
MCE Article
12
Review The Polycystic Ovary Syndrome and recent human evolution Stephen Corbett a,b,⇑ , Laure Morin-Papunen c,1 a Population Health, Clinical Support Division (Western), Gungurra (Building 68) Cumberland Hospital, Locked Bag 7118, Parramatta, BC NSW 2124, Australia b School of Public Health and Western Clinical School, University of Sydney, Australia c Department of Gynecology and Obstetrics, PL 23, FIN 90029 OYS, Oulu, Finland article info Article history: Available online xxxx Keywords: Polycystic Ovary Syndrome Fertility selection Fecundability Reproductive ecology Evolution Thrifty genotype abstract The Polycystic Ovary Syndrome (PCOS) is a complex endocrine disorder characterised both by reproduc- tive and metabolic disturbance, and is the most common cause globally of ovarian infertility. It is also a familial polygenic condition, linked genetically to both Type 2 diabetes and the metabolic syndrome. The striking evolutionary paradox of this prominent genetically-based condition, which impairs fertility, is that not only should it have diminished in prevalence, but it should have done so rapidly – unless there has been some form of balancing selection. The emerging discipline of evolutionary medicine can provide important insights into the causes and patterns of occurrence of common diseases such as PCOS. In this paper we review the impacts of PCOS on infertility, fecundability and lifetime reproductive success and then critically appraise published hypotheses about the evolutionary origins of PCOS and related conditions. Crown Copyright Ó 2013 Published by Elsevier Ireland Ltd. All rights reserved. Contents 1. Introduction .......................................................................................................... 00 2. Fecundability, infertility and lifetime reproductive success in PCOS ............................................................. 00 2.1. Fecundability and infertility in PCOS ................................................................................. 00 2.2. Early pregnancy loss in PCOS ....................................................................................... 00 2.3. Pregnancy and lifetime reproductive success .......................................................................... 00 2.4. Obesity, under-nutrition and fertility in PCOS ......................................................................... 00 3. Fertility selection ...................................................................................................... 00 4. Nutritional history and reproductive ecology of Homo sapiens .................................................................. 00 5. Not necessarily a thrifty gene ............................................................................................ 00 6. Hyperandrogenism, PCOS and human evolution ............................................................................. 00 6.1. Direct effects on growth, behaviour and immune function ............................................................... 00 6.2. Fewer offspring concentrates maternal investment ..................................................................... 00 6.3. Effects of maternal androgens on infant development ................................................................... 00 7. Evolutionary hypotheses and the Polycystic Ovary Syndrome .................................................................. 00 7.1. What is the object of explanation? .................................................................................. 00 7.2. Has the object of explanation been influenced by evolution? ............................................................. 00 7.3. What kind of trait is the object of explanation? ........................................................................ 00 7.3.1. Fixed traits .............................................................................................. 00 7.3.2. Facultative traits .......................................................................................... 00 7.3.3. Human genes ............................................................................................ 00 7.4. What kind of explanation is sought? ................................................................................. 00 7.4.1. Mechanism .............................................................................................. 00 7.4.2. Development ............................................................................................. 00 7.4.3. Phylogeny ............................................................................................... 00 7.4.4. Function ................................................................................................ 00 0303-7207/$ - see front matter Crown Copyright Ó 2013 Published by Elsevier Ireland Ltd. All rights reserved. http://dx.doi.org/10.1016/j.mce.2013.01.001 ⇑ Corresponding author at: Population Health, Clinical Support Division (Western), Gungurra (Building 68) Cumberland Hospital, Locked Bag 7118, Parramatta, BC NSW 2124, Australia. Tel.: +61 2 97994585; fax: +61 2 98403608. E-mail addresses: [email protected] (S. Corbett), [email protected].fi (L. Morin-Papunen). 1 Fax: +358 8 3154310. Molecular and Cellular Endocrinology xxx (2013) xxx–xxx Contents lists available at SciVerse ScienceDirect Molecular and Cellular Endocrinology journal homepage: www.elsevier.com/locate/mce Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycystic Ovary Syndrome and recent human evolution. Molecular and Cellular En- docrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
Transcript of MCE Article

Molecular and Cellular Endocrinology xxx (2013) xxx–xxx
Contents lists available at SciVerse ScienceDirect
Molecular and Cellular Endocrinology
journal homepage: www.elsevier .com/locate /mce
Review
The Polycystic Ovary Syndrome and recent human evolution
Stephen Corbett a,b,⇑, Laure Morin-Papunen c,1
a Population Health, Clinical Support Division (Western), Gungurra (Building 68) Cumberland Hospital, Locked Bag 7118, Parramatta, BC NSW 2124, Australiab School of Public Health and Western Clinical School, University of Sydney, Australiac Department of Gynecology and Obstetrics, PL 23, FIN 90029 OYS, Oulu, Finland
a r t i c l e i n f o a b s t r a c t
Article history:Available online xxxx
Keywords:Polycystic Ovary SyndromeFertility selectionFecundabilityReproductive ecologyEvolutionThrifty genotype
0303-7207/$ - see front matter Crown Copyright � 2http://dx.doi.org/10.1016/j.mce.2013.01.001
⇑ Corresponding author at: Population Health, Clini2124, Australia. Tel.: +61 2 97994585; fax: +61 2 984
E-mail addresses: [email protected] Fax: +358 8 3154310.
Please cite this article in press as: Corbett, S., Mdocrinology (2013), http://dx.doi.org/10.1016/j.
The Polycystic Ovary Syndrome (PCOS) is a complex endocrine disorder characterised both by reproduc-tive and metabolic disturbance, and is the most common cause globally of ovarian infertility. It is also afamilial polygenic condition, linked genetically to both Type 2 diabetes and the metabolic syndrome. Thestriking evolutionary paradox of this prominent genetically-based condition, which impairs fertility, isthat not only should it have diminished in prevalence, but it should have done so rapidly – unless therehas been some form of balancing selection. The emerging discipline of evolutionary medicine can provideimportant insights into the causes and patterns of occurrence of common diseases such as PCOS. In thispaper we review the impacts of PCOS on infertility, fecundability and lifetime reproductive success andthen critically appraise published hypotheses about the evolutionary origins of PCOS and relatedconditions.
Crown Copyright � 2013 Published by Elsevier Ireland Ltd. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 002. Fecundability, infertility and lifetime reproductive success in PCOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
2.1. Fecundability and infertility in PCOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 002.2. Early pregnancy loss in PCOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 002.3. Pregnancy and lifetime reproductive success . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 002.4. Obesity, under-nutrition and fertility in PCOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
3. Fertility selection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 004. Nutritional history and reproductive ecology of Homo sapiens . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 005. Not necessarily a thrifty gene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 006. Hyperandrogenism, PCOS and human evolution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
6.1. Direct effects on growth, behaviour and immune function . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 006.2. Fewer offspring concentrates maternal investment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 006.3. Effects of maternal androgens on infant development . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
7. Evolutionary hypotheses and the Polycystic Ovary Syndrome . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
7.1. What is the object of explanation? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.2. Has the object of explanation been influenced by evolution? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.3. What kind of trait is the object of explanation? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.3.1. Fixed traits . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.3.2. Facultative traits . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.3.3. Human genes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
7.4. What kind of explanation is sought? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
7.4.1. Mechanism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.4.2. Development. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.4.3. Phylogeny . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.4.4. Function . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00013 Published by Elsevier Ireland Ltd. All rights reserved.
cal Support Division (Western), Gungurra (Building 68) Cumberland Hospital, Locked Bag 7118, Parramatta, BC NSW03608..au (S. Corbett), [email protected] (L. Morin-Papunen).
orin-Papunen, L. The Polycystic Ovary Syndrome and recent human evolution. Molecular and Cellular En-mce.2013.01.001

2 S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx
Pleasedocrin
7.5. Are all viable hypotheses considered ? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
7.5.1. Are there different vulnerabilities to PCOS in different populations? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.5.2. What categories of explanation are under consideration?. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.6. Could multiple explanations be correct? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.7. How can these hypotheses be tested? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
7.7.1. Understanding variation in PCOS susceptibility . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.7.2. Establishing the existence and intensity of positive or negative fertility selection for an insulin resistant/PCOS phenotype in human
populations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 007.7.3. Investigating the mechanisms and circumstances by which insulin resistance or sensitivity can optimise reproductive outcomes 00
8. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
1. Introduction
The Polycystic Ovary Syndrome is a complex endocrine disordercharacterised by reproductive and metabolic disturbance – poly-cystic ovaries, anovulation and hyperandrogenism, and insulinresistance, hyperinsulinaemia, pancreatic ß cell dysfunction andhyperlipidaemia. It is the most common cause globally of ovarianinfertility and is a familial polygenic condition (Franks et al.,1997; Franks and McCarthy, 2004; Vink et al., 2009), linked genet-ically to both Type 2 diabetes and the metabolic syndrome. Theprevalence of PCOS is between 6% and 10% in developed countries(Broekmans et al., 2006), although this clinical syndrome sits atopa spectrum of disordered polycystic ovarian morphology and met-abolic function with an estimated prevalence of 20–30% in devel-oped countries (Balen and Michelmore, 2002) including up to52% of South Asian immigrant women in Britain (Rodin et al.,1998).
The evolutionary paradox of the high global prevalence of PCOSis stark. RA Fisher, one of the three architects of the modern syn-thesis of Darwinian evolution, recognised that the intensity ofselection for genes which influence fertility has the potential tobe 1 to 2 orders of magnitude greater than those which confer aviability advantage (Fisher, 1930a,b). So PCOS, a genetically basedcondition which impairs fertility, should, in the absence of balanc-ing selection, not only diminish in prevalence, but do so rapidly.This paradox has stimulated the publication of at least 11 hypoth-eses about the evolutionary origins of PCOS, a majority of whichposit a mechanism for positive balancing fertility or viability selec-tion as a counterweight to the fertility disadvantages associatedwith this condition (Table 1, hypotheses numbered for reference).
Evolutionary hypotheses about disease vulnerability have thepotential to frame and synergise with scientific endeavours tosearch for proximate causes of disease. They can also be, as Ste-phen Jay Gould opined, evolutionary ‘‘just so’’ stories – unsup-ported but plausible verbal arguments which purport to explainobservable natural phenomena (Gould, 1985). Nesse (2011) has re-cently proposed a series of 10 questions which can be asked of evo-lutionary hypotheses about disease vulnerability, and which arelikely to enhance the coherence and utility of these propositions.A list of these questions, appropriately abbreviated for this pur-pose, is presented in Table 2.
In this paper we review the evidence relating to the magnitudeof the intensity of selection against the PCOS phenotype in contem-porary populations, as measured by impacts on fertility, fecundityand lifetime reproductive success (Byars et al., 2010; Cavalli-Sforzaand Bodmer, 1971). This evidence is fundamental to an under-standing current and recent evolution of the PCOS phenotype.
To frame our review and analysis of current evolutionaryhypotheses, we present a brief account of four issues germane tothis review; the potential of fertility selection to effect rapid evolu-tionary changes, the reproductive ecology and nutritional historyof the human species, role of metabolic thrift and energy efficiency,and hyperandrogenism as important factors (Lotka, 1922a,b; Neel,
cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
1962, 1999) in the evolutionary biology of PCOS and Type 2diabetes.
We then critically review published hypotheses about the evo-lutionary origins of PCOS through the prism of Nesse’s 10 ques-tions, and draw some conclusions about the validity of thesehypotheses, how they might be tested, and their relevance for evo-lutionary, clinical, epidemiological and population geneticsresearch.
2. Fecundability, infertility and lifetime reproductive success inPCOS
The reproductive dysfunction in women with PCOS relates bothto subfertility secondary to anovulation, and to early pregnancyloss. Anovulation is the cause of infertility in about one third ofcouples seeking for infertility treatment, and PCOS accounts for90% of those cases (Balen and Rutherford, 2007).
The exact pathogenesis of anovulation is complex, and manypossible mechanisms have been postulated. Ovarian function isdisturbed, with resultant abnormal folliculogenesis and steroido-genesis (see chapter. . . in this issue of the journal) and lack ofdevelopment of a dominant follicle. Hyperandrogenaemia, hyper-insulinaemia and obesity seem to have the greatest impact on ovu-lation and pregnancy survival. Other factors, such as abnormalitiesin gonadotrophin secretion (increased secretion of LH and de-creased secretion of FSH), derangements in plasminogen activatorinhibitor activity and in ovarian paracrine and autocrine function,decreased apoptosis in the ovary, and abnormal vascular reactivitysecondary to endothelial dysfunction have been shown to play alsoa role in the pathogenesis of anovulation. All these disorders mayalso disturb implantation and increase the risk of early pregnancyloss through alterations in the endometrial environment (van derSpuy and Dyer, 2004).
2.1. Fecundability and infertility in PCOS
Fecundability estimates the probability of conception of a clin-ically detectable pregnancy within 12 months. Fecundability is cal-culated by using the time to pregnancy (TTP), which measures inmonths the time from the beginning of unprotected intercourseuntil the first pregnancy.
A large Finnish cohort (Koivunen et al., 2008) of 4535 women isthe only cohort study to compare the fecundability of women report-ing symptoms of PCOS – oligo-amenorrhea and/or hirsutism - tothe fecundability of non-symptomatic women. Only comparisonsat age 31 have been published.
Oligo-amenorrhea and/or hirsutism (fecundability rate,FR = 0.74, p < 0.001) and obesity (FR = 0.68, p = 0.002) were bothindependently associated with decreased fecundability. Womenreporting both hirsutism and oligo-amenorrhea had the lowestchance of conceiving within 12 months (FR 0.39, 95% CI 0.27–0.54). Those with oligo-amenorrhea alone also had significantly re-
Ovary Syndrome and recent human evolution. Molecular and Cellular En-

Table 1PCOS evolutionary hypotheses.
No. Authors Year Hypothesis Summary
1 Robinson et al. 1992 Metabolic thrift Reduced post-prandial thermogenesis in PCOS and Type2 Diabetes reduces overall energy expenditure
2 Holte 1998 Metabolic thrift Metabolic thrift, muscular strength and increased fatstorage are advantages under low energy conditions
3 Abbott and Dumesic 2002 Developmental origins Foetal exposure to excess androgens programmeshypothalamic and pituitary control of LuteinisingHormone excretion, enhancing abdominal fatdistribution, thereby predisposing to insulin resistanceand anovulation
4 Barnett and Abbott 2003 Reproductive adaptations to a large brained foetus The development of a large brained foetus requiresextended priming of the uterine endometrium (duringthe a prolonged follicular phase of the menstrual cycle),to meet the demands of a growing brain afterimplantation. This adaptation makes humans vulnerableto disrupted follicle development and anovulation
5 Kirchengast 2005 Sexual selection Decreased mating and reproductive success due to themasculinizing features of PCOS
6 Gleischer and Barad 2006 Fertility storage An increase in the number of immature follicles in PCOSis evidence of ‘‘fertility storage’’, conferring an advantageboth by becoming available in lean times and by latermenopause
7 (a) Escobar-Morreale et al. 2005 Androgenisation and reproduction infection defence Early Reproductive Maturation and increased intervalbetween pregnancies could aid maternal and infantsurvival. Production of pro-inflammatory cytokines,protecting against infection
7 (b) Insulin resistance Insulin resistance, metabolic thrift, and fat storage8 Eggers 2007 Kin selection Preferential survival of families in which less fertile
PCOS mothers could contribute to child rearing andsurvival
9 (a) Shaw and Elton 2008 Food deprivation Women with PCOS phenotype have a quicker ovulationtrigger in times of food deprivation
9 (b) Refeeding Women with a PCOS phenotype more readily return tomenses with refeeding after a period of food deprivation
9 (c) Transgenerational privation In challenging nutritional environments maternalandrogens in women with the PCOS phenotype willconfer benefits for the foetus, such as increased fatstorage and increased follicular readiness in adulthood
10 Corbett 2009 Fertility first – PCOS, T2D and MS are manifestations of a genotypeattuned to the environmental and dietary conditions ofthe late Pleistocene– Selection for an insulin sensitive phenotype wasconstrained by a fertility advantage of PCOS phenotypeduring the Agrarian period– The direction of this selection was reversed with thepopulation wide improvements in nutrition whichbegan in Europe in the 18th century and have nowextended to 60% of the world’s population– Rapid and recent decreases in the prevalence of thegenes underlying this condition are likely to have beenachieved through the powerful mechanism of fertilityselection
11 (a) Azziz 2010 Benefits of androgenisation Increased bone and muscle strength fromandrogenisation conferred a survival benefit in huntergatherer societies
11 (b) Insulin resistance enhances fertility Insulin resistance under low energy conditions maydivert available glucose towards ovulatory function
S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx 3
duced fecundability compared with non-symptomatic women(FR = 0.75, 95% CI 0.62–0.91), whereas hirsutism alone was notassociated with decreased fecundability, which may well be ex-plained by the subjectivity of this symptom (it was defined bythe complaint of the patient without any objective score).
As expected, in this same cohort, women with PCOS symptomssuffered significantly more often from infertility that non-symp-tomatic women (26.5% versus 17.1%, p < 0.01). For those who hadsought help for infertility, the main reason for infertility was attrib-utable to the women in 52.5% of the couples in the symptomaticgroup and in 26.3% in the non-symptomatic group (p < 0.001)(Koivunen et al., 2008). Similarly, in two retrospective Swedishstudies, one conducted in women who had undergone ovarianwedge resection in the past (22–31 years previously) (Dahlgren
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
et al., 1992) and the other conducted on 91 of women with a diag-nosis of PCOS done 14 to 22 years before, (Hudecova et al., 2009) aneven greater rate of women with PCOS (70–76%) had attendedgynecological clinics because of infertility problems, and 64–67%had been treated medically for infertility.
In this Finnish study self-reporting of symptoms alone, ratherthan in combination with either measured androgen levels orabdominal ultrasonography were used in the reporting of infertil-ity by phenotype. Although misclassification could bias these re-sults, the authors of this study had previously shown a strongcorrelation between symptoms and hormonal profiles (Taponenet al., 2003) and a positive predictive value of oligomenorrhoeaand hirsutism of 70.4% for the presence of polycystic ovaries(Taponen et al., 2004). The addition of hormonal profiles and
Ovary Syndrome and recent human evolution. Molecular and Cellular En-

Table 2Ten Questions for evolutionary studies of disease vulnerability.
Task 1: Define the object of explanation preciselyQ1. Is the object of explanation a uniform trait in the species, or is the goal to explain variations in a trait among groups or individuals?Q2. Has the object of explanation been influenced by evolution?Q3. What kind of trait is the object of explanation?
a. A fixed human trait, such as the narrow birth canalb. A facultative trait, such as the capacity for sweatingc. Human genes, in the most general sense of the term
Task 2: Specify the kind of explanation soughtQ4. Is the goal to explain the evolution of the trait, or its proximate mechanisms?Q5. Is the goal to explain the trait’s phylogeny, or the evolutionary forces that shaped it?
Task 3: List and consider all viable hypothesesQ6. Are all viable hypotheses considered and given fair consideration, or are some hypotheses neglected, while others receive favoured treatment?Q7. Could different vulnerabilities cause the disease in different individuals or subgroups?Q8. What categories of explanation are under consideration?
a. Mismatch of bodies with environments they did not evolve inb. Constraints on selection, such as time required, genetic drift, and mutationc. Trade Offsd. Reproductive success at the expense of health
Q9. Could multiple explanations be correct?
Task 4: Describe the methods used to test the hypothesesQ10. What methods are used to test the hypotheses?
a. Consistency with evolutionary theoryb. Modelling using quantitative methodsc. Comparative methodsd. Experimental methods
4 S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx
information on ovarian morphology will enhance the precisionwith which the anovulatory and PCOS phenotypes can be described(Dewailly et al., 2006; Sanchon et al., 2012).
2.2. Early pregnancy loss in PCOS
The decreased number of deliveries in women with PCOS mayalso been explained by an increased rate of early pregnancy losses.Hospital based studies of women seeking treatment for infertilitysuggest that early pregnancy loss occurs in 30–50% in women withPCOS (Balen, 1993; Glueck et al., 1999; Homburg et al., 1988; Sagleet al., 1988), a 3-fold increase higher than reported in retrospectivestudies in healthy women (Gray and Wu, 2000; Regan et al., 1989).Most of these studies, however, have focused on hospital-basedsubjects, including women seeking infertility treatment and thusdo not necessarily represent the general population. Other re-ported series have also noted normal early pregnancy loss ratesin women with the syndrome (Dahlgren et al., 1992; Winteret al., 2002). In the population-based cohort study mentionedabove, symptomatic women had a similar cumulative incidenceof at least one spontaneous abortion as non-symptomatic womenand the number of spontaneous abortions/recurrent spontaneousabortions per woman did not differ between symptomatic andnon-symptomatic women (Koivunen et al., 2008).
2.3. Pregnancy and lifetime reproductive success
In the Finnish cohort study, symptomatic women became preg-nant at least once as frequently as non-symptomatic women,although the time to pregnancy was longer. At the age 31 theyhad given birth successfully to 1–2 children as often as referencewomen, suggesting that fecundability could be restored by infertil-ity treatment. At the age of 31 years, however, women with bothhirsutism and irregular menstruation had a smaller family size(7.7% with more than 3 deliveries) than non-symptomatic women(16.8% with more than 3 deliveries) or women with only one PCOSsymptom (Koivunen et al., 2008). This data is in keeping with re-sults of a retrospective study, showing that 24% of women with
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
PCOS who had undergone ovarian wedge resection remained child-less compared with 16% of the referents (Dahlgren et al., 1992). Itseems therefore that women with PCOS exhibit a decreased nativ-ity compared with a reference population.
In a Swedish study, there was no difference between the PCOSpatients and controls in number of women who had given birth.Among women who had attempted a pregnancy, a similar propor-tion of the PCOS patients (87%) and control subjects (92%) had gi-ven birth to at least one child. The average number of children inwomen with PCOS was 1.67 and in controls was 1.87 (Hudecovaet al., 2009). Although statistically not significant in this smallgroup, a difference in LRS of this magnitude sustained over gener-ations could result in a selection intensity of 20% per generation(Bodmer, 1964).
A recent study by Sanchon et al. (2012) recruited 592 pre-men-opausal women blood donors and assessed clinical and biochemi-cal features of hyperandrogenism. Women with PCOS had 0.5(p < 0.05) pregnancies per woman less than healthy women,although all women by definition had not completed their repro-ductive lives and healthy women were on average 5 years olderthan women with PCOS.
Ovarian reserve seems to be better preserved in women withPCOS, as the mean ovarian volume, the number of antral folliclesand the serum level of anti-Müllerian hormone in premenopausalPCOS women have been shown to be significantly higher than incorresponding control subjects (Hudecova et al., 2009; Nikolaouand Gilling-Smith, 2004).
In summary, women with symptoms of PCOS and/or PCOS havelower fecundability and more frequent infertility problems thanasymptomatic women. Whether they have an increased risk ofspontaneous abortion remains controversial. In the North FinlandBirth Cohort women with a diagnosis of PCOS, followed until theage of 31, an approximate mid-point of their reproductive lives,had had approximately 0.3 children less than normal women.There were no differences however in the proportion having 1–2children, suggesting that fecundability can be restored, at leastpartly, by infertility treatment. This cohort has recently been fol-lowed up at age 45, presaging a more precise estimate of lifetimereproductive success when these results are available.
Ovary Syndrome and recent human evolution. Molecular and Cellular En-
S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx 5
2.4. Obesity, under-nutrition and fertility in PCOS
Obesity is common in PCOS, and almost all obese subjects withthe syndrome have insulin resistance (Dunaif et al., 1987; Holteet al., 1994). It seems that obesity and PCOS play an independentand synergistic role in the etiology of infertility. In a populationof 1281 consecutive patients examined for symptoms related toandrogen excess, 82% of them had PCOS and 30% infertility prob-lems, and patients with infertility were significantly more obese(body mass index, BMI 34.0 ± 8.9 kg/m2) than their non-infertilecounterparts (32.5 ± 9.2 kg/m2, p < 0.03) (Azziz et al., 2004a,b). Arecent cohort of 3029 consecutive sub-fertile couples shown that,after adjusting for other confounding variables and excluding cou-ples with anovulation, tubal pathology and male factors, the prob-ability of spontaneous pregnancy declined linearly with thewomen’s BMI over 29 kg/m2 (HR 0.95 per kg/m2 above 29 kg/m2,95%CI 0.91–0.99). In case of a woman with a BMI of 35 kg/m2,the probability of pregnancy was 26% lower, an in the case of a wo-man with a BMI of 40 kg/m2, it was 43% lower compared with wo-men with a BMI between 21 and 29 kg/m2 (van der Steeg et al.,2008). In the Finnish cohort study mentioned above PCOS symp-toms and obesity, but not overweight, had independent and ad-verse effects on fecundability, and women suffering from bothobesity and PCOS symptoms experienced the lowest fecundability(FR = 0.33, 95% CI 0.16–0.68) (Koivunen et al., 2008).
Obesity and insulin resistance appear also to have independentroles in the etiology of early pregnancy losses (Hamilton-Fairleyet al., 1992; Jakubowicz et al., 2004; Wang et al., 2002). Obesityper se seems to be an independent risk factor for spontaneous abor-tion after infertility treatments (Bellver and Pellicer, 2004; Bruniet al., 2003; Lashen et al., 2004) and has been shown to increasespontaneous abortion rates in the general population. A recentmeta-analysis suggested an increased risk of miscarriage in womenwith a BMI of P25 kg/m2 (odd ratio 1.67, 95% CI 1.25–2.25) (Metw-ally et al., 2008). Some data even suggest that elevated spontane-ous abortion risk is connected only to obesity and not to thediagnosis of PCOS. On the other hand, a large retrospective studyon 1293 women undergoing IVF failed to confirm an associationbetween BMI and rates of clinical pregnancy or live birth (Dokraset al., 2006). In the previous cited population-based cohort study,neither obesity nor even abdominal obesity increased the sponta-neous abortion risk (Koivunen et al., 2008), and a study conductedin women treated by in vitro fertilization, including also very early
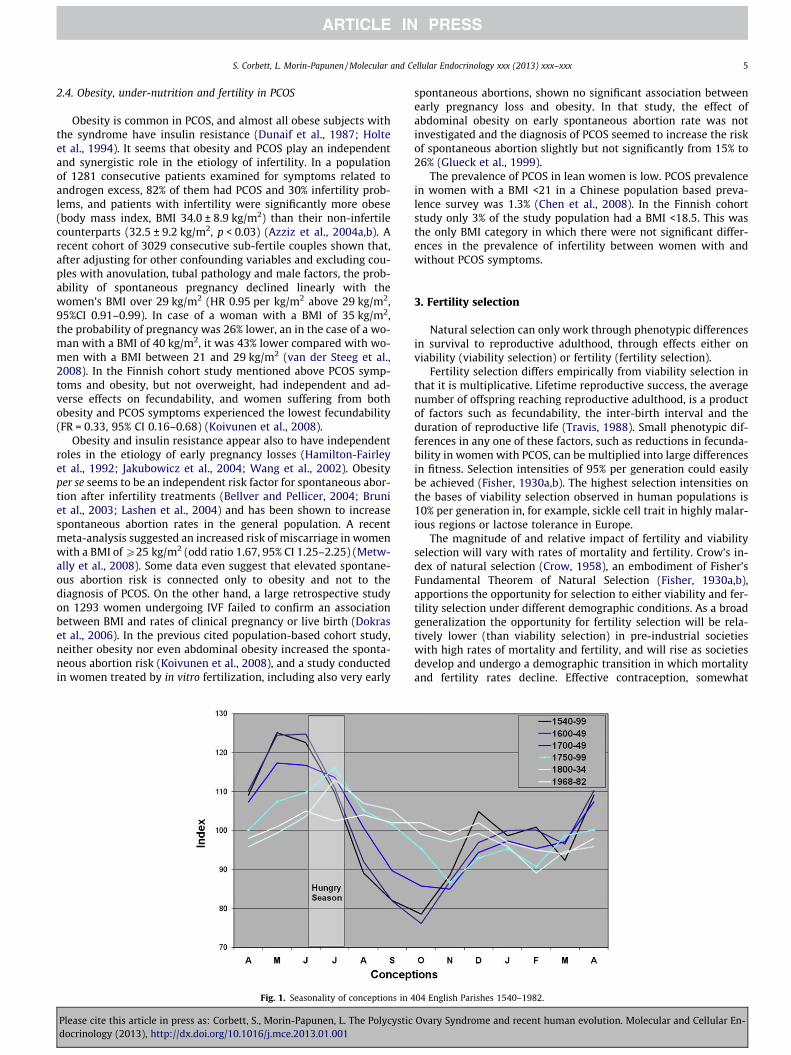
Fig. 1. Seasonality of conceptions in 4
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
spontaneous abortions, shown no significant association betweenearly pregnancy loss and obesity. In that study, the effect ofabdominal obesity on early spontaneous abortion rate was notinvestigated and the diagnosis of PCOS seemed to increase the riskof spontaneous abortion slightly but not significantly from 15% to26% (Glueck et al., 1999).
The prevalence of PCOS in lean women is low. PCOS prevalencein women with a BMI <21 in a Chinese population based preva-lence survey was 1.3% (Chen et al., 2008). In the Finnish cohortstudy only 3% of the study population had a BMI <18.5. This wasthe only BMI category in which there were not significant differ-ences in the prevalence of infertility between women with andwithout PCOS symptoms.
3. Fertility selection
Natural selection can only work through phenotypic differencesin survival to reproductive adulthood, through effects either onviability (viability selection) or fertility (fertility selection).
Fertility selection differs empirically from viability selection inthat it is multiplicative. Lifetime reproductive success, the averagenumber of offspring reaching reproductive adulthood, is a productof factors such as fecundability, the inter-birth interval and theduration of reproductive life (Travis, 1988). Small phenotypic dif-ferences in any one of these factors, such as reductions in fecunda-bility in women with PCOS, can be multiplied into large differencesin fitness. Selection intensities of 95% per generation could easilybe achieved (Fisher, 1930a,b). The highest selection intensities onthe bases of viability selection observed in human populations is10% per generation in, for example, sickle cell trait in highly malar-ious regions or lactose tolerance in Europe.
The magnitude of and relative impact of fertility and viabilityselection will vary with rates of mortality and fertility. Crow’s in-dex of natural selection (Crow, 1958), an embodiment of Fisher’sFundamental Theorem of Natural Selection (Fisher, 1930a,b),apportions the opportunity for selection to either viability and fer-tility selection under different demographic conditions. As a broadgeneralization the opportunity for fertility selection will be rela-tively lower (than viability selection) in pre-industrial societieswith high rates of mortality and fertility, and will rise as societiesdevelop and undergo a demographic transition in which mortalityand fertility rates decline. Effective contraception, somewhat
04 English Parishes 1540–1982.
Ovary Syndrome and recent human evolution. Molecular and Cellular En-

6 S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx
counter-intuitively, does not automatically reduce the potential forfertility selection as the theoretical potential for fertility selectionactually rises as the average number of births per woman falls,all other factors being equal (Crow, 1958).
The conversion of this potential for fertility selection into a real-ity is more likely in the context of the unprecedented change to hu-man reproductive ecology which has accompanied modernizationand development. Two inevitable consequences of developmentin particular, a reduction in seasonal under-nutrition and in theamplitude of birth seasonality, and a reduction in the total fertilityrate, have the potential to enhance or alter previous patterns of fer-tility selection (Crow, 1958; Vitzhum, 2009).
The hypotheses under consideration here imply that fertilityselection for the PCOS phenotype acted in one of two ways.Hypotheses 8 (Eggers et al., 2007) and 11 (Azziz et al., 2010) sug-gest that any negative impact of the PCOS phenotype has been inconstant balance with a number of other factors. All other hypoth-eses assume that before modernization and improvements innutrition there was likely to be net positive fertility selection forthe PCOS genotype, and negative selection thereafter.
4. Nutritional history and reproductive ecology of Homo sapiens
The fossil record suggests that in the cool and dry Pleistoceneepoch, beginning about two million years ago, the hominid dietchanged from being predominantly vegetarian to one based largelyon meat and animal products (Cordain et al., 2005; McMichael,2001).
There then occurred a gradual shift, beginning around12,000 years ago, from the consumption of root plants, wild pulses,various nuts and fruit and of hoofed mammals (gazelle, antelopeand deer) to cultivated wheat and barley and domesticated sheep,goats, cattle, and pigs (Larsen, 2000). Driven variously by popula-tion increase, megafaunal extinction and climatic change, agricul-ture emerged independently over five millennia in fivecontinents (Cavalli-Sforza and Bodmer, 1971; Flannery, 1994).
In almost all pre-industrial agrarian societies, chronic and espe-cially seasonal food shortage, punctuated by famine, was the nor-mal condition (Fogel, 2004). Obesity was rare, average calorieintake was low, and food quality was poor. Average daily caloriesper capita in England and France fell short of 2400 kcal/day, thecurrent FAO recommended minimum energy intake, until 1850.The energy intake of the typical diet in France in the eighteenthcentury was equivalent to that in present day Ethiopia, and in Eng-land in 1850 to that of rural India today (Fogel, 2004).
A striking characteristic of these pre-industrial societies was theseasonality of food availability, with a hunger season of one tothree months duration occurring before the harvest. This hungerseason was associated seasonal with reduction in the number ofovulatory cycles and conceptions. The amplitude of these seasonalvariations in rates of conception reduced over centuries as systemsfor food distribution and storage improved, as seen in data fromfamily reconstruction of demographic data from 404 English par-ishes between 1542 and 1834 (Wrigley and Schofield, 1981). Dataon English birth seasonality between 1968 and 1982 (Lam and Mir-on, 1994) have been added for comparison (Fig. 1).
Beginning in Europe in the eighteenth century improvements inagriculture, transport and food distribution unshackled cycles ofmortality and fertility from the price of grain (Fogel, 2004; Wrigleyand Schofield, 1981). Improved nutrition followed gradually innineteenth-century Europe, but increased rapidly and on a globalscale after World War II, with a 25% increase—550 calories per capi-ta—between 1961 and 2003. While these increases have reduced theprevalence of chronic energy deficiency they have also led tounprecedented levels of obesity in children and adults in both devel-
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
oped and developing countries. Between 1961 and 1999, and againsta background of declining per capita energy expenditure, the num-ber of countries consuming a per capita calorie intake greater than3,200 calories per day increased from 7 to 31 and the number of peo-ple from 110 million to 1 billion (Schmidhuber and Shetty, 2004).
5. Not necessarily a thrifty gene
Evolutionary approaches to PCOS and related common chronicconditions have assumed that ancestral traits, more attuned tothe ecological conditions of hunter gatherers in the late Pleisto-cene, leave us vulnerable to disease in the wake of radical and rel-atively recent change in selective pressures, particularly to diet andto average levels of energy expenditure.
Perhaps the most influential account of this kind has beenJames Neel’s thrifty gene hypothesis, which proposed that meta-bolic thrift and resistance to the action of insulin were appropriatemetabolic adaptations to the seasonality and uncertainty of thefood supply in the past (Neel, 1962, 1999). There are numerous cri-tiques of this hypothesis, including an admission by Neel himselfthat the idea had probably outlived its usefulness (Neel, 1999).Thrift may not necessarily be a solely a metabolic characteristic(Prentice et al., 2005), but could equally be linked genetically toadipogenesis, an ability to switch off non-essential energy expen-diture, slothfulness, gluttony, and other forms of energy storingor saving behaviours. Any of these factors would enhance the flex-ibility with which an organism responded to changing environ-mental circumstances. Despite this, the concept of metabolicthrift and metabolic efficiency linked to insulin resistance pervadesevolutionary thinking in relation to PCOS. (Hypotheses 1,2, 7(b)and 11(b)).
But is insulin resistance, a hallmark of both PCOS and Type 2Diabetes, thrifty? The evidence that insulin resistance itself in-creases either energy storage (as fat) or energy efficiency is incon-sistent, perhaps because the associated hyperinsulinaemia canboth increase fat storage and cause central (hypothalamic) sup-pression of appetite(Porte, 2006). Evidence from longitudinal stud-ies in adult Pima Indians (Swinbum et al., 1991). Caucasians,Mexican Americans, Creoles, Chinese, Asian Indians (Hodge et al.,1996) and in pregnant women (Catalano, 1999) suggest that themost insulin-sensitive (the least insulin-resistant) individuals andthose with low insulin levels may have the greater potential forweight gain. A recent meta-analysis of glucose regulation andchanges in weight showed that for people without diabetes therelationship was complex and depended on factors such as age,sex, race and methods of measurement (Chiu et al., 2010).
Similarly, it is not at all clear that the differences in post-pran-dial thermogenesis seen in both Type 2 diabetes and PCOS, (Robin-son et al., 1992; Robinson and Johnston, 1995) and invoked inhypotheses 1 and 2, represent a true energy saving or an increasein fat storage (Cooney and Storlein, 1994).
A somewhat obvious corollary of the thrifty gene hypothesis isthe existence of a non-thrifty (Allen and Cheer, 1996), insulin sen-sitive or energetically profligate (Prentice et al., 2005) phenotype.Furthermore this phenotype is likely to be a more recent adapta-tion to changed nutritional and environmental circumstances (Al-len and Cheer, 1996). This perspective has received surprisinglylittle attention in the literature on either Type 2 Diabetes or PCOS.Prentice suggest that evolutionary benefits may accrue from traitswhich demand energy profligacy, particularly in environmentswith a secure food supply; enhanced metabolic control, thermo-regulatory thermogenesis, quenching oxidative stress and the con-centration of nutrient deficient diets. There may also, under somecircumstances, be advantages for fertility.
The Fertility First (Corbett et al., 2009) (Hypothesis 10), FoodDeprivation and Refeeding (Shaw and Elton, 2008) (Hypotheses
Ovary Syndrome and recent human evolution. Molecular and Cellular En-

S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx 7
9(a) and 9(c) hypotheses all suggest that the primary evolutionaryadvantage of an insulin resistant phenotype comes not from anassociation with energy storage or efficiency, but from its role insustaining or reinvigorating ovulation under low energy condi-tions. The relatively higher levels of glucose and insulin associatedwith the PCOS phenotype, even under low energy conditions,would help to sustain ovulation during the annual hungry season,or to presage an earlier return to menses in women with lacta-tional amenorrhoea (Shaw and Elton, 2008). Support for this ideacomes from longitudinal studies of young women with anorexianervosa with nutritional amenorrhoea (Dei et al., 2008) and fromToba women in Northern Argentina with lactational amenorrhoea.(Valeggia and Ellison, 2009) In both groups insulin levels predict areturn to menses.
Vitzthum (2001) has advanced a Flexible Response Model of hu-man reproduction which considers the specific conditions underwhich a reduction in reproductive effort decreases or increasesDarwinian fitness. She suggests that there are two reproductivestrategies in uncertain nutritional environments – Obligates andDelayers – and poses the question;
. . .under what conditions does it make sense to ovulate no matterwhat environmental conditions exist (a phenotype of obligatereproductive investment at every opportunity). Or, when is it betterto adjust functioning according to the quality of the environment?. . .In other words, when would a delayer have the selective advan-tage over the obligates.
The reproductive consequences of PCOS/insulin resistant, andinsulin sensitive phenotypes therefore have many of the character-istics of these putative obligate and delayer reproductive strategiesrespectively, and selection for one or the other may depend on theexact nutritional circumstances of a population.
6. Hyperandrogenism, PCOS and human evolution
Hyperandrogenism in women, whether linked to PCOS or not,has been hypothesised to confer selective advantage by threemechanisms;
6.1. Direct effects on growth, behaviour and immune function
Witchel et al. (1997) proposed that elevated adrenal androgensseen in heterozygote carriers of 21-hydroxylase deficiency wouldpresage the early onset of puberty, and assertive behaviour. Shealso suggested that the brisk cortisol response she observed inthese subjects would enable a rapid return to homeostasis in re-sponse to infectious, inflammatory, or other environmental stres-ses and may protect from inappropriate immune responses.
Villuendas et al. (2002) and Escobar-Morreale et al. (2005) havebuilt on this insight to develop an evolutionary hypothesis forPCOS which posits that an interplay between genes for insulinsecretion and resistance, pro-inflammatory genes, and genes forandrogen synthesis, regulation and action, acted in concert innutritionally challenged populations to improve survival byimproving defences against infection and more efficiently storingenergy as fat for lean times.
6.2. Fewer offspring concentrates maternal investment
The assertion that relative infertility, attributed to hyperan-drogenism, concentrates maternal investment in fewer offspring,thereby enhancing the probability of survival is repeated in a num-ber hypotheses considered here (Azziz et al., 2010; Escobar-Morre-ale et al., 2005). The evidence in humans that large family sizereduces, and conversely that small family size improves, survival
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
is mixed (Lawson et al., 2012). These studies are confounded byfactors such as maternal health and level of education, and replace-ment or insurance phenomena whereby early or anticipated infantdeaths are compensated for by additional births. A recent studywhich attempted to address these sources of confounding con-cluded that in the context of ancestral, pre-industrial or developingworld populations with high extrinsic mortality risks from infec-tious diseases (Black, 2010), limiting fertility as a strategy to in-crease levels of parental investment is more likely to reducereproductive success (Lawson et al., 2012).
6.3. Effects of maternal androgens on infant development
Female rhesus monkeys exposed prenatally to excess androgenshad as adults a PCOS-like phenotype including hyperinsulinaemicandrogen excess, polycystic ovaries, elevated levels of circulatingLH, dyslipidemia, visceral adiposity, and a reduced ovulation rateresponsive to insulin sensitization (Abbott et al., 1998, 2002). The‘‘prenatal androgen excess’’ hypothesis as to the developmentalorigins of PCOS had its origins in these experiments, although theplausibility of this idea has been questioned because unborn girlsare thought to be protected from excess maternal androgens byabundant placental aromatase activity (de Zegher and Ibanez,2009). Findings from a longitudinal study (Hickey et al., 2009) of244 unselected girls recruited prenatally, along with their mothers,did not find that maternal androgens, within the normal range forpregnancy, directly program PCOS in the offspring.
In summary, the primacy of hyperandrogenism in the evolutionof the PCOS phenotype, particularly in ancestral populations whoin the main had BMI in the 17–22 range, is unclear. There seemsto be a consensus both that in PCOS hyperinsulinaemia affectsandrogen secretion and metabolism, rather than vice versa (Ehr-mann, 2005; Franks, 1995), and that defects in both insulin andandrogen secretion and action are likely to be involved in PCOSexpression (Ehrmann, 2005; Franks, 1995).
7. Evolutionary hypotheses and the Polycystic Ovary Syndrome
7.1. What is the object of explanation?
Most medical research focuses on how bodily mechanisms work,and how differences among individuals account for why somebecome ill. An evolutionary approach focuses on how bodily mech-anisms evolved, and how traits shared by most members of the spe-cies leave them vulnerable to a disease. The difference becomesclear only if the object of explanation and kind of explanationsought are described explicitly (Nesse, 2011).
The appropriate object of explanation is, in the case of PCOS, theparadox of a very common but also familial condition which causesinfertility and reduces lifetime reproductive success.
Alternatively the object of explanation could be viewed morebroadly as an apparent variation in the prevalence of fundamentalmetabolic and reproductive traits in the population, traits whichare likely to be normally distributed (Plomin et al., 2009). PCOSand related conditions can then be thought of as extreme manifes-tations of these traits occurring in an environment characterised byhigh energy intake and reduce levels of physical activity.
Two variants might be the ancestral insulin resistance pheno-type and a ‘‘non -thrifty’’ (Allen and Cheer, 1996) or profligate (Pre-ntice et al., 2005) phenotype characterised by greater insulinsensitivity. In PCOS the primary anomaly is likely to be either inthe central hypothalamic axis, involving increased LH secretion,overproduction of androgens in either ovary or adrenal, or insulinresistance and hyperinsulinaemia (Dunaif and Finegood, 1996;Escobar-Morreale et al., 2005; Sam and Dunaif, 2003).
Ovary Syndrome and recent human evolution. Molecular and Cellular En-

8 S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx
7.2. Has the object of explanation been influenced by evolution?
The essence of evolutionary change are phenotypic differencesin fertility and mortality prior to reproductive adulthood. In theOrigin of Species Darwin recognised that ‘‘the reproductive systemis eminently susceptible to changes in the conditions of life’’ (Dar-win, 1859).
The hypothesis that natural selection favours efficient energyutilization also has a long history (Lotka, 1922a,b). The confluenceof disturbances of energy homeostasis and reproduction and fertil-ity in PCOS make it likely that evolution, perhaps recent and rapidevolution (Corbett et al., 2009), has played a part in its appearancein populations in both the developed and developing world.
7.3. What kind of trait is the object of explanation?
7.3.1. Fixed traitsBarnett and Abbott (Barnett and Abbott, 2003) (hypothesis 4)
posit that a fixed universal trait of all primates which may haveestablished a vulnerability for follicular disruption and anovula-tion; the metabolic demands of a large and rapidly growing brainimmediately upon implantation required a prolongation of the per-iod of endometrial priming. The resulting compression of the pro-cess of follicular recruitment created this vulnerability.
7.3.2. Facultative traitsAbbott et al. (2002) (hypothesis 3) have proposed that PCOS is a
developmental disorder of ovarian androgen production origin forPCOS in which androgen excess in utero programmes the hypotha-lamic-pituitary control of LH, enhancing visceral fat distributionand predisposing to insulin resistance, anovulation and hyperan-drogenaemia. Shaw and Elton (2008) (hypothesis 9(c)) propose asimilar transgenerational effect of maternal androgens in whichfat storage and follicular readiness provide a fertility advantage un-der poor nutritional conditions. What is of great evolutionaryinterest about both of these hypotheses is whether, under low en-ergy conditions, these developmental phenomena constitute facul-tative adaptations – a predictive adaptive response – unwittingsignals to the unborn child that it is about to enter a harsh worldand which may result in characteristics which may help it growand in adulthood reproduce under these conditions (Batesonet al., 2004; Gluckman et al., 2005).
7.3.3. Human genesA large familial component to the aetiology of PCOS has long
been acknowledged (Cooper et al., 1968) and both dominant andmultigenic modes of inheritance have been proposed (Urbanek,2007). In twin studies the heritability of PCOS has been estimatedas 0.72 (Vink et al., 2009). Attempts to identify causative loci forPCOS in over 100 candidate genes involved in androgen biosynthe-sis and secretion, gonadotrophin secretion, the secretion and ac-tion of insulin and folliculogenesis has been largely disappointing(Escobar-Morreale et al., 2005; Urbanek, 2007). A systematic re-view of both candidate gene and genome wide approaches to thedetection of functional genetic polymorphisms in PCOS (Simoniet al., 2008) identified only one polymorphism – SNPs in exon 10of the FSH receptor gene which can be used as markers to predictdifferences in FSHR function and ovarian response to FSH – whichwas consistently associated with increased susceptibility to PCOS,although never strongly enough to correlate alone with suscepti-bility to the disease.
More recently genome-wide studies (Chen et al., 2011; Good-arzi et al., 2012; Welt et al., 2012) have identified one locus onchromosome 9, linked to the DENND1A gene, in which the minorallele confers an increased risk of PCOS in both Han Chinese,
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
Caucasian and Icelandic populations (Strauss et al., 2012). Otherloci linked to PCOS in Han populations (THADA and LHGCR) arenot found consistently in European populations.
Most hypotheses under consideration here (1, 2, 6, 8, 9, 10, 11)posit some advantage for the PCOS phenotype in ancestral popula-tions, which, have been ‘‘rendered detrimental by progress’’ (Neel,1962). A corollary of this idea is that genes or combinations ofgenes which determine PCOS risk are likely to be ancestral, ratherthan derived polymorphisms. This ancestral susceptibility model ofevolution (Di Rienzo and Hudson, 2005) has different implicationsfor the frequency spectrum of disease alleles and their geographicdistribution, which in turn determine the optimal strategies forfinding susceptibility variants.
Two of these hypotheses (9 and 10) suggest that selectionagainst the PCOS phenotype is likely to have accompanied theunprecedented changes to human nutrition, lifespan and fertilitywhich began in Europe in the 18th century. Signals of very recentpositive selection for alleles which have not yet reached fixationhave identified genes involved in fertilisation and metabolism(Voight et al., 2006) as targets of adaptive evolution.
7.4. What kind of explanation is sought?
There are the four kinds of questions that must be answered toprovide a full biologic explanation for any trait: mechanism, devel-opment, function, and phylogeny. The first two are proximateexplanations, the latter two, evolutionary (Tinbergen, 1963).
7.4.1. MechanismConfusion often arises when proximate mechanisms are in-
voked to test a proposal about the adaptive significance of a trait(Nesse, 2011). The phenotypic and clinical characteristics of PCOS,mostly observed in developed countries with a high prevalence ofobesity and over-nutrition, are invoked in a number of the hypoth-eses under consideration as having adaptive significance for ances-tral foraging or agrarian populations in which the prevalence ofoverweight or obesity was usually less than 5% and average BMIlevels ranged between 15 and 21.(Jenike, 2001; O’Dea et al.,1988) Seasonal food shortage, accompanied by 2–8 kg weight loss(Prentice and Cole, 1994), was the norm in these populations. Forexample, the adaptive significance of excess androgens such as in-creased muscle mass and strength invoked in Hypothesis 11 (Azzizet al., 2010)) or as an influence on fat patterning and sexual selec-tion (Hypothesis 5 (Kirchengast, 2005)) both assume that theexpression of hyper-androgenism would be similar in pre-indus-trial and more affluent populations.
7.4.2. DevelopmentEvidence from studies in rhesus monkeys and sheep that expo-
sure to excess androgen during key developmental windows re-sults, in adult life, in pathophysiological changes that closelyresemble the features of PCOS in women suggest that PCOS is adevelopmental disorder, primarily ovarian in origin, in whichandrogen hyper-secretion begins in foetal life, resulting in a pro-gramming of the reproductive and metabolic features of PCOS.(Franks and Berga, 2012; Shaw and Elton, 2008) Proponents ofthe predictive adaptive function of the PCOS phenotype extend thisargument by suggesting that this foetal programming may contrib-ute to optimal reproduction and development in an uncertainnutritional environment (Gluckman et al., 2005).
7.4.3. PhylogenyBarnett and Abbott have suggested that progressive develop-
ment of pre-implantation endometrial proliferation and a rapidly
Ovary Syndrome and recent human evolution. Molecular and Cellular En-

S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx 9
invading placenta across the Primate order, necessary to accom-modate differentiation and growth of an increasingly large foetalbrain, put the latter stages of ovarian follicle selection at risk ofexposure to excessive pituitary secretion of Luteinising Hormone.This excessive secretion of LH during follicle selection could resultin impaired follicle development, excessive ovarian androgensecretion and ovulation failure. (Hypothesis 3, (Barnett and Abbott,2003)).
7.4.4. FunctionLife history traits such as growth rates, age at menarche and
menopause, number and size of offspring, and lifespan are compo-nents of reproductive success which are under strong continualselection, (Stearns, 1992) and women with PCOS have been shownto have delayed menarche (Sadrzadeh et al., 2003) and menopause.(Dahlgren et al., 1992) Hypothesis 6 (Gleischer and Barad, 2006)suggests that the PCOS phenotype may be associated with in-creased ovarian reserve and women with PCOS undergoing fertilitytreatment have sustained oocyte counts with advancing age com-pared to women with infertility due to fallopian tube obstruction(Mellembakken et al., 2011). In preindustrial societies practicingnatural fertility schedules, age specific fertility rates were highestin younger women (Henry, 1961). It is unclear whether in thesepopulations maintenance of ovarian reserve would augment life-time reproductive success.
Both kin selection (Hypothesis 8 (Eggers et al., 2007)), wherebyinfertile mothers contribute to the survival of related children, andsexual selection (Hypothesis 5 (Kirchengast, 2005)) whereby fatpatterning contribute to sexual allure, have been invoked.
The high global prevalence of PCOS suggests that the underlyinggenes represent a standing polygenic variation in human popula-tions (Pritchard and Di Rienzo, 2010). The alignment in large epide-miological studies of average weight and the weight at whichmaximum ovarian fertility occurs (Rich-Edwards et al., 2002) sug-gests that the evolutionary mechanism of fertility selection is ableto tailor the fecundability of a population to average energetic con-ditions. Alterations in the relative proportion in a population of aPCOS/ insulin resistant and an insulin sensitive phenotype maybe a means of doing this (Corbett et al., 2009).
7.5. Are all viable hypotheses considered ?
Hypotheses specific to PCOS have been considered, but need tobe viewed in the context of the slew of hypotheses proposed inrelation to the evolutionary origins of Type 2 Diabetes (Allen andCheer, 1996; Colagiuri and Brand Miller, 2002; Prentice et al.,2005; Speakman, 2006).
7.5.1. Are there different vulnerabilities to PCOS in differentpopulations?
All human populations seem to be at risk of PCOS once theyachieve permanent food security. Informative variations in diseaseprevalence between populations are rare, because, until recently,there was little agreement about an appropriate case definition(The Rotterdam ESHRE/ASRM PCOS consensus, 2004). Populationbased estimates of the prevalence of a diagnosis of PCOS fromten population based studies from nine countries (Asuncionet al., 2000; Azziz et al., 2004a,b; Chen et al., 2008; Diamanti-Kand-arakis et al., 1999; Goodarzi et al., 2005; Kumarapeli et al., 2008;Lindholm et al., 2008; March et al., 2010; Moran et al., 2010; San-chon et al., 2012; Yildiz et al., 2012) show a prevalence varying be-tween 2% and 13%. The lowest prevalence was in Southern China,and here the mean BMI of the study population was 21, consider-ably less than in all other studies.
There is no evidence of a relative immunity to PCOS in Europidpopulations which might parallel that seen with Type 2 diabetes
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
(Diamond, 2003), although at present such a comparison is notpossible on the basis of published information.
7.5.2. What categories of explanation are under consideration?7.5.2.1. Mismatch. Hypotheses 9 and 10 imply that PCOS in modernpopulations is an example of phylogenetic inertia, a slowness ofadaptation to changing environmental conditions, and in particulara slowness to adapt to a diet of cereals rather than meat and fat.
7.5.2.2. Constraints. Hypotheses 5,6,7(a), 8 and 11(a) in Table 1 as-sume that characteristics of PCOS observed in contemporary pop-ulations, such as androgenisation or increased muscle mass willbe an object of explanation. Again it is informative to imagine con-ditions before and after modernisation and improved nutrition. Be-fore modernisation with obesity a rarity and average BMI in the18–20 range, the expression of many of these phenotypic featuresis likely to have been be uncommon and subtle, and in the samepositive direction as fertility selection for PCOS. After modernisa-tion these putative non reproductive phenotypic features, althoughexpressed more strongly, would be unlikely to be able to match thestrong negative selection intensity associated with impairments tofertility. Kin selection (Eggers et al., 2007) as described would onlybe relevant after modernisation and again would be unlikely to bea match for the strong negative fertility selection.
The intensity of selection on the basis of fertility, and the conse-quent speed at which change could occur in the prevalence ofgenes involved in fundamental metabolic and reproductive pro-cesses opens up avenues of explanation for hitherto poorly under-stood variations in the occurrence of diabetes and cardiovasculardisease in human populations. Examples include the apparentimmunity of Europids to Type 2 diabetes, the low levels of cardio-vascular disease in southern Europe – the so called Mediterraneanparadox (de Lorgeril et al., 2002) – and in Japan, countries whichotherwise have a high prevalence of cardiovascular risk factors.
7.5.2.3. Trading off reproductive success at the expense of health.Trade-offs occur when a change in one trait which increases fitnessis connected to a change in another trait which decreases fitness.The concept of a genetically based trade-off between investmentof energetic resources in growth and reproduction forms the basisof the Disposable Soma Theory of Health and Ageing(Holliday,1989; Shanley and Kirkwood, 2000). The later life cardiovascularsequelae of insulin resistance, PCOS and T2D could be viewed asthe life-shortening consequences of this disinvestment in somato sustain reproduction, although these consequences almost allconfined to the post reproductive years.
7.6. Could multiple explanations be correct?
Given the central role that nutrition and energetics have playedin sustaining human fertility and fuelling demographic expansion(Ellison, 2003; Leonard and Ulijaszek, 2002), the concurrence ofreproductive and metabolic features in a very common (PCOS)phenotype may be an important clue as to how our speciesadapted to changes in reproductive ecology, in diet and the ener-getic demands of food gathering, in altitude, climate and tempera-ture, and in the burden of infectious illness in their traverse of fivecontinents over 200,000 years. Wells has suggested that the devel-opmental plasticity of the human metabolic phenotype bufferedhuman populations against the need for genetic commitment toa particular genotype (Wells, 2010). For Type 2 Diabetes, there isgood evidence of this ‘‘genetic commitment’’ to variations in met-abolic phenotype (Chiu et al., 2010) with demonstrable ethnicitybased differences in prevalence, particularly with increasing aver-age weight. For PCOS this commitment is suggestive but not con-clusive (see Section 7.5.1).
Ovary Syndrome and recent human evolution. Molecular and Cellular En-

10 S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx
The hypotheses considered here take two basic approaches toexplain the apparent paradox of PCOS occurrence. The first group(Hypotheses1–8)) speculate as to how one or all of the phenotypicfeatures of PCOS observed in contemporary populations could actindividually or in concert to attain to a survival or fertility advan-tage in ancient populations. The second group (Hypotheses 9–11)make explicit attempts to link the metabolic characteristics ofthe PCOS phenotype to the exquisite sensitivity of ovulation in hu-mans to the availability of metabolic fuel. These latter hypothesesprovide an account of the potential evolutionary advantage of thePCOS phenotype which is more closely aligned with human nutri-tional, reproductive and demographic history.
7.7. How can these hypotheses be tested?
Ultimately these hypotheses will need to survive, like all scien-tific postulates, the rigours of falsification. Some avenues for futureresearch into the evolutionary origins of PCOS could include;
7.7.1. Understanding variation in PCOS susceptibilityA corollary of many of these hypotheses is that significant var-
iation in susceptibility to PCOS should be evident in human popu-lations, but this is not well described. Inter-ethnic comparisons ofPCOS prevalence which avoid selection bias, which use standarddiagnostic definitions, and which are able to adjust for age andobesity are rare, but are necessary to understand the extent of thisvariation, and in particular whether the variation mirrors that ob-served in Type 2 diabetes (Chiu et al., 2011; Corbett et al., 2009).
The recent identification of polymorphisms linked to the DEN-ND1A gene which confer a risk of PCOS in Han Chinese and Euro-pean populations provides opportunities to assess the impact ofthese variants on adaptive evolution (Helgason et al., 2007).
7.7.2. Establishing the existence and intensity of positive or negativefertility selection for an insulin resistant/PCOS phenotype in humanpopulations
Currently large cohorts such as the Million Women study (UK),the Nurses’ Health Study (US) and the 45 and Up cohort (Australia)contain enough women who have both completed their reproduc-tive lives and who have a history of diabetes and/or PCOS to esti-mate selection intensities of 3–5% or greater, sufficient for thehypothesis of rapid and recent evolution to be falsified.
7.7.3. Investigating the mechanisms and circumstances by whichinsulin resistance or sensitivity can optimise reproductive outcomes
Young women with anorexia nervosa have nutritional amenor-rhoea, and menstrual resumption is an important milestone inrecovery. In some senses this recovery simulates in a closely ob-served clinical setting the physiological responses to re-feedingwhich are characteristic of many contemporary hunter gathererand agrarian populations subject to seasonal under-nutrition.
Longitudinal studies of metabolic phenotype and lifetime repro-ductive success in populations in different environments and withdiffering agricultural systems (Koivunen et al., 2008; Moore et al.,2001; Vallegia and Ellison, 2004) may help to resolve the relativeadvantages and metabolic characteristics of obligate and delayerreproductive strategies.
8. Conclusion
The emerging discipline of evolutionary medicine can providesome important insights into causes and patterns of disease occur-rence. Firstly, organisms are not perfectly formed machines but abundle of compromises shaped by natural selection to maximizereproduction, not health. Secondly, much disease arises from the
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
mismatch of our bodies to modern environments and thirdly theassumption that common heritable diseases are caused by a fewdefective genes is usually incorrect (Stearns et al., 2010).
Understanding the evolutionary origins of PCOS has a particularpoignancy for global efforts to reduce the vulnerability of thosepopulations to emerging global epidemics of diabetes and cardio-vascular disease in both the developed and the developing world.As some of the hypotheses reviewed here imply (Corbett et al.,2009; Shaw and Elton, 2008) the elimination of under-nutritionmay, by both relaxing selection pressure for and increasing selec-tion pressure against a PCOS/insulin resistant phenotype, reducethe prevalence of the underlying genotypes within decades ratherthan the centuries or millennia of evolutionary time.
References
Abbott, D., Dumesic, D., Eisner, J., Colman, R., Kemnitz, J., 1998. Insights into thedevelopment of PCOS from studies of prenatally androgenized female rhesusmonkeys. Trends Endocr Metab. 9, 62–67.
Abbott, D., Dumesic, D.A., Franks, S., 2002. Developmental origin of polycystic ovarysyndrome an hypothesis. J. Endocrinol. 174, 1–5.
Allen, J., Cheer, S., 1996. The non-thrifty genotype. Curr. Anthropol. 37, 831–842.Asuncion, M., Calvo, R., San Millan, J., Sancho, J., Avila, S., Escobar-Morreale, H., 2000.
A prospective study of the prevalence of the polycystic ovary syndrome inunselected Caucasian women from Spain. J. Clin. End. Metab. 85, 2434–2438.
Azziz, R., Dumesic, D., Goodarzi, M., 2010. Polycystic ovary syndrome: an ancientdisorder? Fertil. Steril.
Azziz, R., Sanchez, L.A., Knochenhauer, E.S., Moran, C., Lazenby, J., Stephens, K.C.,Taylor, K., Boots, L.R., 2004a. Androgen excess in women: experience with over1000 consecutive patients. J. Clin. Endocrinol. Metab. 89, 453–462.
Azziz, R., Woods, K.S., Reyna, R., Key, T.J., Knochenhauer, E.S., Yildiz, B.O., 2004b. Theprevalence and features of the polycystic ovary syndrome in an unselectedpopulation. J. Clin. Endocrinol. Metab. 89, 2745–2749.
Balen, A., 1993. Hypersecretion of luteinizing hormone and the polycystic ovarysyndrome. Hum. Reprod. 8, 123–128.
Balen, A., Michelmore, K., 2002. What is polycystic ovary syndrome? Are nationalviews important? Hum. Reprod. 17, 2219–2227.
Balen, A.B., Rutherford, A.J., 2007. Managing anovulatory infertility and polycysticovary syndrome. BMJ 335, 608–611.
Barnett, D., Abbott, D., 2003. Reproductive adaptations to a large-brained fetus opena vulnerability to anovulation similar to polycystic ovary syndrome. Am. J. Hum.Biol. 15, 296–319.
Bateson, P., Barker, D., Clutton-Brock, T., Deb, D., D’Udine, B., Foley, R., Gluckman, P.,Godfrey, K., Kirkwood, T., Lahr, M., McNamara, J., Neil, B., Metcalfe, N.,Monaghan, P., Spencer, H., Sultan, S., 2004. Developmental plasticity andhuman health. Nature 439, 419–421.
Bellver, J., Pellicer, A., 2004. Impact of obesity on spontaneous abortion. Am. J.Obstet. Gynecol. 190, 293–294.
Black, R.E.e.a., 2010. Global, regional and national causes of child mortality in 2008:a systematic analysis. Lancet 375, 1969–1987.
Bodmer, W., 1964. Differential fertility in population genetics models. Genetics 51,411–424.
Broekmans, F., Knauff, E., Valkenburg, O., Laven, J., Eijkemans, M., Fauser, B., 2006.PCOS according to the Rotterdam consensus criteria: change in prevalenceamong WHO-II anovulation and association with metabolic factors. BJOG 113,1210–1217.
Bruni, V., Dei, M., Pontelo, V., Vangelisti, P., 2003. The management of polycysticovary syndrome. Ann. N. Y. Acad. Sci. 997, 307–321.
Byars, S., Ewbank, D., Govindarajuc, D., Stearns, S., 2010. Natural selection in acontemporary human population. PNAS 170, 1787–1792.
Catalano, P., 1999. Pregnancy and lactation in relation to range of acceptablecarbohydrate and fat intake. Eur. J. Clin. Nutr. 53 (suppl. 1), S124–S135.
Cavalli-Sforza, L., Bodmer, W., 1971. The Genetics of Human Populations. TheGenetics of Human Populations. W Freeman and Co., San Francisco, pp. 184.
Chen, X., Yang, D., Mo, Y., Li, L., Chen, Y., Huang, Y., 2008. Prevalence of polycysticovary syndrome in unselected women from southern China. Eur. J. Obstet.Gynecol. Reprod. Biol. 139, 59.
Chen, Z.-J., Zhao, H., He, L., Shi, Y., Qin, Y., Shi, Y., Li, Z., You, L., Zhao, J., Liu, J., Liang,X., Zhao, X., Zhao, J., Sun, Y., Zhang, B., Jiang, H., Zhao, D., Bian, Y., Gao, X., Geng,L., Li, Y., Zhu, D., Sun, X., Xu, J.-e., Hao, C., Ren, C.-e., Zhang, Y., Chen, S., Zhang, W.,Yang, A., Yan, J., Li, Y., Ma, J., Zhao, Y., 2011. Genome-wide association studyidentifies susceptibility loci for polycystic ovary syndrome on chromosome2p16.3, 2p21 and 9q33.3. Nat. Genet. 43, 55–59.
Chiu, C., Wray, L., Beverly, E., 2010. Relationship of glucose regulation to changes inweight: a systematic review and guide to future research. Diabetes Metab. Res.Rev. 26, 323–335.
Chiu, M., Austin, P., Manuel, D., Shah, B., Tu, J., 2011. Deriving ethnic-specific BMIcutoff points for assessing diabetes risk. Diabetes Care 34, 1741–1748.
Colagiuri, S., Brand Miller, J., 2002. The ‘carnivore connection’-evolutionary aspectsof insulin resistance. Eur. J. Clin. Nutr. 56, 30–35.
Ovary Syndrome and recent human evolution. Molecular and Cellular En-

S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx 11
Cooney, C., Storlein, L., 1994. Insulin action, thermogenesis and obesity. BaillieresClin. Endocrinol. Metab. 8, 481–507.
Cooper, H., Spellacy, W., Prem, K., Cohen, W., 1968. Hereditary factors in Stein-Leventhal syndrome. Am. J. Obstet. Gynecol. 100, 371–387.
Corbett, S., McMichael, A., Prentice, A., 2009. Type 2 diabetes, cardiovascular diseaseand the evolutionary paradox of the polycystic ovary syndrome: a fertility firsthypothesis. Am. J. Hum. Biol. 21, 587–598.
Cordain, L., Boyd Eaton, S., Sebastian, A., Mann, N., Lindeberg, S., Watkins, B.,O’Keefe, J., Brand-Miller, J., 2005. Origins and evolution of the Western diet:health implications for the 21st century. Am. J. Clin. Nutr. 81, 341–354.
Crow, J., 1958. Some possibilities for measuring selection intensities in man. Hum.Biol. 30, 1–13.
Dahlgren, E., Johansson, S., Lindstedt, G., Knutsson, F., Oden, A., Janson, P., Mattson,L., Crona, N., Lundberg, P., 1992. Women with polycystic ovary syndrome wedgeresected in 1956 to 1965: a long-term follow-up focusing on natural history andcirculating hormones. Fertil. Steril. 57, 505–513.
Darwin, C., 1859. The Origin of Species, Paperback Edition, 1968 ed. Penguin Books,New York.
de Lorgeril, M., Salen, P., Paillard, P., Laporte, F., Boucher, F., de Leiris, J., 2002.Mediterranean diet and the French paradox: two distinct biogeographicconcepts for one consolidated scientific theory on the role of nutrition incoronary heart disease. Cardiovasc. Res. 54, 503–515.
de Zegher, F., Ibanez, L., 2009. Early origins of polycystic ovary syndrome:hypotheses may change without notice. J. Clin. Endocrinol. Metab. 94, 3682–3685.
Dei, M., Seravalli, V., Bruni, V., Balzi, D., Pasqua, A., 2008. Predictors of recovery ofovarian function after weight gain in subjects with amenorrhea related torestrictive eating disorders. Gynecol. Endocrinol. 24, 459–464.
Dewailly, D., Catteau-Jonard, S., Reyss, A., Leroy, M., Pigny, P., 2006.Oligoanovulation with Polycystic Ovaries But Not Overt Hyperandrogenism. J.Clin. Endocrinol. Metab. 91, 3922–3927.
Di Rienzo, A., Hudson, R., 2005. An evolutionary framework for common diseases:the ancestral-susceptibility model. Trends Genet. 21, 596–601.
Diamanti-Kandarakis, E., Kouli, C.R., Bergiele, A.T., Filandra, F.A., Tsianateli, T.C.,Spina, G.G., Zapanti, E.D., Bartzis, M.I., 1999. A survey of the polycystic ovarysyndrome in the greek island of lesbos: hormonal and metabolic profile. J. Clin.Endocrinol. Metab. 84, 4006–4011.
Diamond, J., 2003. The double puzzle of diabetes. Nature 423, 599–602.Dokras, A., Baredziak, L., Blaine, J., Syrop, C.H., VanVoorhis, B.J., Sparks, A., 2006.
Obstetric outcomes after in vitro fertilization in obese and morbidly obesewomen. Obstet Gynecol. 108, 61–69.
Dunaif, A., Finegood, D., 1996. Beta-cell dysfunction independent of obesity and glucoseintolerance in the polycystic ovary syndrome. J. Clin. End. Metab., 942–947.
Dunaif, A., Graf, M., Mandeli, J., Laumas, V., Dobrjansky, A., 1987. Characterization ofgroups of hyperandrogenic women with acanthosis nigricans, impaired glucosetolerance, and/or hyperinsulinemia. J. Clin. Endocrinol. Metab. 65, 499–507.
Eggers, S., Hashimoto, D., Kirchengast, S., 2007. An evolutionary approach to explain thehigh frequency of the polycystic ovary syndrome. Anthropol. Anz. 65, 169–179.
Ehrmann, D.A., 2005. Polycystic ovary syndrome. N. Engl. J. Med. 352, 1223–1236.Ellison, P., 2003. Energetics and reproductive effort. Am. J. Hum. Biol. 15, 342–351.Escobar-Morreale, H.F., Luque-Ramírez, M., San Millán, J.L., 2005. The molecular-
genetic basis of functional hyperandrogenism and the polycystic ovarysyndrome. Endocr. Rev. 26, 251–282.
Fisher, R., 1930a. The Fundamental Theorem of Natural Selection. The GeneticalTheory of Natural Selection. Oxford University Press, Oxford, pp. 22–47.
Fisher, R., 1930b. The Inheritance of Human Fertility. Genetical Theory of NaturalSelection. Clarendon Press, Oxford, pp. 188–209.
Flannery, T., 1994. The Future Eaters: An Ecological History of the AustralasianLands and People. Grove Press, New York.
Fogel, R., 2004. The Escape from Hunger and Premature Mortality 1750–2100.Cambridge University Press.
Franks, S., 1995. Polycystic ovary syndrome. N. Engl. J. Med., 853–861.Franks, S., Berga, S.L., 2012. Does PCOS have developmental origins? Fertil. Steril. 97,
2–6.Franks, S., Gharani, M., Waterworth, D., Batty, S., White, D., Williamson, R.,
McCarthy, M., 1997. The genetic basis of polycystic ovary syndrome. Hum.Reprod. 12, 2641–2648.
Franks, S., McCarthy, M., 2004. Genetics of ovarian disorders: polycystic ovarysyndrome. Rev. End. Metab. Disorders 5, 69–76.
Gleischer, N., Barad, D., 2006. An evolutionary concept of polycystic ovarian disease:does evolution favour reproductive success over survival. Reprod. Biomed.Online 12, 587–589.
Gluckman, P., Hanson, M., Spencer, H., 2005. Predictive adaptive responses andhuman evolution. Trend Ecol. Evol. 20, 527–533.
Glueck, C., Wang, P., Fontaine, R., Sieve-Smith, L., Tracy, T., 1999. Plasminogenactivator inhibitor activity: an independent risk factor for the high miscarriagerate during pregnancy in women with polycystic ovary syndrome. Metabolism48, 1589–1595.
Goodarzi, M., Quiñones, M., Azziz, R., Rotter, J., Hsueh, W., Yang, H., 2005. Polycysticovary syndrome in Mexican–Americans: prevalence and association with theseverity of insulin resistance. Fertil. Steril. 84, 766–768.
Goodarzi, M.O., Jones, M.R., Li, X., Chua, A.K., Garcia, O.A., Chen, Y.-D.I., Krauss, R.M.,Rotter, J.I., Ankener, W., Legro, R.S., Azziz, R., Strauss, J.F., Dunaif, A., Urbanek, M.,2012. Replication of association of DENND1A and THADA variants withpolycystic ovary syndrome in European cohorts. J. Med. Genet. 49, 90–95.
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
Gould, S., 1985. Not necessarily a wing. Natur. History 94, 12–25.Gray, R., Wu, L., 2000. Subfertility and risk of spontaneous abortion. Am. J. Public
Health 90, 1452–1454.Hamilton-Fairley, D., Kiddy, D., Watson, H., Paterson, C., Franks, S., 1992. Association
of moderate obesity with a poor pregnancy outcome in women with polycysticovary syndrome treated with low dose gonadotrophin. Br. J. Obstet Gynaecol.99, 128–131.
Helgason, A., Pálsson, S., Thorleifsson, G., Struan, F.A., Emilsson, V., Gunnarsdottir, S.,Adeyemo, A., et al., 2007. Refining the impact of TCF7L2 gene variants on type 2diabetes and adaptive evolution. Nat. Genet. 39, 218–225.
Henry, L., 1961. Some data on natural fertility. Eugen. Quart. 8, 81–91.Hickey, M., Sloboda, D., Atkinson, H., Doherty, D., Franks, S., Norman, R., Newnham,
J., Hart, R., 2009. The relationship between maternal and umbilical cordandrogen levels and ovarian function in adolescence. A prospective cohortstudy. J. Clin. Endocrinol. Metab. 94, 3714–3720.
Hodge, A., Dowse, G.K., Alberti, K.G., Tuomilehto, J., Gareeboo, H., Zimmet, P.Z., 1996.Relationship of insulin resistance to weight gain in non-diabetic Asian Indian,Creole, and Chinese Mauritians. Metabolism 45, 627–633.
Holliday, R., 1989. Food reproduction and longevity: is the extended lifespan ofcalorie restricted animals an evolutionary adaptation. BioEssays 10, 125–127.
Holte, J., Bergh, T., Berne, C., Berglund, L., Lithell, H., 1994. Enhanced early insulinresponse to glucose in relation to insulin resistance in women with polycysticovary syndrome and normal glucose tolerance. J. Clin. Endocrinol. Metab. 78,1052–1058.
Homburg, R., Armar, N., Eshel, A., Adams, J., Jacobs, H., 1988. Influence of serumluteinising hormone concentrations on ovulation, conception, and earlypregnancy loss in polycystic ovary syndrome. BMJ 297, 1024–1026.
Hudecova, M., Holte, J., Olovsson, M., Sundstrom-Poromaa, I., 2009. Long-termfollow-up of patients with polycystic ovary syndrome: reproductive outcomeand ovarian reserve. Hum. Reprod. 24, 1176–1183.
Jakubowicz, D., Essah, P., Seppala, M., Jakubowicz, S., Baillargeon, J., Koistinen, R.,Nestler, J., 2004. Reduced serum glycodelin and insulin-like growth factor-binding protein-1 in women with polycystic ovary syndrome during firsttrimester of pregnancy. J. Clin. Endocrinol. Metab. 89, 833–839.
Jenike, M., 2001. Nutritional ecology; diet, physical activity and body size. In:Panter-Brick, C. et al. (Eds.), Hunter Gatherers. An Inter-DisciplinaryPerspective. Cambridge University Press, Cambridge, pp. 205–238.
Kirchengast, S., 2005. Evolutionary and medical aspects of body compositioncharacteristics in subfertile and infertile women. Acta Medica. Lituanica. 12,22–27.
Koivunen, R., Pouta, A., Franks, S., Martikainen, H., Sovio, U., Hartikainen, A.-L.,McCarthy, M., Ruokonen, A., Bloigu, A., Jarvelin, M., Morin-Papunen, L., 2008.Fecundability and spontaneous abortions in women with self-reported oligo-amenorrhea and/or hirsutism: Northern Finland Birth Cohort 1966 Study. Hum.Reprod. 23, 2134–2139.
Kumarapeli, V., Seneviratne, R.d.A., Wijeyaratne, C., Yapa, R., Dodampahala, S., 2008.A simple screening approach for assessing community prevalence andphenotype of polycystic ovary syndrome in a semiurban population in SriLanka. Am. J. Epidemiol. 168, 321–328.
Lam, D., Miron, J., 1994. Global patterns of seasonal variation in human fertility.Ann. NY Acad. Sci. 709, 9–28.
Larsen, C., 2000. Dietray reconstruction and nutritional assessment of past peoples:the bioanthropological record. In: Kiple, K., Ornelas, K. (Eds.), Cambridge WorldHistory of Food. Cambridge University Press, Cambridge, pp. 13–34.
Lashen, H., Fear, K., Sturdee, D., 2004. Obesity is associated with increased risk offirst trimester and recurrent miscarriage: matched case-control study. Hum.Reprod. 19, 1644–1646.
Lawson, D.W., Alvergne, A., Gibson, M.A., 2012. The life-history trade-off betweenfertility and child survival. Proc. R Soc. B: Biol. Sci..
Leonard, W., Ulijaszek, S., 2002. Energetics and evolution: an emerging researchdomain. Am. J. Hum. Biol. 14, 547–550.
Lindholm, Å., Andersson, L., Eliasson, M., Bixo, M., Sundström-Poromaa, I., 2008.Prevalence of symptoms associated with polycystic ovary syndrome. Int. J.Gynecol. Obstet. 102, 39.
Lotka, A., 1922a. Contribution to the energetics of evolution. Proc. Natl. Acad. Sci.USA 8, 147–151.
Lotka, A., 1922b. Natural selection as a physical principle. Proc. Natl. Acad. Sci. USA8, 151–154.
March, W., Moore, V., Willson, K., Phillips, D., Norman, R., Davies, M., 2010. Theprevalence of polycystic ovary syndrome in a community sample assessedunder contrasting diagnostic criteria. Hum. Reprod. 25, 544–551.
McMichael, A., 2001. Human Frontiers, Environments and Disease Past Patterns,Uncertain Futures. Cambridge University Press.
Mellembakken, J.R., Berga, S.L., Kilen, M., Tanbo, T.G., Åbyholm, T., Fedorcsák, P.,2011. Sustained fertility from 22 to 41 years of age in women with polycysticovarian syndrome. Hum. Reprod. 26, 2499–2504.
Metwally, M., Ong, K.J., Ledger, W.L., Li, T.C., 2008. Does high body mass indexincrease the risk of miscarriage after spontaneous and assisted conception? Ameta-analysis of the evidence. Fertil. Steril. 90, 714–726.
Moore, S., Halsall, I., Howarth, D., Poskitt, E., Prentice, A.M., 2001. Glucose, insulinand lipid metabolism in rural Gambians exposed to early malnutrition. DiabeticMed. 18, 646–653.
Moran, C., Tena, G., Moran, S., Ruiz, P., Reyna, R., Duque, X., 2010. Prevalence ofpolycystic ovary syndrome and related disorders in Mexican women. Gynecol.Obstet. Invest. 69, 274–280.
Ovary Syndrome and recent human evolution. Molecular and Cellular En-

12 S. Corbett, L. Morin-Papunen / Molecular and Cellular Endocrinology xxx (2013) xxx–xxx
Neel, J., 1962. Diabetes mellitus: a thrifty genotype rendered detrimental byprogress. Am. J. Hum. Genet. 14, 353–362.
Neel, J., 1999. The ‘‘thrifty genotype’’ in 1998. Nutr. Rev. 57, S1–S8.Nesse, R., 2011. Ten questions for evolutionary studies of disease vulnerability. Evol.
Appl. 4, 264–277.Nikolaou, D., Gilling-Smith, C., 2004. Early ovarian ageing: are women with
polycystic ovaries protected? Hum. Reprod. 19, 2175–2179, %R 10.1093/humrep/deh419.
O’Dea, K., Traianedes, K., Hopper, J.L., Larkins, R.G., 1988. Impaired glucosetolerance, hyperinsulinemia, and hypertriglyceridemia in Australianaborigines from the desert. Diabetes Care 11, 23–29.
Plomin, R., Haworth, C., Davis, O., 2009. Common disorders are quantitative traits.Nature 10, 872–879.
Porte, D., 2006. Central regulation of energy homeostasis: the key role of insulin.Diabetes 55, S155.
Prentice, A., Rayco-Solon, P., Moore, S.E., 2005. Insights from the developing world:thrifty genotypes and thrifty phenotypes. Proc. Nutr. Soc. 64, 153–161.
Prentice, A.M., Cole, T.J., 1994. Seasonal changes in growth and energy status in theThird World. Proc. Nutr. Soc. 53, 509–519.
Pritchard, J.K., Di Rienzo, A., 2010. Adaptation – not by sweeps alone. Nat. Rev.Genet. 11, 665–667.
Regan, L., Braude, P., Trembath, P., 1989. Influence of past reproductive performanceon risk of spontaneous abortion. BMJ 299, 541–545.
Rich-Edwards, J., Spiegelman, D., Garland, M., Hertzmark, E., Hunter, D.J., Colditz,G.A., Willett, W.C., Wand, H., Manson, J.E., 2002. Physical activity, body massindex, and ovulatory disorder infertility. Epidemiology 13, 184–190.
Robinson, S., Chan, S.P., Spacey, S., Anyaoku, V., Johnston, D.G., Franks, S., 1992. Postprandial thermogenesis is reduced in polycystic ovary syndrome and isassociated with increased insulin resistance. Clin. Endocrinol. 36, 537–543.
Robinson, S., Johnston, D., 1995. Advantage of diabetes? Nature 375, 640.Rodin, D., Bano, G., Bland, J.M., Taylor, K., Nussey, S.S., 1998. Polycystic Ovaries and
associated metabolic abnormalities in Indian subcontinent Asian women. Clin.Endocrinol. 1, 91–99.
Sadrzadeh, S., Klip, W.A.J., Broekmans, F.J.M., Schats, R., Willemsen, W.N.P., Burger,C.W., van Leeuwen, F.E., Lambalk, C.B., O.P.g., 2003. Birth weight and age atmenarche in patients with polycystic ovary syndrome or diminished ovarianreserve, in a retrospective cohort. Hum. Reprod. vol. 18, pp. 2225–2230, %R10.1093/humrep/deg409.
Sagle, M., Bishop, K., Ridley, N., Alexander, F., Michel, M., Bonney, R., Beard, R.,Franks, S., 1988. Recurrent early miscarriage and polycystic ovaries. BMJ 297,1027–1028.
Sam, S., Dunaif, S., 2003. Polycystic ovary syndrome: syndrome XX? TrendsEndocrinol. Metab. 14, 365–370.
Sanchon, R., Gambineri, A., Alpanes, M., Martinez-Garcia, M., Pasquali, R., Escobar-Morreale, H., 2012. Prevalence of functional disorders of androgen excess inunselected premenopausal women: a study in blood donors. Hum. Reprod. 27,1209–1216.
Schmidhuber, J., Shetty, P., 2004. Nutrition transition, obesity & noncommunicablediseases: drivers, outlook and concerns. SCN News. pp. 13–19.
Shanley, D., Kirkwood, T., 2000. Calorie restriction and ageing: a life history analysis.Evolution 54, 740–750.
Shaw, L., Elton, S., 2008. Polycystic ovary syndrome: a transgenerationalevolutionary adaptation. BJOG 115, 144–148.
Simoni, M., Tempfer, C.B., Destenaves, B., Fauser, B.C.J.M., 2008. Functional geneticpolymorphisms and female reproductive disorders. Part I: Polycystic ovarysyndrome and ovarian response. Hum. Reprod. Update 14, 459–484, %R10.1093/humupd/dmn024.
Speakman, J., 2006. Thrifty genes for obesity and the metabolic syndrome – time tocall off the search? Diabetes Vascul. Dis. Res. 3, 7–11.
Stearns, S., 1992. The Evolution of Life Histories. Oxford University Press, New York.Stearns, S.C., Nesse, R.M., Govindaraju, D.R., Ellison, P.T., 2010. Evolutionary
perspectives on health and medicine. Proc. Nat. Acad. Sci. 107, 1691–1695.Strauss, J.F., McAllister, J.M., Urbanek, M., 2012. Persistence pays off for PCOS gene
prospectors. J. Clin. Endocrinol. Metab. 97, 2286–2288.Swinbum, B., Nyomba, B.L., Saad, M.F., Zurlo, F., Raz, I., Knowler, W.C., Lillioja, S.,
Bogardus, C., Ravussin, E., 1991. Insulin resistance associated with lower rates ofweight gain in Pima Indians. J. Clin. Invest. 88, 168–173.
Taponen, S., Ahonkallio, S., Martikainen, H., Koivunen, R., Ruokonen, A., Sovio, U.,Hartikainen, A.L., Pouta, A., Laitinen, J., King, V., Franks, S., McCarthy, M.I.,Järvelin, M.R., 2004. Prevalence of polycystic ovaries in women with self-reported symptoms of oligomenorrhoea and/or hirsutism: Northern FinlandBirth Cohort 1966 study. Hum. Reprod. 19, 1083–1088.
Taponen, S., Martikainen, H., Järvelin, M.-R., Laitinen, J., Pouta, A., Hartikainen, A.-L.,Sovio, U., McCarthy, M.I., Franks, S., Ruokonen, A., 2003. Hormonal profile ofwomen with self-reported symptoms of oligomenorrhea and/or hirsutism:Northern Finland Birth Cohort 1966 study. J. Clin. Endocrinol. Metab. 88, 141–147.
Please cite this article in press as: Corbett, S., Morin-Papunen, L. The Polycysticdocrinology (2013), http://dx.doi.org/10.1016/j.mce.2013.01.001
The Rotterdam ESHRE/ASRM PCOS consensus, 2004. Revised 2003 consensus ondiagnostic criteria and long-term health risks related to polycystic ovarysyndrome (PCOS). Hum. Reprod. 19, 41–47.
Tinbergen, N., 1963. On the aims and methods of ethology. Zeitschrift fürTierpsychologie 20, 410–463.
Travis, J., 1988. Differential fertility as a major mode of selection. Tree 3, 227–230.Urbanek, M., 2007. The genetics of the polycystic ovary syndrome. Nat. Clin. Pract.
Endocrinol. Metab. 3, 103–111.Valeggia, C., Ellison, P., 2009. Interactions between metabolic and reproductive
functions in the resumption of postpartum fecundity. Am. J. Hum. Biol. 21, 559–566.
Vallegia, C., Ellison, P., 2004. Lactational Amenorrhoea in well nourished Tobawomen of Formosa, Argentina. J. Biosocial Sci. 36, 573–595.
van der Spuy, Z.M., Dyer, S.J., 2004. The pathogenesis of infertility and earlypregnancy loss in polycystic ovary syndrome. Best Pract. Res. Clin. Obstet.Gynaecol. 18, 755–771.
van der Steeg, J.W., Steures, P., Eijkemans, M.J., Habbema, J.D., Hompes, P.G.,Burggraaff, J.M., Oosterhuis, G.J., Bossuyt, P.M., van der Veen, F., Mol, B.W., 2008.Obesity affects spontaneous pregnancy chances in subfertile, ovulatory women.Hum. Reprod. 23, 324–328.
Villuendas, G., San Millan, J.L., Sancho, J., Escobar-Morreale, H.F., 2002. The -597 G->A and -174 G->C polymorphisms in the promoter of the IL-6 gene areassociated with hyperandrogenism. J. Clin. Endocrinol. Metab. 87, 1134–1141.
Vink, J., Sadrzadeh, S., Lambalk, C., Boomsma, D., 2009. Heritability of polycysticovary syndrome in a dutch twin-family study. J. Clin. Endocrinol. Metab. 91,2100–2104.
Vitzhum, V., 2009. The ecology and evolutionary endocrinology of reproduction inthe human female. Am. J. Phys. Anthropol. 52, 95–136.
Vitzthum, V., 2001. Why not so great is still good enough: flexible responsiveness inhuman reproductive functioning. In: Ellison, P. (Ed.), Reproductive Ecology andHuman Evolution. Aldine de Gruyter, New York, pp. 179–202.
Voight, B., Kudaravalli, S., Wen, X., Pritchard, J., 2006. A map of recent positiveselection in the human genome, PLoS Biol, vol. 4(3), e72.4, pp. 447–458.
Wang, J., Davies, M., et al., 2002. Obesity increases the risk of spontaneous abortionduring infertility treatment. Obes Res. 10, 551–554.
Wells, J., 2010. Adiposity in Human Evolution. The Evolutionary Biology of HumanBody Fatness Thrift and Control. Cambridge University Press, Cambridge.
Welt, C.K., Styrkarsdottir, U., Ehrmann, D.A., Thorleifsson, G., Arason, G.,Gudmundsson, J.A., Ober, C., Rosenfield, R.L., Saxena, R., Thorsteinsdottir, U.,Crowley, W.F., Stefansson, K., 2012. Variants in DENND1A are associated withpolycystic ovary syndrome in women of European ancestry. J. Clin. Endocrinol.Metab. 97, E1342–E1347.
Winter, E., Wang, J., Davies, M., Norman, R., et al., 2002. Early pregnancy lossfollowing assisted reproductive technology treatment. Hum. Reprod. 17, 3220–3223.
Witchel, S.F., Lee, P.A., Suda-Hartman, M., Trucco, M., Hoffman, E.P., 1997. Evidencefor a heterozygote advantage in congenital adrenal hyperplasia due to 21-hydroxylase deficiency. J. Clin. Endocrinol. Metab. 82, 2097–2101.
Wrigley, E., Schofield, R., 1981. The Population History of England 1541–1871; aReconstruction. Blackwell, Oxford, Oxford.
Yildiz, B., Bozdag, G., Yapici, Z., Esinler, I., Yarali, H., 2012. Prevalence, phenotype,and cardiometabolic risk of polycystic ovary syndrome under differentdiagnostic criteria. Hum. Reprod. 27, 3067–3073.
Glossary
Facultative traits: are those like callous formation on the hands and feet which areindividual responses to environmental variation
Fertility selection: is Darwinian selection based upon phenotypic differences inlifetime reproductive success
Fixation: is the change in a gene pool from a situation where there exist at least twovariants of a particular gene (allele) to a situation where only one of the allelesremains
Fixed traits: are those that are unvarying in differing environmentsKin selection: refers to apparent strategies in evolution that favour the reproductive
success of an organism’s relatives, even at a cost to their own survival and/orreproduction
Phylogeny: is the evolutionary history of an organismReproductive ecology: is a sub-field of human evolutionary biology which focuses on
the responsiveness of the human reproductive system to ecological factors. Aparticular focus of this field the role that energetics has played in humanevolution
Viability Selection: is Darwinian selection based upon phenotypic differences insurvival until reproductive adulthood
Ovary Syndrome and recent human evolution. Molecular and Cellular En-




















