germination ................................................ 165
6.3.5 Conditioning of Striga seed ................................................................................... 166
6.3.6 The assay set up ..................................................................................................... 166
6.5 Results .......................................................................................................................... 167
6.5.1 Maximum germination distance (MGD) ................................................................... 167
6.5.2 Tillering ..................................................................................................................... 168
6.5.3 Striga counts .............................................................................................................. 169
6.5.3 Correlations between maximum germination distance, tillering and Striga counts . 170
6.6 Discussion .................................................................................................................... 171

xiv
6.7 Conclusion .................................................................................................................... 173
6.8 References ............................................................................................................... 173
CHAPTER SEVEN: GENERAL DISCUSSION, CONCLUSIONS AND
RECOMMENDATIONS ................................................................................................... 178
7.1 Introduction .................................................................................................................. 178
7.2 Discussion .................................................................................................................... 178
7.3 Conclusions .................................................................................................................. 180
7.4. Recommendations for further research ....................................................................... 181

xv
List of tables
Table 2.1: Zimbabwe sorghum production by year from 2010 - 2014 .................................... 16
Table 2.2: The distribution and occurrence of Striga spp in sub-Saharan Africa .................... 18
Table 2.3: Agro-ecological regions of Zimbabwe and agricultural activities carried out in the
various regions. ........................................................................................................................ 22
Table 2.4: Different methods of controlling Striga asiatica ..................................................... 40
Table 3.1: Sorghum genotypes effects on chlorophyll content at 6 and 10 WACE ................ 76
Table 3.2: Moisture stress effects on NDVI at 6 and 10 WACE ............................................. 77
Table 3.3: Effect of S. asiatica infection on chlorophyll content of sorghum ......................... 79
Table 3.5: Effect of sorghum genotypes on head weight and head index ............................... 82
Table 3.6: The effect of infection on head weight and head index .......................................... 83
Table 3.6: The effect of water availability on head weight and head index across the two
experiments .............................................................................................................................. 83
Table 3.7: Effect of sorghum genotypes on leaf dry matter, leaf index, stem weight and index
and total dry matter in both experiments ................................................................................. 86
Table 3.8: The effect of infection on leaf weight and index, stem weight and index and total
dry matter in both experiments. ............................................................................................... 86
Table 3.9: The effects of water availability on leaf weight and index, stem weight and index
and total dry matter. ................................................................................................................. 87
Table 4.1: Sorghum genotypes tested for tolerance to Striga in the 2013/14 and 2014/15
seasons ................................................................................................................................... 107
Table 4.2: Effect of mulching on chlorophyll concentration for the 2013/14 and 2014/15
summer seasons ..................................................................................................................... 109
Table 4.6: Effect of sorghum variety on Striga counts .......................................................... 113
Table 4.7: Effects of infestation on plant height at 8 and 12 WACE in the year 2013/14 and
2014/15 seasons. .................................................................................................................... 117
Table 5.4: Effect of sorghum genotypes on head index, root index, stem index and leaf index
at BUSE and Henderson sites. ............................................................................................... 146
Table 5.5: Effect of Striga strains on head, root, stem and leaf index at BUSE and Henderson.
................................................................................................................................................ 147

xvi

xvii
List of figures
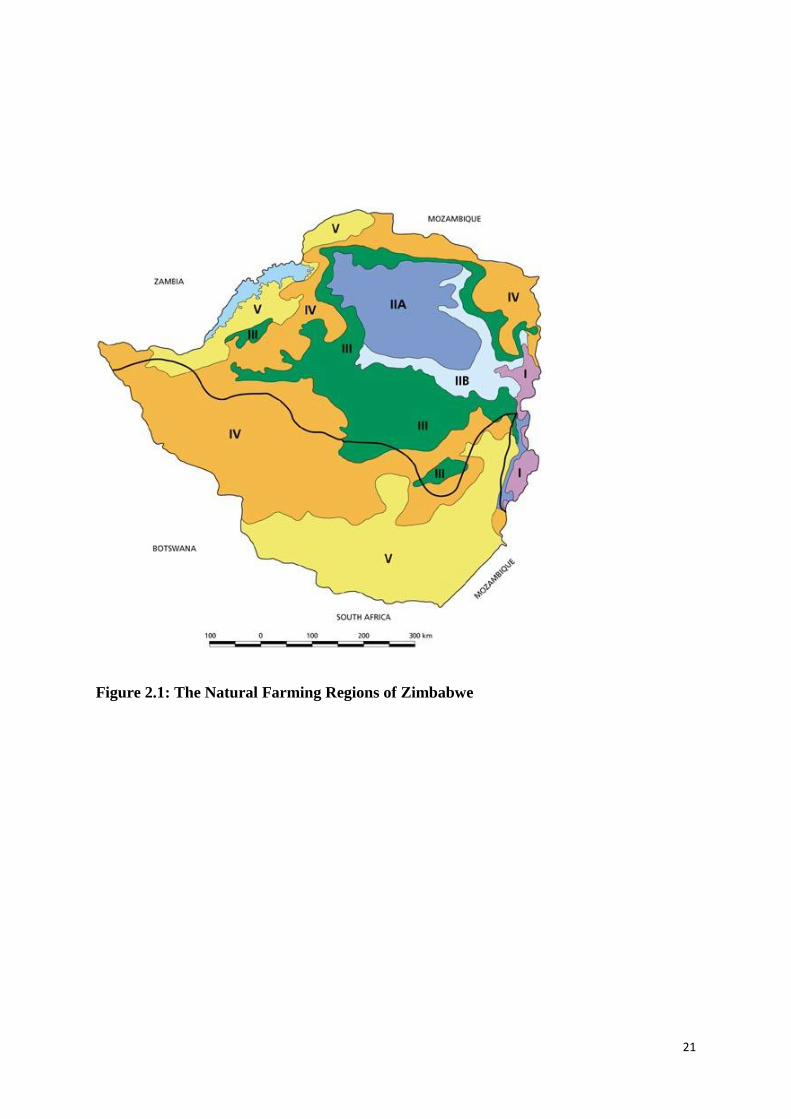
Figure 2.1: The Natural Farming Regions of Zimbabwe ......................................................... 21
Figure 2.2: The life cycle of S. asiatica (Rich and Ejeta, 2007). ............................................. 27
Figure 3.1: Interaction effects of sorghum genotype and moisture availability on NDVI at 10
WACE in Experiment II. ......................................................................................................... 78
Figure 3.2: Interaction effects of sorghum genotypes and water availability on chlorophyll
concentration at 6 WACE in Experiment II. ............................................................................ 79
Figure 3.3: Effect of Striga infestation on internode length. ................................................... 80
Figure 3.4: Interaction effects of Striga infestation and water availability on sorghum
internode length. ...................................................................................................................... 81
Figure 3.3: The response of sorghum genotypes yield to moisture availability. ..................... 84
Figure3.6: Interaction effects of water availability and Striga infestation on leaf index. ....... 88
Figure 3.7: Interaction effects of sorghum genotypes and Striga asiatica infestation on stem
weight ....................................................................................................................................... 89
Figure 4.1: Effect of infestation on chlorophyll concentration at 6 WACE during the 2014/
15 season. ............................................................................................................................... 109
Figure 4.2: Interaction between variety and Striga infestation a) 6 WACE andvariety and
mulch b) 8 WACE on chlorophyll concentration ................................................................. 110
Figure 4.3: Effect of infestation status on stomatal conductance for the sorghum genotypes
during the 2013/14 season. .................................................................................................... 111
Figure 4.4: Effect of season on tiller numbers over two seasons........................................... 112
Figure 4.6: Interaction effects of sorghum genotype and S. asiatica infection on sorghum
height in the 2014/15 season at 8 and 12 WACE. ................................................................. 115
Figure 4.7: Interaction effects of sorghum genotype, mulching and infestation on plant height
in the 2014/15 season. ............................................................................................................ 116
Figure 4.8: Interaction effects of sorghum variety and infestation on plant height at 4, 8 and
12 WACE during the 2013 season. ........................................................................................ 118
Figure 4.9: Interaction effects of sorghum genotypes and infestation on sorghum yield for the
2013/14 season ....................................................................................................................... 119
Figure 4.10: Effect of sorghum genotypes, infestation and mulching on sorghum yield in the
2014/15 season ....................................................................................................................... 120

xviii
Figure 5.1: Interaction effects of sorghum genotypes and Striga strain on sorghum height at 6,
8 and 12 WACE at Henderson research station. .................................................................... 138
Figure 5.2: Interaction effects of sorghum genotype and Striga strains at 12 WACE at BUSE
................................................................................................................................................ 139
Figure 5.3: Interaction effects of sorghum genotype and Striga strains on chlorophyll content
at 10 WACE at both sites ....................................................................................................... 143
Figure 5.4: Effect of sorghum variety on tiller number at 12 WACE at both sites ............... 144
Figure 5.5: Effect of Striga strains on tiller number. ............................................................. 145
Figure 5.7: Interaction effects of sorghum genotypes and Striga strain at BUSE and
Henderson .............................................................................................................................. 149
Figure 5.8: Total dry matter of sorghum genotypes for BUSE and Henderson .................... 150
Figure 5.9: Interaction effects of sorghum genotype and Striga strain on total dry mass at
BUSE ..................................................................................................................................... 151
Figure 6.1: Maximumgermination distances for various sorghum genotypes……….….....168
Figure 6.2: Effect of Striga asiatica on tillering of sorghum genotypes ................................ 169
Figure 6.3: Effect of Sorghum genotypes on Striga counts ................................................... 169
Figure 6.4 : The relationship between germination distance and tiller numbers in
sorghum.................................................................................................................................170
Figure 6.5: Relationship between sorghum yield and tiller numbers………..…………..…171

xix
List of Acronyms
WACE – Weeks after crop emergence
FC – Field capacity
NDVI—Normalised vegetation difference index
BUSE – Bindura University of Science Education
HRS—Henderson research Station
MGD—Maximum germination distance

1
CHAPTER ONE: INTRODUCTION
1.1 Statement of the problem
Sorghum (Sorghum bicolor L. Moench) is an important crop in traditional farming systems
and in the diet of millions of people in the semi-arid tropics (Haussmann et al., 2001). It is an
annual C4 crop commonly consumed as human food and livestock feed and it feeds more than
500 million people in 98 countries in Africa, Asia and America (Pennissi, 2009). It is one of
the top five cereal grains produced worldwide (Burdette, 2007). In Zimbabwe, it is ranked the
third most important cereal after maize and wheat (FAO, 1996). It can maintain remarkable
yield potential in environments that are normally too extreme for other C4 plants (Tari et al.,
2013).One of the remarkable characteristics of sorghum is its drought tolerance, which has
made it an important cereal grown for food and beverages by resource poor farmers in sub-
Saharan Africa. . Sorghum represents a large proportion of the calorie intake of the people in
sub-Saharan Africa. White sorghum is ground into flour mostly for making sadza (a stiff
dumpling), beverages and porridge (Mutengwa, 2004). It can be popped like popcorn to
create a delicious snack food.
Among a myriad of factors constraining sorghum production, Striga is the major biological
constraint that hinders increased sorghum production in the small-holder sector of sub-
Saharan Africa. Striga species are a major parasitic weedy pest throughout the semi-arid sub-
Saharan Africa and many parts of Asia (Rubiales et al., 2009). Many cropping fields in
African countries including, Tanzania, Kenya, Malawi, Madagascar, Botswana, Zimbabwe,
Gabon, Nigeria, Ethiopia, Niger, Togo, Benin and Burkina Faso are highly infested with
Striga causing serious yield losses that are as high as 100 % at some sites (Lagoke et al.,
1988; Badu-Apraku et al., 2014; Bozkurt et al., 2014). In Kenya, crop losses have been
reported to be as high as 100 % in sorghum. For maize, losses of 50 % under ‘moderate’

2
infestation and 87 % under heavy infestation have also been reported (Manyong et al., 2007).
In Zimbabwe, complete crop failure has been reported by Mabasa (2003). The parasite is a
major constraint to subsistence agriculture in Africa such that resource poor farmers are
sometimes forced to abandon their fields with grave consequences to their families (Berner et
al., 1995; Ejeta, 2007). In particular, major crops that supply the bulk of the energy and
protein needs of the poor in the African savannah, namely maize, sorghum, millets, upland
rice and cowpeas have been severely vulnerable (Ejeta, 2005).
Losses from Striga are compounded because of the tendency of crops grown under severe
moisture and poor fertility conditions to show significant predisposition to Striga. According
to Timko et al., (2012), two thirds of the farmland under cultivation in sub-Saharan Africa is
infested with one or more Striga spp directly affecting livelihoods of more than 300 million
people in 25 countries. According to Scholes and Press (2008) and Ejeta (2007), over 50
million hectares of arable farmland under cultivation with cereals and legumes in sub-
Saharan Africa are infested with one or more Striga species. In many of these places, the
Striga has reached epidemic proportions presenting a desperate situation in subsistence
agriculture (Ejeta and Butler, 1993). The weed causes annual losses of yield estimated to be
in excess of US$10 billion (Ejeta, 2007). Striga spp affects the welfare and livelihoods of
over 100 million people in Africa (Rubiales et al., 2009). According to Parker (2009), the
weed has impacted on the sub-region’s economy. Striga spp stand as a major constraint that
prevents attainment of household food security for some of the world’s poorly resourced
people.
The Striga problem in sub-Saharan Africa is made worse by its exquisite adaptation to the
climatic conditions of the semi-arid tropics, its high fecundity and longevity of its seed
reserves in the tropical soils (Ejeta, 2007). The problem is tending to increase rather than
decrease as intensive land use and lack of fertilizers leads to continued decline in soil fertility

3
which greatly favours Striga (Parker, 2012). The climatic conditions of sub-Saharan Africa
permit timely break down of seed dormancy and conditioning of Striga seeds. Striga asiatica
has a high reproductive capacity, producing 10 000 to 20 000 seeds per plant (Hearne, 2009).
Striga seeds have very small dimensions (0.3 nm * 0.15 nm) and are light (4 – 7 ug) such that
they are easily dispersed by wind, water and animals. The viability of Striga seed goes
beyond 20 years in the soil and this creates seed banks that are difficult to manage.
The life cycle of the noxious cereal weed S.asiatica is complex and has co-evolved with
many hosts to comprise a series of discrete steps that are tightly coupled with the host’s
biochemistry, life cycle and genotype (Bouwmeester et al., 2003). The parasitic plant grows
underground for 4 – 7 weeks prior to emergence and utilizes host water, nutrients and
photosynthates (Jamil et al., 2012). Yoneyama et al., (2010) reported that seeds of parasitic
Striga only germinate after perceiving a germination stimulant (strigolactones) from their
host. The concentration of the stimulant required to initiate germination are as low as 10-18
M
soil solution (Stewart and Press, 1990, Yoneyama et al., 2007).
No single completely effective and practical method to eliminate Striga is known (Bozkurt et
al., 2014). Over the years, many promising Striga control strategies have been suggested in
various formats, with some suggestions appearing in multiple incarnations (Hearne, 2009).
Inspite of all this valuable work, adoption and utility of control methods is limited. The
control measures are neither fully effective nor easy to apply. Only marginal successes have
been obtained to date (Joel et al., 2006). As a result, yield loss attributable to Striga is acute,
as noted before (De Groote et al., 2008). Hence Striga continues to present a challenge for
the forseable future, not only in the areas already affected but also in terms of introduction
into new areas (Parker, 2012).

4
Host plant defense against Striga spp is the only sustainable route for Striga control. It
constitutes two complimentary mechanisms and these are resistance and tolerance. According
to Kim (1994) and Badu-Apraku et al., (2006) resistance to Striga refers to the ability of a
host plant to stimulate the germination of Striga seeds but prevent the attachment of the
parasite to its roots or kill the attached parasite. Host resistance is multi-dimensional with
both general and specific defence mechanisms that can disrupt critical steps throughout the
parasite’s life. Kim (1994) and Rodenburg and Bastiaans (2011) reported that a Striga
tolerant genotype germinates and supports many Striga plants as the intolerant ones, but
produces more grain and stover and shows fewer damage symptoms. Different cultivars may
differ in their capacity to tolerate the physiological and pathological effects caused by Striga
parasitism, finally resulting in milder or stronger impacts on crop yield (Cardoso et al., 2011).
There is only a marginal role of tolerance in the exploitation of host plant defense in sorghum
against Striga asiatica. Resistance is neither complete nor everlasting. The high level of
genetic variation in Striga populations, combined with typical high seed production rates
threatens the durability of resistance. The loss of resistance can be disastrous as the parasite
depends on the staple crops of rural farmers. Tolerance will act as a ‘safety net’ that prevents
a sudden and unforeseen collapse in food supply (Rodenburg and Bastiaans, 2011).
According to Gurney et al., (2003), the control of Striga has proved challenging, mostly as a
result of intricate life cycle of the parasite with the host. Much research has therefore focused
on the development of cereals resistant to infection as a sustainable long term solution.
Complete resistance has not been identified in sorghum although varieties differ in their
sensitivity to infection (Gurney et al., 1995). Therefore, the traits that reduce fitness impacts
of damage such as increased photosynthesis, compensatory growth, architecture of auxillary
buds and carbon storage organs are very critical in varietal selection under Striga infestation.

5
Understanding the mechanisms that plants use to defend themselves and the ecological
drivers thereof has been a major research problem (Hartmann, 2008; Agrawal, 2011).
1.2 Justification of the study
Despite cultivating the sorghum crop in S. asiatica infested fields, some level of yield has
been maintained in sorghum due to tolerance. Tolerance gives the ability to produce yield
despite Striga infestation. The stability of tolerance may be affected by exogeneous
environmental drivers such as athropogenically induced climate change, changes of farming
systems or cultural practices, or the existence of genetic variance in Striga asiatica virulence.
According to Stringer et al., (2009), agricultural systems face the increasing risk of water
stress and that can affect the nature of parasitism between Striga and its host. The stability of
tolerance may vary across genotypes and ecological contexts. The most common scenario is
for Sorghum to deal with both water stress and S. asiatica infestation. Given that global
change involves modification of a series of environmental factors concurrently and changes
in the severity of different stress factors, knowledge on how plants acclimate to multiple
successive or multiple combined stresses is of key significance in understanding the effects of
future climates on the parasite. A combination of two or more stresses such as drought and
Striga infestation is a common occurence to many agricultural areas around the world and
impacts negatively on crop productivity. According to Suzuki et al., (2014) there is an urgent
need to generate crops with enhanced tolerance to stress combinations. Information is limited
in literature on the stability of sorghum tolerance under both drought and Striga asiatica
stresses.
Additionally, human interventions to curb the effects of climate change can modify the
agricultural landscape to the demise or promotion of the parasite. Mulch based farming
systems being promoted as a component of conservation agriculture in sub Saharan Africa
may affect the expression of tolerance traits in some sorghum genotypes. Mulch increases

6
soil moisture and fertility, decreases soil temperature, light transmittance and these are
antagonistic to weed development (Carsky et al., 1994; Oswald et al., 2002). Information is
not available for Zimbabwe on the effects of this practice on the Striga epidemic and whether
it enhances the expression of tolerance in sorghum. Weed responses to such cultural practices
are usually species and genotype specific.
Sorghum arundinaceum (wild sorghum) is increasing in Zimbabwe’s cropping systems as a
weed and occurs in all crops, even where sorghum has never been grown. Near relatives of
cereals could provide new sources of tolerance and resistance (Ejeta et al., 2000, Gurney et
al., 2001, 2002). Studies done by Gurney et al., (2002) indicated that the Sorghum
arundinaceum strain they used demonstrated tolerance to infections by S. asiatica in terms of
growth, biomass accumulation and grain production and this contrasted with Sorghum
bicolor, in which infestations had deleterious effects. Such genotypes can be sources of
resistance and tolerance traits which can be exploited in breeding for elite genotypes. Mwenje
(2006) reported high cross compatibility between cultivated Sorghum spp and their wild
relatives. This provides the opportunity for gene flow between S. arundinaceaum and
cultivated sorghums. Rich et al., (2004) found wild sorghum lines that rarely stimulated the
development of haustoria in Striga. Currently it is not known whether the Zimbabwean wild
sorghum ecotypes are resistant, tolerant or susceptible to Striga species such that their role in
the spread of Striga species is unknown.
Genetic variation for virulence can enable the parasite to adapt to new host resistance alleles.
Hence a better understanding of the virulence variability of S. asiatica populations is essential
for deployment of resistant varieties in integrated Striga control. According to Bozkurt et al.,
(2014), geographic distance plays a more important role in population differentiation than
specialization to a host species. The high evolutionary potential of the parasite populations
was also reported by Karltz and Shykoff (1998). Information on the existence of

7
physiological speciation and the stability of tolerance to various S. asiatica accessions is
unavailable Zimbabwe but can enhance efforts to breed elite sorghum varieties with broad
spectrum and durable resistance through better understanding of the host parasite interactions.
Musimwa et al., (2001) observed wide genetic distance among S.asiatica strains sourced in
Zimbabwe.
Production of low germination stimulants results in low numbers of Striga asiatica
attachments, thus producing a resistant phenotype. Jamil et al., (2011) found significant
variation in strigolactones production in New Rice for Africa (NERICA) and the low
producers gave resistant phenotypes. Information is not available for Zimbabwe on the
resistance through reduced strigolactones production and the link between strigolactones and
tillering.
1.3 Main objective
The overall objective of this research was to examine the stability of tolerance of Striga
asiatica infested Sorghum spp under drought, mulch and different Striga strains. Also to
determine the existence of reduced strigolactones production as a resistance mechanism, and
establish its link to sorghum tillering.
1.4 Specific objectives
The objectives of this research were:
1. to determine the effect of drought stress and Striga asiatica infection on the stability
of morpho-physiological traits which impart tolerance to Sorghum bicolor and
Sorghum arundinaceum;
2. to determine the effect of mulching on Striga infected sorghum growth, tillering and
dry matter partitioning;

8
3. to determine the variable effects of two Striga asiatica strains on the stability of
tolerance of S. bicolor varieties and S. arundinaceaum; and
4. to quantify strigolactones produced in Sorghum bicolor and Sorghum arundinaceaum
and correlate it to sorghum tillering.
1.5 Alternate Hypothesis
1. Reduced water availability and Striga asiatica infestation have an effect on the
morpho-physiological traits of Sorghum bicolor and Sorghum arundinaceaum.
2. Mulch reduces S. asiatica incidence on S. bicolor and S. arundinaceaum and
enhances the expression of tolerance traits in sorghum.
3. There are variable effects of S. asiatica strains sourced from geographically distant
places on sorghum varieties and sorghum is able to maintain its tolerance in the face
of physiological speciation of S. asiatica.
4. There is pre-attachment resistance through reduced strigolactones production in
Sorghum bicolor varieties and Sorghum arundinaceaum.

9
References
1. Agrawal AA (2011). Current trends in evolutionary ecology of plant defence.
Functional Ecology 25: 420 – 432.
2. Badu-Apraku B, Akinwale RO, Oyikunle M (2014). Efficiency of secondary traits in
selecting for improved grain yield in extra-early maize under Striga-infested and
Striga-free environments. Plant Breeding 133: 373 – 380.
3. Badu-Akrapu B, Fakorede MAB, Menkir A, Lum, A, Obeng-Antwi, K (2006).
Multivariate analysis of the genetic diversity of forty seven Striga resistant tropical
early maize inbred lines. Maydica 51: 551 – 559.
4. Berner DK, Kling JG, Singh BB (1995). Striga research and control- a perspective
from Africa. Plant Disease 79: 652 – 660.
5. Bozkurt ML, Muth P, Parzies HK, Haussmann BIG (2014). Genetic diversity of East
and West Africa Striga hermonthica and virulence effects on a contrasting set of
sorghum cultivars. Weed Research 55: 71 – 81.
6. Burdette AL (2007). Nutraceutical uses of sorghum bran (Sorghum bicolor). Doctor
of Philosophy thesis. The University of Georgia. Graduate Faculty. 176 pages.
7. Bouwmeester HJ, Matusova R, Sun Zhongkui and Beale MH (2003). Secondary
metabolite signaling in host-parasitic plant interactions. Current Opinions in Plant
Biology 6: 358 – 384.
8. Cardoso C, Ruyter-Spira C, Bouwmeester HJ (2011). Strigolactones and root
infestation by plant parasitic Striga Orobanche and Phelipanche spp. Plant Science
180: 414 - 420

10
9. Carsky, RJ, Singh L and Ndikawa R (1994). Suppression of Striga hermonthica on
sorghum using a cowpea intercrop. Experimental Agriculture 30 (03): 349 – 358. Doi
10.1017/50014479700024467.
10. De Groote H, Wangare L, Kanampiu F, Odando M, Diallo A, Karaya H (2008). The
potential of herbicide resistant maize technology for Striga control in Africa.
AgriSystems 97: 83 - 94
11. Ejeta G (2005). Integrating biotechnology, breeding and agronomy in the control of
the parasitic weed Striga spp in sorghum. In Tuberosa, R, Philips, R.L., Gate M (eds).
Proceedings of the International Congress. In the wake of the double helix: from the
green revolution to the gene revolution. 27 – 31 May 2003, Bologna. Italy 239 –
2005.
12. Ejeta G (2007). The Striga scourge in Africa: a growing pandemic. In Integrating new
technologies for Striga control: towards ending the witch hunt (Ejeta G, Gressel J
Eds), pp 3 – 16. World Scientific publishing company Private Limited.
13. Ejeta G and Butler, LG (1993). Host parasite interaction throughout the Striga
lifecycle and their contribution to Striga resistance. African crop Science Journal 1:
75 – 80.
14. Ejeta G, Mohammed P, Rich A, Merlake-Berhan TL, Housely TL, Hess DE (2000).
Selection for specific mechanisms of resistance to Striga in sorghum. In breeding for
Striga resistance in cereals. Proceedings of a workshop held at IITA, IBADAN,
Nigeria. (Eds BIG Haussman, Hess DE, Koyama LM, Grivet L, Rattunde HFW,
Geiger HH, (29 – 37). Margravelag. Wekersheim, Germany.

11
15. FAO (1996). FAO production Yearbook, 1996 Volume 50, FAO statistics series
number 135, Rome, ITALY, 536 pages.
16. Gurney AL, Grimanelli D, Kanampiu FK, Hoisinhton DA, Scholes JD, Press MC
(2001). Can wild relatives of cereals provide new sources of resistance to the parasitic
angiosperm Striga? In Proceedings of the seventh International parasitic weed
Symposium (Eds A Fer, P Thoalouran, Joel DM, Mussellman LJ, C Parker and J
Verleif. 140 nantes. France.
17. Gurney AL, Press, MC and Scholes JD (2002). Can wild relatives of sorghum provide
new sources of resistance or tolerance against Striga spp. Weed Research 42: 317 –
324.
18. Gurney AL, Grimanelli D, Kanampiu F, Hoisington D, Scholes DJ, Press MC (2003).
Novel sources resistance to Striga hermonthica in Tripsacum dactyloides, a wild
relative of maize. New Phytologist 160: 557 – 568.
19. Gurney AL, Ransom JK and Press MC (1995). The parasitic angiosperm Striga
hermonthica can reduce photosynthesis of its sorghum and maize hosts in the field.,
Journal of Experimental Botany 46: 1817 – 1823.
20. Hartmann, T (2008). The lost origin of chemical ecology in the late 19th
century.
Proceedings of the Natural academy of Sciences, USA, 105: 4541 – 4546
21. Haussmann BIG, Hess DE, Reddy BVS, Mukuru SZ, Kayentao M, Weltz HG,
Geiger, HH (2001). Quantitative genetic parameters of Sorghum growth under Striga
infestation in Mali and Kenya. Plant Breeding 120: 49 - 56
22. Hearne SJ (2009). The Striga conundrum. Pest Management Science 65: 603 – 614
23. Jamil M, Rodenburg J, Charnikhova T, Bouwmeester HJ (2012) Pre-attachment
Striga hermonthica resistance of new Rice for Africa (NERICA) cultivars based on
strigolactone production. New Phytologist 192: 964 – 975.

12
24. Jamil M, Charnikhova T, Houshyani B, van Aast A, Bouwmeester HJ (2011). Genetic
variation in strigolactones production and tillering in rice and its effects on Striga
hermonthica infection. Planta DOI 10.1007/s00425-011-1520y.
25. Joel DM, Hershenhorn Y, Eizenberg H, Aly R, Ejeta G, Rich PJ, Ranson JK,
Sauerborn J, Rubiales D (2006). Biology and management of weedy root parasites.
Horticultural reviews 38. John Wiley and Sons.
26. Lagoke, ST, Parkinson V and Agunbiade, RM (1988). In Proceedings of the
International workshop. Kim, S.K (Ed), Ibadan, IITA, ICRISAT and IDRC, Nigeria.
27. Karltz O and Shykoff JA (1998). Local adaptation in host-parasite systems. Heredity
81: 361 – 370.
28. Kim, SK (1994). Genetics of maize tolerance of S. hermonthica. Crop Science 34:
900 – 907.
Mabasa S (2003). Nitrogen and the control of Striga: timing and dose of organic and
inorganic forms. A PhD thesis submitted to the University of Sheffield, United Kingdom.
29. Manyong VM, Alene AD, Olanrewaju A, Ayedun V, Rweyendala AS, Wesonga G,
Omanya HD, Mignouna, Botanga M (2007). Baseline study of Striga control using IR
maize in western Kenya. http//aatf-africa.org.pdf
30. Mutengwa C (2004) The search for molecular markers, parental characterization and
inheritance studies of witch weeds (Striga asiatica Kuntze) resistance in Sorghum
bicolor. D. Phil Thesis, University of Zimbabwe, Crop science Department.

13
31. Mwenje E (2006). Sorghum and its wild relatives: ecological implications in the
event of the release of transgenic sorghum in SADC region. Biotechnology, Biosafety
and Environment in South Africa 2: 5 - 6
32. Musimwa C, Tongoona P, CS Mutengwa and Chivinge OA (2001). Genetic variation
in witchweed inferred from RAPD-PCR markers. African crop Science Conference
Proceedings, Volume 5: 51 – 55. African crop Science Society.
33. Oswald A, Ransom J.K, Kroschel J and Sauerborn J (2002). Intercropping control
Striga in maize based farming systems. Crop protection 21 (5): 367 – 374. DOI
10.1016/50261-2194(01)001041
34. Parker C (2012). Parasitic weeds: A world challenge. Weed science 60: 269 - 276
35. Parker C (2009). Observations on the current status of Orobanche and Striga
problems worldwide. Pest management Science 65: 453 – 459
36. Pennissi E (2009) Plant genetics: how sorghum withstand heat and drought. Science
323: 573.
37. Rich PJ, Greinier C and Ejeta G (2004). Striga resistance in the wild relatives of
Sorghum. Crop Science 44: 2221 – 2229.
38. Rodenburg J and Bastiaans L(2011). Host plant defence against Striga spp:
reconsidering the role of tolerance. Weed Research 51: 438 – 441
39. Rubiales. D, Verkleij. J, Vurro. M , Murdoch, A.J and Joel, D.M (2009). Parasitic
plant management in sustainable agriculture. Weed Research 49: 1 – 5.
40. Scholes JD and Press MC (2008). Striga infestation of cereals crops-an unresolved
problem in resource limited agriculture. Current opinion on Plant Biology 11: 180 –
186

14
41. Stringer L.C, Dyer J.C, Read M.S, Dongil A.J, Twymen C, Mkwambisi D (2009).
Adaptations to climate change, drought and dersertifications: local insights to enhance
policy in southern Africa. Environmental Science Policy,
doi10.1016/jenvsci.209.04.0002
42. Suzuki N, Revero RM, Shulaev V, Blumwald E, Mittler, R (2014). Abiotic and biotic
stress combinations. New Phytologist 203: 32 - 43
43. Tari, G; Lasckay G, Takacs Z, and Poor P (2013). Response of sorghum to abiotic
stresses: a review. Journal of Agronomy and Crop Science 199: 264 – 274.
44. Timko MP, Huang K, Lis KE (2012). Host resistance and parasitic virulence in
Striga-host plant interactions: a shifting balance of power. Weed Science 60: 307 –
315.
45. Yoneyama Kaori, Xie X, Kusumoto D, Sekimoto H, Sugimoto Y, Takeuchi Y and
Yoneyama Koichi (2007). Nitrogen deficiency as well as phosphorus deficiency in
sorghum promotes the production and exudation of 5-de-oxystrigol, the host
recognition signal for arbuscular mycorhizzal fungi and root parasites. Planta 227:
125 – 132
46. Yoneyama K, Awad A.A, Xie X, Yoneyama Kaori and Takeuchi Y (2010).
Strigolactones as germination stimulants for root parasitic plants. Plant and cell
physiology 51(7): 1095 – 1103

15
CHAPTER TWO: LITERATURE REVIEW
2.1. Introduction to literature review
The literature review provides a detailed background to the issues studied in this thesis. The
major aim of this study was determination of the stability of tolerance traits of Striga asiatica
infested Sorghum bicolor and Sorghum arundinaceaum under drought conditions, under
mulch, and under two different Striga asiatica strains sourced in Zimbabwe. The study also
sought to quantify strigolactones produced by sorghum varieties as a resistance mechanism
and also to find the relationship between strigolactones quantities and sorghum tillering. It
was thus necessary to explore literature on sorghum production in Zimbabwe, the genus
Striga, the extent of the S. asiatica problem in sub Saharan Africa and Zimbabwe, Striga
research in Zimbabwe, Striga life cycle and management and functions of strigolactones. The
mechanisms of resistance and tolerance were also explored, including Striga management,
sorghum response to stress and mulching. The discussions on Striga management were
biased towards subsistence agriculture as it was an important goal of this study to enhance
productivity in this agricultural sector.
2.2 Sorghum production in Zimbabwe
Sorghum bicolor is an important staple crop in Africa, Asia and central America. It is the fifth
major cereal crop after wheat, rice, maize and barley. Sorghum is adapted to tropical and
subtropical climates but the greatest area of the crop is cultivated in drought prone semi-arid
tropical environments with 400 – 600 mm rainfall that are too dry for maize (Mwenje, 2006).
Sorghum bicolor is one of the most drought tolerant small cereal grain crop grown under the
smallholder sector in Zimbabwe’s natural farming region III, IV and V (Mutengwa, 2004)
which covers more than 70 % of the area in Zimbabwe. These regions are the driest in
Zimbabwe, receiving less than 500 mm of rainfall per year. The crop is grown traditionally in

16
Zimbabwe to ensure food security even in drought years. Sorghum is therefore a calorie
source for the millions who reside in Zimbabwe’s semi-arid areas. The major producing areas
are Matebeleland North, Matebeleland South and Masvingo provinces. The Mashonaland
province produces only 10 % of the total tonnage in Zimbabwe. The production of sorghum
in Zimbabwe has been on the increase since 2010 for about five years (Table 2.1) with the
exception of 2012 when there was a decrease caused by reduced sorghum prices by the Grain
Marketing Board.
Table 2.1: Zimbabwe sorghum production by year from 2010 - 2014
Year Tonnage (MT) Growth rate (%)
2010 74 000 5.71
2011 95 000 28.38
2012 65 000 -31.58
2013 69 000 6.15
2014 95 000 37.68
Source: FAOSTAT 2015
2.3 The genus Striga
Striga is a latin word for ‘witch’ presumably because plants infested by Striga display stunted
growth and an overall drought like phenotype long before the weed appears. The genus was
previously grouped within the family Scrophulariaceae but more recent analysis has placed
Striga under the family Orobanchaceae (Ejeta, 2007; Spallek et al., 2013). Striga possibly
originates from a region between Semien mountains of Ethiopia and the Nubian Hills of
Sudan (Atera and Itoh, 2011). The same is the centre of cultivated sorghum which is the
major host species for several Striga spp (Spallek et al., 2013).

17
The main agriculturally important Striga spp in cereal crops are S. hermonthica (Del) benth
and S. asiatica (L.) Kuntze, while S. gesneroides (Willd) Vatke is a major pest in cowpeas.
Striga aspera (Willd) Benth and S. forbesii are also significant problems in cereals in limited
locations (Parker, 2009). The distribution of various Striga spp across the African continent
is shown on Table 2.2.
2.3.1 Striga hermonthica
This is the most damaging of all Striga spp affecting most staple cereal crops grown in
Africa. It has purple flowers. It is found mainly in the tropical and northern sub tropical
regions from Gambia to western Kenya, Tanzania and Ethiopia in the East (Parker, 2009).
According to Mohammed et al., (2001) S. hermonthica is widespread in sub Saharan Africa
and is found throughout west Africa to Ethiopia and east Africa.S. hermonthica is particularly
harmful to sorghum, maize and millet but is also found in sugarcane and rice fields (Atera
and Itoh, 2011).
2.3.2 Striga asiatica
Striga asiatica has red flowers and occurs sporadically across west Africa but becomes pre-
dominant species towards the east African coast, southwards to southern Africa (Parker,
2009). This is the Striga spp that wrecks havoc in Zimbabwe. Kroschel (1999) reported that a
countrywide survey in Malawi found 63 % of maize fields to be infested with this species. De
Groote et al., (2008) reported that 10 % and 25 % of the maize crop in Namibia and in
Angola, respectively, was infested with this species. A survey by Mabasa (1994) in
Zimbabwe found that 79 % of the farmers reported that this Striga species was present in
their fields.

18
2.3.3 Striga gesneroides
Striga gesneroides occur widely in Africa mainly on wild plants only although it has been
found in cowpeas in west Africa (Parker, 2009). It has been found in northern Zimbabwe
infesting cowpeas and weeds.
Table 2.2: The distribution and occurrence of Striga spp in sub-Saharan Africa
Striga spp Host Plants Distribution
Striga
asiatica
Rice, Sorghum and
maize
Angola, Lesotho, Malawi, Mozambique, Namibia,
Tanzania, Madagascar, South Africa, Zanzibar,
Zambia, Botswana, Burundi, DRC, Zimbabwe
Striga aspera Rice, maize, sorghum,
finger millet, wild
grasses and sugarcane
Burkina Faso, Cameroun, Central African republic,
Ethiopia, Gambia, Guinea, Cote’divoire, Nigeria,
Niger, Mali, Ghana, Senegal and Sudan
Striga
forbesii
Sorghum, sugarcane,
maize, rice
Angola, Botswana, DRC, Ethiopia, Kenya, Malawi,
Mozambique, South Africa, Sudan, Swaziland,
Tanzania, Uganda, Zambia, Zimbabwe
Striga
gesneroides
Cowpeas, and legumes,
Nicotiana spp,
Eurphobia spp and
Ipomoea spp
Angola, Botswana, Burkina Faso, Cameroun,
Central Africa Republic, DRC, Ethiopia, Sierra
Leone, Senegal, South Africa, Tanzania,
Zimbabwe, Gambia, Ghana, Kenya, Malawi, Mali,
Mozambique, Somalia, Nigeria, Rwanda, Uganda,
Zambia.
Striga
hermonthica
Maize, millet, rice,
sorghum,Pearl millet,
Fingermillet, sugarcane
Angola, Cameroun, Central Africa Republic,
Djibouti, Eritrea, Gambia, Guinnea Bissau,
Ethiopia, Nigeria

19
2.4 Extent of the Striga problem in sub-Saharan Africa
Parasitic weeds are fast becoming a major constraint to many crops in sub Saharan Africa and
yet the efficacy of available means to control them are minimal. Parasitic weeds have become
one of the greatest biological constraints to food production in the drier parts of Africa,
probably a more serious problem than insects, birds or plant diseases. The C4 cereals
consisting of maize, sorghum and millet are the preferred hosts and the infection of these
plants by Striga spp can result in severe grain losses. S hermonthica causes up to 100 % grain
yield losses. Losses associated with this pest are estimated to be more than US$7 billion in
sub Saharan Africa only (Berner et al., 1995) with most affected being the resource poor
subsistence farmers (Gurney et al., 2006). According to Rubiales et al., (2009), typical yield
losses vary from 15 to 20 % at a regional level, but can be more severe at local scales,
sometimes resulting in total crop failure. The losses largely depend on the level of infection,
stage of crop growth when infestation occurs, crop variety, soil fertility and rainfall (Menkir
and Kling, 2007).
In addition to the yield losses estimated to exceed US$7 billion in value, the weed also causes
adverse effects on the welfare and livelihoods of over 100 million people in Africa (Rubiales
et al., 2009). The most affected are the resource poor, small scale subsistence farmers and
severe infestations cause serious food shortages. There are contrasting statistics on the extent
of the Striga infestations by individual species but according to Mwakaboko (2003), 40 % of
arable land in sub Saharan Africa and 67 % of the 73 million hectares in cereal zones is
infested by Striga. Harsh conditions mean that few alternative crops can be grown, and the
use of high-cost inputs such as herbicides is generally not affordable to the resource poor
farmers. Sorghum is generally grown by small scale farmers whose cash investments in crop
production are low. According to Webb and Smith (1996), inputs are low, rainfall is erratic
and soils are poor. These conditions are likely to be the most suitable for the Striga epidemic.

20
In other areas, the weed has reached epidemic proportions, presenting a desperate scenario to
small scale farmers. Where the scenario has worsened to these proportions, the farmers are
left with no option except to abandon the land. According to Evans et al., (2012),
demographic pressure has led to monocropping, thus increasing the frequency of Striga spp
host crops in the cropping system, an ideal condition for Striga to thrive.
Ahmed et al., (2001) reported that research in Africa on the control of Striga has been going
on for >70 years and despite these efforts, limited success has been achieved. Effective
control of Striga has been elusive due to the fact that the weed produces thousands of seeds
that can remain viable in the soil for a long time, combined with the complicated mode of
parasitism where vascular connections occur underground (Midega et al., 2013). This is
partly due to the complex life cycle of Striga, which is intimately linked to its host and
depends on the response to chemical and tactile cues, posing a challenge to control both
before and after attachment to the host.
2.5 Striga problem in the smallholder sector of Zimbabwe
Zimbabwe is a land locked country which lies entirely in the tropics. It is situated between
15o30
I and 30
o05
I east longitudes. It has been divided into five agro-ecological zones- I, II.
III, IV and V defined largely by rainfall distribution (Figure 2.1) (Vincent and Thomas,
1961). The agricultural regions are therefore an indication of the agricultural potential of the
various regions in Zimbabwe (Table 2.2) (Rambakudzibga, 2000). A summary of the rainfall
characteristics of the five agro-ecological zones of Zimbabwe and the suitable agricultural
activities are shown in Table 2.3.
21
Figure 2.1: The Natural Farming Regions of Zimbabwe

22
Table 2.3: Agro-ecological regions of Zimbabwe and agricultural activities carried out
in the various regions.
Agro-
ecological
region
Area
(Km2)
% of
total
Rainfall characteristics Agricultural activities
I 7.000 2 More than 1050 mm per
annum with some rain in all
months
Specialized diversified farming region.
Suitable for forestry, temperate fruit
production and intensive livestock
production
II 58.
600
15 700 – 1050 mm confined to
summer. Infrequent heavy
rainfall. Subject to seasonal
droughts
Flue–cured tobacco, maize, soyabean, cotton,
sugar beans and coffee can be grown.
Sorghum, groundnuts, seed maize, wheat and
barley can be grown. Wheat and barley are
grown in winter under irrigation. Mixed
cropping with poultry, beef and dairy
production very common in the region.
III 72.
900
18 500 – 7000 mm per annum.
Infrequent heavy rainfall.
Subject to periodic seasonal
droughts, prolonged mid
season dry spells and
unreliable starts of the season.
A semi-intensive farming area. Smallholder
farmers occupied 39 % of this prior to land
reform and most of the land was used for
intensive ranching. Maize production
dominated commercial production. Irrigation
played an important role in sustaining crop
production in commercial farming areas
IV 147.
800
38 450- 600 mm per annum Suitable for intensive ranching and wild life
management. Too dry for successful crop
production and most crops suitable are

23
sorghum and millets and other drought
tolerant crops. Maize is commonly grown
under smallholder farmers. Sugarcane and
cotton are produced under irrigation in large
estates
V 104.
400
27 Normally less than 500 mm per
annum
Extensive ranching and wildlife management
are the most suitable activities
TOTAL 390.
700
100
Source: Rambakudzibga, 2000.
Zimbabwe is considered an agricultural country because the majority of the population
indirectly or directly depends on agriculture (Jasi and Mabasa, 2001). Prior to the land reform
program, most of the large scale commercial farmers were located in Natural Regions I and II
which are high potential areas for crop production. At independence, nearly 75 % of the
communal farmers were located in natural regions IV and V (Whitlow, 1980) and up to now
most of the communal areas are located in these regions. Crop production in these low
rainfall areas is risky, yields are low and they experience periodic crop failure.
Soils in the communal areas are mainly light sands with low fertility because they contain
low levels of nitrogen and phosphorus (Mashiringwani, 1983). Farmers in these areas have
limited access to capital for the purchase of inorganic fertilisers, pesticides and seed.
Agriculture in the small scale farming area is, therefore, characterized by unfavourable soil
and rainfall as well as constraints in input procurements. Striga is a problem mainly in this
farming sector.

24
2.6 History and distribution of Striga asiatica in Zimbabwe
Striga asiatica was first recorded in Zimbabwe in 1916 when it was found in farms around
Mazowe District (Weinmann 1972). Weinmann (1975) reported that 22 % of the total land
area in Mazowe District was infested between 1929 – 1930. A weed survey conducted in
1970 in the commercial farms showed that Striga asiatica was only a problem in isolated
areas in Mazowe (Thomas, 1970). Subsequent surveys done showed that Striga asiatica was
a problem in both the commercial sector (Chivinge, 1983) and the smallholder sector
(Chivinge, 1988; Mabasa, 1993).
In Zimbabwe, the weed is mostly found in the low rainfall areas where 75 % of the rural
farmers reside (Mabasa, 1993). In a survey done by Chivinge (1988), Striga asiatica was
found infesting maize, sorghum, pearl millet and finger millet in all the then eight provinces
of Zimbabwe. Chivinge (1988) reported the weed as the second most aggressive weed in
Mashonaland Central and third most aggressive in the Midlands and Masvingo Provinces in
the large scale commercial farms.
Another survey by Mabasa (1994) showed that 79 % of the farmers reported that Striga was
present in their fields. Mabasa (1993) noted that Striga is a significant production constraint
in three of the five agro-ecological regions (III, IV and V) where nearly 75 % of the
communal farmers are based. Efforts to combat the Striga should therefore be intensified to
come up with sustainable means of combating the weed.
2.7 Striga asiatica research in Zimbabwe
According to Timson (1945), Striga asiatica research in Zimbabwe dates back to the 1930s
and 1940s. After realizing the threat of Striga in the 1920s, an experimental farm was set up
in the Concession-Glendale area in the Mazowe valley in the 1930s with the aim of carrying
out both experimental and demonstrative work on Striga asiatica control (Anon, 1938).

25
Between 1945 and 1980, there was no research that was done on Striga because it was
assumed that the problem had been solved (Mabasa, 1993). This could be because research
by then targeted mostly white commercial farmers and neglected the black farmers who
might have been suffering due to the effects of the weed.
In 1986-87, the International Crop Research Institute for the Semi-Arid Tropics (ICRISAT)
initiated Striga research and they specifically looked at the screening of sorghum cultivars for
resistance and tolerance to Striga forbesii (Mabasa, 1993). A survey done in 1988 by the
Weed Research Team (Agronomy Research Institute) found that 79 % of the interviewed
farmers reported that Striga was in their fields with the highest infestations in Zaka and
Chiwundura (Jasi and Mabasa, 2001).
Research was initiated on the management of Striga and agronomic trials were conducted by
the Weed Research Team on the effects of planting dates, herbicides (Dicamba), manure and
fertiliser on Striga in maize sorghum and millet. Early planted crops had the highest Striga
infestations than late crops and maize was found to tolerate Striga when manure was applied
at 30 tonsha-1
and nitrogen at 90 – 140 kgha-1
(Agronomy Institute, 1988/89). The level of
manure and fertiliser was found to be beyond the reach of Zimbabwe’s smallholder farmers.
The tested herbicide was Dicamba (3,6 dichloromethoxybenzoic acid) and it was found to be
effective in the control of Striga but it was unaffordable to the small scale farmers and the
numeracy that was needed for calibration made it complex for the smallholder farmers.
Maize cultivars that were tested for resistance were found to be susceptible. For sorghum,
cultivars SAR29, SAR33, SAR35, SAR37 and SAR16 supported low Striga asiatica counts
compared to SV1 and SV2 (Mabasa, 1996). However, their yields were very low. There was
therefore a need to improve the yielding capability of the resistant varieties which were
otherwise the least preferred by farmers.

26
A comprehensive study by Musambasi (1997) compared herbicide 2.4 dichlorophenoxyacetic
acids (2.4 D) and 3.6 dichloromethoxybenzoic acid (dicamba) to hand weeding at 2 and 5
weeks after crop emergence and found that dicamba resulted in the best control of Striga
asiatica and 2.4 D was effective in suppressing the weed at 12 weeks after crop emergence
and that hand weeding was uneconomic. In the same study, intercropping with Vigna
subterranean and Vigna unguiculata, field beans (Phaseolus vulgaris) and groundnut
(Arachis hypogeal) resulted in reduced Striga asiatica numbers. Another study by Mabasa
(2003) revealed that the source and timing of nitrogen application did not influence Striga
asiatica tolerance in maize and recommended supportive hand weeding to prevent build up of
seeds in the weed seed bank.
There are various constraints in the use of available technologies which should be taken into
account when working with smallholder farmers. Most technologies will need 3 – 4 years
before there is any appreciable reduction in Striga infestation. No technology has been able to
eliminate Striga hence research continues. Consequently, some farmers have come to accept
the problem as something they have to live with.

27
2.8 Striga asiatica lifecycle
Figure 2.2: The life cycle of S. asiatica (Rich and Ejeta, 2007).
2.8.1 Dormancy and conditioning
Striga seeds must go through a phase of conditioning before they can be able to germinate
(Spallek et al., 2013). If no strigolactone is received during this time, the Striga seeds will
eventually fall into secondary dormancy (Cardoso et al., 2011). Germination is linked to the
presence of the host that is nearby as the endosperm of Striga can sustain growth for only 3 –
7 days (Berner et al., 1995).
After ripened Striga seeds may not germinate until they have passed through a pre-
conditioning process (Figure 2.2). According to Ejeta and Butler (1993), peak germination of
S asiatica seed occurs in vitro after 10 – 15 days of soaking in water at a temperature of 28
oC. According to Sun et al., (2008), preconditioning strongly affects the responsiveness of

28
seeds to the stimulants. Pre-conditioning at 30oC releases the dormancy within 2 – 3 weeks
and increases the sensitivity to strigolactones by several orders of magnitude.
2.8.2 Germination
The biology of Striga parasitism at its various stages is a series of signal exchanges between
the host and the parasite that leads to successful establishment (Rich and Ejeta, 2008). The
germination of Striga is tightly defined by spatial relations with potential hosts. Striga seed
germination is reflected by the distance from the host root where strigolactones are still
active, that is, concentrated enough to elicit germination (Fate et al., 1990). Striga seeds can
not germinate without specific germination stimulants released by the potential host (Shen et
al., 2006). The strigolactone concentrations required to stimulate germination are in the
concentration range of 10-10
– 10-15
MoleM3 (Cechin and Press, 1994). Strigolactones have
been shown to induce germination of Striga at concentrations that are as low as 10-16
M
(Mussellman, 1980). Hearne et al., (2008) reported that the concentration dependent spatial
limitation of Striga seed germination ensures nearness to a potential host. This is critical
because once germinated, the seedlings will lose their capacity to form competent haustoria
within 3 – 7 days (Berner et al., 1995). This is because Striga seed has small amounts of food
reserves which can only support limited growth and the Striga would die in a few days
without attaching to the host (Shen et al. 2006).
2.8.3 Haustorium development
After seed germination has been triggered, the radicle of the germinating seed penetrates the
host root and forms haustoria to establish a xylem to xylem connection with the host to
withdraw water and nutrients (Jamil et al., 2011). A haustorium is a multifunctional organ
that attaches to a host, establishes a xylem and/or phloem continuum, a physiological bridge
between the parasite and its host, directs the unidirectional flow of resources to the parasite
and functions at multiple stages in the parasitism (Estabrook and Yoder, 1998). Generally,

29
haustorium formation consists of three phases, initiation (formation of the haustorium
primordial), invasion (the penetration of haustorium primordial) and maturation (the
establishment of xylem continuum linking the host to the parasite vacular systems
(Musselman and Dickison, 1975). The radicle tip grows chemotropically towards potential
host roots after germination.
Haustoria are the invasive structures that develop at the tips of Striga radicle in response to a
host root contact. Certain phenolics, flavonoids and quinines have been identified that induce
haustorium development when added to the parasite in vitro (Riopel and Timko, 1995).
Chang and Lynn (1986) reported that the only haustorium inducing factor isolated directly
from the host root is 2.4-dimethoxy-p-benzoquinone (DMBQ). DMBQ is a product of lignin
oxidation and decarboxylation of phenolic acids found in plant cell walls (Spallek et al.,
(2013). Within hours after the parasite roots come into contact with haustorial initiation
factors, the growth and division of cortical cells is altered, resulting in localized swelling that
develops into haustorium (Yoder, 1999). Xylem formation only occurs upon contact with the
host stele (Yoder, 1998). Striga therefore elicits the host to produce a signal necessary for
parasite development in the process called semagenesis (Keyes et al., 2007). A synthetic
haustorium initiation factor known as syringic acid has been made in the laboratory. Within
24 hours after contact, rapid cell division of the radical tip stops and a hypertrophic growth
phase begins (Hood et al., 1998). Penetration of the host epidermis is mediated by the
elongation of distal cells in the protoderm or epidermis and the underlying tissue, followed by
rounds of periclinal and anticlinal divisions of the cells leading to the growth into the cortex
of the host plants (Spallek et al., 2013).
The haustorium is a physical and physiological connection between the parasite and its host
and its interaction with host tissues is important in the translocation of molecules (Aly, 2013).
The haustoria initially adheres to the host root by a secreted, mucilaginous substance (Joel

30
and Kesner-Gashen, 1974) and penetrates by pushing between the host cells. Penetration is
aided by digestive enzymes secreted by the parasite that include pectin methylesterase,
polygalacturonase and endocellulase (Benhod et al., 1993, Losner-goshen et al., 1998). In
general, penetration is completed 40 – 72 hours after contact with a host root (Hood et al.,
1998). Molecular translocation between host and parasite ranges from the movement of sugar
(Aber et al., 1983), herbicides (Joel et al., 1995, Nandular et al., 1999) to movement of
proteins (Hamamouch et al., 2005; Aly, 2007).
2.8.4 Nutrient transfer
Hibberd and Jeschke (2001) reported that most of the parasitic plants make a beeline for their
host’s vascular system and they are able to tap into a large flux of amino acids, organic
sugars, ions and water in the host’s xylem vessels or sugars, ions or amino acids in the host’s
phloem. Press and Whittaker (1993) found that most of the hemi-parasites have very high
transpiration rates and that can be double that of their host especially under low moisture
availability. This then creates a strong mass flow that facilitates transfer of water, carbon,
nitrogen and mineral nutrients from the host plant. Shen et al., (2006) reported that the
stomata of the parasite remains more open than that of the host and results in higher
transpiration rates and lower water potentials in the parasite than the host. This was also
confirmed by Press (1995). Low water potentials are maintained by the biosynthesis of
polyhydric alcohols like mannitol (Press and Graves, 1995) in the parasite.
2.8.5 Establishment of parasitism and flowering
Once xylem to xylem connections are established, the cotyledons of Striga enlarge and break
free from the seed coat within 24 hours (Hood et al., 1998). The Striga grows adventitious
roots and are able to form secondary haustoria on the same host. Once a strong sink has
developed between the host and the parasite, water and nutrients begin to flow and this

31
damages the crops’ development by reducing crop yield. When the Striga plant starts to
photosynthesise, the low carbon dioxide fixation and high dark respiration rates of Striga
asiatica result in negative carbon gain over a 24 hour period, thus making the weed
dependent on the host when growing above ground (Press et al., 1987). Striga leaves are
characterised by degenerated palisade cell layers and a relatively small number of
chloroplasts per cell (Spallek et al., 2013). This leads to lower photosynthetic rates by Striga.
A stem develops and emerges above ground to flower and disseminate seed after
approximately 4 – 7 weeks of growth (Aly, 2013). Flowers of Striga asiatica are red and after
pollination, seeds mature within 4 weeks in seed pods, which contain 250 – 500 of dust-like
seeds of 200 – 300 µm in size. According to Berner et al., (1995) under optimal conditions
each Striga plant can produce between 50 000- 500 000 seeds. Eventually the seed pods open
up and the seeds spread on the soil adding to the seed bank.
2.9. Strigolactones in plants
The frequently asked question is why plants exude strigolactones that enables their
locationby parasitic plants. Xie et al., (2010) concluded that the strigolactones may
have other roles that outweigh the risks of parasitism. According to Akiyama et al.,
(2005), Akiyama and Hayashi (2006) and Akiyama et al., (2007) strigolactones
function as branching factors for the symbiotic abuscular mycorrhizae (AM) fungi
from which plants benefit. The AM fungi are soil inhabiting microorganisms that
form symbiotic associations with plants. The fungi penetrate and colonise plant roots
where they develop highly branched arbuscules which serve as nutrient exchange
sites. According to Xie et al., (2010) the fungi supply their host with water and
nutrients, especially nitrogen and phosphorus that are obtained through the hyphae
that is found on the outside in the soil.

32
In the absence of the host, the AM hyphae differentiate into morphological structures
characterized by extensive branching (Buee et al., 2000). Strigolactones therefore appear to
trigger a cascade of molecular and cellular events necessary for hyphae to become more
effective. In host plants, production of strigolactones is stimulated by phosphate shortages
(Yoneyama et al., 2007a). This demonstrates that strigolactones are molecules for use in the
successful establishment of plants with hyphal fungi- plant symbiosis.
Strigolactones have also been implicated in fungal/host shoot branching (Umehara et al.,
2008). When there is a host plant root in the vicinity of the germinating spores, signaling
molecules are released by the roots into the rhizosphere and they reach the hyphae and the
fungus responds to this with increased growth and intensive hyphal branching.
During symbiosis, the AM fungi obtain carbohydrates from the host plant and at the same
time the host obtains water and minerals from their fungal partners enabling them to perform
better under stressful conditions (Lopez Raez, et al., 2008a). This probably explains why
strigolactones are produced inspite of the risk of infection by parasitic plants. It is likely that
the parasitic plant has evolved the capacity to perceive the presence of a host plant by taking
advantage of the already existing signaling between plants and AM fungi.
Strigolactones have been classified as a new group of plant hormones that inhibit shoot
branching by preventing the outgrowth of leaf auxiliary buds. Like other plant hormones, the
strigolactones require a receptor in order to exert their activity; they are active at low
concentrations and can be transported over some distance (Umehara et al., 2008). Increased
synthesis of strigolactones under low phosphate and nitrogen conditions leads to the
proposition that under low inorganic phosphate conditions, plants increase biosynthesis of

33
strigolactones to reduce shoot branching and increase root growth to maximize interactions
with AM fungi to facilitate the uptake of mineral nutrients (Lopez Raez et al., 2008b).
2.10 The parasite as a sink
There is evidence that parasitic plants generally obtain carbon and nitrogen from their hosts
(Press, 1989; Press, 1995). However, the level of dependence may differ. For example hemi-
parasitic species that are facultative can survive in the absence of a host whilst holo-parasitic
angiosperms rely exclusively on their hosts for their carbon and nitrogen supplies (Press,
1995). The Striga parasite has roots and pigmented leaves and that suggests that the root
hemi-parasite may receive and incorporate water and solutes from both autotrophic and
heterotrophic sources (Press, 1996).
Mabasa (2003) reported that the rates of photosynthesis in hemi-parasites are generally
towards the low end as observed in C3 species and are usually lower than those of their hosts.
Lowly concentrated chlorophyll, poorly developed mesophyll cells and fewer air spaces
between the spongy mesophyll cells (Press et al., 1988) and low levels of ribulose 1,5-
biphosphate carboxylase (Press et al., 1986) in some root hemi-parasite may account for the
low rates of carbon dioxide fixation. According to Graves et al., (1989), carbon budget
models suggest that S hermonthica would be unable to maintain any appreciable positive
carbon balance in the absence of carbon from the host. This makes the parasite a strong sink
for the carbohydrates manufactured by the hosts.
2.11 Mechanisms of sorghum tolerance to Striga asiatica
2.11.1 Host photosynthesis
Host responses to infestation cannot be attributed solely to resource competition between the
host and the parasite (Gurney et al., 1999). Differences in dry matter accumulation between

34
infested and uninfested plants partly results from the parasite acting as a sink for carbon,
inorganic solutes and water, particularly in the later stages of infection, but also lower rates of
carbon gain by the infested cereals (Cechin and Press, 1993; Gurney et al., 1995; Smith et al.,
1995).
A study by Gurney et al., (2002) found lower rates of photosynthesis for all parasite-host
associations. However, the same study revealed that the tolerant Sorghum arundinaceaum
was not affected by the parasite with respect to dry matter accumulation. This demonstrated
the uncoupling of photosynthesis and biomass accumulation as was observed by Frost et al.,
(1997) in some sorghum–Striga associations.
In a study by Van Aast and Bastiaans (2006), it was found that two sorghum varieties
Tiemarifing and CK60-B had differential response to infection by Striga asiatica.
Tiemarifing’s reduction in panicle weight was proportional to overall reduction in total plant
dry weight but for CK60-B panicle weight was much more strongly reduced in total weight
than total dry mass. It was then concluded that the ability to maintain a constant harvest index
under Striga infestation conditions might be another aspect of tolerance in this cultivar.
According to Haussmann et al., (2001), genetic variation for tolerance to Striga under field
conditions exist in cultivated sorghum especially in local African cultivars. The different
responses of wild sorghum to infection by S. hermonthica and S. asiatica suggest that these
Striga spp possibly influence host growth through different mechanisms or differential
disruption of host metabolism.
Aly (2007) reported export of proteins from the host plant to the parasite. This movement of
may lower protein availability hence chlorophyll.Therefore the ability of a genotype to
maintain chlorophyll content irrespective of the infection is key to tolerance. Since
chlorophyll concentration is connected to nitrogen availability, chlorophyll concentration

35
becomes a key parameter in the measurement of plant canopies and subsequently
carbohydrate accumulation.
2.11.2 Sorghum tillering as Striga tolerance mechanism
Tillering is generally recognized as one of the most plastic traits affecting accumulation of
biomass and ultimately grain yield in many field crops (Kim et al., 2010a). Tillering is one of
the most important agronomic traits in poaceous crops and plays a major role in determining
plant architecture and grain yield (Wu et al., 1998). Tillering confers environmental plasticity
to grain crops. High tiller production capacity improves the chances of persistence after
periods of unfavourable conditions during which a plant may experience biotic and abiotic
stress (Assuero and Togretti, 2010). According to Bartholomew (2009), when a tiller dies as a
consequence of stress, it is replaced by another tiller in order to keep forage production. The
control of tillering is affected by endogeneous factors and environmental supplies (Mcsteen,
2009). According to Hammer et al., (2006), genetic variation in tillering affects the dynamics
of canopy development. Yoshida (1976) reported that one of the most critical characteristics
of successful high yielding varieties for rice and wheat is a semi-dwarf plant type with high
tillering ability.
Although tillering is considerably less in Sorghum bicolor, it nonetheless has a major
influence on plant leaf area development (Larfage et al., 2002). Modern sorghum hybrids
produce from zero to four fertile tillers under field conditions such that at plant densities
below 4 m-2
, around 70 – 80 % of the total plant leaf area and grain yield is attributable to
tillers. Kim et al., (2010a) asserted that the differences in tillering could be associated with
differences in carbon supply-demand balance and the propensity to tiller could possibly be
associated with hormonal signaling. Differences in tillering could be caused by differences in
hormonal signaling or responsiveness to sugar level. Studies done have found novel
hormones that trigger branching and are known to affect tiller growth in sorghum (George-

36
Jaeggli, 2009). Therefore, the capacity of a variety to produce tillers is important when
considering tolerance to Striga.
2.11.3 Environmental regulation of tillering
Studies by Kim et al., (2010b) found that the rate of appearance of successive leaf tillers and
the leaf appearance rate were similar to the appearance rate of leaves of the main shoot.
Therefore, the environmental effects on plant phenology can be important in explaining
tillering dynamics in terms of topological location, appearance and fertility frequency.
Environmental conditions affect tillering through their effects on the carbohydrate
supply/demand framework. Environmental effects on sorghum tillering have been reported
by Kaitaniemi et al., (1999) and were due to changes in leaf elongation rate in response to
temperature, vapour pressure deficit and water availability. In sorghum that is under parasite
attack, there is intense nitrogen competition and carbon usage is more prominent. For a
variety that uses tillering as a tolerance mechanism, the tillers should therefore appear after
the Striga has completed its life cycle. Striga generally takes a total of 10 – 12 weeks to
complete its life cycle. Once the Striga is dead and new tillers come up they are likely to
result in more yield as there is an extensive root system that will have been established.
Assuero et al., (2000) found that tiller number was reduced by endophyte infection in two
Fastuca arundinacea that had been infested by two fungal endophyte strains Neotyphodium
coenophilum strains. The same should be applied to plants parasitised by Striga as they lead
to resource competition.
2.12.1 Propensity to tiller
Studies by Kim et al., (2010b) revealed that high tillering was also associated with the
propensity to tiller. This is associated with hormonal signaling and responsiveness to sugar
level. Umehara et al., (2008) identified the hormone strigolactone that triggers branching.

37
The hormones are known to affect tiller outgrowth in sorghum (George-Jaeggli, 2009).
Research by Jamil et al., (2011) in rice cultivars found that low tillering rice cultivars have
lower strigolactones in the root exudates compared to high tillering.
2.13 Host plant resistance against Striga
Host resistance to parasitic plants is a multicomponent process that occurs at different stages
of a parasite’s life cycle (Estabrook and Yoder, 1998). Rich and Ejeta (2008) reported that the
biology of Striga at its various stages is a series of signal exchanges between host and
parasite that leads to successful establishment. Yoder and Scholes (2010) combined the
various stages of parasite development into the three general periods: pre-attachment, parasite
establishment and post attachment maturation. Parasitic plants use chemical communication
in the rhizosphere to trigger certain stages to occur in their life cycle. Therefore any genetic
differences in the biosynthesis or release of signaling molecules can reduce parasite viability.
The types of resistance occurring in parasitic plant hosts are classified into reduced host
germination stimulant production, reduced haustorium initiation factor production, host
resistance to parasite establishment and host resistance after parasite establishment.
2.13.1 Reduced host stimulant production
According to Jamil et al., (2011), there is a promising opportunity to minimize losses through
avoiding triggering of Striga seed germination through reduced strigolactone production.
Low production of host plant root exudates compounds that are essential for Striga
germination is the understood mechanism for Striga resistance (Lynn and Chang, 1990). In a
study by Sun et al., (2008), and Jamil et al., (2011), germination of Striga hermonthica seed
was dependent upon the quantity and quality of strigolactone production and any genetic
variation in this trait could potentially confer pre-attachment resistance. In a study by Jamil
et al., (2011), rice varieties CG14, WAB56-104 and NERICA 1 produced the smallest
amounts of strigolactones. The same varieties were shown to have differences in the

38
composition of the strigolactone blend. In another study, Jamil et al., (2011b) found that the
concentration of strigolactones in root exudates varied strongly in rice cultivars and that in
the study, Super Basmati, TN1, Anakila and Agee produced the least strigolactones. Low
germination stimulant genotypes of sorghum have enhanced resistance to Striga because of
the reduction in Striga germination (Ejeta, 2007). Low germination stimulant was also
implicated in some legume and sunflower accessions that showed enhanced resistance to
Orobanche species.
According to Ejeta et al., (2001) all susceptible Sorghum spp appear to be high stimulant
producers. Vogler et al., (1996) found that low germination stimulant (lgs) production of
sorghum is controlled by a single recessive gene. According to Ejeta et al., (1997), the lgs
gene found in a source germplasm has been transferred into high yielding and broadly
adapted sorghum cultivars.
2.13.2 Reduced haustorium inducing factors (Low haustorial factor)
Haustoria are the structures that invade the host root and causes the formation of xylem-
xylem connections. Riopel and Timko (1995) identified that phenolics, flavonoids and
quinones as compounds that induce haustorial development.
The success of parasitic plants results largely from strategies that tightly couple
developmental transitions with host recognition signals. This means in the absence of specific
signals by a potential host, successful infestation by the parasite is prevented. Studies by
Gurney et al., (2003) found that Tripscum dactyloides, a wild relative of maize does not
produce primary haustorium inducing factors as a small number of parasites initiated
haustorial formation. Rich and Ejeta (2004) found sorghum lines that rarely developed
haustoria.

39
Low haustorial factor (Lhf) is inherited as one dominant gene. According to Ejeta (2005) the
lhf gene found in sorghum has been transferred into improved sorghum cultivars and has
been pyramided with other resistant genes.
2.13.3 Parasite establishment resistance
In some cultivars, the parasite penetrates the host root cortex but is unable to form vascular
continuity with the host and it dies (Gurney et al., 2005). The same study revealed that by day
9 after infection, the parasite could not form a parasite-host xylem-xylem connection. In
many cases, the endophyte passed straight through the root cortex and emerged from the
other side of the root. Vascular continuity allows movement of water and nutrients from host
to parasite. It also provides factors required for further differentiation of the haustorium.
In a study by Gurney et al., (2006), the haustorium of parasites attached to the sorghum
variety Noponbare did not mature and differentiate. Their results indicated that by day 21, 49
% of the attached parasites were either dead or showed signs of necrosis. In other sorghum
varieties (Framida and Dobbs), a proportion of the parasites died owing to the onset of the
rapid hypersensitive reaction (Mohamed et al., 2003).
2.13.4 Post-attachment resistance
Resistance can occur following successful connection of parasites to the host vascular system
(Yoder and Scholes, 2010). Perez-de-Luque et al., (2006) found that in some pea genotypes,
host vascular cells fill with mucilage-like compounds that block the transport of nutrients to
the parasite leading to death of tubercules.
2.13.5 Incompatible response (IR)
This response is similar to the hypersensitive response in that it discourages the development
of Striga beyond attachment. There is however no apparent necrosis in host root tissue
surrounding the attachment site. In the IR based resistance, Striga seedlings that penetrated

40
the host tissue may not develop beyond first emergence of leaf primordia (Ejeta, 2005). Some
Striga appear to develop normally at first but show signs of stunted growth with time.
2.13.6 Hypersensitive response
In some sorghum genotypes, necrotic areas appear on roots at the site of Striga attachment.
Ejeta (2005) reported that necrotic lesions start as red becoming brownish with time and can
spread to 2 mm from the centre of attachment but most remain more localized. This
discourages further advancement of attached Striga which does not develop normally and
eventually dies on the host. Sorghum varieties with this phenomenon have been observed and
they include Framida, CK32 and CK33 (Ejeta, 2005). Hypersensitive reaction is conditioned
by two complementary dominant genes.
2.14 Managing Striga asiatica in sub-Saharan Africa
Despite the concerted efforts to come up with a sustainable method of Striga asiatica control,
there is no sustainable control method available for managing the parasite. It remains as the
number one biological constraint limiting the production of cereal grains in sub-Saharan
Africa. Several methods have been studied but have been accompanied by minimum success.
The numerous methods that have been suggested or developed together with their technical
limitations are shown in Table 2.4.
Table 2.4: Different methods of controlling Striga asiatica
Control
strategy
Mechanism Limitation References
Increased soil
fertility
Shortage of nitrogen
and phosphorus leads to
more active production
of strigolactone to
Most of the
affected farmers
cannot afford the
levels of fertiliser
Williams (1961), Raju et al., (1990)
Mabasa (2003), Jamil et al., (2011),
Lopez-Raez et al., (2008),
Yoneyama et al., (2007a, 2007b))

41
attract mychorhizal
fungi for increased
nutrient absorption.
required to reduce
production of
strigolactones
Intercropping
with legumes
The mechanism
according to Fernandez-
Aparicio et al., (2011)
involves smothering by
cover crops and
increase in soil moisture
and fertility for legumes
and decrease soil
temperature.
Jasi and Mabasa (2001),
Rambabkudzibga and Mabasa
(1993), Kabambe (1991),
Musambasi (1997), Kasembe
(1995), Mbwaga (1995), Adipala et
al., (1997), Kuchinda et al., (2003),
Carsky et al., (1994), Oswald et al.,
(2002), Khan et al., (2006), Midega
et al., (2010), Picket et al., (2010),
Khan et al., (2008) Tsanuo et al.,
2003; Hooper et al., 2010.
Trap crops These crops stimulate
germination of the
parasitic weed but is not
compatible with
subsequent infection
process
Trap crops cannot
exhaust the seed
banks and their
cultivation may not
be economic.
Rao and Gacheru (1998), Oswald et
al., (1996), Odhiambo and Ransom
(1996).
Catch crops These are true hosts that
promote Striga
germination but will be
burnt or chopped down
as soon as Striga
germinates
Not economic for
the farmer
Oswald et al., (1997)
Host
resistance or
Low strigolactone
production, low
This is a
sustainable
Jamil et al., (2011), Gurney et al.,
(2003), Rich and Ejeta (2008), El

42
tolerance haustorial initiation
factors, plugging of
xylem-xylem
connection between
host and parasite and
antibiosis.
approach but not
even one sorghum
cultivar has shown
complete
resistance.
Heweris (1987).
Herbicide
seed dressing
Herbicide coated seed
maize uses germplasm
resistant to
Imidazolinone group of
accetolactate synthase
(ALS) inhibiting family
of herbicides.
Successful so far Samb and Chamel (1992), Chivinge
et al., (1999), Odhiambo and
Ransom (1993), Kanampiu et al.,
(1997), Manyong et al., (2008),
Kanampiu et al., (2002)
2.15. Sorghum response to drought stress
About a third of the world’s agricultural land currently suffers from chronically inadequate
water availability (Ghannoun, 2009). Adaptations to drought is a quantitative trait controlled
by many genes (Fenta et al., 2012). According to Mitchell et al., (2013), plants exposed to
low intensity but long duration droughts may maintain water status above the critical water
potential thresholds but deplete stored carbohydrates. High intensity drought and incapability
to regulate plant water status above critical thresholds will lead to plant death. The ability of a
plant to maintain high rates of photosynthesis is an important determinant of the ability of a
crop plant to maintain growth and indicates tolerance of a crop to drought and Striga
infections.
Fenta et al., (2012) found differences in root biomass under water replete conditions with one
variety showing the greatest biomass accumulation. The same trend was demonstrated for

43
root biomass while no differences were shown in shoot/root biomass. The same study found
that tolerance was often concentrated largely on shoot parameters, particularly those
associated with photosynthesis. Gilbert et al., (2011) found that considerable genetic
differences exist in the ability of soyabeans to maintain high water use efficiency and
photosynthesis during drought. Genetic differences in stomatal conductance are considered to
exert the greatest effect on the intrinsic water use efficiency (Gilbert et al., 2011). The
stability of photosynthesis under conditions of water deprivations is also considered to be an
important aspect of drought tolerance.
The sorghum plant has the capability of withstanding drought conditions through adjustments
in stomatal aperture, maintenance of cell turgor, hydraulic conductivity and maintenance of
cell photosynthesis. However, the introduction of an extra sink from the parasite may alter
this delicate balance. Even if sorghum varieties may be tolerant to drought, the occurrence of
drought to a host attached to the parasite may aggravate the situation. The ability of a
sorghum plant to withstand both the parasite and drought may be compromised.
2.16 Existence of Striga physiological speciation and differential virulence
A lot of Striga management systems have been suggested, including introduction of resistant
sorghum varieties. According to Mohammed et al., (2007), crops with some measure of
resistance have been integrated into Striga management programmes and the new material
gets challenged by the Striga seed bank. The newly introduced sorghum variety must be able
to cope with the great potential genetic diversity in the seed bank. One of the major problems
in Striga control is the possible existence of physiological speciation within the species of
Striga as was first suggested by Jones (1955). Ten years later, Doggett (1965), was the first
one to report varietal differences in sorghum susceptibility to Striga hermonthica in east
Africa. It was observed that varieties of sorghum that were resistant in one location became
susceptible in another suggesting the existence of physiological strains of the parasite.

44
According to Bozkurt et al. (2015), genetic variation for virulence can enable the parasite to
adapt to new host resistance alleles and several studies have been undertaken to understand
the parasite’s genetic diversity and variability for virulence.
Rao and Musselmann (1987) suggested the possible causes of the differential response to be
Striga intensity differences, instability of host resistance or other several soil and
environmental factors which affect resistance of the host cultivars. Bebawi (1981)
investigated the germination response of eleven Striga hermonthica samples from sorghum
and pearl millet exudates of 27 sorghum cultivars and confirmed the suspiscions of
physiological speciation in Striga which had been alluded to by Parker and Reid (1979).
Botanga and Timko (2006), suggested that geographical isolation and host driven selection
are important factors in the formation of races of Striga gesneriodes in West Africa. In Striga
asiatica and Striga hermonthica populations from Kenya, 96.8 % and 84 % of the Amplified
fragment length polymorphism (AFLP) bands were polymorphic respectively (Gethi et al.,
2005). The observed polymorphism indicated that different strains were in each of the Striga
populations. A study in Benin discovered a high degree of host speciation within the 14
analysed Striga asiatica populations (Botanga et al., 2002). According to Spallek et al.,
(2013), Striga asiatica isolated from wild grasses species were unable to successfully
parasitise sorghum and maize plants susceptible to other Striga asiatica collections. The same
authors found a significant relationship between genetic and geographic distances in Striga
species.
2.17 Mulch effects on Striga asiatica incidence
There is a growing worldwide concern about soil health, usage of fossil fuels and overall
economics of field crop production. Conservation agriculture (CA) is being promoted

45
globally as a farming system that can address many of those concerns and increase the overall
economic productivity of mechanised agriculture (Norsworthy and Oleivera, 2007; Hobbs et
al., 2008; Sanyal et al., 2008). Conservation agriculture is a broad term, which encompasses
activities such as minimum and zero tillage, tractor powered, animal powered and manual
methods, integrated soil and water management and includes conservation farming. It is
generally defined as any tillage sequence that minimises soil and water loss and achieves at
least 30 % soil cover by crop residues.
According to Bilalis et al., (2003), the correct use of residues helps to conserve moisture and
deal with weeds while improving soil structure. Teasdale and Mohler (1993) indicated that
annual weed infestation decreases as crop residues increase on the soil surface offering direct
evidence that crop residues control weed emergence. The effect of the surface mulch on weed
seedling emergence is controlled by several factors such as quality of residue, weed species,
position in the soil, soil type and environmental conditions. According to Liebman et al.,
(2001), the mechanisms for weed suppression include changes in soil moisture, reduced
temperatures at the soil surface, physical impedance and allelopathy.
Weed response to residue depends on quality, quantity of residue and biology of a particular
weed. Residues of small grains have been shown to inhibit weed emergence and growth in
cropping systems (Putnam et al., 1983). Mulch therefore exerts a selection pressure on weeds
and the weeds that possess traits that enable them to germinate on the surface will increase in
numbers.
Crop residues used as mulch substantially decrease maximum daily soil temperature (Mohler
and Teasdale, 1993). Differences in temperature between mulched and unmulched plots can
be as great as 14 oC. Mulch shades weed seeds from light and prevents shoot growth. The
presence of light might indicate proximity to the soil surface or absence of overstorey canopy

46
and assurance for successful establishments and growth. Mohler and Teasdale (1993)
reported that large quantities of residues were needed to effect weed control as total weed
emergence paralleled light transmittance through the residues. The ability of mulch to
suppress weeds is also correlated to light extinction coefficient of the mulch. However no
study has sought to determine the effects of mulch on Striga asiatica establishments.
2.18 Photosynthesis in a Striga-infested plant
According to Gurney et al., (1999), an increased understanding of the parasitic mode of
nutrition has demonstrated that host response to infestation cannot be attributed to more
competition between the host and the parasite. Reduced photosynthesis may account for the
lower rates of host productivity. Graves et al., (1989) found that 80 % of the biomass not
accumulated by the host compared with uninfested plants can be accounted for by lower
carbon fixation in infested plants compared to controls. Gurney et al., (2002) suggested that
an ability to maintain high rate of photosynthesis while infested may be an important factor
of tolerance to the parasite.
According to Frost et al., (1997) lower rates of photosynthesis are a consequence of lower
stomatal conductance, particularly in the early stages of infection. Increased levels of the
plant growth regulator abscissic acid in infested plants cause stomatal closure (Taylor et al.,
1996; Gurney et al., 1997). The effects of plant parasites on their hosts vary from
undetectable levels where a plant proceeds to reproduction as if there is no pest to extreme
cases where the host dies. Some Striga-cereal associations result in stunting of the internode
elongation leading to unpacking of the leaves within the canopy (Walting and Press, 1997,
1998).

47
2.18.1 Stomatal conductance
One of the shoot related physiological traits that may affect stress tolerance is the decline in
whole plant water use during soil water deficit events (Manavalan et al., 2009). In most
cases, stomatal conductance affects leaf gas and water vapour exchange. According to Liu et
al., (2003), drought stress decreases relative leaf expansion rate, stomatal conductance and
leaf turgor whereas it increases abscissic acid content in the leaf and xylem. There are
obviously genetic differences in the ability to keep stomata open despite the presence of
stress in some plants. The control of stomatal conductance under stress is a physiological trait
that can be exploited when developing drought resistant materials in sorghum.
Adjustments in stomatal arpeture act to reduce transpiration in response to the declining
hydraulic conductance and/or reductions in leaf turgor and help plants to avoid water
potentials that can induce hydraulic failure (Sperry, 2000). Phosphorus and potassium are the
most essential nutrients. Ruiz-Lozamo et al., (1995) asserted that in particular, potassium
plays a key role in plant water stress and has been found to be the cationic solute responsible
for stomatal movement. Ge et al., (2012) found that the decline in nutrient uptake especially
P and K was because of low transpiration rate under soil water deficit. This had earlier been
observed by Sinha (1978) that stress tolerant wheat varieties can accumulate more potassium
than susceptible ones and that plants well supplied with potassium have higher stomatal
resistance which resulted in low transpiration rates.
2.19 Conclusions
A comprehensive up-to-date review of literature was conducted. In this review, Striga was
found to cause devastating effects on sorghum production. The life cycle of Striga spp was
discussed paying attention to its multi-stage nature, and depedance on host signals. The
current management methods were reviewed together with the limitations of each of them.

48
Tolerance was determined as central to Striga management since there is no single genotype
that has been found to be completely resistant. However, there is limited information on the
response of sorghum spp to various ecological factors such as mulching, drought and the
existence of physiological speciation in Striga.
References
1. Adipala E, Byabagambi S, Osiru D.S.O (1997). Effect of weeding frequency on the
growth and yield of cowpea. Pages 107 – 111 in Proceedings of the 16th
Biennial East
African Weed Science Society (1997). Kampala. Uganda.
2. Ahmed N.E, Sugimoto Y, Babiker A.G, Mahomed O.E, Ma Y, Inanaga S (2001).
Effects of Fusarium solani isolates and metabolites on Striga germination. Weed
Science 49: 354 – 358
3. Atera E , Itoh K (2011). Evaluation of ecologies and severity of Striga weed on rice in
sub Saharan Africa. Agricultural and Biology Journal of North America 2: 752 - 760
4. Aly R (2013). Trafficking molecules between parasitic plants plants and their hosts.
Weed Research 53: 231 - 241
5. Akiyama K (2007). Chemical identification and functional analysis of apocarotenoids
involved in the development of arbscular mycorhizal symbiosis. Bioscience,
Biotechnological and Biochemistry 71: 1405 – 1414
6. Akiyama K, Hayashi H (2006). Strigolactones: chemical signals for fungal
antisymbionts and parasitic weeds in plant roots. Annals of Botany 97: 925 – 931

49
7. Akiyama K, Matsuzaki K, Hayashi H (2005). Plant sesquiterpenes induce hyphal
branching in arbsucular mycorhizal fungi. Nature 435: 824 – 827
8. Assuero SG, Tognetti JA (2010). Tillering regulation by endogeneous and
environmental factors and its agricultural management. The Americas Journal of plant
science and biotechnology 4 : 35 – 48
9. Assuero SG, Matthew C, Kemp PD, Latch GCM, Baker DJ and Haslett S (2000).
Morphological and physiological effects of water deficit and endophyte infection on
contrasting tall fescue cultivars. New Zealand Journal of Agricultural Research 43: 49
– 61
10. Berner DK, Kling JG, Singh BB (1995). Striga rsearch and control: a perspective
from Africa. Plant Disease 79: 652 - 660
11. Bartholomew PE (2009) Growth of the bgrass plant. Kwazulu-Natal department of
Agriculture. Available at http://agriculture.kzntl.gov.za/publications. Accessed on
12. Bozkurt ML, Muth P, Parzies HK, Hausmann BIG (2015). Genetic diversity of east
and West African Striga hermonthica populations and virulence effects on a
contrasting set of sorghum cultivars. Weed Research 55: 71 - 81
13. Buee M, Rossignol M, Jauneau A, Ranjeva R and Becard G (2000). The presymbiotic
growths of absucular mycorhizal fungi is induced by a branching factor partially
purified from plant root exudates. Molecular Plant Microbe Interactions 13: 693 –
698
14. Bebawi FF, Eplee RE, Harris CE (1984) Longevity of witch weed (Striga asiatica)
seeds. Weed Research 32: 494 - 497

50
15. Botanga CJ., Timko MP (2006). Phenetic relationship among different races of S
gesneroides (Willd) vatke from west Africa. Genome 49: 1351 - 1365
16. Botanga C, Kling J, Berner D, Timko M (2002). Genetic variability of Striga asiatica
based on AFLD analysis and host parasite interactions. Euphytica 128: 375 - 388
17. Bilalis D, Sidiras N, Economou G, Vikali, C. (2003). Effect of different levels of
wheat straw surface coverage on weed flora of Vicia faba. Journal of Agronomy and
Crop Science 189: 233 - 241
18. Cardoso C, Ruyter-spira C, Bouwmeester HJ (2011). Strigolactones and root
infestation by plant parasitic Striga, Orobanche and Philipanche spp. Plant Science
180: 414 – 420.
19. Cechin I, Press M.C (1994). Nitrogen relations of the sorghum-Striga hermonthica
host-parasite association: germination, attachment and early growth. New Phytologist
124: 681 – 687
20. Carsky, RJ, Singh L, Ndikawa R (1994). Suppression of Striga hermonthica on
sorghum using a cowpea intercrop. Experimental Agriculture 30 (03): 349 – 358. Doi
10.1017/50014479700024467.
21. Chang M, Lynn D.L. (1986) The haustorium and parasitic angiosperm. Journal of
Chemical Ecology 12: 561 - 579
22. Chivinge O.A (1983). A weed survey of arable lands oin the commercial farming
sector of Zimbabwe. Zimbabwe Agricultural Journal 80 (4): 139 – 141
23. Chivinge O.A (1988). A weed survey of arable lands in the small scale farming sector
of Zimbabwe. Zambvezia XV (ii): 167 - 179

51
24. Chivinge O.A, Musambasi D, Mariga I.K (1999). The effect of 2.4D, dicamba and
hand weeding on witch weed density and crop yield. African Crop Science Journal 7
25. De Groote H, Wangare L, Kanampiu F, Odando M, Diallo A, Karaya H (2008). The
potential of herbicide resistant maize technology for Striga control in Africa.
Agricultural Systems 97: 83 – 94
26. Doggett H (1965). Striga hermonthica on Sorghum in East Africa. Journal of
Agricultural Sciences 65: 183 - 194
27. Ejeta G and Butler LG (1993). Host parasite interaction throughout the Striga
lifecycle and their contribution to Striga resistance. African crop Science Journal 1:
75 – 80
28. Ejeta G (2005). Integrating biotechnology, breeding and agronomy in the control of
the parasitic weed Striga spp in sorghum. In Tuberosa, R, Philips, R.L., gate M (eds).
Proceedings of the International Congress. In the wake of the bouble helix: from the
green revolution to the gene revolution. 27 – 31 May 2003, Bologna. Italy 239 –
2005.
29. Ejeta G (2007). The Striga scourge in Africa: a growing pandemic. Integrating new
technologies for Striga control: towards ending the witch hunt. (Eds Ejeta G and
Gressel J), World Scientific publishers.
30. Evans A.A, Kazuyuki I, Tetsushi A, Takashiga I (2012). Farmer’s perspectives on the
biotic constraint of Striga hermonthica and its control in Western Kenya. Weed
biology and management 12: 53 - 62
31. Estabrook E.M, Yoder J.I (1998). Plant-plant communications: rhizosphere signaling
between parasitic angiosperms and their hosts. Plant Physiology 116: 1 – 7

52
32. El Heweris S.O (1987). Nature of resistance to Striga hermonthica (del) Benth
parasitism in some Sorghum vulgare Pers cultivars. Weed Research 27: 305 - 311
33. Fate G, Chang M, Lynn D.G. (1990). Control of germination in Striga asiatica;
chemistry of spatial definition. Plant Physiology 93: 201 - 207
34. Fenta BA, Driscoll SP, Kunrt KJ, Foyer CJ (2013) Characterisation of drought
tolerance traits in nodulated soya beans; the importance of maintaining photosynthesis
and shoot biomass under drought. Journal of Agronomy and Crop Science 178: 92 –
103.
35. Frost DL, Gurney AL, Press MC, Scholes J (1997). Striga hermonthica reduces
photosynthesis in sorghum: the importance of stomatal limitations and potential role
of ABA. Plant Cell and Environment 20: 483 – 492
36. George-Jaeggli (2009). The physiology and genetics of height, yield associations in
sorghum. PhD thesis, University of Queensland. Australia.
37. Gilbert, M.E, Zwieniecki M.A, Holbrook N.M (2011). Independent variation in
photosynthetic capacity and stomatal conductance leads to differences in intrinsic
water use efficiency in soya bean genotypes before and during mild drought. Journal
of Experimental Botany 62: 2875 – 2887.
38. Graves J.D, Press M.C and Stewart G.R. (1989). A carbon balance model of the
sorghum-Striga hermonthica host parasite association. Plant, Cell and Environment
12: 101 - 107
39.

53
40. Gurney AL, Press MC and Scholes JD (1999). Infection time and density influence
the response of sorghum to the parasitic angiosperm Striga hermonthica. New
Phytologist 143: 573 – 580
41. Gurney AL, Slate J, Press MC, Scholes JD (2006). A novel form of resistance in rice
to the angiosperm parasite Striga hermonthica. New Phytologist 169: 199 - 208
42. Gurney A.L, Ransom J.K and Press M.C (1995). The parasitic angiosperm Striga
hermonthica can reduce photosynthesis of its sorghum and maize hosts in the field.
Journal of experimental Botany 46: 1817 – 1823
43. Gurney A.L, press, M.C and Scholes J.D (2002). can wild relatives of sorghum
provide new sources of resistance or tolerance against Striga spp. Weed Research 42:
317 - 324
44. Gethi J.G, Smith S.E, Kresovich S (2005). Genetic diversity of Striga hermonthica
and Striga asiatica populations in Kenya. Weed Research 45: 64 – 75.
45. Gurney A.L, Grimanelli D, Kanampiu F, Hoisington D, Scholes D.J, Press M.C
(2003). Novel sources resistance to Striga hermonthica in Tripsacum dactyloides, a
wild relative of maize. New Phytologist 160: 557 - 568
46. Ghannoun, O. (2009). C4 photosynthesis and eater stress. Annals of botany 103: 635
– 644.
47. Ge Ti-Du, Sun Ning-BO, Bai Li-Ping, Tong Chen-Li, Sui-Gong (2012). Effects of
drought stress on phosphorus and pottassiuum uptake dynamics in summer maize
(Zea mays) throughout the growth cycle. Acta Physiologia Plantarum 34: 2179 –
2186
48. Hearne S.J (2008). The Striga conundrum. Pest Management Science 65: 603 – 614
49. Hibberd JM and Jeschkle WD (2001). Solute influx into the parasitic plants. Journal
of Experimental Botany 52: 2043 - 2049

54
50. Hood M.E, Condon J.M, Timko M.P (1998). Primary haustorial development of
Striga asiatica on host and non host spp. Phytopathology 88: (1): 70 – 75.
51. Hamamouch N, Westhood J.H, Benner I, Cramer C.L, Gepstein S, Aly R (2005). A
peptide from insect transgenic tobacco from parasitic weed. Transgenic Research 14:
227 - 236
52. Haussman B.I.G, Hess D.E, Welz H.G, and Geiger, H.H (2001). Improved
technologies for breeding Striga resistant Sorghums. Field crops Research 66: 195 -
211
53. Hammer G, Cooper M and Tardieu F (2006). Models for navigating biological
complexity in breeding improved crop plants. Trends in Plant Science 11: 587 - 593
54. Hooper, A.M, Tasnuo, M.K, Chamberlain K, Toittcomb K, scholes J and Hasanali J
(2010). Isoschaftoside, a C-glucosylflavanoid from Desmodium uncinatum root
exudate, an allelochemical against development of Striga. Phytochemistry 71 (8-9):
904 - 908
55. Hobbs P.R, Sayre K, Gupta R (2008). The role of conservation agriculture in
sustainable agriculture. Philo Trans Royal society B 363: 543 – 555.
56. Jasi L, Mabasa, S. (2001). The status of Striga (witchweed research and extension in
Zimbabwe. In Striga research in southern Africa and strategies for regionalized
control options ((eds) Mgonja M.A, Chivinge O.A, and Monyo, E.S). International
Crop research Institute for the semi-arid tropics. Pp 35 – 47.
57. Jamil M, Charnikhova T, Cardoso C, Jamil T, Ueno K, Verstappen F, Asami T,
Bouwmeeter H.J (2011). Quantification of the relationship between Strigolactones

55
and Striga hermonthica infection in rice under varying levels of Nitrogen and
phosphorus. Weed Research 51: 373 – 385
58. Joel DM (2000). The long term approach to parasitic weed control: manipulation of
specific developmental mechanisms of the parasite. Crop protection 19: 753 - 758
59. Joel D.M, Steffens, J.C, Matthews D.E (1995). Germination of weed root parasites. In
‘Seed development and germination’. Eds Kigel J, and Galili G. Marcel bekker Incl,
New York Pp 567 - 597
60. Jones, W.(1955) . Further experiments in witch weed control. 11. The existence of
physiological strains of Striga hermonthica. Empire Journal of Experimental
Agriculture 23: 206
61. Krosthel J, Sauerborn J (1996). Farming systems and the problem methods: A
comparison of case studies from Northen Ghana, Tanzania and Malawi. In Advances
in parasitic Weed research. Ed Moreno M.T, Berner D, Joel D, Musselman L.T,
Parker C. Junta de antalucia, Consejeria de agricutura Pesca.
62. Keyes WJ, Palmer AG, Erbil BWK, Taylor JV, Apkarian RP, Weeks ER, Lynn DG
(2007) Sernagenesis and the parasitic angiosperm Striga asiatica. Plant Journal 51:
707 – 716
63. Kim H.K, Luquet D, Oosterom, Dingkuhn M, Hammer G (2010a). Regulation of
tillering in sorghum: genotypic effects. Annals of Botany 106: 69 – 78
64. Kim H.K, Oostrom E.V, Dingkuhn M, Luquet D, Hammer G (2010b). Regulation of
tillering in Sorghum: environmental effects. Annals of Botany 106:57 - 67

56
65. Kaitameni P, Room P.M, Hana J.S (1999). Architecture and morphogenesis of grain
sorghum, Sorghum bicolor (L) Moench. Field Crops Research 61: 51 – 60
66. Kabambe V (1991). The development of cultural methods for the control of Striga in
maize in malawi. In proceedings of the 5th
international symposium of parasitic weeds
(eds) JK Ransom, Musselman LJ, Worsham AD, Parker C, Pp 46 – 50. CIMMYT,
Nairobi, Kenya.
67. Kasembe, E (1995). The effect of different cowpeas (Vigna anguiculata) cultivars and
time of ridging on witch weed emergence, density and maize grain yield in the
smallholder sector of Zimbabwe. Journal of Plant protection in the Tropics 12(1): 15 -
26
68. Kuchinda, N.C, Kureh I, Tarfa, B.D, Shinggu C, Omolehin R (2003). Onfarm
evaluation of improved maize varieties intercropped with some legumes in the control
of Striga in the Northern Guinnea Savvana of Nigeria. Crop Protection 22(3): 533 -
538
69. Khan, Z.R, Midega, C.A.O, Hasanali A, Pickett J.A, Wadhams L.J and Wanjoya A
(2006). Management of Witchweed Striga hermonthica and stem borers in Sorghum
bicolor, through intercropping with green leaf Desmodium intortum. International
Journal of Pest Management 52 (4) 297 – 302
70. Khan Z.R, Pickett J.A, Hassanali A, Hooper A.M and Midega C.A.O (2008).
Desmodium spp and associated biochemical traits for controlling Striga spp: present,
and future prospects. Weed Research 48 (4): 302 – 306
71. Kanampiu F.K, Ransom J.K and Gressel J (1997). Advantages of seed primed
Imazapoyr for Striga hermonthica control in Maize bearing target site resistance.

57
Pages 255 – 259: In Proceddings of the 16th
East Africa Biennial weed Science
conference
72. Kanampiu F.K, Ransom J, Friesen D and Gressel J (2002). Imazapyr and pyrithobac
movement in the soil and from maize seed coats to control Striga in Legume
intercropping. Crop Protection 21: 611 – 619
73. Liu F, Andersen, M.N and Jensen, C.R (2003). Loss of pod set caused by drought
stress is associated with water status and ABA content of reproductive structures in
soyabean. Functional Plant biology 30: 271 - 280
74. Lopez-Raez J.A, Charnikhova T, Gomez Riolden V, Matusova R, kohlen W, and De
Vos R (2008a). Tomato strigolactones are derived from carotenoids and their
biosynthesis is promoted by phosphate starvation. New phytologist 178: 863 – 874
75. Lopez-Raez J.A, Matusova R, Cardoso C, Jamil M, Charnikhova T, Kohlen W,
Ruyter-spira C, Verstappen F and Bouwmeester H (2008b). Strigolactones: ecological
significance and use as a target for parasitic plant control. Pest management science
64: 471 - 477
76. Losner-Gorshen D, Portnoy VH, Mayer, AM and Joel DM (1998). Pectolytic activity
by the haustorium of the parasitic plant Orobanche in host root. Annals of Botany 81:
319 – 326.
77. Larfage T.A, Broad I, Hammer G.L (2002). Tillering in grain sorghum over a wide
range of population densities: identification of a common hieracrh for tiller
emergence, leaf area emergence, leaf area development and fertility. Annals of Botany
90: 87 - 98

58
78. Liebman M, Mohler CL, Staver C.P (2001). Ecological management of agricultural
weeds. Cambridge University Press.
79. Mbwaga AM (1996). On farm verification of Striga control components in Tanzania.
Paper presented at the Parasitic plants: Biiology and resistance International
workshop. June 1995. Long Ashton Research station. Bristol. UK.
80. Midega CAO, Ittchar J, Salifu D, Picketti JA, Khan ZR (2013). Effects of mulching,
N fertilization and intercropping with Desmodium uncinatum on Striga hermonthica
infestation in maize. Crop Protection 44: 44 - 49
81. Mutengwa C (2004) The search for molecular markers, parental characterization and
inheritance studies of witch weeds (Striga asiatica Kuntze) resistance in Sorghum
bicolor. D. Phil Thesis, University of Zimbabwe, Crop Science Department.
82. Mohammed K.I; Musselman, L.J Riches C.R (2001). The genus Striga
(scrophulariaceae) in Africa. Annual Missouri Botannical gard
83. Menkir A, Kling J.G (2007). Response to concurrerent selection for resistance to
Striga hermonthica in tropical maize population. Crop Science 47: 674 – 684
84. Mashiringwani NA (1983). The present nutrient status of the soils in the communal
areas of Zimbabwe. Zimbabwe Agriculture Journal 80: 73 – 75.
85. Mabasa S (2003). Nitrogen and the control of Striga: timing and dose of organic and
inorganic forms. A PhD thesis submitted to the University of Sheffield, United
Kingdom.
86. Mabasa S (1994). Striga activity in Zimbabwe. Pages 143 – 146. In Lagoke, S.T.O..
Hoevers, R; M’boob, S.S. and Traboulsi (Eds). Imporoving Striga management in

59
Africa. Proceedings of the second general workshop of the pan African Striga control
netqwork (PASCON). 23 – 29 June, 1991), Nairobi, Kenya, Accra. FAO
87. Mabasa S (1996). Screening sorghum varieties for resistance to Striga asiatica in
Zimbabwe. Pages 201 – 209. Inn Leuschner, K and Manthe C.S (Eds). Drought
tolerant crops for Southern Africa. Proceedings of the SADC ICRISAT regional
sorghum and millet workshop. 25 – 29 July 1994. \Gaborone, Botswana. Patancheru,
India. Plenum Press, United Kingdom.
88. Musambasi D (1997). Maize witch weed (Striga asiatica L Kuntze) management
technologies in the smallholder sector of Zimbabwe. M Phil Thesis. Crop Science
department, University of Zimbbabwe.
89. Musselman L.J (1980). The biology of Striga, Orobanche and other root parasites.
Annual Reviews of Phytopathology 18: 463
90. Midega, C.A.O, Khan Z.R, Amudai D.M, Pittchar J and Pickett J.A (2010). Integrated
management of Striga hermonthica and cereal stem borers in finger millet (Eleucine
coracana L) through intercropping with Desmodium Intortum. International Journal
of Pest Management 56 (2): 145 – 151
91. Manavalan L.P, Guttikonda, S.K, Tran P.L, Guyen H.T (2009). Physiological and
molecular approaches to improve drought resistance in soyabeans. Plant cell
Physiology 50 (7): 1260 – 1276.
92. Manyong V.M, Nindi S.J, Alene A.D, Odhiambo G.D, Omanya G, Mignouna H.D
and Bokanga M (2008). Farmer perception of Imazapyr-Resistant (IR) maize
technology on the control of Striga in Western Kenya. African Agriculturel
Technology Foundation. Available at www.aatf-africa.org). Nairobi, Kenya.

60
93. Mohamed A, Ellicot A, Houseley T.C, Ejeta G (2003). Hypersensitive response of
Striga infection in Sorghum. Crop Science 43: 1320 – 1324
94. Musselman LJ and Dickison W.J (1975). The strudture and development of the
haustorium in the parasitic scrophulariaceae. Botanical ajournal of the Linnean
Society 70: 183 - 212
95. Mwenje E (2006). Sorghum and its wild relatives : ecological implications in the
event of a release of transgenic sorghum in the SADC region. Biotechnology,
Biosafety and Environment in Southern Africa 2 : 5 - 6
96. Nandulla V.K, Foy C.L, Orcutt (1999). Glyphosate for Orobanche aegyptiaca control
in Vicia faba and Brassica napus. Weed Science 47: 486: - 491.
97. Norsworthy J.K, Oleivera M.J (2007). Tillage and canopy effects on cocklebur
(Xanthium strumarium) emergence. Weed Science 55: 474 - 480
98. Odhiambo, G.D, Ransom J.K (1996). Effect of continuos cropping of trap crops and
maize under varying manage,ent systems on the restoration of land infested with
Striga hermonthica. In Advances in Parasitic Plant research Proceedings of the 6th
Parasitic weed symposium, Cardoba, Spain 16 – 18 April 1996. Edited by Moreno
M.T, Cubero J.I, Berner D, Joel D, Musselman L.J Parker C. Cardoba Spain, PP 834 –
842
99. Odhiambo, G.D Ransom J.K (1993). Effect of Dicamba in the control of Striga
hermonthica in Maize in Western Kenya. African crop Science Journal 1: 105 -107

61
100. Oswald A, Abayo A, Ransom J.K, Kroschel J Sauerborn J (1996). Catch
cropping with Sudan grass: an option for the control of Striga in subsistsence
agriculture. Brighton Crop Protection Conference: Weeds: 1: 227 - 232
101. Oswald A, Ransom JK, Kroschel J, Sauerborn J (2002). Intercropping control
Striga in maize based farming systems. Crop Protection 21 (5): 367 – 374. DOI
10.1016/50261-2194(01)001041
102. Parker, C (2009). Observations on the current status of Orobanche and Striga
problems worldwide. Pest management Science 65: 453 – 459
103. Press, M.C (1989). Autotrophy and heterotrophy in root hemiparasites. Trends
in Ecology and Evolution 4 (4): 258 – 263
104. Press MC and Graves JD (1995). Parasitic plants. London, Chapman Hall,
292 pp
105. Press MC, Whittaker JB (1993). Exploitation of the xylem stream by parasitic
organisms. Philosophical transactions of the Royal society of London, Serries B-
biological sciences 341: 101 – 111.
106. Press, M.C (1995). How do parasitic Striga and Orobanche influence host
carbon relations? Aspects of Applied Biology: Physiological responses of plants to
pathogens 42: 63 – 70.
107. Press M.C, Graves, J.D and Stewart G.R (1988). Transpiration and carbon
acquisition in root hemi-parasitic angiosperms. Journal of experimental Botany 39:
1009 – 1014.

62
108. Press MC, shah N, Stewart, G.R (1986). The parasitic habit: trends in
metabolic reductionism. In Biology and control of Orobanche. EDS S.J. ter Borg). Pp
96 – 106. Wageningen, The Netherlands.
109. Perez-de-Luque A, Doleres Lozamo M, Cubero J, Gonzalez-Melendi P,
Risueno M, Rubiales D (2006). Mucilage production during the incompatible
interaction between Orobanche cronata and Vicia sativa. Journal of Experimental
Botany 57: 931 - 942
110. Pickett, JA, Hamilton, ML, Hooper AM, Khan A, Midega C.A.O (2010).
Companion cropping to manage parasitic plants. Annual Reviews of Phytopathology
48(1): 161 – 177
111. Parker C, Reid, DC. (1979). Host specificity in striga species-some
preliminary observations, In proceedings of the second symposium on parasitic
weeds, musselman, LJ; Worsham, AD, Eplee RE. North Carolina State University Pp
79
112. Putnam AR, De Frank, J, Barnes J.P. (1983). Exploitation of allelopathy foe
weed control in annual and perennial cropping systems. Journal of chemical Ecology
9: 1001 – 1010.
113. Raju, PS, Osman MA, Soman P, Peacock J.M (1990). Effects of N, P and K
on Striga asiatica (L) Kunzte seed germination and infestation in sorghum. Weed
Science 30: 139 – 144
114. Rambakudzibga, A.M (2000). Aspects of the host-parasite association
between the grain legume, Vigna unguiculata (L) (WALP) and the parasitic

63
angiosperm Alectra vogelii Benth. PhD thesis der Justus-Liebig-Universitat Gieben
(2000).
115. Rambakudzibga AM, Mabasa S (1993). Aspects of Alectra vogelli and Striga
asiatica germination in root exudates of Bambara nuts {Vigna subterranean} and cow
peas (vigna anguiculkata varieties in mazoe, Zimbabwe. Agronomy Research
Institute, Henderson research institute. Zimbabwe
116. Rao, M.J.V, Musselman, L.J. (1987). Host specificity in striga spp and
physiological strains. In Parasitic weeds in agriculture olume 1 Ed Musselman L.J.;
CRC Press, Florida.
117. Rao, M.R, Gancheru E (1998). Prospects of agroforestry in Striga
management. Agroforestry Forum 9: 22 - 27
118. Rich, P.J, Ejeta G (2008). Towarsd an effective resistance to Striga in African
maize. Plants Signalling and Behaviour 3(9): 618 – 621
119. Rich, P.J, Greinier C , Ejeta G (2004). Striga resistance in the wild relatives of
Sorghum. Crop Science 44: 2221 – 2229.
120. Riopel J.L, Timko M.P (1995). Haustorial initiation and differentiation. In
Parasitic plants. Edited by Press M.C, Graves J.D. Chapman and Hall. Pages 39 - 79
121. Rubiales. D, Verkleij. J, Vurro. M , Murdoch, A.J, Joel, D.M (2009). Parasitic
plant management in sustainable agriculture. Weed Research 49: 1 – 5.
122. Samb P.I Chamel A (1992). Foliar absorption and translocation of 14C-
dicamba into host (Pearl millet) and Cow pea and parasite plants of the genus Striga.
Weed Research 32: 129 - 136

64
123. Sanyal D, Bhowmik, P.C, Anderson, R.L, Shrestha A. (2008). Revisiting the
perspective and progress of integrated weed management. Weed Science 56: 161 –
167.
124. Sinha SK (1978). Influence of potassium on tolerance to stress. In Sekhon GS
(ed). Pottassium in soils and crops. Pottassium Research Institute. New Delhi pp 223
– 240.
125. Shen H, Ye W, Hong L, Wang Z, Deng X, Yang Q, Xu Z (2006). Proress in
parasitic plant biology: host selection and nutrient transfer. Plant biology 8: 175 - 185
126. Smith, L.H, Keys A.J, Evans M.C.W (1995). Striga hermonthica decreases
photosynthesis in Zea mays through effects on leaf structure. Journal of Experimental
Botany 46: 759 - 765
127. Spallek T, Mutuku M, Shirasu K. (2013). The genus Striga; a witch profile.
Molecular Plant pathology. DOI:10.1111/mpp.12058
128. Sperry, J.S. (2000). Hydraulic constraints on plant gas exchange. Agricultural
and Forest Meterology 104: 13 – 23.
129. Steward G.R and Press M.C (1990). The physiology and biochemistry of
parasitic angiosperms. Annual Rev of plant physiology Plant Mol Biology 41: 127 -
151
130. Sun, Z, Hans J, Walter M.H, Matusova R, Beekwilder R, Vestapen F.W.A,
Ming Z, van Echtelt E, Strack D, Bisseling T, Boumeester H.J (2008). Cloning and
Characterisation of a maize characterisation of a maize carotenoid cleavage
dioxygenase (ZmCCDI) and its involvement in the biosynthesis of apocarotenoids
with various roles in mutualistic and parasitic interactions. Planta 228: 789 - 801

65
131. Taylor, A, Martin J and Seel W.E (1996). Psychology of the parasitic
association between maize and witch weed Striga hermonthica; Is ABA involved.
Journal of Experimental Botany 47: 1057 – 1065
132. Teasdale C.A and Mohler JR (1993). Interaction of light, soil moisture and
temperature in weed suppression by hairy vetch residue. Weed Science 41: 46 – 51
133. Thomas P.E.L (1970). A survey of arable weeds in Rhodesia. Rhodesia Agric
Journal 67: 25 – 42.
134. Timson S.D. (1945). Wicth weed demonstration farm, final progress report.
December, 1944. Rhodesia Agric journal 42 (5): 404 - 409
135. Tsanuo, M.K, Hassanali A, Hooper, A.M, Khan Z.R, Kabeira F, Pichett, J.A
Wadhams L.J (2003). Isoflavanones from the allelopathic aqeous root exudates of
desmodium uncinatum. Phytochemistry 64(1): 265 – 273
136. Umehara M, Hanada A, Yoshida S, Akiyama K, Arite T (2008). Inhibition of
shoot branching by new terpernoid plant hormones. Nature 455: 195 - 200
137. Van Ast A. Bastiaans, L. (2006). The role of infection time in the differential
response of sorghum cultivars to Striga hermonthica infection. Weed Research 46:
264 - 274
138. Vincent V, Thomas R.G (1961). An agricultural survey of Sothern Rhodesia.
Part 1: Ag o-ecological regions survey. Salisbury. Rhodesia. Ministry of agriculture.
139. Vogler R.K, Ejeta G, Butler L.G (1996). Inheritance of low production of
Striga germination stimulant in sorghum. Crop Science 36: 1185 – 1191.

66
140. Webb M, Smith M.C (1996). Biology of Striga in sahelian Mali: effects in
pearl millet and prospects for control. Weed research 36:203 – 211
141. Walting J.R, Press M.C (1997). How is the relationship between C4 cereal
Sorghum bicolor and C3 hemi-parasitic Striga hermonthica and Striga asiatica
affected by elevated carbon dioxide levels. Plant, cell and environment 20: 1292 -
1300
142. Walting J.R, Press M.C (1998). How does the C4 grass Eragrostis pilosa
respond to elevated carbon dioxide and infection with parasitic angiosperm Striga
hermonthica? New Phytologist 140: 667 - 675
143. Weinmann, H (1972). Agricultural research and development in southern
Rhodesia, 1890 – 1923.. Occassional paper No 4:pp 1 – 161
144. Weinmann H (1975). Agricultural research and development in southern
Rhodesia, 1924 – 1950. Serries in science Number 2: 1 – 240
145. Williams C.N. (1961). Effect of innoculum size and nutrition on the host
parasite relations of Striga senegalensis in sorghum. Plant and Soil XV: 1 - 12
146. Whitlow, J.R (1980). Environmental constraints and population pressures in
tribal areas of Zimbabwe. Zimbabwe Agric Journal 77: 173 – 180
147. Wu G.W, Wilson L.T, Mclung A.M. (1998). Contribution of rice tillers to dry
matter accumulation and yield. Agronomy Journal 90: 317 - 323
148. Xie X, Yoneyama K, Yoneyama K. (2010). The strigolactones story. Annual
reviews of phytopathology 48: 93 – 117`
149. Yoder JI (1999). Parasitic plant reponses to host plant signals: a model for
subterranean plant-plant interactions. Current Opinion in plant biology 2: 65 – 70

67
150. Yodder JI (1998). Self and cross compatibility in three spp of hemiparasite
triphysaria. Environment and experimental botany 39: 77 - 83
151. Yoder J.I and Scholes J.D. (2010). Host plant resistance to parasitic weeds,
recent progress and bottlenecks. Current opinions on plant biology 13: 478 – 484.
152. Yoshida S (1978). Physiologicalspects of grain yield. Annals of plant
physiology 23: 437 - 464
153. Yoneyama K, Yoneyama K, Takeuchi Y, Sekimoto H (2007a). Phosphorus
deficiency in red clover promotes exudation of orobanchol, the signal for mycorhizal
symbionts and germination stimulant for root parasites. Planta 225: 1031 – 1038
154. Yoneyama Kaori, Xie X, Kusumoto D, Sekimoto H, Sugimoto Y, Takeuchi Y
Yoneyama Koichi (2007b). Nitrogen deficiency as well as phosphorus deficiency in
sorghum promotes the production and exudation of 5-de-oxystrigol, the host
recognition signal for arbscular mycorhizzal fungi and root parasites. Planta 227: 125
– 132

68
Chapter Three:
The effect of witchweed (Striga asiatica L. Kuntze) and moisture
stress on selected morpho-physiological traits which impart
tolerance to Sorghum spp
3.1 Abstract
Sorghum production is hampered by the parasite Striga asiatica and recurring droughts.
However, the morpho-physiological effects of these combined stresses on tolerance of
sorghum traits are poorly understood. Two pot experiments were set up to determine the
effects of the two stresses on morpho-physiological traits of sorghum. A 2 * 2 * 5 factorial
experiment laid down as a completely randomised design replicated three times was carried
out twice at Bindura University of Science Education (BUSE) nursery. The first factor was
water availability at two levels: 50 % field capacity (FC) and 100 % FC. Infection was the
second factor at two levels: infested and uninfested, and all these were imposed on five
sorghum lines, including wild sorghum (Sorghum arundinaceaum). Sorghum chlorophyll
content, normalized difference vegetation index (NDVI) and dry matter traits were analysed
using Genstat version 14 to compare treatment effects. Watering at 100 % FC gave the higher
(P<0.01) NDVI across all the measured periods compared to 50 % FC. The results indicated
that sorghum genotypes differed sharply with respect to chlorophyll content and the NDVI
with the genotype Mukadziusaende having the most chlorophyll and NDVI (P<0.05), whilst
the least was wild sorghum. Chlorophyll content and NDVI differed significantly among
genotypes while limited water availability did not reduce chlorophyll content of

69
Mukadziusaende, wild sorghum and Chiredhi. Striga infection when it co-occurred with
water stress did not affect chlorophyll content. Mukadziusaende had the highest (P<0.05)
head weight and head index. Infestation with Striga significantly reduced (P<0.05) head
weight across all treatments. Drought stress and Striga infection had mutually exclusive
effects on chlorophyll content and NDVI. However, both infection and drought stress reduced
head weight illustrating the two factors were synergistic on their effects on sorghum head
weight.
3.2 Introduction
Sorghum is an important cereal crop that feeds more than a third of the population in southern
Africa. One of the remarkable characteristics of sorghum is its drought tolerance, that has
made it an important cereal grown for food and beverages in sub-Saharan Africa (SSA).
Among the major constraints of sorghum production are drought and infestation with Striga
asiatica. Striga is a parasitic weed that attaches itself to the roots of sorghum from where it
draws its moisture and nutrient requirements thus inhibiting host plant growth, reducing yield
and in severe cases, causing plant death. Striga affects the major crops that supply the bulk of
the carbohydrate and protein needs of the poor who reside in SSA (Ejeta, 2005). Scholes and
Press (2008) and Ejeta (2007) reported that over 50 million hectares of arable farmland under
cultivation with cereals and legumes are infested with one or more Striga species in SSA.
About a third of the world’s agricultural land currently suffers from chronically inadequate
water availability (Ghanounn et al., 2009) and this situation is predicted to worsen (Jury and
Vaux, 2007) due to climate change. Global warming, changes in rainfall abundance and
frequency and severity of rainfall events may exert a significant pressure on agricultural
water use, with several regions currently experiencing water deficits likely to face further
shortages (Padgam, 2009). Infact, many of the world’s poorest people farm in areas with
inadequate and unreliable rainfall. Even in traditionally irrigated areas, water stress is

70
becoming a serious threat to crop production due to water scarcity resulting from the growing
and competing demands for water uses. Despite all this, agricultural productivity must be
increased to provide food for the world’s ever increasing population. Future food demand for
the rapidly increasing population pressures is likely to further aggravate the effects of drought
(Somerville and Briscoe, 2001).
Under natural conditions, a combination of two or more stresses such as drought, salinity and
heat are common to many agricultural areas around the world and negatively impacts crop
productivity (Suzuki et al., 2014). Information on plant performance under a more complex
environment where multiple stresses co-occur is fragmentary (Farooq et al., 2010). Cramer et
al., (2011) asserted that the major crops of the world are likely to be exposed to a wide range
and a number of abiotic and biotic stress conditions as well as their combinations. Stress
combinations represent one of the most critical challenges facing sorghum production today
and improved theory and practice are needed for quantification of genotype responses. As an
example, studies done elsewhere have revealed that the molecular responses of plants to a
combination of heat stress and drought is unique and cannot be directly extrapolated from the
response of plants to stresses such as drought or heat when applied individually (Rhizhsky et
al., 2002, Suzuki et al., 2005; Mittler et al., 2006).
There has been contrasting responses of different plants to different stress combinations.
Demirevska et al., (2010) found that tobacco showed the same physiological responses to
drought and heat and their combinations. In barley, the effect of drought or heat stress
reduced plant growth with a more severe effect coming from drought. The combination of
drought and heat stress reduced plant growth to a much greater extent than drought or heat
applied individually (Suzuki et al., 2014). However, Iyer et al., (2013) reported that
Medicago truncatula showed contrasting responses to a combination of ozone and drought
stress. Ozone stress caused development of chlorotic and necrotic tissue and drought alone

71
caused wilting and collapse of leaves but a combination of the two stresses cancelled the
effects of both stresses. Drought leads to stomatal closure and reduce the uptake of pollutants
via stomata thereby ameliorating the effect of gaseous pollutants like ozone (Olinger et al.,
1997; Low 2006). Actually, Suzuki et al., (2014) posited that some stress combinations might
have beneficial effects compared with the occurrence of separate stresses. Understanding the
limits of stress tolerance and acclimation to stress is of great importance and practical value
in predicting the potential limit of plant productivity (Isebrands et al., 2000).
Wahid and Rasul (2005) found that the major effect of drought is reduction in photosynthetic
machinery and pre-mature leaf senescence culminating in reduction of food production.
Drought stress produces changes in photosynthetic pigments and components (Anjom et al.,
2003) and diminishes the activities of the Calvin cycle enzymes which reduce yields (Fu and
Huang, 2001). According to Cramer et al., (2011), the hormones abscisic acid (ABA) and
ethylene have been found to be important regulators of plant responses to both abiotic and
biotic stresses. Striga has been shown to increase ABA in infested maize and sorghum plants
(Frost et al., 1997; Taylor and Frost, 1997). ABA induces stomatal closure which allows a
reduction in water loss and as a consequence, the maintenance of beneficial water potential.
Farooq et al., (2010) reported that the stoma close gradually as drought progresses, followed
by the parallel decline in net photosynthesis. Studies done on maize have shown that drought
stress leads to morphological, physiological and biochemical changes, including reduced
photosynthesis (Pervez et al., 2004; Zhao et al., 2010). Drought stress frequently enhances
allocation of dry matter to the roots which enhance water uptake (Leport et al., 2006).
Although the sorghum crop has evolved appropriate stress tolerance strategies, they are
largely incompatible with the exploitative root parasitic strategy of Striga species (Tesitel et
al., 2015). Given that global change involves a series of environmental factors occurring
concurrently and changes in the severity of different stress factors; knowledge on how plants

72
acclimate to multiple stresses is of key importance in understanding the effects of the future
climate on crops (Niinemets, 2010). An urgent need to generate crops with enhanced
tolerance to stress combinations therefore exists (Suzuki et al., 2014). It is necessary to select
for sorghum genotypes with enhanced tolerance to Striga asiatica, drought, and a
combination of the stresses. To determine the response of sorghum to a combination of
abiotic and biotic stresses applied simultaneously, the effects of Striga asiatica parasite and
drought on chlorophyll content, internode length, dry matter traits and productivity of
sorghum were studied. A combination of drought and Striga stress represent conditions
encountered by many cereal crops growing in the semi-arid environments of the sub tropical
regions of Africa. It becomes necessary to select for sorghum genotypes with enhanced
tolerance to drought and Striga asiatica and their combinations to ensure food security for the
poorly resourced farmers. The objectives of this study were:
i) To determine the effect of Striga infestation and reduced water availability on
normalized difference vegetation index (NDVI), chlorophyll concentration and
sorghum height as tolerance parameters to Striga.
ii) To determine the effects of Striga asiatica infestation and water availability on
sorghum yield and dry matter partitioning of sorghum.
The associated alternate hypotheses were:
i) Striga infestation and reduced water availability, lower chlorophyll content, NDVI
and sorghum height.
ii) Striga infestation and reduced water availability reduces sorghum yield and alters dry
matter partitioning of the host species.

73
3.3 Methodology
3.3.1 Experimental site
The pot experiments were carried out at Bindura University of Science Education (BUSE)
Astra Campus nursery, Bindura (17o 18
1 58
11 S and 31
o 19
1 23
11 East). Bindura is located 89
km north of the city of Harare. The soil type used was sandy, with 4.3 % clay content and a
pH of 4.4. The area receives an annual rainfall of about 700 mm per annum, with an average
temperature of 25 oC in the summer months.
3.3.2 Seed sources
Striga asiatica seeds were obtained from Henderson Research Station (Weed Research team)
at Mazowe in Zimbabwe. The seeds had been collected from Chiwundura communal lands in
the Midlands Province in Zimbabwe from farmers’ fields in the 2009 summer season.
Sorghum seed was obtained from the gene bank at the Department of Research and Specialist
Services in Harare. Wild sorghum seeds were collected from Gwebi Agricultural College
fields, 27 km west of Harare.
3.3.3 Experimental design and treatments
The experiment was a 2 * 2 * 5 factorial experiment laid down as a randomized complete
block design replicated three times. The first factor was sorghum genotype at five levels, the
second factor was infestation at two levels, infested and uninfested. The third factor was
irrigation at two levels, 50 % and 100 % of field capacity. The moisture level of 50 % Field
Capacity (FC) was included to mimic the low rainfall areas in SSA where total rainfall is
usually below 400 mm and that is where Striga has deleterious effects. Irrigation scheduling
was done using the 100 % field capacity application. The experiment was repeated twice over
time, and denoted as experiments I and II.

74
3.3.4 Experimental procedures
Plastic pots with a height of 27 cm and diameters of 25 cm and 17.5 cm for the top and
bottom, respectively, were filled with 8 kg of soil. All pots had six drainage holes at the
bottom. Half the pots were infested with 1 gram of Striga asiatica seeds and mixed with the
top 10 cm of the soil. Fertilizer was applied at a rate of 5 g maizefert (8 N: 14 P2O5: 7 K2O)
per pot. Top dressing was done at 4 weeks after crop emergence (WACE) by applying 2.5
grammes of ammonium nitrate (34.5 % N). Ten sorghum seeds were planted and germinated
after 6 days and were thinned to one plant per pot at 2 WACE. Weeds other than Striga were
hand pulled as soon as they emerged.
3.3.5 Irrigation
The soil had its water holding capacity determined and half the pots were watered with water
that gave the field capacity (FC) and the other by half that amount. To determine field
capacity, five pots with the same oven dried soil were weighed and gradually filled with
water until the addition of any extra water created a tiny flood layer. The pots were then left
to drain freely for 48 hours and weighed again. This method was according to Kabiri et al.,
(2014). The amount required to reach field capacity was 1.5 litres per pot. The pots were
irrigated to a moisture content of 100 % and 50 % field capacity according to Webster and
Grey (2008) and Chauhan and Johnson (2010).
3.3.6 Data collection
Data collected during crop growth were: normalized difference vegetation index (NDVI),
chlorophyll content and sorghum internode length. The NDVI was measured using a
handheld green-seeker optical sensor unit (NTech industries, Inc, USA). Chlorophyll content
was measured using a chlorophyll meter (SPAD 502, KONICA MINOLTA Incl) starting

75
from 6 WACE. At the end of the experiment, head weight and total dry matter were
determined using a sensitive scale. At crop maturity, the sorghum plants were harvested and
partitioned into roots, leaves and stems. They were put in the drier at 104 oC for 48 hours for
dry matter determination using a sensitive weight scale. Total dry matter constituted the total
weights of roots, leaves, stems and head for each treatment. Head, stem, leaf and root indices
were computed as follows:
Part (e.g. roots, leaves stems) Index = Part weight/Total dry weight.
3.3.7 Statistical analysis
Data was assessed by analysis of variance using Genstat version 14. Means that were
significantly different were separated using LSD at 0.05 probability level.Barlett’s test was
applied and variances for the two experiments were not homogeneous, hence the data were
analysed separately.
3.4 Results
3.4.1 Chlorophyll concentration and NDVI
Sorghum varieties differed sharply with respect to chlorophyll concentration (P<0.01).
Across all the measured periods (6 and 10 WACE) in both experiments, the sorghum
genotype Mukadziusaende gave the highest chlorophyll content and the least was recorded
for wild sorghum (Table 3.1).

76
Table 3.1: Sorghum genotypes effects on chlorophyll content at 6 and 10 WACE
Experiment I
Chlorophyll concentration
(mmolcm-2
)
Experiment II
Chlorophyll concentration
(mmolcm-2
)
Sorghum
genotype
6 WACE 10 WACE 6 WACE 10 WACE
SC Sila 40.21±2.14a 40.5±2.025
a 38.39±2.035
a 31.04±3.152
a
Mukadziusaende 43.00±2.14a 47.23±2.025
b 42.33±2.035
b 33.55±3.152
a
Wild Sorghum 34.9±2.14b 36.3±2.025
c 32.39±2.035
c 29.78±3.152
a
Chiredhi 40.07±2.14a 42.09±2.025
a 38.49±2.035
a 32.95±3.152
a
Isifumbathe 43.77±2.14a 41.46±2.025
a 39.8±2.035
a 31.97±3.152
a
At increased moisture availability, there were significantly higher (P<0.005) NDVI values
compared to 50 % FC across the measured periods in both experiments (Table 3.2).

77
Table 3.2: Moisture stress effects on NDVI at 6 and 10 WACE
Experiment I Experiment II
6 WACE 10 WACE 6 WACE 10 WACE
100 % FC 0.525±0.067a 0.594±0.0242
a 0.59±0.0242
a 0.528±0.0267
a
50 % FC 0.464±0.067b 0.523±0.0242
b 0.523±0.0242
b 0.464±0.0267
b
At 10 WACE in experiment II, there was a significant interaction of genotype and water
availability on NDVI (Figure 3.1). For the genotype Chiredhi, higher NDVI were found at
100 % FC compared to 50 % FC. All the other genotypes had similar NDVI despite different
moisture availabilities (Figure 3.1).

78
Sorghum varieties
SC Sila
Mukadziusaende
Wild SorghumChiredhi
Isifumbathe
ND
VI
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7100 %
50 %
Figure 3.1: Interaction effects of sorghum genotype and moisture availability on NDVI at 10
WACE in Experiment II.
Chlorophyll content was not significantly affected by infection except at 6 WACE in
Experiment II (Table 3.3). Infection did not affect chlorophyll content in Experiment I and at
10 WACE in Experiment II. At 6 WACE, uninfested sorghum had a significantly higher
chlorophyll content compared to infested (Table 3.3).

79
Table 3.3: Effect of S. asiatica infection on chlorophyll content of sorghum
Chlorophyll content (mmolcm-2
)
Experiment I Experiment II
6 WACE 10 WACE 6 WACE 10 WACE
Infected 40.99±1.359a 41.54±1.295
a 36.73±1.458
a 31.35±1.994
a
Uninfected 39.79±1.359a 41.59±1.295
a 39.8±1.458
b 32.37±1.994
a
Figure 3.2: Interaction effects of sorghum genotypes and water availability on chlorophyll
concentration at 6 WACE in Experiment II.
The genotypes Mukadziusaende, Wild sorghum and Chiredhi maintained their chlorophyll
content despite variations in moisture availability. The chlorophyll concentration of
genotypes Isifumbathe and SC Sila was significantly (P<0.05) lowered by reduced moisture
availability (Figure 3.2).

80
Sorghum genotypes
Sc Sila
Mukadziusaende
Wild
Sorghum
Chiredhi
Isifumbathe
Ch
loro
ph
yll
co
nte
nt (m
mo
lcm
-2)
0
10
20
30
40
50
60
100 % FC
50 % FC
Figure 3.2: Interaction effects of sorghum genotypes and water availability on
chlorophyll concentration at 6 WACE in experiment II.
3.4.2 Internode length
Sorghum internode length was significantly lowered by infection (P<0.01). The uninfested
sorghum genotypes gave longer internode lengths compared to infested ones (Figure 3.3).
Infestation status
Infested Uninfested
Inte
rnod
e le
ngth
(cm
)
0
2
4
6
8
10
Figure 3.3: Effect of Striga infestation on internode length.

81
A significant interaction of infection and drought (P<0.01) showed that under Striga
infestation, internode length was the same both for 100 % FC and 50 % FC, whilst under non
infested conditions, 100 % FC increased sorghum internode compared to 50 % FC (Figure
3.4)
Infection status
Infested Uninfested
Inte
rnod
e leng
th (
cm
)
0
2
4
6
8
10
12
100 % FC
50 % FC
Figure 3.4: Interaction effects of Striga infestation and water availability on sorghum
internode length.
3.4.2 Dry matter traits
Striga infested sorghum significantly (P<0.05) increased both root weight and root index in
experiment I. In experiment II, infestation did not affect either root weight or root index
(Table 3.4). In both experiments, irrigation at 100 % FC increased root weight compared to
50 % FC.

82
Table 3.4: Effect of infestation and moisture availability on root weight and root index
Experiment I Experiment II
Root weight (g) Root Index Root weight Root index
Striga infested 39.6±4.53a 0.489±0.029a 34±3.99a 0.479±0.028a
Uninfested 29.4±4.53b 0.429±0.029b 28.8±3.99a 0.426±0.028a
100 % FC 41.4±4.53a 0.464±0.029a 37.4±2.28a 0.448±0.028a
50 % FC 27.6±4.53b 0.453±0.029a 25.4±2.28b 0.457±0.028a
Head weight and head index were significantly affected by sorghum genotype across the two
experiments (Table 3.4).
Table 3.5: Effect of sorghum genotypes on head weight and head index
Experiment I Experiment II
Sorghum
genotype
Head weight (g) Head index Head weight (g) Head index
SC Sila 4.04±1.304a 0.056±0.0266
a 5.52±1.228
a 0.0659±0.0211
a
Mukadziusaende 8.12±1.304b 0.165±0.0266
b 8.05±1.228
b 0.1446±0.0211
b
Wild Sorghum 0.44±1.304c 0.005±0.0266
a 1.96±1.228
c 0.0207±0.0211
c
Chiredhi 3.22±1.304a 0.0544±0.0266
a 3.4±1.228
c 0.0486±0.0211
ac
Isifumbathe 1.85±1.304ac
0.0339±0.0266a 1.94±1.228
c 0.0268±0.0211
dc

83
Head weight was highest on the genotype Mukadziusaende, which had head indices of 0.16
and 0.45 in experiment I and II respectively. The least head weight and head index were
recorded for wild sorghum in both experiments (Table 3.5).
Table 3.6: The effect of infection on head weight and head index
Experiment I Experiment II
Head weight (g) Head index Head weight (g) Head index
Infested 2.31±0.825a 0.0385±0.0168
a 2.83±0.777
a 0.0432±0.0134a
Uninfested 4.76±0.825b 0.0874±0.0168
b 5.52±0.777
b 0.0795±0.0134
b
The results revealed that infestation significantly reduced head weight and head index in both
experiments. Non infestation led to increase in head weight and head index in both
experiments (Table 3.6).
Table 3.6: The effect of water availability on head weight and head index across the two
experiments
Experiment I Experiment II
Head weight (g) Head index Head weight (g) Head index
100 % FC 4.93±0.825a 0.0755±0.034
a 6.14±0.777
a 0.777±0.0133
a
50 % FC 2.14±0.825b 0.0504±0.034
a 2.21±0.777
b 0.045±0.0133
b

84
Increased water availability significantly increased head weight and head index in both
experiments except head index in experiment I (Table 3.6). Increased moisture availability
significantly increased (P<0.05) head weight for SC Sila and Mukadziusaende, whilst the rest
of the sorghum genotypes did not respond to moisture availability (Figure 3.3).
Sorghum genotypes
Sc Sila
Mukadziusaende
Wild
Sorghum
Chiredhi
Isifumbathe
So
rgh
um
yie
ld (
g)
0
2
4
6
8
10
12
14
16
100 % FC
50 % FC
Figure 3.3: The response of sorghum genotypes yield to moisture availability.
The yields of wild sorghum, Chiredhi and Isifumbathe were not affected by water
availability. However, yields of SC Sila and Mukadziusaende were lowered by reduced
moisture availability although they remained higher than the other genotypes (Figure 3.5).
The yield of Mukadziusaende at 50 % water availability was still higher compared to wild
sorghum at 100 % FC (Figure 3.5).
In experiment II, leaf index, stem weight and stem indices were significantly affected by
sorghum genotypes (Table 3.7). However, SC Sila had a significantly (P<0.01) higher dry
weight and the least was Mukadziusaende. This trend was repeated for Experiment 11 (Table

85
3.7). Infestation did not affect leaf weight, leaf index, stem weight and stem index and total
dry matter for both experiments (Table 3.8). However, irrigation at 100 % field capacity gave
a significantly higher leaf weight, leaf index, stem weight, stem index and total dry matter in
both experiments (Table 3.9).

86
Table 3.7: Effect of sorghum genotypes on leaf dry matter, leaf index, stem weight and index and total dry matter in both experiments
Experiment I Experiment II
Sorghum genotype Leaf Index Leaf dry matter
(g)
Stem weight
(g)
Stem index Total dry
matter (g)
Leaf weight (g) Leaf index Stem weight
(g)
Stem index Total dry matter
(g)
Sc Sila 0.197±0.028a 14.54±1.754a 21.2±3.2a 0.284±0.032a 78.4±9.21a 14.92±1.684a 0.2055±0.024a 19.0±2.91a 0.262±0.034a 75.8±7.83a
Mukadziusaende 0.1888±0.028a 9.20±1.754b 15.9±3.2a 0.312±0.032a 49.7±9.21b 8.85±1.684b 0.1767±0.024a 14.84±2.91a 0.291±0.034a 49.8±7.83b
Wild sorghum 0.2028±0.028a 16.81±1.754a 18.4±3.2a 0.248±0.032a 76.3±9.21a 15.7±1.684a 0.2314±0.024a 17.12±2.91a 0.238±0.034a 71.5±7.83a
Chiredhi 0.1972±0.028a 14.62±1.754a 22.9±3.2a 0.293±0.032a 77.9±9.21a 13.32±1.684a 0.1954±0.024a 21.27±2.91a 0.303±0.034a 71.4±7.83a
Isifumbathe 0.222±0.028a 15.94±1.754a 18.4±3.2a 0.248±0.032a 74.1±9.21a 12.91±1.684a 0.1925±0.024a 19.3±2.91a 0.292±0.034a 66.7±7.83a
Table 3.8: The effect of infection on leaf weight and index, stem weight and index and total dry matter in both experiments.
Experiment I Experiment II
Leaf weight (g) Leaf index Stem weight (g) Stem index Total dry matter
(g)
Leaf weight (g) Leaf index Stem weight (g) Stem Index Total dry matter
(g)
Infested 14.63 0.205±0.018a 19.4±2.03a 0.267±0.014a 75.9±5.82a 13.48±1.06a 0.2028±0.015a 17.99±1.84a 0.269±0.02a 68.3±4.95a
Uninfested 13.82 0.197±0.018a 19.3±2.03a 0.287±0.014a 66.7±5.82a 12.8±1.06a 0.1978±0.015a 18.62±1.84a 0.285±0.02a 65.7±4.95a

87
Table 3.9: The effects of water availability on leaf weight and index, stem weight and index and total dry matter.
Experiment I Experiment II
Water
availability
Leaf weight (g) Leaf index Stem weight (g) Stem Index Total dry
matter (g)
Leaf weight (g) Leaf Index Stem weight (g) Stem index Total dry
matter (g)
100 % FC 15.81±1.109a 0.1947±0.0176a 22.1±2.03a 0.266±0.302a 84.2±5.82a 15.08±1.065a 0.1943±0.015a 21.3±1.842a 0.269±0.02a 79.9±4.95a
50 % FC 12.64±1.109b 0.1922±0.0176a 16.6±2.03b 0.287±0.302b 58.4±5.82b 11.20±1.065b 0.2063±0.015a 15.32±1.842b 0.285±0.02a 54.1±4.95b

88
There was a significant effect of infection and drought on leaf index (P<0.05) (Figure 3.7)
Infestation status
Infested Uninfested
Leaf
inde
x
0.00
0.05
0.10
0.15
0.20
0.25
0.30
100 % irrigation
50 % Irrigation
Figure3.6: Interaction effects of water availability and Striga infestation on leaf index.
Under infestation, 100 % irrigation had a lower leaf index compared to 50 % and under
infestation there were no significant differences (Figure 3.7). There was a significant
interaction of variety and infection on stem dry matter (P <0.05). Stem weight of wild
sorghum was reduced by Striga infestation (P<0.05) whilst it was vice versa for Chiredhi
(Figure 3.7).

89
Sorghum genotypes
Sc Sila
Mukadziusaende
Wild sorghumChiredhi
Isifumbathe
Stem
weig
ht (g
)
0
5
10
15
20
25
30
35
Infested
Uninfested
Figure 3.7: Interaction effects of sorghum genotypes and Striga asiatica infestation on stem
weight
3.5 Discussion
The objective of the study was to determine the effects of Striga asiatica and water stress
occurring simultaneously on sorghum productivity. The sorghum genotype Mukadziusaende
had the highest chlorophyll concentration of 47.33 mmolcm-2
, and the least was recorded for
wild sorghum, with 29.78 mmolcm-2
. This trend for chlorophyll concentration was the same
for both experiments. These values are in the range commensurate with Gurney et al.,
(2002)’s findings, where a maximum of 47.44 and a minimum of 32.33 mmolcm-2
were
reported.
Chlorophyll concentration was lowered by moisture deficit when irrigated at 50 % FC
compared to 100 % FC (Table 3.2) but was not affected by infection (Table 3.4). This
contrasts with the findings of Gurney et al., (2002), where Striga asiatica infection alone

90
reduced chlorophyll concentration. Similar results were also found by Wahid and Rasul
(2005) and Fu and Huang (2001) who reported that drought impaired the photosynthetic
machinery of the plant which eventually reduces food production. Likewise, Anjum et al.
(2003) also reported changes in photosynthetic pigments and their components as a result of
drought. According to Niinemets et al., (2010), measurements of chlorophyll provides an
important tool to gaining insight into modifications of foliage physiological activity. The
sensitivity of photosynthesis to both biotic and abiotic stresses varies with plant genotype
tolerance. This study revealed that sorghum genotypes vary greatly with respect to
chlorophyll concentration when exposed to the same environmental limitations. Palta et al.,
(1994) and Zhang et al., (1998) reported that water deficits result in early senescence which
results in reduced chlorophyll concentrations. The results of this study suggested that drought
stress takes precedence over Striga asiatica stress when they co-occur in sorghum. This may
be attributed to the fact that water has to be available prior to Striga asiatica infection in
sorghum. The results may also suggest that the two are mutually exclusive on their effects on
chlorophyll concentration in sorghum.
The responses of sorghum genotypes to chlorophyll content under 50 and 100 % FC tended
to differ (Figure 3.2). The genotypes Mukadziusaende, wild sorghum and Chiredhi had
similar chlorophyll content at both irrigation regimes. However, reduced water availability
lowered the chlorophyll content of genotypes SC Sila and Isifumbathe. Similar results were
found by Gurney et al., (2002) who reported a maize variety, ‘Staha’, whose foliar
chlorophyll concentration was unaffected by the parasite. In the current study, it was
hypothesized that the genotypes Mukadziusaende, wild sorghum and Chiredhi showed
resilience to both stresses hence photosynthesis was maintained in these genotypes despite
the presence of both stresses, which may help maintain sorghum productivity. This may be
due to the limited sensitivity of the genotypes towards drought. According to Cameron et al.,

91
(2006), it is known that the responses of the genotypes to reduced water availability might be
high osmotic adjustments that help maintain leaf water potential. Bloom et al., (1985)
reported that even in limited supply of resources, plants have to maintain a balanced
investment such that all functions and organs are limited to the same degree. Across all the
two experiments, NDVI was higher at 100% compared to 50 %. NDVI is a measurement of
amalgamated plant growth that reflects the effects of various plant growth factors and is
highly correlated with plant available soil moisture (Verhulst and Govaerts, 2010). For the
genotypes SC Sila, Mukadziusaende, wild sorghum and Isifumbathe, NDVI was lowered by
drought treatments. Bjorkman and Powles (1984) reported that the effect of S. asiatica on
both photosynthetic performance and photo-inhibition of maize plants under light conditions
is similar to the effects observed when abiotic factors such as water shortage are imposed. For
the genotype Chiredhi, NDVI was higher at 100 % moisture compared to 50 %, whilst the
rest of the genotypes were not affected. Irrigation at 50 % of field capacity could have limited
nitrogen assimilation and consequently lowered chlorophyll concentration in the affected
genotypes.
Under infested conditions, moisture availability did not affect internode length. However, 100
% FC under uninfested conditions increased internode length (Figure 3.4). The fact that
drought reduced internode length in sorghum is in tandem with Deligoz and Gur (2015)’s
findings who reported that drought stress causes physiological and metabolic changes which
negatively affects growth and development of plants. Actually, Farooq et al., (2009) reported
that growth is accomplished by cell division, enlargement and differentiation. Nonami (1998)
posited that under water deficient conditions, cell elongation can be inhibited by interruption
of water flow from xylem vessels to surrounding cells. This study revealed that in relation to
internode length, the effect of reduced water availability is equal to the effect of Striga.

92
Under 50 % FC, non infested sorghum had limited growth and it only grew when water was
made available at 100 % FC.
Striga infestation increased dry matter allocated to the roots in Experiment I, but had no
effect on experiment II. This agrees with Poorter et al., (2011), who reported that plants
allocate more dry matter to the roots as the limiting factor is below the ground. Similar results
were also found by Farooq et al., (2009 and Liu et al., (2011). The results indicated that root
dry mass decreased under drought, which was also reported by Luttschwager et al., (2016),
who found decreased root mass under drought in Populus tremula.
Head weight and head index were lowered by S. asiatica infestation and drought (Tables 6
and 7). The results are in tandem with the findings of Baker et al., (1996) and Vasey et al.,
(2005) in which infestation reduced dry matter allocated to the head. Similar results were
found by Pandey et al., (2000) who found that the harvest index was lowered by increased
water stress. Groene (2008) concurs with the assertions and reported that drought has an
effect on pollen viability, pollen tube germination and increases in ovule abortion rates as a
result of reduction in assimilate supplies which are required for grain development.
According to Ober et al., (1991), water stress resulted in diminished grain set and kernel
growth in wheat and decreased rate of endosperm cell division. Striga asiatica causes
increases in abscissic acid (Taylor et al., 1996; Frost et al., 1997). Also, increases in ABA
concentration as a result of drought had been previously documented by Aldesuquy and
Ibrahim, (2001) and Gniazdowska et al., (2007). Cramer et al., (2011) asserted that ABA is
an important regulator of plant responses to both abiotic and biotic stresses. Both drought and
Striga infestation have been reported to lead to an increase in ABA production and
consequently cause stomatal closure reducing carbon dioxide entrance into the leaf, hence
reduced productivity.

93
Leaf and stem indices were not affected by sorghum genotypes (Table 3.8) and infestation
(Table 3.9) whereas they were both reduced by irrigation at 50 % of field capacity. These
results are in disagreement with the findings of Aflakpui et al., (1998), who found that Striga
infestation reduced leaf and stem indices. However, their study on maize was subjected to
Striga only whereas in this study, drought was also a factor that was added to S. asiatica
infection. Taken together, these results indicated that the response of sorghum to S. asiatica
and drought is complex and cannot be extrapolated from the results of each stress applied
singly. This confirms the assertion by Mittler (2005) that two or more stresses may require a
unique response on the hosts and that the responses may have synergistic or antagonistic
effects on each other. From the results, drought effects got preference when they co-existed
with Striga infestation. The simultaneous occurrence of different biotic and abiotic stresses
was shown to result in a high degree of complexity in plant responses as the responses to
these combined stresses are largely controlled by different signaling pathways that may
interact or inhibit one another (Artkinson and Urwin, 2012; Rasmussen et al., 2013).
Consequently, the fact that drought only had a significant influence on leaf and stem biomass
indicated that its influence was greater than that of S. asiatica or the response pathways to
drought suppresses the effects of S. asiatica. The effects of S. asiatica are inhibited when it
co-exists with drought in sorghum. Results from this study confirmed the findings of
Artkinson and Urwin (2012), who reported that plants respond in a specific manner when
they have to face more than one stress simultaneously, and the response cannot be predicted
based on the plant’s response to the individual stresses.
Leaf index was higher under infested conditions at 50 % irrigation compared to uninfested
conditions (Figure 3.4). This demonstrated that the two stresses resulted in more dry matter

94
being channeled to the leaves. These results confirmed the assertion by Suzuki et al., (2014)
that stress combinations might have beneficial effects on plants compared to each stress
applied separately.
Stem weight was not significantly affected by infections for all varieties except for wild
sorghum where infection lowered stem weight but increased stem weight for Chiredhi.
Reduced allocation of dry matter to the stems combined with increased allocation to the roots
was reported by Frost et al., (1995) and Graves et al., (1990). It is now known that the
parasite acts as a sink for carbon, inorganic solutes and water and also because of the reduced
carbon gain in infested hosts as reported by Smith et al., (1995) and Cechin and Press (1993).
Under infestation, reduced water availability increased leaf index and this is likely an issue of
overcompensation.
3.5 Conclusions
The study found that with respect to chlorophyll content, reduced water availability and
infestation were mutually exclusive. With respect to sorghum internode length, the resilience
of sorghum to Striga asiatica is reduced when sorghum is affected by drought which shows
synergistic effects of drought and S. asiatica infestation..
References
Aldesuquy HS, Ibrahim AH (2001) Water relations, abscissic acid and yield of wheat plants
in relation to the interactive effect of sea water and growth bioregulators. Journal of
Agronomy and Crop Science 187: 97 – 104
Anjum F, Yaseen M, Rasul E, Wahid A, Anjum S (2003) Drought stress in barley
(Hordeum vulgare) .1. Effect of chemical composition and chlorophyll content. Pakistan
Journal of Agric Sciences 40: 45 – 49.

95
Atkinson N, Urwin PE (2012) The interaction of plant abiotic and biotic stresses: from genes
to the field. Journal of Experimental Botany 63: 3523 - 3544
Baker ER, Press MC, Scholes JD, Quick WP (1996) Interactions between the parasite
angiosperm Orobanche aegyptiana and it tomato host: growth and biomass allocation. New
Phytologist 133: 632 - 642
Bjorkman O and Powles SP (1984) Inhibition of photosynthetic reactions under water stress:
interactions with light level. Planta 153: 376 – 387.
Bloom AJ, Chapin FS, Mooney HA (1985) Resource limitation in plants- an economic
analogue. Annual Review of Ecology and Systematics 16: 363 – 392.
Cameron DP, Coats AM, Seel WE (2006) Differential resistance among host and non host
spp underlies that variable success of the hemi-parasitic plant Rhinathus minor. Annals of
Botany 98: 1289 – 1299.
Cechin I and Press MC (1993) Nitrogen relations of sorghum –Striga hermonthica host
parasite association: growth and photosynthesis. Plant, Cell and Environment 16: 237 - 247
Chauhan BS and Johnson, DE (2010) Growth and reproduction of jungle rice (Echnochloa
colona) in response to water stress. Weed Science 58 (2): 132 –135
Cramer GR, Urano K, Delrot S, Pezzotti M, Shinozaki K (2011) Effects of abiotic stress on
plants: a systems biology perspective. BMC Plant biology 11: 163 – 179.
Deligoz A and Gur M (2015). Morphological, physiological and biochemical reponses to
drought stress of stone pine seedlings. Acta Physiologia Plantarum 37: 243 - 251

96
Demirevska K, Simova-Stoilova L, Fedina I, Georgieva K, Kunert K (2010). Response of
Oryza cystatin 1 transformed tobacco plant to drought, heat and light stress. Journal of
Agronomy and Crop Science 196: 90 – 99.
Ejeta G (2005). Integrating biotechnology, breeding and agronomy in the control of parasitic
weed Striga spp in sorghum. In Tuberosa R, Philips R.L; Gate M (eds). Proceedings of the
international congress, In the wake of a double helix: from green revolution to gene
revolution. 27 – 31 may 2003. Bologna, Italy
Ejeta G (2007) The Striga scourge in Africa: a growing pandemic. In integrating new
technology for Striga control: towards ending the witch hunt (Ejeta G and Gressel J Eds).
Pages 1 – 16. World scientific publishing company.
Farooq M, Wahid A, Kaboyashi N, Fujita D, Basraa SMA (2009) Plant drought stress:
effects, mechanisms and management. Agronomy for Sustainable Development 29: 185 –
212.
Frost DL, Gurney AL, Press MC, Scholes JD (1997). Striga hermonthica reduces
photosynthesis in sorghum: the importance of stomatal limitations and potential role of
ABA?. Plant Cell and Environment 20: 483 - 492
Fu J, Huang B (2001) Involvement of antioxidants and lipid peroxidation in the adaptation of
two cool season grasses in localized drought stress. Environmental Experimental Botany 45:
105 – 114.
Ghanounn O (2009) C4 photosynthesis and water stress. Annals of Botany 103: 635 – 644
Gniazdowska A, Oracz K, Bogatek R (2007) Phytotoxic effects of sunflower (Helianthus
annuus L.) leaf extracts on germinationg mustard seed (Sinapsis alba. L.). Allelopathy
Journal 19: 215 – 226.

97
Graves JD, Press MC, Stewart GR (1990) A carbon balance model of the Sorghum-Striga
hermonthica host parasite association. Plant, Cell and Environment 12: 101 – 107.
Gurney AL, Taylor A, Mbwaga A, Scholes JD, Press MC (2002). Do maize cultivars
demonstrate tolerance to the parasitic weed Striga asiatica. Weed Research 42: 299 – 306.
Groene, G.A (2008). Evaluating sorghum and maize Germplasm for post anthesis drought
tolerance. A thesis submitted to Kansas State University, USA, For the Partial Fulfillment of
the Degree of Master of Science.
Isebrands JG, Dickson RE, Rebbeck J, Karnosky DF (2000). Interacting effects of multiple
stresses on growth and physiological processes in northern forest trees. In Mickler RA,
Birdsey RA, Hom J (eds). Responses of northern US forests to environmental changes.
Springer verlag. New York pp 149 – 180.
Iyer NJ, Tang Y, Mahalingam R (2013) Physiological, biochemical and molecular responses
to a combination of drought and ozone in Medicago truncatula. Plant, Cell and Environment
36: 706 - 720
Jury WA, Vaux HJ (2007) The emerging global water crisis: managing conflict between
water users. Advances in Agronomy 95: 1 – 76.
Kabiri S, Rodenburg J, Kayeke J, Van Aast A, Makokha DW, Msangi SH, Bastiaans L
(2014). Can the parasitic weeds Striga asiatica and Rhamphicarpa fistulosa co-occur in
rainfed rice?. Weed Research DOI:10:1111/wre.12124
Leport L,Turner NC, French RJ, Barr MD, Duda R, Davis SL (2006) Physiological responses
of chickpea genotypes to terminal drought in a Meditterannean type environment. European
Journal of Agronomy 11: 279 – 292.

98
Low M, Herbinger K, Nunn AJ, Harbele KH, Leuchner M, Heerdt C, Werner H, Wipfler P,
Pretzsch H, Taisz M (2006). Extraordinary drought of 2003 overules ozone impact on adult
beech trees (Fugus sylvatica). Trees 20: 539 – 548.
Luttschwager D, Ewald D, Alia LA (2016). Consequences of moderate drought stress on net
photosynthesis, WUE and biomass production of three poplar clones. Acta Physio Plantarum
38: DOI10.1007/s11738-015-2057-7.
Mittler R (2005). Abiotic stress, the field environment and stress combination. Trends in
Plant Science 11: 15 – 19.
Mittler R, Blumwald E (2010) Genetic engineering for modern agriculture: challenges and
perspectives. Annual Review of Plant Biology 61: 443 - 462
Nakashima K, Ito Y, Yamaguchi-Shinozak K (2009). Transcriptional regulatory networks in
response to abiotic stresses in Arabidopsis and grasses. Plant physiology149: 88 – 95
Niinemets U (2010). Response of forest trees to single and multiple environmental stresses
from seedlings to mature plants: Past stress history, stress interactions, tolerance and
acclimation. Forest Ecology and Management 260: 1623 – 1639.
Nonami H (1998). Plant water relations and control of cell elongation at low water potentials.
Journal of Plant Research 111: 373 – 382
Ober ES, Settler TL, Madison JT, Thompson JF, Shapiro PS. (1991). Influence of water
deficit on maize endosperm development: enzyme activities and RND transcripts of starch
and zein synthesis, ABA and cell division. Plant Physiology 97: 154 - 164

99
Ollinger SV, Aber JB, Reich PB (1997) Simulating ozone effects on forest productivity:
interaction among leaf-canopy and standard level processes. Ecological Applications 7: 1237
- 1251
Padgham J (2009). Agriculture and development under climate change: opportunities and
challenges for adaptation. The World Bank, August 2009. Issue 1.
Palta JA, Kobata T, Turner NC, Fillery IR (1994). Remobilisation of nitrogen in wheat as
influenced by post anthesis water deficits. Crop Science 34: 118 - 124
Pandey RK, Maranville JW, Chetima MM (2000). Deficit irrigation and nitrogen effects on
maize in a Sahelian environment 11. Shoot growth, nutrient uptake and water extraction.
Agric Water Management 46: 15 - 27
Pervez HZ, Srinivasan G, Cardova HS, Sanchez C (2004). Gains from improvement for mid
season tolerance in tropical Zea mays. Field Crops Research 89: 135 – 152.
Poorter H, Nicklas KJ, Reich PB, Oleksyn J, Poot P, Mommer L (2011). Biomass allocation
to leaves, stems and roots: meta-analysis of interspecific variation and environmental control.
New Phytologist 193: 30 – 50.
Prasad PVV, Pisipati SR, Momcilovic I, Ristic Z (2011). Independent and combined effects
of high temperature and drought stress during grain filling on plant yield and chloroplast EF-
Tu expression in spring wheat. Journal of Agronomy and crop Science 197: 430 - 441
Prasch CM, Sonnewald U (2013). Simultaneous application of heat, drought and virus to
Arabidopsis plant reveals significant shifts in signaling networks. Plant physiology 162: 1849
- 1866

100
Rasmussen S, Barah P, Suarez-Rodriguez MC, Bresserndoff S, Friis P, Constantino, P, Bones
AM, Nielson HB, Mundy J (2013). Transcroptome responses to combinations of stresses in
Arabidopsis. Plant Physiology 161: 1783 – 1794.
Rejeb, I.B, Pastor, V and Mauch-Mani, B (2014). Plant responses to simultaneous biotic and
abiotic stress: molecular mechanisms. Plants 3: 458 – 475
Rhizhsky L, Liang H, Mittler R (2002). The combined effect of drought and heat shock on
gene expression in tobacco. Plant physiology 130: 1143 – 1151
Rhizhsky L (2004). When defence pathways collide: the response of Arabidopsis to a
combination of drought and heat stress. Plant Physiology 134: 1683 - 1696
Scholes JD, Press MC (2008). Striga infestation of cereals crops: an unresolved problem in
resource limited agriculture. Current Opinions in Plant Biology 11: 180 – 186.
Smith LH, Keys A, Evans MCW (1995) Striga hermonthica decreases photosynthesis in Zea
mays through effects on leaf structure. Journal of Experimental Botany 46: 759 – 765.
Somerville C, Briscoe J (2001). Genetic engineering and water. Science 292: 2217.
Suzuki N, Riveiro RM, Shulaev V, Blumwald E, Mittler R (2014) Abiotic stress and biotic
stress combinations. New Phytologist 203: 32 – 43.
Suzuki N (2005) Enhanced tolerance to environmental stress in transgenic plants expressing
transcriptional co-activator Multiprotein bridging factor1c. Plant physiology 134: 1313 –
1322.
Tari I, Laskay G, Takacs Z, Poor P (2013) The response of sorghum to abiotic stresses: A
review. Journal of Agronomy and Soil science 199: 264 274

101
Taylor, A, Martin J, Seel WE (1996) Physiology of the parasitic association between maize
and the witch weed Striga hermonthica: is ABA involved. Journal of Experimental Botany
47: 1057 – 1065.
Tesitel J, Tesitolova T, Fisher JP, Leps J, Cameron DD (2015) Integrating ecology and
physiology of root hemi-parasitic interaction: interactive effects of biotic resources shape the
interplay between parasitism and autotrophy. New Phytologist 205: 350 – 360.
Vasey RA, Scholes JD, Press MC (2005) Wheat (Triticum aestivum) is susceptiple to the
parasitic angiosperm Striga hermonthica, a major cereal pathogen in Africa. Genetics and
Resistance 95: 1294 – 1300.
Velhurst N, Govaerts B (2010) The normalized difference vegetation index (NDVI)
greenseeker handheld sensor: toward the integrated evaluation of crop management. Part A:
concepts and case studies. Mexico, D.F.; CIMMYT.
Zhang J, Sui X, Li B, Su B, Li J, Zhou D (1998). An improved water use efficiency for
winter grown wheat under deficit irrigation. Field Crops Research 59: 91 - 98
Vile D, Pervent M, Belluau M, Vasseur F, Bresson J, Muller B, Granier C, Simonneau T
(2012). Arabidopsis growth under prolonged high temperature and water deficit: independent
or interactive effects. Plant, Cell and Environment 35: 702 – 718
Wahid A, Rasul E (2005) Photosynthesis in leaf, stem, flower and fruit. In Pessarakli, M.
(Ed), handbook of photosynthesis, 2nd
Edition. CRC Press, Florida Pp 479 – 497.
Webster TM and Grey TL (2008) Growth and reproduction of benghal day flower
(Commelina benghalensis) in response to drought stress. Weed science 56: 561 – 566.

102
Zhao W, Liu B, Zhang Z (2010). Water requirements of maize in the middle Heihe river
basin, China. Agric water Management 97: 215 - 223

103
CHAPTER FOUR:
The response of tolerance traits of Sorghum bicolor (L) Moench
and Sorghum arundinaceum (Desv) Stapf to Striga asiatica (L)
Kuntze infestation under mulch
4.1 Abstract
The production of sorghum is hampered by the parasitic weed Striga asiatica. Studies are
lacking on the effect of cultural techniques of managing the weed such as mulching.
Mulching is promoted as a component of the conservation agriculture systems in sub Saharan
Africa. Nine Sorghum bicolor cultivars and one Sorghum arundinaceum line were evaluated
under Striga infestation and under Striga free conditions with half being mulched and the
other half unmulched for two years at Bindura University of Science Education (BUSE)
nursery in Zimbabwe. The objective was to assess the effects of mulching and infestation on
the tolerance of Sorghum bicolor and Sorghum arundinaceum to Striga asiatica. The
experiment was a 2*2*10 factorial, replicated three times in both seasons and arranged as a
completely randomized design. Results indicated that mulching increased chlorophyll content
in the 2014/15 season compared to 2013/14. Infestation reduced chlorophyll content in both
seasons. For the 2014 season, mulching increased chlorophyll content in all varieties except
Ruzangwaya, Mukadziusaende and SC Sila. Stomatal conductance and tiller numbers were
higher (P<0.01) in uninfested sorghum compared to infested in both seasons. The varieties
Mukadziusaende, Chiredhi and Hlubi were able to maintain height despite infestation by
Striga. When the same varieties were infested under mulch and infested without mulch, the

104
results showed that mulching overcomes the effect of infestation in some varieties. Varieties
Chiredhi, Mukadziusaende and Mashava were able to maintain yield despite infestation by
Striga in the 2013 season. Wild sorghum was highly susceptible to Striga. Mulching negates
the effect of Striga parasitism in the drier season compared to wetter season and results in
yield maintenance in some varieties.
4.2 Introduction
Sorghum (Sorghum bicolor L) is a multipurpose crop belonging to the Poaceae family, which
are C4 carbon cycle plants with high photosynthetic efficiency and productivity (Tari et al.,
2013). Sorghum is a preferred crop in sub tropical Africa as it can maintain yields in
environments normally regarded as too hostile for other crops such as maize. Many people in
Southern Africa live in chronic food deficit regions including the semi-arid zones where
rainfall shortage causes recurring food shortages and consequent malnutrition. In these
regions, sorghum is a critical strategic grain crop for a vast number of farmers. It is an
essential component of sustainable agricultural systems in the region’s extensive semi-arid
areas. Improvement of sorghum productivity is an immediate priority in these areas.
Striga species are obligate hemi-parasitic plants that attach to the root of the host to obtain
water, nutrients and carbohydrates (Parker and Riches, 1993). The life cycle of Striga
asiatica is complex, and co-evolved with many hosts to comprise a series of discrete steps
that are tightly coupled to the host’s biochemistry and life cycle (Bouwmeester et al., 2003).
The parasite grows underground for 4 – 8 weeks prior to emergence (Jamil et al., 2013). The
severe infestations appear to render African farmers helpless even though they are otherwise
very resilient and adaptive (Ejeta, 2007). The extent and intensity of Striga species
infestations has rapidly increased and become a threat to food production in practically the
entire semi-arid region farming systems of southern Africa.

105
Effective control of S asiatica has proved challenging, mostly as a result of the intricate life
cycle of the parasite (Gurney et al., 2003). Complete resistance has not been identified in
sorghum although varieties vary in their sensitivity to the parasite (Gurney et al., 1995).
Therefore, the traits that reduce fitness impacts of damage such as maintenance of
chlorophyll, increased photosynthesis, compensatory growth and tiller production despite
infection are very critical in varietal selection under Striga infestations. Also, any cultural
practice that negates Striga parasitism is critical to sorghum productivity.
Cultural practices like mulching may enable expression of tolerance since the parasite affects
the water economy of the plant. Mulch tends to develop and strengthen the top soil structure
through soil protection, microfauna activities and the incorporation of organic matter which
usually provides a high infiltration rate (Rao et al., 1998, Scopel et al., 1998). Mulch reduces
surface runoff due to increased roughness (Gilley et al., 1991; Gilley and Kottwitz, 1992).
Adekalu et al., (2007) reported increased water infiltration with increasing levels of mulch.
The same mulch has low thermal conductivity such that soil temperature is reduced
sometimes with consequent decrease in root development (Riddle et al., 1996). Generally
mulching is known to reduce weeds through physical effects and allelopathy. However, this
information relates to non-parasitic weeds and there is limited information on the effects of
mulching on Striga incidence and parasitism.
Sorghum arundinaceum is increasing in Zimbabwe’s cropping systems as a weed and occurs
in all crops. Studies by Gurney et al., (2002) and Rich et al., (2004) found a Sorghum
arundinaceum strain that was tolerant to Striga. In Zimbabwe, Mwenje (2006) reported high
compatibility between cultivated sorghum and its wild relatives. Therefore, the objectives of
this study were:

106
i) To determine the effect of mulching and Striga infestation on sorghum chlorophyll
concentration, stomatal conductance, sorghum height and Striga counts.
ii) To determine the effect of mulching and Striga infestation on sorghum grain yield and
dry matter traits.
The corresponding alternative hypothesis were
i) Mulching and Striga infestation affect the expression of tolerance traits of sorghum
genotypes with respect to chlorophyll concentrations, stomatal conductance,
sorghum height and Striga counts.
ii) Mulching and Striga infestation affect the expression of tolerance traits in sorghum
with respect to sorghum grain yield and dry matter partitioning.
4.3 Materials and methods
4.3.1 Experimental site
The pot experiments were carried out at Bindura University of Science Education (BUSE)
Astra Campus nursery, Bindura (GPS coordinates: 17o 181 5811 S and 31
o 19
1 2311 East) during
the 2013/14 and 2014/15 summer seasons. The area is located in agro-ecological region 11b
according to the Zimbabwean classification system (Vincent and Thomas, 1961) and it
receives a total of 800 mm of rainfall per season from November to April. The pot
experiment was done in medium grained sands with a pH of 4.2.
4.3.2 Experimental design and pot layout
For both seasons, the experiment was 2*2*10 factorial experiment. The first factor was
mulch at two levels: mulched at a rate equivalent to 3 t ha-1
and 0 t ha-1
. The second factor
was infestation at two levels: infested and uninfested. The third factor was sorghum genotype

107
at 10 levels (Table 4.1). The experiment was laid down as a completely randomized design
replicated three times.
Table 4.1: Sorghum genotypes tested for tolerance to Striga in the 2013/14 and 2014/15
seasons
Sorghum variety/code* Region Collected/grown Local name
1551 Matebeleland Isifumbathe
1773 Chiredzi Chiredhi
1836 Mrewa Mashava
1555 Matebeleland Tswetha
1556 Matebeleland Hlubi
1697 Masvingo Mukadziusaende
1669 Matebeleland Zambia
4487 Masvingo Ruzangwaya
SC Sila Zimbabwe
Wild sorghum Mashonaland West
*Codes refer to accession numbers at the Department of Research and Specialist Services,
gene bank in Harare, Zimbabwe
4.3.4 Source of Seeds
Sorghum and S. asiatica seeds were obtained from a source as stated in Section 3.3.2.
4.3.5 Experimental details
The soil was obtained from a farmer’s field where no Striga was reported before. The pots
were filled with soil and the top 10 cm of the soil was thoroughly mixed with 5.7 g of maize
fert (7N: 14 P2O5: 7 K2O) compound fertiliser and 1 g of conditioned S. asiatica seed. The
soil/S. asiatica mixture was placed back into the bucket after mixing thoroughly. Ten

108
sorghum seeds were planted in the bucket at a depth of 0.5 cm. The pot filling and planting
process was started on the uninfested pots to avoid contamination. The experiment was rain-
fed and the sorghum seedlings were thinned to one plant per pot at two weeks after crop
emergence (WACE). Sorghum plants were top-dressed by applying 3 grams of ammonium
nitrate (34.5 % N) per pot representing 90 kg ha-2
at 4 WACE. Non Striga weeds which
emerged were manually pulled and this was done continuously throughout the experimental
period.
4.3.6 Data collection
Height of the plants was measured and tiller number counted at 8 and 12 weeks after crop
emergence. Chlorophyll content was measured by a chlorophyll meter (SPAD-502 KONICA
MINOLTA INCL) at 6, 8 and 12 WACE. Sorghum yield was estimated by harvesting the
head when the sorghum plant had reached maturity and was weighed using a sensitive scale.
Stomatal conductance was measured using a porometer (DECAGON INCL).
4.3.7 Data analysis
Analysis of variance was performed to examine the effects of treatments and their
interactions on chlorophyll content, stomatal conductance, plant height and yield. The
Bartlett’s test for homogeneity of variance procedure showed that variances were not
homogeneous and hence, the data were analysed separately. The analysis was performed
using Genstat statistical software, Version 14. S. asiatica counts were subjected to square root
transformation prior to analysis.
4.4 Results
4.4.1 Chlorophyll content

109
Mulching increased (P<0.001) chlorophyll concentration at 6, 8 and 12 WACE for the
2014/15 season while it had no significant effect (P>0.05) during the 2013/14 season (Table
4.2).
Table 4.2: Effect of mulching on chlorophyll concentration for the 2013/14 and 2014/15
summer seasons
Condition Chlorophyll content for the 2013/2014
summer season
Chlorophyll content (mmolcm-2
) for 2014/2015 summer
season
6 WACE 8 WACE 12 WACE 6 WACE 8 WACE 12 WACE
Mulched 19.83a 31.62
a 41.06
a 20.69±0.883
a 46.25±1.014
a 37.69±1.376
a
Unmulched 20.3a 31.55
a 39.87
a 17.11±0.883
b 39.23±1.014
b 31.09±1.376
b
Infestation also lowered chlorophyll concentration at 6 WACE in the 2014/15 season but was
not significant at 8 and 12 WACE during the same season (Figure 4.1).
Figure 4.1: Effect of infestation on chlorophyll concentration at 6 WACE during the 2014/
15 season.

110
Sorghum genotypes
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
TshwethaSc Sila
Wild Sorghum
Chl
orop
hyll
conc
entr
atio
n (m
mol
cm-2
)
5
10
15
20
25
30
35
Infested
Uninfested
8 WACE in 2014 season
Sorghum genotypes
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
TshwethaSc Sila
Wild Sorghum
Chl
orop
hyll
conc
entra
tion
(mm
olcm
-2)
20
25
30
35
40
45
50
55
60 Mulched
Unmulched
Figure 4.2: Interaction between variety and Striga infestation a) 6 WACE andvariety and
mulch b) 8 WACE on chlorophyll concentration
There was a significant interaction of variety and mulching on chlorophyll content in the
2013/14 season (P<0.01) (Figure 2) and variety and Striga infestation (P<0.001).

111
At 6 WACE in the 2013, infestation significantly (P<0.05) lowered chlorophyll content of SC
Sila whilst all the other varieties were not affected. At 6 WACE in the 2014/15 season,
Isifumbathe, Chiredhi, Zambia and Wild sorghum had their chlorophyll concentration
significantly lowered by infection. Mulching significantly increased (P<0.01) chlorophyll
content in all varieties except Ruzangwaya, Mukadziusaende, Mashava and SC Sila. At 8
WACE, infestation lowered chlorophyll content (P<0.05) of wild sorghum whilst the content
for Tshwetha was not affected.
Figure 4.3: Effect of infestation status on stomatal conductance for the sorghum genotypes
during the 2013/14 season.
There was a significant effect of infection on stomatal conductance (P<0.001) (Figure 4.3).
The results revealed lowered stomatal conductance due to infestation compared to uninfested
conditions.
4.4.2 Tillering

112
The results showed that Mukadziusaende, Mashava and Isifumbathe had the highest tillers in
the 2013/14 summer season. The same varieties had the highest tiller numbers in the 2014/15
season. The 2013/14 season had less tillers for Chiredhi than 2013/14 season while 2014/15
had more tillers for Mukadziusaende and the same was true for Tshwetha (Figure 4.4).
Sorghum genotype
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
TshwethaSC Sila
Wild Sorghum
Till
er n
umbe
rs
0
1
2
3
4
5
2014 -15 season
2013 - 14 season
Figure 4.4: Effect of season on tiller numbers over two seasons.
Infestation status
Infested Uninfested
Till
er
nu
mb
er
0
1
2
3
4
Figure 4.5: Effect of infestation status on tiller numbers

113
For the variety Mukadziusaende and Tswetha, tiller number was significantly higher in the
2013/14 season compared to the 2014/15. Also infestation significantly (P<0.001) lowered
tillering across all the sorghum genotypes (Figure 4.5).
Sorghum genotypes differed significantly (P <0.05) at 10 WACE with regard to S. asiatica
incidence. The sorghum genotype Hlubi supported the highest number of S. asiatica plants
while Isifumbathe and Mukadziusaende supported the lowest at 10 WACE during both
seasons (Table 4.6).
Table 4.6: Effect of sorghum variety on Striga counts
Sorghum variety Striga counts at 10 WACE in 2013-
14 season
Striga counts at 10 WACE in 2014-
15 season
Hlubi 3.4 (12.88)±0.632d 5.08 (29.5)±0.71
e
Isifumbathe 1.28 (3.5)±0.632a 2.59 (8.2)±0.71
ab
Ruzangwaya 1.85 (4.67)±0.632abc
4.11 (21.7)±0.71cde
Chiredhi 1.73 (4.33)±0.632abc
2.34 (6.8)±0.71a
Mukadziusaenda 1.49 (3.67)±0.632ab
2.29 (5.5)±0.71a
Mashava 2.81 (8.67)±0.632cd
3.54 (13.7)±0.71abcd
Zambia 2.02 (4.17)±0.632abc
2.77 (8.5)±0.71abc
Tswetha 2.19 (5.87)±0.632abc
4.68 (23.2)±0.71de
SC Sila 2.66 (7.5)±0.632bcd
3.82 (14.8)±0.71bcde
Wild Sorghum 1.72 (5)±0.632abc
3.65 (13.5)±0.71abcd
The numbers in brackets represent the actual counts and those not in brackets are figures after square root transformation.

114
4.4.4 Plant height
The effects of Striga infection on sorghum height are shown in Figure 4.6. At 6 WACE,
Striga infection significantly reduced the height of some sorghum varieties. It was clear that
the impact of Striga infection on sorghum height was different among sorghum varieties at 12
WACE. It was interesting to note that Striga infection did not significantly reduce the
heights of Isifumbathe, Mukadziusaende and Chiredhi. In contrast, Striga infection reduced
the height of other sorghum cultivars (Figure 4.6).
8 wace in 2014 - 15 season
Sorghum genotype
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
TshwethaSC Sila
Wild Sorghum
Sor
ghum
hei
ght (
cm)
20
30
40
50
60
70
80
90
Infested
Uninfested
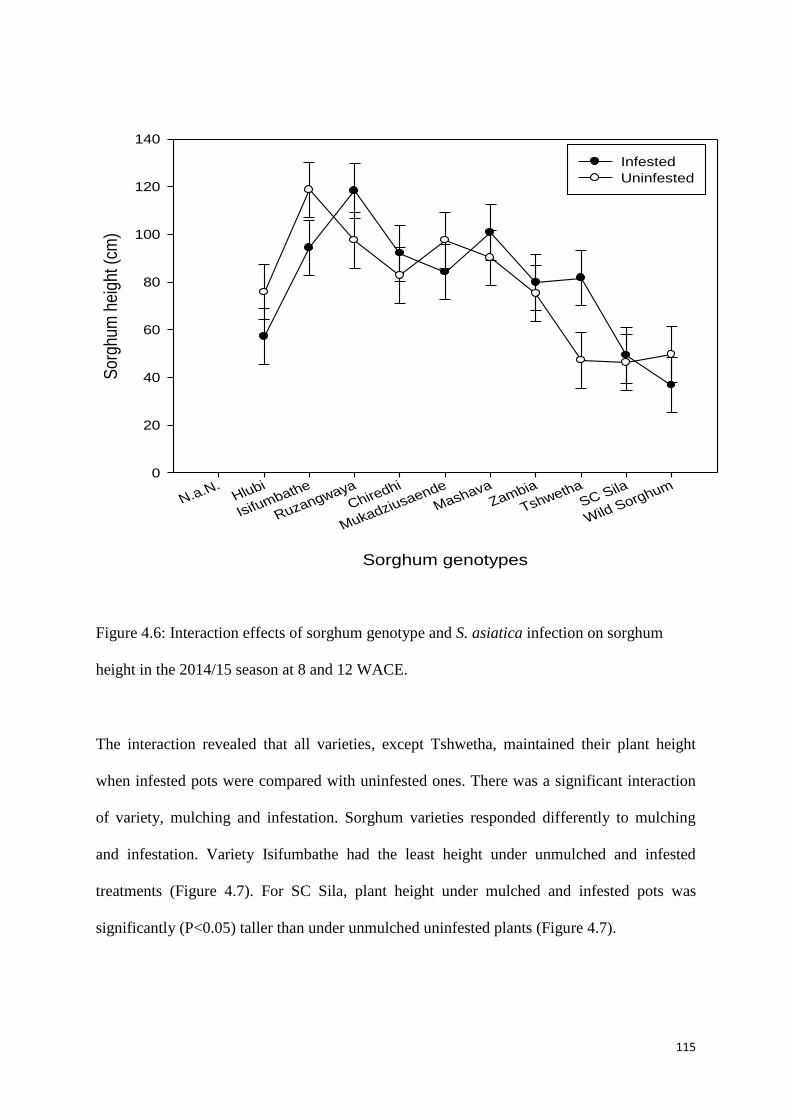
115
Sorghum genotypes
N.a.N.Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
TshwethaSC Sila
Wild Sorghum
So
rghu
m h
eig
ht (
cm)
0
20
40
60
80
100
120
140
Infested
Uninfested
Figure 4.6: Interaction effects of sorghum genotype and S. asiatica infection on sorghum
height in the 2014/15 season at 8 and 12 WACE.
The interaction revealed that all varieties, except Tshwetha, maintained their plant height
when infested pots were compared with uninfested ones. There was a significant interaction
of variety, mulching and infestation. Sorghum varieties responded differently to mulching
and infestation. Variety Isifumbathe had the least height under unmulched and infested
treatments (Figure 4.7). For SC Sila, plant height under mulched and infested pots was
significantly (P<0.05) taller than under unmulched uninfested plants (Figure 4.7).

116
Sorghum genotypes
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
TshwethaSC Sila
Wild Sorghum
Pla
nt heig
ht (c
m)
0
20
40
60
80
100
120
140
160
180
Mulched infested
Mulched uninfested
Unmulched infested
Unmulched uninfested
Figure 4.7: Interaction effects of sorghum genotype, mulching and infestation on plant height
in the 2014/15 season.
For the varieties Ruzangwaya, Tshwetha and SC Sila, mulched and infested pots gave taller
plants compared to unmulched and infested pots (Figure 4.7). For the variety Isifumbathe,
mulched and infested pots were taller better compared to infested but unmulched pots in the
2014/15 season (Table 4.7).
In the 2013/14 season infestation significantly (P<0.001) reduced affected plant height.

117
Table 4.7: Effects of infestation on plant height at 8 and 12 WACE in the year 2013/14
and 2014/15 seasons.
Infestation 2013/14 8 WACE 12 WACE 2014/15 8 WACE 12WACE
Infested 53.24±1.995a 97.3±6.61
a 48.94±1.955
a 79.5
a
Uninfested 60.68±1.995b 125.4±6.61
b 54.5±1.955
b 78.1
a
In the 2013/14 season there were significant interactions between variety and infestation at 6,
8 and 12 WACE.
Sorghum genotypes
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
Mashava
Zambia
Tshwetha
SC Sila
Sc Sila
Sor
ghum
hei
ght (
cm)
0
10
20
30
40
50
60
Infested
Uninfested
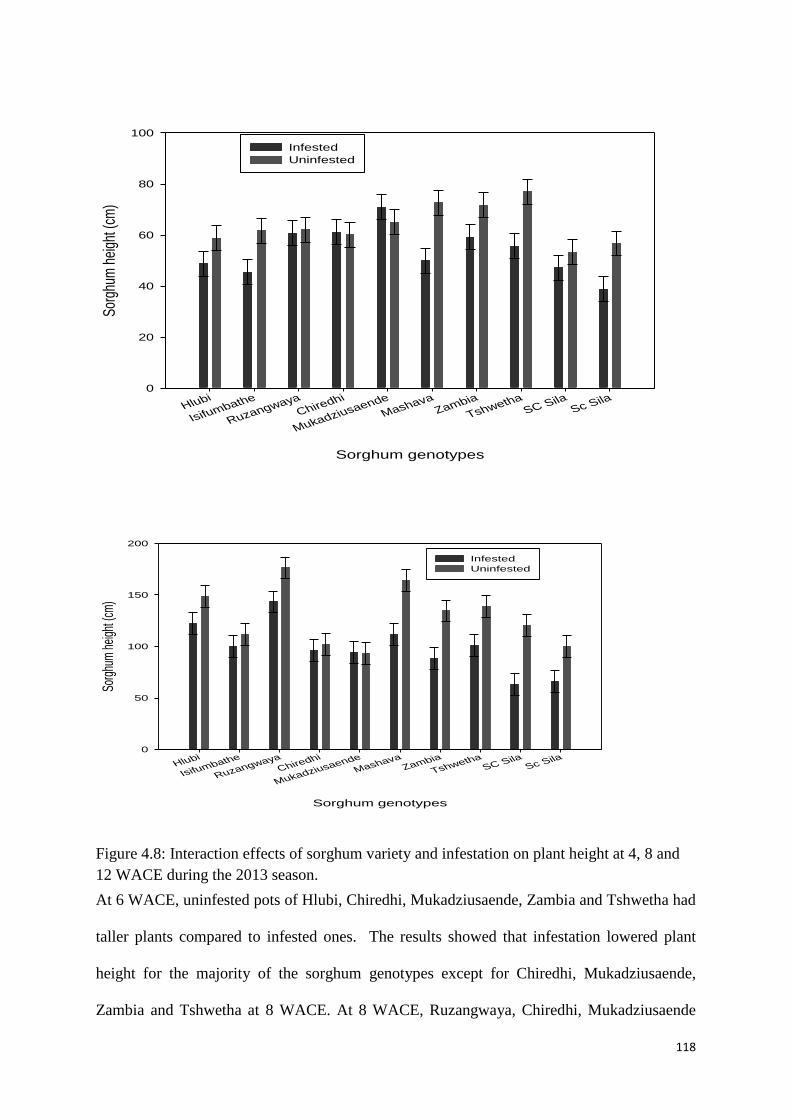
118
Sorghum genotypes
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
TshwethaSC Sila
Sc Sila
Sor
ghum
hei
ght (
cm)
0
20
40
60
80
100
Infested
Uninfested
Sorghum genotypes
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
TshwethaSC Sila
Sc Sila
Sorg
hum
hei
ght (
cm)
0
50
100
150
200
Infested
Uninfested
Figure 4.8: Interaction effects of sorghum variety and infestation on plant height at 4, 8 and
12 WACE during the 2013 season.
At 6 WACE, uninfested pots of Hlubi, Chiredhi, Mukadziusaende, Zambia and Tshwetha had
taller plants compared to infested ones. The results showed that infestation lowered plant
height for the majority of the sorghum genotypes except for Chiredhi, Mukadziusaende,
Zambia and Tshwetha at 8 WACE. At 8 WACE, Ruzangwaya, Chiredhi, Mukadziusaende

119
and SC Sila were not significantly affected by infestation (Figure 4.8). Isifumbathe,
Mukadziusaende and SC Sila resisted the dwarfing effects of Striga at 12 WACE.
4.4.5 Grain yield
The interaction of sorghum variety and Striga infection was significant (P<0.01) on sorghum
grain yield (P < 0.05). Striga infection significantly reduced the grain yield for most of the
sorghum varieties with the exception of Chiredhi, Mukadziusaenda and Mashava (Fig 4.9). In
fact, Mukadziusaende had significantly higher grain yield in the Striga infected plants
compared to uninfected plants (P < 0.05) for the 2013/14 season (Figure 4.9).
Sorghum varieties
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
Tswetha
SC Sila
Wild Sorghum
Sorg
hum
yie
ld (g
plan
t-1)
0
20
40
60
80
100
Infested
Uninfested
Figure 4.9: Interaction effects of sorghum genotypes and infestation on sorghum yield for the
2013/14 season
Uninfested pots had a higher head weight (P<0.05) compared to infested pots for Hlubi,
Isifumbathe, Ruzangwaya, Zambia, Tswetha and wild sorghum (Figure 4.10). Chiredhi and
Mashava maintained their yield despite infection. For the 2014/15 season head weight was

120
significantly affected by sorghum variety (P<0.001), infestation (P<0.001) and mulch (P=
0.001) (Figure 4.10).
sorghum yield in 2014 - 15 season
Infestation
Infested Uninfested
So
rgh
um
yie
ld (
g/p
lan
t)
0
10
20
30
40
2014 - 15 season
Mulching status
Mulched Unmulched
Yie
ld (
g/p
lan
t)
0
10
20
30
40
50
Yield for the 2014 - 15 season
Sorghum varieties
Hlubi
Isifumbathe
Ruzangwaya
Chiredhi
Mukadziusaende
MashavaZambia
TshwethaSC Sila
Wild Sorghum
Sorg
hum
yie
ld (g
/pla
nt)
0
10
20
30
40
50
60
Figure 4.10: Effect of sorghum genotypes, infestation and mulching on sorghum yield in the
2014/15 season
It was noted that whilst there were no significant interactions on yield, infestation reduced
yield while mulching increased yield (Figure 4.10). The sorghum genotypes Isifumbathe and
SC Sila had the highest yield. Mulching significantly (P<0.001) increased yield in the
2014/15 season.

121
4.5 Discussion
In this study, chlorophyll concentration was reduced by infection with S. asiatica.
Chlorophyll concentration is indicative of photosynthetic functioning and potential maximum
carbon dioxide assimilation rates. The fact that chlorophyll concentration was reduced by
infection is in agreement with the findings of Gurney et al., (2011) who found lower
chlorophyll content in Striga infested maize than uninfested susceptible maize genotypes.
Mulching increased chlorophyll concentrations at 6, 8 and 12 WACE for the 2014/15 season
which was not the case for 2013/14 season. The 2013/14 season was a wet season whilst the
2014/15 season had less rainfall with a lot of mid-season droughts. According to Chakraborty
et al., (2008), mulching is a suitable agronomic practice for conserving soil and water and
controlling soil temperature regimes. Mupangwa et al., (2012); Rao et al. (1998); Scopel et
al., (1998); and Adekalu et al., (2007) reported that the presence of mulch residue at the soil
atmosphere interface has a direct influence on infiltration of rainwater into the soil and
evaporation from the soil leading to improved water supply for crops. The advantage of
mulching with respect to chlorophyll content was apparent in the 2014/15 season which had
erratic rainfall.
Chlorophyll concentration is connected to nitrogen availability and is a key parameter in
measurement of plant canopies (Gitelson et al., 2003). Any perturbations in nitrogen
assimilation as a result of water limitations could in turn lower chlorophyll content as
reflected by the results of this study. The rest of the varieties whose chlorophyll content was
not affected by mulching may have high osmotic adjustment that helps maintain higher leaf
water potential. Blum (2005) reported that osmotic adjustment helps to sustain growth while
the plant is meeting transpirational demand by reducing its leaf water potential.

122
Stomatal conductance was lower in infested compared to uninfested sorghum genotypes. The
reduced stomatal conductance was probably caused by changes in the concentrations of
growth regulators in the host which are stimulated by Striga infestations (Frost et al., 1997).
Taylor and Frost (1997) revealed that increased levels of abscissic acid (ABA) in infested
plants resulted in stomatal closure. The wide variations of stomatal conductance following
infection across different varieties were probably a reflection of variable responses to ABA
accumulation in the plant.
The genotypes that maintain higher stomatal conductance are likely going to maintain
photosynthesis despite Striga infection. The varieties Chiredhi, Zambia and Wild sorghum
for the 2013/14 season and Hlubi, SC Sila and Tshwetha for the 2014/15 season maintained
opening of stoma under infestation and this is a useful trait for maintenance of
photosynthesis. It therefore enhances tolerance of the genotype to Striga infestations.
The genotypes that had the highest tiller numbers were Mukadziusaende, Mashava,
Isifumbathe and Ruzangwaya for both seasons. Tillers were more in infested sorghum
cultivars compared to uninfested ones. Tillering is one of the most important agronomic traits
in poaceous crops and plays a major role in determining plant architecture and grain yield
(Wu et al., 1998). According to Assuero and Tognetti (2010), high tiller production capacity
improves the chances of persistence after periods of unfavourable environmental conditions
during which a plant can experience biotic or abiotic stress. It means that there are higher
chances of survival after a biotic stress as one or more tillers can take over after stress has
affected the initial stems.
Strigolactones are involved in the regulation of above ground plant architecture by inhibiting
tiller development or shoot branching (Umehara et al., 2010). Jamil et al., (2011) found an
inverse relationship between number of tillers per plant and strigolactone production. This

123
may mean that high tillering varieties are producers of low strigolactones which gives an
advantage of supporting a lower number of S. asiatica parasites. Tillering therefore becomes
a useful trait that can be incorporated into other cultivars. According to Khush et al., (1999),
tillering increased the widespread adoption of wheat and rice varieties in the 1960s because
the altered plant architecture averted severe food shortages and was an essential component
of the green revolution. Jahn et al., (2011) found that continuous tillering is associated with
high grain yield and is characteristically selected for in advanced varieties in most crops. This
study showed that tillering was higher in uninfested pots than infested ones. This coincides
with the findings of Cruz and Boval (2000) who found a positive effect of nitrogen
availability on tillering in Lolium perenne. In this study, it could be possible that the nitrogen
levels in the Striga infested pots were not sufficient to sustain the direct demands of the
parasite and the tillering processes of the sorghum plants.
The germination of S. asiatica is elicited by strigolactones from the hosts. Therefore the
number of Striga attached is a reflection to some extent of the quantity of strigolactones
produced by the host. In a study by Jamil et al., (2011), the germination of Striga was
dependent on the quantity and quality of strigolactones produced by the host. As a result, any
genetic variation in this trait could potentially confer pre-attachment resistance. Varieties
Hlubi, Mashava, Ruzangwaya and Tshwetha are likely to be high strigolactone producers as
they stimulated a lot of seeds to germinate. Mukadziusaende and Chiredhi are likely to be
low strigolactone producers as they elicited the germination of a few parasites.
The heights of varieties Chiredhi, Mukadziusaende and Isifumbate were not significantly
lowered by Striga infestation. Mukadziusaende and Chiredhi resisted the dwarfing effects of
S. asiatica. Reduction in plant growth rate is caused by changes in the growth regulators in
the host (Frost et al., 1997). Taylor et al., (1996) reported that the increased levels of the
plant growth regulator, ABA in infested plants may result in stomatal closure. The reduction

124
in photosynthesis and ultimately growth would explain the reduction in height of infested
plants that was observed in this study.
The differences in plant height observed between infested and uninfested genotypes for some
varieties reflected that the parasite acted as a sink for carbon, inorganic solutes and water.
This was confirmed by Gurney et al., (2002), Cechin and Press (1993), and Gurney et al.,
(1995). The sorghum genotypes that maintained plant height irrespective of infestation
demonstrated tolerance. The results showed that at 6, 8 and 12 WACE for the 2013/14
season, the varieties Mukadziusaende, Chiredhi and Isifumbathe maintained their plant
height. The same genotypes were able to maintain high chlorophyll content, were
independent of mulch and infection with respect to chlorophyll content, maintained high
stomatal conductance and supported low Striga numbers. The first two parameters constitute
the photosynthetic machinery, implying that photosynthesis was not affected by infection in
those genotypes and hence they were able to maintain plant height despite the parasite load.
For the varieties Tshwetha, SC Sila and Ruzangwaya, mulched and infested pots had taller
plants than unmulched and uninfested ones demonstrating that in drier seasons, moisture
availability can negate the effects of infestation. The absence of mulch caused the plant to be
affected more than by infection alone. Despite the fact that sorghum is a drought tolerant
crop, absence of mulch which regulates soil temperature and conserve soil moisture had
deleterious effects on plant height. This is supported by Chakraborty et al., (2008) and
Mupangwa et al., (2012) who reported the merits of mulching in moisture conservation and
temperature regulation in hot and dry environments.
The ability of the varieties to maintain grain yield when infested by Striga asiatica may result
from the ability of the cultivars to maintain higher rates of photosynthesis (Gurney et al.,
2002). Striga asiatica therefore has little effect on the physiology of these varieties. Infact,

125
Mukadziusaende had higher yields in infested than uninfested pots, a phenomenon that was
attributed to profuse tillering. Jahn et al., (2011) reported on rice varieties that continued to
tiller after the main stem begins senescence. Striga spends about 7 weeks underground and
about 3 weeks above ground, after which it sets seed and dies. Mukadziusaende and Chiredhi
matured early and the main stem dried, many tillers formed and these matured very early
which lead to increased yield. Tillering seems to be the major parameter for tolerance under
infested conditions. Mukadziusaende had the highest number of tillers and chlorophyll
content and all these added together may provide a suite that overcompensates for Striga
infestation to the extent that yield becomes higher under infested than uninfested conditions.
The ability to tiller when the main stem approaches senescence therefore becomes a
beneficial trait in response to Striga infestation.
Varieties Isifumbathe, Mukadziusaende and Mashava had the highest number of tillers. The
same varieties supported the least number of S. asiatica. The data showed that there was an
inverse relationship between the number of tillers per plant and S. asiatica parasitism. Similar
results were found by Jamil et al., (2011) in rice cultivars where there was an inverse
relationship between number of tillers in rice and the number of Striga supported. Their study
quantified strigolactones and found that they were also inversely related to tillering in rice
cultivars. This research did not quantify strigolactones but their quantities can be inferred
from the Striga asiatica attached together with tillering of sorghum. The results imply that
Mukadziusaende is a low strigolactone producer which is why it had the highest tiller
number. Hlubi, Ruzangwaya and SC Sila supported the highest number of Striga asiatica and
had lower number of tillers so they are likely to be producers of low quantities of
strigolactones. The tillering of sorghum can therefore be used as a measure of sorghum
tolerance when selecting genotypes for use. Chlorophyll content and growth rate can be used

126
as parameters for tolerance to Striga. When all these traits are combined, they can
characterise a sorghum variety in terms of its ability to resist and tolerate Striga.
This study demonstrated substantial sorghum genetic variation for chlorophyll content, Striga
number, tillering, stem height, internode length and grain yield in response to mulching. The
sorghum varieties Mukadziusaende and Chiredhi were Striga tolerant, because of their ability
to maintain high chlorophyll content, stem height, high internode length values, high grain
yields and supporting low Striga attachments. The other two sorghum varieties were
Ruzangwaya with high chlorophyll content and maintenance of stem heights under Striga
infection, and Mashava with high tillering ability and high grain yield production. The results
clearly indicated that varieties Mukadziusaende and Chiredhi were able to perform well in
parasite infested pots.
4.6 Conclusions
Mulching increased chlorophyll content and sorghum height in the drier season of 2014/15
compared to the 2013/14 wetter season. Yield and dry matter traits were maintained under S.
asiatica infestation in the drier season of 2014/15. Therefore, mulching negated the effects of
drought spells in the drier seasons. In the wetter season the effects of mulching were not
apparent.
6.0 References
Adekalu, KO, Olorinfemi IA, Osunbitan JA (2007). Grass mulching effect on on infiltration,
surface runoff from two southwestern Nigerian agricultural soils. Bioresource Technology
98: 912 - 917

127
Assuero SG and Tognetti JA (2010). Tillering regulation by endogeneous and environmental
factors and its agricultural management. The Americas Journal of Plant Science and
Biotechnology. Global Science Books.
Blum A (2005). Drought resistance, water use efficiency and yield potential- are they
compatible, dissonant r mutually exclusive. Australian Journal of Agric Research 56: 1159 -
1168
Bouwmeester, HJ, Matusova, R, Sun Zhongkui and Beale, MH (2003). Secondary metabolite
signalling in host parasitic plant interactions. Current Opinions in Plant Biology 6: 358 –
384.
Cechin I and Press M.C (1993) Nitrogen relationships of the Sorghum-Striga hermonthica
host parasite associations: growth and photosynthesis. Plant cell and Environment 16: 237 -
247
Chakraborty D, Nagarajan S, Agrawal P, gupta VK, Tomar RK, Garg RN, Sahoo RN, Sarkar
A, Chopra UK, Samar KSS, Kalra N (2008). Effect of mulching on soil and plant water and
the growth and yield of wheat (Triticum aestivum) in a semi-arid environment. Agric Water
Management 95: 1323 - 1334
Cruz P and Boval M (2000). Effect of nitrogen on some morphogenic traits of temperate and
tropical forage grasses. In Lemaire G’ Hogson J, De MOraes A, Cavacho P.C, de F, Nabinger
C (eds). Grassland ecophysiology and grazing ecology. CAB International. Wallingford, PP
134 – 150
Ejeta G (2007). The Striga scourge in Africa: a growing pandemic. In Integrating new
technologies for Striga control: towards ending the witch hunt (Ejeta G, Gressel J Eds), pp 3
– 16. World Scientific publishing company Private Limited.

128
Frost. DL, Gurney AL, Press, MC, Scholes J (1997). Striga hermonhica reduces
photosynthesis in sorghum: the importance of stomatal limitations and potential role of ABA.
Plant Cell and environment 20: 483 – 492.
Gitelson, AA; Gritz U, Merzlyak, MN (2003). Relationship between leaf chlorophyll content
and spectral reflectance and algorithms for non destructive chlorophyll assessment in higher
plants. Journal of plant Physiology 160: 271 – 282.
Gilley, JE and Kottwitz, ER (1992). Beginning of motion for selected unanchored residue
materials. Journal of irrigation and drainage engineering 118: 619 – 630
Gilley JE, Knowittz ER, Wieman GA (1991). Roughness coefficients for selected residue
materials. Journal of irrigation and drainage engineering 117: 503 – 514.
Gurney A.L, Ransom J.K and Press M.C (1995). The parasitic angiosperm Striga
hermonthica can reduce photosynthesis of its sorghum and maize hosts in the field. Journal
of Experimental Botany 46: 1817 – 1823
Gurney AL, Grimanelli D, Kanampiu F, Hoisington D, Scholes DJ, Press MC (2003). Novel
sources resistance to Striga hermonthica in Tripsacum dactyloides, a wild relative of maize.
New Phytologist 160: 557 – 568
Gurney AL, press, MC and Scholes JD (2002). Can wild relatives of sorghum provide new
sources of resistance or tolerance against Striga spp. Weed Research 42: 317 - 324
Jahn CE, McKay JK, Mauleon R, Stephens J, McNally KL, Bush DR, Leung H and Leach JE
(2011). Genetic variation in biomass traits among 20 diverse rice varieties. Plant Physiology
155 (1): 157-168

129
Jamil, M; Van Mourik, TA, Charnikhova T and Bouwmester, HJ (2013). Effect of
diammonium phosphate application on strigolactone production and Striga hermonthica
infection in three sorghum cultivars. Weed Research 53: 121 – 130.
Jamil M, Charnikhova T, Houshyani B, van Ast A, Boumeester HJ (2011) Genetic variation
in Strigolactone production and tillering in rice and its effects on Striga hermonthica
infection. Planta. DOI10:1007/s00425-011-1520-y.
Khush GS (1999). Green revolution: preparing for the 21st century. Genome 42: 646 – 655
Mupangwa W, Twomlow S, Walker S (2012). Reduced tillage, mulching and rotational
effects on maize, cowpea and sorghum yields under semi-arid conditions. Field Crops
Research 132: 139 - 148
Mwenje E (2006). Sorghum and its wild relatives: ecological implications in the event of the
release of transgenic (herbicide/Bt) sorghum in the SADC region. Biotechnology, biosafety
and Environment in Southern Africa 2: (1): 5 - 6
Parker C. and Riches, CR (1993). Parasitic weeds of the world: biology and control. CAB
International, Wallingford, UK.
Rao, K, Steenhuis, T, Cogle A, Srinivasan, S, Yule D, Smith G (1998). Rainfall infiltration
and runoff from an alfisol in semi-arid india. 1. No till systems. Soil and Tillage Res 48: 51 -
59
Riddle WC, Gillespie TJ, Swanton CJ (1996). Rye mulch characterisation for purposes of
microclimatic modeling. Agric for Meteorology 78: 67 - 81
Rich, PJ; Greinier, C. and Ejeta, G. (2004). Striga resistance in the wild relatives of Sorghum.
Crop Science 44: 2221 – 2229.

130
Scopel E, Muller, B, Areola Tostado J, Chavez Guera E, Maraux F (1998). Quantifying and
modeling the effects of a light crop residue on the water balance: an application to rainfed
maize in western Mexico. World congress on soil science, France
Tari G, Lasckay G, Takacs Z, and Poor P. (2013). Response of sorghum to abiotic stresses: a
review. Journal of Agronomy and Crop Science 199: 264 – 274.
Taylor A, Martin J, Seel WE (1996). Psychology of the parasitic association between maize
and witch weed Striga hermonthica; Is ABA involved. Journal of Experimental Botany 47:
1057 - 1065
Umehara M, Hanada A, Yoshida S, Akiyama K, Arite T, Takeya-Kamiya N, Magome H,
Kamiya Y, Shirasu K, Yoneyama K, Kyozuka J and Yomaguchi S (2008). Inhibition of shoot
branching branching by new terpenoid plant hormones. Nature 455: 195 – 200.
Vincent V, Thomas, RG (1961). An agricultural survey of southern Rhodesia. Part 1:
Agroecological regions survey. Salisbury. Rhodesia. Ministry of Agriculture.
Wu GW, Wilson LT, Mclung AM (1998). Contribution of rice tillers to dry matter
accumulation and yield. Agronomy Journal 90: 317 - 323

131
CHAPTER FIVE
The existence of different physiological ‘strains’ of Striga asiatica
(L.) kuntze on Sorghum bicolor (L.) Moench and Sorghum
arundinaceum (desv) Stapf in Zimbabwe
5.1 Abstract
A better understanding of the virulence variability of S. asiatica populations and host parasite
interactions is essential for more efficient resistant material deployment. This study was
stimulated by the observation that sorghum that is tolerant to Striga asiatica in a particular
area could still lack the same tolerance to a strain native to remote areas.Therefore an
experiment was designed with the objective of determining the stability of Sorghum spp
tolerance to two Striga strains. Nine Sorghum bicolor and one Sorghum arundinaceaum
(Wild sorghum) genotypes were subjected to two Striga asiatica strains sourced from
Rushinga and Chiundura, which are 500 km apart, and a control. A 3*10 factorial experiment
was set up, arranged in a completely randomized design with three replications at two sites.
The data collected were sorghum height, chlorophyll content, tiller number and dry matter
traits. The results showed that sorghum genotypes differed significantly (P<0.05) in their
response to the stunting effects of Striga strains at both sites. The Chiundura strain was more
virulent to Isifumbathe, Zambia, Wild sorghum and Mashava at Henderson research station
whilst at BUSE, the Chiundura strain was more virulent on wild sorghum and Mashava. The
two Striga strains were generally similar on their effects on chlorophyll content. At
Henderson, the Chiundura strain reduced chlorophyll content of Chiredhi, Zambia, Tshwetha
and SC Sila. At BUSE both strains were not significantly different (P<0.01) from each other

132
at 10 weeks after crop emergence (WACE). The effects of the two strains were similar for
head index, root index, stem index and leaf index at both sites. The Chiundura strain reduced
the total dry weight at BUSE but not at Henderson. Overally, the Chiundura strain had more
deleterious effects on sorghum traits compared to the Rushinga strain confirming the
existence of physiological speciation on Striga asiatica in Zimbabwe.
Key words: Striga asiatica, sorghum, Striga strains, dry matter partitioning
5.2 Introduction
Sorghum (Sorghum bicolor) is an important food crop in sub Saharan Africa where plant
available water is often a limiting factor for crop production. Sorghum allows for
maintenance of yield stability in arid environments. Despite cultivating the crop in more than
60 % of the world hectarage, African sorghum constitutes 37 % of the world’s production and
the average yield in most African countries is about 0.9 tha-1
which is substantially lower than
the world average of 1.4 t ha-1
(Jamil et al., 2012). The major biotic constraint to sorghum
production in the southern Africa sub-region is Striga asiatica, a hemi-parasitic weed. Striga
competes effectively with the host for carbon, nitrogen and inorganic solutes (Gurney et al.,
1999) and also causes phytotoxic effects on the host plants within days of attachment (Frost
et al., 2005; Frost et al., 1997). The witchweeds are obligate hemi-parasites and although
they contain some photosynthetic capacity, Striga species have an absolute requirement for
the host in order to develop and complete their life cycle (Aly, 2007).
Host resistance to Striga has been proposed as the best method of control. According to
Mohammed et al., (2007), crops with some measure of resistance have been integrated into
Striga management programmes and the new material got challenged by the Striga seed
bank. The newly introduced sorghum genotype must be able to cope with the great potential
genetic diversity in the seed bank. Lewin (1939) was the first to observe the possible

133
existence of biological strains in Striga to explain the differential damage and distribution of
Striga on wild and cultivated hosts in sub Saharan Africa. Jones (1955) reported that the
major problem in Striga control is the possible existence of physiological speciation within
the species. Ten years later, Doggett (1965), reported varietal differences in sorghum
susceptibility to Striga hermonthica sourced from different places in east Africa. It was
observed that varieties of sorghum resistant in one location became susceptible in another
location thus suggesting the existence of physiological strains of the parasite.
Some workers have reported that resistance could be dependent on the virulence of the Striga
strains (Parker and Reid, 1979; Ramaiah, 1987). A study by Riches et al., (1992) on cowpeas
found occurrence of distinct varietal specificity among races of the S. gesneroides. This
meant existence of genetic divergence in Striga. Reda et al., (2010) found that there were
populations of Striga that represented a threat to resistant varieties. Virulence could depend
on the history of agriculture and specific characteristics of the source environment
(Polniaszek et al., 1991).
Botanga and Timko (2006) suggested that geographical isolation and host driven selection are
important factors in the formation of races of Striga gesneriodes in West Africa. S. asiatica
and S. hermonthica populations from Kenya were respectively polymorphic for 96.8 % and
84 % of the amplified fragment length polymorphic (AFLP) bands that were assessed (Gethi
et al., 2005). A study in Benin discovered a high degree of host speciation within the 14 S.
asiatica populations that were analysed (Botanga et al., 2002). According to Spallek et al.,
(2013), S. asiatica isolated from wild grasses spp were unable to successfully parasitise
sorghum and maize plants susceptible to other Striga asiatica collections.

134
In Zimbabwe, studies conducted by Musimwa et al. (2001) using RAPD-PCR markers found
great genetic distances among Striga from three places which are sparsed from one another
by about 400 kilometres.
The objective of this study were
i) to determine the differential virulence effects of two srains of S. asiatica to sorghum
genotypes with respect to growth and dry matter traits in sorghum.
ii) to establish the existence of different strains of S. asiatica in Zimbabwe
There the alternative hypothesis were
i) the two S. asiatica strains were differentially virulent on sorghum growth and
dry matter parameters.
ii) there exists physiological strains in S. asiatica in Zimbabwe
5.3 Materials and Methods
5.3.1 Experimental sites
Two experiments were carried out at Henderson Research Station in Mazowe and the other
one at Bindura University of Science Education (BUSE) nursery. Henderson Research station
is located at latitude17.340S and longitude 30.58
0E. BUSE is located at the coordinates: 17
o
18I 58II S and 31o 19
I 23IIE. The two sites are located in agro-ecological region IIa of
Zimbabwe (Vincent and Thomas, 1961) and receive about 800 mm rainfall per annum. The
average temperature for both areas was 25oC. The experiments were conducted in the 2013 -
14 cropping season. The soil used at both sites was medium grained sands with a pH of 4.1
(CaCl2) and with a phosphorus content of 20 parts per million (ppm), 4 ppm nitrogen.

135
Exchangeable cations in milligram equivalent per 100 g of soil were 0.06 for potassium, 0.67
calcium and 0.48 magnesium.
5.3.2 Source of sorghum genotypes and Striga asiatica strains
Sorghum bicolor genotypes were sourced from the gene bank at the Department of Research
and specialist services, Harare research station, Zimbabwe. The Chiundura S. asiatica strain
was sourced from the Midlands province of Zimbabwe, which is 500 km from Rushinga in
Mashonaland province where the Rushinga strain was sourced. Sorghum arundinaceum seed
was collected in arable lands at Gwebi Agricultural College, 27 km west of Harare.
5.3.3 Experimental design
The experiments were conducted in pots with a 3*10 factorial treatment structure. The
sorghum genotypes that were used are shown in Section 4.3.2.
5.3.4 Experimental details
Medium grained sandy soil was obtained from the top 15 cm of a farmer’s field where Striga
has never been reported. Six litre plastic pots with six drainage holes at the bottom were filled
with the soil, and the top 10 cm of the soil were thoroughly mixed with 5.7 g of Compound D
fertilizer (8%N, 14% P2O5, 7 K2O) and 1 g of Striga asiatica seed. Ten sorghum seeds were
planted in the plastic pots at a depth of 0.5 cm and watered. The two strains were each used
to infest a third of the buckets and the other third was uninfested. The pot filling and planting
process started with the uninfested pots to avoid contamination. The sorghum seedlings were
thinned to one plant/pot at two WACE. The sorghum plants were topdressed at a rate of 3 g
per pot using ammonium nitrate (34.5 %N) at 4 WACE.

136
5.3.5 Data Collection
Height of the sorghum plants was measured using a metre ruler from the soil level to the
growing point. Tiller number was counted on each plant. Chlorophyll content was measured
by a chlorophyll meter (SPAD-502 KONICA MINOLTA INCL) at 6, 8 and 12 WACE.
Striga numbers were determined by counting the number of Striga plants that emerged from
the pots. The whole sorghum plant was carefully uprooted at maturity and the total dry matter
was separated into roots, leaves and stems. Head weight was determined by drying the
sorghum head in the sun until moisture content was 14 %. Roots, leaves and stems were dried
in an oven at 78oC for 48 hours for dry mass determination. Various indices were calculated
by dividing mass of the part, like leaf mass, by total mass.
5.3.6 Data Analysis
Barlett’s test for homogeneity of variance was done and the variances were not
homogeneous. Hence, the data from the two sites were analysed separately. Data was
subjected to analysis of variance (ANOVA) to determine the treatment effects using Genstat
release 14 (VSN International, UK).
5.4 Results
5.4.1 Sorghum plant height
Sorghum genotypes differed significantly (P< 0.01) in plant height in response to the two
different Striga strains. There was a significant interaction (P<0.05) of Striga strain and
sorghum genotype. The Chiundura strain lowered the height of Isifumbathe, Zambia and wild
sorghum 6 WACE at Henderson research station (Figure 5.1).

137
Henderson plant height at 6 WACE
Sorghum genotypes
ChiredhiHlubi
IsifumbatheZambia
Tshwetha
Mukadziusaende
RuzangwayaSC Sila
Wild Sorghum
Mashava
Sor
ghum
hei
ght (
cm)
15
20
25
30
35
40
45
50
Rushinga
Chiundura
Uninfested

138
Sorghum genotypes
ChiredhiHlubi
IsifumbatheZambia
Tshwetha
Mukadziusaende
RuzangwayaSC Sila
Wild Sorghum
Mashava
Pla
nt h
eigh
t (cm
)
10
20
30
40
50
60
70
80
90
Rushinga
Chiundura
Uninfested
Sorghum genotypes
ChiredhiHlubi
IsifumbatheZambia
Tshwetha
Mukadziusaende
RuzangwayaSC Sila
Wild Sorghum
Mashava
Pla
nt h
eigh
t (cm
)
0
20
40
60
80
100
120
140
160
180
200
220
Rushinga
Chiundura
Uninfested
Figure 5.1: Interaction effects of sorghum genotypes and Striga strain on sorghum
height at 6, 8 and 12 WACE at Henderson research station.

139
At 8 WACE, the Chiundura strain significantly lowered the height of genotype Isifumbathe.
At 12 WACE, the same strain significantly lowered the height of Zambia, Wild sorghum and
Mashava genotypes compared to Rushinga strain and uninfested genotypes. The Rushinga
strain significantly (P<0.05) dwarfed Tshwetha compared to Chiundura and uninfested pots
(Figure 5.1). Mukadziusaende and Ruzangwaya genotypes resisted the dwarfing effects of the
Striga strains. The two strains had similar effects on Chiredhi and Isifumbathe genotypes
(Figure 5.1).
Sorghum genotype
Chiredhi
Hlubi
Isifumbathe
Zambia
Tshwetha
Mukadziusaende
Ruzangwaya
SC Sila
Wild Sorghum
Mashava
Pla
nt h
eigh
t (cm
)
-50
0
50
100
150
200
250
Rushinga
Chiundura
Uninfested
Figure 5.2: Interaction effects of sorghum genotype and Striga strains at 12 WACE at BUSE
A significant interaction (P<0.05) of sorghum genotype and Striga strains was observed for
plant height. The Chiundura strain dwarfed wild sorghum and Mashava genotypes compared
to Rushinga and the control at BUSE. Rushinga lowered the height of Tshwetha genotype
compared to Chiundura strain (Figure 5.2).

140
Table 5.2: Effect of Striga strain on plant height at 6 and 12 WACE at Henderson and
BUSE
6 WACE 12 WACE
Henderson BUSE Henderson BUSE
Rushinga 32.67±1.029a 32.51
a 114.6±2.461
a 101.4±9.58
a
Chiundura 28.00±1.029b 30.62
a 111.6±2.461
a 98.9±9.58
a
Uninfested 34.23±1.029c 33.86
a 138.3±2.461
b 122.9±9.58
b
Values followed by different superscripts are significantly different at P<0.05
Across the measured periods Chiundura elicited the lowest sorghum height at all sites (Table
5.2).
5.4.2 Chlorophyll concentration
Chlorophyll content was significantly affected (P<0.001) by Striga strains at 8 and 12 WACE
(Table 5.3). The results revealed that the two strains were equally virulent on chlorophyll
content but were significantly lower compared to uninfested pots at both sites at 8 and 12
WACE (Table 5.3)

141
Table 5.3: Effect of Striga strains on chlorophyll content at 8 and 12 WACE
Chlorophyll content (mmolcm-2
)
Accesions 8WACE 12 WACE
BUSE Henderson BUSE Henderson
Rushinga 26.48±0.749a 35.67±0.815
a 29.54±0.784
a 30.66±0.975
a
Chiundura 26.56±0.749a 34.53±0.815
a 30.74±0.784
a 31.21±0.975
a
Uninfested 29.45±0.749b 40.16±0.815
b 33.01±0.784
b 37.45±0.975
b
Values followed by different superscripts are significantly different at P<0.001
The genotype Mukadziusaende resisted the effects of the two strains with respect to
chlorophyll content (Figure 5.3). The two strains were equally virulent on Hlubi, SC Sila,
Mashava and Ruzangwaya (Figure 5.3) and the results were similar at both sites. The
Rushinga strain elicited the lowest chlorophyll content (Figure 5.3) across all sorghum
genotypes.

142
Sorghum genotypes
Chiredhi
Hlubi
Isifumbathe
Zambia
Tshwetha
Mukadziusaende
Ruzangwaya
SC Sila
Wild Sorghum
Mashava
Chl
orop
hyll
cont
ent (
mm
olcm
-2)
0
10
20
30
40
50
60Rushinga
Chiundura
Uninfested

143
Sorghum genotypes
Chiredhi
Hlubi
Isifumbathe
Zambia
Tshwetha
Mukadziusaende
Ruzangwaya
SC Sila
Wild Sorghum
Mashava
Chl
orop
hyll
cont
ent (
mm
olcm
-2)
0
10
20
30
40
50
Rushinga
Chiundura
Uninfested
Figure 5.3: Interaction effects of sorghum genotype and Striga strains on chlorophyll
content at 10 WACE at both sites
5.4.3 Sorghum tillering
At 8 WACE, tiller number was significantly (P<0.001) affected by sorghum genotype and by
Striga strain (P<0.05) (Figure 5.4). The genotypes Chiredhi, Ruzangwaya and Mashava had
the biggest tiller numbers, while Hlubi, Zambia and SC Sila had the least.

144
Sorghum varieties
Chiredhi
Hlubi
Isifumbathe
Zambia
Tswetha
Makadziusaende
RuzangwayaSC Sila
Wild Sorghum
Mashava
Tille
r num
ber
0
2
4
6
8
10
Sorghum varieties
ChiredhiHlubi
IsifumbatheZambia
Tswetha
Makadziusaende
RuzangwayaSC Sila
Wild Sorghum
Mashava
Tille
r num
ber
0
2
4
6
8
10
12
Figure 5.4: Effect of sorghum variety on tiller number at 12 WACE at both sites
At Henderson research Station, the sorghum genotypes with the highest tiller numbers were
Chiredhi, Ruzangwaya, Mashava and Wild sorghum. At BUSE, Wild sorghum, Mashava and
Ruzangwaya had the highest tiller numbers. At both sites the lowest tillers were on genotypes
Zambia and SC Sila. The Chiundura strain elicited the highest number of tillers compared to
infestation using Rushinga strain and uninfested sorghum genotypes (Figure 5.5).

145
Striga strains
Rushinga Chiwundura Uninfested
Tille
r num
ber
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Figure 5.5: Effect of Striga strains on tiller number.
5.4.4 Sorghum dry matter traits
Sorghum genotype had a significant (P<0.001) effect on head, root, stem and leaf indices at
both sites (Table 5.4)

146
Table 5.4: Effect of sorghum genotypes on head index, root index, stem index and leaf index at BUSE
and Henderson sites.
Head Index Root Index Stem Index Leaf Index
Sorghum
variety or
landrace
Henders
on
BUSE Henders
on
BUSE Henders
on
BUS
E
Henders
on
BUSE
Chiredhi 0.31d 0.40
e 0.19
a 0.29
a 0.42
c 0.23
bc 0.074
a 0.073
b
c
Hlubi 0.18b 0.21
c 0.29
bcd 0.48
cd 0.458
cd 0.24
bc 0.069
a 0.074
b
c
Isifumbathe 0.24c 0.319
d 0.287
abcd 0.413
b 0.412
bc 0.216
bc
0.057a 0.051
a
Zambia 0.208bc
0.239c 0.207
ab 0.367
a 0.497
cd 0.289
c
0.088b 0.104
e
Tshwetha 0.089a 0.142
a 0.315
cd 0.531
d 0.527
d 0.244
bc
0.068a 0.082
c
d
Mukadziusaen
de
0.232bc
0.315
5d
0.287abcd
0.409b 0.394
bc 0.215
bc
0.066a 0.06
ab
Ruzangwaya 0.171b 0.200
c 0.342
d 0.498
b
cd
0.392b 0.207
b
0.094b 0.094
d
e
SC Sila 0.1881b 0.185
b 0.285
abcd 0.514
c
d
0.408bc
0.176d
0.118c 0.1230
e
Wild Sorghum 0.077a 0.082
7a
0.493ef 0.676
e 0.32
b 0.161
a
0.108bc
0.079b
cd
Mashava 0.168b 0.119
a 0.518
f 0.688
e 0.213
a 0.118
a
0.10b 0.074
b
c
Sed 0.023 0.037 0.05 0.048 0.053 0.037 0.009 0.01
Means followed by different superscripts in the same column are significantly different at P<0.001.
Head index was significantly higher on Chiredhi, Isifumbathe, Zambia and Mukadziusaende
while the lowest indices were recorded for Tshwetha and wild sorghum genotypes at both
sites. Root index was highest on Tshwetha, Mashava and wild sorghum genotypes (Table 5.4)
whilst the lowest were Chiredhi, Mukadziusaende, Zambia and SC Sila genotypes (Table
5.4). Stem index was lowest on Mukadziusaende, Ruzangwaya and Wild sorghum genotypes.
The Striga strains significantly affected the four indices at Henderson but not at BUSE (Table
5.5).

147
Table 5.5: Effect of Striga strains on head, root, stem and leaf index at BUSE and
Henderson.
Head Index Root index Stem index Leaf index
Striga
strain
Henderson BUSE Henderson BUSE Henderson BUSE Henderson BUSE
Rushinga 0.1658±0.0123a 0.1785±0.02a 0.388±0.027a 0.566±0.027a 0.37a 0.181±0.029a 0.076±0.005a 0.0741a
Chiundura 0.1861±0.0123a 0.213±0.02a 0.349±0.027a 0.516±0.027a 0.366a 0.185±0.029a 0.1±0.005b 0.0856a
Uninfested 0.2151±0.0123b 0.272±0.02b 0.229±0.027b 0.377±0.027b 0.477b 0.2646±0.029b 0.078±0.005a 0.0857a
Means followed by different letters are significantly different at P<0.001
Both strains led to more dry matter being allocated to the root at both sites. The two Striga
strains reduced the biomass allocated to the stems at both sites compared to uninfested hosts.
Head index was also lowered by Striga infestations. Leaf index was independent of Striga
strain at BUSE but it increased significantly (P<0.001) under the Chiundura strain compared
to Rushinga strain and uninfested genotypes.
There was a significant interaction (P<0.05) of sorghum genotype and Striga strain on leaf
index at the Henderson site (Figure 5.6).

148
. Sorghum genotypes
Chiredhi
Hlubi
Isifumbathe
Zambia
Tshwetha
Mukadziusaende
Ruzangwaya
SC Sila
Wild Sorghum
Mashava
Leaf
inde
x
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0.20
Rushinga
Chiundura
Uninfested
Figure 5.6: Interaction effects of sorghum genotype and Striga strains at Henderson.
Leaf indices were similar for Chiredhi, Hlubi, Isifumbathe, Zambia, Mukadziusaende and
Ruzangwaya genotypes. The genotypes SC Sila, Wild sorghum and Mashava increased their
leaf indices under infestation by Chiundura strain compared to uninfested sorghum
genotypes. The Chiundura strain stimulated formation of more leaf tissue in wild sorghum
and SC Sila because of the higher leaf indices observed for these genotypes (Figure 5.6).
5.4.5 Sorghum head weight
There was a significant (P<0.05) interaction of Striga strain and sorghum genotype on
sorghum head weight at Henderson research Station and at BUSE (P<0.05) (Figure 5.7).

149
Sorghum genotypes
Chiredhi
Hlubi
Isifumbathe
Zambia
Tshwetha
Mukadziusaende
Ruzangwaya
SC Sila
Wild Sorghum
Mashava
Hea
d w
eigh
t (g)
0
10
20
30
40
50
60
70
Rushinga
Chiundura
Uninfested
Head weight at BUSE
Sorghum genotypes
ChiredhiHlubi
IsifumbatheZambia
Tswetha
Mukadziusaende
RuzangwayaSC Sila
Wild Sorghum
Mashava
Sor
ghum
hea
d w
eigh
t (g)
0
10
20
30
40
50
60
70
Rushinga
Chiundura
Uninfested
Figure 5.7: Interaction effects of sorghum genotypes and Striga strain at BUSE and
Henderson

150
At both sites, the head weight of Hlubi, Mukadziusaende and Wild sorghum genotypes were
not affected by the Striga strains. The effects of Rushinga and Chiundura strains were similar
for Ruzangwaya and SC Sila genotypes and they were lower than uninfested genotypes
(Figure 5.7).
5.4.6 Total dry weight
The total dry weight was significantly affected by Striga strain (P<0.05) at BUSE, but was
not affected at Henderson Research Station (Figure 5.8). The Rushinga strain gave the
highest total dry weight at BUSE compared to the Chiundura strain and uninfested genotypes.
At Henderson, the total dry weight was not affected by the Striga strains (Figure 5.8).
Striga strains
Rushinga Chiundura Uninfested
Tota
l dry
mat
ter (
g)
0
50
100
150
200
250
BUSE
Henderson
Figure 5.8: Total dry matter of sorghum genotypes for BUSE and Henderson

151
total dry mass at BUSE
Sorghum genotype
ChiredhiHlubi
IsifumbatheZambia
Tswetha
Mukadziusaende
RuzangwayaSC Sila
Wild Sorghum
Mashava
Tota
l dry
mass
(g)
0
50
100
150
200
250
300
350
Rushinga
Chiundura
Uninfested
Figure 5.9: Interaction effects of sorghum genotype and Striga strain on total dry mass
at BUSE
There was a significant interaction (P <0.05) between variety and Striga strain for total dry
matter. Uninfested Isifumbathe, Zambia, Ruzangwaya, SC Sila and Mashava genotypes had
significantly higher (P<0.05) head weight compared to infested genotypes. The Chiwundura
strain had higher virulence on head weight for the varieties Mashava, Zambia and Chiredhi.
The head weight of genotypes Hlubi, Tshwetha, Mukadziusaende, Ruzangwaya and wild
sorghum was not significantly reduced by the Striga strains (Figure 5.9).
Total dry matter was not significantly (P >0.05) affected by both Striga strains for the
genotypes Chiredhi, Hlubi, Isifumbathe, Zambia, Tshwetha, Hlubi, Isifumbathe,

152
Mukadziusaende, Ruzangwaya and SC Sila. The Chiwundura strain significantly (P<0.05)
lowered the total dry matter of the genotypes wild sorghum and Mashava.
5.5 Discussion
The objective of this study was to determine the stability of sorghum tolerance when infested
with two Striga asiatica strains. The ability to express tolerance in a wider range of
environments indicates plasticity which is an important component for yield maintenance.
According to Poorter et al., (2012), the plant has to balance the allocation of dry matter to
leaves, stems and roots in a way that matches the physiological activities and functions
performed by these organs.
Generally, the Chiundura strain was more virulent in dwarfing the sorghum genotypes
compared to the Rushinga strain. The sorghum genotypes which resisted the dwarfing effects
of the Striga were Hlubi, Zambia, Mukadziusaende, and Ruzangwaya. The two strains
differed in their effects on sorghum lines. The results concur with the findings of Campos et
al., (2004) who reported that the response of plants to stress is genotype specific. Other
studies by Taylor et al., (1996) found that the plant growth regulator abscissic acid (ABA)
increases in Striga infested plants and this leads to a reduction of stomatal conductance which
consequently reduced carbon assimilation. In tolerant maize genotypes, Gurney et al., (2002)
found two maize varieties in which Striga had limited effect on photosynthesis despite
infection. In this study, failure by the Striga strains to dwarf Zambia, Mukadziusaende and
Ruzangwaya may be a demonstration that the genotypes are less responsive to ABA hence
may be labelled as tolerant with respect to height. In sorghum genotypes such as Mashava,
wild sorghum, SC Sila and Chiredhi, differential effects of the Striga strains were observed.
Flexas et al., (2006) attributed reduction in plant growth from imposition of stress to changes
in the partitioning of assimilates between different organs and the balance between
photosynthesis and respiration.

153
The differential effects of the Striga strains on plant height was demonstrated in this study.
In this case, the Chiundura strain was the more virulent than the Rushinga strain. However,
Richards et al., (2006) asserted that a given host genotype may be plastic for a certain trait in
a certain set of environments but not plastic for other traits in the same environments.
The effects of the two strains on chlorophyll content was not significantly different but all
were significantly less than the uninfested genotypes. The chlorophyll content of the
genotypes Ruzangwaya, Mukadziusaende and Mashava was not affected by the Striga strains
at both sites. These results were similar to observations by Gurney et al., (2002), who
reported that a tolerant maize variety was able to maintain high chlorophyll content levels
despite being infested. Chlorophylls are the light harvesting complexes and tolerant
genotypes should adjust chlorophyll content to be independent to keep photosynthesis at
optimal rate. Aly (2007) reported that the host proteins are exported to the parasite. The
ability of a genotype to maintain chlorophyll content irrespective of infection is key to
tolerance. Chlorophyll content is a key parameter in tolerant genotypes as it depicts the extent
of plant canopies and the subsequent carbon assimilation.
The varieties which lacked tolerance could have been caused by perturbations in host carbon
assimilation that could also limit nitrogen assimilation and in-turn lower chlorophyll
synthesis. This may explain the lower chlorophyll content of the infested Tshwetha,
Isifumbathe, Hlubi and SC Sila genotypes.
Across all genotypes, the root index for Rushinga was 0.388 at Henderson and 0.566 at
BUSE. Chiundura produced 0.349 at Henderson and 0.516 at BUSE against 0.22 and 0.377
for uninfested genotypes at Henderson and BUSE respectively. This indicated that sorghum
genotypes allocated more dry matter to roots in response to Striga infestations. This
behaviour coincides with that established by Poorter and Nagel (2000) and, Shipley and

154
Meziane (2002) who reported a proportional increase in root fraction as soil available
moisture decreases. A study by Acciaressi and Guiamet (2010) on Sorghum halepense found
that the witchweeds incite a host to increase the amount of biomass allocated to roots when
they are stressed. The root parameter is important because the spatial deployment of roots
determines the ability of a plant to secure edaphic resources. According to Campos et al.,
(2004) and Reynolds et al., (2007), there is evidence to suggest that adverse effects of stress
can successfully be avoided by changing the carbon allocation patterns to allow formation of
a deep root system before the onset of a growth limiting stress. In this study, the sorghum
genotypes differed in the allocation of dry matter to the roots. The lowest root indices were
recorded for genotypes Chiredhi, Zambia, Isifumbathe and Mukadziusaende. The highest
performing genotype was Mashava with 0.518 and wild sorghum with 0.493. The varieties
that respond to Striga infestations by allocating more dry matter to roots at the expense of
yield are non tolerant to Striga in terms of grain yield. The genotypes which show plasticity
to Striga infestation and do not alter the allocated carbohydrates to the roots show plasticity
to yield. Therefore, the varieties Chiredhi, Zambia, Mukadziusaende, Isifumbathe and SC
Sila are plastic to Striga infestations in terms of dry matter allocated to roots.
Striga strain did not lower sorghum head weight compared to uninfested sorghum for
genotypes Chiredhi, Hlubi, Tswetha, Mukadziusaende, wild sorghum and Mashava at both
sites. However, Tswetha and wild sorghum had low yields. This is expected for wild sorghum
as it has not undergone any improvement to increase its yield. The genotypes have the
capacity to diminish the consequences of infection. According to Swabrick et al., (2008),
different cultivars differ in their capacity to tolerate the physiological and pathological effects
of Striga parasitism finally resulting in milder or stronger impacts on crop yield. Despite the
fact that several studies (Gurney et al., 2002; Rodenburg et al., 2005), have shown a
reduction in yield after Striga infestation, it may be that these varieties uncouple the process

155
of photosynthesis and dry matter allocation. Similar results were found by Frost et al., (1997)
in sorghum: Striga hermonthica associations. The results confirm the assertion by
Haussmann et al., (2001) that genetic variation for tolerance to Striga under field conditions
exists in cultivated sorghums especially in local African cultivars. This may mean that Striga
does not cause serious changes in the metabolism of some hosts.
Genotypes Isifumbathe, Zambia, Ruzangwaya and SC Sila had their head weight depressed
by Striga. The results showed a depression in the dry matter allocated to the head. The
reduced head weight index may partly result from the parasite acting as a sink of carbon,
inorganic solutes and water and also lower rates of carbon gain by infested cereals (Cechin
and Press, 1993). According to Joel (2000), the parasite develops as a strong metabolic sink,
relative to the host and channels the flow of water and nutrients to itself thereby damaging the
crops’ development. In this study, the stems made up the bulk of the biomass. The same
results were found by Jahn et al., (2010) in rice cultivars.
The Chiundura and Rushinga strains had similar effects on head, root, and stem indices, and
stem weight. This is in agreement with the findings of Dube and Belzile (2010) who found
very low levels of genetic diversity among Striga gesneroides and this was attributed to a
high degree of selfing. The results point to the fact that there are minor genetic variations in
the virulence of S. asiatica strains on the stated sorghum traits. According to Loveless and
Hamrick (1984), S. asiatica is mainly autogamous and this can limit genetic diversity. Studies
done in Kenya by Gethi et al., (2005) showed very low genetic diversity and the reason that
was advanced was that the parasitic weed had recently been introduced in Kenya. In Benin,
Botanga et al., (2002) showed host speciation of Striga asiatica. In our study, the results
indicated that there are particular processes and parameters of sorghum which were affected
more by a particular strain in a particular genotype. These results may indicate that the strain
seriously reduce dry matter accumulation in the affected varieties. When the same varieties

156
are grown in areas in which the Rushinga strain dominates the genotypes may be labelled as
resistant only to lose resistance when grown where the Chiwundura strain dominates. It can
therefore be inferred that the Striga strains each has physiological processes that it affects
more in a particular genotype. In some sorghum genotypes, particular strains affected
chlorophyll content, in some height; in some they caused allocation of dry matter to be more
to the roots than the harvestable part.
5.6 Conclusion
It can therefore be concluded that there was differential virulence of Striga asiatica strains on
plant height, chlorophyll content, tiller production, different sorghum indices and yield. This
study confirms the variable virulence and the existence of physiological strains of Striga
asiatica in Sorghum species in Zimbabwe.
5.7 References
Acciaresi HA, Guiamet JJ (2010).Below and above ground growth and biomass allocation in
maize and Sorghum halepense in response to water competition. Weed Research 50: 481 –
492.
Aly R (2007) Conventional and biotechnological approaches for control of parasitic weeds.
In vitro cell Dev Biology Plant 43: 304 – 317
Bebawi FF (1981). Interspecific physiological variants of Striga hermonthica. Experimental
Agriculture 17: 419 - 423
Botanga C, Kling J, Berner D,Timko M (2002). Genetic variability of Striga asiatica L.
Kuntze based on AFLP analysis and host parasite interactions. Euphytica 128: 375 – 388
Botanga CJ,Timko MP (2006) Phenetic relationship among different races of Striga
gesneroides (Willd) vatke from West Africa. Genome 49: 1351 1365

157
Campos H, Cooper A, Halburn JE, Edmead GO, Schussler J.R. (2004). Improving drought
tolerance in maize: view from industry. Field Crops Research 90: 19 – 34.
Cechin I and Press M.C (1993). Nitrogen relations of the Striga hermonthica host parasite
associations: growth and photosynthesis. Plant cell and environment 16: 237 - 247
Doggett H (1965). Striga hermonthica on Sorghum in East Africa. Journal of Agricultural
Science 65: 183 – 194.
Dube M-P and Belzile FJ (2010). Low genetic variability of Striga gesneroides population on
cowpeas might be explained by recent origin. Weed Research 50: 493 - 502
Flexas J, Bota J, Galmes J, Medrano H, Ribus-Carbo M (2006). Keeping a positive carbon
balance under adverse conditions: response of photosynthesis and respiration to water
stress.PhysiologiaPlantarum 127: 343 - 352
Frost DL; Gurney AL., Press MC, Scholes JD (1997). Striga hermonthica reduces
photosynthesis in sorghum: the importance of stomatal limitations and potential role of ABA.
Plant cell and environment 20: 4873 – 4492
Gethi JG Smith ME Mitchell SE and Kresovich S (2005).Genetic Structure of Striga
hermonthica and Striga asiatica population of Kenya. Weed Research 45: 64 – 73.
Gurney, AL, Press M.C and Scholes J.D (1999).Infection time and density influence the
response of Sorghum to the parasitic angiosperm, Striga hermonthica. New Phytologist 143:
573 – 580
Gurney, A.L, taylor A, Mbwaga A, Scholes J.D, Press M.C. (2002). Do maize cultivars
demonstrate tolerance to the parasitic weed Striga asiatica. Weed research 42: 299- 306

158
Gurney, A.L, Slate J, Press M.C and Scholes J.D. (2006). A novel form of resistance in rice
to the angiosperm parasite Striga hermonthica. New Phytologist 169: 199 – 208.
Hausmann B.I.G, Hess, D.E’ Welz H.G, Geiger, H.H (2000). Improved technologies for
breeding Striga resistant sorghums. Field crops Research 66: 195 - 211
Jamil M, Charnikhova T, Houshyani B, van Ast, A, Bouwmesster, H (2012), Genetic
variation in strigolactones production and tillering in rice and its effects on Striga
hermonthica infection. Planta. DOI10.1007/s00425-011-1520-y
Jahn C.E, McKay J.K, Mauleon R, Stephens J, McNally K.L, Bush D.R, Leung H and Leach
J.E (2011). Genetic variation in biomass traits among 20 diverse rice varieties. Plant
physiology 155 (1): 157-168
Joel D.M (2000). The long term approach to parasitic weed control: manipulation of specific
developmental mechanisms of the parasite. Crop protection 19: 753 – 758
Jones W (1955). Further experiments in witch weed control 11 Existence of strans of Striga
hermonthica. Empire Journal of Exp Agric 23: 206
Lewin C.H. (1939). Witchweeds (Striga lutea, Lour, Var Bicolor, O. Kuntze. Northern
Rhodesia Department of Agriculture Bulletin 2: 51 – 52
Lichtenhaler A.K and Wellburn L.R. (1983). Determination of total carotenoids and
chlorophyll a and b of leaf extracts in different solvents. Biochemical Society Transactions
11: 591 - 592
Loveless M.D and Hamrick J.K (1984). Annual Reviews of Ecological systems 15: 65 - 73

159
Musimwa C, Tongoona P, Mutengwa C.S. and Chivinge O.A (2001). Genetic variations in
the witch weed inferred from RADP-PCR markers. African crop Science Proceedings 5: 51 –
55.
Pinheiro C and Chaves M.M (2011). Photosynthesis and drought: can we make metabolic
connections from available data. Journal of Experimental Botany 62: 869 – 882.
Polniazek T.I, Parker C, Riches C.R (1991). Variation in virulence of Alectra vogelii
populations in cowpeas. Tropical Pest Management 37: 152 - 154
Poorter H and Nagel O (2000). The role of biomass allocation in the growth response of
plants to different levels of light, carbon dioxide, nutrients and water: a quantitative review.
Australian journal of Plant Physiology 27: 595 – 607
Poorter H, Niklas K.J, Reich P.B, Olekysyn J, Poot P and mommer L (2012). Biomass
allocation to leaves, stems and roots: meta analysis of interspecific variation and
environmental control. New Phytologist 193: 30 - 50
Ramaiah K.V (1987). Breeding cereals grains for resistance to wicthweeds. In Parasitic
weeds in agriculture ed Musselman L.J .CRC Press. Pages 227 - 242
Reda F, DierickA,andVerkleij J.A.C (2010). Virulence study of Striga hermonthica
populations from the Tigray Region (northern Ethiopia). World Journal of Agricultural
Sciences 6 (6): 676 - 682
Reynolds, M., Calderini, D., Condon, A. & Vargas, M., 2007. Association of source-sink
traits with yield, biomass and radiation use efficiency among random sister lines from three
wheat crosses in a high yield environment. Journal of Agricultural Science, 145: .3-16.

160
Richard C.L, Bossdorf O, Muth N.Z, gurevitch J and Pigliucci (2006).Jack of all trades,
master of some?On the role of phenotypic plasticity in plant invasions. Ecology letters 9: 981
- 993
Riches C.R, Hamilton K.A, Parker C (1992). Parasitism in grain legumes by Alectra spp.
Annals of Applied biology 121: 362 - 370
Parker C and Reid D.C (1979).Host specificity in striga spp, some preliminary observation.
Supplement to the proceedings of the second International Symposium on Parasitic weeds.
Pages 79 - 90
Rodenburg J, Bastiaans L, Weltzien E and Hess D (2005. How can field selection of for
Striga resistance and tolerance in Sorghum be improved. Field Crops Research 93: 34 – 50
Spallek T, Mutuku J.M and Shirasu K (2013). The genus Striga: a witch profile. Molecular
Plant Pathology.DOI10.1111.mpp.12058.
Swarbrick P.J; Huang K and Liu G (2008). Global patterns of gene expression in rice
cultivars undergoing a susceptible or resistant interaction with the parasitic Striga
hermonthica. New Phytologist 179: 515 – 529.
Taylor A, Martin J, Seel W.E, (1996). Physiology of parasitic associations between maize
and witchweed (Striga hermonthica): Is ABA involved. Journal of Experimental botany 47:
1057 – 1065.
Vincent V and Thomas R.G (1961). An Agricultura survey of southern Rhodesia: agricultural
regions survey. Salisbury, Rhodesia. Ministry of agriculture.

161
CHAPTER SIX
Sorghum response to Striga asiatica based on maximum
germination distance, Striga counts and sorghum tillering in
Zimbabwe
6.1 Abstract
Resistance through reduced strigolactones is one of the sustainable ways of managing Striga
asiatica. To verify the existence of reduced strigolactone production in sorghum genotypes,
an agar gel assay was carried out on seven Sorghum bicolor lines and one Sorghum
arundinaceaum sourced in Zimbabwe. The eight sorghum genotypes were also grown in sand
and Striga attachments and sorghum yield were recorded. The results indicated that sorghum
genotypes varied significantly (P<0.05) with respect to maximum germination distance
(MGD) with wild sorghum and SC Sila having the largest MGDs. The genotype
Mukadziusaende had the highest tiller numbers (P<0.05), while SC Sila had the lowest.
Striga counts were highest on Wild Sorghum, Ruzangwaya and Hlubi. There was a negative
correlation coefficient (R2 = 0.2225) between Mgd and tiller number, showing that the
highest strigolactone producers had low tiller numbers. A positive correlation coefficient (P=
0.843) was found between sorghum yield and tiller numbers and it indicates that the more the
tiller numbers the more the yield. It can therefore be concluded that resistance through
reduced strigolactones was found in the sorghum genotype Mukadziusaende. The direct
relationship between MGD and tillering means that tiller number can be used to select for
reduced strigolactone production in the field.

162
6.2 Introduction
The average sorghum productivity in the sub-Saharan Africa is low as a result of a myriad of
production constraints of which Striga asiatica is a major obstacle. Striga asiatica is an
obligate hemi-parasite that attaches to the roots of several crop species leading to severe yield
losses (Parker, 1991). Beyond the burden of losing food and water to these parasites, host
plants suffer from a characteristic malady resembling the symptoms of severe drought. Striga
spp parasitise important food crops such as sorghum, maize, millets and rice. They have
functional chloroplasts but their photosynthates provide only part of the energy required for
survival, hence are regarded as hemi-parasites (Xie et al. 2010). This hemi-parasite has long
been recognized as the greatest biological constraint to food production in sub Saharan
Africa, causing annual losses in excess of US$7 billion (Samejina et al., 2016).
Striga asiatica is geographically the most widespread species with large populations having
been reported throughout sub Saharan Africa, south east China and the Indian subcontinent
while smaller isolated populations have been reported in Arabia, Indonesia, Phillipines, north
and east Carolina (USA) and Australia (Cochrane and Press, 1997). Striga asiatica is the
predominant species towards the east African coast and southern Africa (Parker, 2009).
According to Bouwmeester et al., (2003), these parasites infest about two thirds of the 70
million hectares used for cereal production in Africa. Jamil et al., (2011) asserted that about
20 – 80 % yield losses or even complete crop failure can occur due to Striga parasitism.
The life cycle of the noxious cereal weed has co-evolved with many hosts to comprise a
series of discrete steps that are closely linked to the host’s biochemistry (Hearne, 2009). The
root parasitic weed has developed the ability to germinate only when they are exposed to
germination stimulants released from the host roots, thus syncronising their life cycle to those
of their potential hosts. Thus, they only germinate when a suitable host seed is in proximity to
the Striga seed (Fernandez-Aparacio, et al., 2009). Fernadez-Aparacio et al., (2011) reported

163
that the synchrony is vital for parasitic weed survival because they have an absolute
requirement for nutritional support from the host. This complex life cycle also presents
opportunities for disruption (Pierce et al. 2003).
According to Bouwmeester et al., (2003) and Akiyama and Hayashi (2006), the first critical
step in the life cycle of Striga, the germination of its seed, is regulated by strigolactones. The
dependence on strigolactones could be exploited for Striga management through breeding for
low strigolactones producing cultivars. The seeds of these parasitic plants will only germinate
after perceiving a germination stimulant of their host (Yoneyama et al., 2010). After radicle
emergence, the haustoria attaches and penetrates the host roots (Yoder, 2001). Cardoso et al.,
(2011) reported that once germination has been triggered, the radicle protrudes from the testa,
elongates towards the root and develops haustorium, an organ that can attach to and penetrate
roots of the host plant. The parasitic plant grows underground for 4 – 7 weeks prior to
emergence and utilises host water, nutrients and photosynthates (Jamil et al., 2011). Much of
the damage will have occurred by the time the Striga emerges above the ground.
Jamil et al., (2011) found significant variation among NERICA rice cultivars and their
parents for strigolactones production and Striga germination. Production of low germination
stimulants results in low numbers of Striga asiatica attachments thereby producing a resistant
phenotype. Low germination stimulant producing genotypes have enhanced resistance to
Striga because of the reduction in Striga germination (Dun et al., 2009). Striga resistance is
the mechanism that ensures lower field infestation and allows for satisfactorily high yields
than fully susceptible ones (Rodenburg et al., 2005). Studies done in sorghum have also
shown that genotypes with low production of germination stimulants have demonstrated
resistance to Striga in the field (Ramaiah, 1987; Hess et al., 1992; Ejeta, 2007). In the past
decade, several sorghum varieties with Striga resistance based on low germination stimulant
production have been introduced such as SRN 39 and IS9830 in Sudan and Gobiye in

164
Ethiopia (Tefera et al., 2012). However, resistance alone is not sufficient for adoption of a
variety as adaptability to the environment and farmer preference have to be taken into
account (Mohemed et al., 2016). This raises the need to identify resistant genotypes among
those cultivated by farmers locally. Since the root parasites affect the crop from the time they
attach to the root, the development of new control strategies should focus on the initial steps
in host parasite interaction (Lopez-Raez et al., 2008).
Ejeta et al., (2000), Wilson et al., (2000), Gurney et al., (2001), and Gurney et al., (2002)
demonstrated that the near relatives of cereals could provide new sources of tolerance and or
resistance to parasite infection and may provide the way forward for the control of Striga spp.
According to Doggett (1976, 1988) and De Wet (1978), the cultivated sorghums of today
primarily originated in Africa from the wild Sorghum bicolor spp arundinaceaum. Southern
Africa has more sorghum landraces and whilst the quest to find a landrace that produces the
least strigolactones is still on, there is need to look at the wild sorghum and the vast number
of sorghum landraces that are under cultivation in the African savannah. Given the high
genetic diversity of the Sorghum spp, including wild sorghum, there is need to quantify
strigolactones in most of these cultivated lines as they are grown in Striga infested fields.
According to Ejeta et al., (1993), different sorghum genotypes differed by as much as a
billion fold in the amount of germination stimulants they produce. Jamil et al., (2011),
asserted that strigolactones have a triple role which is underground communication between
the plant, AM fungi, parasitic plants and the regulation of tillering. According to Umehara et
al., (2008), and Lopez-Raez et al., (2008), strigolactones inhibit tillering in plants and
therefore the ability to tiller could be used as a selection criteria for reduced strigolactones
production.Therefore, the objectives of this study were to:
i) identify low strigolactone producing sorghum lines using agar gel analysis.

165
ii) establish the relationships between maximum germination distance (MGD), sorghum
tillering, Striga asiatica counts and sorghum yield.
The corresponding alternate hypothese were:
i) the Sorghum genotypes vary in strigolactones production, resulting in variable
resistance to Striga asiatica.
ii) maximum germination distance is inversely related to sorghum tillering, while it is
positively related to Striga asiatica counts.
iii) MGD is inversely related to sorghum yield.
6.3 Materials and methods
6.3.1 Experiment 1: Agar jel assays
6.3.2 Sorghum germplasm and Striga asiatica seed sources
Refer to section 3.2.2
6.3.3 Experimental design
The treatments for this experiment were the sorghum genotypes (Table 3.1). They were
arranged in a completely randomized design replicated four times.
6.3.4 Surface Sterilisation and sorghum seed germination
Sorghum seeds were soaked in 1 % sodium hypohlorite solution for 60 minutes and rinsed in
double deionised water. The seeds were soaked in an aqeous solution of 10 % captan
overnight. Seeds were rinsed with deionised water three times and then incubated in moist
filter paper at 27 oC. After 48 hours, germinating seeds were placed in agar plates as outlined
by Hess et al.(1992).

166
6.3.5 Conditioning of Striga seed
Striga asiatica seeds were placed in 30 ml sample bottles and rinsed three times by adding 3-
5 drops of the detergent tween 20 into 10 ml of distilled water. Sonication was done using an
ultra sonic cleaner three minutes during the first rinse (Mutengwa, 2004). The Striga seeds
were incubated at 27 oC for three days prior to transferring them into the fresh sterile flasks
containing 15 ml of 0.001 % acqeous benomyl solution. The sample bottles were re-
incubated at 25 oC for 35 days before they were ready for use in the agar jel assay.
6.3.6 The assay set up
Pre-conditioned Striga seeds were pipetted into petri dishes. Water agar was then poured over
the seed. The roots of the germinating sorghum seeds were placed in the solidifying agar with
the root tip pointing across the plate. The plates were incubated in the dark for five days. The
MGD (distance between the host root and the furthest germinated Striga seed) were used as
indicators of the quantities of strigolactones produced. The MGDs were recorded using a
graduated microscope at 120 hours of incubation time at 30oC. .
6.4 Experiment 2: Pot screening
About 1g of S. asiatica seeds were weighed for every treatment and mixed thoroughly with 2
Kg of washed river sand. The sand was sterilized before the start of the experiment by
heating in an oven at 120 oC for 48 hours to kill any Striga weed seeds that could be in the
sand. The method was adapted from Jamil et al., (2011). Plastic pots of dimensions 18 cm
diameter and 20 cm height were used.
6.4.1 Experimental design, planting and data collection
There were 8 genotypes (Table 3.1) and the experiment was replicated four times and laid
down as a completely randomized design. Five sorghum seeds were planted in pots at a depth

167
of 0.5 cm. The seedlings were thinned to one plant per pot at 2 WACE. The sorghum plants
were allowed to grow in pots for 20 weeks and the Striga that emerged were counted. The
number of tillers, maximum germination distance were also recorded. Yield data was also
recorded at the end of the experiment.
6.4.2 Data analysis
The data was analysed according to the model:
Yij = µ + Ti +eij
Yij was the measured parameter e.g tiller number, µ = general mean, Ti effect of the treatment
and eij is the error term. Correlations of the measured parameters were determined using
Satistical Package for Social Sciences and Genstat version 12.
6.5 Results
6.5.1 Maximum germination distance (MGD)
The sorghum genotypes differed strongly (P=0.001) with regard to maximum germination
distance, which is indicative of the strigolactones quantity produced (Figure 6.1). The
minimum germination distance was 1.225 cm and it was for Mukadziusaende whilst the
maximum was 2.775 cm for SC Sila (Figure 6.1).

168
Sorghum genotypes
Mukadziusaende
Wild sorghumSC Sila
Chiredhi
Ruzangwaya
Isifumbathe
Hlubi
Zambia
Ger
min
atio
n di
stan
ce (c
m)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Figure 6.1: Maximum germination differences for various sorghum genotypes
6.5.2 Tillering
Sorghum genotypes varied strongly (P <0.01) in tillering, and the average number of tillers varied
from 2 for SC Sila to 5 for Mukadziusaende (Figure 6.2). The genotypes that had the highest number
of tillers were Mukadziusaende and wild sorghum while the lowest were SC Sila and Zambia (Figure
6.2).

169
Sorghum genotypes
Mukadziusaende
Wild sorghumSC Sila
Chiredhi
Ruzangwaya
Isifumbathe
Hlubi
Zambia
Tille
r num
ber
0
1
2
3
4
5
6
Figure 6.2: Effect of Striga asiatica on tillering of sorghum genotypes
6.5.3 Striga counts
Sorghum genotypes
Mukadziusaende
Wild sorghumSC Sila
Chiredhi
Ruzangwaya
IsifumbatheHlubi
Zambia
Strig
a co
unts
0
2
4
6
8
10
12
14
16
18
Figure 6.3: Effect of Sorghum genotypes on Striga counts

170
The sorghum genotypes varied strongly in eliciting Striga germination (P<0.01). The
genotypes that had the highest number of Striga counts were Ruzangwaya and Hlubi, while
the lowest were Mukadziusaende and Chiredhi (Figure 6.3).
6.5.3 Correlations between maximum germination distance and tillering
The results indicate a negative relationship between MGD and tillering (R2 = 0.2225) (Figure
6.4). The relationship indicates that as MGD increases tillering decreases. A positive
relationship between tiller numbers and yield (R2
=0.8436) indicates that as tiller numbers
increase yield also increases (Figure 6.5).

171
Figure 6.4: The relationship between germination distance and tiller numbers
Figure 6.5: Relationships between sorghum yield and tiller numbers
6.6 Discussion
The objectives of this study were to identify sorghum genotypes that produced the lowest
strigolactones, and then correlate strigolactones production to tillering, Striga counts and
yield. It is noteworthy that, among all the genotypes evaluated n the present study the
genotype Mukadziusaende was identified as the resistant genotype producing low amount of
strigolactones and sustained lower Striga emergence. The sorghum genotypes with lowest
maximum germination distances were Mukadziusaende, Chiredhi and Isifumbathe, whilst
wild sorghum and SC Sila had the biggest maximum germination distances. However, the
level of susceptibility differed among the genotypes. This is supported by Haussmann et al.,
(2000) who reported that the Striga species negatively affects the growth and yield of crops
they infect but the extent of the negative effects is a function of the environment and genetic
makeup of the host and the parasite.
These results are consistent with previous observations in cereals like rice (Jamil et al., 2011).
The results confirm the existence of large genetic variation among the sorghum cultivars.

172
According to Jamil et al., (2011), up to about 500-fold differences exist in the amounts of
strigolactones exuded by rice Germplasm. Ejeta et al., (1993) confirmed the same results in
sorghum and reported differences as much as a billion fold in the amounts of stimulants
produced by sorghum.
Tillering varied among sorghum genotypes with Mukadziusaende having the highest tiller
number whilst the lowest was SC Sila (Figure 6.2). In sorghum, tillering has been proposed to
be under genetic and environmental control and according to Kim et al., (2010), tillering has
not been comprehensively addressed by the carbohydrate supply and demand framework. In
this study the sorghum genotypes were subjected to the same environmental conditions hence
the environmental influences were eliminated. Therefore, the differences suggested that
sorghum genotypes may also differ in their propensity to tiller which is independent of the
carbon supply demand (Kim et al., 2010). The differences could be due to differences in
hormonal signaling as the plants were in the same environment. According to Umehara et al.,
(2008), the hormone strigolactone reduces tillering in plants, such that plants that produce
less strigolactones have profuse tillering compared to those that produce more. The results of
this study support the propensity to tiller hypothesis as the genotypes were grown in the same
conditions.
Increase in MGD indicates susceptibility of the genotype to Striga infestations. This means
reduced MGD gives a resistant phenotype which inturn gives higher grain yields. The results
corroborates previous findings by Mohemed et al., (2016) on the relationship between
germination stimulant activity and Striga infestation. Smaller MGDs mean low amounts of
active stimulants which confer pre-attachment resistance and accounts for yield mantainance
in the sorghum genotypes. There was a significant correlation between MGD and Striga
counts. To the contrary, Haussmann et al., (2000) compared the results of pot trials and only
weak correlations between germination distance in agar jel assay and the number of emerged

173
Striga plants. However, Rodenburg et al., (2005) reported high correlation coefficient
between root exudates and infection levels as illustrated by Striga counts.
6.7 Conclusion
Among the set of eight sorghum genotypes screened under pot conditions in Zimbabwe,
significant differences were found in their levels of resistance. The genotype
Mukadziusaende showed excellent levels of resistance as illustrated by lower MGDs and
Striga counts. The genotypes SC Sila and wild sorghum were the most susceptible. Yield of
sorghum was inversely related to germination distance and tiller number increased as
sorghum yield.
6.8 References
Akiyama K and Hayashi H (2006). Strigolactones: chemical signals for fungal symbionts and
parasitic weeds in plant roots. Annals of Botany 97: 925 – 931.
Bouwmeester, H.J, Matusova R, Sun Z, Beale M.H (2003). Secondary metabolic signaling in
host-parasite interaction. Current Opinions in Plant Biology 6: 358 – 384.
Cardoso C, Ruyteer-Spira C, Bouwmeester H.J (2011). Strigolactones and root infestation by
plant parasiti Striga, Orobanche and Phillipanche spp. Plant Science 180: 414 - 420
Cochrane V and Press M.C (1997). Geographical distribution and aspects of ecology of the
hemiparasitic angiosperm Striga asiatica (L) Kuntze: a herbarium study. Journal of Tropical
Ecology 13: 371 - 380
De Wet J.M.J (1978). Systematics and evolution of sorghum sect.Sorghum (graminae).
American Journal of Botany 65: 477 – 484
Doggett H (1976). Sorghum. In evolution of Crop Plants (eds Simmonds N.W) 112 – 117.
Longman group, new York. USA.

174
Doggett H (1988). Sorghum.2nd
.longman group. London. UK.
Dun EA, Brewer PB, Beveridge CA (2009). Strigolactones: discovery of the elusive shoot
branching hormone. Trends in Plant Science 14: 364 - 372
Ejeta G, Mohammed P, Rich A, merlake-berhan T.L, Housely T.L, Hess D.E (2000).
Selection for specific mechanisms of resistance to Striga in sorghum.In breeding for Striga
resistance in cereals. Proceedings of a workshop held at IITA, Ibadan, Nigeria (eds B.I.G
Haussmann, Hess D.E, Koyama L.M, L. Grivet, Rattunde H.F.W and Geiger H.H. 29 – 37.
Margragverlag. Welkersheim, Germany.
Ejeta G (2007). Breeding for resistance in sorghum: exploitation of an intricate host-parasite
biology. Crop Science 47: 216 - 227
Fernandez-Aparicio M, Westwood J.H and Rubiales D (2011). Agronomic, breeding and
biotechnological approaches to parasitic plant management through manipulation of
germination stimulant level in agricultural soils. Botany 89: 813 – 826.
Fernandez-aparacio M, Flores F and Rubiales D (2009). Recognition of root exudates by
seeds of boomrape, orobanche and phelipanche spp. Annals of Botany (London) 103: 423 -
431
Gurney A.L, Press M.C and Scholes D.J (2002). Can wild relatives of sorghum provide new
sources of resistance or tolerance against Striga spp. Weed Research 42: 317 – 324
Gurney A.L, Grimanelli D, Kanampiu F.K, Hoisinhton, D.A, Scholes J.D, press M.C (2001).
Can wid relatives of cereals provide new sources of resistance to the parasitic angiosperm
Striga?. In proceedings of the Seventh international parasitic weed Symposium (eds A. Fer, P
Thoalouran, D.M. Joel, L.J. Mussellman, C. parker, and J verleij. 140 Nantes. France.

175
Haussman BIG, Hess DE, Weltz HG, Geiger HH (2000). Improved methodologies for
breeding Striga-resistant sorghum. Field Crops Research 66: 195 – 201.
Hearne S.J. (2009). Control-Striga conundurum. Pest Management Science 65: 603 – 614
Hess D.E, Ejeta G, Butler L.G (1992).Selecting sorghum genotypes expressing a quantitative
expressing a quantitative biosynthetic trait that confers resistance to Striga. Phytochemistry
31: 493 – 497
Jamil M, Rodeburg J, Charnikhova T, and Bouwmeester H.J (2011).Pre-attachment Striga
hermonthica resistance of New Rice for Africa (NERICA) cultivars based on low
strigolactones production. New Phytologist 192: 964 – 975.
Jamil M, charnikhova T, Houshyani B, van Ast A, Bouwmeester H.J (2011). Genetic
variation in strigolactones production and tillering in rice and its effect on Striga hermonthica
infection. Planta DOI10.1007/s00425-011-1520-y.
Lopez-Raez J.A, Charnikhova, T, Gome-Rolden V, Matusova R, Kohlen, W, de Vos R
(2008). Tomato strigolactones are derived from the carotenoides and their biosynthesis is
promoted by phosphate starvation. New Phytologist 178: 863 – 874.
Mohemed N, Charnikhova T, Bakker E.J, van Aast A, Babiker AGT, Bouwmeester HJ
(2016). Evaluating of field resistance to Striga hermonthica in Sorghum bicolor: the
relationship with strigolactones. Pest Management Science 72: 2082 – 2090.
Parker C (1991).Protection of crops against parasitic weeds. Crop Protection 10: 6 – 22.
Parker C (2009).Observations on the current status of orobanche and Striga problems
worldwide. Pest Management Science 65: 453 – 459.

176
Pierce S, Mbwaga AM, Press MC and Schles JD (2003). Xenognosin production and
tolerance to Striga asiatica infection of high yielding maize cultivars. Weed Research 43: 139
– 145
Ramaiah K.V. (1987). Breeding cereal grains for resistance to witchweed. New York, USA.
Academic Press.
Rodenburg J, Batiaans L, Weltzien E, Hess DE (2005). How can field selection for Striga
resistance and tolerance in sorghum be improved? Field Crops Research 93: 34 – 50.
Rodenburg J, Cissoko M, kayeke J, Dieng I, Khan ZR, Midega C.A.O, Onyuka E.A, Scholes
J.D (2015). Do NERICA rice cultivars express resistance to S. hermontica and S asiatica
under field conditions. Field Crops Research 170: 83 – 94.
Rodenburg J (2005). The role of sorghum genotypes in the interaction with the parasitic weed
Striga hermonthica. PHD thesis. Wageningen University. The Netherlands.
Samejina H, Babiker AG, Takikawa H, Sasaki M, Sugimoto Y (2016). Practicality of suicidal
germination approach for controlling Strga hermonthica. Pest management Science
DOI10.1002/ps4215.
Tefera H, Reston VA, Dahlberg J, Smale M (2012). External evaluation report on sorghum,
millets and other grains. Instormil collaborative research Support programme.
Umehara, M, Hanada A, Magome H, Takeda-Kamiya, Yomaguchi S (2010). Contribution of
strigolactones on the inhibition of tiller bud outgrowth under phosphate deficiency conditions
in rice. Plant Cell Phydsiology 51 (7): 1118 - 1126

177
Wilson J.P, Hess D.E, Hanna W.W (2000).Resistance to Striga hermonthica in wild
accessions of the primary gene pool of Pennisetumglaucum. Phytopathology 90: 1169 –
1172.
Xie X, Yoneyama K, Yoneyama K (2010). The strigolactones story. Annual Review of
Phytopathology 48: 93 - 117
Yoder J.T (2001). Host-plant recognition by parasitic scrophulariaceae. Current Opinions on
Plant Biology 4: 359 – 365.
Yoeneyama K, Awad A.A, Xie X, Yoneyama K, takeuchi Y (2010). Strigolactones as
germination stimulants for root parasitic plants. Plant Cell Physiology 51 (7): 1095 – 1103.

178
CHAPTER SEVEN: GENERAL DISCUSSION, CONCLUSIONS AND
RECOMMENDATIONS
7.1 Introduction
This chapter is a synopsis of the whole study in relation to the objectives and the major
findings. The main objectives of the study are presented together with major findings and
conclusions. Lastly, a section on recommendations for further research and for farmers who
produce sorghum in Striga asiatica infested fields are presented.
7.2 Discussion
The ‘niche area’ of the study or the recommendation domain is the small holder sorghum
producers in Zimbabwe and the whole of tropical sub Saharan Africa. Literature review
showed that communal farmers have serious problems in sorghum production as a result of
Striga infestation sometimes resulting in 100 % losses (Berner et al. 1995, Mabasa, 2003,
Rubiales et al. 2009). Management of Striga still remains a major challenge faced by small
holder farmers in Zimbabwe. Results reported elsewhere especially for rice has promising
results through resistance and tolerance (Jamil et al., 2012, Sun et al. 2007, Jamil et al. 2011)
Sorghum genotypes were evaluated on their resistance and tolerance to Striga. This study
investigated the resilience of Striga infested sorghum under drought, mulch and under
various Striga asiatica strains. The study also sought to determine sorghum resistance to S.
asiatica with respect to reduced strigolactone production.
When growing sorghum in Striga infested fields which are prone to drought, the drought
exacerbates the effects of Striga in sorghum. However, there are genotypic differences on
sorghum responses to the two stresses. The sorghum line Mukadziusaende emerged as a
better performer when exposed to both stresses. On growth, water availability and Striga
infestation had the same effect on sorghum. Reduced water availability on Striga infested
sorghum strongly reduced sorghum growth. It is therefore important to subject sorghum

179
varieties to multiple stresses common in the field especially at evaluation stages as
simultaneous occurrence of the stress is common in the subtropical sub Saharan Africa.
Mulching at a rate of 3 tons/ha enables expression of tolerance traits in some sorghum
genotypes and negates the effects of Striga parasitism. Mulching at 3 ton/ha as promoted in
CA has been reported to suppress weeds through physical impedance and allelopathy. When
mulch is aplied, there is increased water penetration into the soil with subsequent prevention
of excessive evapo-transpiration losses (Scopel et al. 1998, Rao et al. 1998, Adekalu et al.
2007). Striga affects the water economy of the sorghum plant hence increased moisture
availability through mulch protects sorghum from the effects of Striga. Tolerance expression
is improved under mulch. Mulching is currently being promoted as a component of
conservation agriculture so adoption can cushion the small scale farmers from the effects of
Striga asiatica parasitism. Mulching increased chlorophyll concentration compared to
unmulched pots which in-turn maintained sorghum productivity. Mulching was advantageous
in the drier season compared to the wetter season. Drier seasons are more common in sub
Saharan Africa due to the effects of climate change. Again, the ability to tiller gave
genotypes higher yield as some genotypes continued to produce tillers after being affected by
Striga.
The problem of the existence of Striga asiatica strains was confirmed in Zimbabwe in this
study. Differential effects were observed when similar genotypes were subjected to the two
strains. This dimension complicates Striga asiatica management in the smallholder sector.
Trials done on supposedly resistant materials should be done all over the country to
determine if they can withstand local Striga asiatica diversity. Sorghum genotypes tended to
vary in their response to the strains.

180
No genotype was completely resistant to Striga but were differentially sensitive. The
genotype Mukadziusaende produced the least strigolactones which in turn produces a
resistant phenotype. Wild sorghum and SC Sila (registered variety) were the highest
producers of strigolactones. Sorghum tillering was inversely related to strigolactones and this
confirmed what is found in literature (Jamil et al. 2012). It implies that farmers may use
tillers to predict strigolactone production in the field. The researchers may select for resistant
lines using tiller numbers. Higher tillers are lower producers of strigolactones.
7.3 Conclusions
The following conclusions can be made from the results of this study
1. Reduced water availability caused by droughts increase the effects of Striga asiatica
on sorghum although some genotypes may withstand the pressure.
2. Reduced water availability and Striga asiatica infestation had mutually exclusive
effects on chlorophyll concentration and NDVI.
3. The use of mulch benefits small holder farmers who grow their sorghum in Striga
infested fields since the mulch negates the effects of Striga especially under low
rainfall conditions.
4. There is existence of Striga asiatica physiological strains in Zimbabwe and this calls
for widespread trials on any material to be deemed resistant to the parasite.
5. Mukadziusaende had the least MGD and this means it was the least strigolactones
producer and got fewer Striga attachments and hence it produced the resistant
phenotype.

181
6. Higher tiller producers in sorghum indicates reduced strigolactones producers hence
researchers can use it to select for resistant phenotypes.
7. Sorghum arundinaceaum was found to be susceptible to Striga asiatica and may not
be a source of resistant material in sorghum breeding.
7.4. Recommendations for further research
1. More studies are needed on the link between reduced moisture availability to abscissic
acid need to be done to gain full understanding of the combined effects of drought on Striga
infested sorghum.
2. There is need to quantify the link between mulching, moisture availability and the effects
of parasitism.
3. Striga asiatica strains from the major sorghum producing regions in Zimbabwe to be tested
on any genotypes deemed resistant.
4. More genotypes from the genebank and from farmers’ fields need to be evaluated for
Striga resistance and tolerance.

182
References
1. Adekalu, KO, Olorinfemi IA, Osunbitan JA (2007). Grass mulching effect on on
infiltration, surface runoff from two southwestern Nigerian agricultural soils.
Bioresource Technology 98: 912 - 917
2. Badu-Apraku B, Akinwale RO, Oyenkule M (2014). Efficiency of secondary traits in
selecting for improved grain yield in extra early maize under Striga infested and
Striga free environments. Plant Breeding 133: 373 - 380
3. Berner, DK, Kling JG, Singh BB (1995). Striga research and control- a perspective
from Africa. Plant Disease 79: 652 – 660.
4. Huang K, Whitlock R, Press MC, Scholes JD (2012). Variations of host range within
and among populations of the parasitic plant, Striga hermonthica. Heredity 108: 96 -
104
5. Jamil M, Rodenburg J, Charnikhova T, Bouwmeester H.J (2012) Pre-attachment
Striga hermonthica resistance of new Rice for Africa (NERICA) cultivars based on
strigolactone production. New Phytologist 192: 964 – 975.
6. Jamil M, Charnikhova T, Houshyani B, van AAst A, Bouwmeester HJ (2011).
Genetic variation in strigolactones production and tillering in rice and its effects on
Striga hermonthica infection. Planta DOI 10.1007/s00425-011-1520y.
7. Kountche BA, Hash CT, Dodo H, Laovaly O, Sanogo MD, Timbeli A, Vigouroux Y,
This D, Nokamp D, Hausmann BIG (2013). Development of pearl millet Striga
resistant gene pool, response to five cycles of recurrent selection under Striga infested
field conditions in West Africa. Field Crop Research 154: 82 - 90

183
8. Mabasa S (2003). Nitrogen and the control of Striga: timing and dose of organic and
inorganic forms. A PhD thesis submitted to the University of Sheffield, United
Kingdom.
9. Rao, K, Steenhuis, T, Cogle A, Srinivasan, S, Yule D, Smith G (1998). Rainfall
infiltration and runoff from an alfisol in semi-arid India. 1. No till systems. Soil and
Tillage Res 48: 51 – 59.
10. Rubiales. D, Verkleij. J, Vurro. M , Murdoch, A.J and Joel, D.M (2009). Parasitic
plant management in sustainable agriculture. Weed Research 49: 1 – 5.
11. Scopel E, Muller, B, Areola Tostado J, Chavez Guera E, Maraux F (1998).
Quantifying and modeling the effects of a light crop residue on the water balance: an
application to rainfed maize in western Mexico. World congress on soil science,
France.
12. Sun, Z, Hans J, Walter M.H, Matusova R, Beekwilder R, Vestapen F.W.A, Ming Z,
van Echtelt E, Strack D, Bisseling T, Boumeester H.J (2008). Cloning and
Characterisation of a maize characterisation of a maize carotenoid cleavage
dioxygenase (ZmCCDI) and its involvement in the biosynthesis of apocarotenoids
with various roles in mutualistic and parasitic interactions. Planta 228: 789 - 801

184
APPENDIX
APPENDICES
Appendix for Chapter 3: Analysis of variance tables for experiment 1 and 11
Analysis of variance
Variate: Head_DM
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 252.555 63.139 9.07 <.001
Infection 1 83.733 83.733 12.02 0.001
Drought 1 178.124 178.124 25.57 <.001
Variety.Infection 4 5.909 1.477 0.21 0.930
Variety.Drought 4 23.180 5.795 0.83 0.513
Infection.Drought 1 2.481 2.481 0.36 0.554
Variety.Infection.Drought
4 11.714 2.928 0.42 0.793
Residual 40 278.599 6.965
Total 59 836.294
Analysis of variance
Variate: Head_DM_adj
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 328.220 82.055 9.07 <.001
Infection 1 108.819 108.819 12.02 0.001
Drought 1 231.490 231.490 25.57 <.001
Variety.Infection 4 7.679 1.920 0.21 0.930
Variety.Drought 4 30.125 7.531 0.83 0.513
Infection.Drought 1 3.224 3.224 0.36 0.554
Variety.Infection.Drought
4 15.223 3.806 0.42 0.793
Residual 40 362.067 9.052
Total 59 1086.847
Analysis of variance
Variate: Root_dry
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 2850.7 712.7 2.99 0.030
Infection 1 416.0 416.0 1.74 0.194
Drought 1 2163.5 2163.5 9.07 0.004
Variety.Infection 4 408.4 102.1 0.43 0.788
Variety.Drought 4 496.7 124.2 0.52 0.721
Infection.Drought 1 147.7 147.7 0.62 0.436
Variety.Infection.Drought
4 328.4 82.1 0.34 0.847
Residual 40 9544.2 238.6
Total 59 16355.7

185
Analysis of variance
Variate: Rut_index
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.13483 0.03371 2.78 0.040
Infection 1 0.04188 0.04188 3.45 0.071
Drought 1 0.00140 0.00140 0.12 0.735
Variety.Infection 4 0.01746 0.00436 0.36 0.836
Variety.Drought 4 0.03239 0.00810 0.67 0.618
Infection.Drought 1 0.01862 0.01862 1.54 0.223
Variety.Infection.Drought
4 0.02271 0.00568 0.47 0.759
Residual 40 0.48511 0.01213
Total 59 0.75439

186
Analysis of variance for spad in experiment 2 at 10 wace
Variate: Spad_3ex
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 108.45 27.11 0.45 0.768
Infection 1 15.50 15.50 0.26 0.613
Drought 1 41.50 41.50 0.70 0.409
Variety.Infection 4 52.84 13.21 0.22 0.925
Variety.Drought 4 45.59 11.40 0.19 0.942
Infection.Drought 1 28.15 28.15 0.47 0.496
Variety.Infection.Drought
4 151.34 37.83 0.63 0.641
Residual 40 2385.09 59.63
Total 59 2828.47
Analysis of variance
Variate: Total_DM
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 4976.0 1244.0 3.38 0.018
Infection 1 103.4 103.4 0.28 0.599
Drought 1 9986.5 9986.5 27.17 <.001
Variety.Infection 4 2860.0 715.0 1.94 0.122
Variety.Drought 4 231.4 57.9 0.16 0.959
Infection.Drought 1 58.9 58.9 0.16 0.691
Variety.Infection.Drought
4 582.3 145.6 0.40 0.810
Residual 40 14704.8 367.6
Total 59 33503.4
Analysis of variance
Variate: dry_stems
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 283.43 70.86 1.39 0.254
Infection 1 5.94 5.94 0.12 0.734
Drought 1 536.29 536.29 10.53 0.002
Variety.Infection 4 687.27 171.82 3.37 0.018
Variety.Drought 4 101.22 25.31 0.50 0.738
Infection.Drought 1 0.01 0.01 0.00 0.989
Variety.Infection.Drought
4 335.88 83.97 1.65 0.181
Residual 40 2036.62 50.92
Total 59 3986.66
Analysis of variance at 6 wace in experiment 1
Variate: green_1_exp
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.010077 0.002519 0.37 0.829
Infection 1 0.029927 0.029927 4.38 0.043

187
Drought 1 0.390427 0.390427 57.18 <.001
Variety.Infection 4 0.018423 0.004606 0.67 0.614
Variety.Drought 4 0.010123 0.002531 0.37 0.828
Infection.Drought 1 0.023207 0.023207 3.40 0.073
Variety.Infection.Drought
4 0.044577 0.011144 1.63 0.185
Residual 40 0.273133 0.006828
Total 59 0.799893
Analysis of variance
Variate: green_2_exp_2
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.108593 0.027148 4.23 0.006
Infection 1 0.009882 0.009882 1.54 0.222
Drought 1 0.101682 0.101682 15.86 <.001
Variety.Infection 4 0.006060 0.001515 0.24 0.916
Variety.Drought 4 0.031327 0.007832 1.22 0.317
Infection.Drought 1 0.008402 0.008402 1.31 0.259
Variety.Infection.Drought
4 0.027407 0.006852 1.07 0.385
Residual 40 0.256467 0.006412
Total 59 0.549818
Analysis of variance
Variate: head_index
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.119577 0.029894 11.16 <.001
Infection 1 0.019735 0.019735 7.37 0.010
Drought 1 0.016000 0.016000 5.97 0.019
Variety.Infection 4 0.003059 0.000765 0.29 0.886
Variety.Drought 4 0.002850 0.000713 0.27 0.898
Infection.Drought 1 0.000630 0.000630 0.24 0.630
Variety.Infection.Drought
4 0.002811 0.000703 0.26 0.900
Residual 40 0.107182 0.002680
Total 59 0.271845
Analysis of variance
Variate: leaf_index
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.019685 0.004921 1.45 0.235
Infection 1 0.000386 0.000386 0.11 0.738
Drought 1 0.002152 0.002152 0.63 0.430
Variety.Infection 4 0.015046 0.003761 1.11 0.366
Variety.Drought 4 0.007571 0.001893 0.56 0.694
Infection.Drought 1 0.004273 0.004273 1.26 0.268
Variety.Infection.Drought
4 0.000867 0.000217 0.06 0.992
Residual 40 0.135712 0.003393

188
Total 59 0.185692
Analysis of variance
Variate: leaves_DM
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 338.93 84.73 4.98 0.002
Infection 1 6.98 6.98 0.41 0.525
Drought 1 226.40 226.40 13.30 <.001
Variety.Infection 4 160.65 40.16 2.36 0.070
Variety.Drought 4 4.32 1.08 0.06 0.992
Infection.Drought 1 6.66 6.66 0.39 0.535
Variety.Infection.Drought
4 28.12 7.03 0.41 0.798
Residual 40 680.87 17.02
Total 59 1452.93
Analysis of variance for experiment 1 at 6 wace
Variate: spad1_Exp2
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 641.20 160.30 5.03 0.002
Infection 1 141.53 141.53 4.44 0.041
Drought 1 440.38 440.38 13.81 <.001
Variety.Infection 4 74.74 18.68 0.59 0.675
Variety.Drought 4 351.77 87.94 2.76 0.041
Infection.Drought 1 23.60 23.60 0.74 0.395
Variety.Infection.Drought
4 105.80 26.45 0.83 0.514
Residual 40 1275.09 31.88
Total 59 3054.11
Analysis of variance
Variate: stem_index
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.033732 0.008433 1.38 0.257
Infection 1 0.004114 0.004114 0.68 0.416
Drought 1 0.003641 0.003641 0.60 0.444
Variety.Infection 4 0.024635 0.006159 1.01 0.413
Variety.Drought 4 0.029982 0.007496 1.23 0.313
Infection.Drought 1 0.001804 0.001804 0.30 0.589
Variety.Infection.Drought
4 0.020288 0.005072 0.83 0.513
Residual 40 0.243676 0.006092
Total 59 0.361872

189
Analysis of variance
Variate: Height_1
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 5431.7 1357.9 11.83 <.001
Infection 1 268.8 268.8 2.34 0.134
Drought 1 163.4 163.4 1.42 0.240
Variety.Infection 4 97.6 24.4 0.21 0.930
Variety.Drought 4 403.4 100.9 0.88 0.485
Infection.Drought 1 421.3 421.3 3.67 0.063
Variety.Infection.Drought
4 72.4 18.1 0.16 0.958
Residual 40 4590.0 114.8
Total 59 11448.6
Analysis of variance
Variate: Height_2
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 3903.6 975.9 6.60 <.001
Infection 1 777.6 777.6 5.26 0.027
Drought 1 614.4 614.4 4.16 0.048
Variety.Infection 4 116.2 29.1 0.20 0.939
Variety.Drought 4 759.1 189.8 1.28 0.293
Infection.Drought 1 77.1 77.1 0.52 0.474
Variety.Infection.Drought
4 352.4 88.1 0.60 0.668
Residual 40 5913.3 147.8
Total 59 12513.7
Analysis of variance
Variate: Height_3
Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr.
Variety 4 1872.3 468.1 2.15 0.093
Infection 1 908.7 908.7 4.17 0.048
Drought 1 1096.5 1096.5 5.03 0.031
Variety.Infection 4 911.3 227.8 1.05 0.396
Variety.Drought 4 1583.0 395.7 1.82 0.145
Infection.Drought 1 139.5 139.5 0.64 0.428
Variety.Infection.Drought
4 890.6 222.7 1.02 0.408
Residual 39 (1) 8496.5 217.9
Total 58 (1) 15753.5

190
Analysis of variance
Variate: Tillers
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 46.567 11.642 5.87 <.001
Infection 1 5.400 5.400 2.72 0.107
Drought 1 0.067 0.067 0.03 0.855
Variety.Infection 4 11.433 2.858 1.44 0.238
Variety.Drought 4 2.433 0.608 0.31 0.872
Infection.Drought 1 0.267 0.267 0.13 0.716
Variety.Infection.Drought
4 10.900 2.725 1.37 0.260
Residual 40 79.333 1.983
Total 59 156.400
Analysis of variance
Variate: Greeness2
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.090640 0.022660 3.19 0.023
Infection 1 0.003375 0.003375 0.47 0.495
Drought 1 0.001042 0.001042 0.15 0.704
Variety.Infection 4 0.005300 0.001325 0.19 0.944
Variety.Drought 4 0.023133 0.005783 0.81 0.524
Infection.Drought 1 0.004002 0.004002 0.56 0.457
Variety.Infection.Drought
4 0.049140 0.012285 1.73 0.163
Residual 40 0.284333 0.007108
Total 59 0.460965
Analysis of variance
Variate: Spad_1
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 523.73 130.93 4.73 0.003
Infection 1 11.97 11.97 0.43 0.515
Drought 1 17.93 17.93 0.65 0.426
Variety.Infection 4 66.47 16.62 0.60 0.665
Variety.Drought 4 182.16 45.54 1.64 0.182
Infection.Drought 1 10.09 10.09 0.36 0.550
Variety.Infection.Drought
4 36.34 9.08 0.33 0.857
Residual 40 1107.59 27.69
Total 59 1956.27
Analysis of variance
Variate: Spad_2

191
Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr.
Variety 4 734.04 183.51 7.29 <.001
Infection 1 0.23 0.23 0.01 0.924
Drought 1 10.46 10.46 0.42 0.523
Variety.Infection 4 10.32 2.58 0.10 0.981
Variety.Drought 4 133.83 33.46 1.33 0.276
Infection.Drought 1 25.94 25.94 1.03 0.316
Variety.Infection.Drought
4 32.42 8.10 0.32 0.861
Residual 39 (1) 981.38 25.16
Total 58 (1) 1922.27
Analysis of variance for experiment 11 at 6wace
Variate: greeness
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.072640 0.018160 2.07 0.103
Infection 1 0.005042 0.005042 0.57 0.453
Drought 1 0.066002 0.066002 7.51 0.009
Variety.Infection 4 0.042333 0.010583 1.20 0.324
Variety.Drought 4 0.019673 0.004918 0.56 0.693
Infection.Drought 1 0.009882 0.009882 1.12 0.295
Variety.Infection.Drought
4 0.041060 0.010265 1.17 0.339
Residual 40 0.351533 0.008788
Total 59 0.608165

192
Analysis of variance at 10 WACE in experiment 11
Variate: greeness_3
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.03739 0.00935 0.88 0.486
Infection 1 0.09048 0.09048 8.49 0.006
Drought 1 0.06080 0.06080 5.71 0.022
Variety.Infection 4 0.05143 0.01286 1.21 0.323
Variety.Drought 4 0.10554 0.02639 2.48 0.060
Infection.Drought 1 0.00160 0.00160 0.15 0.700
Variety.Infection.Drought
4 0.01591 0.00398 0.37 0.826
Residual 40 0.42627 0.01066
Total 59 0.78942
Analysis of variance
Variate: spad_3
Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr.
Variety 4 346.18 86.54 1.05 0.395
Infection 1 12.97 12.97 0.16 0.694
Drought 1 843.00 843.00 10.22 0.003
Variety.Infection 4 121.81 30.45 0.37 0.829
Variety.Drought 4 203.81 50.95 0.62 0.653
Infection.Drought 1 36.97 36.97 0.45 0.507
Variety.Infection.Drought
4 246.88 61.72 0.75 0.565
Residual 39 (1) 3217.59 82.50
Total 58 (1) 4845.74
Analysis of variance
Variate: Head_index
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.177163 0.044291 10.37 <.001
Infection 1 0.035755 0.035755 8.37 0.006
Drought 1 0.009396 0.009396 2.20 0.146
Variety.Infection 4 0.014336 0.003584 0.84 0.509
Variety.Drought 4 0.018068 0.004517 1.06 0.390
Infection.Drought 1 0.000334 0.000334 0.08 0.781
Variety.Infection.Drought
4 0.008320 0.002080 0.49 0.745
Residual 40 0.170891 0.004272
Total 59 0.434264
Analysis of variance
Variate: Head_wei
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 404.89 101.22 9.92 <.001

193
Infection 1 90.28 90.28 8.85 0.005
Drought 1 116.82 116.82 11.45 0.002
Variety.Infection 4 32.75 8.19 0.80 0.531
Variety.Drought 4 111.80 27.95 2.74 0.042
Infection.Drought 1 4.98 4.98 0.49 0.489
Variety.Infection.Drought
4 15.97 3.99 0.39 0.814
Residual 40 408.14 10.20
Total 59 1185.64
Analysis of variance
Variate: Internode
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 64.077 16.019 1.66 0.178
Infection 1 75.264 75.264 7.80 0.008
Drought 1 14.017 14.017 1.45 0.235
Variety.Infection 4 37.273 9.318 0.97 0.437
Variety.Drought 4 33.353 8.338 0.86 0.494
Infection.Drought 1 97.283 97.283 10.09 0.003
Variety.Infection.Drought
4 14.067 3.517 0.36 0.832
Residual 40 385.813 9.645
Total 59 721.147
Analysis of variance
Variate: Leaf_index
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.007335 0.001834 0.39 0.812
Infection 1 0.000971 0.000971 0.21 0.650
Drought 1 0.002985 0.002985 0.64 0.428
Variety.Infection 4 0.003511 0.000878 0.19 0.943
Variety.Drought 4 0.003462 0.000866 0.19 0.944
Infection.Drought 1 0.026219 0.026219 5.64 0.022
Variety.Infection.Drought
4 0.004498 0.001124 0.24 0.913
Residual 40 0.186057 0.004651
Total 59 0.235038
Analysis of variance
Variate: LeavesDM
Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr.
Variety 4 421.57 105.39 5.71 0.001
Infection 1 9.91 9.91 0.54 0.468
Drought 1 151.02 151.02 8.18 0.007
Variety.Infection 4 180.02 45.00 2.44 0.063
Variety.Drought 4 13.98 3.50 0.19 0.943
Infection.Drought 1 8.19 8.19 0.44 0.509
Variety.Infection.Drought
4 68.25 17.06 0.92 0.460

194
Residual 39 (1) 719.91 18.46
Total 58 (1) 1566.35
Analysis of variance
Variate: Root_dry
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 5019.6 1254.9 4.08 0.007
Infection 1 1567.0 1567.0 5.10 0.029
Drought 1 2840.2 2840.2 9.25 0.004
Variety.Infection 4 535.5 133.9 0.44 0.782
Variety.Drought 4 615.4 153.9 0.50 0.735
Infection.Drought 1 890.5 890.5 2.90 0.096
Variety.Infection.Drought
4 265.4 66.3 0.22 0.928
Residual 40 12288.3 307.2
Total 59 24022.0
Analysis of variance
Variate: Root_index
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.29315 0.07329 5.75 <.001
Infection 1 0.05435 0.05435 4.26 0.045
Drought 1 0.00155 0.00155 0.12 0.729
Variety.Infection 4 0.01544 0.00386 0.30 0.874
Variety.Drought 4 0.01643 0.00411 0.32 0.861
Infection.Drought 1 0.04829 0.04829 3.79 0.059
Variety.Infection.Drought
4 0.02045 0.00511 0.40 0.807
Residual 40 0.50983 0.01275
Total 59 0.95949
Analysis of variance
Variate: Stem_index
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 0.039043 0.009761 1.60 0.193
Infection 1 0.005657 0.005657 0.93 0.341
Drought 1 0.006670 0.006670 1.09 0.302
Variety.Infection 4 0.039305 0.009826 1.61 0.190
Variety.Drought 4 0.029976 0.007494 1.23 0.314
Infection.Drought 1 0.005795 0.005795 0.95 0.335
Variety.Infection.Drought
4 0.012942 0.003235 0.53 0.714
Residual 40 0.243833 0.006096
Total 59 0.383221
Analysis of variance
Variate: Total_DM

195
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 7122.8 1780.7 3.50 0.015
Infection 1 1287.7 1287.7 2.53 0.119
Drought 1 9983.1 9983.1 19.62 <.001
Variety.Infection 4 3449.9 862.5 1.70 0.170
Variety.Drought 4 412.6 103.2 0.20 0.935
Infection.Drought 1 942.5 942.5 1.85 0.181
Variety.Infection.Drought
4 1254.5 313.6 0.62 0.653
Residual 40 20350.3 508.8
Total 59 44803.3
Analysis of variance
Variate: dry_stems
Source of variation d.f. s.s. m.s. v.r. F pr.
Variety 4 357.61 89.40 1.45 0.234
Infection 1 0.24 0.24 0.00 0.951
Drought 1 456.34 456.34 7.42 0.010
Variety.Infection 4 850.16 212.54 3.46 0.016
Variety.Drought 4 76.22 19.05 0.31 0.870
Infection.Drought 1 13.34 13.34 0.22 0.644
Variety.Infection.Drought
4 427.87 106.97 1.74 0.160
Residual 40 2460.59 61.51
Total 59 4642.37
APPENDIX 2: Analysis of variance tables for Chapter 4
Analysis of variance
Variate: Chlorophy_I_3 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 1862.54 206.95 5.30 <.001 Mulch 1 0.16 0.16 0.00 0.949 infestati 1 22.06 22.06 0.57 0.454 Variety.Mulch 9 461.28 51.25 1.31 0.244 Variety.infestati 9 440.13 48.90 1.25 0.276 Mulch.infestati 1 109.92 109.92 2.82 0.097 Variety.Mulch.infestati 9 417.35 46.37 1.19 0.315 Residual 79 (1) 3084.37 39.04

196
Total 118 (1) 6314.94
Analysis of variance for chlorophyll content at 6 WACE in 2013-14 season
Variate: Chlorophyl1 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 343.60 38.18 1.81 0.080 Mulch 1 6.42 6.42 0.30 0.583 infestati 1 46.94 46.94 2.23 0.140 Variety.Mulch 9 201.91 22.43 1.06 0.399 Variety.infestati 9 375.21 41.69 1.98 0.053 Mulch.infestati 1 1.11 1.11 0.05 0.819 Variety.Mulch.infestati 9 172.52 19.17 0.91 0.522 Residual 77 (3) 1623.73 21.09 Total 116 (3) 2740.53
Analysis of variance for chlorophyll content at 8 WACE in the 2013 – 14 season
Variate: Chlorophyl_2 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 488.52 54.28 3.60 <.001 Mulch 1 42.96 42.96 2.85 0.096 infestati 1 3.47 3.47 0.23 0.633 Variety.Mulch 9 108.70 12.08 0.80 0.617 Variety.infestati 9 166.80 18.53 1.23 0.290 Mulch.infestati 1 16.88 16.88 1.12 0.294 Variety.Mulch.infestati 9 130.23 14.47 0.96 0.481 Residual 79 (1) 1192.75 15.10 Total 118 (1) 2145.72
Analysis of variance
Variate: Chlorophyl_4 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 856.97 95.22 1.32 0.242 Mulch 1 51.16 51.16 0.71 0.403 infestati 1 0.16 0.16 0.00 0.963 Variety.Mulch 9 1560.43 173.38 2.40 0.019 Variety.infestati 9 734.54 81.62 1.13 0.353 Mulch.infestati 1 36.14 36.14 0.50 0.482 Variety.Mulch.infestati 9 1033.34 114.82 1.59 0.134 Residual 73 (7) 5270.21 72.19 Total 112 (7) 8981.34
Analysis of variance
Variate: Chlorophyl_5

197
Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 842.61 93.62 1.04 0.420 Mulch 1 375.77 375.77 4.16 0.045 infestati 1 81.43 81.43 0.90 0.346 Variety.Mulch 9 660.95 73.44 0.81 0.606 Variety.infestati 9 881.51 97.95 1.08 0.385 Mulch.infestati 1 145.31 145.31 1.61 0.209 Variety.Mulch.infestati 9 984.42 109.38 1.21 0.302 Residual 72 (8) 6505.40 90.35 Total 111 (8) 10367.63
Analysis of variance for stomatal conductance in the 2013 – 14 season
Variate: Conductance_1 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 6839.8 760.0 3.73 <.001 Mulch 1 0.3 0.3 0.00 0.969 infestati 1 793.9 793.9 3.89 0.052 Variety.Mulch 9 1525.5 169.5 0.83 0.590 Variety.infestati 9 2044.1 227.1 1.11 0.364 Mulch.infestati 1 57.8 57.8 0.28 0.596 Variety.Mulch.infestati 9 2063.8 229.3 1.12 0.357 Residual 74 (6) 15095.3 204.0 Total 113 (6) 26690.3
Analysis of variance
Variate: Height_1 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 2586.89 287.43 5.17 <.001 Mulch 1 12.50 12.50 0.22 0.637 infestati 1 191.83 191.83 3.45 0.067 Variety.Mulch 9 770.53 85.61 1.54 0.150 Variety.infestati 9 951.86 105.76 1.90 0.065 Mulch.infestati 1 39.99 39.99 0.72 0.399 Variety.Mulch.infestati 9 540.70 60.08 1.08 0.388 Residual 73 (7) 4059.35 55.61 Total 112 (7) 8943.01
Analysis of variance
Variate: Height_2 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 4060.7 451.2 3.78 <.001 Mulch 1 0.5 0.5 0.00 0.948

198
infestati 1 1658.9 1658.9 13.89 <.001 Variety.Mulch 9 1780.5 197.8 1.66 0.116 Variety.infestati 9 1270.9 141.2 1.18 0.320 Mulch.infestati 1 69.3 69.3 0.58 0.449 Variety.Mulch.infestati 9 605.0 67.2 0.56 0.823 Residual 70 (10) 8360.9 119.4 Total 109 (10) 15730.4
Analysis of variance
Variate: Height_4 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 69831. 7759. 5.92 <.001 Mulch 1 241. 241. 0.18 0.670 infestati 1 23801. 23801. 18.15 <.001 Variety.Mulch 9 7446. 827. 0.63 0.767 Variety.infestati 9 16268. 1808. 1.38 0.214 Mulch.infestati 1 1665. 1665. 1.27 0.264 Variety.Mulch.infestati 9 4834. 537. 0.41 0.926 Residual 71 (9) 93108. 1311. Total 110 (9) 200388.
Analysis of variance
Variate: Leaf_length Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 14022.9 1558.1 15.37 <.001 Mulch 1 7.8 7.8 0.08 0.783 infestati 1 55.4 55.4 0.55 0.462 Variety.Mulch 9 1109.3 123.3 1.22 0.299 Variety.infestati 9 734.0 81.6 0.80 0.614 Mulch.infestati 1 0.1 0.1 0.00 0.975 Variety.Mulch.infestati 9 407.5 45.3 0.45 0.905 Residual 71 (9) 7198.8 101.4 Total 110 (9) 23007.7
Analysis of variance for effect of sorghum genotypes on tiller number in the 2013 – 14 season
Variate: Tiller_1 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 106.135 11.793 6.33 <.001 Mulch 1 5.852 5.852 3.14 0.080 infestati 1 13.002 13.002 6.98 0.010 Variety.Mulch 9 20.335 2.259 1.21 0.299 Variety.infestati 9 29.185 3.243 1.74 0.093 Mulch.infestati 1 1.302 1.302 0.70 0.406

199
Variety.Mulch.infestati 9 31.052 3.450 1.85 0.072 Residual 79 (1) 147.167 1.863 Total 118 (1) 353.933
Analysis of variance
Variate: height_3 Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. Variety 9 17558.5 1950.9 3.38 0.002 Mulch 1 218.7 218.7 0.38 0.540 infestati 1 6453.3 6453.3 11.17 0.001 Variety.Mulch 9 4393.1 488.1 0.84 0.578 Variety.infestati 9 2637.4 293.0 0.51 0.865 Mulch.infestati 1 224.1 224.1 0.39 0.535 Variety.Mulch.infestati 9 1299.2 144.4 0.25 0.985 Residual 72 (8) 41597.0 577.7 Total 111 (8) 70805.4
Analysis of variance
Variate: leaves_1 Source of variation d.f. s.s. m.s. v.r. F pr. Variety 9 34.000 3.778 1.76 0.090 Mulch 1 1.200 1.200 0.56 0.457 infestati 1 8.533 8.533 3.97 0.050 Variety.Mulch 9 28.967 3.219 1.50 0.163 Variety.infestati 9 37.300 4.144 1.93 0.060 Mulch.infestati 1 0.833 0.833 0.39 0.535 Variety.Mulch.infestati 9 18.333 2.037 0.95 0.489 Residual 80 172.000 2.150 Total 119 301.167
Analysis of variance for head weight in the 2014 – 15 season
Variate: Headweight_15 Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 8424.1 936.0 7.95 <.001 Infestation 1 1780.9 1780.9 15.13 <.001 Mulch 1 4179.7 4179.7 35.50 <.001 variety.Infestation 9 1503.0 167.0 1.42 0.194 variety.Mulch 9 834.7 92.7 0.79 0.628 Infestation.Mulch 1 36.4 36.4 0.31 0.580 variety.Infestation.Mulch 9 554.2 61.6 0.52 0.854 Residual 80 9419.6 117.7 Total 119 26732.5

200
Appendix 3: Analysis of variance table for Chapter 5 experiment 1 and 11
Analysis of variance for head weight at Henderson research station
Variate: HEAD_WEI_g Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 6378.43 708.71 14.86 <.001 Strain 2 1098.93 549.46 11.52 <.001 variety.Strain 18 1541.34 85.63 1.79 0.047 Residual 60 2862.33 47.71 Total 89 11881.03
Analysis of variance
Variate: Head_weight_g Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 7776.93 864.10 16.87 <.001 Strain 2 596.40 298.20 5.82 0.005 variety.Strain 18 2058.11 114.34 2.23 0.011 Residual 60 3074.01 51.23 Total 89 13505.45
Analysis of variance
Variate: LEAF_WEI_g Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 527.380 58.598 7.64 <.001 Strain 2 1.550 0.775 0.10 0.904 variety.Strain 18 216.699 12.039 1.57 0.098 Residual 60 460.220 7.670 Total 89 1205.849
Analysis of variance
Variate: L_LENGTH Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 418.23 46.47 1.36 0.226 Strain 2 16.07 8.03 0.24 0.791 variety.Strain 18 965.93 53.66 1.57 0.097 Residual 60 2046.67 34.11 Total 89 3446.90
Analysis of variance
Variate: Leaf_weight_g

201
Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 1425.37 158.37 9.02 <.001 Strain 2 4.31 2.15 0.12 0.885 variety.Strain 18 456.57 25.36 1.44 0.145 Residual 60 1053.60 17.56 Total 89 2939.85
Analysis of variance for sorghum height at Buse at 12 WACE
Variate: P_HEIGHT Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 53351. 5928. 4.30 <.001 Strain 2 10432. 5216. 3.79 0.028 variety.Strain 18 35048. 1947. 1.41 0.159 Residual 60 82631. 1377. Total 89 181463.
Analysis of variance for root weight at BUSE
Variate: Root_weight Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 36290.6 4032.3 5.56 <.001 Strain 2 16331.9 8165.9 11.26 <.001 variety.Strain 18 17350.5 963.9 1.33 0.204 Residual 60 43531.7 725.5 Total 89 113504.6
Analysis of variance for root index at BUSE
Variate: Rutindex Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 1.29520 0.14391 13.68 <.001 Strain 2 0.57482 0.28741 27.32 <.001 variety.Strain 18 0.25376 0.01410 1.34 0.197 Residual 60 0.63115 0.01052 Total 89 2.75493
Analysis of variance
Variate: STEM_WEI_g Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 3080.2 342.2 3.20 0.003 Strain 2 1268.5 634.3 5.93 0.004 variety.Strain 18 3796.0 210.9 1.97 0.026 Residual 60 6413.5 106.9 Total 89 14558.2
Analysis of variance

202
Variate: S_COUNT Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 474.27 52.70 1.19 0.319 Strain 2 424.09 212.04 4.78 0.012 variety.Strain 18 1021.47 56.75 1.28 0.234 Residual 60 2660.00 44.33 Total 89 4579.82
Analysis of variance for stem index at Buse
Variate: Stem_Inde Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 0.188103 0.020900 3.46 0.002 Strain 2 0.133236 0.066618 11.03 <.001 variety.Strain 18 0.171405 0.009522 1.58 0.096 Residual 60 0.362323 0.006039 Total 89 0.855066
Analysis of variance
Variate: Stem_weight_g Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 48728. 5414. 5.35 <.001 Strain 2 8141. 4070. 4.03 0.023 variety.Strain 18 28045. 1558. 1.54 0.108 Residual 60 60675. 1011. Total 89 145589.
Analysis of variance
Variate: Striga_Counts Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 792.90 88.10 1.69 0.111 Strain 2 687.09 343.54 6.59 0.003 variety.Strain 18 809.80 44.99 0.86 0.622 Residual 60 3126.67 52.11 Total 89 5416.46

203
Analysis of variance
Variate: TILLER_No Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 440.267 48.919 5.35 <.001 Strain 2 43.889 21.944 2.40 0.099 variety.Strain 18 103.667 5.759 0.63 0.862 Residual 60 548.667 9.144 Total 89 1136.489
Analysis of variance
Variate: Total Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 28960.4 3217.8 3.32 0.002 Strain 2 8709.5 4354.8 4.49 0.015 variety.Strain 18 27440.5 1524.5 1.57 0.097 Residual 60 58132.2 968.9 Total 89 123242.6
Analysis of variance
Variate: chloro_1 Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 1112.223 123.580 12.41 <.001 Strain 2 532.241 266.120 26.71 <.001 variety.Strain 18 712.641 39.591 3.97 <.001 Residual 60 597.693 9.962 Total 89 2954.798
Analysis of variance for chlorophyll content at 8 wace at BUSE
Variate: chloro_2 Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 602.738 66.971 7.26 <.001 Strain 2 186.027 93.013 10.08 <.001 variety.Strain 18 454.389 25.244 2.74 0.002 Residual 60 553.773 9.230 Total 89 1796.927
Analysis of variance for chlorophyll content at 12 wace at BUSE
Variate: chloro_3 Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 917.362 101.929 12.11 <.001 Strain 2 171.811 85.905 10.21 <.001 variety.Strain 18 412.843 22.936 2.73 0.002 Residual 60 504.973 8.416

204
Total 89 2006.989
Analysis of variance
Source d.f. s.s. m.s. v.r. F pr. variety ignoring Strain 9 359.48 39.94 1.17 0.333 variety eliminating Strain 9 369.94 41.10 1.20 0.312 Strain ignoring variety 2 194.15 97.07 2.84 0.067 Strain eliminating variety 2 204.60 102.30 2.99 0.058 variety.Strain 18 609.21 33.84 0.99 0.485 Residual 58 1984.81 34.22 Total 87 3158.10 36.30
Analysis of variance for head index at BUSE
Variate: headindex Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 0.812826 0.090314 14.78 <.001 Strain 2 0.135071 0.067535 11.05 <.001 variety.Strain 18 0.141500 0.007861 1.29 0.229 Residual 60 0.366666 0.006111 Total 89 1.456063
Analysis of variance for plant height at 6 wace at Buse
Source d.f. s.s. m.s. v.r. F pr. variety ignoring Strain 9 1874.00 208.22 6.99 < 0.001 variety eliminating Strain 9 1907.52 211.95 7.11 < 0.001 Strain ignoring variety 2 112.02 56.01 1.88 0.162 Strain eliminating variety 2 145.54 72.77 2.44 0.096 variety.Strain 18 512.05 28.45 0.95 0.522 Residual 56 1669.17 29.81 Total 85 4200.76 49.42
Analysis of variance for leaf index at BUSE
Variate: lifindex Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 0.0361770 0.0040197 8.15 <.001 Strain 2 0.0026811 0.0013405 2.72 0.074 variety.Strain 18 0.0109778 0.0006099 1.24 0.264 Residual 60 0.0296068 0.0004934 Total 89 0.0794427

205
Analysis of variance
Variate: tiller_num Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 71.833 7.981 6.41 <.001 Strain 2 10.422 5.211 4.19 0.020 variety.Strain 18 15.800 0.878 0.71 0.792 Residual 60 74.667 1.244 Total 89 172.722
Analysis of variance
Variate: HEAD_WEI_g Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. VARIETY 9 8979.0 997.7 9.65 <.001 STRI_STR 2 1055.4 527.7 5.10 0.009 VARIETY.STRI_STR 18 2633.8 146.3 1.41 0.161 Residual 56 (4) 5791.6 103.4 Total 85 (4) 18103.9
Analysis of variance
Variate: LEAF_WEI_g Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. VARIETY 9 523.919 58.213 7.09 <.001 STRI_STR 2 1.806 0.903 0.11 0.896 VARIETY.STRI_STR 18 218.240 12.124 1.48 0.134 Residual 56 (4) 459.493 8.205 Total 85 (4) 1184.259
Analysis of variance
Variate: STEM_WEI_g Source of variation d.f. (m.v.) s.s. m.s. v.r. F pr. VARIETY 9 3432.8 381.4 2.78 0.009 STRI_STR 2 411.5 205.7 1.50 0.231 VARIETY.STRI_STR 18 3839.5 213.3 1.56 0.103 Residual 59 (1) 8086.3 137.1 Total 88 (1) 15765.2
Analysis of variance
Variate: Head_index Source of variation d.f. s.s. m.s. v.r. F pr. VARIETY 9 0.404695 0.044966 18.37 <.001 STRI_STR 2 0.036723 0.018362 7.50 0.001

206
VARIETY.STRI_STR 18 0.062302 0.003461 1.41 0.159 Residual 60 0.146895 0.002448 Total 89 0.650616
Analysis of variance
Variate: Head_weight_g Source of variation d.f. s.s. m.s. v.r. F pr. VARIETY 9 6677.58 741.95 16.79 <.001 STRI_STR 2 1329.86 664.93 15.05 <.001 VARIETY.STRI_STR 18 1631.83 90.66 2.05 0.020 Residual 60 2651.25 44.19 Total 89 12290.51
Analysis of variance for leaf index for the Henderson experiment
Variate: Leaf_index Source of variation d.f. s.s. m.s. v.r. F pr. VARIETY 9 0.0332699 0.0036967 10.10 <.001 STRI_STR 2 0.0092419 0.0046209 12.63 <.001 VARIETY.STRI_STR 18 0.0128970 0.0007165 1.96 0.027 Residual 60 0.0219552 0.0003659 Total 89 0.0773640
Analysis of variance
Variate: Leaf_weight_g Source of variation d.f. s.s. m.s. v.r. F pr. VARIETY 9 1444.88 160.54 12.59 <.001 STRI_STR 2 23.77 11.89 0.93 0.399 VARIETY.STRI_STR 18 530.42 29.47 2.31 0.008 Residual 60 764.80 12.75 Total 89 2763.87
Analysis of variance for Root index at henderson
Variate: Root_Index Source of variation d.f. s.s. m.s. v.r. F pr. VARIETY 9 0.92416 0.10268 9.22 <.001 STRI_STR 2 0.40930 0.20465 18.37 <.001 VARIETY.STRI_STR 18 0.24271 0.01348 1.21 0.283 Residual 60 0.66855 0.01114 Total 89 2.24472
Analysis of variance
Variate: Root_weight

207
Source of variation d.f. s.s. m.s. v.r. F pr. VARIETY 9 36290.6 4032.3 5.56 <.001 STRI_STR 2 16331.9 8165.9 11.26 <.001 VARIETY.STRI_STR 18 17350.5 963.9 1.33 0.204 Residual 60 43531.7 725.5 Total 89 113504.6
Analysis of variance for stem index for the Henderson experiment.
Variate: Stem_index Source of variation d.f. s.s. m.s. v.r. F pr. VARIETY 9 0.63802 0.07089 5.52 <.001 STRI_STR 2 0.23920 0.11960 9.32 <.001 VARIETY.STRI_STR 18 0.30578 0.01699 1.32 0.206 Residual 60 0.77000 0.01283 Total 89 1.95300
Analysis of variance
Variate: Stem_weight_g Source of variation d.f. s.s. m.s. v.r. F pr. VARIETY 9 48626. 5403. 5.34 <.001 STRI_STR 2 8114. 4057. 4.01 0.023 VARIETY.STRI_STR 18 28040. 1558. 1.54 0.108 Residual 60 60677. 1011. Total 89 145458.
Analysis of variance for total dry weight at henderson
Variate: Total_Dry Source of variation d.f. s.s. m.s. v.r. F pr. VARIETY 9 86549. 9617. 4.70 <.001 STRI_STR 2 10423. 5211. 2.54 0.087 VARIETY.STRI_STR 18 68784. 3821. 1.87 0.037 Residual 60 122889. 2048. Total 89 288645.
Analysis of variance for chlorophyll content at 8 WACE at Henderson
Variate: chloro_1 Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 1112.223 123.580 12.41 <.001 Strain 2 532.241 266.120 26.71 <.001 variety.Strain 18 712.641 39.591 3.97 <.001 Residual 60 597.693 9.962

208
Total 89 2954.798
Analysis of variance FOR CHLOROPHYLL CONTENT AT buse AT 10 WACE
Variate: chloro_2 Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 602.738 66.971 7.26 <.001 Strain 2 186.027 93.013 10.08 <.001 variety.Strain 18 454.389 25.244 2.74 0.002 Residual 60 553.773 9.230 Total 89 1796.927
Analysis of variance
Variate: chloro_3 Source of variation d.f. s.s. m.s. v.r. F pr. variety 9 917.362 101.929 12.11 <.001 Strain 2 171.811 85.905 10.21 <.001 variety.Strain 18 412.843 22.936 2.73 0.002 Residual 60 504.973 8.416 Total 89 2006.989

209
Appendix 6: ANOVA tables for Chapter 6
Analysis of variance for tiller number for quantification trial Variate: Tiller Source of variation d.f. s.s. m.s. v.r. F pr. treatment 7 24.0000 3.4286 4.70 0.002 Residual 24 17.5000 0.7292 Total 31 41.5000
Analysis of variance for maximum germination distance
Variate: germ_dist Source of variation d.f. s.s. m.s. v.r. F pr. treatment 7 7.40219 1.05746 14.16 <.001 Residual 24 1.79250 0.07469 Total 31 9.19469
Analysis of variance for Striga counts in the quantification experiments
Variate: stri_counts Source of variation d.f. s.s. m.s. v.r. F pr. treatment 7 492.969 70.424 19.26 <.001 Residual 24 87.750 3.656 Total 31 580.719
Correlations

210
germ distance tiller striga counts
Kendall's tau_b
germ distance
Correlation Coefficient 1.000 -.191 .564**
Sig. (2-tailed) . .172 .000
N 32 32 32
tiller
Correlation Coefficient -.191 1.000 .090
Sig. (2-tailed) .172 . .518
N 32 32 32
striga counts
Correlation Coefficient .564** .090 1.000
Sig. (2-tailed) .000 .518 .
N 32 32 32
Spearman's rho
germ distance
Correlation Coefficient 1.000 -.258 .741**
Sig. (2-tailed) . .154 .000
N 32 32 32
tiller
Correlation Coefficient -.258 1.000 .062
Sig. (2-tailed) .154 . .735
N 32 32 32
striga counts
Correlation Coefficient .741** .062 1.000
Sig. (2-tailed) .000 .735 .
N 32 32 32
**. Correlation is significant at the 0.01 level (2-tailed).

211




















