
Measures of structural complexity in digital images for monitoring the ecological signature of an...
15
Measures of structural complexity in digital images for monitoring the ecological signature of an old-growth forest ecosystem Raphae ¨l Proulx, Lael Parrott * Complex Systems Laboratory, Department of Geography, University of Montreal, C.P. 6128 succursale Centre-ville, Montreal, Que., Canada H3C 3J7 1. Introduction It is well established that although mechanisms driving ecological dynamics are not well understood at the commu- nity (intermediate) level, ecosystem theory is nonetheless supported by robust principles occurring at either larger (e.g., landscape scaling laws; Lawton, 1999; Gaston, 2000) or smaller (e.g., individual based allometric laws; Turchin, 2001; Marquet et al., 2005; West and Brown, 2005) levels of taxonomic resolution (see also Simberloff, 2004). Notwithstanding this fact, ecologists are now faced with the additional challenge of uncovering organizing principles governing ecological dynamics across multiple spatial and temporal scales (Sole ´ et al., 1999; Brown et al., 2002; Storch and Gaston, 2004). In this context, organizing principles in natural systems may be regarded as an epiphenomenon of heuristic ecological goal functions (Wilhelm and Bru ¨ ggeman, 2000) which encom- pass the formation of higher-level structures emerging from lower-level interactions (Margalef, 1963; Christensen, 1995; Ulanowicz, 2004; Mu ¨ ller, 2005). In the last century the search for goal functions, also more properly termed ecological orientors (EO) in Mu ¨ ller and Leupelt (1998), has spread so ecological indicators 8 (2008) 270–284 article info Article history: Received 28 September 2006 Received in revised form 14 February 2007 Accepted 19 February 2007 Keywords: Ecological orientor Structural complexity Imagery Forest Spatiotemporal dynamics Shannon entropy Mean information gain abstract Conducting field samples for monitoring ecological dynamics across multiple spatiotem- poral scales is a difficult task using standard protocols. One alternative is to measure a restricted set of variables which can serve as an ecological orientor (EO) for quantifying habitat change. The objective of this article is to derive from digital images a measure of structural complexity that may serve as a proximate EO for monitoring forest dynamics in space and time. The mean information gain (MIG) index was used as a measure of structural complexity in photographs taken directly in the field over the entire growing season. At a small scene extent, the complexity of light intensity variations in digital images was positively related to species richness. At larger scene extents, forest understorey and overstorey layers showed predictable ecological signatures in structural complexity. In general, intensity and chroma were the two color space components which yielded the greatest sensitivity to habitat change through time. Within the framework of a standardized photographic protocol, it seems therefore reasonable to consider MIG as a suitable EO for monitoring forest dynamics in both space and time. Our results support the idea that it is possible on one hand to adopt a more holistic view of ecological processes to gain, on the other hand, spatial and temporal degrees of freedom for testing multiple scale hypotheses in the field. # 2007 Elsevier Ltd. All rights reserved. * Corresponding author. Tel.: +1 514 343 8064; fax: +1 514 343 8008. E-mail addresses: [email protected] (R. Proulx), [email protected] (L. Parrott). available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/ecolind 1470-160X/$ – see front matter # 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.ecolind.2007.02.005
Transcript of Measures of structural complexity in digital images for monitoring the ecological signature of an...

Measures of structural complexity in digital images formonitoring the ecological signature of an old-growthforest ecosystem
Raphael Proulx, Lael Parrott *
Complex Systems Laboratory, Department of Geography, University of Montreal, C.P. 6128 succursale Centre-ville, Montreal, Que.,
Canada H3C 3J7
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4
a r t i c l e i n f o
Article history:
Received 28 September 2006
Received in revised form
14 February 2007
Accepted 19 February 2007
Keywords:
Ecological orientor
Structural complexity
Imagery
Forest
Spatiotemporal dynamics
Shannon entropy
Mean information gain
a b s t r a c t
Conducting field samples for monitoring ecological dynamics across multiple spatiotem-
poral scales is a difficult task using standard protocols. One alternative is to measure a
restricted set of variables which can serve as an ecological orientor (EO) for quantifying
habitat change. The objective of this article is to derive from digital images a measure of
structural complexity that may serve as a proximate EO for monitoring forest dynamics in
space and time. The mean information gain (MIG) index was used as a measure of structural
complexity in photographs taken directly in the field over the entire growing season. At a
small scene extent, the complexity of light intensity variations in digital images was
positively related to species richness. At larger scene extents, forest understorey and
overstorey layers showed predictable ecological signatures in structural complexity. In
general, intensity and chroma were the two color space components which yielded the
greatest sensitivity to habitat change through time. Within the framework of a standardized
photographic protocol, it seems therefore reasonable to consider MIG as a suitable EO for
monitoring forest dynamics in both space and time. Our results support the idea that it is
possible on one hand to adopt a more holistic view of ecological processes to gain, on the
other hand, spatial and temporal degrees of freedom for testing multiple scale hypotheses in
the field.
# 2007 Elsevier Ltd. All rights reserved.
avai lable at www.sc iencedi rec t .com
journal homepage: www.e lsev ier .com/ locate /ecol ind
1. Introduction
It is well established that although mechanisms driving
ecological dynamics are not well understood at the commu-
nity (intermediate) level, ecosystem theory is nonetheless
supported by robust principles occurring at either larger (e.g.,
landscape scaling laws; Lawton, 1999; Gaston, 2000) or smaller
(e.g., individual based allometric laws; Turchin, 2001; Marquet
et al., 2005; West and Brown, 2005) levels of taxonomic
resolution (see also Simberloff, 2004). Notwithstanding this
fact, ecologists are now faced with the additional challenge of
* Corresponding author. Tel.: +1 514 343 8064; fax: +1 514 343 8008.E-mail addresses: [email protected] (R. Proulx), lael.pa
1470-160X/$ – see front matter # 2007 Elsevier Ltd. All rights reservedoi:10.1016/j.ecolind.2007.02.005
uncovering organizing principles governing ecological
dynamics across multiple spatial and temporal scales (Sole
et al., 1999; Brown et al., 2002; Storch and Gaston, 2004).
In this context, organizing principles in natural systems
may be regarded as an epiphenomenon of heuristic ecological
goal functions (Wilhelm and Bruggeman, 2000) which encom-
pass the formation of higher-level structures emerging from
lower-level interactions (Margalef, 1963; Christensen, 1995;
Ulanowicz, 2004; Muller, 2005). In the last century the search
for goal functions, also more properly termed ecological
orientors (EO) in Muller and Leupelt (1998), has spread so
[email protected] (L. Parrott).
d.

e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4 271
widely among ecosystem theories that reviewing all of them
would necessitate an article length task. Any proximate EO (as
opposed to an absolute or universal one) is defined as the
quantity a living system tends to optimize, from a nonteleo-
logical view, in the course of its development. Contemporary
examples include concepts of: fitness, productivity, stability,
resilience, exergy, emergy, ascendancy, network efficiency,
metabolic activity, and criticality among others (Odum, 1969;
Kauffman, 1995; Muller and Leupelt, 1998; Jorgensen and
Muller, 2000; Fath et al., 2001). All these concepts are useful
descriptors of ecological communities, but share the common
drawback that their estimation requires excessively large
amounts of input data and variables that are difficult to
measure in the field.
Consequently a trade-off exists between the number of
variables, replicates, and visits that one can include in a field
sampling protocol, where each aspect (i.e., descriptive, spatial
and temporal) is constrained by extent and resolution limits
(Fig. 1). It follows that most ecological field protocols are
misbalanced towards maximizing the number of descriptive
variables, counterbalanced by poor spatial (or temporal)
extent and resolution (Proulx, in press). In other words,
proximate EO are often constructed from a single-scaled
multivariate standpoint (e.g., matrices of environmental or
community descriptors), preventing us from collecting suffi-
ciently long spatiotemporal datasets in situ. Thus, to incorpo-
rate more spatial and temporal degrees of freedom in field
protocols for monitoring ecosystems, a simple, rapid and
preferably cost effective EO sampling should be performed.
In a recent meta-analysis, Tews et al. (2004) reviewed a
proximate EO known as the habitat heterogeneity hypothesis
which arises from the empirical positive relationship between
habitat and species diversity (e.g., MacArthur and MacArthur,
1961; Roth, 1976). The authors showed how modifying scales
can affect our interpretation of this relationship and how
sampling constraints have systematically favored the descrip-
tive component of field protocols; i.e., measuring a large
number of variables (Fig. 1). Another important idea in Tews
et al. (2004) is that habitat heterogeneity is better quantified by
its structural complexity rather than its diversity per se. This
Fig. 1 – The sampling triangle illustrates, on one hand, the
necessity of increasing descriptive, spatial and temporal
resolution and extent for studying ecosystem changes at
multiple scales. On the other hand, for practical reasons a
trade-off always exists between the number of variables,
the sample size and the visiting frequency anyone can
achieve within the framework of a field protocol.
idea links with the arguments of Anand and Tucker (2003) who
call for a shift of emphasis from diversity (counts of biological
objects at a given time and place) to complexity measures
(spatiotemporal structure of a set of biological objects at a
given scale). For instance, two sites may well contain the same
proportion of the same biological objects but nonetheless
show very different complexity. Measures of complexity thus
require more than just a statistical distribution of parts.
Forest ecology has a long tradition of linking overstorey and
understorey vegetation structures to historical succession
dynamics (Bazzaz, 1975; Denslow, 1987; Whitney and Foster,
1988; reviewed in Millet et al., 1998). Contemporary measures
of vegetation structure were devised to quantify plant
architecture in a strict geometrical sense (Jennings et al.,
1999; Jonckheere et al., 2004; Parker et al., 2004) as opposed to
other measures of community composition. Considering that
forest light regimes in overstorey and understorey layers is an
important determinant of ecological processes at various
scales (Endler, 1993; Trichon et al., 1998; Thery, 2001;
Valladares et al., 2002; Montgomery, 2004) and that the
dynamics of forest light can be captured by photographs, it
appears logical to consider digital images as a starting point for
monitoring forest dynamics. In particular, we hypothesize
that heterogeneity in forest light can serve as an indicator of
the structural complexity of the vegetation.
The principal objective of this article is to derive from
digital images a measure of structural complexity that may
serve as a proximate EO for monitoring forest dynamics in
space and time. This objective relies on two key assumptions:
(1) mean information gain (MIG) represents a relevant
measure of the structural complexity in digital images, and
(2) structural complexity is a proximate EO of a forest
ecosystem. More specifically we aim to show the existence
of a positive relationship between MIG and species richness at
small scene extents. Furthermore, we expect to find pre-
dictable spatial and seasonal patterns of structural complexity
in forest understorey and overstorey vegetation layers at
larger extents.
2. Methodology
This study was conducted at the Gault Nature Reserve (Mount
St-Hilaire, Quebec, Canada), an old-growth forest which
comprises about 700 of the 1600 regional species of vascular
plants in a 10 km2 area, where dominant trees are sugar maple
(Acer saccharum) and American beech (Fagus grandifolia). The
Reserve shelters a gradient of species assemblages and
geographic conditions which have been extensively studied
by various research groups (www.mcgill.ca/gault/research/
bibliography). Field experiments were carried out over two
growing seasons and involved two types of experimental
protocols; hereafter described as snapshot and trajectory
experiments. The two experiments were designed to address
complementary questions regarding the relevance of our
approach at different temporal and scene extents. The
snapshot experiment aimed to evaluate the existence of a
positive relationship between structural complexity and
species richness at small scene extents (<1 m2). The goal of
the trajectory experiment was to demonstrate the sensitivity

Fig. 2 – Schematic plane view of photographic settings in
snapshot and trajectory experiments. See Table 1 for
complementary information.
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4272
of our structural complexity measure in detecting habitat
related features and temporal variations at a larger scene
extent (up to 900 m2).
2.1. Photographic settings
Digital images were taken with a digital camera (EOS Rebel 6.3
MP, Canon Inc., Tokyo, Japan) equipped with a 18–55 mm lens
(EF-S f3.5-5.6, Canon Inc., Tokyo, Japan), which is roughly
equivalent to a 30–90 mm lens under 35 mm standards. The
photographic settings for each experiment are given in Table 1
and Fig. 2. Since both the aperture and the focal length were
fixed, the shutter speed was used as a measure of the
luminance (i.e., sum of all illumination sources reaching the
lens). Under the aperture priority mode, the camera uses its
internal light meter to optimally expose the scene by adjusting
the shutter speed. The exact relationship between scene
luminance and shutter speed is a specific function of the
settings in Table 1. The more illuminated the scene is; the
faster the shutter speed and vice versa. The illumination of a
forest scene is sensitive to the sun orientation (daylight hour),
the shooting direction, weather conditions, and canopy
closure. It is important to stress that the average intensity
in an image is independent of the scene luminance (see
below). Discussions and field excursions with a professional
photographer confirmed the above settings.
Commercial digital cameras, such as the one described
above, typically record images in the RGB (red, green and blue)
color space. Because in commercial cameras the distribution of
transmittances overlaps considerably among these three
spectral bands, the representation of each image was converted
to the HSV (hue, saturation and value) color space following
Smith (1978). This color representation is more natural to a
human observer since it separates the pure color component
(hue) from chroma (saturation) and intensity (value) compo-
nents. Color intuitively refers to the dominance of wavelengths
in the light signal. Intensity is the grey tone, that is, the
departure of a hue from black, the color of zero energy. Finally,
chroma is defined as how much the light spectrum differs from
boththepurecolor and theachromatic component (i.e., the grey
of the same intensity). The three components are expected be of
ecological relevance for quantifying structural complexity in
natural scenes (cf., Endler, 1993; Thery, 2001).
Table 1 – Photographic settings for snapshot (summer 2004) anMount St-Hilaire Reserve, Quebec, Canada
Setting Snapshot e
Focal length 55.0 mm
Light metering mode 35 segment
Aperture diameter 5.6 mm
Focus distance 1.25 m
Tripod’s head above ground 0.25 m
Depth of field (DF) 1.05–1.45 m
Exposure mode Aperture pr
Camera pointing direction Inward
Time window for shooting 11 h30–12 h
Visual obstruction < DF Manually cl
White balance mode Manual
Image size 3.2MP (ca. 1
2.2. Structural complexity measures
Measures of structural complexity were derived from digital
images by applying information theoretic indices on each
component of the HSV color space. Prior to the analysis, each
pixel value was linearly rescaled to an integer between 1 and
10. From the relative distribution of rescaled pixel values gi in
the image it is possible to calculate an index of aspatial
heterogeneity (dominance sensu O’Neill et al., 1988) using
Shannon’s formula for entropy:
H½g� ¼ �XN
i¼1
pðg iÞ log pðg iÞ; (1)
where p(gi) is the probability of observing a pixel value inde-
pendently of its location in the image (i.e., aspatial hetero-
geneity) and N is the number of frequency bins (categories) of
pixel values. Similarly, it is possible to calculate an index of
spatial heterogeneity (contagion sensu O’Neill et al., 1988) as
follows:
H½x� ¼ �XN4
i¼1
pðxiÞ log pðxiÞ; (2)
d trajectory (summer 2005) experiments performed at the
xperiment Trajectory experiment
18.0 mm
s evaluative 35 segments evaluative
6.3 mm
15.0 m
1.0 m
2.0 m–infinity
iority Aperture priority
Outward
30 9 h30–15 h30
eared Avoided
Natural light
000 � 3000) 1.6MP (ca. 1000 � 1500)

e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4 273
whereherep(xi)denotestheprobabilityoffindingaspecific2 � 2
colored combinationxi in the image andN4 represents the num-
ber of theoretical 2 � 2 combinations. Since H[x] also expresses
the joint entropy (JE) among neighbor pixels we can write:
JE = H[x] = H[g(x,y), g(x+1,y), g(x+1,y+1), g(x,y+1)]. Superscripts in par-
entheses indicate relative locations to the right (x + 1, y), on the
diagonal (x + 1, y + 1), and underneath (x, y + 1) pixel coordinate
(x, y), thus returning different 2 � 2 squares of color combina-
tions. Following the same reasoning, H[g] represents the mar-
ginal entropy (ME) of the joint distribution. We can appreciate
from theabove observations that spatial heterogeneity depends
on the degree of aspatial heterogeneity in the image. For this
reason, to quantify structural complexity one needs an index
which is able to partition these two heterogeneity fractions.
The mean information gain (MIG) index was used to
quantify structural complexity (Andrienko et al., 2000) and
calculated as follows:
MIG ¼ H½x� �H½g�log N4 � log N1 ¼
JE�ME
log N4 � log N1 : (3)
MIG determines the amount of spatial heterogeneity (i.e.,
JE) that excludes the fraction devoted to aspatial heterogeneity
(i.e., ME). The fixed quantity (log N4 � log N1) represents the
maximum obtained for equiprobable distributions of both ME
and JE and it serves to normalize MIG estimates over the range
0–1. MIG is zero for uniform patterns and is maximal (MIG = 1)
for random ones. Although formula (3) predicts the existence
of a linear 1:1 relationship between ME and MIG for random
(structureless) processes, it is far from obvious why any linear
trend should hold in natural scenes, particularly at high ME
values (e.g., Fig. 3). Therefore, the presence of a relationship
other than 1:1 between structural complexity (i.e., MIG) and
aspatial heterogeneity (i.e., ME) in natural scenes might
suggest an intrinsic organizing process.
To avoid biasing the MIG index because not all possible 2 � 2
combinations are observable (sufficiently frequent) in the
image, the ratio of the total number of pixels to N4 should not
fallunder 100.Thisapproximation is based uponWolf’s formula
for a 1% relative precision (Wolf, 1999). Herein, we used N = 10,
which yielded a ratio of ca. 150 for the 1.6MP images (snapshot
experiment) and a ratio of ca. 600 for the 3.2MP ones (trajectory
experiment). The MIG index has been successfully used before
to study time-series in various ecological contexts (Klemm and
Lange, 1999; Lange, 1999) and is a well known complexity
measure in statistical physics (Wackerbauer et al., 1994).
Because MIG increases monotonously with spatial ran-
domness it may be difficult to assert which value amongst
several is the more complex in terms of its distance to
simplicity (see Parrott, 2005), as MIG can be anywhere between
the extremes of order (uniformity) and disorder (randomness).
To further investigate this aspect, the fourth-order mean
mutual information (MMI) was calculated on our images
(Wackerbauer et al., 1994) as:
MMI ¼ 4H½g� �H½x�4 log N1 � log N1 ¼
4ME� JE
4 log N1 � log N1 ; (4)
where the notation is the same as in formula (3). The MMI
function is maximal (MMI = 1) for uniform patterns and zero
for random ones; it indicates the presence of inherent spatial
correlations in the image. Lopez-Ruiz et al. (1995) proposed a
simple complexity index which tends to zero towards the
extremes of ordered and disordered patterns (see also Shiner
et al., 1999). Here, we can use their formalism to construct a
convex complexity index G from the interplay between MIG (as
a measure of disorder) and MMI (as a measure of order) as
follows:
G ¼ MIG�MMI (5)
Fig. 3 reports MIG, MMI, and G estimates obtained for a
gradient of spatial autocorrelation scales. For the present
purposes, G will be used to determine an intermediate MIG
value which identifies the critical state between order and
disorder in the dataset. This intermediate value will then serve
as a structural complexity orientor for interpreting the results.
MIG has two main advantages over G: (1) it is a well defined
index in statistical physics, and (2) it allows to the classifica-
tion of spatial patterns above (i.e., towards disorder) and below
(i.e., towards order) a critical state, a property not shared by G
(Shiner et al., 1999). All digital images were routinely processed
using batch routines in Matlab 6.5 (MathWorks Inc., Natick,
MA).
2.3. Snapshot experiment
From 7 June to 16 August 2004, diversity measures and digital
images were obtained at 225 sites. The sampling frame
consisted of a 0.5 m wide cubic quadrat made of small plastic
tubes. The frame was easy to dismount and to carry from place
to place in the Reserve. When a site was selected, the frame
was carefully mounted over the vegetation without disturbing
the material inside. Nine sites were simultaneously sampled
in 1 day. Each site was visited once during the season (i.e.,
snapshot) and purposely chosen to capture a mixture of biotic–
abiotic conditions (Fig. 4). These conditions ranged from rocky
summits to boggy lowlands, and from rangeland to forest
shade stands. More extreme conditions such as sites full of
woody debris, crowded with small boulders, or simply lying on
bare soils were also included to assess the robustness of the
method.
Each site was photographed around noon on two adjacent
sides of the cubic quadrat; the camera held outside and
pointing inward towards the quadrat (Fig. 2). For each image
the shutter speed was recorded and used as a proxy for the
luminance. To quantify biological diversity a complete survey
of the vegetation at each site was conducted. The survey
involved the identification of all fern, herb, shrub, and tree
species. A given species was counted whenever one of its
parts was visible inside the cubic quadrat. Additionally, the
total percent filling was estimated for all species together
using 10% increments. While percent cover serves to evaluate
the visual obstruction of a two-dimensional surface by
vegetation, percent filling aims at the same in three dimen-
sions. Thus, sites full of abiotic material received a low
percent filling value and heavily vegetated sites received high
values. Total percent filling inside each quadrat was eval-
uated by a single person from lateral obstruction estimates of
the vegetation.

Fig. 3 – Artificial ornaments created to return increasing (decreasing) MIG (MMI) values in the interval (0.2–0.7). Although
each image has a different MIG value, all have the maximal ME. For this reason, one cannot predict the presence of a
relationship between MIG and ME in a specific set of digital images, particularly at high ME values for non-random patterns,
since at this level the measure of structural complexity is less severely constrained by the aspatial heterogeneity (see text).
The convex complexity index G is maximal at intermediate MIG values and vanishes towards the extremes.
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4274
2.4. Trajectory experiment
This experiment was designed for monitoring temporal
fluctuations (i.e., trajectory) in structural complexity. Eighteen
sites were visited on a weekly basis for a total of 26 weeks from
May 10 to November 10 2005. The sites were visited in the same
sequence each week and sampled within a 48 h period from
Tuesday to Thursday. The 18 sampling sites were selected
from a larger set of 69 permanent stations put in place for
quantifying multivariate relationships between community,
environmental and geographic descriptors at the Gault
Reserve (Gilbert and Lechowicz, 2004). The final selection
was based on plant species lists (Gilbert, pers. commun.) and
direct observations in the field to ensure a suitable topography
for photographic purposes. Nine sites were located directly
above or right next to small creeks of varying width (ca. 0.2–
1.5 m), whereas another group of nine were located in drier
sites (Fig. 4). At the center of each site a steel rod was planted in

Fig. 4 – (a) An example of images captured in high and low diversity sites in 2004. (b) Images taken from the side view in dry
and humid habitats in 2005. The white dashed line (horizon line) separates overstorey and understorey layers in these
images. (c) Top view images of the same scene at different moments in 2005.
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4 275
a vertical position and at a fixed height above the ground.
These rods served as markers for positioning the tripod when
revisiting sites. Sampling always occurred within the 9 h30–
15 h30 daylight interval to control for large periodic variations
in illumination, both in direction and luminance.
Photographs were taken as shown in Fig. 2, with the camera
held at the center of the site and pointing outwards in five
directions: north, east, south, west and top. In addition to the
use of steel rods for positioning the tripod, multiple markers
and reference points on the tripod were used to ensure that
the same scene was photographed over time.
For this experiment, time, luminance, zenith and azimuth
angles were used as quantitative covariables in the analyses,
whereas habitat (humid or dry), layer (understorey or over-
storey) and orientation (side or top view) were used as factor
variables (Fig. 4). The zenith angle from the vertical and the
azimuth angle eastward from the north were obtained for
each image using the algorithm of Reda and Andreas (2005).
Among factor variables, habitat distinguished the two major
groups of sites (i.e., humid or dry; Fig. 4) and layer determined
the portion of the image (i.e., overstorey or understorey; Fig. 4)
being analysed. Our choice of photographic settings ensured
that the horizon line would split the scene in half, vertically
separating overstorey and understorey layers in side view
images (Fig. 4). Finally, the orientation factor regroups images
captured with the camera positioned for shooting either side
(i.e., north, east, south and west) or top views (Figs. 2 and 4).
Although overstorey and understorey layers have no parti-
cular meaning in top view images, MIG estimates were
nonetheless calculated to provide a balanced statistical
design, but one should not expect a significant effect of this
variable in this case.

Fig. 5 – Convex relationship between MIG and G for the
three color space components, for both trajectory and
snapshot experiments. Hue: solid (—); chroma: dotted
(- - -); intensity: dashed (– –). G is maximal when MIG is
close to the identified critical state of 0.4. The sample size
of each curve is n = 4705. Curves were smoothed using a
weighted moving average (LOWESS interpolator; tension
sets to 0.3) on raw estimates.
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4276
2.5. Statistical analyses
For both the snapshot and trajectory experiments, bivariate
regressions were assessed between ME and MIG for each
image component to determine their deviation from the 1:1
relationship predicted for random processes.
For the snapshot experiment, relationships were inspected
between: (1) MIG estimates on two adjacent sides of the same
cubic quadrat, (2) percent filling and MIG, and (3) species
richness and MIG, where MIG was independently calculated
for the color (hue), chroma (saturation) and intensity (value)
components of each image. Luminance, as measured by
shutter speed, was not significantly related to MIG in the
snapshot experiment. Consequently, to simplify the presenta-
tion of the results, luminance was not considered in relation-
ships between species richness (or percent filling) and MIG.
For the trajectory experiment, general linear models (GLM)
were obtained to predict the main effects (and interaction) of
habitat and layer factors on MIG, with time, luminance, zenith
and azimuth as covariables. Individual bivariate regressions
were performed between each covariable and MIG variables to
detect significant dependency. Only covariables showing
significant dependency with MIG were retained for statistical
modeling. One GLM was performed for each dependent MIG
color space (i.e., color, chroma and intensity) and each
orientation (i.e., side or top view), for a total of six statistical
models. The number of terms included in the model was kept
at the minimum and stepwise selection procedures were
avoided.
The luminance variable was log transformed prior to
statistical analysis. Omnibus (Legendre and Legendre, 1998)
and log transformations were applied on time to linearize the
variable. The temporal order of the images was reorganized in
a systematic fashion by permuting blocks of weeks as follows:
the original series of 26 weeks [(1, 2, 3, 4), (5, 6, 7, 8), (9, 10, 11, 12,
13, 14), (15, 16, 17, 18), (19, 20, 21, 22), (23, 24, 25, 26)] was
omnibus transformed to [(1, 2, 3, 4), (23, 24, 25, 26), (19, 20, 21,
22), (5, 6, 7, 8), (15, 16, 17, 18), (9, 10, 11, 12, 13, 14)]. This
permutation positions early-spring and late-fall samples at
the beginning of the series, and mid-summer samples at the
end.
Since our objective was to demonstrate the sensitivity of
our structural complexity measure in detecting habitat and
temporal changes using common statistical tools, MIG
estimates for multiple images across space were averaged
to avoid dealing with intricate autocorrelation functions and
pseudo-replication that could be linked to sampling loca-
tions. Each week, the results for the four directions in each
site and the nine sites in each habitat-layer combination
were averaged to conserve only four estimates per week.
The overall number of degrees of freedom in the analyses
was decreased to 104, thus reducing the probability of doing
a type I error and buffering occasional outliers (if any) in
this large dataset. Each of the 104 points therefore
represents the MIG average of either 36 side view or 9 top
view images. This statistical framework may be overly
conservative but possesses the advantage of providing
straightforward interpretable results. The complete inves-
tigation of the spatiotemporal autocorrelation structure in
these sequences will be the focus of a forthcoming article.
All statistical tests were performed with Systat 11.0 (Systat
Software Inc., Richmond, CA).
3. Results
3.1. Relationships between G, MMI and ME in the images
Fig. 5 shows the convex relationship between MIG and G for
each of the three color space components. For the chroma and
intensity components, G is maximal for MIG values in the
range (0.35–0.45). The hue curve did not behave as a convex
function and was found below the two other curves, probably
because of low estimates of aspatial diversity in this
component. Therefore, for the remainder of this text the
notions of high and low structural complexity in digital images
will refer to their relative distance from MIG = 0.4.
For the three color space components, linear regression
slopes between ME and MIG in both experiments deviated
significantly from one (Fig. 6), indicating a non-random spatial
structure in the images. For the snapshot experiment, slope
estimates were 0.22, 0.25 and 0.26 for color, chroma and
intensity, respectively. For the trajectory experiment, slopes
for color, chroma and intensity were respectively 0.36, 0.59
and 0.41 in side view images while being 0.41, 0.46 and 0.25 in
top view images. Lower R2 were the result of MIG estimates
obtained for a narrow range of high ME values and forming
clumps visible in Fig. 6.
3.2. Snapshot experiment
Because the MIG estimates obtained on two adjacent sides of
the same quadrat were well correlated (Pearson’s r: 0.65, 0.73
and 0.82 for color, chroma and intensity) only one of the two
estimates was retained for the ensuing analyses. The above

Fig. 6 – Relationships between ME and MIG, for side view (*) and top view (*) images obtained from snapshot (n = 225) and
trajectory (n = 936) experiments. The first column of panels shows results for the snapshot experiment. Relationships are
reported for each of the three image components in the HSV color space. The expected relationship for a random process is
represented by the 1:1 dashed line. Least-square determination coefficients (R2) were low for chroma and intensity (0.20 and
0.11), but moderate for color (0.64) in the snapshot experiment. In the trajectory experiment, R2 varied among components as
follows—side view: 0.58, 0.57, 0.32 and top view: 0.76, 0.67, 0.33, respectively for color, chroma and intensity.
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4 277
correlations suggest that MIG efficiently captures site specific
signatures, probably in relation to the structural complexity of
the vegetation.
3.2.1. Relationship between species richness and structuralcomplexitySpecies richness alone was a better determinant of structural
complexity than percent filling which yielded correlations to
MIG close to zero (Fig. 7). A positive association was observed
between species richness and MIG (mean � 1S.E.) in the
intensity component, whereas no such behaviors were
detected for color and chroma (Fig. 7b). Results from the
snapshot experiment at a small scene extent indicate that
structural complexity was maximal for more diversified sites.
3.3. Trajectory experiment
Among our set of covariables time and luminance showed
significant dependency on MIG using raw observations
(n = 4680), whereas zenith and azimuth did not (R2 < 0.05).

Fig. 7 – Relationships (mean W 1S.E.) between (a) percent
filling and MIG, (b) species richness and MIG, for the
snapshot experiment. The sample sizes are reported in
the top margins for each semi-quantitative category.
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4278
Even when forcing zenith and azimuth variables into our
models to check if these could explain some of the residual
variance, their specific contribution remained at trivial levels.
The time omnibus transformation was well fitted by a third-
order polynomial function and constituted an acceptable
linearization of time in all statistical models. Usual statistical
prerequisites of the GLM such as residual heteroscadicity and
normality were inspected and fulfilled.
3.3.1. Effect of time, luminance, habitat and layer on
structural complexityIn addition to an intercept, six more terms were included in
our GLM: time, luminance, habitat, layer, habitat � layer and
habitat � time. The layer � time interaction was initially
included but explained minor fractions of the variation in
MIG and was removed before fitting final models. Transformed
time and luminance covariables were found to be respectively
negatively and positively associated to MIG. However, lumi-
nance did not always show significant relationships because a
part of its variation was shared by other variables in the model.
In fact, when removing luminance from models the MIG
fraction of the variation explained by this term was almost
entirely transferred to the time explanatory fraction. Such an
observation suggests that luminance differences in response
to weather conditions have only a weak effect on structural
complexity estimates. Note that this assertion is also true for
the snapshot experiment. Structural complexity was at a
minimum during early-spring and fall surveys and at a
maximum during weeks 7–10 (i.e., late-June to early-July 2005).
For side view images, we found a significant effect of
habitat and layer factors in the intensity component (overall
R2 = 0.73). These two factors explained little of the variation in
MIG for the color and chroma models (Table 2). The MIG
estimate for intensity was the most complex in understorey
layers of dry habitats (Fig. 8; time adjusted mean
MIG � 1S.E. = 0.39 � 0.01) and the less complex in overstorey
layers of humid habitats (Fig. 8; time adjusted mean
MIG � 1S.E. = 0.48 � 0.01). An interaction between habitat
and time for intensity was also detected (Table 2; Fig. 8). A
significant difference in structural complexity between humid
and dry habitats was present in spring and fall, but remained
untraceable in summer samples as indicated by the habi-
tat � time interaction. For top view images, the GLM showed a
significant effect of habitat in all color space components
(Table 3). Habitat effects are depicted in Fig. 9 for intensity and
chroma, as well as for the habitat � time interaction in chroma
(Fig. 9). As expected, the layer factor was not significant in top
view images. It is interesting to note that a MIG value of about
0.4 behaved as an orientor in both sets of images. MIG
estimates of 0.4 were reached in early-spring and late-fall
samples among top view images, while reached in mid-
summer samples among side view images (Figs. 8 and 9).
4. Discussion
Major findings of this study revealed that MIG, as a measure of
structural complexity, can capture site specific differences
across space and time in an old-growth temperate forest. At a
small scene extent, the complexity of light intensity variations
in digital images was positively related to species richness.
The understorey and overstorey (side views) as well as the
canopy (top view) vegetation patterns showed trends in their
structural complexity signatures through time. Intensity and,
to a lesser degree, chroma, were the two image components
which yielded the greatest sensitivity to habitat changes
independently of the scene luminance and sun orientation.
Within the framework of a standardized photographic proto-
col, it seems therefore reasonable to consider MIG as a suitable
EO (ecological orientor) for monitoring forest dynamics.
4.1. Relationship between aspatial heterogeneity (ME) andstructural complexity (MIG)
It is intriguing that within an experimental treatment a linear
relationship between ME and MIG was found in many color
space components. Furthermore, the slope of these relation-
ships varied significantly among and between experimental
treatments (each color space component, in each set of
images, in each experiment). One may therefore conjecture
that once photographic settings are fixed then habitat
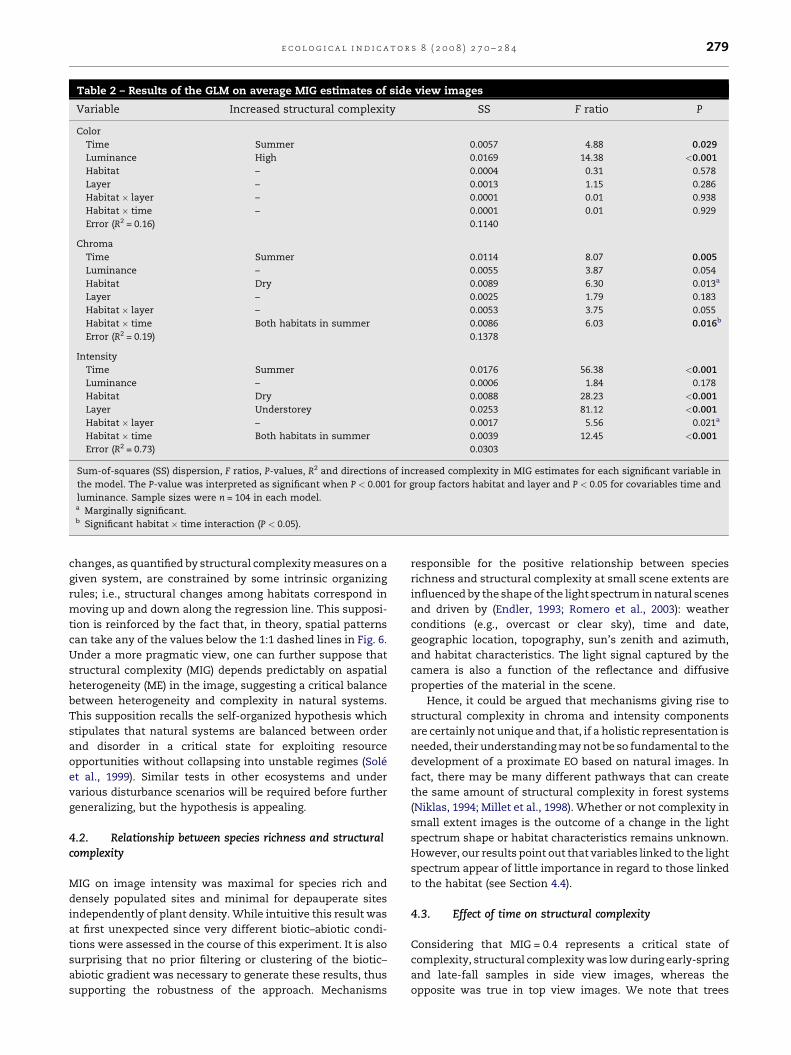
Table 2 – Results of the GLM on average MIG estimates of side view images
Variable Increased structural complexity SS F ratio P
Color
Time Summer 0.0057 4.88 0.029
Luminance High 0.0169 14.38 <0.001
Habitat – 0.0004 0.31 0.578
Layer – 0.0013 1.15 0.286
Habitat � layer – 0.0001 0.01 0.938
Habitat � time – 0.0001 0.01 0.929
Error (R2 = 0.16) 0.1140
Chroma
Time Summer 0.0114 8.07 0.005
Luminance – 0.0055 3.87 0.054
Habitat Dry 0.0089 6.30 0.013a
Layer – 0.0025 1.79 0.183
Habitat � layer – 0.0053 3.75 0.055
Habitat � time Both habitats in summer 0.0086 6.03 0.016b
Error (R2 = 0.19) 0.1378
Intensity
Time Summer 0.0176 56.38 <0.001
Luminance – 0.0006 1.84 0.178
Habitat Dry 0.0088 28.23 <0.001
Layer Understorey 0.0253 81.12 <0.001
Habitat � layer – 0.0017 5.56 0.021a
Habitat � time Both habitats in summer 0.0039 12.45 <0.001
Error (R2 = 0.73) 0.0303
Sum-of-squares (SS) dispersion, F ratios, P-values, R2 and directions of increased complexity in MIG estimates for each significant variable in
the model. The P-value was interpreted as significant when P < 0.001 for group factors habitat and layer and P < 0.05 for covariables time and
luminance. Sample sizes were n = 104 in each model.a Marginally significant.b Significant habitat � time interaction (P < 0.05).
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4 279
changes, as quantified by structural complexity measures on a
given system, are constrained by some intrinsic organizing
rules; i.e., structural changes among habitats correspond in
moving up and down along the regression line. This supposi-
tion is reinforced by the fact that, in theory, spatial patterns
can take any of the values below the 1:1 dashed lines in Fig. 6.
Under a more pragmatic view, one can further suppose that
structural complexity (MIG) depends predictably on aspatial
heterogeneity (ME) in the image, suggesting a critical balance
between heterogeneity and complexity in natural systems.
This supposition recalls the self-organized hypothesis which
stipulates that natural systems are balanced between order
and disorder in a critical state for exploiting resource
opportunities without collapsing into unstable regimes (Sole
et al., 1999). Similar tests in other ecosystems and under
various disturbance scenarios will be required before further
generalizing, but the hypothesis is appealing.
4.2. Relationship between species richness and structuralcomplexity
MIG on image intensity was maximal for species rich and
densely populated sites and minimal for depauperate sites
independently of plant density. While intuitive this result was
at first unexpected since very different biotic–abiotic condi-
tions were assessed in the course of this experiment. It is also
surprising that no prior filtering or clustering of the biotic–
abiotic gradient was necessary to generate these results, thus
supporting the robustness of the approach. Mechanisms
responsible for the positive relationship between species
richness and structural complexity at small scene extents are
influenced by the shape of the light spectrum in natural scenes
and driven by (Endler, 1993; Romero et al., 2003): weather
conditions (e.g., overcast or clear sky), time and date,
geographic location, topography, sun’s zenith and azimuth,
and habitat characteristics. The light signal captured by the
camera is also a function of the reflectance and diffusive
properties of the material in the scene.
Hence, it could be argued that mechanisms giving rise to
structural complexity in chroma and intensity components
are certainly not unique and that, if a holistic representation is
needed, their understanding may not be so fundamental to the
development of a proximate EO based on natural images. In
fact, there may be many different pathways that can create
the same amount of structural complexity in forest systems
(Niklas, 1994; Millet et al., 1998). Whether or not complexity in
small extent images is the outcome of a change in the light
spectrum shape or habitat characteristics remains unknown.
However, our results point out that variables linked to the light
spectrum appear of little importance in regard to those linked
to the habitat (see Section 4.4).
4.3. Effect of time on structural complexity
Considering that MIG = 0.4 represents a critical state of
complexity, structural complexity was low during early-spring
and late-fall samples in side view images, whereas the
opposite was true in top view images. We note that trees
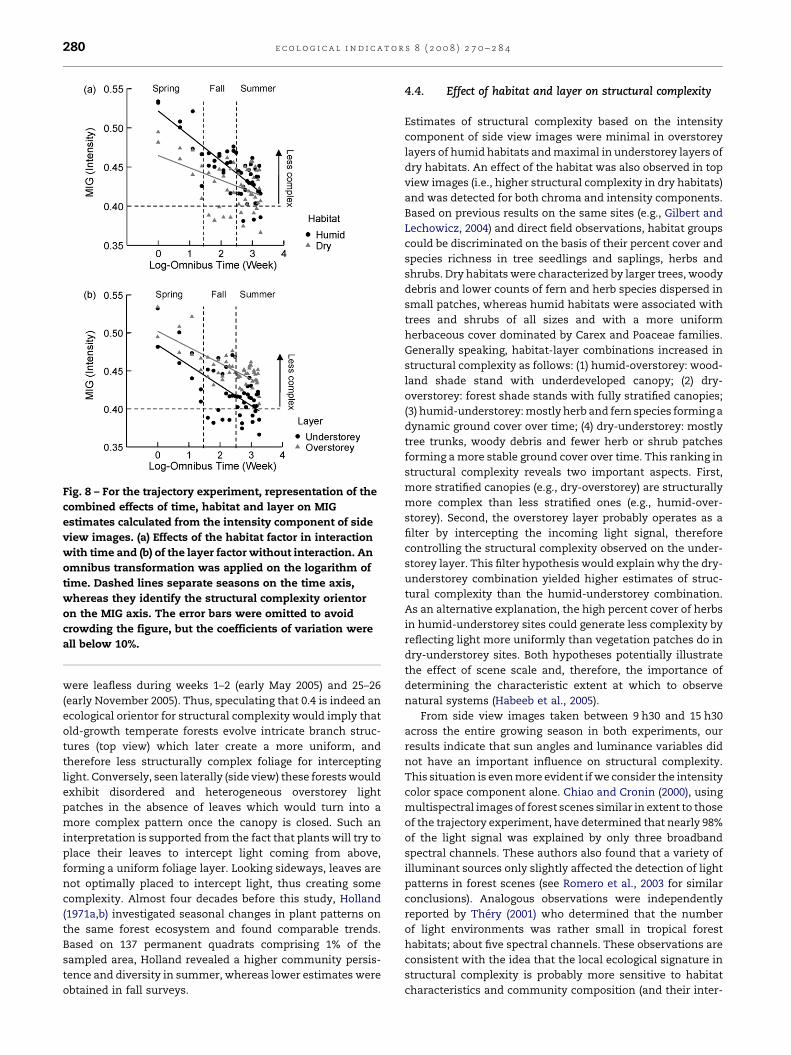
Fig. 8 – For the trajectory experiment, representation of the
combined effects of time, habitat and layer on MIG
estimates calculated from the intensity component of side
view images. (a) Effects of the habitat factor in interaction
with time and (b) of the layer factor without interaction. An
omnibus transformation was applied on the logarithm of
time. Dashed lines separate seasons on the time axis,
whereas they identify the structural complexity orientor
on the MIG axis. The error bars were omitted to avoid
crowding the figure, but the coefficients of variation were
all below 10%.
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4280
were leafless during weeks 1–2 (early May 2005) and 25–26
(early November 2005). Thus, speculating that 0.4 is indeed an
ecological orientor for structural complexity would imply that
old-growth temperate forests evolve intricate branch struc-
tures (top view) which later create a more uniform, and
therefore less structurally complex foliage for intercepting
light. Conversely, seen laterally (side view) these forests would
exhibit disordered and heterogeneous overstorey light
patches in the absence of leaves which would turn into a
more complex pattern once the canopy is closed. Such an
interpretation is supported from the fact that plants will try to
place their leaves to intercept light coming from above,
forming a uniform foliage layer. Looking sideways, leaves are
not optimally placed to intercept light, thus creating some
complexity. Almost four decades before this study, Holland
(1971a,b) investigated seasonal changes in plant patterns on
the same forest ecosystem and found comparable trends.
Based on 137 permanent quadrats comprising 1% of the
sampled area, Holland revealed a higher community persis-
tence and diversity in summer, whereas lower estimates were
obtained in fall surveys.
4.4. Effect of habitat and layer on structural complexity
Estimates of structural complexity based on the intensity
component of side view images were minimal in overstorey
layers of humid habitats and maximal in understorey layers of
dry habitats. An effect of the habitat was also observed in top
view images (i.e., higher structural complexity in dry habitats)
and was detected for both chroma and intensity components.
Based on previous results on the same sites (e.g., Gilbert and
Lechowicz, 2004) and direct field observations, habitat groups
could be discriminated on the basis of their percent cover and
species richness in tree seedlings and saplings, herbs and
shrubs. Dry habitats were characterized by larger trees, woody
debris and lower counts of fern and herb species dispersed in
small patches, whereas humid habitats were associated with
trees and shrubs of all sizes and with a more uniform
herbaceous cover dominated by Carex and Poaceae families.
Generally speaking, habitat-layer combinations increased in
structural complexity as follows: (1) humid-overstorey: wood-
land shade stand with underdeveloped canopy; (2) dry-
overstorey: forest shade stands with fully stratified canopies;
(3) humid-understorey: mostly herb and fern species forming a
dynamic ground cover over time; (4) dry-understorey: mostly
tree trunks, woody debris and fewer herb or shrub patches
forming a more stable ground cover over time. This ranking in
structural complexity reveals two important aspects. First,
more stratified canopies (e.g., dry-overstorey) are structurally
more complex than less stratified ones (e.g., humid-over-
storey). Second, the overstorey layer probably operates as a
filter by intercepting the incoming light signal, therefore
controlling the structural complexity observed on the under-
storey layer. This filter hypothesis would explain why the dry-
understorey combination yielded higher estimates of struc-
tural complexity than the humid-understorey combination.
As an alternative explanation, the high percent cover of herbs
in humid-understorey sites could generate less complexity by
reflecting light more uniformly than vegetation patches do in
dry-understorey sites. Both hypotheses potentially illustrate
the effect of scene scale and, therefore, the importance of
determining the characteristic extent at which to observe
natural systems (Habeeb et al., 2005).
From side view images taken between 9 h30 and 15 h30
across the entire growing season in both experiments, our
results indicate that sun angles and luminance variables did
not have an important influence on structural complexity.
This situation is even more evident if we consider the intensity
color space component alone. Chiao and Cronin (2000), using
multispectral images of forest scenes similar in extent to those
of the trajectory experiment, have determined that nearly 98%
of the light signal was explained by only three broadband
spectral channels. These authors also found that a variety of
illuminant sources only slightly affected the detection of light
patterns in forest scenes (see Romero et al., 2003 for similar
conclusions). Analogous observations were independently
reported by Thery (2001) who determined that the number
of light environments was rather small in tropical forest
habitats; about five spectral channels. These observations are
consistent with the idea that the local ecological signature in
structural complexity is probably more sensitive to habitat
characteristics and community composition (and their inter-

Table 3 – Results of the GLM on average MIG estimates of top view images
Variable Increased structural complexity SS F ratio P
Color
Time Spring and fall 0.0151 16.69 <0.001
Luminance High 0.0231 25.48 <0.001
Habitat Dry 0.0098 10.86 <0.001
Layer – 0.0002 0.21 0.644
Habitat � layer – 0.0127 14.08 <0.001a
Habitat � time – 0.0009 1.07 0.303
Error (R2 = 0.66) 0.0878
Chroma
Time Spring and fall 0.0387 41.63 <0.001
Luminance High 0.0389 41.88 <0.001
Habitat Dry 0.0286 30.79 <0.001
Layer – 0.0014 1.53 0.218
Habitat � layer – 0.0234 25.21 <0.001a
Habitat � time – 0.0056 6.09 0.015b
Error (R2 = 0.80) 0.0903
Intensity
Time Spring and fall 0.0339 114.23 <0.001
Luminance High 0.0143 48.08 <0.001
Habitat Dry 0.0117 39.31 <0.001
Layer – 0.0001 0.14 0.411
Habitat � layer – 0.0109 36.94 <0.001a
Habitat � time – 0.0001 0.44 0.508
Error (R2 = 0.88) 0.0288
Sum-of-squares (SS) dispersion, F ratios, P-values, R2 and directions of increased complexity in MIG estimates for each significant variable in
the model. The P-value was interpreted as significant when P < 0.001 for group factors habitat and layer and P < 0.05 for covariables time and
luminance. Sample sizes were n = 104 in each model.a Since there is a significant effect of habitat and that no effect was expected for layer (see text), the significance value of their interaction
should not be interpreted.b Significant habitat � time interaction (P < 0.05).
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4 281
action) than to the shape of the light spectrum itself. In fact,
recent studies suggest that many different sets of functional
trade-offs in plants may well translate into an equal overall
fitness within a habitat (Niklas, 1994; Lei and Lechowicz, 1998;
Marks and Lechowicz, 2006). Using a three-dimensional
modeling program of leaf and crown characteristics, Valla-
dares et al. (2002) examined the light capture efficiency of 24
seedling and herbaceous species in a lowland rainforest.
Despite important differences in all morphological (structural)
variables, the species were highly similar in their light capture
efficiency based on measures of photon flux densities
(Valladares et al., 2002). Consequently, although the number
of different light spectra may be relatively low in forest scenes,
the variety of living structures evolved locally by the ecological
community is expected to be much broader.
4.5. General comments on the photographic approach
The present study is not the first to consider digital images to
characterize habitat features. Marsden et al. (2002) used
filtered and binarized side view images for measuring the
complexity of understorey vegetation in tropical forests
following logging activities. Alados et al. (1999) investigated
the effect of grazing through fractal and lacunarity analyses of
preprocessed, filtered and binarized side view images of a
Mediterranean shrub (Anthullis cytisoides). Using hemispherical
photography in the tropical rainforest, Trichon et al. (1998)
reported differences in gap patterns which they related to
three disturbance phases; gap, building and mature phases.
Spatial patterns were therein quantified by canopy openness,
spherical variance and plant area indices. The above-cited
studies revealed interesting results in regards to their
hypotheses, but required specifically designed image seg-
mentation protocols. These studies were also directed to
answer specific questions and, as a consequence, were not
discussed in the context of a proximate EO for monitoring
ecosystem dynamics in space and time.
To incorporate more spatial and temporal degrees of
freedom in field protocols, a simple, rapid and preferably cost
effective EO sampling should be performed. In this context,
satellite based sensors have led to the greatest advances,
including standardized measures like the Normalized Differ-
ence Vegetation Index (cf., Badeck et al., 2004; Stockli and
Vidale, 2004) and the Leaf Area Index (cf., Jonckheere et al.,
2004). Although satellites can return data on a landscape scale
their limits are reached at local scales, and they are not always
appropriate for detecting vertical structures under the canopy
cover. Moreover, airborne remote sensing is not what one
would call a simple and cost effective approach for monitoring
changes in space and time. Atmospheric and geometric
corrections are often tedious and the low temporal resolution
of most remotely sensed data makes monitoring short term
dynamics difficult. Alternatively, the deployment of field
sensor networks (Baldocchi et al., 2001; Green et al., 2005)
such as ARTS (Automated Radio Telemetry System), NEON
(National Ecological Observatory Network) and FLUXNET will
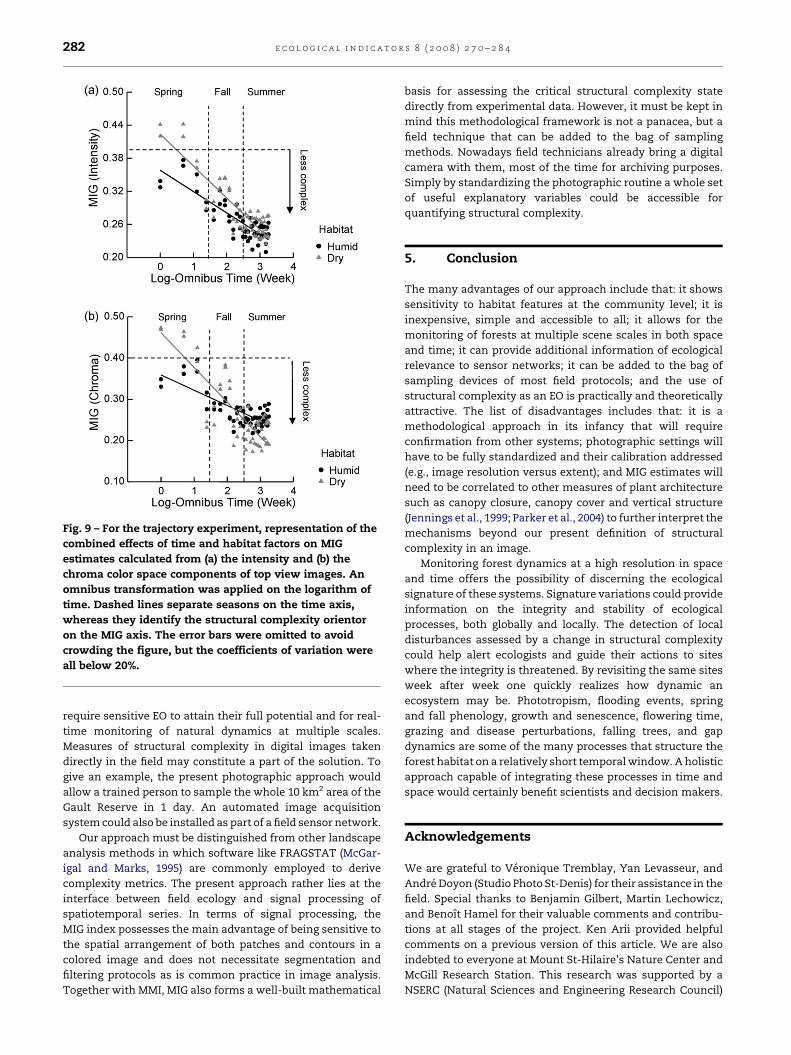
Fig. 9 – For the trajectory experiment, representation of the
combined effects of time and habitat factors on MIG
estimates calculated from (a) the intensity and (b) the
chroma color space components of top view images. An
omnibus transformation was applied on the logarithm of
time. Dashed lines separate seasons on the time axis,
whereas they identify the structural complexity orientor
on the MIG axis. The error bars were omitted to avoid
crowding the figure, but the coefficients of variation were
all below 20%.
e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4282
require sensitive EO to attain their full potential and for real-
time monitoring of natural dynamics at multiple scales.
Measures of structural complexity in digital images taken
directly in the field may constitute a part of the solution. To
give an example, the present photographic approach would
allow a trained person to sample the whole 10 km2 area of the
Gault Reserve in 1 day. An automated image acquisition
system could also be installed as part of a field sensor network.
Our approach must be distinguished from other landscape
analysis methods in which software like FRAGSTAT (McGar-
igal and Marks, 1995) are commonly employed to derive
complexity metrics. The present approach rather lies at the
interface between field ecology and signal processing of
spatiotemporal series. In terms of signal processing, the
MIG index possesses the main advantage of being sensitive to
the spatial arrangement of both patches and contours in a
colored image and does not necessitate segmentation and
filtering protocols as is common practice in image analysis.
Together with MMI, MIG also forms a well-built mathematical
basis for assessing the critical structural complexity state
directly from experimental data. However, it must be kept in
mind this methodological framework is not a panacea, but a
field technique that can be added to the bag of sampling
methods. Nowadays field technicians already bring a digital
camera with them, most of the time for archiving purposes.
Simply by standardizing the photographic routine a whole set
of useful explanatory variables could be accessible for
quantifying structural complexity.
5. Conclusion
The many advantages of our approach include that: it shows
sensitivity to habitat features at the community level; it is
inexpensive, simple and accessible to all; it allows for the
monitoring of forests at multiple scene scales in both space
and time; it can provide additional information of ecological
relevance to sensor networks; it can be added to the bag of
sampling devices of most field protocols; and the use of
structural complexity as an EO is practically and theoretically
attractive. The list of disadvantages includes that: it is a
methodological approach in its infancy that will require
confirmation from other systems; photographic settings will
have to be fully standardized and their calibration addressed
(e.g., image resolution versus extent); and MIG estimates will
need to be correlated to other measures of plant architecture
such as canopy closure, canopy cover and vertical structure
(Jennings et al., 1999; Parker et al., 2004) to further interpret the
mechanisms beyond our present definition of structural
complexity in an image.
Monitoring forest dynamics at a high resolution in space
and time offers the possibility of discerning the ecological
signature of these systems. Signature variations could provide
information on the integrity and stability of ecological
processes, both globally and locally. The detection of local
disturbances assessed by a change in structural complexity
could help alert ecologists and guide their actions to sites
where the integrity is threatened. By revisiting the same sites
week after week one quickly realizes how dynamic an
ecosystem may be. Phototropism, flooding events, spring
and fall phenology, growth and senescence, flowering time,
grazing and disease perturbations, falling trees, and gap
dynamics are some of the many processes that structure the
forest habitat on a relatively short temporal window. A holistic
approach capable of integrating these processes in time and
space would certainly benefit scientists and decision makers.
Acknowledgements
We are grateful to Veronique Tremblay, Yan Levasseur, and
Andre Doyon (Studio Photo St-Denis) for their assistance in the
field. Special thanks to Benjamin Gilbert, Martin Lechowicz,
and Benoıt Hamel for their valuable comments and contribu-
tions at all stages of the project. Ken Arii provided helpful
comments on a previous version of this article. We are also
indebted to everyone at Mount St-Hilaire’s Nature Center and
McGill Research Station. This research was supported by a
NSERC (Natural Sciences and Engineering Research Council)

e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4 283
grant to L. Parrott and a FQRNT (Fond Quebecois de Recherche
sur la Nature et les Technologies) scholarship to R. Proulx.
r e f e r e n c e s
Alados, C.L., Escos, J., Emlen, J.M., Freeman, D.C., 1999.Characterization of branch complexity by fractal analyses.Int. J. Plant Sci. 160, S147–S155.
Anand, M., Tucker, B.C., 2003. Defining biocomplexity: anecological perspective. Comments Theor. Biol. 8, 497–510.
Andrienko, Y.A., Brilliantov, N.V., Kurths, J., 2000. Complexity oftwo-dimensional patterns. Eur. Phys. J. 15, 539–546.
Badeck, F.-W., Bondeau, A., Bottcher, K., Dokor, D., Lucht, W.,Schaber, J., Sitch, S., 2004. Responses of spring phenology toclimate change. New Phytol. 162, 295–309.
Baldocchi, D.D., Falge, E., Gu, L., Olson, R., Hollinger, D.,Running, S., Anthoni, P., Bernhofer, Ch., Davis, K., Evans, R.,Fuentes, J., Goldstein, A., Katul, G., Law, B., Lee, X., Malhi, Y.,Meyers, T., Munger, J.W., Oechel, W., Pilegaard, K., Schmid,H.P., Valentini, R., Verma, S., Vesala, T., Wilson, K., Wofsy,S., 2001. FLUXNET, a new tool to study the temporal andspatial variability of ecosystem-scale carbon dioxide, watervapor and energy flux densities. Bull. Am. Meteorol. Soc. 82,2415–2434.
Bazzaz, F.A., 1975. Plant species diversity in old-fieldsuccessional ecosystems in Southern Illinois. Ecology 56,485–488.
Brown, J.H., Gupta, V.K., Li, B.-L., Milne, B.T., Restrepo, C., West,G.B., 2002. The fractal nature of nature: power laws,ecological complexity and biodiversity. Phil. Trans. R Soc.Lond. B: Biol. Sci. 357, 619–626.
Chiao, C.-C., Cronin, T.W., 2000. Color signal in natural scenes:characteristics of reflectance spectra and effect of naturalilluminants. J. Opt. Soc. Am. A 17, 218–224.
Christensen, V., 1995. Ecosystem maturity-towardsquantification. Ecol. Model. 77, 3–32.
Denslow, J.S., 1987. Tropical rainforest gaps and tree speciesdiversity. Annu. Rev. Ecol. Syst. 18, 431–451.
Endler, J.A., 1993. The color of light in forest and its implication.Ecol. Monogr. 63, 1–27.
Fath, B.D., Patten, B.C., Choi, J.S., 2001. Complementarity ofecological goal functions. J. Theor. Biol. 208, 493–506.
Gaston, K.J., 2000. Global patterns in biodiversity. Nature 405,220–227.
Gilbert, B., Lechowicz, M.J., 2004. Neutrality, niches, anddispersal in a temperate forest understorey. Proc. Natl.Acad. Sci. 101, 7651–7656.
Green, J.L., Hasting, A., Arzberger, P., Ayala, F.J., Cottingham,K.L., Cuddington, K., Davis, F., Dunne, J.A., Fortin, M.-J.,Gerber, L., Neubert, M., 2005. Complexity in ecology andconservation, mathematical, statistical, and computationalchallenges. Bioscience 55, 501–510.
Habeeb, R.L., Trebilco, J., Wotherspoon, S., Johnson, C.R., 2005.Determining natural scales of ecological systems. Ecol.Monogr. 75, 467–487.
Holland, P.G., 1971a. Seasonal change in the shoot flora diversityof hardwood forest stands on Mount St. Hilaire, Quebec.Can. J. Bot. 49, 1713–1720.
Holland, P.G., 1971b. Seasonal change in the plant patterns ofdeciduous forest in southern Quebec (Canada). Oikos 22,137–148.
Jennings, S.B., Brown, N.D., Sheil, D., 1999. Assessing forestcanopies and understorey illumination: canopy closure,canopy cover and other measures. Forestry 72, 59–73.
Jonckheere, I., Fleck, S., Nackaerts, K., Muys, B., Coppin, P.,Weiss, M., Baret, F., 2004. Review of methods for in situ leaf
area index determination. Part I. Theories, sensors andhemispherical photography. Agric. For. Meteorol. 121,19–35.
Jorgensen, S.E., Muller, F. (Eds.), 2000. Handbook of EcosystemTheories and Management. CRC Press/Lewis Publishers,Boca Raton.
Kauffman, S., 1995. At home in the Universe: The Search for theLaws of Self-organization and Complexity. OxfordUniversity Press, New York.
Klemm, O., Lange, H., 1999. Trends of air pollution in theFichtelgebirge Mountains, Bavaria. Environ. Sci. Pollut. Res.6, 193–199.
Lange, H., 1999. Are ecosystems dynamical systems? Int. J.Comput. Anticipatory Syst. 3, 169–186.
Lawton, J.H., 1999. Are there general laws in ecology? Oikos 84,177–192.
Legendre, P., Legendre, L., 1998. Numerical Ecology, 2nd Englished. Elsevier, New York.
Lei, T.T., Lechowicz, M.J., 1998. Diverse responses of maplesaplings to forest light regimes. Ann. Bot. 82, 9–19.
Lopez-Ruiz, R., Mancini, H.L., Calbert, X., 1995. A statisticalmeasure of complexity. Phys. Lett. A 209, 321–326.
MacArthur, R.H., MacArthur, J.W., 1961. On bird speciesdiversity. Ecology 42, 594–598.
Margalef, R., 1963. On certain unifying principles in ecology. Am.Nat. 97, 357–374.
Marquet, P.A., Quinones, R.A., Abades, S., Labra, F., Tognelli, M.,Arim, M., Rivadeneira, M., 2005. Scaling and power-laws inecological systems. J. Exp. Biol. 208, 1749–1769.
Marks, C.O., Lechowicz, M.J., 2006. Alternative designs and theevolution of functional diversity. Am. Nat. 167, 55–66.
Marsden, S.J., Fielding, A.H., Mead, C., Hussin, M.Z., 2002. Atechnique for measuring the density and complexity ofunderstorey vegetation in tropical forests. For. Ecol.Manage. 165, 117–123.
McGarigal, K., Marks, B.J., 1995. FRAGSTATS: Spatial PatternAnalysis Program for Quantifying Landscape Structure,PNW-GTR-351. United States Department of Agriculture,Pacific Northwest Research Station, Oregon.
Millet, J., Bouchard, A., Edelin, C., 1998. Plant succession andtree architecture: an attempt at reconciling two scales ofanalysis of vegetation dynamics. Acta Biotheor. 46, 1–22.
Montgomery, R.A., 2004. Effects of understorey foliage onpatterns of light attenuation near the forest floor. Biotropica36, 33–39.
Muller, F., 2005. Indicating ecosystem and landscapeorganisation. Ecol. Indicat. 5, 280–294.
Muller, F., Leupelt, M. (Eds.), 1998. Eco Targets, Goal Functionsand Orientors. Springer, Berlin.
Niklas, K.J., 1994. Morphological evolution through complexdomains of fitness. Proc. Natl. Acad. Sci. 91, 6772–6779.
Odum, E.P., 1969. The strategy of ecosystem development.Science 164, 262–270.
O’Neill, R.V., Krumme1, J.R., Gardner, R.H., Sugihara, G., Jackson,B., DeAngelist, D.L., Milne, B.T., Turner, M.G., Zygmunt, B.,Christensen, S.W., Dale, V.H., Graham, R.L., 1988. Indices oflandscape pattern. Landscape Ecol. 3, 153–162.
Parker, G.G., Harmon, M.E., Lefsky, M.A., Chen, J., Van Pelt, R.,Weiss, S.B., Thomas, S.C., Winner, W.E., Shaw, D.C.,Franklin, J.F., 2004. Three-dimensional structure of an old-growth Pseudotsuga-tsuga canopy and its implications forradiation balance, microclimate, and gas exchange.Ecosystems 7, 440–453.
Parrott, L., 2005. Quantifying the complexity of simulatedspatiotemporal population dynamics. Ecol. Complex. 2,175–184.
Proulx, R., in press. Ecological complexity for unifying ecologicaltheory across scales, a field ecologist’s perspective. Ecol.Complex.

e c o l o g i c a l i n d i c a t o r s 8 ( 2 0 0 8 ) 2 7 0 – 2 8 4284
Reda, I., Andreas, A., 2005. Solar position algorithm for solarradiation application. National Renewable EnergyLaboratory (NREL) Technical report NREL/TP-560-34302,Colorado.
Romero, J., Hernandez-Andres, J., Nieves, J.L., Garcia, J.A., 2003.Color coordinates of objects with daylight changes. ColorRes. Appl. 28, 25–35.
Roth, R.R., 1976. Spatial heterogeneity and bird species diversity.Ecology 57, 773–782.
Shiner, J.S., Davidson, M., Landsberg, P.T., 1999. Simple measurefor complexity. Phys. Rev. E 59, 1459–1464.
Simberloff, D., 2004. Community ecology: is it time to move on?Am. Nat. 163, 787–799.
Smith, A.R., 1978. Color Gamut Transform Pairs. In: Beatty, J.C.,Booth, K.S. (Eds.), 1982. Tutorial, Computer Graphics, 2nded. IEEE Computer Society Press, Silver Spring, pp. 376–383.
Sole, R.V., Manrubia, S.C., Benton, M., Kauffman, S., Bak, P.,1999. Criticality and scaling in evolutionary ecology. TrendsEcol. Evol. 14, 156–160.
Stockli, R., Vidale, P.L., 2004. European plant phenology andclimate as seen in a 20-year AVHRR land-surface parameterdataset. Int. J. Remote Sens. 25, 3303–3330.
Storch, D., Gaston, K.J., 2004. Untangling ecological complexityon different scales of space and time. Basic Appl. Ecol. 5,389–400.
Tews, J., Brose, U., Grimm, V., Tielborger, K., Wichmann, M.C.,Schwager, M., Jeltsch, F., 2004. Animal species diversitydriven by habitat heterogeneity/diversity, the importance ofkeystone structures. J. Biogeogr. 31, 79–92.
Thery, M., 2001. Forest light and its influence on habitatselection. Plant Ecol. 163, 251–261.
Trichon, V., Walter, J-M.N., Laumonier, Y., 1998. Identifyingspatial patterns in the tropical rain forest usinghemispherical photographs. Plant Ecol. 137, 227–244.
Turchin, P., 2001. Does population ecology have general laws?Oikos 94, 17–26.
Ulanowicz, R.E., 2004. On the nature of ecodynamics. Ecol.Complex. 1, 341–354.
Valladares, F., Skillman, J.B., Pearcy, R.W., 2002. Convergence inlight capture efficiencies among tropical forest understoryplants with contrasting crown architectures: a case ofmorphological compensation. Am. J. Bot. 89, 1275–1284.
Wackerbauer, R., Witt, A., Atmanspacher, H., Kurths, J.,Scheingraber, H., 1994. A comparative classification ofcomplexity measures. Chaos Solitons Fractals 4, 133–173.
West, G.B., Brown, J.H., 2005. The origin of allometric scalinglaws in biology from genomes to ecosystems, towards aquantitative unifying theory of biological structure andorganization. J. Exp. Biol. 208, 1575–1592.
Whitney, G.G., Foster, D.R., 1988. Overstorey composition andage as determinant of the understorey flora of woods ofcentral New England. J. Ecol. 76, 867–876.
Wilhelm, T., Bruggeman, R., 2000. Goal functions for thedevelopment of natural systems. Ecol. Model. 132, 231–246.
Wolf, F., 1999. Berechnung von information and komplexitat inZeitreihen: Analyse des wasserhaushalts von bewaldeteneinzugsgebieten. PhD Thesis, University of Bayreuth,Bayreuth (in German).




















