
Mate Choice, Parental Care and Foraging Behaviour of the ...
211
Mate Choice, Parental Care and Foraging Behaviour of the northern trout gudgeon Mogurnda mogurnda (Pisces: Eleotridae) A thesis submitted for the degree of Master of Philosophy at the Australian National University Michelle A. Shackleton March 2007
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Mate Choice, Parental Care and Foraging Behaviour of the ...

Mate Choice, Parental Care and Foraging
Behaviour of the northern trout gudgeon
Mogurnda mogurnda (Pisces: Eleotridae)
A thesis submitted for the degree of Master of Philosophy
at the Australian National University
Michelle A. Shackleton
March 2007

DECLARATION
I declare that this thesis is all my own work and that all sources used have been
acknowledged.
M A Shackleton

ACKNOWLEDGEMENTS
I would like to thank my supervisor Michael Jennions for all his help over the many
years he has had the dubious honour of being my supervisor, as well as Alan Muir and the
other support staff for their help with all my numerous logistical issues. Thanks also to my
supervisory panel members Pat Backwell, Scott Keogh and Rob Magrath. A special thank
you to Bob Wong for showing me the ropes and always being so generous with his time and
invaluable advice. I’d also like to thank Dave Wilson for providing me with the gudgeons from
up in Darwin, and David Linterman for his patience and support with the two-spined
blackfish.
A very special thanks to Leeann Reaney without whom I would never have managed
to finish this, and for whose friendship I am eternally grateful. Thanks also to Golo Maurer for
doing it first so I could see what I was in for, and for always somehow finding the time to be
there if I needed him. Thanks to my other colleagues and friends Rachel Sims, Stuart
Cooney, Jean Drayton and Erika Alacs for their support both in and out of university. And
special appreciation too goes to my personal motivator Heather, who always knows the right
thing to say.
The biggest thank you of all goes to my parents for educating me in all aspects of life:
they have never wavered in their total support of everything I’ve done, and have always been
there when I needed them: this is for you.

TABLE OF CONTENTS
ABSTRACT.............................................................................................................................................1
CHAPTER 1 GENERAL INTRODUCTION................................................................ 31.1 Sexual selection.........................................................................................................................3
1.1.1. An overview............................................................................................................................3
1.2 Female choice and male competition...................................................................................... 4
1.3 Direct benefits model................................................................................................................ 6
1.4 Indirect benefits of female choice............................................................................................ 6
1.4.1. Genetic compatability.............................................................................................................7
1.4.2. “Good genes” model.............................................................................................................. 7
1.4.3. The Fisher process................................................................................................................ 7
1.4.4. The Lek paradox.................................................................................................................... 8
1.4.5. Strength of indirect selection................................................................................................. 9
1.5 Sexual conflict...........................................................................................................................9
1.6 Male-male dominance............................................................................................................111.7 Fish as models for parental care............................................................................................ 12
1.8 Parental care behaviours.........................................................................................................131.9 Variation in parental care.......................................................................................................151.10 Costs and benefits of parental care......................................................................................151.11 Honest Signalling...................................................................................................................161.12 Foraging: maximising energy gain.......................................................................................18
1.13 Aims of this study...................................................................................................................201.14 Study species..........................................................................................................................21References............................................................................................................................................24
CHAPTER 2 MATE CHOICE IN THE NORTHERN TROUT GUDGEON (MOGURNDA MOGURNDA).....................................................................................472.1 Introduction.............................................................................................................................47
2.1.1. Female mating preferences.................................................................................................. 47
2.1.2. Life history traits and female mating preferences................................................................47
2.1.3. Male dominance and female mating preferences................................................................48
2.1.4. Parental Care and Female Mating Preferences...................................................................50
2.1.5. Measuring female mating preference...................................................................................50
2.1.6. Aims of this study.................................................................................................................52
2.2 Methods...................................................................................................................................53
2.2.1. Pilot studies on male dominance.......................................................................................... 53
2.2.2. Mate choice trials..................................................................................................................54
2.2.3. Preference Criteria...............................................................................................................55

2.2.4. Statistical Analyses - Mate Choice Analysis........................................................................ 56
2.3 Results.....................................................................................................................................582.4 Discussion...............................................................................................................................71
2.4.1. Females preferred larger males........................................................................................... 71
2.4.2. Size assortative mating........................................................................................................72
2.4.3. Condition-dependent female choice.....................................................................................72
2.4.4. Comparing measures of mating preference.........................................................................74
References............................................................................................................................................75
CHAPTER 3 SPAWNING AND PARENTAL CARE IN THE NORTHERN TROUT GUDGEON (MOGURNDA MOGURNDA) ................................................................. 883.1 Introduction............................................................................................................................. 88
3.1.1. How much to invest in parental care: balancing costs and benefits....................................88
3.1.2. Paternal care in fish...............................................................................................................89
3.1.3. Sexual conflict over parental care.........................................................................................90
3.1.4. Adjusting parental investment: responding to partner effort................................................ 91
3.1.5. Adjusting parental investment: responding to partner condition..........................................92
3.1.6. Parental care and mate choice..............................................................................................93
3.1.7. Variation in parental care behaviours....................................................................................95
3.1.8. Aims of this study................................................................................................................. 97
3.2 Methods................................................................................................................................... 98
3.2.1. Parental Care........................................................................................................................98
3.2.2. Statistical Analyses - Parental Care Analysis...................................................................... 98
3.3 Results................................................................................................................................... 1003.4 Discussion............................................................................................................................. 108
3.4.1. Variation in parental care quality......................................................................................... 108
3.4.2. Variation in mate choice..................................................................................................... 110
References.......................................................................................................................................... 113
CHAPTER 4 FORAGING BEHAVIOUR AND SUBSTRATE DISCRIMINATION IN THE NORTHERN TROUT GUDGEON (MOGURNDA MOGURNDA)................... 1244.1 Introduction........................................................................................................................... 124
4.1.1 Food density and distribution............................................................................................. 124
4.1.2 Food density and Individual variation.................................................................................. 126
4.1.3 Microhabitat complexity...................................................................................................... 127
4.1.4 Discriminating between microhabitats................................................................................ 128
4.1.5 Aims of this study............................................................................................................... 130
4.2 Methods................................................................................................................................. 131
4.2.1 Binary Substrate Choice..................................................................................................... 131
4.2.2 Statistical Analyses - Foraging Analysis............................................................................ 132
4.3 Results................................................................................................................................... 134
4.4 Discussion............................................................................................................................. 145
References.......................................................................................................................................... 148

CHAPTER 5 GENERAL DISCUSSION...............................................................1745.1 Aims of this study................................................................................................................. 174
5.2 Condition-dependent female choice................................................................................... 175
5.3 Parental care behaviours in M. Mogurnda......................................................................... 176
5.4 Maintaining a maladaptive preference............................................................................... 177
5.5 Comparing female choice tests........................................................................................... 178
5.6 Variation in expression of mating preference................................................................... 178
5.7 Cues to mating preference.................................................................................................. 179
5.8 Foraging................................................................................................................................. 180
5.9 Conclusion............................................................................................................................ 181
References......................................................................................................................................... 182

ABSTRACT
The parental role males play ranges from no male parental care to extensive male-only care.
Females may acquire either direct or indirect fitness benefits from mate choice. Direct fitness gains
include access to male parental care, and researchers have traditionally emphasized this benefit in
species with extensive male parental care. Larger, more dominant males are assumed to be better
mates that provide higher quality parental care but many new studies dispute these assumptions. The
assumption that females prefer dominant males is questionable because few studies fully control for
male-male competition. Also, higher parental quality is not always correlated with male size, although
evidence for other honest signals of parental quality is growing. Moreover, the accuracy of traditional
simultaneous choice tests is now disputed because recent studies show that when the results of such
association preference tests are compared to latency to mating tests, preferred characteristics are
often no longer preferred or different characteristics become indicators of male attractiveness.
Here I test how male size, female choice, and the quality of male parental care interact in the
northern trout gudgeon Mogurnda mogurnda. I aimed to identify any traits that present an honest
signal of male parental quality, and whether females were choosing males based on these
characteristics. I found that female M. mogurnda prefer larger males in simultaneous choice tests, and
some evidence for size-assortative mating as larger females tended to spend proportionally more time
with larger males. Larger females also exhibited a stronger preference for generally preferred males
than smaller females, implying that choice is condition-dependent. Comparing mate choice tests,
however, I found that male traits that were preferred in traditional association tests were not preferred
in latency tests. Instead, females took longer to spawn with preferred males, which directly
contradicted the results based on association time.
There was no correlation between male size and parental care. The only trait that correlated
with male care was male attractiveness (based on association tests). In the latency to mating tests, it
is unclear what traits females based their preference response on, but they spawned sooner with non
preferred males (from association tests) who showed higher quality parental care (higher fanning rate)
than preferred males. Females could therefore still be choosing males based on an unmeasured trait
correlated with quality of care.
Foraging efficiency has an important effect on body size and condition so it is closely
associated with sexual selection as mate choice and parental are energetically expensive activities.
Recent work shows that some fish preferentially foraging around complex habitats, presumably due to
greater potential prey density, although other factors also influence foraging behaviour. I found that in
1 .

M. mogurnda there was no preference for “complex” over “simple” substrates. I conclude that the
greater potential for food created by the higher incidence of interstitial spaces indicates that increased
search and handling time associated with complex microhabitats and the lower energetic award from
smaller prey items may have balanced the preference between substrates.

CHAPTER 1 General Introduction
1.1 S e xu a l se lec tio n
1.1.1. An overview
The nature of sexual reproduction has lead to conflict between mates as they each try to
maximise their own lifetime reproductive output in the face of competition with members of the same
sex, and differences in interest between the sexes. Competition is inherent in the mechanism of
reproduction by anisogamy (the differentiation of the sexes by the size of their gametes) as many
small male gametes compete for a limited number of larger female gametes. Conflict is also present in
any situation where individuals who manipulate their mates stand to significantly increase their own
lifetime reproductive success by so doing, even if this results in a reduction in their mate’s lifetime
fitness.
Sexual selection occurs when members of each sex maximise their reproductive success in
different ways. Males produce much smaller and more numerous gametes than females and maximise
their reproductive output by mating with more females; mating with females that have greater
reproductive potential; and attaining a greater proportion of fertilisation for each reproductive event.
Females produce fewer and much larger gametes and maximise their reproductive success by mating
at an intermediate optimal rate and selecting the best “quality” males as their mates and, ultimately, as
the fertilisers of their eggs. This conflict between the sexes (and between individuals of the same sex)
leads to sexual selection.
Anisogamy results in many more male gametes (sperm) competing for far fewer female
gametes (eggs) and this creates competition between males. Males maximise their reproductive
success by maximising their number of mates, and by investing less into their current mating event
increase their future reproductive output. Successful males (i.e. those that females choose as mates
or that consistently out-compete other males) therefore tend to have a higher potential reproductive
rate (PRR) than the average female. As a result, these males are therefore inclined to invest less into
each reproductive event, so that females often have to compensate by investing more (Fisher 1930).
Females initially invest more than males into each offspring, as the production of eggs is
relatively more costly than producing sperm. This cost is small, however, in comparison to the costs of
parental care. Traditionally, females provide parental care while males invest more into finding mates.
The resulting male-biased operational sex ratio (OSR) exacerbates competition between males, and
3 .

completes a positive feedback loop by reducing competition between females for mates. This allows
females to invest more into parental care and, importantly, facilitates females being choosier. As
females become choosier, the pool of males that are likely to mates becomes smaller, so the PPR of
these males becomes even higher than that of females. This makes it even more worthwhile for these
males to invest less in care and more into seeking extra matings.
This, however, is not the whole story. Every offspring has one male and one female parent
(Fisher 1930) and therefore, given an equal adult sex ratio, overall males do not participate in more
matings than females. Thus greater competition between males due to a male-biased OSR does not
explain why males do not generally invest more into parental care (Kokko and Jennions 2003). Queller
(1997) shed some light on the situation, and suggested that multiple matings and sperm competition
leads to uncertainty in paternity and decreased benefits of investing in parental care for a male
compared to a female. For example, Neff (2003) showed experimentally that male bluegill sunfish
Lepomis macrochirus adjusted their parental effort in accordance with the perceived paternity of the
eggs in their nest, favouring clutches that they identified as being more closely related to them. Queller
(1997) also suggested that if a small proportion of males selected for by sexual selection enjoy higher
mating success, they have a higher breeding rate than the average male or the average female and
the costs of investing in parental care rather than seeking additionally mating opportunities are higher
for such males.
Fish have significantly contributed to research on mate choice behaviour. For example, three
spined sticklebacks Gasterosteus aculeatus have been used in ground-breaking research relating to
functions of visual signals (Tinbergen 1951) and were, in fact, the first species used to perform mate
choice experiments by artificially enhancing male secondary sexual characteristics (Semler 1971).
Studies on guppies Poecilia reticulata have been both prolific and innovative, including Endler’s
demonstration that predation selects for less colourful males even though females prefer more
colourful males (Endler 1980; 1983; 1988). Fish are also an excellent taxa for studying parental care
as they show all care systems (no care, biparental care, maternal care and paternal care) and care
behaviours in many forms, such as preparing nests [male common gobies, Pomatoschistus micropa
(Nyman 1953)] and caring for eggs [e.g., mouth brooding (Oppenheimer 1970)].
1.2 Female choice and male competition
Classical sex roles are that males compete, and females care and choose. This is not always
the case, however, and can even change during a breeding season (Lengkeek and Didderen 2006;
Shibata and Kohda 2006; Silva et al. 2006). To predict which sex will choose and which will compete
4 .

requires models that consider not only the OSR and PRR as models prior to Queller (1997) did (e.g.,
Clutton-Brock and Parker 1992; Parker and Simmons 1996), but also include variance in mate quality
(Johnstone et al. 1996) and, most importantly, the relatively new inclusion of life-history traits such as
the cost of breeding.
Mating is costly (Daly 1978; Pomiankowski 1988; Iwasa et al. 1991). Costs from mating
include an increased risk of predation (Sakaluk and Belwood 1984; Magnhagen 1991), and time and
energy costs from searching for males (e.g., Alatalo et al. 1988; Gibson and Bachman 1992).
Furthermore, mate choice increases these costs, which include energy spent to search for an
“acceptable” individual or fend off undesirable mates, lost mating opportunities from being selective, or
predation from increased searching behaviour (reviewed by Jennions and Petrie 1997). For example,
mate choice involves, by necessity, mate rejection, and male harassment and rejection of courting
males has shown to be is costly to female Drosophila melanogaster (Partridge and Fowler 1990;
Holland and Rice 1999). Godin and Briggs (1996) showed that female Trinidadian guppies Poecilia
reticulata reduced both their preference for bright males and their overall level of sexual activity when
the risk of predation was increased by the presence of a pike cichlid at a high predation site.
Interestingly, mate preference should be differentially costly with respect to female quality (Cotton et
al. 2006). Low quality females should suffer higher costs of mate preference than higher quality
females making the same mating decision, or higher quality females may be able to be more
discerning than poorer quality females for the same cost.
The question of how female mating preferences initially evolved and is maintained is still a key
question in the sexual selection literature. In a recent review, Kokko et al. (2006) outlined and detailed
various models of the evolution of sexual selection which I will discuss below. There is assumedly
something to be gained from the expression of mating biases or else it would be more beneficial to
mate randomly. If females choose males that provide increased benefits, selection will favour the
persistence of a female preference for the most beneficial males. This, in turn, will favour males that
provide these benefits, so the net result may be mutualistic coevolution driven by benefits (Eberhard
and Cordero 1995). Material reproductive gains that increase female lifespan reproductive success
are the most obvious, but not the only, benefits obtained from female choice. In addition, indirect
genetic benefits can lead to choosy females producing more viable and/or attractive offspring and
ultimately the production of relatively more grandchildren due to the greater reproductive success of
sexy sons (reviewed in Kokko et al. 2003).
5 .

1.3 Direct benefits model
Preference for direct benefits is favoured by natural selection because the preference itself
confers an increase in direct female fitness (Heywood 1989; Hoelzer 1989). For example, female
choice leading to direct material gains, such as nutritional gifts in spermatophores (e.g., Simmons and
Parker 1989), is under positive direct selection as it will increase female lifetime egg production and
imposes directional selection on males to signal this benefit (Price et al. 1993; Andersson 1994;
Kirkpatrick 1996). Male offerings can also include parental care, and exclusive paternal care may have
evolved in arthropods (Tallamy 2000) and territorial fish (Ah-King et al. 2005) because females prefer
males that provide parental care and paternal care was under direct selection. In extreme cases, male
investment in reproduction may exceed that of females [e.g., crickets (Simmons 1992)] and lead to
sex role reversal and male mate choice (e.g., Berglund and Rosenqvist 2003; LeBas et al. 2003). An
increase in female reproductive output offers the most straightforward explanation as to how mate
choice benefits can outweigh any costs of choice (Iwasa and Pomiankowski 1999), which explains
why it has been the main focus of research in many studies (e.g., Pizzari and Snook 2003; Wagner
and Harper 2003).
Alternately, the “sensory drive” or “sensory bias” model (West-Eberhard 1984; Ryan and Rand
1990; Endler and Basolo 1998; reviewed in Fuller et al. 2005) assumes that natural selection operates
directly on pre-existing sensory biases in circumstances other than mate choice, and may be the
initiating factor in the evolution of female choice (Kokko et al. 2003). For example, male guppies
Poecilia reticulata with greater orange colouration are more attractive to females possibly due to a
sensory bias toward orange food objects (Rodd et al. 2002). Female guppies, however, also use these
orange spots as indicators of male foraging ability and health (Endler 1980) as colour saturation is
condition-dependent (Houde and Torio 1992; Grether 2000; Kodric-Brown and Nicoletto 2005). So,
even when female choice has evolved from exploitation of a pre-existing sensory bias, direct or
indirect selection on the female preference may occur due to benefits accruing from better-quality
mates.
1.4 Indirect benefits of female choice
Situations where there appear to be no direct material benefits to mate choice are of particular
interest as they imply that there are compensatory indirect benefits to being choosy. Indirect benefits
arise due to the increased viability and/or the reproductive success of offspring, which subsequently
results in a greater number of descendents (Kokko et al. 2003).
6.

1.4.1. Genetic compatability
Choosy females can increase their fitness by reproducing with males that are more genetically
complimentary or compatible (e.g., Zeh and Zeh 1996; Brown 1997; 1997; Tregenza and Wedell
2000) due to such effects as increasing major histocompatibility complex heterozygosity (e.g., Landry
et al. 2001; Hoffman et al. 2007), inbreeding avoidance (e.g., Tregenza and Wedell 2002) and greater
offspring genetic diversity (Garant et al. 2005). For example, female fur seals go a greater distance in
a colony to conceive with males with lower relatedness, creating offspring with high heterozygosity
(Hoffman et al. 2007). Female choice for males on the basis of their genetic compatibility is, however,
not based on a linear directional preference for a male phenotypic trait (e.g., longer tails) and is
therefore difficult to observe.
1.4.2. “Good genes” model
Female choice can evolve rapidly if females show a preference for traits that are genetically
correlated with any trait favoured by direct selection such as male attractiveness or fecundity (and
ultimately with fitness) (Kokko et al. 2003). For example, females would receive indirect benefits of
female choice if they preferred males that are more colourful, and colourful males sire offspring that
have increased attractiveness or survival. Costly female mate choice might have originated and been
maintained by indirect genetic viability benefits to offspring provided by males (‘good genes’ model)
(e.g., Zahavi 1977; Andersson 1986; Pomiankowski 1987; Kirkpatrick and Ryan 1991; Moore 1994;
Kirkpatrick and Barton 1997). In support of this, female cockroaches Nauphoeta cinerea who mate
with attractive males produce offspring that hatch faster and reach sexual maturity sooner (Moore
1994). For numerous other cases of females mated to attractive males producing offspring with
increased viability, see Möller and Alatalo (1999). The good genes model predicts that female choice
imposes directional selection on males and the preference is under indirect positive selection through
the viability advantage conferred to the young.
1.4.3. The Fisher process
Instead of female choice being driven by the indirect benefits to offspring from inheriting
increased viability from the preferred male, its evolution may be driven by the simple fact that offspring
inherit the preference itself. Male offspring may therefore inherit genes for whatever trait is preferred
as well the genes for the preference for that male trait (which are only expressed in females). This
process is generally known as the Fisher process (Fisher 1958; Andersson 1994). Linkage
disequilibrium may occur because females carrying genes determining female preference for preferred
7.

male traits (e.g., genes for preferring peacocks with long trains) mate with males carrying the genes
that underlie the male traits that fulfil female mating criteria (e.g., genes for long trains in males). For
example, an artificial selection experiment female guppies were selectively mated with more orange
coloured males and after a few generations it was found that female offspring showed a stronger
preference for males with greater amounts of orange colouring (Houde 1994). Houde (1994) showed
that there is a genetic correlation between orange colour in males and female preference for orange
males, supporting Fisher’s runaway hypothesis. Other studies have shown similar genetic correlations
in sticklebacks (Bakker 1993), fruit flies (Blows 1999), moths Uthesia ornatrix (Iyengar et al. 2002) and
crickets (Gray and Cade 1999). The Fisherian model, however, lacks an explanation for how female
choice initially arose (Kirkpatrick 1987) and, although linkage disequilibrium is often used to define the
Fisher process, the two are not mutually inclusive (Lande 1980; Pomiankowski 1988).
Models encompassing both ‘good gene’ and Fisherian principles (termed the ‘Fisher-Zahavi’
process) are more inclusive and probably more realistic than trying to differentiate between these
models as this distinction is often a matter of how they are defined (Kokko et al. 2006).
1.4.4. The Lek paradox
In species where females receive little from males but sperm (and assuming that indirect
benefits are a relatively weak force in compensating for direct fitness costs), how does female choice
persist (see Kirkpatrick and Ryan 1991)7 The related question of how variation in preferred male traits
persists in a population, despite being selected against by strong directional selection, is known as the
‘lek paradox’ (e.g., Borgia 1979; Taylor and Williams 1982). Genes for preferred male traits should
become fixed in a population; nevertheless there is evidence of additive genetic variance in spite of
direction selection (Houle 1992; Pomiankowski and Möller 1995 ). One explanation is that preferred
male traits are condition dependent and the numerous loci influencing overall condition could shelter
large amounts of genetic variation (Rowe and Houle 1996). Another explanation also assumes that
preferred male traits are condition-dependent, but rejects the notion that the “best” genotype is static
in time or space. Rather, the “best” genotypes vary among environments therefore additive genetic
variation in condition may actually be high (Lesna and Sabelis 1999; Kotiaho et al. 2001; Kokko et al.
2002). Male attractiveness is subject to life-history trade-offs like all other functional traits (Grafen
1990; Kokko 2001), and males change the allocation of resources into attractiveness in response to
their environment (Candolin 2000; Badyaev and Qvarnström 2002; Hunt et al. 2004b), but due to
genotype by environment interactions allocation strategies may differ among genotypes.
8.

It is important to note that direct and indirect benefits are often treated as mutually exclusive
reasons for choosing mates, but a female can receive any mixture of the two benefits (Cordero and
Eberhard 2003; Kokko et al. 2003). As long as the sum of the benefits outweighs the costs, the
behaviour of female choice will persist (i.e. female choice ultimately results in a greater number of
descendents), otherwise it would be more advantageous to mate randomly.
1.4.5. Strength of indirect selection
Indirect selection, however, is often thought to be a weak evolutionary force in comparison to
direct selection on mating preferences (Kirkpatrick 1996; Kirkpatrick and Barton 1997; Houle and
Kondrashov 2002) but the relative contribution of direct and indirect selection has not been quantified
(Hall et al. 2000) though there have been some recent attempts (Arnqvist and Rowe 2005). Empirical
data suggest that the heritability of fitness in nature is generally low (Burt 1995; 2000). In one of the
few field studies to measure additive genetic variation of fitness, Merilä and Sheldon (2000) found no
heritability of lifetime reproductive success in male collared flycatchers Ficedula albicollis. Möller and
Alatalo (1999) reviewed 22 studies showing viability benefits to offspring of attractive males and found,
on average, that 1.5% of variation in viability was explained by male traits preferred by females. This
implies that the influence of ‘good genes’ on female choice is reasonably slight, though it appears to
be pervasive across taxa.
1.5 Sexual conflict
Strict genetic monogamy generates the same reproductive interests for both sexes, however
most species are to some extent sexually promiscuous (reviewed in Birkhead and Möller 1992) and
the lack of monogamy allows for divergent interests between mates. An alternative theory to the
traditional models of mutualistic evolution generating female choice incorporates sexual conflict (the
divergence of reproductive interests between the sexes over mating). Sexual conflict theory covers the
gamut of direct costs on females resulting from male manipulation to maximise his own reproductive
success (Chapman et al. 2003; Arnqvist and Rowe 2005).
Direct costs of sexual antagonism arise from males who evolve specialised means of enticing
females to mate (by expressing high levels of ‘stimuli’), or ways to increase their fertilisation success
at the expense of lowering a female’s lifetime reproductive output. These direct costs of sexually
antagonistic coevolution have been demonstrated to arise by a variety of mechanisms such as the
spiked intromittent organ of the bean weevil Callosobruchus maculatus that wounds females during
intromission: when females are prevented from kicking males and terminating copulations, they incur
9 .

even more damage (Crudgington and Siva-Jothy 2000). There is also evidence in the bed bug Cimex
lectularius of insemination by piercing the female body wall with modified male genitalia that result in
females remating at suboptimal rates and thereby reduces their longevity (Stutt and Siva-Jothy 2001).
In common dungfly Sepsis cynipsea, copulating pairs have long, violent precopulatory struggles,
which may be caused by females trying to avoid injuries inflicted by males (Blanckenhorn 2002). The
male intromittent organ damages the female reproductive tract and copulations greatly increases the
likelihood of female death. Aggression towards females is frequently seen among primates in
promiscuous mating systems, and in chimpanzees sexual coercion to constrain female choice and
gain higher mating success is the most probable function of male aggression toward females (Muller
et al. 2007).
The best studied case of the occurrence of male manipulation of female reproductive
physiology is the reduction in female lifespan in fruitflies Drosophila melanogaster due to harmful
proteins transferred in seminal fluid (Chen et al. 1988; e.g., Chapman and Partridge 1996; but see
Rice 1996; Holland and Rice 1999; Civetta and Clark 2000; reviewed in Chapman 2001). Seminal
fluids reduce the competitive ability of sperm from other males, thereby increasing male fertilisation
success (Harshman and Prout 1994). Additional evidence for sexual conflict in D. melanogaster is that
when females are prevented from coevolving with males, males become even more damaging to
females (Rice 1996; see also Holland and Rice 1999). This leads to the next aspect of sexually
antagonistic coevolution: evolution of female resistance to male manipulation.
A recent review by Wedell et al. (2006) examined which aspects of life histories are most likely
to be most influenced by sexual conflict. Conflict is inherent in all aspects of investment in
reproduction from gamete production to parental care. Even the expression of genes is under conflict
between the sexes: intralocus conflict is when the same allele has opposite effects on the fitness of
the sexes (Fisher 1930; Lande 1980). For example, males benefit from narrow pelvises that increase
locomotion performance while females benefit from broader pelvises for the purpose of childbearing
(Rice and Chippindale 2001). Therefore, genes for pelvic width are under sexually antagonistic
selection which acts to stabilise evolution on the trait, and also selects for sex-limited expression (Rice
1984) either through modifiers or translocation of alleles to the sex chromosome. This idea was tested
experimentally with D. melanogaster. Prasad et al. (2007) showed a marked increase in male fitness
and sexually dimorphic trait expression over 25 generations of limited genome-wide gene expression,
but when these male-evolved genomes were expressed in females, their fitness displayed a nearly
symmetrical decrease.
10.

Unlike intralocus conflicts, interlocus conflict cannot be solved through sex-limited expression.
Conflict arises from the evolutionary advantage of individuals that manipulate their mate to maximise
their own reproductive output. Genotypes that influence mates to increase their investment in the
current reproductive event will be selected for (Lessells 1999). Interlocus conflict between the sexes
arises from each sex having a different optima in terms of life history aspects of offspring production,
such as mating rate. Sexual conflict over mating is well documented: females have lower optimal
mating rates than males (Arnqvist and Nilsson 2000) as mating can be costly (Daly 1978). Males can
gain additional matings in several ways: (1) offering direct benefits [e.g. nutrients in ejaculate (Vahed
1998)]; (2) offering indirect benefits (Kokko et al. 2003); (3) exploiting sensory biases [orange
colouration in guppies (Rodd et al. 2002)]; (4) imposing costs to resisting matings [e.g. cost to females
of dislodging males in water striders (Watson et al. 1998), and; (5) by physically forcing copulations
with females [e.g. grasping and anti-grasping structures in water striders (Arnqvist and Rowe 1995).
Conflict exists over the timing of remating as well as remating rates (reviewed in Wedell et al. 2006).
Conceptually interesting, these strategies offer an alternate perspective on the evolutionary
drivers of female choice. Female preference for not mating is often termed female resistance, although
this is still a form of choice {sensu Wiley and Poston 1996) that leads to direct selection on a subset of
males (Holland and Rice 1998). In this scenario, female preferences simply act to reduce the number
of times females mate favouring the more persistent or manipulative males (Gavrilets et al. 2001;
Rowe et al. 2005). In this sense, sexual conflict is intrinsically driven by the same forces as the direct
benefits model of female mate choice. Mating preference in both models leads to an increase in
female reproductive output and sexual conflict is still inherent in the direct benefits model (some males
provide more benefits than others). The lost opportunity of gaining any extra benefit from mating
selectively is functionally the same as reducing a cost of mating unselectively.
1.6 Male-male dominance
In most taxa, larger males are more likely to win fights (spiders: Kotiaho et al. 1997; e.g. birds:
Hagelin 2002; lizards: Lopez et al. 2002; Reichard et al. 2005). It is generally assumed that stronger,
more dominant males are better quality mates (Berglund et al. 1996). One possible direct benefit of
mating with these males is that they are more likely to protect the female during and after mating.
Another benefit is that they may provide access to superior resources (e.g., better quality territories in
birds). In addition, there is a potential genetic benefit if traits that increase success during male-male
competition are heritable (Cordero and Eberhard 2003).
1 1 .

Although there is a mating bias favouring larger males in a variety of animal taxa, including
many insects (reviewed in Choe and Crespi 1997), the assumption that females prefer larger,
presumably more dominant, males has been challenged (Qvarnstrom and Forsgren 1998). A small but
growing number of studies report no correlation between male fighting ability and attractiveness
(insects: Moore and Moore 1999; Moore et al. 2001; frogs: Morrison et al. 2001; birds: Andersson et
al. 2002; lizards: Lopez et al. 2002; fish: Wong 2004). This could occur because dominant males
impose direct costs on females making them less attractive as mates. For example, they may provide
less or lower quality parental care (Forsgren 1997; Wong 2004), increase the risk of female injury
while mating (Leboeuf and Mesnick 1991), be more likely to transmit diseases (Freeland 1981) or be
sperm depleted (Pitnick and Markow 1994; Preston et al. 2001). There may also be genetic costs if,
for example, male size or dominance is negatively genetically correlated with female fitness, leading to
the production of lower quality daughters, i.e. inheriting sexually antagonistic genes (Chippindale et al.
2001). It is also possible that females do not prefer dominant males because dominance has no effect
on female or offspring fitness, or because there are other traits, uncorrelated with fighting ability, that
are better predictors of a male’s effect on female fitness. Ultimately, the relative magnitude of any
direct effects on female lifetime fecundity and indirect effects on net offspring fitness will determine
whether females should prefer to mate with dominant males (Fedorka and Mousseau 2004).
1.7 Fish as models for parental care
One trait that females may use for selecting their mates is the quality of the male parental care
that they will provide (Forsgren 1997; Wong 2004). Parental care is any form of behaviour that
increases the survival or fitness of that parent’s offspring (Clutton-Brock 1991). Parental care exists in
many forms, from preparing nests [e.g., male common gobies, Pomatoschistus micropa (Nyman
1953)], producing larger eggs [e.g., the skipper, Parnara guttata guttata, (Seko and Nakasuji 2004)],
caring for eggs [e.g., incubation in birds (Hanssen et al. 2003) and mouth-brooding in fish
(Oppenheimer 1970)], to caring for offspring [e.g., feeding nestlings (Drent and Daan 1980b)], and
fully independent young [e.g., Kloss’s gibbon, Hylobates klossii (Tilson 1981)].
Fish present highly suitable but comparatively rarely utilised model systems for studies of
parental care. Fishes are the most numerous and varied of the vertebrates, with over 28 600 species
listed by Fishbase (2004). Despite this abundance, fish are hugely under-represented in behavioural
studies. Studies on birds make up 67% of papers on the two taxa, yet there are three times as many
species of fish as there are of birds (Amundsen 2003). A lot less is known about the patterns of mate
choice and parental care in fish compared to birds. A series such as Birds of the World (del Hoyo et al.
12.

1992-2003) summarise comprehensive knowledge about almost all extant bird species, while there is
nothing comparable for fish. Fish biologists must either stick to a few well-known fish species, or
spend a lot of time learning about the fish they are studying. There are a few reasons for this: (1)
aquatic habitats are difficult to conduct research in; (2) many studies where the focus is on monitoring
variation among individuals require closed, preferably small, populations but aquatic habitats are
usually continuous; and (3) fish are difficult to mark individually.
In teleost (bony) fish, 57% of freshwater breeding families show parental care of eggs or
young (Baylis 1981). Within ray-finned fish, 20% of the 400+ existing families have species that
display some form of parental care (Blumer 1979; 1982). Fish are an interesting taxa for parental care
research because they show a wide range of often dynamic care patterns ranging from no-care, to
uniparental, paternal and maternal care (Gross and Sargent 1985; Reynolds et al. 2002). Among fish,
the majority of external fertilisers (76%) display male parental care which usually involves defence of a
territory containing eggs from more than one female (Gross and Sargent 1985).
A recent study by Mank et al. (2005) reviewed the phylogenetic data for the whole ray-finned
fish group (Actinopterygii). Mank et al. found parental care in 31% (62 of 174) of families, and in 25%
(49 of 193) of external fertilising (ovuliparous) fishes in the group. They showed that all forms of
parental care (biparental, maternal and paternal) have arisen repeatedly and independently during
Actinopterygii evolution. Paternal care arose from external fertilisation, and maternal care arose via
internal fertilisation from external fertilisation. Foundation phylogenetic studies of the evolution of
parental care in a few specific groups of fish (Gross and Sargent 1985) and anuran frogs (Weygoldt
1987) proposed a stepping stone model of the evolution of care from external fertilisation to paternal
care, biparental care and finishing with female-only care. The results from Mank et al (2005) contradict
these stepping stone models, however, and this is not the first study to do so (Goodwin et al. 1998;
Summers et al. 1999).
1.8 Parental care behaviours
Egg guarding is the most common form of care in fish, followed by nest building and then egg
fanning (Blumer 1979). Reduced egg predation or increased hatching success are probably the two
main benefits of male guarding (Baylis 1981). In teleosts, egg guarding is associated with relatively
large egg size and comparatively low fecundity possibly because the longer development period of
larger eggs favours guarding to reduce mortality. In fish, fanning is the most studied parental care
behaviour as oxygen availability is the predominant factor affecting embryos development and survival
(Davis 1975). Embryos subjected to low oxygen (hypoxic) conditions suffer reduced development rate
13.

(Kinne and Kinne 1962; Mihelakakis and Kitajima 1994) and lower survivorship (Dudley and Eipper
1975; Spoor 1984) and egg fanning primarily functions to provide more oxygen for developing
embryos (Tinbergen 1951; van lersel 1953; Sevenster 1961). At low oxygen levels, males will increase
their fanning rate (Reebs et al. 1984; Jones and Reynolds 1999; Green and McCormick 2005) but the
costs of care such as fanning also increase (Hale et al. 2003; Lissaker et al. 2003). As conditions
change and the cost to benefit ratio changes, males often respond by eating all the eggs especially if
they are tending a small clutch (Lissaker et al. 2003). Parents do have ways to alleviate these costs,
such as building nests with larger opening for increased ventilation under low oxygen conditions
(Takegaki and Nakazono 1999; Lissaker and Kvarnemo 2006). It is therefore clear that increasing
oxygen availability to offspring due to fanning by a parent is very important for the offspring’s
development and survival (Zoran and Ward 1983; Kramer 1987; Takegaki and Nakazono 1999;
Lissaker et al. 2003).
Studies measuring filial cannibalism (eating one’s own offspring) show that this behaviour
appears to be common among paternally caring fish (Fitzgerald 1992; Hoelzer 1995; DeWoody et al.
2001; Manica 2004). Partial filial cannibalism has been suggested to have evolved as an adaptive trait
that helps parents to recover the energetic costs of parental care, assuming parents are food-limited
and offspring represent an adequate alterative energy source (Rohwer 1978). Complete filial
cannibalism may have evolved to allow parents to compensate from ‘doomed’ reproductive events
(Fitzgerald 1992; Manica 2002). Studies on Cortez Damselfish Stegastes rectifraenu (Hoelzer 1992)
and gobies Pomatoschistus microps (Kvarnemo et al. 1998) have shown that supplementary feeding
reduces the rates of filial cannibalism, supporting the claim that parental care is costly and that partial
filial cannibalism helps to alleviate these costs. That filial cannibalism is a means to obtain additional
provisions is not an undisputed explanation though. Other studies have shown no decrease in filial
cannibalism by males provided with supplementary food [e.g., in sticklebacks (Belles-lsles and
Fitzgerald 1991), fantail darters (Lindstrom and Sargent 1997), river blennies (Vinyoles et al. 1999),
and sand gobies (Kvarnemo 1997)].
The duration and frequency of parental care behaviours has been shown to vary with
changing parental care cost-to-benefit ratios. The incidence of filial cannibalism, for example, has
been studied by Manica (2004) in the scissortail sergeant Abudefdul sexfasciatus. The cost of care
was reduced by supplementary feeding, and the apparent benefits were reducing by changing the
perceived relatedness of the male to the offspring by showing the caring male a video recording of
rival males to simulate intrusions into the parent’s nest. Supplementary feeding (lowering the cost of
14.

care) reduced the rate of filial cannibalism, while decreased certainty of paternity (reduced benefit of
care) increased the rate of filial cannibalism.
1.9 Variation in parental care
A foundation assumption of studies on the evolution of parental care is that caring for offspring
reduces the survival or future reproductive success of parents (for a comprehensive discussion see
Clutton-Brock 1991). Models of parental care assume that the cost in expenditure in a current
reproductive event reduce the survival or future reproductive success of parents, despite increasing
the current offspring’s reproductive value (Williams 1966a; 1966b; Schaffer 1974b; 1974a; Pianka
1976; Carlisle 1982; Sargent and Gross 1985; 1986; Winkler 1987; Curio 1988; Montgomerie and
Weatherhead 1988). Conversely, saving resources for future reproduction reduces the fitness of the
present offspring but increases subsequent reproductive success. Individuals are expected to
maximise their overall reproductive success by trading off investment between current and future
reproductive events (Winkler 1987). For example, when dissolved oxygen levels decrease, the cost to
ratio benefit changes as care becomes more costly (Hale et al. 2003; Lissäker et al. 2003), and males
often respond by abandoning their current reproductive investment (eating all the eggs). Although
parents would be expected to adjust parental care to maximise their own fitness, to predict variation in
parental care accurately there must be an understanding of the costs and benefits associated with
parental care under different situations.
1.10 Costs and benefits of parental care
Parental care is demonstrably costly. There are energetic costs to building a nest or burrow to
house offspring, for example, black-billed magpies Pica hudsonia have to increase their daily intake by
0.7% - 1.0% to cover the energetic costs of nest construction (Stanley 2002). Although this cost is
relatively small, producing eggs is very costly; female black-billed magpies egg laying is estimated to
require a full 23% of the daily energy expenditure (Stanley 2002). Caring for eggs is also costly, a
clutch-size manipulation experiment in female common eiders Somateria mollissima showed that
females of low quality were unable to increase incubation effort and were more likely to abandon the
nest (Hanssen et al. 2003). On the other hand, females of higher quality apparently had the capacity
both to increase incubation effort and investment in brood care (Hanssen et al. 2003). Male
cardinalfish (Apogonidae) care for their eggs by keeping them in their mouths until they hatch
(Thresher 1984). Males therefore do not feed during this time (Okuda and Yanagisawa 1996) and
mouth-brooding has been found to severely reduce the male’s ability to take up oxygen at reduced
15 .

dissolved oxygen levels (Östlund-Nilsson and Nilsson 2004). Caring for young after they hatch may be
even more costly, feeding nestlings required quadrupling of the basal metabolic rate in five species of
bird (Drent and Daan 1980b). These examples all highlight the importance of body condition as a
factor that could affect the quality of parental care an individual can provide.
In most species, the costs of care are high and for it to have evolved there must, therefore, be
appreciable benefits in terms of improving offspring survival or performance (e.g., in situations of high
predation risk, under high competition for resources or under harsh conditions). For example, female
treehoppers Publilia concava exhibit a range of guarding durations, and females that remained to
guard eggs had fewer and smaller future broods than those that abandoned their offspring, suggesting
a high cost to future reproductive output (Zink 2003). On the other hand, experimentally removing the
female resulted in loss of half of the eggs by predation. Estimates of lifetime fecundity of females that
abandon eggs early versus remain to guard eggs showed roughly equivalent levels of fitness.
Females may therefore only choose to guard when their eggs are under immediate high predation risk
(Zink 2003). Similarly, in the burying beetles Nicrophorus vespilloides parents provide elaborate
parental care to their offspring by guarding a resource from competing conspecifics that has a major
beneficial effect for offspring. Parental beetles bury a small vertebrate carcass (which constitutes the
sole food source for the larvae) and this complex parental care immediately prior to larval hatching
greatly improved offspring survival rates. In addition, continued parental presence for several days
after eggs hatch, during which adults regurgitate pre-digested carrion to their young (Fetherston et al.
1990), also improved larval growth (Eggert et al. 1998).
1.11 Honest Signalling
Parental care can be seen as a direct benefit of female choice, and if a male can signal his
quality as a caregiver, he may increase his mating and reproductive success. A female who uses that
signal to choose a high quality male will also gain from this choice. If males of inferior quality,
however, also displayed this signal they stand to gain from the increase in mating success but females
would suffer from the decreased parental care. The question is then, in the face of these conflicting
interests between signallers and receivers, can signals of quality (care or otherwise) be honest?
Sexual displays (or signals) are under selection by female choice and many studies have
shown a positive correlation between sexual displays and either parental investment or resources
(e.g., Linville et al. 1998; Candolin 2000; Senar et al. 2002; Voltura et al. 2002; Kolm 2004; Halupka
and Borowiec 2006). Other studies show the contrary result, in that preferred or actively courting
males actually provide less parental investment (e.g., Qvarnström 1997; Wong 2004; Mitchell et al.
16.

2007) or that there is no relationship between parental investment and a sexual display signal (e.g.,
Mountjoy and Lemon 1997; Smiseth et al. 2001).
Secondary sexual characteristics are expensive, as already discussed, and the “handicap
principle” suggests that only males of high quality and in good condition can bear the cost of
displaying them, therefore ensuring that sexual advertisement will be honest (Zahavi 1975). The
requirement that signalling is costly therefore appears essential for signalling of quality to be honest
(Getty 1998). Some studies have shown, however, that costly advertisement does not always lead to
honest signals. Parasitised male Drosophila melanogaster with reduced life expectancy increased
their courtship effort (Polak and Starmer 1998) while three spined sticklebacks in poor condition
displayed greater nuptial colouring that their higher quality counterparts. This can come about when,
at the end of a male’s lifetime, future reproductive chances become diminished and the trade-off
between current and future reproductive is diminished. This terminal effort in signalling could lead to
temporary dishonest signalling, although ‘on average’ across a lifetime the positive correlation
between signal expression and male quality persists (Kokko 1998b). There is the possibility, however,
that this terminal energy expenditure in signalling also correlates with an increase in expenditure on
parental care, and the signal then remains honest in terms of signalling parental investment.
There are two hypotheses that model whether or not signalling will be an honest predictor of
parental quality. The good parent hypothesis (Heywood 1989; Hoelzer 1989) suggests that male
secondary sexual characteristics will signal a male's ability to provide parental care due to selection
imposed by female choice and therefore predicts honest signalling. The ‘differential resource
allocation’ hypothesis predicts a reduction in care from attractive males because females will increase
their parental investment to obtain or retain an attractive mate (Burley 1986). In a key model, Kokko
(1998) shows that the predictions of both models can occur and it is the extent to which males stand to
gain from additional matings that will decide on the direction of signal honesty. If the benefits of
additional matings are only marginal, signals are expected to honestly convey genetic and phenotypic
(parental care) quality. If the gains from additional matings are increased, males are likely to allocate
greater resources to displaying the signal and find more mates, and provide less care. More generally,
the effort that males invest in parental care is often negatively associated with mating opportunities
(Westneat et al. 1990).
As an aside, the evolution and maintenance of honest signaling may be partially affected by
condition. Mate preferences for honest signals should be more beneficial for high quality females if low
quality females have less to gain from discrimination (Cotton et al. 2006b). Hypothetically, in a species
in which male display rate honestly signaled male investment in parental care, high quality females
17.

who can produce more offspring than the average male can care for should prefer well-ornamented
mates, as this will directly increase their fitness. In contrast, if low quality females produce few
offspring that can be adequately cared for by an average male, then such females should mate at
random, and avoid paying the costs of mate choice where there is a much smaller benefit (Cotton et
al. 2006).
Signals of parental care need not necessarily be sexually selected traits. Females might
directly observe cues of parenting instead of relying on the honesty of correlated traits. Studies have
been conducted that shown females prefer males of high parental quality, rather than basing their
choice on male dominance or size (e.g., sand gobies Forsgren 1997) although what cues females
base this choice on isn’t always clear. Three spined sticklebacks have exclusive paternal care and
males have larger pectoral fins than females, probably to facilitate better care for eggs when fanning.
The size of a male’s pectoral fin is condition-dependent and females could use fin size to directly
select males of higher parental quality (Bakker and Mundwiler 1999), although this still leaves open
questions about fanning rate which also affects the quality of the care provided.
In some species, males invest a greater amount into parental care than females, and where
nesting sites or territories are limited, the operational sex ratio becomes skewed toward females.
Termed ‘sex role reversal’ (Gwynne 1991; Vincent et al. 1992), this lead to a situation where females
compete for mates and males become choosy. This has been demonstrated in many species (e.g.,
pipefish, Silva et al. 2006; and blennys, Lengkeek and Didderen 2006; Shibata and Kohda 2006). So
do females display honest signalling as well? Some studies show they do (Möller 1993; Langmore et
al. 1996; Svensson et al. 2006) and some that they do not (Hill 1994; Telia et al. 1997).
1.12 Foraging: maximising energy gain
With so much literature focusing on energetically expensive activities such as presenting and
maintaining sexually selected traits, mate searching, interspecific competition, mate choice, mating,
and parental care, it is logical to examine means by which animals obtain energy for these activities.
The efficiency with which individuals gain energy is of immense importance and affects their
competitiveness in competition for access to mates (e.g., dominance struggles) and the expression of
traits that are condition-dependent (e.g., honest signals of quality)
Different habitat structures present animals with differences in food density and associated
food values, such as search times. As discussed above, several species of generalist feeding fish
have been shown to selectively choose food based on their size (e.g., Ivlev 1961; Galbrait 1967;
Brooks 1968; Hall et al. 1970; Munk 1992; Forrester et al. 1994; Luo et al. 1996; Graeb et al. 2006;
18.

Hyvarinen and Huusko 2006). Food size is important to fish growth and may studies demonstrate a
positive relationship between the two (Leeren 1958; Parker and Larkin 1959; Paloheim and Dickie
1966; Graeb et al. 2004; Harter and Heck 2006; Persson and De Roos 2006). Food size is also
important when trying to maximise energy gain, as optimal allocation time spent searching for and
handling prey will change with the energy return for different sized food items
Microhabitat choice (specifically substrate discrimination) is one aspect of patch choice
behaviour that was the subject of a recent study by Webster and Hart (2004). Webster and Hart
(2004) presented three spined sticklebacks Gasterosteus aculeatus simultaneously with food items on
simple and complex substrates, repeating the experiment with different densities of food items. The
sticklebacks showed a marked preference for complex microhabitats when food densities were equal
between substrates, and when density was greater on the complex substrate. Webster and Hart
(2004) cite the link between increased substrate complexity with higher macroinvertebrate densities as
the cause. Increased structural complexity, however, increases the handling and searching times of
smaller prey and this will decrease the associated value of that food item and therefore the net intake
of energy for that patch. I wanted to emulate this study to see if this result was repeatable and if the
decrease in food value from a more complex microhabitat would yield a different result in a different
species.
19.

1.13 Aims of this study
The overall aims of my study are:
1. Investigate what female Mogurnda mogurnda prefer in relation to male size and fighting
ability. Fighting ability (or dominance) is often automatically regarded as being a predictor of female
mate choice but recent papers do not support this claim. My study is designed to add to the growing
list of literature that document whether or not there is a female preference for dominant males when
male-male competition is controlled for.
2. To compare female mate preferences between traditional association preference tests and
latency to mating (spawning time) trials. Recent studies suggest that association preference tests
are not necessarily robust, when the results are compared to latency to mating tests, either preferred
characteristics are no longer preferred or different characteristics become indicators of male
attractiveness.
3. To investigate whether male M. mogurnda display honest signalling of their parental quality,
and if females chose males based on these characteristics. To do this I looked for any correlations
between male size, dominance or attractiveness (as defined by female preference trials) and
parental care.
4. To study a relatively new aspect of foraging behaviour, substrate discrimination. I wanted to
see if individuals showed preference for the greater potential for food presented by the higher
incidence of interstitial spaces in “complex” substrates. By comparison, “simple” substrates hold less
potential for hiding food items. I wanted to see if M. mogurnda showed a preference for foraging
grounds based on these topographical differences.
20.

1.14 Study species
Mogurnda mogurnda, commonly known as the northern trout gudgeon, is widely distributed
across northern Queensland and the Northern Territory (Allen et al. 2002). They can be found from
Mossman (QLD) to Mitchell River (WA), in the Lake Eyre drainage, and in some areas of southern
Papua New Guinea (Fig. 1.1.1). M. mogurnda inhabits slow moving waters in rivers, creeks and
billabongs, congregating around vegetation or rocks. Adult fish prey upon insects, crustaceans,
worms, molluscs, fishes and plant materials.
With wide physiochemical tolerances, M. mogurnda can withstand short periods of low oxygen
levels, water temperatures from 5 to at least 32°C and salinities up to 10,000 parts per million.
Growing to a maximum size of 17cm, their appearance is very similar to that of the better-known
purple-spotted gudgeon Mogurnda adspersa. Dark brown dorsally and lightening to a cream on the
belly, M. mogurnda has dark bars running vertically with numerous reddish spots featuring prominently
on the body and head Fig. 1.1.2). Three diagonal bands of spot run from under the eye across the
cheek, and spots featuring on the dorsal, caudal and anal fins. Somewhat non-descript when stressed,
M. mogurnda displays iridescent blues and purples when in breeding colours.
M. mogurnda spawns during the rainy season (November to March). Sexual dimorphism is
displayed during the breeding season; the male urinogenital papilla is pointed and may attain an
intense blue colouration, and in females the papilla is shorter and broader with a fringed margin. I
used this difference to sex individuals in my study. Spawning involves the adults swimming closely
together and the female produces several batches of 100-500 eggs that are deposited on rocks or
logs. The demersal adhesive eggs are attached, in a cluster of around 50mm in diameter, to the
substrate by small threads and waft in the current. The males guards and fans the eggs until hatching,
which anecdotal evidence suggests takes between 8 and 10 days in the wild, although they will hatch
much more rapidly in aquaria (on average after about 5 days).
21 .

Figure 1.1.1 The distribution of Mogurnda mogurnda across Australia is highlighted across northern
Australian and the Lake Eyre drainage. They are also found in southern Papua New Guinea (not
shown) (NFA, 2006).
22 .

Figure 1.1.2 Male Mogurnda mogurnda guarding his egg mass in an aquarium.
23.

References
(2004) Fishbase (www.fishbase.org). In: Froese R, Pauly D (eds)
Ah-King M, Kvarnemo C, Tullberg BS (2005) The influence of territoriality and mating system on the evolution of male care: a phylogenetic study on fish. Journal of Evolutionary Biology 18:371- 382
Ahnejsö I, Kvarnemo C, Merilaita S (2001) Using potential reproductive rates to predict mating competition among individuals qualified to mate. Behavioral Ecology 12:397-401
Alatalo RV, Carlson A, Lundberg A (1988) The search cost in mate choice of the Pied Flycatcher. Animal Behaviour 36:289-291
Allen GR, Midgley SH, Allen M (2002) Field Guide to the Freshwater Fishes of Australia. Western Australian Museum, Perth
Amundsen T (2003) Fishes as models in studies of sexual selection and parental care. Journal of Fish Biology 63:17-52
Amundsen T, Forsgren E (2001) Male mate choice selects for female coloration in a fish. Proceedings of the National Academy of Sciences, U.S.A. 98:13155-13160
Andersson M (1994) Sexual Selection. Princeton University Press, Princeton, New Jersey
Andersson S, Pryke SR, Ornborg J, Lawes MJ, Andersson M (2002) Multiple receivers, multiple ornaments, and a trade-off between agonistic and epigamic signalling in a widowbird. American Naturalist 160:683-691
Arnqvist G, Nilsson T (2000) The evolution of polyandry, multiple mating and female fitness in insects. Animal Behaviour 60:145-164
Arnqvist G, Rowe L (1995) Sexual Conflict and Arms Races between the Sexes - a Morphological Adaptation for Control of Mating in a Female Insect. Proceedings of the Royal Society of London Series B-Biological Sciences 261:123-127
Arnqvist G, Rowe L (2005) Sexual Conflict. Princeton University Press, Princeton, NJ
Arnqvist G, Rowe L, Krupa J J, Sih A (1996) Assortative mating by size: a meta-analysis of mating patterns in water striders. Evolutionary Ecology 10:265-284
BakkerTCM (1993) Positive correlation between female preference and preferred male ornament in sticklebacks. Nature 363
BakkerTCM, Kunzler R, Mazzi D (1999) Condition-related mate choice in sticklebacks. Nature 401:234
BakkerTCM, Mundwiler B (1999) Pectoral fin size in a fish species with paternal care: a condition- dependent sexual trait revealing infection status. Freshwater Biology 41:543-551
Balshine-Earn S (1995) The costs of parental care in Galilee St Peter's fish, Sarotherodon galilaeus. Animal Behaviour 50:1-7
24 .

Barlow G, Francis RC, Baumgartner FV (1990) Do the colours of parents, companions and selfinfluence assortative mating in the polychromatic Midas cichlid? Animal Behaviour 40:713-722
Bateman AJ (1948) Intra-Sexual Selection In Drosophila. Heredity 2:349-368
Baylis JR (1981) The evolution of parental care in fishes, with reference to Darwin’s rule of male sexual selection. Environmental Biology of Fishes 6:223-251
Belles-lsles JC, FitzGerald GJ (1991) Filial cannibalism in sticklebacks - a reproductive management strategy. Ethology Ecology & Evolution 3:49-62
Berejikian BA, Tezak EP, Larae AL (2000) Female mate choice and spawning behaviour of Chinook salmon under experimental conditions. Journal of Fish Biology 57:647-661
Berglund A, Bisazza A, Pilastro A (1996) Armaments and ornaments: An evolutionary explanation of traits of dual utility. Biological Journal of the Linnean Society 58:385-399
Birkhead TR, Möller AP (1992) Sperm Competition in Birds. Academic Press, New York.
Bisazza A, Marconato A (1988) Female mate choice, male-male competition and parental care in the river bullhead, Cottus gobio L. (Pisces, Cottidae). Animal Behaviour 36:1352-1360
Blanchfield PJ, Ridgway MS (1997) Reproductive timing and use of reed sites by lake-spawning brook trout (Salvelinus fontinalis). Canadian Journal of Fisheries and Aquatic Sciences 54:747-756
Blanckenhorn WU (2002) The costs of copulating in the dung fly Sepsis cynipsea. Behavioral Ecology and Sociobiology 13
Blanckenhorn WU, Kraushaar U, Reim C (2003) Sexual selection on morphological and physiological traits and fluctuating asymmetry in the yellow dung fly. Journal of Evolutionary Biology 16:903- 913
Blows MW (1999) Evolution of the genetic covariance between male and female components of mate recognition: an experimental test. Proceedings of the Royal Society B: Biological Sciences 266:2169-2174
Blumer LS (1979) Male parental care in the bony fishes. Quarterly Review of Biology 54:149-161
Blumer LS (1982) A bibliography and categorization of bony fish exhibiting parental care. Zoological Journal of the Linnean Society 76:1-22
Blumstein DT, Evans CS, Daniel JC (2000) JWatcher. In, 0.9 edn. Animal Behaviour Laboratory,Macquarie University, Sydney, pp A powerful tool for the quantitative analysis of behaviour.
Bonduriansky R, Rowe L (2005) Sexual selection, genetic architecture, and the condition dependence of body shape in the sexually dimorphic fly Prochyliza xanthostoma (Piophilidae). Evolution 59:138-151
Borgia G (1979) Sexual selection and the evolution of mating systems. In: Blum MS, Blum NA (eds) Sexual selection: testing the alternatives. Academic Press, New York, pp 19-80
Brooks JL (1968) Effects of prey size selection by lake planktivores. Systematic Zoology 17:273-&
Brown JL (1997) A theory of mate choice based on heterozygosity. Behavioral Ecology 8:60-65
25.

Brown WD (1990) Size-assortative mating in the blister beetle Lytta magister (Coleoptera: Meloidae) is due to male and female preference for larger mates. Animal Behaviour 40:901-909
Brown WD (1993) The cause of size-assortative mating in the leaf beetle Trirhabda canadensis (Coleoptera: Chrysomelidae). Behavioral Ecology and Sociobiology 33:151-157
Buchanan KL, Catchpole CK (2000) Song as an indicator of male parental effort in the sedge warbler. Proceedings of the Royal Society of London Series B-Biological Sciences 267:321-326
Budaev SV, Zworykin DD, Mochek AD (1999) Individual differences in parental care and behaviour profile in the convict cichlid: a correlation study. Animal Behaviour 58:195-202
Bukowski TC, Linn CD, Christenson TE (2001) Copulation and sperm release in Gasteracantha cancriformis (Araneae : Araneidae): differential male behaviour based on female mating history. Animal Behaviour 62:887-895
Bulmer M (1989) Structural instability of models of sexual selection. Theoretical Population Biology 35:195-206
Burley N (1986) Sexual selection for aesthetic traits in species with biparental care. American Naturalist 127:415-445
Burley N (1988) The differential-allocation hypothesis: an experimental test. American Naturalist 132:611-628
Burley NT, Parker PG, Lundy K (1996) Sexual selection and extrapair fertilization in a sociallymonogamous passerine, the zebra finch (Taeniopygia guttata). Behavioral Ecology 7:218-226
Burt A (1995) Perspective: the evolution of fitness. Evolution 49:1-8
Burt A (2000) Natural selection in the wild. Trends in Ecology & Evolution 15:306-307
Candolin U (1999a) Male-male competition facilitates female choice in sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 266:785-789
Candolin U (1999b) The relationship between signal quality and physical condition: is sexual signalling honest in the three spined stickleback? Animal Behaviour 58:1261-1267
Candolin U (2000) Changes in expression and honesty of sexual signalling over the reproductive lifetime of sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 267:2425-2430
Candolin U (2003) The use of multiple cues in mate choice. Biological Review 78:575-595
Carlisle TR (1982) Brood success in variable environments - Implications for parental care allocation. Animal Behaviour 30:824-836
Chellappa S, Huntingford SA (1989) Depletion of energetic reserves during reproductive aggression in male three spined stickleback, Gasterosteus aculeatus. Journal of Fish Biology 35:275-286
Chippindale AK, Gibson JR, Rice WR (2001) Negative genetic correlation for adult fitness between sexes reveals ontogenetic conflict in Drosophila. Proceedings of the National Academy of Sciences of the United States of America 98:1671-1675
26.

Choe JC, Crespi BJ (1997) Mating systems in insects and arachnids. Cambridge University Press, Cambridge
Clark DL, Biesiadecki B (2002) Mating success and alternative reproductive strategies of thedimorphic jumping spider, Maevia inclemens (Aaneae, Salticidae). Journal of Arachnology 30:511-518
Clutton-Brock TH (1991) The Evolution of Parental Care. Princeton University Press, Princeton
Clutton-Brock TH, Parker GA (1992) Potential Reproductive Rates And The Operation Of Sexual Selection. Quarterly Review of Biology 67:437-456
Coleman RM, Fischer RU (1991) Brood Size, Male Fanning Effort And The Energetics Of ANonshareable Parental Investment In Bluegill Sunfish, Lepomis-macrochirus (Teleostei, Centrarchidae). Ethology 87:177-188
Coleman RM, Gross MR, Sargent RC (1985) Parental Investment Decision Rules - A Test In Bluegill Sunfish. Behavioral Ecology and Sociobiology 18:59-66
Cooke SJ, Schreer JF, Philipp DP, Weatherhead PJ (2003) Nesting activity, parental care behavior, and reproductive success of smallmouth bass, Micropterus dolomieu, in an unstable thermal environment. Journal of Thermal Biology 28:445-456
Cordero C, Eberhard WG (2003) Female choice of sexually antagonistic male adaptations: a critical review of some current research. Journal of Evolutionary Biology 16:1-6
Cote IM, Hunte W (1989) Male And Female Mate Choice In The Redlip Blenny - Why Bigger Is Better. Animal Behaviour 38:78-88
Cotton S, Fowler K, Pomiankowski A (2004) Condition dependence of sexual ornament size andvariation in the stalk-eyed fly Cyrtodiopsis dalmanni (Diptera : Diopsidae). Evolution 58:1038- 1046
Cotton S, Rogers DW, Small J, Pomiankowski A, Fowler K (2006a) Variation in preference for a male ornament is positively associated with female eyespan in the stalk-eyed fly Diasemopsis meigenii. Proceedings of the Royal Society of London Series B-Biological Sciences 273:1287-1292
Cotton S, Small J, Pomiankowski A (2006b) Sexual Selection and Review Condition-Dependent Mate Preferences. Current Biology 16:755-765
Crudgington HS, Siva-Jothy MT (2000) Genital damage, kicking and early death - The battle of the sexes takes a sinister turn in the bean weevil. Nature 407:855-856
Cunningham EJA, Russell AF (2000) Egg investment is influenced by male attractiveness in the mallard. Nature 404:74-77
Curio E (1988) Relative realized life span and delayed cost of parental care. American Naturalist 131:825-836
Daly M (1978) Cost of mating. American Naturalist 112:771-774
Davis JC (1975) Exchange of oxygen at gills of fish in response to oxygen availability. Journal of the Electrochemical Society 122:C101-C101
Dawkins R, Carlisle TR (1976) Parental investment, mate desertion and a fallacy. Nature 262:131-133
27.

de Lope F, Möller AP (1993) Female reproductive effort depends on the degree of ornamentation of their mates. Evolution 47:1152-1160
del Hoyo J, Elliott A, Sargatal J (1992-2003) Handbook of the Birds of the World. Lynx Edicions, Barcelona
Demartini EE (1987) Paternal Defense, Cannibalism And Polygamy - Factors Influencing TheReproductive Success Of Painted Greenling (Pisces, Hexagrammidae). Animal Behaviour 35:1145-1158
DeWoody JA, Fletcher DE, Wilkins SD, Avise JC (2001) Genetic documentation of filial cannibalism in nature. Proceedings of the National Academy of Sciences of the United States of America 98:5090-5092
Downhower JF, Brown L (1980) Mate preferences of female mottled sculpins, Cottus bairdi. Animal Behaviour 28:728-734
Drent RH, Daan S (1980a) The prudent parent: Energetic adjustments in avian breeding. Ardea 68:225-252
Drent RH, Daan S (1980b) The prudent parent: Energetic adjustments in avian breeding. Ardea 68:225-252
Dudley RG, Eipper AW (1975) Survival of largemouth bass embryos at low dissolved oxygen concentrations. Transactions of the American Fisheries Society 104:122-128
Eberhard WG (1996) Female control: sexual selection by cryptic female choice. Princeton University Press, Princeton, New Jersey
Eberhard WG, Cordero C (1995) Sexual Selection By Cryptic Female Choice On Male SeminalProducts - A New Bridge Between Sexual Selection And Reproductive Physiology. Trends in Ecology & Evolution 10:493-496
Eggert AK, Reinking M, Muller JK (1998) Parental care improves offspring survival and growth in burying beetles. Animal Behaviour 55:97-107
Emlen ST, Oring LW (1977) Ecology, sexual selection, and the evolution of mating systems. Science 197:215-223
Endler JA (1980) Natural selection on color in Poecilia reticulata. Evolution 34:76-91
Endler JA (1983) Natural and sexual selection on color patterns in Poeciliid fishes. Environmental Biology of Fishes 9:173-190
Endler JA (1988) Sexual selection and predation risk in Guppies. Nature 332:593-594
Endler JA, Basolo AL (1998) Sensory ecology, receiver biases and sexual selection. Trends in Ecology & Evolution 13:415-420
Evans JP, Kelly JL, Bisazza A, Finazzo E, Pilastro A (2004) Sire attractiveness influences offspring performance in guppies. Proceedings of the Royal Society of London Series B-Biological Sciences 271:2035-2042
Fedorka KM, Mousseau TA (2004) Female mating bias results in conflicting sex-specific offspring fitness. Nature 429:65-67
28 .

Fetherston IA, Scott MP, Traniello JFA (1990) Parental Care in Burying Beetles - the Organization of Male and Female Brood-Care Behavior. Ethology 85:177-190
Fisher RA (1930) The Genetical Theory of Natural Selection. Clarendon Press, Oxford
Fisher RA (1958) The Genetical Theory of Natural Selection. Dover, New York
Fitzgerald GJ (1992a) Filial cannibalism in fishes - why do parents eat their offspring. Trends in Ecology & Evolution 7:7-10
FitzGerald GJ (1992b) Filial cannibalism in fishes: Why do parents eat their offspring? Trends in Ecology & Evolution 7:7-10
Fitzpatrick S, Berglund A, Rosenqvist G (1995) Ornaments or offspring: costs to reproductive success restrict sexual selection processes. Biological Journal of the Linnean Society 55:251-260
Forrester GE, Chace JG, McCarthy W (1994) Diel and density-related change in food consumption and prey selection by Brook Charr in a New Hampshire stream. Environmental Biology of Fishes 39:301-311
Forsgren E (1997) Female sand gobies prefer good fathers over dominant males. Proceedings of the Royal Society of London Series B-Biological Sciences 264:1283-1286
Forsgren E, Karlsson A, Kvarnemo C (1996) Female sand gobies gain direct benefits by choosing males with eggs in their nests. Behavioral Ecology and Sociobiology 39:91-96
Freeland WJ (1981) Parasitism and behavioral dominance among male-mice. Science 213:461-462
Friberg U, Arnqvist G (2003) Fitness effects of female mate choice: preferred males are detrimental for Drosophila melanogasterfemales. Journal of Evolutionary Biology 16:797-811
Fuller RC (2003) Disentangling female mate choice and male competition in the rainbow darter, Etheostoma caeruleum. Copeia 1:138-148
Fuller RC, Houle D, Travis J (2005) Sensory bias as an explanation for the evolution of mate preferences. American Naturalist 166:437-446
Gabor C (1999) Association patterns of sailfin mollies (Poecilia latipinna): alternative hypotheses. Behavioral Ecology and Sociobiology 46:333-340
Galbrait MG (1967) Size-Selective Predation On Daphnia By Rainbow Trout And Yellow Perch. Transactions of the American Fisheries Society 96:1 -&
Garant D, Dodson JD, Bernatchez L (2005) Offspring genetic diversity increases fitness of female Atlantic salmon (Salmo salar). Behavioral Ecology and Sociobiology 57:240-244
Gavrilets S, Arnqvist G, Friberg U (2001) The evolution of female mate choice by sexual conflict. Proceedings of the Royal Society of London Series B-Biological Sciences 268:531-539
Gerhardt HC (1994) The Evolution Of Vocalization In Frogs And Toads. Annual Review of Ecology and Systematics 25:293-324
Gibson RM, Bachman GC (1992) The costs of female choice in a lekking bird. Behavioral Ecology 3:300-309
29 .

Godin JGJ, Briggs SE (1996) Female mate choice under predation risk in the guppy. Animal Behaviour 51:117-130
Goodwin NB, Balshine-Earn S, Reynolds JD (1998) Evolutionary transitions in parental care in cichlid fish. Proceedings of the Royal Society B: Biological Sciences 165
Goto A (1987) Polygyny in the river sculpin, Cottus hangiongensis (Pisces: Cottidae), with special reference to male mating success. Copeia 1:32-40
Goto A (1993) Male Mating Success And Female Mate Choice In The River Sculpin, Cottus-Nozawae (Cottidae). Environmental Biology of Fishes 37:347-353
Goulet D (1998) Spawning success in the damselfish Amblyglyphidodon leucogaster. the influence of eggs in the nest. Animal Behaviour 55:651-664
Gowaty PA, Steinichen R, Anderson WW (2002) Mutual interest between the sexes and reproductive success in Drosophila pseudoobscura. Evolution 56:2537-2540
Graeb BDS, Mangan MT, Jolley JC, Wahl DH, Dettmers JM (2006) Ontogenetic changes in preypreference and foraging ability of yellow perch: Insights based on relative energetic return of prey. Transactions of the American Fisheries Society 135:1493-1498
Grafen A (1990) Biological Signals As Handicaps. Journal of Theoretical Biology 144:517-546
Gray DA, Cade WH (1999) Quantitative genetics of sexual selection in the field cricket, Gryllus integer. Evolution 53:848-854
Green BS, McCormick Ml (2005) 02 replenishment to fish nests: males adjust brood care to ambient conditions and brood development. Behavioral Ecology and Sociobiology 16:389-397
Grether GF (2000) Carotenoid limitation and mate preference evolution: A test of the indicator hypothesis in guppies (Poecilia reticulata). Evolution 54:1712-1724
Griffth SC, Owens IPF, Burke T (1998) Sexual selection in the house sparrow: the best males are the worst fathers. (In preparation.)
Gross MR, Sargent RC (1985) The evolution of male and female parental care in fishes. American Zoologist 25:807-822
Guderley H, Couture P (2005) Stickleback fights: why do winners win? Influence of metabolic and morphometric parameters. Physiological and Biochemical Zoology 78:173-181
Gwynne DT (1991) Sexual competition among females: what causes courtship-role reversal? . Trends in Ecology & Evolution 6:118-121
Hagelin JC (2002) The kinds of traits involved in male-male competition: a comparison of plumage, behavior, and body size in quail. Behavioral Ecology 13:32-41
Hale RE, St Mary CM, Lindström K (2003) Parental responses to changes in costs and benefits along an environmental gradient. Environmental Biology of Fishes 67:107-116
Hall DJ, Cooper WE, Werner EE (1970) An experimental approach to the production dynamics and structure of freshwater animal communities. Limnology and Oceanography 15:829-928
30.

Hall DW, Kirkpatrick M, West B (2000) Runaway sexual selection when female preferences are directly selected. Evolution 54:1862-1869
Halupka K, Borowiec M (2006) Male whitethroats, Sylvia communis, advertise their future contribution to parental care. Behaviour 143:1-14
Hanssen SA, Erikstad KE, Johnsen V, Bustnes JO (2003) Differential investment and costs duringavian incubation determined by individual quality: an experimental study of the common eider (Somateria mollissima). Proceedings of the Royal Society of London Series B-Biological Sciences 270:531-537
Harari AR, Handler AM, Landolt PJ (1999) Size-assortative mating, male choice and female choice in the curculionid beetle Diaprepes abbreviatus. Animal Behaviour 58:1191-1200
Harshman LG, Prout T (1994) Sperm displacement without sperm transfer in Drosophila melanogasta. Evolution 48:758-766
Hastings PA (1988) Female Choice And Male Reproductive Success In The Angel Blenny, Coralliozetus-Angelica (Teleostei, Chaenopsidae). Animal Behaviour 36:115-124
Hegde SN, Krishna MS (1997) Size-assortative mating in Drosophila malerkotliana. Animal Behaviour 54:419-426
Heywood JS (1989) Sexual selection by the handicap mechanism. Evolution 43:1387-1397
Hill GE (1994) Trait Elaboration Via Adaptive Mate Choice - Sexual Conflict In The Evolution Of Signals Of Male Quality. Ethology Ecology & Evolution 6:351-370
Hoelzer G (1988) Filial Cannibalism In A Non-Brood Cycling Marine Fish. Environmental Biology of Fishes 21:309-313
Hoelzer GA (1989) The good parent process of sexual selection. Animal Behaviour 38:1067-1078
Hoelzer GA (1990) Male-male competition and female choice in the cortez damselfish, Stegastes rectifraenum. Animal Behaviour 40:339-349
Hoelzer GA (1992) The ecology and evolution of partial clutch cannibalism by paternal Cortez Damselfish. Oikos 65:113-120
Hoelzer GA (1995) Filial cannibalism and male parental care in damselfishes. Bulletin of Marine Science 57:663-671
Hoffman Jl, Forcada J, Trathan PN, Amos W (2007) Female fur seals show active choice for males that are heterozygous and unrelated. Nature 445:912-914
Holland B, Rice WR (1998) Perspective: Chase-away sexual selection: Antagonistic seduction versus resistance. Evolution 52:1-7
Holland B, Rice WR (1999) Experimental removal of sexual selection reverses intersexual antagonistic coevolution and removes a reproductive load. Proceedings of the National Academy of Sciences of the United States of America 96:5083-5088
Houde AE (1994) Effect of artificial selection on male colour patterns on mating preference of female guppies. Proceedings of the Royal Society B: Biological Sciences 256:125-130
31.

Houde AE, Endler JA (1990) Correlated evolution of female mating preferences and male color patterns in the guppy Poecilia reticulata. Science 248:1405-1408
Houde AE, Torio AJ (1992) Effect of Parasitic Infection on Male Color Pattern and Female Choice in Guppies. Behavioral Ecology 3:346-351
Houle D (1992) Comparing evolvability and variability of quantitative traits. Genetics 130:195-204
Houle D, Kondrashov AS (2002) Coevolution of costly mate choice and condition-dependent display of good genes. Proceedings of the Royal Society of London Series B-Biological Sciences 269:97-104
Houston AI, Szekely T, McNamara JM (2005) Conflict between parents over care. Trends in Ecology & Evolution 20:33-38
Howard DJ, Reece M, Gregory PG, Chu J, Cain ML (1998) The evolution of barriers to fertilization between closely related organisms. In: Howard DJ, Berlocher SH (eds) Endless forms: species and speciation. Oxford University Press, New York, pp 279-288
Hunt J, Brooks R, Jennions MD (2005) Female mate choice as a condition-dependent life-history trait. American Naturalist 166:78-92
Hunt J, Brooks R, Jennions MD, Smith MJ, Bentsen CL, Bussiere LF (2004) High-quality male field crickets invest heavily in sexual display but die young. Nature 432:1024-1027
Hyvarinen P, Huusko A (2006) Diet of brown trout in relation to variation in abundance and size of pelagic fish prey. Journal of Fish Biology 68:87-98
Irschick DJ, Garland TJ (2001) Integrating function and ecology in studies of adaptation: investigations of locomotor capacity as a model system. Annual Review of Ecology and Systematics 32:367- 396
Itzkowitz M, Makie D (1986) Food Provisioning And Foraging Behavior Of The Snowy Egret, Egretta- Thula. Biology of Behaviour 11:111-115
Ivlev VS (1961) Experimental ecology of the feeding of fishes. Yale University Press, New Haven, Connecticut
Iwasa Y, Pomiankowski A (1999) Good parent and good genes models of handicap evolution. Journal of Theoretical Biology 200:97-109
Iwasa Y, Pomiankowski A, Nee S (1991) The evolution of costly mate preferences. II. The ‘handicap’ principle. Evolution 45:1431-1442
Iyengar VK, Reeve HK, Eisner T (2002) Paternal inheritance of a female moth’s mating preference: consequences for sexual selection. Nature 419:830-832
Jennions MD, Backwell PRY, Passmore Nl (1995) Repeatability of mate choice: the effect of size in the African painted reed frog, Hyperolius marmoratus. Animal Behaviour 49:181-186
Jennions MD, Petrie M (1997) Variation in mate choice and mating preferences : a review of causes and consequences. Biological Review 72:283-327
Jennions MD, Polakow DA (2001) The effect of partial brood loss on male desertion in a cichlid fish: an experimental test. Behavioral Ecology 12:84-92
32.

Jivoff P (1997) The relative roles of predation and sperm competition on the duration of the post-copulatory association between the sexes in the blue crab, Callinectes sapidus. Behavioral Ecology and Sociobiology 40:175-185
Johnstone RA, Reynolds JD, Deutsch JC (1996) Mutual mate choice and sex differences in choosiness. Evolution 50:1382-1391
Jones JC, Reynolds JD (1999) Costs of egg ventilation for male common gobies breeding in conditions of low dissolved oxygen. Animal Behaviour 57:181-188
Jones TM, Quinnell RJ (2002) Testing predictions for the evolution of lekking in the sandfly, Lutzomyia longipalpis. Animal Behaviour 63:605-612
Kamler E (1992) Early life histories of fish: an energetics approach. Chapman & Hall, London
Karino K (1995) Male-male competition and female mate choice through courtship display in the territorial damselfish Stegastes nigricans. Ethology 100:126-138
Karino K (1997) Influence of brood size and offspring size on parental investment in a biparental cichlid fish, Neolamprologus moorii. Journal of Ethology 15:39-43
Karino K, Arai R (2006) Effect of clutch size on male egg-fanning behavior and hatching success in the goby, Eviota prasina (Klunzinger). Journal of Experimental Marine Biology and Ecology 334:43-50
Keenleyside MHA, Rangeley RW, Küppers BU (1985) Female mate choice and male parental defense behaviour in the cichlid fish Cichlasoma nigrofasciatum. Canadian Journal of Zoology 63:2489-2493
Keller L, Reeve HK (1995) Why do females mate with multiple males - the sexually selected sperm hypothesis. Advances in the Study of Behavior 24:291-315
Kemp DJ, Alcock J (2003) Lifetime resource utilization, flight physiology, and the evolution of contest competition in territorial insects. American Naturalist 162:290-301
Kinne O, Kinne EM (1962) Rates of development in embryos of a cyprinodont fish exposed to different temperature salinity-oxygen combinations. Canadian Journal of Zoology 40:231-&
Kirkpatrick M (1985) Evolution of female choice and male parental investment in polygynous species: the demise of the “sexy son”. American Naturalist 125:788-810
Kirkpatrick M (1987) The evolutionary forces acting on female mating preferences in polygynous animals. In: Bradbury JW, Andersson MB (eds) Sexual selection: testing the alternatives. Wiley, Chichester, UK, pp 67-82
Kirkpatrick M (1996) Good genes and direct selection in evolution of mating preferences. Evolution 50:2125-2140
Kirkpatrick M, Barton NH (1997) The strength of indirect selection on female mating preferences.Proceedings of the National Academy of Sciences of the United States of America 94:1282- 1286
Klug H, Chin A, St Mary CM (2005) The net effects of guarding on egg survivorship in the flagfish, Jordanella floridae. Animal Behaviour 69:661-668
33 .

Knapp RA, Kovach JT (1991) Courtship as an honest indicator of male parental quality in the bicolor damselfish, Stegastes partitus. Behavioral Ecology 2:295-300
Knapp RA, Warner RR (1991) Male parental care and female choice in the bicolor damselfish, Stegastes partitus: bigger is not always better. Animal Behaviour 41:747-756
Kodric-Brown A, Nicoletto P (2005) Courtship behaviour, swimming performance, and microhabitat use of Trinidadian guppies. Environmental Biology of Fishes 73:299-307
Kokko H (1998a) Good genes, old age and life-history trade-offs. Evolutionary Ecology 12:739-750
Kokko H (1998b) Should advertising parental care be honest? Proceedings of the Royal Society of London Series B-Biological Sciences 265:1871-1878
Kokko H (2001) Fisherian and "good genes" benefits of mate choice: how (not) to distinguish between them. Ecology Letters 4:322-326
Kokko H, Brooks R, Jennions MD, Morley J (2003) The evolution of mate choice and mating biases. Proceedings of the Royal Society of London Series B-Biological Sciences 270:653-664
Kokko H, Brooks R, McNamara JM, Houston AI (2002) The sexual selection continuum. Proceedings of the Royal Society of London Series B-Biological Sciences 269:1331-1340
Kokko H, Jennions M (2003) It takes two to tango. Trends in Ecology & Evolution 18:103-104
Kokko H, Jennions MD, Brooks R (2006) Unifying and testing models of sexual selection. Annual Review of Ecology Evolution and Systematics 37:43-66
Kokko H, Monaghan P (2001) Predicting the direction of sexual selection. Ecology Letters 4:159-165
Kolm K (2004) Female courtship in the Banggai cardinalfish: honest signals of egg maturity and reproductive output? Behavioral Ecology and Sociobiology 56:59-64
Kolm N (2001) Females produce larger eggs for large males in a paternal mouthbrooding fish.Proceedings of the Royal Society of London Series B-Biological Sciences 268:2229-2234
Koref-Santibanez S (2001) Effects of age and experience on mating activity in the sibling species Drosophila pavani and Drosophila gaucha. Behavior Genetics 31:287-297
Kotiaho J, Alatalo RV, Mappes J, Parri S (1997) Fighting success in relation to body mass anddrumming activity in the male wolf spider Hygrolycosa rubrofasciata. Canadian Journal of Zoology-Revue Canadienne De Zoologie 75:1532-1535
Kotiaho JS (2001) Costs of sexual traits: a mismatch between theoretical considerations and empirical evidence. Biological Reviews 76:365-376
Kotiaho JS, Simmons LW, Tomkins JL (2001) Towards a resolution of the lek paradox. Nature 410:684-686
Kraak SBM (1996) Female preference and filial cannibalism in Aidablennius sphynx (Teleostei, Blenniidae); A combined field and laboratory study. Behavioural Processes 36:85-97
Kraak SBM, Groothuis TGG (1994) Female Preference For Nests With Eggs Is Based On The Presence Of The Eggs Themselves. Behaviour 131:189-206
34.

Kraak SBM, Videler JJ (1991) Mate Choice In Aidablennius-Sphynx (Teleostei, Blenniidae) - Females Prefer Nests Containing More Eggs. Behaviour 119:243-266
Kramer DL (1987) Dissolved oxygen and fish behavior. Environmental Biology of Fishes 18:81-92
Kunzler R, Bakker TCM (2000) Pectoral fins and paternal quality in sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 267:999-1004
Kuwamura T, Yugo Y, Nakakshima Y (1993) Size-assortative monogamy and paternal egg care in a coral goby Paragobioon echinocephalus. Ethology 95:65-75
Kvarnemo C (1995) Size-Assortative Nest Choice In The Absence Of Competition In Males Of The Sand Goby, Pomatoschistus-minutus. Environmental Biology of Fishes 43:233-239
Kvarnemo C (1997) Food affects the potential reproductive rates of sand goby females, but not of males. Behavioral Ecology 8:605-611
Kvarnemo C, Ahnejsö I (1996) The dynamics of operational sex ratios and competition for mates. Trends in Ecology & Evolution 11:404-408
Kvarnemo C, Svensson O, Forsgren E (1998) Parental behaviour in relation to food availability in the common goby. Animal Behaviour 56:1285-1290
Lailvaux SP, Irschick DJ (2006) No evidence for female association with high-performance males in the green anole lizard, Anolis carolinensis. Ethology 112:707-715
Lande R (1980) Sexual dimorphism, sexual selection, and adaptation in polygenic characters. Evolution 34
Langmore NE, Davies NB, Hatchwell BJ, Hartley IR (1996) Female song attracts males in the alpine accentor Prunella collaris. Proceedings of the Royal Society of London Series B-Biological Sciences 263:141-146
Lavery RJ, Keenleyside MHA (1990) Parental Investment Of A Biparental Cichlid Fish, Cichlasoma- niqrofasciatum, In Relation To Brood Size And Past Investment. Animal Behaviour 40:1128- 1137
Leboeuf BJ, Mesnick S (1991) Sexual behavior of male Northern Elephant Seals 1. Lethal injuries to adult females. Behaviour 116:143-162
Lengkeek W, Didderen K (2006) Breeding cycles and reproductive behaviour in the river blenny Salaria fluviatilis. Journal of Fish Biology 69:1837-1844
Lesna I, Sabelis MW (1999) Diet-dependent female choice for males with ‘good genes’ in a soil predatory mite. Nature 401:581-584
Lessells CM (1999) Sexual conflicts in animals. In: Keller L (ed) Levels of Selection. Princeton University Press, Princeton, New Jersey, pp 75-99
Lifjeld JT, Dunn PO, Westneat DF (1994) Sexual selection by sperm competition in birds: male-male competition or female choice? Journal of Avian Biology 25:244-50
Lindström K (1988) Male-Male Competition For Nest Sites In The Sand Goby, Pomatoschistus- minutus. Oikos 53:67-73
35.

Lindström K (1998) Effects of costs and benefits of brood care on filial cannibalism in the sand goby. Behavioral Ecology and Sociobiology 42:101-106
Lindström K, Sargent RC (1997) Food access, brood size and filial cannibalism in the fantail darter, Etheostoma flabellare. Behavioral Ecology and Sociobiology 40:107-110
Linville SU, Breitwisch R, Schilling AJ (1998) Plumage brightness as an indicator of parental care in northern cardinals. Animal Behaviour 55:119-127
Lissaker M, Kvarnemo C (2006) Ventilation or nest defense - parental care trade-offs in a fish with male care. Behavioral Ecology and Sociobiology 60:864-873
Lissaker M, Kvarnemo C, Svensson O (2003) Effects of a low oxygen environment on parental effort and filial cannibalism in the male sand goby, Pomatoschistus minutus. Behavioral Ecology 14:374-381
Lopez P, Munoz A, Martin J (2002) Symmetry, male dominance and female mate preferences in the Iberian rock lizard, Lacerta monticola. Behavioral Ecology and Sociobiology 52:342-347
Lopez S (1999) Parasitized female guppies do not prefer showy males. Animal Behaviour 57:1129- 1134
Luo JG, Brandt SB, Klebasko MJ (1996) Virtual reality of planktivores: A fish's perspective of prey size selection. Marine Ecology - Progresses Series 140:271-283
Mackereth RW, Noakes DLG, Ridgway MS (1999) Size-based variation in somatic energy reserves and parental expenditure by male smallmouth bass, Micropterus dolomieu. Environmental Biology of Fishes 56:263-275
Maekawa K, Iguchi K, Katano O (1996) Reproductive success in male Japanese minnows,Pseudorasbora parva: Observations under experimental conditions. Ichthyological Research 43:257-266
Magnhagen C (1991) Predation risk as a cost of reproduction. Trends in Ecology & Evolution 6:183- 185
Manica A (2002a) Alternative strategies for a father with a small brood: mate, cannibalise or care. Behavioral Ecology and Sociobiology 51:319-323
Manica A (2002b) Filial cannibalism in teleost fish. Biological Reviews 77:261-277
Manica A (2003) The effect of brood size and age on partial filial cannibalism in the scissortail sergeant. Journal of Fish Biology 63:37-47
Manica A (2004) Parental fish change their cannibalistic behaviour in response to the cost-to-benefit ratio of parental care. Animal Behaviour 67:1015-1021
Mank JE, Promislow DEL, Avise JC (2005) Phylogenetic perspectives in the evolution of parental care in ray-finned fishes. Evolution 59:1570-1578
Marconato A, Bisazza A (1986) MALES WHOSE NESTS CONTAIN EGGS ARE PREFERRED BY FEMALE COTTUS-GOBIO L (PISCES, COTTIDAE). Animal Behaviour 34:1580-1582
Marconato A, Bisazza A, Fabris M (1993) The Cost Of Parental Care And Egg Cannibalism In The River Bullhead, Cottus-Gobio L (Pisces, Cottidae). Behavioral Ecology and Sociobiology 32:229-237
36.

Marden JH, Waage JK (1990) Escalated damselfly territorial contests are energetic wars of attrition. Animal Behaviour 39:954-959
Markman S, Yomtov Y, Wright J (1995) Male Parental Care In The Orange-Tufted Sunbird -Behavioral-Adjustments In Provisioning And Nest Guarding Effort. Animal Behaviour 50:655- 669
Marques PAM (2003) Male mate desertion in the Spanish sparrow Passer hispaniolensis. Ardea 91:245-250
Marques PAM (2004) Parental care, male desertion, and reproductive success in the Spanish sparrow, Passer hispaniolensis. Zoological Studies 43:123-131
Mazzi D (2004) Parasites make male pipefish careless. Journal of Evolutionary Biology 17:519-527
McLister JD (2003) The metabolic cost of amplexus in the gray tree frog (Hyla versicolor): assessing the energetics of male mating success. Canadian Journal of Zoology 81:388-394
McNamara JM, Szekely T, Webb JN, Houston AI (2000) A dynamic game-theoretic model of parental care. Journal of Theoretical Biology 205:605-623
Mehla J, Sheldon BC (2000) Lifetime reproductive success and heritability in nature. American Naturalist 155:301-310
Mihelakakis A, Kitajima C (1994) Effects of salinity and temperature on incubation period, hatching rate, and morphogenesis of the silver sea bream, Sparus sarba (Forskal, 1775). Aquaculture 126:361-371
Mitchell DP, Dunn PO, Whittingham LA, Freeman-Gallant CR (2007) Attractive males provide less parental care in two populations of the common yellowthroat. Animal Behaviour 73:165-170
Möller AP (1993) Female Preference For Apparently Symmetrical Male Sexual Ornaments In The Barn Swallow Hirundo-Rustica. Behavioral Ecology and Sociobiology 32:371-376
Möller AP, Alatalo RV (1999) Good-genes effect in sexual selection. Proceedings of the Royal Society of London Series B-Biological Sciences 266:85-91
Möller AP, Thornhill R (1998) Male parental care, differential parental investment by females and sexual selection. Animal Behaviour 55:1507-1515
Montgomerie RD, Weatherhead PJ (1988) Risks and rewards of nest defense by parent birds. Quarterly Review of Biology 63:167-187
Moore AJ, Gowaty PA, Wallin WG, Moore PJ (2001) Sexual conflict and the evolution of female mate choice and male social dominance. Proceedings of the Royal Society of London Series B- Biological Sciences 268:517-523
Moore AJ, Moore PJ (1999) Balancing sexual selection through opposing mate choice and male competition. Proceedings of the Royal Society of London Series B-Biological Sciences 266:711-716
Morrison C, Hero JM, Smith WP (2001) Mate selection in Litoria chloris and Litoria xanthomera: Females prefer smaller males. Austral Ecology 26:223-232
Mountjoy DJ, Lemon RE (1997) Male song complexity and parental care in the European starling. Behaviour 134:661-675
37.

Munk P (1992) Foraging Behaviour and prey size spectra of larval herring Clupea harengus. Marine Ecology - Progresses Series 80:149-158
Neat FC, Taylor AC, Huntingford FA (1998) Proximate costs of fighting in male cichlid fish: the role of injuries and energy metabolism. Animal Behaviour 55:875-882
Neff BD (2003) Decisions about parental care in response to perceived paternity. Nature 422:716
Neff BD, Gross MR (2001) Dynamic adjustment of parental care in response to perceived paternity. Proceedings of the Royal Society of London Series B-Biological Sciences 268:1559-1565
Nelson CM, Nolen TG (1997) Courtship song, male agonistic encounters, and female mate choice in the house cricket, Acheta domesticus (Orthoptera: Gryllidae). Journal of Insect Behavior 10:557-570
Nicoletto PF (1995) Offspring quality and female choice in the guppy, Poecillia reticulata. Animal Behaviour 49:377-387
Nilsson SO, Nilsson GE (2000) Free choice by female sticklebacks: lack of preference for maledominance traits. Canadian Journal of Zoology-Revue Canadienne De Zoologie 78:1251-1258
Noonan K (1983) Female mate choice in a cichlid fish Cichlasoma nigrofasciatum. Animal Behaviour 31:1005-1010
Nyman KJ (1953) Observations on the behaviour of Gobius microps. Acta Societatis po Fauna et Flora Fennica 69:1-11
Okuda N (1999) Sex roles are not always reversed when the potential reproductive rates is higher in females. American Naturalist 153:540-548
Okuda N, Ito S, Iwao H (2002) Female spawning strategy in Rhinogobius sp OR: how do females deposit their eggs in the nest? Ichthyological Research 49:371-379
Okuda N, Yanagisawa Y (1996) Filial cannibalism by mouthbrooding males of the cardinal fish,Apogon doederleini, in relation to their physical condition. Environmental Biology of Fishes 45:397-404
Oppenheimer JR (1970) Mouthbrooding in fishes. Animal Behaviour 52:307-314
Osorno JL, Szekely T (2004) Sexual conflict and parental care in magnificent frigatebirds: full compensation by deserted females. Animal Behaviour 68:337-342
Ostlund-Nilsson S (2001) Fifteen-spined stickleback (Spinachia spinachia) females prefer males with more secretional threads in their nests: an honest-condition display by males. Behavioral Ecology and Sociobiology 50:263-269
Östlund-Nilsson S, Nilsson GE (2004) Breathing with a mouth full of eggs: respiratory consequences of mouthbrooding in cardinalfish. Proceedings of the Royal Society of London Series B- Biological Sciences 271:1015-1022
Östlund S, Ahnesjö I (1998) Female fifteen-spined sticklebacks prefer better fathers. Animal Behaviour 56:1177-1183
Pampoulie C, Lindstrom K, St Mary CM (2004) Have your cake and eat it too: male sand gobies show more parental care in the presence of female partners. Behavioral Ecology 15:199-204
38 .

Parker GA (1974) Assessment strategy and the evolution of fighting behaviour. Journal of Theoretical Biology 47:223-243
Parker GA, Royle NJ, Hartley IR (2002) Intrafamilial conflict and parental investment: a synthesis. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 357:295-307
Parker GA, Simmons LW (1996) Parental investment and the control of sexual selection: predicting the direction of sexual competition. Proceedings of the Royal Society of London Series B- Biological Sciences 263:315-321
Parker TH, Garant D (2004) Quantitative genetics of sexually dimorphic traits and capture of genetic variance by a sexually-selected condition-dependent ornament in red junglefowl (Gallus gallus). Journal of Evolutionary Biology 17:1277-1285
Parker TH, Ligon JD (2003) Female mating preferences in red junglefowl: a meta-analysis. Ethology Ecology & Evolution 15:63-72
Parri S, Alatalo RV, Mappes J (1998) Do female leaf beetles Galerucella nymphaeae choose their mates and does it matter? Oecologia 114:127-132
Partridge L, Fowler K (1990) Non-mating costs of exposure to males in female Drosophila melanogaster Journal of Insect Physiology 36:419-425
Payne AG, Smith C, Campbell AC (2002) Filial cannibalism improves survival and development of beaugregory damselfish embryos. Proceedings of the Royal Society of London Series B- Biological Sciences 269:2095-2102
Payne AG, Smith C, Campbell AC (2004) A model of oxygen-mediated filial cannibalism in fishes. Ecological Modelling 174:253-266
Perry G, Levering K, Girard l, Garland TJ (2004) Locomotor performance and dominance in male Anolis cristatellus. Animal Behaviour 67:37-47
Peters A, Michiels NK (1996) Evidence for lack of inbreeding avoidance by selective simultaneous hermaphrodite. Invertebrate Biology 115:99-103
Petersen CW (1990) The occurrence and dynamics of clutch loss and filial cannibalism in twoCaribbean damselfishes. Journal of Experimental Marine Biology and Ecology 135:117-133
Petrie M (1983) Female moorhens compete for small fat males. Science 220:413-415
Pianka ER (1976) Natural selection of optimal reproductive tactics. American Zoologist 16:775-784
Pilastro A, Evans JP, Sartorelli S, Bisazza A (2002) Male phenotype predicts insemination success in guppies. Proceedings of the Royal Society of London Series B-Biological Sciences 269:1325- 1330
Pitnick S, Markow TA (1994) Male gametic strategies - Sperm size, testes size, and the allocation of ejaculate among successive mates by the sperm-limited fly Drosophila-Pachea and its relatives. American Naturalist 143:785-819
Plaistow S, Siva-Jothy MT (1996) Energetics, constraints, and male mate-securing tactics in thedamselfly Calopteryx splendens xanthostoma (Charpentier). Proceedings of the Royal Society of London Series B-Biological Sciences 263:1233-1238
39.

Polak M, Starmer WT (1998) Parasite-induced risk of mortality elevates reproductive effort in male Drosophila. Proceedings of the Royal Society of London Series B-Biological Sciences 265:2197-2201
Polis GA (1981) THE EVOLUTION AND DYNAMICS OF INTRASPECIFIC PREDATION. Annual Review of Ecology and Systematics 12:225-251
Pomfret J, Knell R (2006) Sexual selection and horn allometry in the dung beetle Euoniticellus intermedius. Animal Behaviour 71:567-576
Pomiankowski A (1987) The costs of choice in sexual selection. Journal of Theoretical Biology 128:195-218
Pomiankowski A (1988) The evolution of female mating preferences for male genetic quality. Oxford Surveys in Evolutionary Biology 5:136-184
Pomiankowski A, Iwasa Y, Nee S (1991) The evolution of costly mate preferences. I. Fisher and biased mutation. Evolution 45:1422-1430
Pomiankowski A, Möller AP (1995 ) A resolution of the lek paradox. Proceedings of the Royal Society B: Biological Sciences 260:21-29
Poulin R (1994) Mate choice decisions by parasitized female upland bullies, Gobiomorphus breviceps. Proceedings of the Royal Society of London Series B-Biological Sciences 256:183-187
Prasad NG, Bedhomme S, Day T, Chippindale AK (2007) An evolutionary cost of separate genders revealed by male-limited evolution American Naturalist 169:29-37
Preston BT, Stevenson IR, Pemberton JM, Wilson K (2001) Dominant rams lose out by spermdepletion - A waning success in siring counters a ram's high score in competition for ewes. Nature 409:681-682
Price T, Schlüter D, Heckman NE (1993) Sexual selection when the female directly benefits. Biological Journal of the Linnean Society 48:187-211
Queller DC (1997) Why do females care more than males? Proceedings of the Royal Society of London Series B-Biological Sciences 264:1555-1557
Qvarnström A (1997) Experimentally increased badge size increases male competition and reduces male parental care in the collared flycatcher. Proceedings of the Royal Society of London Series B-Biological Sciences 264:1225-1231
Qvarnstrom A, Forsgren E (1998) Should females prefer dominant males? Trends in Ecology & Evolution 13:498-501
Radesater T, Halldorsdottir H (1993) 2 Male types of the common earwig - male-male competition and mating success. Ethology 95:89-96
Raouf SA, Parker PG, Ketterson ED, Nolan V, Ziegenfus C (1997) Testosterone affects reproductive success by influencing extra-pair fertilizations in male dark-eyed juncos (Aves: Junco hyemalis). Proceedings of the Royal Society of London Series B-Biological Sciences 264:1599-1603
Reebs SG, Whoriskey FG, FitzGerald GJ (1984) DIEL PATTERNS OF FANNING ACTIVITY, EGGRESPIRATION, AND THE NOCTURNAL BEHAVIOR OF MALE 3-SPINED STICKLEBACKS,
40 .

GASTEROSTEUS-ACULEATUS L (F-TRACHURUS). Canadian Journal of Zoology-Revue Canadienne De Zoologie 62:329-334
Reichard M, Bryja J, Ondrackova M, Davidova M, Kaniewska P, Smith C (2005) Sexual selection for male dominance reduces opportunities for female mate choice in the European bitterling (Rhodeus sericeus). Molecular Ecology 14:1533-1542
Reimchen TE (1989) Loss of nuptial color in threespine sticklebacks (Gasterosteus aculeatus). Evolution 43:450-460
Reynolds JD, Goodwin NB, Freckleton RP (2002) Evolutionary transitions in parental care and live bearing in vertebrates. Philosophical Transactions of the Royal Society of London Series B- Biological Sciences 357:269-281
Reynolds JD, Jones JC (1999) Female preference for preferred males is reversed under low oxygen conditions in the common goby (Pomatoschistus microps). Behavioral Ecology 10:149-154
Rice WR (1984) Sex chromosomes and the evolution of sexual dimorphism. Evolution 38:735-742
Rice WR, Chippindale AK (2001) Intersexual ontogenetic conflict. Journal of Evolutionary Biology 14:685-693
Rintämaeki PT, Lundberg A, Alatalo RV, Höeglund J (1998) Assortative mating and female clutch investment in black grouse. Animal Behaviour 56:1399-1403
Robson MA, Miles DB (2000) Locomotor performance and dominance in tree lizards, Urosaurus ornatus. Functional Ecology 14:338-344
Rodd FH, Hughes KA, Grether GF, Baril CT (2002) A possible non-sexual origin of mate preference: are male guppies mimicking fruit? Proceedings of the Royal Society of London Series B- Biological Sciences 269:475-481
Rogers W, Barlow GW (1991) Sex differences in mate choice in a monogamous biparental fish, the midas cichlid (Cichlasoma citrinellum). Ethology 87:249-261
Rohwer S (1978) Parent cannibalism of offspring and egg raiding as a courtship strategy. American Naturalist 112:429-440
Rowe L, Arnqvist G (1996) Analysis of the causal components of assortative mating in water striders. Behavioural Ecology and Sociobiology 38:279-286
Rowe L, Cameron E, Day T (2005) Escalation, retreat, and female indifference as alternative outcomes of sexually antagonistic coevolution. American Naturalist 165:S5-18
Rowe L, Houle D (1996) The lek paradox and the capture of genetic variance by condition dependent traits. Proceedings of the Royal Society B: Biological Sciences 263:1415-1421
Ryan MJ, Rand AS (1990) The Sensory Basis Of Sexual Selection For Complex Calls In The Tungara Frog, Physalaemus-Pustulosus (Sexual Selection For Sensory Exploitation). Evolution 44:305-314
Sakaluk SK, Belwood JJ (1984) Gecko phonotaxis to cricket calling song: a case of satellite predation. Animal Behaviour 32:659-662
41.

Sanz JJ, Kranenbarg S, Tinbergen JM (2000) Differential response by males and females tomanipulation of partner contribution in the great tit (Parus major). Journal of Animal Ecology 69:74-84
Sargent RC (1985) Territoriality and reproductive tradeoffs in the 3-spine stickleback, Gasterosteus- aculeatus. Behaviour 93:217-226
Sargent RC (1988) Paternal care and egg survival both increase with clutch size in the fathead minnow, Pimephales-promelas. Behavioral Ecology and Sociobiology 23:33-37
Sargent RC (1992) Ecology of filial cannibalism in fish: theoretical perspectives. In: Elgar MAC, B. J.(ed) Cannibalism: ecology and evolution among diverse taxa. Oxford University Press, pp 38- 62
Sargent RC (1997) Parental care. In: Godin JGJ (ed) Behavioural Ecology of Teleost Fishes. Oxford University Press, Oxford, pp 292-315
Sargent RC, Crowley PH, Huang C, Lauer M, Neergaard D, Schmoetzer L (1995) A dynamic program for male parental care in fishes: Brood cycling and filial cannibalism. Behaviour 132:1059- 1078
Sargent RC, Gebier JB (1980) Effects of nest site concealment on hatching success, reproductive success, and paternal behavior of the threespine stickleback, Gasterosteus-aculeatus. Behavioral Ecology and Sociobiology 7:137-142
Sargent RC, Gross MR (1985) Parental investment decision rules and the concorde fallacy. Behavioral Ecology and Sociobiology 17:43-45
Sargent RC, Gross MR (1986) Williams' principle: An explanation of parental care in teleost fishes. In: Pitcher TJ (ed) The Behaviour of Teleost Fish. Crrom Helm, London, pp 275-293
Schaffer NM (1974a) Optimal reproductive effort in fluctuating environments. American Naturalist 108:783-798
Schaffer NM (1974b) Selection for optimal life histories: The effects of age structure. Ecology 55:291- 303
Schwagmeyer PL, Mock DW, Parker GA (2002) Biparental care in house sparrows: negotiation or sealed bid? Behavioral Ecology 13:713-721
Seko T, Nakasuji F (2004) Effect of egg size variation on survival rate, development and fecundity of offspring in a migrant skipper, Parnara guttata guttata (Lepidoptera : Hesperiidae). Applied Entomology and Zoology 39:171-176
Semler DE (1971) Some aspects of adaptation in a polymorphism for breeding colours in Threespine Stickleback (Gasterosteus-Aculeatus). Journal of Zoology 165:291 -&
Senar JC, Figuerola J, Pascual J (2002) Brighter yellow blue tits make better parents. Proceedings of the Royal Society of London Series B-Biological Sciences 269:257-261
Sevenster P (1961) A causal analysis of a displacement activity (fanning in Gasterosteus aculeatus L.). Behaviour 9 (Suppl.)
Shackleton MA, Jennions MD, Hunt J (2005) Fighting success and attractiveness as predictors of male mating success in the black field cricket, Teleogryllus commodus: the effectiveness of no-choice tests. Behavioral Ecology and Sociobiology 58:1-8
42 .

Sheldon BC (2000) Differential allocation: tests, mechanisms and implications. Trends in Ecology and Evolution 15:397-402
Shibata J, Kohda M (2006) Seasonal sex role changes in the blenniid Petroscirtes breviceps, a nest brooder with paternal care. Journal of Fish Biology 69:203-214
Silva K, Monteiro NM, Vieira MN, Almada VC (2006) Reproductive behaviour of the black-stripedpipefish Syngnathus abaster (Pisces; Syngnathidae). Journal of Fish Biology 69:1860-1869
Simmons LW (1992) Quantification of role reversal in relative parental investment in a bush cricket. Nature 358:61-63
Simmons LW, Parker GA (1989) Nuptial feeding in insects - mating effort versus paternal investment. Ethology 81:332-343
Skolbekken R, Utne-Palm AC (2001) Parental investment of male two-spotted goby, Gobiusculus flavescens (Fabricius). Journal of Experimental Marine Biology and Ecology 261:137-157
Slagsvold T, Lifjeld JT (1988) Plumage color and sexual selection in the pied flycatcher ficedula- hypoleuca. Animal Behaviour 36:395-407
Sloman KA, Motherwell G, O’Connor Kl, Taylor AC (2000) The effect of social stress on the standard metabolic rate (SMR) of brown trout, Salmo trutta. Fish Physiology and Biochemistry 23:49-53
Smiseth PT, Amundsen T (2000) Does female plumage coloration signal parental quality? A male removal experiment with the bluethroat (Luscinia s. svecica). Behavioral Ecology and Sociobiology 47:205-212
Smiseth PT, Ornborg J, Andersson S, Amundsen T (2001) Is male plumage reflectance correlated with paternal care in bluethroats? Behavioral Ecology 12:164-170
Smith C (1992) Filial cannibalism as a reproductive strategy in care-giving teleosts. Netherlands Journal of Zoology 42:607-613
Smith C, Wootton RJ (1995) THE COSTS OF PARENTAL CARE IN TELEOST FISHES. Reviews in Fish Biology and Fisheries 5:7-22
Soler JJ, de Neve L, Martinez JG, Soler M (2001) Nest size affects clutch size and the start of incubation in magpies: an experimental study. Behavioral Ecology 12:301-307
Spence R, Smith C (2006) Mating preference of female zebrafish, Danio rerio, in relation to male dominance. Behavioral Ecology 17:779-783
Spoor WA (1984) Oxygen requirements of larvae of the smallmouth bass, Micropterus dolomieui Lacepede. Journal of Fish Biology 25:587-592
St Mary CM, Noureddine CG, Lindström K (2001) Environmental effects on male reproductive success and parental care in the Florida flagfish Jordanella floridae. Ethology 107:1035-1052
Stanley TR (2002) How many kilojoules does a Black-billed Magpie nest cost? Journal of Field Ornithology 73:292-297
Stott MK, Poulin R (1996) Parasites and parental care in male upland bullies (Eleotridae). Journal of Fish Biology 48:283-291
43.

Stutt AD, Siva-Jothy MT (2001) Traumatic insemination and sexual conflict in the bed bug Cimexlectularius. Proceedings of the National Academy of Sciences of the United States of America 98:5683-5687
Suk HY, Choe JC (2002) The presence of eggs in the nest and female choice in common freshwater gobies (Rhinogobius brunneus). Behavioral Ecology and Sociobiology 52:211-215
Summers K, Weigt LA, Boag P, Bermingham E (1999) The evolution of female parental care in poison frogs of the genus Dendrobates: evidence form mitochondrial DNA. Herpetologica 55:254- 270
Svensson OLA, Magnhagen C, Forsgren E, Kvarnemo C (1998) Parental behaviour in relation to the occurrence of sneaking in the common goby. Animal Behaviour 56:175-179
Svensson PA, Pelabon C, Blount JD, Surai PF, Amundsen T (2006) Does female nuptial coloration reflect egg carotenoids and clutch quality in the Two-Spotted Goby (Gobiusculus flavescens, Gobiidae)? Functional Ecology 20:689-698
Syriatowicz A, Brooks R (2004) Sexual responsiveness is condition-dependent in female guppies, but preference functions are not. BMC Ecology 4
Takahashi D, Kohda M (2004) Courtship in fast water currents by a male stream goby (Rhinogobius brunneus) communicates the parental quality honestly. Behavioral Ecology and Sociobiology 55:431-438
Takegaki T, Nakazono A (1999) Responses of the egg-tending gobiid fish Valenciennes longipinnisXo the fluctuation of dissolved oxygen in the burrow. Bulletin of Marine Science 65:815-823
Tallamy DW (2000) Sexual selection and the evolution of exclusive paternal care in arthropods. Animal Behaviour 60:559-567
Taylor PD, Williams GC (1982) The lek paradox is not resolved. Theoretical Population Biology 22:392-409
Telia JL, Forero MG, Donazar JA, Hiraldo F (1997) Is the expression of male traits female lesser kestrels related to sexual selection? Ethology 103:72-81
Thresher RE (1984) Reproduction in reef fishes. T. F. H. Publications, Neptuen City, New Jersey
Tilson RL (1981) Family formation strategies of Kloss's gibbons. Folia Primatologica 35:259-281
Tinbergen N (1951) The Study of Instinct. Oxford University Press, London
Tokarz RR (2002) An experimental test of the importance of the dewlap in male mating success in the lizard Anolis sagrei. Herpetologica 58:87-94
Tomkins JL, Radwan J, Kotiaho JS, Tregenza T (2004) Genic capture and resolving the lek paradox. Trends in Ecology & Evolution 19:323-328
Torricelli P, Lugli M, Gandolfi G (1985) A quantitative-analysis of the fanning activity in the male Padogobius martensi (pisces, gobiidae). Behaviour 92:288-301
Tregenza T, Wedell N (1997) Definitive evidence for cuticular pheromones in a cricket. Animal Behaviour 54:979-984
44 .

Tregenza T, Wedell N (2000) Genetic compatibility, mate choice and patterns of parentage: Invited review. Molecular Ecology 9:1013-1027
Trivers R (1985) Social Evolution. The Benjamin/Cummings Publishing Company, New York
Trivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Sexual Selection and the Descent of man. Aldine Publishing Company, London, pp 136-179
Unger LM, Sargent RC (1988) Allopaternal care in the fathead minnow, Pimephales promelas - Females prefer males with eggs. Behavioral Ecology and Sociobiology 23:27-32
Vahed K (1998) The function of nuptial feeding in insects: a review of empirical studies. Biological Reviews 73:43-78
van lersel JJA (1953) An analysis of parental behaviour of the male three spined stickleback (Gasterosteus aculeatus L.). Behaviour 3 (Suppl.):1-159
Vincent A, Ahnejsö I, Berglund A, Rosenqvist G (1992) Pipefishes and seahorses: are they all sex role reversed? Trends in Ecology & Evolution 7:237-241
Vinyoles D, Cöte IM, de Sostoa A (1999) Egg cannibalism in river blennies: the role of natural prey availability. Journal of Fish Biology 55:1223-1232
Voltura KM, Schwagmeyer PL, Mock DW (2002) Parental feeding rates in the house sparrow, Passer domesticus: Are larger-badged males better fathers? Ethology 108:1011-1022
Wagner WEJ (1998) Measuring female mating preferences. Animal Behaviour 55:1029-1042
Watson PJ, Arnqvist G, Stallmann RR (1998) Sexual conflict and the energetic costs of mating and mate choice in water striders. American Naturalist 151:46-58
Webb JN, Houston AI, McNamara JM, Szekely T (1999) Multiple patterns of parental care. Animal Behaviour 58:983-993
Webster MM, Hart PJB (2004) Substrate discrimination and preference in foraging fish. Animal Behaviour 68:1071-1077
Wedell N, Kvarnemo C, Lessells CM, Tregenza T (2006) Sexual conflict and life histories. Animal Behaviour 71:999-1011
West-Eberhard MJ (1984) Sexual selection, competitive communication and species-specific signals in insects. In: Lewis T (ed) Insect Communication Academic Press, London, pp 283-324
Westneat DF, Sherman PW, Morton ML (1990) The ecology and evolution of extra-pair copulations in birds. Current Ornithology 7:331-369
Weygoldt P (1987) Evolution of parental care in dart poison frogs (Amphibia: Anura: Dendrobatidae). Z. Zool. Syst. Evol. 25:51-67
White DJ, King AP, West MJ (2002) Facultative development of courtship and communication in juvenile male cowbirds (Molothrus ater). Behavioral Ecology 13:487-496
Whittingham LA, Dunn PO, Robertson RJ (1994) Female response to reduced male parental care in birds - an experiment in tree swallows. Ethology 96:260-269
45.

Wiley RH, Poston J (1996) Indirect mate choice, competition for mates, and coevolution of the sexes. Evolution 50:1371-1381
Williams GC (1966a) Adaptation and natural selection. Princeton University Press, Princeton
Williams GC (1966b) Natural selection costs of reproduction and a refinement of Lacks principle. American Naturalist 100:687-&
Winkler DW (1987) A general model for parental care. American Naturalist 130:526-543
Wong BBM (2004) Superior fighters make mediocre fathers in the Pacific blue-eye fish. Animal Behaviour 67:583-590
Wong BBM, Candolin U (2005) How is female choice affected by male competition? Biological Review 80:559-571
Wootton RJ (1998) Ecology of teleost fishes. Kluwer, Dordrecht
Wright J, Cuthill I (1989) Manipulation of sex-differences in parental care. Behavioral Ecology and Sociobiology 25:171 -181
Yenisetti SC, Hegde SN (2003) Size-related mating and reproductive success in a drosophilid: Phorticella striata. Zoological Studies 42:203-210
Zahavi A (1975) Mate selection: a selection for a handicap. Journal of Theoretical Biology 53:205-214
Zeh JA, Zeh DW (1996) The evolution of polyandry I: Intragenomic conflict and genetic incompatibility. Proceedings of the Royal Society of London Series B-Biological Sciences 263:1711-1717
Zeh JA, Zeh DW (1997) The evolution of polyandry 2: Post-copulatory defences against genetic incompatibility. Proceedings of the Royal Society of London Series B-Biological Sciences 264:69-75
Zink AG (2003) Quantifying the costs and benefits of parental care in female treehoppers. Behavioral Ecology 14:687-693
Zoran MJ, Ward JA (1983) Parental egg care behavior and fanning activity for the orange chromide, Etroplus maculatus. Environmental Biology of Fishes 8:301-310
46.

CHAPTER 2 Mate choice in the northern trout gudgeon
(Mogurnda mogurnda)
2.1 Introduction
2.1.1. Female mating preferences
In many taxa, females appear to choose between potential sexual partners. How and why mate
choice evolves remains elusive in most cases (Andersson 1994). In general, mating can be costly (Daly
1978), and mate choice often aggravates these costs. The costs of mating include suffering an increased
risk of predation (Sakaluk and Belwood 1984; Magnhagen 1991), and time and energy costs while
searching for males (e.g., Alatalo et al. 1988; Gibson and Bachman 1992). The risk of predation can also
be greater on copulating pairs, sexually transmitted disease can have negative fitness consequences, and
mate searching and copulation may hinder activities unrelated to mating such as foraging (e.g., Keller and
Reeve 1995; Jivoff 1997).
Discriminating among males increases the costs associated with mating and, in the absence of
benefits, should lead to selection against female mating biases (Bulmer 1989). For female choice to
persist there must be benefits to compensate for any costs associated with mate choice, otherwise it
would be more beneficial to mate randomly (Friberg and Arnqvist 2003). One of the most straightforward
explanations for the evolution of female choice is that it results from evolutionary processes that favour an
increase in female lifetime reproductive success (e.g., increased lifespan or increased egg production)
compared to mating non-randomly (Heywood 1989; Hoelzer 1989; Price et al. 1993; Kirkpatrick 1996;
Iwasa and Pomiankowski 1999).
2.1.2. Life history traits and female mating preferences
Recent studies have examined sexual selection from a functional perspective, with a particular
focus on the correlations between physiological performance, secondary sexual traits, male-male
competition and female choice. In a recent review, Lailvaux and Irschick (2006) summarised this emerging
field of study and asked two key questions: Are males who enjoy increased mating success also of high
physiological or phenotypic quality? And, are these males consequently good fighters? Lailvaux and
47 .

Irschick (2006) define two physiological measures; whole organism performance capacity and
physiological state. Whole organism performance capacity (“performance”) encompasses how well an
individual performs an ecologically relevant task, e.g., how fast a fish can swim or even how many
offspring a female can produce in a year (Irschick and Garland 2001). Physiological state (“state”) refers to
the measure of the physiological processes of an individual (e.g., metabolic rates or muscle function). In
sexual selection studies, a male’s “condition” is often an important variable and condition is said to be
encompassed within physiological state in the review.
Male secondary sexual characteristics are generally considered to be costly to produce and/or
maintain (Kotiaho 2001), particularly male vocalisations. For example in frogs and toads, calling males
have a metabolic rate exceeding a factor of 100-220% of the baseline rate (see Gerhardt 1994) and this
huge investment in energetics may reduce their future reproductive encounters (McLister 2003). Studies
have also shown that sexual displays are highly sensitive to male condition (Cotton et al. 2004;
Bonduriansky and Rowe 2005). Logically then, males in better condition can afford to invest more into
sexual displays or secondary sexual characteristics but this does not mean that their physiological state
(post-investment) will be positively correlated with sexual display (Grafen 1990; Kokko and Monaghan
2001; Hunt et al. 2004a; Tomkins et al. 2004). Male may need to be in good condition to produce sexual
advertisement but sexual advertisement may trade off against other functional and life history traits.
Recent studies show that males who heavily allocate resources into sexual advertisement suffer a
reduction in other fitness costs, such as survival, and that the optimal returns are sometimes such that the
males will the most resources invest so heavily into sexual displays that they actually have shorter
lifespans (Hunt et al. 2004).
2.1.3. Male dominance and female mating preferences
So are dominant (and assumedly more reproductively successful males) of higher physiological
quality? Physiology has been implicit in some models of male-male competition (e.g., resource holding
potential: Parker 1974) but the underlying physiological basis of why some males are better than others
has rarely been addressed. High metabolic rates are assumed to be a cost of male dominance (Roskaft et
al. 1986; Bryant and Newton 1994; Metcalfe et al. 1995; Whiting et al. 2003) but this may not always be
the case. Subordinate males may be involved in more challenges than dominant males and therefore
forced to maintain a higher metabolic rate (Sloman et al. 2000). Dominant males may not even suffer
higher metabolic costs of fighting. For example in cichlids, there are relatively higher levels of lactate and
48 .

other end product in males that lose fights (Neat et al. 1998). Some performance measures have been
shown to positively predict success in male-male competition, such as locomotor performance in lizards
(Robson and Miles 2000; Perry et al. 2004). In male sticklebacks, muscle condition is positively linked to
fighting ability (Guderley and Couture 2005). Males with higher energetic reserves are more likely to be
successful in male-male interactions in several animal taxa, such as insects (Marden and Waage 1990;
Plaistow and Siva-Jothy 1996; Blanckenhorn et al. 2003; Kemp and Alcock 2003), birds (Petrie 1983) and
fish (Chellappa and Huntingford 1989; Neat et al. 1998).
Weapon size is often a good predictor of fight outcome. For example, male dung beetles with
larger horns are more successful in male-male interactions (Pomfret and Knell 2006). Males have the
opportunity during fights to assess a rival’s horn size (Pomfret and Knell 2006) and probably use that as
an indicator of their fighting ability. Females have also shown preferences for to male traits correlated with
dominance, such as colouration in three spine sticklebacks Gasterosteus aculeatus (Candolin 1999a) and
body size in Japanese medaka Oryzias latipes (Howard et al. 1998). In fish, there is a wide literature
showing females prefer larger males in species that show biparental or paternal care (Downhower and
Brown 1980; Noonan 1983; Keenleyside et al. 1985; Goto 1987; Bisazza and Marconato 1988; Cote and
Hunte 1989; Rogers and Barlow 1991), but although often assumed to, females do not always prefer
males that are better fighters (e.g., Moore and Moore 1999; Reichard et al. 2005; but see Wong and
Candolin 2005). Overall, there is a large variation across taxa in the fitness outcomes for females of
mating with dominant males (Qvarnstrom and Forsgren 1998; Moore et al. 2001; Wong and Candolin
2005).
There is evidence that females prefer males with increased performance capacities. For example,
female guppies prefer male ornaments that are correlated with swimming ability (Nicoletto 1991; 1993;
Kodric-Brown and Nicoletto 2005), although interestingly there was no evidence of indirect genetic
benefits in the form of increased performance in offspring (Nicoletto 1995; Evans et al. 2004). On the
other hand, there was no documented female preference for higher performing males in the lizard Anolis
carolinensis (Lailvaux and Irschick 2006). Insufficient data prohibits any firm conclusions as to general
trends, however, and this field of study needs to be developed further.
Female choice might, however, select for males in better physiological condition, as predicted by
the handicap hypothesis (Zahavi 1975) and this trend is prevalent across taxa. In some fish species, only
males in good condition are able to court females in fast water (Takahashi and Kohda 2004; Kodric-Brown
and Nicoletto 2005). In birds, comb size in red jungle fowl is heritable, correlated with an index of (Parker
49.

and Garant 2004) condition and a trait used as a mate choice cue (Parker and Ligon 2003). In insects,
yellow dung flies with relatively higher lipid and glucose levels achieve higher mating success
(Blanckenhorn et al. 2003). Female choice may indirectly select for males of good physiological quality
and thereby also select for condition-dependence of male traits (Bonduriansky and Rowe 2005).
2.1.4. Parental Care and Female Mating Preferences
Another aspect of male performance that female choice has been shown to select for is parental
care. In fish, females have been show to prefer many traits, such as courtship (Knapp and Kovach 1991;
Knapp and Warner 1991; Karino 1995), nest characteristics (Hoelzer 1990; Kvarnemo 1995; Ostlund-
Nilsson 2001), number of eggs in a nest (Kraak and Groothius 1994; Forsgren et al. 1996; Goulet 1998)
and the incidence of filial cannibalism (Goto 1993), that are possible predictors of parental competence
and might provide an honest signal of male parental care. Some studies have even found that females
directly prefer males who are better fathers (Forsgren 1997; Wong 2004; Lindström et al. 2006). Courtship
rate has been shown to be both preferred and positively correlated with increased hatching success in
several studies of species such as the bicolour damselfish Stegastes partitus (Knapp and Kovach 1991;
Knapp and Warner 1991), fifteen spined sticklebacks Spinachia spinachia (Östlund and Ahnesjö 1998),
and pacific blue-eye fish Pseudomugil signifier (Wong 2004). In fifteen spined sticklebacks, increased
fanning beat rate leads to higher egg hatching success but also appears to be energetically expensive as
males who have a greater increase in fan beat rate lost more weight (Östlund and Ahnesjö 1998).
Females prefer males that court more (which is positively correlated to fan beat rate) and as courtship and
fan beat rate are condition-dependent, they appear to provide honest cues for female mating preference
(Zahavi 1975). Honest signalling of male parental care has been demonstrated in several studies (Kunzler
and Bakker 2000; Ostlund-Nilsson 2001; Senar et al. 2002; Takahashi and Kohda 2004; Halupka and
Borowiec 2006), but not in others (Linville et al. 1998; Smiseth et al. 2001; Mitchell et al. 2007) and there
is even evidence of dishonest signals in a few cases (Candolin 1999b).
2.1.5. Measuring female mating preference
The traditional method of determining female mating preference is through simultaneous choice
tests. Ideally these choice tests should exclude male-male interactions, otherwise male-male competition
may be influencing any increased mating success (or a surrogate thereof) for some males. This is
supported by several studies that report dominant males do not enjoy higher mating success when male-
50 .

male competition is removed (Radesater and Halldorsdottir 1993; Nelson and Nolen 1997; Nilsson and
Nilsson 2000; Reichard et al. 2005; Shackleton et al. 2005; Spence and Smith 2006). To control male-
male competition, many studies tether males or place them behind glass or mesh, which also has its
downfalls. Female preferences for specific male traits can differ when the sexes are allowed full or
restricted interactions (Candolin 1999a; Nilsson and Nilsson 2000). Also, in some species males behave
abnormally when tethered (Teleogryllus commodus, pers. obs.). A lack of close physical contact between
the sexes may also deny females access to essential mate choice cues (e.g., cuticular contact
pheromones in the field cricket Gryllus bimaculatus Tregenza and Wedell 1997)] and simultaneously
presenting females with two extremes in male phenotype can also artificially amplify a preference effect
(Wagner 1998). Female preference may be based on more than one male trait (reviewed in Candolin
2003) and a controlled trait might well be correlated with another uncontrolled trait.
Even in species that are better suited to simultaneous choice tests, female mate choice in
preference trials is often only indirectly measured. A relationship between, for example, visual orientation
(Clark and Biesiadecki 2002) or time spent near a male is only assumed to equate to actual mating
success. In a few cases this assumption is tested and supported (Clayton 1990; Witte 2006) but in others
it is not (Gabor 1999; Fuller 2003). There is also evidence of cryptic female choice, associated with rate of
oviposition (Eberhard 1996), differential allocation of eggs (Burley 1986) and post-copulatory sperm
selection (Lifjeld et al. 1994) that needs to be taken into account. The major dilemma therefore is that to
measure actual mating success individuals must interact, but it is difficult to do this without reintroducing
the problematic effects of male-male competition on male mating success.
Another way to measure male attractiveness that resolves these problems is to conduct no-choice
tests (Pilastro et al. 2002). These tests usually measure the time between the introduction of a single male
and female, and when a female acquiesces to mating (latency to mating). Measuring attractiveness as
mating latency has several benefits, females are allowed to base their choice on the entire physical and
chemical cues males offer, and female mating preference can be determined from an actual mating. No
choice tests measuring mating latency are most often used in fruitfly studies (Hegde and Krishna 1997;
Koref-Santibanez 2001; Gowaty et al. 2002; Yenisetti and Hegde 2003), but they have also been
employed in studies of birds (White et al. 2002), lizards (Tokarz 2002), fish (Houde and Torio 1992;
Blanchfield and Ridgway 1997), and invertebrates (Peters and Michiels 1996; Parri et al. 1998; Bukowski
et al. 2001; Jones and Quinnell 2002). My honours work on the Black Field Cricket (7. commodus) was
one of the few studies to test and confirm that males with a shorter latency to mating in no-choice trials
5 1 .

enjoyed increased longer-term mating success, supporting no-choice tests as an accurate measure of
male attractiveness (Shackleton et al. 2005).
The rate of egg production has been used as another method to assess female mating
preference. Variation in clutch size is one of the few cryptic choice modes employed by external fertilisers
(Eberhard 1996) and has been demonstrate in several taxa (Cote and Hunte 1989; Rintämaeki et al.
1998; Arnqvist and Danielsson 1999; Reyer et al. 1999; Fu et al. 2001; Parker 2003; Smith and Reichard
2005). Adjusting egg volume has also been implicated as a cryptic form of female choice (Kolm 2001), as
well as decreasing the time to spawning (Berejikian et al. 2000). Using egg production as a measure of
female choice, Spence and Smith (2006) have shown that female zebrafish show preference for males by
varying their clutch size and that they do not prefer dominant males.
2.1.6. Aims of this study
In this experiment, I aimed to quantify female mating preferences using the northern trout
gudgeon Mogurnda mogurnda. I tested whether females showed a mating bias and if this was towards
larger (better “condition”) males, or more dominant males. To quantify and record the mating behaviours
of M. mogurnda, I paired males that differed in size and conducted association preference tests with four
females per male pair. For each female, I measured preference variables such as the number of times she
entered a preference zone for each male, and the time spent in close proximity to each male. I then
analysed these variables to determine which characteristics females preferred so that I could then
compare them to any characteristics that might predict the quality of male parental care (Chapter 3).
52.

2.2 Methods
All aquaria were kept at 26° Celsius in climate controlled rooms on a 14:10 hr day: night
illumination cycle. The holding tanks (6’ x 2’ x 2’) where fish were kept between experiments were
connected to sump filters filled with bioballs containing bacteria that break down waste products in the
water. Four holding tanks were used, two for fish before experiments and two for after experiments. The
sexes were housed separately.
2.2.1. Pilot studies on male dominance
As pilot work for this study, I experimented with the best method of measuring male fighting ability
in M. mogurnda and found I was able to qualify relative male dominance but that I was not to quantify it
comprehensively. I wanted to observe and record males engaging in aggressive dominance behaviours to
test the hypothesis that larger males are more aggressive than smaller males. First, I paired males which
visually differed in mass by 5-10% and then I placed them into experimental tanks measuring 2’ x 1’ x 1’
which had two corner filters with carbon and filter wool in each back corner to maintain water quality
during the experiments. A 50% water change was conducted between tests of successive male pairs and
each method described below was conducted with four separate pairs of males.
I placed a pair of males into an experimental tanks, separated by a clear plastic divider that split
the tank into two equal halves with a male on each side. I acclimatised the males for 10 minutes then lifted
the separator and observed them for 10 minutes. I did this for four pairs of males. After 10 minutes of
observation, none of the four male pairs showed any aggressive behaviour towards each other. The
males either ignored each other or engaged in escape behaviour (systematically searching the tank for a
way out). I then repeated the procedure but changed the acclimatisation and observation time to 30
minutes, but still with no success. There were also no observable dominance behaviours when I
acclimatise the males for 30 minutes, one hour, or six hours. They still showed no sign of aggressive
behaviour after I lifted the divider and observed them.
I then tried to acclimatise the males without physically separating them so I acclimatised them
without a divider in the tank and checked on them after 30 minutes, one hour, three hours and six hours.
When I returned after six hours most of the males appeared to have engaged in aggressive or fighting
behaviours. I deduced that a fight had occurred when one male was out in the open and the other was
resting in the corner, often with visual damage to his fins. The fins of the “subordinate” male was often
53.

quite badly harmed, to the point where I had to separate three of the male pairs after three hours.
Observation of pairs showed that the “dominant” male would often chase the other male which quickly
swam to stay away from the dominant male.
The pairs mostly engaged in fighting somewhere between three to six hours, but the fights that
lead to the subordinate males showing the worst damage occurred within the first three hours. I repeated
this method with 12 pairs of males and 11 pairs had engaged in dominance behaviour by the sixth hour
and in all 11 pairs the larger male was the dominant male.
Considering the range and the length of times before the male engaged in dominance behaviours,
I decided that conducting comprehensive male dominance trials was not feasible. The results of the pilot
study which clearly showed that larger males are more dominant is the basis for my assumption that male
size is a good predictor of male fighting ability and dominance in this study on M. mogurnda.
2.2.2. Mate choice trials
I examined mate choice in M. mogurnda by weighing a series of males and then pairing together a
“large” and “small” male who differed in mass by at least 5 g. I randomly assigned four stock females to
each male pair for the duration of the mate choice experiments. Mate choice trials were conducted in
experimental tanks measuring 2’ x T x 1’. They had two corner filters with carbon and filter wool in each
back corner to maintain water quality during the experiments. A 50% water change was conducted
between tests with each female. Each tank had two mesh dividers to create two smaller compartments at
each end and a larger central compartment (Fig. 2.2.1).
On each mesh divider I attached two halves of a plastic tube on either side on the grid to create a
tube that could be used as a possible spawning site and allowed males and females to interact in a
sheltered location. A large and small male from a pair were then placed individually into opposite end
compartment. Males were allowed to acclimate for 10 minutes before the start of each mate choice trial.
The middle compartment was divided into three preference zones; a non-preference zone in the middle
(30 cm wide) and two preference zones at either end adjacent to the male compartments (15 cm wide).
After each mate choice trial, I swapped the males between the end compartments and allowed ten
minutes before the start of the next trial.
At the start of each trial, the female was placed in the centre of the tank under a clear plastic
container with slits in it to allow water flow. The female was then acclimatised for five minutes before the
container was raised. Her position in the tank (e.g., which zone she was in) was monitored for ten minutes
54 .

using the specialised behaviour analysis program JWatcher™ 0.9 from the Animal Behaviour Laboratory,
Macquarie University NSW (Blumstein et al. 2000). I recorded the duration of time spent in each
preference zone (i.e. time spent with each male) as well as the number of times the female left and then
re-entered each zone.
Each pair of males was tested with four females which resulted in four sets of “preference” data
for each male pair. I set an a priori criterion for which experiments were considered successful and which
pairs of males were eligible to proceed to the parental care experiments (see below). I conducted mate
choice experiments until I had obtained a total of 15 pairs of males (n = 60 mate choice trials). I used the
data from these initial mate choice trials to test whether: (1) females prefer larger males; (2) there was a
size-assortative female mating preference; and (3) the strength of preference for attractive males was
consistent for females tested with the same male pair. I then related these characters to male behaviour in
the subsequent parental care experiment.
2.2.3. Preference Criteria
To select pairs of males that differed sufficiently in their attractiveness for the parental care
experiments that followed, I only kept pairs of males that met a set of criteria. There were two
attractiveness categories: “attractive” males were preferred by at least three of the four females; and
“unattractive” males were preferred by none or one of the four female. To determine whether a female
considered one male more attractive than the other, I first excluded the time she spent in the non
preference zone. I then used the relative time the female spent in each male’s preference zone to
calculate which male she preferred. Relative time was the proportion of time the female spent in a male’s
preference zone relative to the total time spent in both preference zones. If a female spent 66% or more of
her time in the preference zone of one male, I recorded this male as being “preferred” by that female, and
the other as “non-preferred”. If a female spent a more equitable amount of time with each male, the trial
was rejected, another stock female was randomly selected and the trial was rerun. If two females failed to
show a preference for a given pair of males I put all the individuals into the “used” stock tanks and started
again the next day with a new pair of males. When all four females had successfully completed mate
choice trials, I compared their preferences. If three or more females preferred the same male, I deemed
that male as the “attractive” male and the other as the “unattractive” male in the pair. Females that agreed
on which male they preferred were then also returned to the “used” stock tanks.
55 .

2.2 .4 . Statistical Analyses - Mate Choice Analysis
In the analysis of female mating preferences, I considered the following variables:
• The number of visits was the number of times a female entered a male preference zone.
• The total duration was the total amount of time a female spent in a male preference zone.
• The mean duration was the average duration per visit a female spent in a male preference zone.
• The duration proportion was the proportion of time a female spent in a male preference zone
relative to the total duration of the observation period (10 minutes).
• The relative proportion was the proportion of time a female spent in a male preference zone
relative to the time a female spent in both preference zones (excludes the time in the non
preference zone).
I used Wilcoxon signed ranks tests to analyse whether the mean number of visits, total duration,
mean duration, duration proportion and relative proportion of time differed between large and small males.
For these tests I used the means per male for the four females. Using linear mixed models for each
response variable, I also treated female identity as a random factor nested within male pair identity to test
whether females preferred one male over the other. I modelled all five response variables using male size
and female mass as fixed factors. I also modelled number of visits, total duration and mean duration with
pair number as a fixed factor to see if females were spending more time with some male pairs.
I also conducted an ANOVA to test whether there was a difference in mean female mass among
male pairs. Finally, I tested if there was a significant difference in the mass of females to the males they
were paired with.
56.

Figure 2.2.1 The design of experimental tanks for female mate choice trials. A female was placed into the
centre section, and a large and small male were placed at opposite sides of the tank (the positions of the
males were alternated between trials). The presence of the female in each of the preference zones shown
was recorded using the specialised behaviour analysis program JWatcher™ 0.9 from the Animal
Behaviour Laboratory, Macquarie University NSW (Blumstein et al. 2000).
57.

2.3 Results
Females showed a mating preference for larger males (Table 2.3.1); they spent a greater amount
of total amount of time (Fig. 2.3.1), and a greater mean duration per visit (Fig. 2.3.2) with larger males.
When analysing the data using the mean values from the four females assigned to each male pair,
females spent a significantly greater total amount of time with larger males (Wilcoxon Signed Ranks, Z =
2.101, p = 0.036, n = 15) and tended to stay longer per visit (greater mean duration) with larger males
(Wilcoxon Signed Ranks, Z = 1.931, p = 0.053, n = 15). There was no significant difference in the number
of visits, duration proportion or relative proportion of time between different sized males (P = 0.1 to 0.32).
Using linear mixed models, male mass correlated positively with female preference behaviour
(Table 2.3.2). Females spent more time overall (Fig. 2.3.3), more time per visit (Fig. 2.3.4), and a greater
overall proportion of her time (Fig. 2.3.5) with the larger of the two males. Females also spent a greater
relative proportion of time with the larger of the two males. Using female mass and male mass as fixed
factors and female identity as a random factor, the total duration (F = 14.216, df = 1,117, p < 0.001), mean
duration (F = 6.141, df = 1,117, p = 0.015), duration proportion (F = 8.451, df = 1,117, p = 0.004) and
relative proportion (F = 11.879, df = 1,117, p = 0.001) of time was greater with larger males. There was,
however, still no significant difference between the number of times a female visited each male’s
preference zone.
The level of female preference did not differ significantly across the male pairs (Table 2.3.3).
Using linear mixed models with male pairs as the fixed factor and female identity nested within male pair
as a random factor, there was no difference in the number of visits (F = 0.599, df = 1,45, p = 0.851), the
total duration (F = 0.168, df = 105, p = 1.000) or the mean duration of visits to males (F = 0.892, df =
1,105, p = 0.569) across male pairs.
Mean female mass did not differ significantly across the male pairs (F = 1.655, df = 1,14, p =
0.101), however, females were assigned to pairs that differed significantly in the size of the test males
(Table 2.3.4), which raises some concerns about females choice being hindered by extremes in male size
forcing a sub-optimal choice. The difference between the mass of the female and the larger male (F =
2.579, df = 1,14, p = 0.008), and also between the mass of the female and the smaller male (F = 3.328, df
= 1, 14, p = 0.001) differed significantly among male pairs.
Interestingly, there was some evidence for a female preference being condition-dependent (Fig.
2.3.6). I calculated the spearman rank correlation between female size and the relative proportion of time
58.

spent with the preferred male for all 15 pairs. The mean correlation was significantly greater than zero
(mean rs = 0.307, n = 15, p = 0.024; one-sample t-test) showing that larger females exhibited a stronger
preference for the preferred male than smaller females. Using the same approach, there was also
evidence of size assortative mating (Fig. 2.3.7). Larger females had a tendency to spend relatively more
time with larger males compared to the time they spent with smaller males (mean rs = 0.253, n = 15, p =
0.072; one-sample t-test).
59.
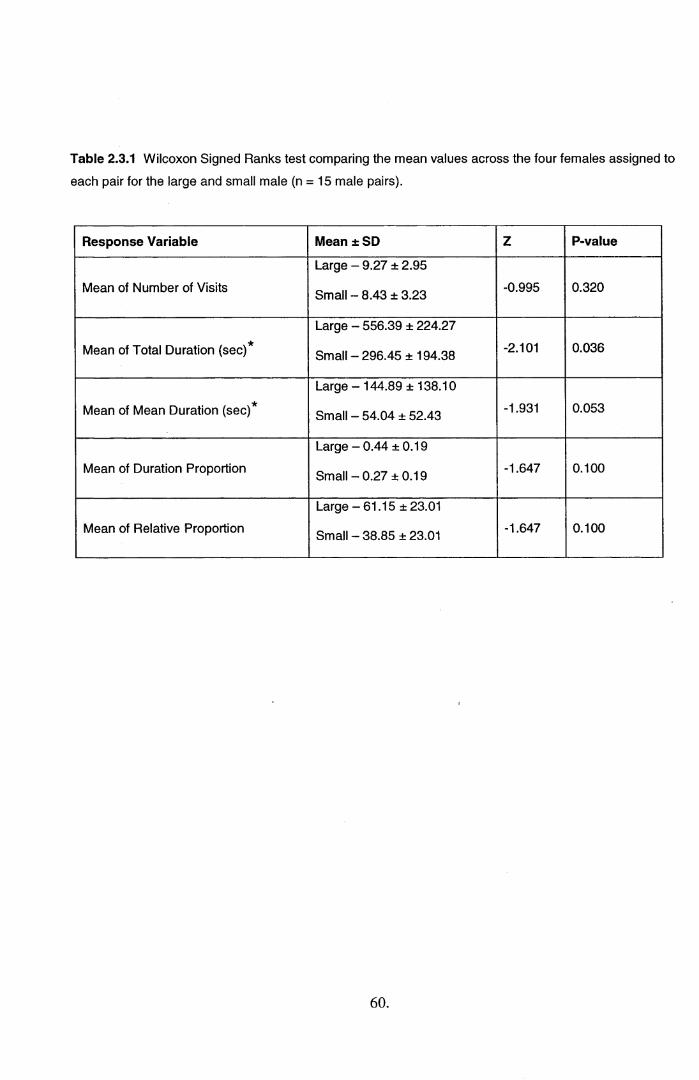
Table 2.3.1 Wilcoxon Signed Ranks test comparing the mean values across the four females assigned to
each pair for the large and small male (n = 15 male pairs).
Response Variable Mean ± SD Z P-value
Mean of Number of Visits
Large - 9.27 ± 2.95
Small-8 .43 ±3.23 -0.995 0.320
Mean of Total Duration (sec)*
Large - 556.39 ± 224.27
Small-296.45 ± 194.38 -2.101 0.036
Mean of Mean Duration (sec)*
Large - 144.89 ± 138.10
Small - 54.04 ± 52.43 -1.931 0.053
Mean of Duration Proportion
Large - 0.44 ±0.19
Small-0 .27 ±0.19 -1.647 0.100
Mean of Relative Proportion
Large-61.15 ±23.01
Small-38.85 ±23.01 -1.647 0.100
60.

Table 2.3.2 Linear mixed models with female mass and male mass as fixed factors and female identity as
a random factor). Variables marked * are significant at the 0.05 level.
Response Variable Dependent Variable df F Sig.
Number of Visits Male Size 1,58 0.773 0.206
Female Mass 1,58 1.634 0.386
Total Duration Male Size* 1,117 14.216 0.000*
Female Mass 1,117 2.215 0.148
Mean Duration Male Size* 1,117 6.141 0.015*
Female Mass 1,117 0.089 0.766
Duration Proportion Male Size* 1,117 8.451 0.004*
Female Mass 1,117 2.032 0.157
Relative Proportion Male Size* 1,117 11.879 0.001*
Female Mass 1,117 0.000 1.000
61.

Table 2.3.3 Linear mixed models with male pair as a fixed factor and female identity nested within male
pair identity as a random factor.
Response Variable df F Sig.
Number of Visits 14,45 0.599 0.851
Total Duration 14,105 0.168 1.000
Mean Duration 14,105 0.892 0.569
62.

Table 2.3.4 One way ANOVA on female mass, the difference between female mass and the mass of the
larger male (Larger Male Difference), and the difference between female mass and the mass of the
smaller male (Smaller Male Difference) across pairs.
Response Variable df F Sig.
Female Mass 14, 59 1.655 0.101
Larger Male Difference 14, 59 2.579 0.008
Smaller Male Difference 14, 59 3.328 0.001
63.

3.0 ~
2.5 -
2.0 ”
(J
4>3cr4)S_
fa
1.5 -
1.0 -
0.5 -
0 0 -----
-1000 -800 -600 -400 -200 0 200 400 600
Difference in Mean Total Duration
I800 11000
Figure 2.3.1 The means of the difference in total duration (average across four females per male)
between the larger and smaller male in a pair (Wilcoxon Signed Ranks, Z = -2.101, p = 0.036; n = 15 male
pairs). Positive values indicate that females spent more time with the larger male.
64 .

0.5 -
Difference in Mean Duration
Figure 2.3.2 The means of the difference in mean duration (average across four females per male)
between the larger and smaller male in a pair (Wilcoxon Signed Ranks, Z = -1.931, p = 0.053; n = 15 male
pairs). Positive values indicate that females spent more time with the larger male.
65.
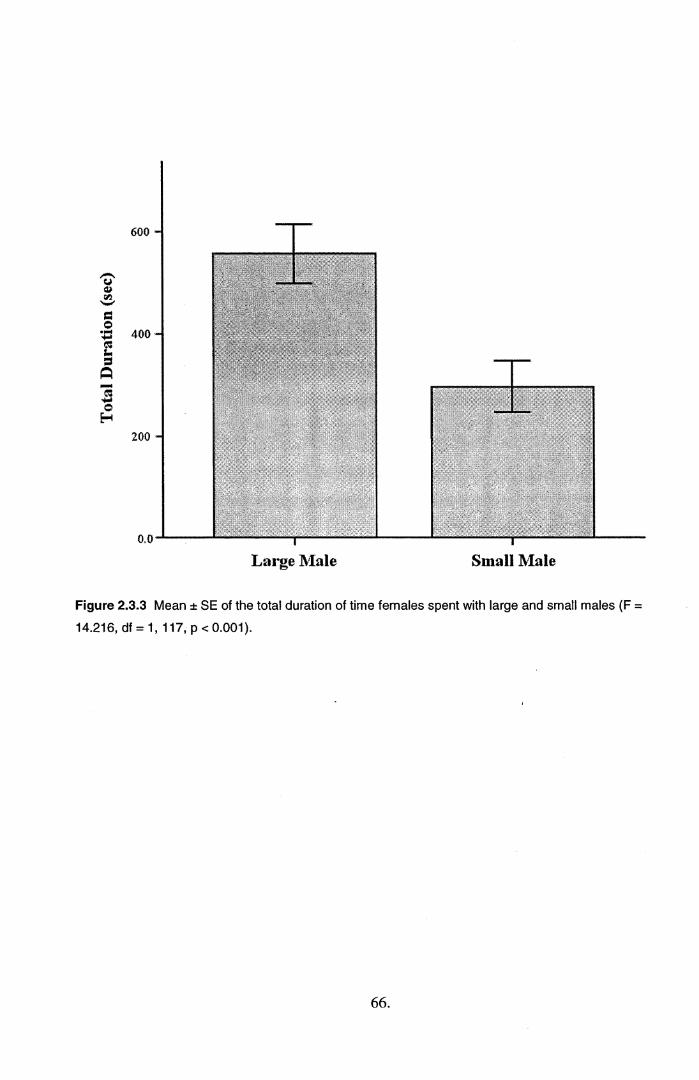
600 -
Large Male Small Male
Figure 2.3.3 Mean ± SE of the total duration of time females spent with large and small males (F =
14.216, df = 1, 117, p< 0.001).
6 6 .
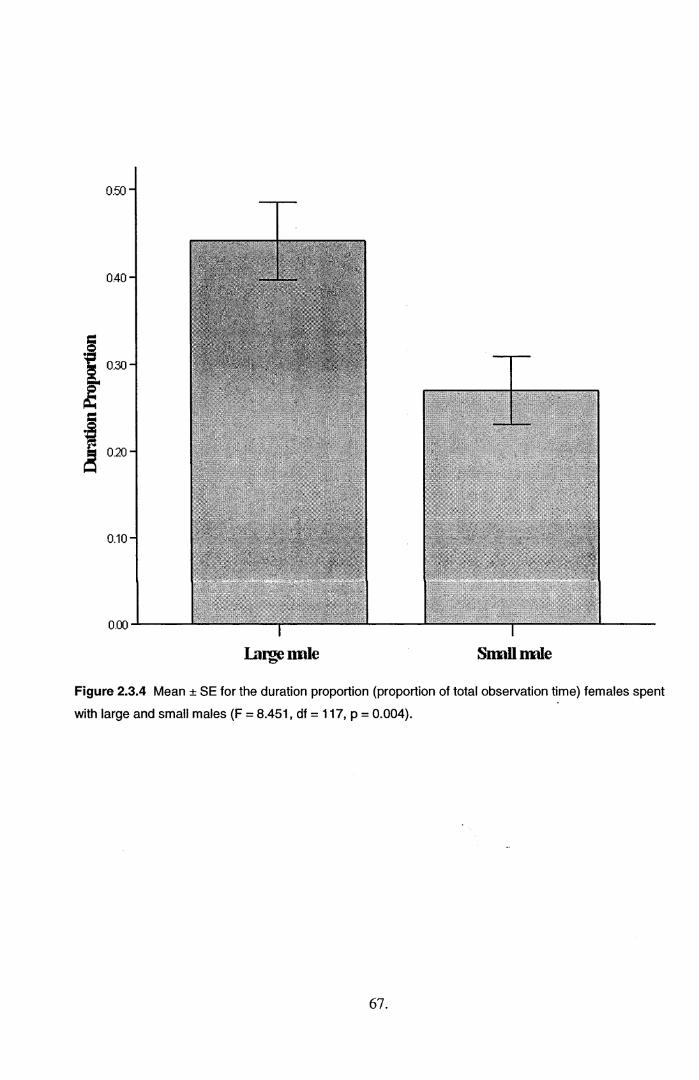
0.50-
Large mile Small male
Figure 2.3.4 Mean ± SE for the duration proportion (proportion of total observation time) females spent
with large and small males (F = 8.451, df = 117, p = 0.004).
67.

Figure 2.3.5 Mean ± SE for the proportion of time females spent with large and small males relative to
the amount of time spent with the other male (F = 11.879, df = 1,117, p = 0.001).
68.

- 1.0 - 0.8 - 0.6 - 0.4 - 0.2 0.0 0.2 0.4 0.6 0.8 1.0
Spearmini’s correlation
Figure 2.3.6 Histogram of Spearman’s rank correlations between female size and time spent with the
preferred male in a pair (n = 4 per correlation) showing that larger females spent relatively more time with
preferred males (one-sample t-test, t = 2.52, p = 0.024, n = 15 male pairs).
69.

5 -
4 “
u 33ss< up N
2 -
1 -H 1* '
t f « - -
4 .
> . ' ,
;
•.
I I I■1.0 - 0.8 - 0.6 - 0.4 - 0.2 0.0 0.2 0.4
Sp eann an*s con- elati on
0.6 0.8 1.0
Figure 2.3.7 Histogram of Spearman’s rank correlations between female size and time spent with the
larger male in a pair (n = 4 per correlation) showing that larger females tended to spend more time with
larger males (one-sample t-test, t = 1.95, p = 0.072, n = 15 male pairs).
70.

2.4 Discussion
I found that female Mogurnda mogurnda discriminated between potential mates along a size
gradient. Females spent a greater amount of total time duration and a longer mean duration with larger
males when measured both as a mean across the four females assigned to each pair, and treating each
females as an individual data point. This result was consistent across all pairs, as was female mass. Of
some concern is the fact that some pairs of males differed significantly in size, raising concerns about
female choice being altered by extremes in male size compelling an artificial choice.
Interestingly, I found that larger females exhibited a stronger preference for the preferred male
than smaller females, implying that female choice in this species is condition-dependent. There was also
evidence of size assortative mating with larger females tending to spend more time with larger males
compared to the time they spent with smaller males.
2.4.1. Females preferred larger males
Females display mating preferences for larger males across many taxa (reviewed in Choe and
Crespi 1997) and this study further supports that trend; female M. mogurnda showed a marked preference
for larger males. There are demonstrated benefits to mating with larger males: larger males are more
likely to win fights (e.g., Kotiaho et al. 1997; Hagelin 2002; Lopez et al. 2002; Reichard et al. 2005) and so
provide access to superior resources. For example in fish, larger males are better at obtaining and
guarding spawning sites (Bisazza and Marconato 1988; Lindström 1988). In addition, there is a potential
genetic benefit if traits that increase success during male-male competition are heritable (Cordero and
Eberhard 2003).
Dominant males are mostly likely to be of higher physiological quality. Some performance
measures have shown to positively predict success in male-male competition, and males with higher
energetic reserves are more likely to be successful in male-male interactions in several animal taxa
(Chellappa and Huntingford 1989; Neat et al. 1998). Female choice for larger, more dominant males might
actually be for males in better physiological condition, thereby also selecting for condition-dependence of
male traits (Bonduriansky and Rowe 2005).
Condition-dependence is most commonly discussed in terms of the increased investment by
males into sexually selected traits to attract mates (see Kokko 1998a and the references therein). The
“handicap principle” suggests that only males of high quality and in a good condition can bear the cost of a
71.

sexually selected trait to attract females and so signal their quality through these secondary sexually
characteristics (Zahavi 1975). Signals of condition, however, don’t necessarily need to be through sexually
selected traits, females might directly observe cues instead of relying on correlated traits. Though M.
mogurnda display no secondary sexual characteristics, increased condition could be expressed as higher
courtship rates (Knapp and Warner 1991; Karino 1995) or higher quality parental care. Parental care is
costly (Drent and Daan 1980a; Stanley 2002; Hanssen et al. 2003) and body condition will affect the
quality of parental care an individual can provide (e.g., Linville et al. 1998, Wilson 1998; Kolm 2004;
Candolin 2000; Voltura et al. 2002; Senar et al. 2002; Halupka and Borowiec 2006). Studies have shown
that females sometimes prefer males of high parental quality, rather than basing their choice on male
dominance or size (e.g., sand gobies; Forsgren 1997) although what cues females base this choice on
isn’t always clear (see 2.4.3.”Comparing measures of mating preference”).
2.4.2. Size assortative mating
In this study I not only found that female M. mogurnda generally preferred larger males, I also
found a weak size assortative mating preference. Size-assortative mating appears to be relatively
common across taxa (e.g., Brown 1990; 1993; Arnqvist et al. 1996; Hegde and Krishna 1997; Harari et al.
1999), and has been found in several species of fish e.g., cichlids (Barlow et al. 1990; Budaev et al.
1999), and coral goby Paragobioon echinocephalus (Kuwamura et al. 1993). Again, when mating is costly
the influence of size appears to be particularly important (Hedrick and Dill 1993; Rowe 1994; Grafe 1997;
Watson et al. 1998). For example, larger males can provide females with more food, are better able to
defend territories or resources, and provide protection from conspecifics and predators (Wiegmann and
Baylis 1995; Ptacek and Travis 1997; Savalli and Fox 1998; Watson 1998). The importance of minimising
these costs may lead to assortative mating (Arnqvist et al. 1996; Rowe and Arnqvist 1996). Although there
are numerous examples of size-assortative mating (inferring a role for female as well as male size in
mating) the question of the specific mechanisms by which this arises is still unclear (e.g., mutual mate
choice, mechanical constraints, arrival at breeding sites).
2.4.3. Condition-dependent female choice
I found that as well as size assortative mating behaviour, larger female M. mogurnda showed a
stronger preference for preferred males, indicating that female mating preferences may be condition-
72.

dependent in this species. This is an interesting result because the evidence of condition-dependent
female preference has received a lot of attention but doesn’t have a great deal of empirical support.
The theories around condition-dependent female choice has gained momentum in the literature in
recent years, and was reviewed recently by Cotton et al. (2006b). There is growing evidence of condition-
dependent female mating preferences in fish, e.g., guppies Poecilia reticulata (Lopez 1999; Syriatowicz
and Brooks 2004), three spined sticklebacks Gasterosteus aculeatus (Bakker et al. 1999), upland bullies
Gobiomorphus breviceps (Poulin 1994) and deep-snouted pipefish Syngnathus typhie (Mazzi 2004) as
well as in insects [e.g., black field crickets Teleogryllus commodus (Hunt et al. 2005)], amphibians [e.g.,
African reed frogs Hyperolius marmoratus, (Jennions et al. 1995)], and birds [e.g., black grouse Tetrao
tetrix (Rintämaeki et al. 1998)].
As discussed, expressing mating preferences exacerbates the costs of mating and logically
females in better condition should be better able to bear these costs than females in worse condition
(Cotton et al. 2006). Alternately, higher quality females may be more discriminating for the same cost than
poorer quality females. As preferences are predicted to be weaker when the costs are higher, the strength
of female preference is expected to vary with female condition by much the same argument that sexually
secondary traits vary with male condition (Pomiankowski 1987; Grafen 1990; Iwasa et al. 1991; Iwasa and
Pomiankowski 1999). For example, Mazzi (2004) showed that experimental infection of both sexes of
deep-snouted pipefish Syngnathus typhie revealed changes in mating preference. Deep-snouted pipefish
are sex-role reversed and males are the choosier sex, so males in good condition showed a strong
preference for uninfected females whereas infected males in worse condition did not discriminate between
infected and not infected females. As such, the first prediction of a system that displays condition-
dependent female choice is that better quality females will showed stronger preferences. This study
satisfied this prediction as larger female M. mogurnda showing stronger preference for the preferred male
in each pair.
Another prediction of condition-dependent choice is size assortative mating (Cotton et al. 2006).
Better quality females should be able to exhibit stronger preferences for the same costs and therefore
have better mates. This may be through more intense searching, or having a higher rejection rate than
lower quality females. This study also satisfied this prediction, with larger female M. mogurnda preferring
larger (higher quality) males than smaller females. Again, using deep-snouted pipefish as an example
species, there is evidence of mutual mate choice with both sexes showing preference for larger partners
(Mazzi 2004). Male condition-dependent preference was particularly sensitive to female infection status
73.

amongst small females who have lower than average fecundity, suggesting that one of the main factors
underlying male preference is the selection of mates in order to increase the number and quality of eggs
received. The influence of size on parental care behaviours in M. mogurnda will be discussed in Chapter
3.
The final prediction is that females of higher genetic quality should also show stronger mating
preferences for better quality males than females with lower genetic quality. This is because females with
high genetic quality will do better in any given environment than females of lower genetic quality and
therefore the mechanisms of condition-dependent choice should be the same across different
environments, i.e. mating decisions should be maintained. The scope of my study did not cover the
genetic quality of my fish across environments but this would be a clear avenue for a subsequent study.
2.4.4. Comparing measures of mating preference
Having argued that females demonstrated mating preferences for larger males across many taxa
(reviewed in Choe and Crespi 1997) and appear to do so in this study, the assumption that females prefer
larger, presumably more dominant, males has been challenged on numerous occasions (Qvarnstrom and
Forsgren 1998). Dominant males may impose direct (e.g., Leboeuf and Mesnick 1991; Preston et al.
2001; Wong 2004) and indirect (e.g., Chippindale et al. 2001) costs on females, making them less
attractive as mates.
As discussed, traditional mate choice tests by means of simultaneous choice tests have their
problems. Even in studies like this one, where the effect of male-male competition has been controlled for,
and physical and chemical contact between the female and males has been maximised, female mate
choice is still being indirectly measured and this indirect measure is only assumed to equate to actual
mating success. Another way to measure male attractiveness is to conduct no-choice tests (e.g., Pilastro
et al. 2002; Shackleton et al. 2005) that measure latency to mating. Measuring attractiveness as mating
latency has several benefits: females are allowed to base their choice on the entire suite of physical and
chemical cues males offer; and female mating preference can be determined from an actual mating. In
Chapter 3, I discuss how I used the days until spawning as alternate measure of mating preference and
compare it to the results from this chapter.
74.

References
Alatalo RV, Carlson A, Lundberg A (1988) The search cost in mate choice of the Pied Flycatcher. Animal
Behaviour 36:289-291
Andersson M (1994) Sexual Selection. Princeton University Press, Princeton, New Jersey
Arnqvist G, Danielsson I (1999) Postmating sexual selection: the effects of male body size and recovery
period on paternity and egg production rate in a water strider. Behavioral Ecology 10:358-365
Arnqvist G, Rowe L, Krupa JJ, Sih A (1996) Assortative mating by size: a meta-analysis of mating patterns
in water striders. Evolutionary Ecology 10:265-284
BakkerTCM, Kunzler R, Mazzi D (1999) Condition-related mate choice in sticklebacks. Nature 401:234
Barlow G, Francis RC, Baumgartner FV (1990) Do the colours of parents, companions and self influence
assortative mating in the polychromatic Midas cichlid? Animal Behaviour 40:713-722
Berejikian BA, Tezak EP, Larae AL (2000) Female mate choice and spawning behaviour of Chinook
salmon under experimental conditions. Journal of Fish Biology 57:647-661
Bisazza A, Marconato A (1988) Female mate choice, male-male competition and parental care in the river
bullhead, Cottus gobio L. (Pisces, Cottidae). Animal Behaviour 36:1352-1360
Blanchfield PJ, Ridgway MS (1997) Reproductive timing and use of redd sites by lake-spawning brook
trout (Salvelinus fontinalis). Canadian Journal of Fisheries and Aquatic Sciences 54:747-756
Blanckenhorn WU, Kraushaar U, Reim C (2003) Sexual selection on morphological and physiological
traits and fluctuating asymmetry in the yellow dung fly. Journal of Evolutionary Biology 16:903-913
Blumstein DT, Evans CS, Daniel JC (2000) JWatcher. In, 0.9 edn. Animal Behaviour Laboratory, Maquarie
University, Sydney, pp A powerful tool for the quantitative analysis of behaviour.
Bonduriansky R, Rowe L (2005) Sexual selection, genetic architecture, and the condition dependence of
body shape in the sexually dimorphic fly Prochyliza xanthostoma (Piophilidae). Evolution 59:138-
151
Brown WD (1990) Size-assortative mating in the blister beetle Lytta magister (Coleoptera: Meloidae) is
due to male and female preference for larger mates. Animal Behaviour 40:901-909
75.

Brown WD (1993) The cause of size-assortative mating in the leaf beetle Trirhabda canadensis
(Coleoptera: Chrysomelidae). Behavioral Ecology and Sociobiology 33:151-157
Bryant DM, Newton AV (1994) Metabolic costs of dominance in dippers, Cinclus cinclus. Animal
Behaviour 48:447-455
Budaev SV, Zworykin DD, Mochek AD (1999) Individual differences in parental care and behaviour profile
in the convict cichlid: a correlation study. Animal Behaviour 58:195-202
Bukowski TC, Linn CD, Christenson TE (2001) Copulation and sperm release in Gasteracantha
cancriformis (Araneae : Araneidae): differential male behaviour based on female mating history.
Animal Behaviour 62:887-895
Bulmer M (1989) Structural instability of models of sexual selection. Theoretical Population Biology
35:195-206
Burley N (1986) Sexual selection for aesthetic traits in species with biparental care. American Naturalist
127:415-445
Candolin U (1999a) Male-male competition facilitates female choice in sticklebacks. Proceedings of the
Royal Society of London Series B-Biological Sciences 266:785-789
Candolin U (1999b) The relationship between signal quality and physical condition: is sexual signalling
honest in the three spined stickleback? Animal Behaviour 58:1261-1267
Candolin U (2003) The use of multiple cues in mate choice. Biological Review 78:575-595
Chellappa S, Huntingford SA (1989) Depletion of energetic reserves during reproductive aggression in
male three spined stickleback, Gasterosteus aculeatus. Journal of Fish Biology 35:275-286
Chippindale AK, Gibson JR, Rice WR (2001) Negative genetic correlation for adult fitness between sexes
reveals ontogenetic conflict in Drosophila. Proceedings of the National Academy of Sciences of
the United States of America 98:1671-1675
Choe JC, Crespi BJ (1997) Mating systems in insects and arachnids. Cambridge University Press,
Cambridge
Clark DL, Biesiadecki B (2002) Mating success and alternative reproductive strategies of the dimorphic
jumping spider, Maevia inclemens (Aaneae, Salticidae). Journal of Arachnology 30:511-518
76.

Clayton NS (1990) Assortative mating in zebra finch subspecies, Taeniopygia-Guttata-Guttata and T-G-
Castanotis. Philosophical Transactions of the Royal Society of London Series B-Biological
Sciences 330:351-370
Cordero C, Eberhard WG (2003) Female choice of sexually antagonistic male adaptations: a critical
review of some current research. Journal of Evolutionary Biology 16:1-6
Cöte IM, Hunte W (1989) MALE AND FEMALE MATE CHOICE IN THE REDLIP BLENNY - WHY
BIGGER IS BETTER. Animal Behaviour 38:78-88
Cotton S, Fowler K, Pomiankowski A (2004) Condition dependence of sexual ornament size and variation
in the stalk-eyed fly Cyrtodiopsis dalmanni (Diptera : Diopsidae). Evolution 58:1038-1046
Cotton S, Small J, Pomiankowski A (2006) Sexual Selection and Review Condition-Dependent Mate
Preferences. Current Biology 16:755-765
Daly M (1978) Cost of mating. American Naturalist 112:771-774
Downhower JF, Brown L (1980) Mate preferences of female mottled sculpins, Cottus bairdi. Animal
Behaviour 28:728-734
Drent RH, Daan S (1980) The prudent parent: Energetic adjustments in avian breeding. Ardea 68:225-252
Eberhard WG (1996) Female control: sexual selection by cryptic female choice. Princeton University
Press, Princeton, New Jersey
Evans JP, Kelly JL, Bisazza A, Finazzo E, Pilastro A (2004) Sire attractiveness influences offspring
performance in guppies. Proceedings of the Royal Society of London Series B-Biological
Sciences 271:2035-2042
Forsgren E (1997) Female sand gobies prefer good fathers over dominant males. Proceedings of the
Royal Society of London Series B-Biological Sciences 264:1283-1286
Forsgren E, Karlsson A, Kvarnemo C (1996) Female sand gobies gain direct benefits by choosing males
with eggs in their nests. Behavioral Ecology and Sociobiology 39:91-96
Friberg U, Arnqvist G (2003) Fitness effects of female mate choice: preferred males are detrimental for
Drosophila melanogasterfemales. Journal of Evolutionary Biology 16:797-811
77.

Fu P, Neff BD, R. GM (2001) Tactic-specific success in sperm competition. Proceedings of the Royal
Society of London Series B-Biological Sciences 268:1105-1112
Fuller RC (2003) Disentangling female mate choice and male competition in the rainbow darter,
Etheostoma caeruleum. Copeia 1:138-148
Gabor C (1999) Association patterns of sailfin mollies (Poecilia latipinna): alternative hypotheses.
Behavioral Ecology and Sociobiology 46:333-340
Gerhardt HC (1994) THE EVOLUTION OF VOCALIZATION IN FROGS AND TOADS. Annual Review of
Ecology and Systematics 25:293-324
Gibson RM, Bachman GC (1992) The costs of female choice in a lekking bird. Behavioral Ecology 3:300-
309
Goto A (1987) Polygyny in the river sculpin, Cottus hangiongensis (Pisces: Cottidae), with special
reference to male mating success. Copeia 1:32-40
Goto A (1993) MALE MATING SUCCESS AND FEMALE MATE CHOICE IN THE RIVER SCULPIN,
COTTUS-NOZAWAE (COTTIDAE). Environmental Biology of Fishes 37:347-353
Goulet D (1998) Spawning success in the damselfish Amblyglyphidodon leucogaster: the influence of
eggs in the nest. Animal Behaviour 55:651-664
Gowaty PA, Steinichen R, Anderson WW (2002) Mutual interest between the sexes and reproductive
success in Drosophila pseudoobscura. Evolution 56:2537-2540
Grafe TU (1997) Costs and benefits of mate choice in the lek-breeding reed frog, Hyperolius marmoratus.
Animal Behaviour S3:1103-1117
Grafen A (1990) BIOLOGICAL SIGNALS AS HANDICAPS. Journal of Theoretical Biology 144:517-546
Guderley H, Couture P (2005) Stickleback fights: why do winners win? Influence of metabolic and
morphometric parameters. Physiological and Biochemical Zoology 78:173-181
Hagelin JC (2002) The kinds of traits involved in male-male competition: a comparison of plumage,
behavior, and body size in quail. Behavioral Ecology 13:32-41
Halupka K, Borowiec M (2006) Male whitethroats, Sylvia communis, advertise their future contribution to
parental care. Behaviour 143:1-14
78.

Hanssen SA, Erikstad KE, Johnsen V, Bustnes JO (2003) Differential investment and costs during avian
incubation determined by individual quality: an experimental study of the common eider
(Somateria mollissima). Proceedings of the Royal Society of London Series B-Biological Sciences
270:531-537
Harari AR, Handler AM, Landolt PJ (1999) Size-assortative mating, male choice and female choice in the
curculionid beetle Diaprepes abbreviatus. Animal Behaviour 58:1191-1200
Hedrick AV, Dill LM (1993) Mate choice by female crickets is influenced by predation risk. Animal
Behaviour 46:193-196
Hegde SN, Krishna MS (1997) Size-assortative mating in Drosophila malerkotliana. Animal Behaviour
54:419-426
Heywood JS (1989) Sexual selection by the handicap mechanism. Evolution 43:1387-1397
Hoelzer GA (1989) The good parent process of sexual selection. Animal Behaviour 38:1067-1078
Hoelzer GA (1990) Male-male competition and female choice in the cortez damselfish, Stegastes
rectifraenum. Animal Behaviour 40:339-349
Houde AE, Torio AJ (1992) Effect of Parasitic Infection on Male Color Pattern and Female Choice in
Guppies. Behavioral Ecology 3:346-351
Howard DJ, Reece M, Gregory PG, Chu J, Cain ML (1998) The evolution of barriers to fertilization
between closely related organisms. In: Howard DJ, Berlocher SH (eds) Endless forms: species
and speciation. Oxford University Press, New York, pp 279-288
Hunt J, Brooks R, Jennions MD (2005) Female mate choice as a condition-dependent life-history trait.
American Naturalist 166:78-92
Hunt J, Brooks R, Jennions MD, Smith MJ, Bentsen CL, Bussiere LF (2004) High-quality male field
crickets invest heavily in sexual display but die young. Nature 432:1024-1027
Irschick DJ, Garland TJ (2001) Integrating function and ecology in studies of adaptation: investigations of
locomotor capacity as a model system. Annual Review of Ecology and Systematics 32:367-396
Iwasa Y, Pomiankowski A (1999) Good parent and good genes models of handicap evolution. Journal of
Theoretical Biology 200:97-109
79.

Iwasa Y, Pomiankowski A, Nee S (1991) The evolution of costly mate preferences. II. The ‘handicap’
principle. Evolution 45:1431-1442
Jennions MD, Backwell PRY, Passmore Nl (1995) Repeatability of mate choice: the effect of size in the
African painted reed frog, Hyperolius marmoratus. Animal Behaviour 49:181-186
Jivoff P (1997) The relative roles of predation and sperm competition on the duration of the post-
copulatory association between the sexes in the blue crab, Callinectes sapidus. Behavioral
Ecology and Sociobiology 40:175-185
Jones TM, Quinnell RJ (2002) Testing predictions for the evolution of lekking in the sandfly, Lutzomyia
longipalpis. Animal Behaviour 63:605-612
Karino K (1995) Male-male competition and female mate choice through courtship display in the territorial
damselfish Stegastes nigricans. Ethology 100:126-138
Keenleyside MHA, Rangeley RW, Küppers BU (1985) Female mate choice and male parental defense
behaviour in the cichlid fish Cichlasoma nigrofasciatum. Canadian Journal of Zoology 63:2489-
2493
Keller L, Reeve HK (1995) Why do females mate with multiple males - the sexually selected sperm
hypothesis. Advances in the Study of Behavior 24:291-315
Kemp DJ, Alcock J (2003) Lifetime resource utilization, flight physiology, and the evolution of contest
competition in territorial insects. American Naturalist 162:290-301
Kirkpatrick M (1996) Good genes and direct selection in evolution of mating preferences. Evolution
50:2125-2140
Knapp RA, Kovach JT (1991) Courtship as an honest indicator of male parental quality in the bicolor
damselfish, Stegastes partitus. Behavioral Ecology 2:295-300
Knapp RA, Warner RR (1991) Male parental care and female choice in the bicolor damselfish, Stegastes
partitus: bigger is not always better. Animal Behaviour 41:747-756
Kodric-Brown A, Nicoletto P (2005) Courtship behaviour, swimming performance, and microhabitat use of
Trinidadian guppies. Environmental Biology of Fishes 73:299-307
Kokko H (1998) Good genes, old age and life-history trade-offs. Evolutionary Ecology 12:739-750
80.

Kokko H, Monaghan P (2001) Predicting the direction of sexual selection. Ecology Letters 4:159-165
Kolm N (2001) Females produce larger eggs for large males in a paternal mouthbrooding fish.
Proceedings of the Royal Society of London Series B-Biological Sciences 268:2229-2234
Koref-Santibanez S (2001) Effects of age and experience on mating activity in the sibling species
Drosophila pavani and Drosophila gaucha. Behavior Genetics 31:287-297
Kotiaho J, Alatalo RV, Mappes J, Parri S (1997) Fighting success in relation to body mass and drumming
activity in the male wolf spider Hygrolycosa rubrofasciata. Canadian Journal of Zoology-Revue
Canadienne De Zoologie 75:1532-1535
Kotiaho JS (2001) Costs of sexual traits: a mismatch between theoretical considerations and empirical
evidence. Biological Reviews 76:365-376
Kraak SBM, Groothius TGG (1994) Female preference for nests with eggs is based on the presence of
the eggs themselves. Behaviour 131:189-206
Kunzler R, Bakker TCM (2000) Pectoral fins and paternal quality in sticklebacks. Proceedings of the Royal
Society of London Series B-Biological Sciences 267:999-1004
Kuwamura T, Yugo Y, Nakakshima Y (1993) Size-assortative monogamy and paternal egg care in a coral
goby Paragobioon echinocephalus. Ethology 95:65-75
Kvarnemo C (1995) SIZE-ASSORTATIVE NEST CHOICE IN THE ABSENCE OF COMPETITION IN
MALES OF THE SAND GOBY, POMATOSCHISTUS-MINUTUS. Environmental Biology of Fishes
43:233-239
Lailvaux SP, Irschick DJ (2006) No evidence for female association with high-performance males in the
green anole lizard, Anolis carolinensis. Ethology 112:707-715
Leboeuf BJ, Mesnick S (1991) Sexual behavior of male Northern Elephant Seals 1. Lethal injuries to adult
females. Behaviour 116:143-162
Lifjeld JT, Dunn PO, Westneat DF (1994) Sexual selection by sperm competition in birds: male-male
competition or female choice? Journal of Avian Biology 25:244-50
Lindström K (1988) MALE-MALE COMPETITION FOR NEST SITES IN THE SAND GOBY,
POMATOSCHISTUS-MINUTUS. Oikos 53:67-73
81.

Lindström K, St Mary C, Pampoulie C (2006) Sexual selection for male parental care in the sand goby,
Pomatoschistus minutus. Behavioral Ecology and Sociobiology 60:46-51
Linville SU, Breitwisch R, Schilling AJ (1998) Plumage brightness as an indicator of parental care in
northern cardinals. Animal Behaviour 55:119-127
Lopez P, Munoz A, Martin J (2002) Symmetry, male dominance and female mate preferences in the
Iberian rock lizard, Lacerta monticola. Behavioral Ecology and Sociobiology 52:342-347
Lopez S (1999) Parasitized female guppies do not prefer showy males. Animal Behaviour 57:1129-1134
Magnhagen C (1991) Predation risk as a cost of reproduction. Trends in Ecology & Evolution 6:183-185
Marden JH, Waage JK (1990) Escalated damselfly territorial contests are energetic wars of attrition.
Animal Behaviour 39:954-959
Mazzi D (2004) Parasites make male pipefish careless. Journal of Evolutionary Biology 17:519-527
McLister JD (2003) The metabolic cost of amplexus in the gray tree frog (Hyla versicolor): assessing the
energetics of male mating success. Canadian Journal of Zoology 81:388-394
Metcalfe NB, Taylor AC, Thorpe JE (1995) Metabolic rate, social status and life-history strategies in
Atlantic salmon. Animal Behaviour 49:431-436
Mitchell DP, Dunn PO, Whittingham LA, Freeman-Gallant CR (2007) Attractive males provide less
parental care in two populations of the common yellowthroat. Animal Behaviour 73:165-170
Moore AJ, Gowaty PA, Wallin WG, Moore PJ (2001) Sexual conflict and the evolution of female mate
choice and male social dominance. Proceedings of the Royal Society of London Series B-
Biological Sciences 268:517-523
Moore AJ, Moore PJ (1999) Balancing sexual selection through opposing mate choice and male
competition. Proceedings of the Royal Society of London Series B-Biological Sciences 266:711-
716
Neat FC, Taylor AC, Huntingford FA (1998) Proximate costs of fighting in male cichlid fish: the role of
injuries and energy metabolism. Animal Behaviour 55:875-882
Nelson CM, Nolen TG (1997) Courtship song, male agonistic encounters, and female mate choice in the
house cricket, Acheta domesticus (Orthoptera: Gryllidae). Journal of Insect Behavior 10:557-570
82.

Nicoletto PF (1991) The relationship between male ornamentation and swimming performance in the
guppy, Poecilia reticulate. Behavioral Ecology and Sociobiology 28:365-370
Nicoletto PF (1993) Female response to condition-dependent ornaments in the guppy, Poecilia reticulata.
Animal Behaviour 46:441-450
Nicoletto PF (1995) Offspring quality and female choice in the guppy, Poecillia reticulata. Animal
Behaviour 49:377-387
Nilsson SO, Nilsson GE (2000) Free choice by female sticklebacks: lack of preference for male
dominance traits. Canadian Journal of Zoology-Revue Canadienne De Zoologie 78:1251-1258
Noonan K (1983) Female mate choice in a cichlid fish Cichlasoma nigrofasciatum. Animal Behaviour
31:1005-1010
Ostlund-Nilsson S (2001) Fifteen-spined stickleback (Spinachia spinachia) females prefer males with more
secretional threads in their nests: an honest-condition display by males. Behavioral Ecology and
Sociobiology 50:263-269
Östlund S, Ahnesjö I (1998) Female fifteen-spined sticklebacks prefer better fathers. Animal Behaviour
56:1177-1183
Parker GA (1974) Assessment strategy and the evolution of fighting behaviour. Journal of Theoretical
Biology 47:223-243
Parker TH (2003) Genetic benefits of mate choice separated from differential maternal investment in red
junglefowl (Gallus gallus). Evolution 57:2157-2165
Parker TH, Garant D (2004) Quantitative genetics of sexually dimorphic traits and capture of genetic
variance by a sexually-selected condition-dependent ornament in red junglefowl (Gallus gallus).
Journal of Evolutionary Biology 17:1277-1285
Parker TH, Ligon JD (2003) Female mating preferences in red junglefowl: a meta-analysis. Ethology
Ecology & Evolution 15:63-72
Parri S, Alatalo RV, Mappes J (1998) Do female leaf beetles Galerucella nymphaeae choose their mates
and does it matter? Oecologia 114:127-132
Perry G, Levering K, Girard I, Garland TJ (2004) Locomotor performance and dominance in male Anolis
cristatellus. Animal Behaviour 67:37-47
83.

Peters A, Michiels NK (1996) Evidence for lack of inbreeding avoidance by selective simultaneous
hermaphrodite. Invertebrate Biology 115:99-103
Petrie M (1983) Female moorhens compete for small fat males. Science 220:413-415
Pilastro A, Evans JP, Sartorelli S, Bisazza A (2002) Male phenotype predicts insemination success in
guppies. Proceedings of the Royal Society of London Series B-Biological Sciences 269:1325-
1330
Plaistow S, Siva-Jothy MT (1996) Energetics, constraints, and male mate-securing tactics in the damselfly
Calopteryx splendens xanthostoma (Charpentier). Proceedings of the Royal Society of London
Series B-Biological Sciences 263:1233-1238
Pomfret J, Knell R (2006) Sexual selection and horn allometry in the dung beetle Euoniticellus
intermedius. Animal Behaviour 71:567-576
Pomiankowski A (1987) The costs of choice in sexual selection. Journal of Theoretical Biology 128:195-
218
Poulin R (1994) Mate choice decisions by parasitized female upland bullies, Gobiomorphus breviceps.
Proceedings of the Royal Society of London Series B-Biological Sciences 256:183-187
Preston BT, Stevenson IR, Pemberton JM, Wilson K (2001) Dominant rams lose out by sperm depletion -
A waning success in siring counters a ram's high score in competition for ewes. Nature 409:681-
682
Price T, Schlüter D, Heckman NE (1993) Sexual selection when the female directly benefits. Biological
Journal of the Linnean Society 48:187-211
Ptacek MB, Travis J (1997) Mate choice in the sailfin molly, Poecilia latipinna. Evolution 51:1217-1231
Qvarnstrom A, Forsgren E (1998) Should females prefer dominant males? Trends in Ecology & Evolution
13:498-501
Radesater T, Halldorsdottir H (1993) 2 Male types of the common earwig - male-male competition and
mating success. Ethology 95:89-96
Reichard M, Bryja J, Ondrackova M, Davidova M, Kaniewska P, Smith C (2005) Sexual selection for male
dominance reduces opportunities for female mate choice in the European bitterling (Rhodeus
sericeus). Molecular Ecology 14:1533-1542
84.

Reyer H-U, Frei G, Som C (1999) Cryptic female choice: frogs reduce clutch size when amplexed by
undesired males. Proceedings of the Royal Society of London Series B-Biological Sciences
266:2101-2107
Rintämaeki PT, Lundberg A, Alatalo RV, Höeglund J (1998) Assortative mating and female clutch
investment in black grouse. Animal Behaviour 56:1399-1403
Robson MA, Miles DB (2000) Locomotor performance and dominance in tree lizards, Urosaurus ornatus.
Functional Ecology 14:338-344
Rogers W, Barlow GW (1991) Sex differences in mate choice in a monogamous biparental fish, the midas
cichlid (Cichlasoma citrinellum). Ethology 87:249-261
Roskaft E, Järvi T, Bakken M, Bech C, Reinertson RE (1986) The relationship between social status and
resting metabolic rate in great tits (Parus major), and pied flycatchers (Ficedula hypoleuca).
Animal Behaviour 34:838-842
Rowe L (1994) The costs of mating and mate choice in water striders. Animal Behaviour 48:1049-1056
Rowe L, Arnqvist G (1996) Analysis of the causal components of assortative mating in water striders.
Behavioural Ecology and Sociobiology 38:279-286
Sakaluk SK, Belwood JJ (1984) Gecko phonotaxis to cricket calling song: a case of satellite predation.
Animal Behaviour 32:659-662
Savalli UM, Fox CW (1998) Sexual selection and the fitness consequences of male body size in the seed
beetle Stator limbatus. Animal Behaviour 55:473-483
Senar JC, Figuerola J, Pascual J (2002) Brighter yellow blue tits make better parents. Proceedings of the
Royal Society of London Series B-Biological Sciences 269:257-261
Shackleton MA, Jennions MD, Hunt J (2005) Fighting success and attractiveness as predictors of male
mating success in the black field cricket, Teleogryllus commodus: the effectiveness of no-choice
tests. Behavioral Ecology and Sociobiology 58:1-8
Sloman KA, Motherwell G, O’Connor Kl, Taylor AC (2000) The effect of social stress on the standard
metabolic rate (SMR) of brown trout, Salmo trutta. Fish Physiology and Biochemistry 23:49-53
Smiseth PT, Ornborg J, Andersson S, Amundsen T (2001) Is male plumage reflectance correlated with
paternal care in bluethroats? Behavioral Ecology 12:164-170
85.

Smith C, Reichard M (2005) Females solicit sneakers to improve fertilisation success in the bitterling
(Rhodeus sericeus). Proceedings of the Royal Society of London Series B-Biological Sciences
272:1683-1688
Spence R, Smith C (2006) Mating preference of female zebrafish, Danio rerio, in relation to male
dominance. Behavioral Ecology 17:779-783
Stanley TR (2002) How many kilojoules does a Black-billed Magpie nest cost? Journal of Field
Ornithology 73:292-297
Syriatowicz A, Brooks R (2004) Sexual responsiveness is condition-dependent in female guppies, but
preference functions are not. BMC Ecology 4
Takahashi D, Kohda M (2004) Courtship in fast water currents by a male stream goby (Rhinogobius
brunneus) communicates the parental quality honestly. Behavioral Ecology and Sociobiology
55:431-438
Tokarz RR (2002) An experimental test of the importance of the dewlap in male mating success in the
lizard Anolis sagrei. Herpetologica 58:87-94
Tomkins JL, Radwan J, Kotiaho JS, Tregenza T (2004) Genic capture and resolving the lek paradox.
Trends in Ecology & Evolution 19:323-328
Tregenza T, Wedelt N (1997) Definitive evidence for cuticular pheromones in a cricket. Animal Behaviour
54:979-984
Wagner WEJ (1998) Measuring female mating preferences. Animal Behaviour 55:1029-1042
Watson PJ (1998) Multi-male mating and female choice increase offspring growth in the spider Neriene
litigiosa (Linyphiidae). Animal Behaviour 55:387-403
Watson PJ, Arnqvist G, Stallmann RR (1998) Sexual conflict and the energetic costs of mating and mate
choice in water striders. American Naturalist 151:46-58
White DJ, King AP, West MJ (2002) Facultative development of courtship and communication in juvenile
male cowbirds (Molothrus ater). Behavioral Ecology 13:487-496
Whiting MJ, Nagy KA, Bateman PW (2003) Evolution and maintenance of status-signalling badges:
experimental manipulations in lizards. In: Fox SF, McCoy JK, Baird TA (eds) Lizard Social
Behavior. Johns Hopkins University Press, Baltimore, pp 47-82
86.

Wiegmann DD, Baylis JR (1995) Male body size and paternal behaviour in smallmouth bass, Micropterus
dolomieui (Pisces: Centrarchidae). Animal Behaviour 50:1543-1555
Witte K (2006) Time spent with a male is a good indicator of mate preference in female zebra finches.
Ethology Ecology & Evolution 18:195-204
Wong BBM (2004) Superior fighters make mediocre fathers in the Pacific blue-eye fish. Animal Behaviour
67:583-590
Wong BBM, Candolin U (2005) How is female choice affected by male competition? Biological Review
80:559-571
Yenisetti SC, Hegde SN (2003) Size-related mating and reproductive success in a drosophilid: Phorticella
striata. Zoological Studies 42:203-210
Zahavi A (1975) Mate selection: a selection for a handicap. Journal of Theoretical Biology 53:205-214
87.

CHAPTER 3 Spawning and parental care in the
northern trout gudgeon (
mogurnda)
3.1 Introduction
Parental investment theories predict that selection favours individuals that maximise their lifetime
reproductive success. There are both costs and benefits to each reproductive event and therefore a trade
off between present and future reproduction (Williams 1966b; Trivers 1972; Gross and Sargent 1985;
Sargent and Gross 1985; Clutton-Brock 1991). The costs of parental care include expenditure of energy
and time, and loss of foraging and mating opportunities (Clutton-Brock 1991; Balshine-Earn 1995; Smith
and Wootton 1995; Mackereth et al. 1999), and the investment in parental care is beneficial by increasing
the current reproductive success of parents (Sargent and Gebier 1980; Zoran and Ward 1983; Kunzler
and Bakker 2000; Karino and Arai 2006). In species with male parental care, it has been suggested that
females pay attention to the willingness of their future mate to invest in offspring and the quality of the
care they will provide (Möller and Thornhill 1998).
3.1.1. How much to invest in parental care: balancing costs and benefits
Parents have to make decisions on how much to invest in each clutch and how to adjust their
expenditure in response to variation in costs and benefits (Winkler 1987). Finding the optimal trade-off of
costs and benefits to maximise net fitness is complex. For example, an increased risk of predation
increases the benefits of caring for young but also increases the potential costs to the parent thereby
resulting in a reduction in future reproductive success. Lindström (1998) found that male sand gobies
Pomatoschistus minutus only continued to guard eggs in the presence of a predator (which increased the
cost of care) when the clutch was large (when the benefit was higher). This suggests that males only
continue caring for eggs if the benefit of investing in the current clutch outweighs the cost to future
reproductive success.
If the costs become too great, parents can decide to abandon the reproductive event completely.
Filial cannibalism and desertion is commonly reported in fish with male parental care (Rohwer 1978;
Petersen 1990; FitzGerald 1992b; Marconato et al. 1993; Okuda and Yanagisawa 1996; Lindström and
8 8 .

Sargent 1997; Lindström 1998; Jennions and Polakow 2001; Manica 2002b; Payne et al. 2002). Females
have, however, developed a strategy to help reduce this loss. Males usually only fully cannibalise clutches
that are small (Sargent 1992) and females have been shown to increase the clutch size of males by
preferentially spawning with those who are already guarding eggs (Marconato and Bisazza 1986; Unger
and Sargent 1988; Petersen 1990; Kraak and Groothuis 1994; Forsgren et al. 1996; Goulet 1998;
Reynolds and Jones 1999). The cost of even partial filial cannibalism will be diluted if a male is guarding
several clutches of eggs (Forsgren et al. 1996).
Other factors that affect the amount of parental investment in care include the reproductive value
of the parents, the age of the offspring and the physical condition of the parents (Trivers 1985; Sargent
1997; Takegaki and Nakazono 1999; Lissaker et al. 2003; Manica 2003; Pampoulie et al. 2004).
Investment in care is also affected by factors such as environmental gradients (Schaffer 1974a; Kramer
1987; St Mary et al. 2001; Hale et al. 2003) and clutch size (Karino and Arai 2006). Parental care
behaviours are discussed in detail in Section 3.1.7 (“Variation in parental care behaviours”).
3.1.2. Paternal care in fish
Although in many taxa female-only care is the predominant form of care (Clutton-Brock 1991), the
majority of fish display male parental care (Gross and Sargent 1985). The majority of external fertilisers
(76%) display male parental care, which usually involves defence of a territory by more than one female
(Gross and Sargent 1985). Mank et al. (2005) showed that paternal care was by far the most common
form of care in ovuliparous fish (41 families) compared to five incidences each for maternal and biparental
care. Individuals control their own level of care and, aside from mammalian offspring that require nutrition
from the mother in the form of lactation, either parent can provide the necessary care in most breeding
systems. Sole parental care by the male seems counter-intuitive, however, in the light of the costs
discussed in Chapter 1. So how is it so feasible?
Mank et al. (2005) showed that paternal care appears to have evolved straight from the
generalised basal condition of external fertilisation at least 22 times. They also showed that Actinopterygii
display phenotypic expressions of sexual selection, most commonly sexual dichromatism (Reimchen
1989; Houde and Endler 1990; Stott and Poulin 1996; Amundsen and Forsgren 2001). Sexual
dichromatism was positively correlated with paternal care, which suggests that the competition for mates
that drives sexual selection also drives the evolution of parental care. Furthermore, nest construction was
also positively correlated to paternal care, and generally preceded it. Parental care may have grown from
89.

the territorial defence of spawning sites by males: nest construction may be a pre-adaptation for male
parental care.
Several reasons have been put forward to explain why paternal care is so common in external
fertilisers. Firstly, males have higher confidence in paternity with external fertilisation (Trivers 1972)
therefore males are more likely to invest care in eggs as the mean benefits are higher (Neff and Gross
2001). Secondly, based on the assumption that females release gametes first, this favours female
desertion leaving the male to either care for the eggs or desert (Trivers 1972; Dawkins and Carlisle 1976).
Finally, even if the benefits of care are equal for both sexes, the costs to males are lower as they can
guard clutches from several females simultaneously with a reduced loss in mating opportunities (Blumer
1979). Males with eggs already in their nests are actually preferred by females in some species further
reinforcing the evolution of paternal care (Unger and Sargent 1988; Kraak and Groothuis 1994; Forsgren
et al. 1996; Goulet 1998; Reynolds and Jones 1999).
When one parent (male or female) can successfully be the sole provider of care, this presents the
opportunity for their mates to manipulate the relative contributions of parental investment. The benefits of
manipulating a partner’s expenditure on care results from sexual conflict between the sexes.
3.1.3. Sexual conflict over parental care
Conflict exists because the costs of parental care are usually disproportionately carried between
individuals but the benefits are shared equally (Trivers 1972; Parker et al. 2002). Therefore, a parent who
manipulates the other parent into increasing his or her investment into the current reproductive event will
enjoy a selective advantage.
Early studies argued that males are more likely to benefit from investing less care as they have
greater potential reproductive rates (Clutton-Brock 1991). Males gain more from investing effort into
gaining additional matings than providing care, and this is the basis of the traditional argument as to why
parental care is predominantly provided by females in most taxa (Bateman 1948; Trivers 1972). As
discussed in Chapter 1, however, every offspring has one mother and one father and the average
reproductive rates are equal between the sexes. Queller (1997) remedies this flaw in the argument by
noting that if a subset of “attractive” males are gaining the majority of matings then the reproductive rate of
the average male is irrelevant because the ‘average’ male does not mate. The reproductive rate of mating
males is higher than that of females and the decision to expend energy seeking new mates rather than
providing care is beneficial and provides an evolutionary stable strategy for males.
90.

This asymmetry provides a feasible model for why females tend to provide more care than males,
particularly as sexual selection (and male-male competition) intensifies (Clutton-Brock and Parker 1992).
When females provide more care, this creates a positive feedback loop if fewer and fewer males can
monopolize an even greater share of the available females due to greater female choosiness or male-
male competition having a stronger effect on determining which males gain access to mates.
3.1.4. Adjusting parental investment: responding to partner effort
Experimental studies suggest that the amount of care each parent provides is likely to be a
responsive dynamic process of negotiation rather than a predetermined decision (Dawkins and Carlisle
1976; Webb et al. 1999; McNamara et al. 2000). Recent studies actually suggest that parents may benefit
from handicapping themselves and thus manipulating their mate to compensate for their decreased efforts
(Marques 2004; Osorno and Szekely 2004; Houston et al. 2005). There is evidence to show that when
one mate decreases their effort in care, the partner will increase their efforts in compensation. For
example, in the starling Sturnus vulgaris when the parental effort of one parent is experimentally
decreased, the other mate responded by increasing their level of care (Wright and Cuthill 1989).
Studies that looked at the amount of compensation a parent provides have, however, produced
varied results. By clipping feathers or attaching small weights, bird researchers can increase the costs of
care and manipulate the parental effort of a mate (measured by feeding rate). These studies aim to
measure the corresponding change in effort of the other partner, which ranged from no difference
(Whittingham et al. 1994; Sanz et al. 2000), to both an increase (Slagsvold and Lifjeld 1988) and a
decrease (Markman et al. 1995) in effort. Whittingham et al. (1994) handicapped both male and female
great tits Parus major, and found no overall effect on the combined feeding rate of pairs but the responses
from birds were complex. The sexes differed in their response, with handicapped males decreasing their
feeding rates while handicapped females did not. In response, females partnered with a handicapped
male fully compensated for their decrease in work rate, but males mated with a handicapped female did
not show any compensation and even tended to decrease their feeding rates (Whittingham et al. 1994).
Theory predicts that compensation should only be partial, not full, otherwise a parent could
decrease it’s own effort without reducing the total effort to the offspring which is clearly an unstable
evolutionary strategy (Houston et al. 2005). Contrary to this, however, Osorno and Szekely (2004) showed
near full compensatory parental effort after a male has deserted the nest in magnificent frigatebird Fregata
91.

magnificens. In this case, rather than responding to the relative effort of a mate, females might instead
respond to the condition of their mates (Schwagmeyer et al. 2002).
3.1.5. Adjusting parental investment: responding to partner condition
Logically, an individual should choose a mate in good condition to provide better care for their
offspring. The good parent hypothesis predicts that secondary sexual characteristics should signal an
individual’s ability to provide care (Heywood 1989; Hoelzer 1989), but studies provide evidence that both
support (Linville et al. 1998; Buchanan and Catchpole 2000; Soler et al. 2001) and contradict (Smiseth
and Amundsen 2000; Smiseth et al. 2001) this prediction. As discussed in Chapter 1, sexually selected
traits are expensive to produce (Kotiaho 2001) and often demonstrate a trade-off with other life history
traits (Hunt et al. 2004). So it may be that males with more attractive traits are in fact not in better
condition (Grafen 1990; Kokko 2001; Hunt et al. 2004; Tomkins et al. 2004), or do not honestly signal their
condition (Candolin 1999b).
Kokko (1998b) provides a model of parental care and attractiveness in which males allocate
energy expenditure to self-maintenance, sexual advertising and parental care to maximise their fitness. If
there are diminishing returns from advertising and care (i.e. reduced benefits to extra matings or reduced
opportunities for extra matings), advertising will be an honest indicator of male care and females will prefer
these males. This model, however, doesn’t predict how much a female will allocate to a reproductive event
due to the attractiveness of the male or the effect of indirect genetic benefits and adding these factors into
the model could change the conclusions (Houston et al. 2005).
As previously mentioned, attractive males may decrease their parental care effort because they
gain more from investing in seeking new mates due to their higher reproductive rate. Females mated to
theses attractive males, however, may increase their effort by allocating a greater investment into those
reproductive events (Cunningham and Russell 2000). Known as the ‘differential allocation hypothesis’
(Burley 1988; Sheldon 2000), females may increase expenditure on offspring because the offspring are
more valuable (Sheldon 2000) or as an attempt to retain their attractive mate (Burley 1988).
The question of what cues females use for mate choice and the honesty of these cues relating to
condition and parental care can be less obvious in species where males do not display secondary sexual
ornaments. In fish, females have been shown to use parental care as an outright cue for mating
preferences, preferring males that exhibit parental care to a greater degree and have a higher associated
hatching success (Downhower and Brown 1980; Unger and Sargent 1988; Knapp and Kovach 1991;
92.

Forsgren et al. 1996; Forsgren 1997). Studies have reported females preferring quality of parental care
over more traditionally traits such as dominance (e.g. pacific blue-eye fish, Wong 2004). In situations
where the trait used for mate attraction is (or is correlated to) male care quality, it is expected to be under
sexual selection (Kirkpatrick 1985). This creates an interesting dynamic in which the traditional trade-off
between current and future reproductive success is lessened because there is less trade-off between
parental care (current reproduction) and expressing traits under sexual selection (future reproduction).
Care might even be used as a courtship display and so increased in the presence of receptive females
(Fitzpatrick et al. 1995; Kokko 1998b).
3.1.6. Parental care and mate choice
Parental care: correlated traits
In fish, there is a wide literature showing females prefer larger males in species that show
biparental or paternal care (Downhower and Brown 1980; Noonan 1983; Keenleyside et al. 1985; Goto
1987; Bisazza and Marconato 1988; Cote and Hunte 1989; Rogers and Barlow 1991). Larger and more
dominant males are more likely to be of higher physiological quality (e.g., Chellappa and Huntingford
1989; Guderley and Couture 2005) and female mate choice for larger males might mask a preference for
better physiological condition, thereby also selecting for condition-dependence of male traits
(Bonduriansky and Rowe 2005).
The condition of a male or his quality in terms of parental care is often related to his size. For
example, guarding is one of the most common forms of parental care in fish and there appears to be a
large male advantage in nest defence. Larger males are better at acquiring and guarding spawning sites
(Bisazza and Marconato 1988; Lindström 1988) and their ability to obtain better quality nesting sites could
also contribute to an increased survival rate of eggs (Hastings 1988; Cote and Hunte 1989; Kvarnemo et
al. 1998). Guarding is a foraging limiting activity (Kraak and Videler 1991; Kraak 1996; Payne et al. 2002),
and males have been shown to lose weight by the end of the caring period (Coleman and Fischer 1991;
Maekawa et al. 1996; Jones and Reynolds 1999; Skolbekken and Utne-Palm 2001) which selects for
males that are initially in better condition. Where dominance does not equate to increased parental care,
however, females often choose overall paternal quality over male fighting ability (Forsgren 1997; Wong
2004; Wong and Candolin 2005).
93.

Other factors effecting female choice - filial cannibalism
Females also choose mates based on other male traits such as courtship (Knapp and Warner
1991; Östlund and Ahnesjö 1998), nest characteristics (Sargent and Gebier 1980), and number of eggs in
a nest. The incidence of females preferentially choosing to lay their eggs with males who already have
eggs in their nest is widespread (Marconato and Bisazza 1986; Unger and Sargent 1988; Cote and Hunte
1989; Petersen 1990; Kraak and Groothuis 1994; Forsgren et al. 1996; Goulet 1998; Reynolds and Jones
1999; Jennions and Polakow 2001; Okuda et al. 2002). There may be several reasons for this: (1) to
reduce the risk of males eating the whole clutch or abandoning the nest by increasing total clutch size
(Rohwer 1978); (2) to reduce the proportion of her eggs that are eaten during care (e.g., Forsgren et al.
1996); (3) to increase male parental investment by increasing clutch size (e.g., Coleman et al. 1985;
Karino and Arai 2006); and (4) to increase the chance that a male is a high quality parent (mate choice
copying) (Kraak and Videler 1991).
If the costs of care are too great (or the benefits too small), males often eat their own eggs (filial
cannibalism) or desert the brood to regain condition to invest in future reproduction (FitzGerald 1992b;
Marconato et al. 1993; Kraak 1996; Okuda and Yanagisawa 1996; Kvarnemo et al. 1998; Marques 2003).
This behaviour is adaptive for males, who benefit in terms of resource consumption but represents a loss
of investment for the female. Expenditure on small clutches presents a low return on investment which
may be better allocated to future broods, so whole clutch filial cannibalism is usually found when males
have small or low quality clutches (Petersen 1990; Sargent 1992; Smith 1992; Svensson et al. 1998;
Manica 2002a; Lissaker et al. 2003; Manica 2003; Karino and Arai 2006). Experimental reductions of
clutches have shown that parents have a threshold below which a clutch is totally cannibalised (Lavery
and Keenleyside 1990) or a male will abandon his eggs (Jennions and Polakow 2001). Females who
preferentially lay eggs in nests where there are already eggs present will increase the clutch size of a
male’s brood and therefore decrease the likelihood of losing her investment to whole clutch filial
cannibalism or desertion.
Males do not always eat the entire clutch; sometimes they eat only a portion of their eggs. Partial
clutch cannibalism is theorised to be a method of regaining energy lost from care to invest into current or
future offspring (Unger and Sargent 1988; Forsgren et al. 1996; Reynolds and Jones 1999). The
behaviour is therefore adaptive as it can reduce the loss of condition and increase a male’s overall
reproductive success by increasing the hatching success of the current reproductive event and
maintaining condition for future clutches (Sargent 1985; Smith and Wootton 1995). As with whole clutch/or
94.

cannibalism, females who choose to lay eggs in nests where there are already eggs present will dilute the
effect of partial cannibalism on her eggs by increasing the total clutch size.
Evidence that males devote greater amounts of parental investment into larger clutches is
widespread in fish (Coleman et al. 1985; Sargent 1988; Forsgren et al. 1996; Karino 1997; Jennions and
Polakow 2001; Suk and Choe 2002; Manica 2003; Karino and Arai 2006) though some studies have found
no relationship between clutch size and effort (Knapp and Warner 1991; Skolbekken and Utne-Palm
2001). Females who preferentially lay their eggs to maximise their mates clutch size should be at a
selective advantage and receive direct benefits for this behaviour. Finally, it has been suggested that a
female preference for males with large broods might be adaptive because the number of eggs present in
the nest is probably a reliable predictor of the survival chances of the eggs (Kraak and Videler 1991).
Overall parental investment is also affected by the environmental conditions, such as temperature
(Skolbekken and Utne-Palm 2001; Cooke et al. 2003) and oxygen level (Lissaker et al. 2003).
3.1.7. Variation in parental care behaviours
Fanning
Variation in specific parental care behaviours are also affected by the costs and benefits of caring.
Fanning behaviour is a very common form of care in fish (Blumer 1979) and entails oxygenation of the
embryos by fanning the eggs with the fish’s pectoral fins. Fanning is often the most important factor in
hatching success (Sargent 1988; Kraak 1996; Lindström 1998; Karino and Arai 2006) and egg
development rate (e.g., Sargent and Gebier 1980; Zoran and Ward 1983; Karino and Arai 2006). Fanning
increases water circulation and gas exchange (Wootton 1998; Green and McCormick 2005), which is
important as reduced oxygen availability retards embryo development and increases mortality (Kunzler
and Bakker 2000). When oxygen levels are low, an increase in ventilation of nests by fanning more water
over the eggs has been noted in many species of fish, e.g., sticklebacks Gasterosteus aculeatus (van
lersel 1953; Reebs et al. 1984) and gobies (Torricelli et al. 1985; Jones and Reynolds 1999; Skolbekken
and Utne-Palm 2001). Fanning is also effected by changes in temperature and salinity (St Mary et al.
2001) and clutch size (Karino and Arai 2006).
Filial cannibalism
As discussed, filial cannibalism has been widely recorded in fish (recorded in 17 families, Manica
2002) , and across other taxa (Polis 1981). Partial clutch cannibalism as a means for males to recuperate
95 .

lost condition and energy assumes that caring males are food limited, and many findings support this
claim (Sargent 1985; Itzkowitz and Makie 1986; Demartini 1987; Hoelzer 1992; Sargent et al. 1995; Smith
and Wootton 1995). Payne et al. (2002), found that males fed a supplementary diet gained more weight
than control males but interestingly, that did not equate into an increase in hatching success. Loss of
condition does appear to affect the filial cannibalism though, Kvarnemo (1998) for example found that
males whose diets where supplemented decreased their rate of filial cannibalism but did not erase it all
together. On the other hand, other studies have found no clear relationship between nutritional status and
filial cannibalism (Belles-lsles and Fitzgerald 1991; Kvarnemo 1997; Lindström and Sargent 1997;
Vinyoles et al. 1999).
Payne et al. (2002) propose an alternate theory for filial cannibalism: increasing available oxygen
to the eggs. Reducing egg density increased development time and hatching success, probably as a
function of increasing oxygen availability to the embryos. Males in lowered dissolved oxygen (DO) levels
consumed relatively more eggs consistent with this explanation. An alternative possibility, however, is that
this could simply be a function of fact the energetic costs of care are increased under low DO (Kamler
1992). A oxygen mediated model showed that males who cannibalise a high fixed proportion of their eggs
had higher hatching success than males who cannibalised a low proportion or none at all (Payne et al.
2004).
Another proposed reason for partial filial cannibalism is control of disease or infection. Males may
not be eating healthy eggs, they may instead be cannibalising dead, unfertilised of diseased eggs
(Hoelzer 1988; Kraak 1996). Though Kraak (1996) showed males selectively ate dead eggs, the rate of
filial cannibalism is mostly well above the natural rate of diseased eggs so this can only offer a partial
explanation for egg predation (Kraak 1996; Klug et al. 2005). It has also been proposed that eggs provide
a source of particular nutrients important to the male (Belles-lsles and Fitzgerald 1991; Sargent 1992) but
experimental evidence shows males fed with other male’s embryos (eggs from another male’s nest) do
not have a lower incidence of cannibalism on their own clutch (Kvarnemo et al. 1998).
96.

3 .1.8 . Aims of this study
In this experiment, I aimed to quantify and record the parental care behaviours of Mogurnda
mogurnda, which as far as I am aware, has not been documented before. For each male, I measured
variables such as spawning time, egg fanning, egg guarding, filial cannibalism, desertion and fanning rate.
Relating these variables back to my results from Chapter 2, I wanted to determine if:
(a) there are honest (i.e. predictable) cues to male parental care quality;
(b) these are the same characters used by females to make their mating preferences; and
(c) male preference shown in Chapter 2 from association preference tests is congruent to the
results in this chapter where I used latency to mating as an alternative measure of female preference for a
male.
97.

3.2 Methods
As described for the mate choice trials (Chapter 2) all aquaria were kept at 26° Celsius in climate-
controlled rooms, and on a 14:10 hr day: night illumination cycle. Holding tanks (6’ x 2’ x 2’) where fish
were kept between experiments were connected to sump filters filled with bioballs containing bacteria that
break down waste products in the water. Four holding tanks were used, two for fish before experiments
and two for after experiments while keeping the sexes separate.
3.2.1. Parental Care
Having selected pairs of males that differ significantly in attractiveness (Chapter 2), I randomly
assigned two similarly sized breeding females to each male pair. Each male was then put into a separate
breeding tank with one female. All breeding tanks (36” x 18” x 18”) had adequate filtration and I kept water
quality high with weekly 25% water changes. Each tank had three spawning sites comprising two white
rectangular tiles at opposite ends and a piece of plastic piping.
I checked every tank for spawning every 1-2 days and I recorded each time that a pair spawned
(or if they had failed to do so after 60 days). I also recorded male parental care behaviour using
JWatcher™ (Blumstein et al. 2000) until the eggs hatched or were eaten. I recorded male behaviour for 10
minutes per recording session, with 1-2 days between sessions. The behaviours I recorded included the
frequency and duration of fanning bouts, the intervals between fanning bouts, fin beat rate (average
number of fin beats per second), time spent guarding (resting on top of the eggs), time guarding (near the
eggs), time away from the eggs and any observations of filial cannibalism. I then analysed the parental
care behaviour data to test whether there was any effect of male size or male “attractiveness” on the
various measures of parental quality.
3.2.2. Statistical Analyses - Parental Care Analysis
To analyse whether mate choice preference inferred from latency to mating (no-choice tests) was
congruent with the preferences seen in the traditional association mate choice tests, I used survival
analyses (Kaplan Meier) to compare the number of days until the first spawning occurred between large
and small males, as well as between preferred and non-preferred males (i.e. survival is equivalent to how
long it was until a male’s status changed from no spawning to spawning). The data was right censored if
the male had still not spawned at the end of the study period (60 days).
98 .

In the parental care behaviour analyses, I analysed sources of variation in the following parental
care behaviours that were recorded during the mating experiments:
• Attack was when a male left the eggs to chase away another fish.
• Away was when a male left the eggs for another reason.
• Fanning was when a male used his pectoral fins to fan the eggs to facilitate the flow of fresh
oxygenated water over them.
• Flare was when a male flared his operculum at another fish in warning.
• Near was when a male was within 10cm of the eggs but not directly over them.
• Rest was when a male was lying directly over the eggs but was not fanning them.
• Fan beat rate was the average number of fanning beats per second.
I first calculated the mean value by pooling across all observations for a given male. I then
compared individual means between large and small males, as well as between preferred and non
preferred males, for each variable using Mann-Whitney U tests. I did not used paired statistical tests (i.e.
pairing by male test pair) as there were too few cases where data was available from both males. I also
compared the proportion of spawning attempts in which filial cannibalism occurred between males that
differed in size class (large or small), as well as the number of spawning attempts because this could be
another measure of male attractiveness since it stands to reason females would more readily remate with
an attractive male. Finally, 1 used linear mixed models to analyse variation in hatching time while
controlling for the fact that some males had multiple clutches and treated male identity and pair identity as
random factors.
99 .

3.3 Results
I found no effect of male size or male attractiveness on guarding or fanning behaviours in
Mogurnda mogurnda. There was no difference in the number of occurrences, the total duration or the
mean duration of a bout between large and small males for guarding or fanning behaviours (Table 3.3.1
and Table 3.3.2). There was also no significant difference in these parental care behaviours between
males that differed in attractiveness (Table 3.3.3 and Table 3.3.4). Disappointingly, there was no effect of
male size class (Mann-Whitney U = 105.500, Z = -0.392, p = 0.775) or attractiveness (Mann-Whitney U =
88.500, Z = -1.343, p = 0.325) on the proportion of clutches where filial cannibalism occurred. There were,
however, only eight incidences of filial cannibalism from seven of the 30 males, with only 13% of all
spawning attempts (8 out of 61) showing any incidences of filial cannibalism and they were all whole
clutch cannibalisms. Small samples sizes may have contributed to these non-significant results for these
behaviours as not all males performed all behaviours (see tables for sample sizes).
Unexpectedly, I found that preferred males had significantly lower fanning rate than non-preferred
males (Mann-Whitney U = 43.00, Z = -2.892, p = 0.004, Fig. 3.3.1). Non-preferred males had an average
fanning rate of 1.21 beats per second, and preferred males having a mean fanning rate of 0.73. There
was no effect of male size class on fanning rate (Mann-Whitney U = 94.00, Z = -0.770, p = 0.461).
There was no effect of male size on time to spawning (Fig. 3.3.2; Kaplan Meier, Chi-square =
0.064, df = 1, p = 0.801). Contrary to expectation however, preferred males actually took significantly
longer to spawn than non-preferred males (Kaplan Meier, Chi-square = 5.396, df = 1, p = 0.020;
Fig. 3.3.3). Preferred males spawned after an average of 27.1 days, while non-preferred males took an
average of 7.9 days. Closer analysis of the data, however, showed that five preferred males did not spawn
at all which could explain the result. Interestingly, three of the five males who did not spawn were from
pairs where the smaller male was preferred over the larger male.
There was no difference in the total number of spawning attempts per male between males that
differed in attractiveness (Mann-Whitney U = 86.00, Z = -1.120, p = 0.285) or in size (Mann-Whitney U =
107.00, Z = - 0.232, p = 0.838). Finally, there was no effect of male mass (treated as a continuous
variable) (F = 81.802, df = 58, p = 0.544) or attractiveness (F = 81.026, df = 58, p = 0.570) on the number
of days until eggs hatched.
100.
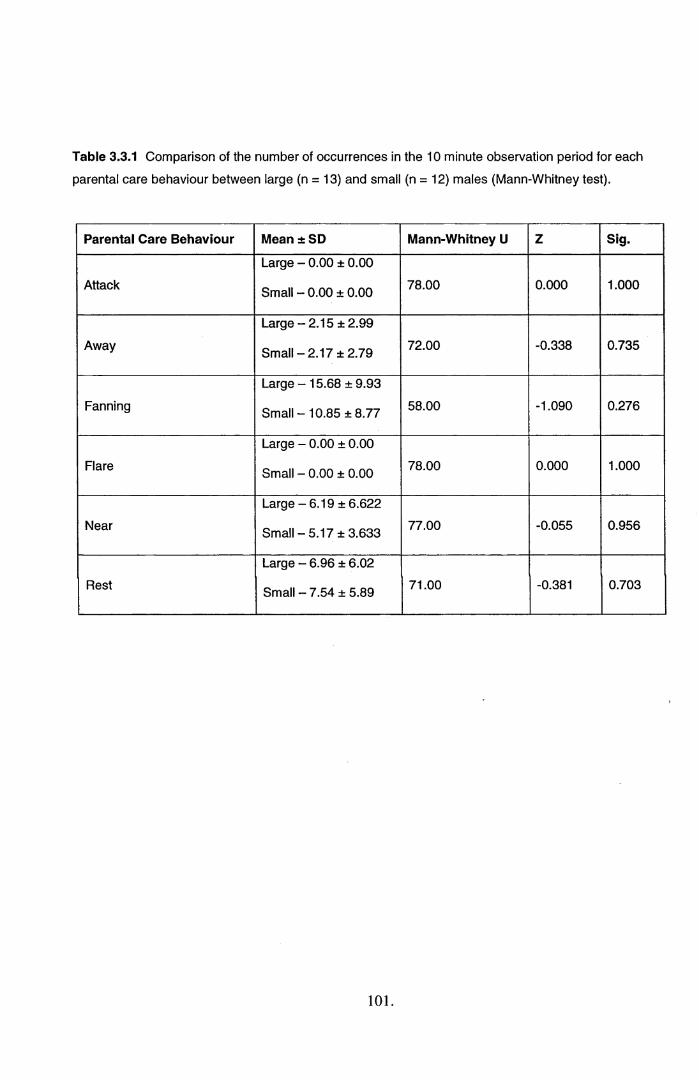
Table 3.3.1 Comparison of the number of occurrences in the 10 minute observation period for each
parental care behaviour between large (n = 13) and small (n = 12) males (Mann-Whitney test).
Parental Care Behaviour Mean ± SD Mann-Whitney U Z Sig.
Attack
Large - 0.00 ± 0.00
Small - 0.00 ± 0.00 78.00 0.000 1.000
Away
Large - 2.15 ± 2.99
Small-2 .17 ±2.79 72.00 -0.338 0.735
Fanning
Large - 15.68 ± 9.93
Small - 10.85 ±8.77 58.00 -1.090 0.276
Flare
Large - 0.00 ± 0.00
Small - 0.00 ± 0.00 78.00 0.000 1.000
Near
Large - 6.19 ± 6.622
Small-5 .17 ±3.633 77.00 -0.055 0.956
Rest
Large — 6.96 ± 6.02
Small-7 .54 ±5.89 71.00 -0.381 0.703
101.

Table 3.3.2 Comparison of the total and mean durations of time (seconds) spent in the 10 minute
observation period engaged in parental care behaviours between large and small males (Mann-Whitney
test).
Variable Behaviour Mean ± SD Mann- Whitney U Z Sig.
Total Away Large - 137.21 ± 205.79 (n=7)
Duration(sec) Small - 137.52 ± 171.31 (n=8) 26.00 -0.231 0.817
Fanning Large - 208.66 ± 177.21 (n=9)
Small - 212.27 ± 199.61 (n=11) 75.50 -0.136 0.892
Near Large - 153.95 ± 107.37 (n=9)
Small - 165.72 ± 134.93 (n=11) 46.00 -0.266 0.790
Rest Large - 197.25 ± 203.08 (n=12)
Small - 122.31 ±87.45 (n=11) 60.00 -0.369 0.712
Mean Away Large - 98.09 ± 220.93 (n=7)
Duration(sec) Small-34.24 ±29.16 (n=8) 21.00 -0.810 0.418
Fanning Large - 19.75 ± 16.55 (n=9)
S m a ll-28.69 ± 24.19 (n=11) 62.50 -0.843 0.399
Near Large - 28.74 ± 28.22 (n=9)
Small - 33.77 ± 35.27 (n=11) 48.00 -0.114 0.909
Rest Large - 49.44 ± 88.44 (n=12)
Small - 17.53 ± 15.20 (n=11) 52.00 -0.862 0.389
102.
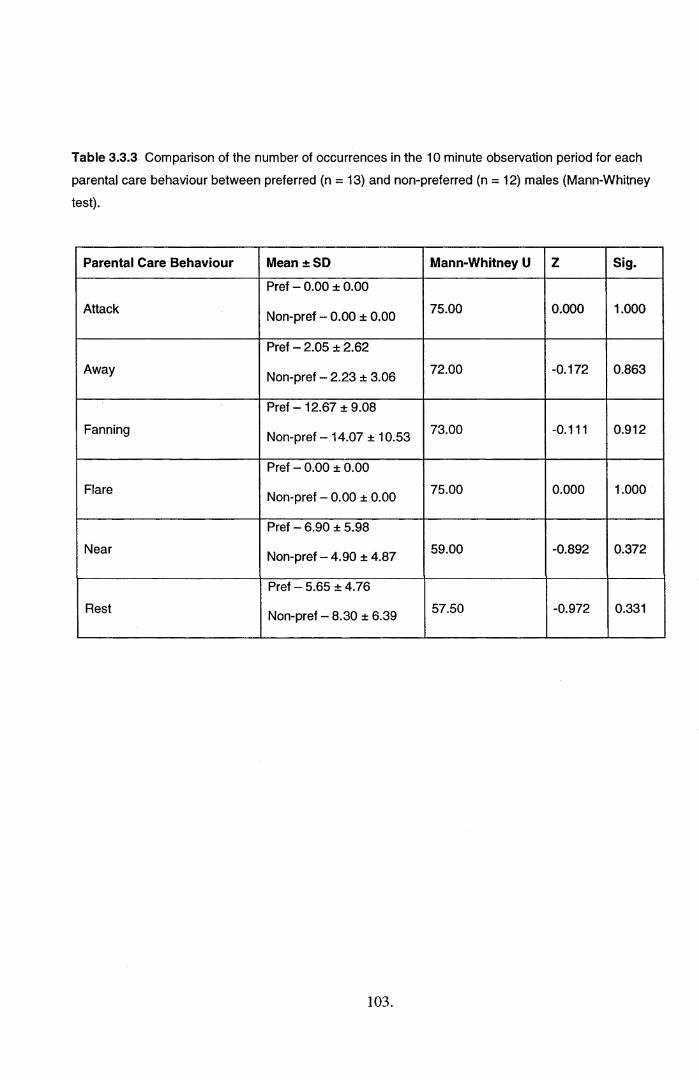
Table 3.3.3 Comparison of the number of occurrences in the 10 minute observation period for each
parental care behaviour between preferred (n = 13) and non-preferred (n = 12) males (Mann-Whitney
test).
Parental Care Behaviour Mean ± SD Mann-Whitney U Z Sig.
Attack
Pref - 0.00 ± 0.00
Non-pref - 0.00 ± 0.00 75.00 0.000 1.000
Away
Pref - 2.05 ± 2.62
Non-pref - 2.23 ± 3.06 72.00 -0.172 0.863
Fanning
Pref-12.67 ±9.08
Non-pref - 14.07 ± 10.53 73.00 -0.111 0.912
Flare
Pref - 0.00 ± 0.00
Non-pref - 0.00 ± 0.00 75.00 0.000 1.000
Near
Pref - 6.90 ± 5.98
Non-pref - 4.90 ± 4.87 59.00 -0.892 0.372
Rest
Pref-5 .65 ±4.76
Non-pref - 8.30 ± 6.39 57.50 -0.972 0.331
103.

Table 3.3.4 Comparison of the total and mean durations of time (seconds) spent engaged in parental
care behaviours between preferred and non-preferred males (Mann-Whitney test).
Variable Behaviour Mean ± SD Mann- Whitney U Z Sig.
TotalDuration(sec)
Away
P re f- 123.12 ±210.67 (n=7)
Non-pref- 149.85 ± 164.99 (n=8) 21.00 -0.810 0.418
Fanning
Pref-223.20 ±191.60 (n=7)
Non-pref - 190.60 ± 182.85 (n=8) 60.50 -0.804 0.421
Near
Pref - 191.12 ±117.00 (n=8)
Non-pref - 139.96 ± 123.06 (n=12) 36.00 -0.926 0.355
Rest
Pref - 162.23 ± 169.08 (n=9)
N on-pref-160.88 ± 160.21 (n=14) 61.00 -0.126 0.900
MeanDuration(sec)
Away
Pref-98.70 ±210.67 (n=7)
Non-pref - 33.70 ± 29.52 (n=8) 24.00 -0.463 0.643
Fanning
Pref-27.37 ±23.69 (n=7)
Non-pref - 19.48 ± 15.47 (n=8) 56.50 -1.026 0.305
Near
Pref - 34.40 ± 30.94 (n=8)
Non-pref - 29.58 ± 33.20 (n=12) 41.00 -0.540 0.589
Rest
Pref-30.28 ±33.81 (n=9)
Non-pref - 36.69 ± 80.78 (n=14) 57.00 -0.378 0.705
104.
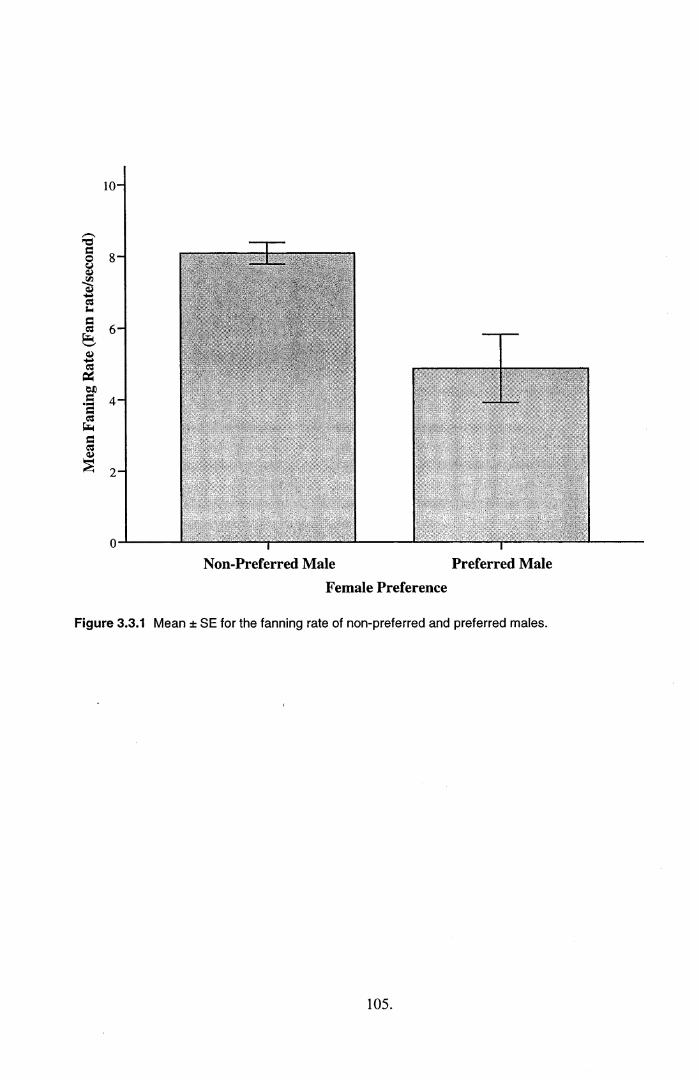
10-
~a
Non-Preferred Male Preferred Male
Female Preference
Figure 3.3.1 Mean ± SE for the fanning rate of non-preferred and preferred males.
105 .
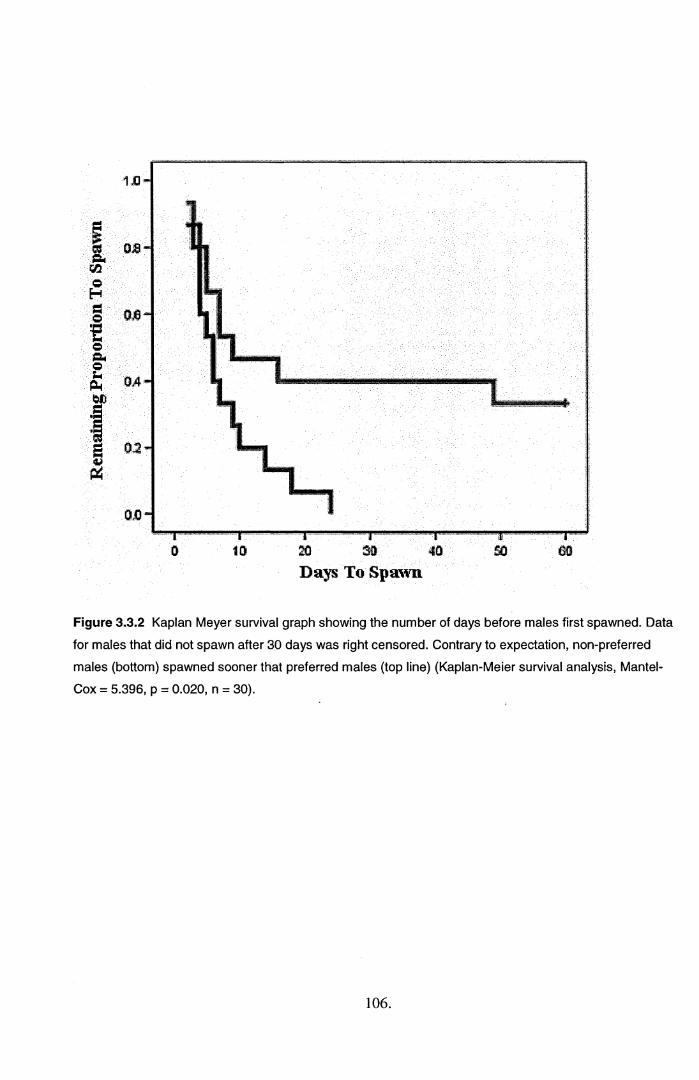
1 0 «
Days To Spawn
Figure 3.3.2 Kaplan Meyer survival graph showing the number of days before males first spawned. Data
for males that did not spawn after 30 days was right censored. Contrary to expectation, non-preferred
males (bottom) spawned sooner that preferred males (top line) (Kaplan-Meier survival analysis, Mantel-
Cox = 5.396, p = 0.020, n = 30).
106.
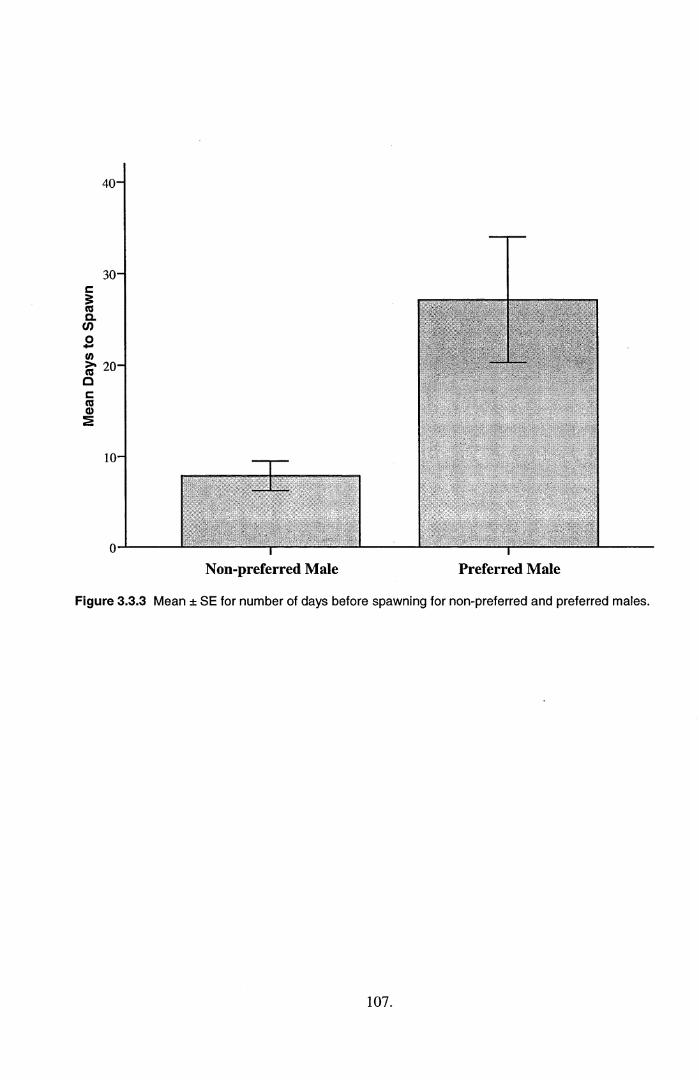
40 -
30 -
c
Non-preferred Male Preferred Male
Figure 3.3.3 Mean ± SE for number of days before spawning for non-preferred and preferred males.
107.

3.4 Discussion
3.4.1. Variation in parental care quality
In this chapter, I first aimed to see if there were any honest (i.e. predictable) cues to male parental
care quality. Interestingly, I did not find any effect of male size on any parental care behaviours, so size
was not a predictor of parental quality. There was no effect of male size on guarding, fanning, filial
cannibalism or egg development time. This suggests that male size (and by association, male dominance)
does not correlate with parental quality.
If there were any cues to male parental care quality, I wanted to compare them to the characters
used by females to select mates in Chapter 2. Male size per se does not predict care, but if we assume
that male attractiveness is correlated to one or more traits that females base their choice on, male
attractiveness could be treated conceptually as a trait when looking for effects on care behaviour. The
non-significant response for all parental care behaviours is repeated for males differing in attractiveness,
except in the case of fanning rate. Unexpectedly, preferred males fanned at a lower rate. This poses a
dilemma as it makes female choice maladaptive, with females actually suffering a cost of decreased male
care from their mating preference. This leads to the question of how female choice is maintained in this
species if females are incurring higher costs for their mating preferences. There are several possible
explanations for the discovery of maladaptive female choice in this study; females could be gaining
material benefits unrelated to parental care or females could be gaining genetic benefits. Another
explanation is that my method for discriminating female preference is not accurate which I discuss later.
Maladaptive female choice
Mogurnda mogurnda joins the list of species in which attractive or more actively courting males
provide less care (Burley 1988; de Lope and Möller 1993; Burley et al. 1996; Qvarnström 1997; Raouf et
al. 1997; Griffth et al. 1998) a prediction generated by the model of differential allocation (Burley 1986). It
is unclear what impact this cost would have on actual reproductive success as I wasn’t able to measure
hatching success. The eggs were too small to count from a distance of 20 cm and males were very
sensitive to predation risk while guarding a clutch and would abandon the eggs if I disturbed them. I had to
be very careful even moving the front cover slightly to observe parental care behaviours, and males
frequently abandoned their eggs during my pilot study until I learned what level of disturbance they would
tolerate. It is difficult to draw quantative conclusions about the cost of this choice but fanning has been
108.

repeatedly demonstrated to effect embryo hatching success so I can safely assume the cost is sizeable.
This therefore raises the question of why female M. mogurnda preferred larger males. As already
discussed, mate choice is costly (Pomiankowski 1987; Iwasa et al. 1991; Pomiankowski et al. 1991) and
especially so in this case, so for female choice to persist there must be benefits to compensate for the
costs associated with mate choice, otherwise it would be more beneficial to mate randomly (Friberg and
Arnqvist 2003). Assuming the measure of preference response is accurate for now, why do female M.
mogurnda prefer larger, more dominant males?
Alternate material benefits
It is generally assumed that stronger, more dominant males are better quality mates (Berglund et
al. 1996) and increased parental care quality is only one possible benefit of mating with them. Parental
care seemed the most likely gain, as females play no role in parental care and males do not provide other
obvious material benefits such as nuptial gifts. Another possible direct benefit of mating with these males,
however, is that they are more likely to protect the female during and after mating. In most taxa, larger
males are more likely to win fights, (e.g., Kotiaho et al. 1997; Hagelin 2002; Lopez et al. 2002; Reichard et
al. 2005) and M. mogurnda are no exception. They are an extremely aggressive species and larger
individuals are better fighters (pers. obs.). Males might therefore protect females during or after spawning
from other aggressive individuals. Another benefit may be that males provide access to superior
resources. Males are very territorial when guarding their eggs, and if males hold territories around
spawning sites, females may benefit from having access to better foraging grounds or better spawning
sites.
Genetic benefits
Indirect genetic benefits could maintain maladaptive female choice if the costs to overall
reproduction success is small. Genetic benefits can lead to choosy females producing more viable and/or
attractive offspring (reviewed in Kokko et al. 2003) and there is a potential genetic benefit if traits that
increase success during male-male competition are heritable (Cordero and Eberhard 2003). The
‘differential resource allocation’ hypothesis predicts a reduction in care from attractive males because
females will increase their parental investment to obtain or retain an attractive mate (Burley 1986) which
presumably results in indirect benefits of mating with that male. Alternately, benefits could be gained by
mating with males that are more genetically complimentary or compatible (e.g., Zeh and Zeh 1996; Brown
1997; 1997; Tregenza and Wedell 2000) due to such effects as increasing major histocompatability
109.

complex heterozygosity (e.g., Hoffman et al. 2007), inbreeding avoidance (e.g., Tregenza & Wedell 2002)
and greater offspring genetic diversity (Garant et al. 2005).
Indirect selection, however, is generally accepted as a weak evolutionary force in comparison to
direct selection on mating preferences (Kirkpatrick 1996; Kirkpatrick and Barton 1997; Houle and
Kondrashov 2002). It is possible that the costs of selecting males that are worse parents are smallas there
was no detectable effect of male attractiveness on any other parental care behaviour, but it is unlikely that
genetic benefits could compensate for this cost.
An alternate interpretation
Several studies on fish have reported a decrease in fanning rate but note that it does not
necessarily result in reduced reproductive success (Sargent and Gebier 1980; Coleman and Fischer
1991; Östlund and Ahnesjö 1998; Jones and Reynolds 1999). Males who fanned for shorter bouts actually
enjoyed increased hatching success in three spined sticklebacks Gasterosteus aculeatus (Sargent and
Gebier 1980), fifteen spined sticklebacks Spinachia spinachia (Östlund and Ahnesjö 1998), bluegill
sunfish Lepomis macrochirus (Coleman and Fischer 1991) and common gobies Pomatoschistus microps
(Jones and Reynolds 1999). Males who fanned for shorter bouts, but more often, enjoyed increased
hatching success implying that the frequency that males fan (not necessarily just the total time fanning)
has a large effect on egg hatching. In this study however, the decrease in fan beat was not matched with
an increased frequency of fanning bout so this explanation is unlikely.
3.4.2. Variation in mate choice
I wanted to see if the males who were preferred in Chapter 2 from traditional mate choice trials
were also preferred using an alternative preference test that is gaining ground as being superior to
traditional tests. Contrary to the results from Chapter 2, females spawned sooner with non-preferred
males. Using a latency to mating measure of female choice (the number of days it took pairs to spawn),
not only did male size have no effect on latency to mating or the number of spawnings, females actually
had shorter latency to mating with non-preferred males than preferred males. There was no effect of
attractiveness on the number of spawnings and no effect of male size on any measure of spawning.
Looking more closely at the data shows that the five males who did not spawn at all were all
preferred males. Interestingly, three of the five males who did not spawn were from pairs where the
smaller male was preferred over the larger male, and there were only four pairs of males where the
110.

smaller male was preferred. In the mate choice trials (see Chapter 2) some females were assigned to
pairs that had males who were significantly larger and smaller than them, so perhaps the mean size of
trialled females was smaller than average in the pairs where the smaller male was preferred. This could
have resulted in artificially smallmales being chosen in those three trials, consequently leading to females
choosing not to spawn with those smallmales in the parental care experiments.
There is a chance that these finding are actually resulting from male exerting mate choice and
delaying or preventing mating with unattractive females. If females are the more competitive sex in this
species, the direction of sexual selection may be the reverse of conventional sex roles i.e. ‘reversed sex
role’; (Gwynne 1991; Vincent et al. 1992; Kvarnemo and Ahnejsö 1996). Sex roles are largely influenced
by the operational sex ratio (OSR) (Emlen and Oring 1977; Clutton-Brock and Parker 1992; Kvarnemo
and Ahnejsö 1996), and the OSR will be skewed towards the sex in which there are more reproductive
adults than the opposite sex (Clutton-Brock and Parker 1992; Kvarnemo and Ahnejsö 1996; Parker and
Simmons 1996; Okuda 1999; Ahnejsö et al. 2001). For example, if suitable spawning sites are scarce for
M. mogurnda, the number of reproductive males will be reduced through the shortage of sites, females will
compete for access to mates, and the OSR may skew towards females (in other words, the sex roles are
reversed). Moreover, the costs of reproduction are much higher for males as they are the sole providers of
care making male mate choice even more probable. Kokko & Monaghan (2001) suggest that the
difference in the cost of breeding between sexes is more important than the OSR in determining the sex
roles and a model by Johnstone et al. (1996) that sex differences in costs of choice are more likely have a
much stronger effect on choosiness.
Alternately, female preference may not actually be for worse fathers because the “preferred”
males in this study may not actually be more attractive that non-preferred males. It is possible that the
female preference responses from traditional choice tests did not tell the whole story. The incongruence of
female preference between the two methods for measuring female choice implies that it is based on more
complex parameters than visual (and possibly chemical) cues, and that its expression into actual
increased mating success for some males over others may not be accurately measured by an indirect
measure such as time association.
Incongruence between the two mating preference tests
More studies are starting to show that simultaneous choice tests are not the most accurate
method of quantifying female choice (Wagner 1998; Shackleton et al. 2005; Cotton et al. 2006a). They do
1 1 1 .

not always exclude male-male interactions that may influence mating success for some males, and those
that do often restrict interactions and cues which can effect the expression of choice (Candolin 1999a;
Nilsson and Nilsson 2000) as female preference may be based on more than one male trait (reviewed in
Candolin 2003). Simultaneously presenting females with two extremes in male phenotype does not
indicate how females respond to intermediates (Wagner 1998). Also, the indirect measures of female
mate choice are usually only assumed to equate to actual mating success (Clark and Biesiadecki 2002).
Latency to mating tests resolves many of these problems (Pilastro et al. 2002), females are allowed full
access to physical and chemical cues males offer, and female mating preference can be determined from
an actual mating.
Are non-preferred males actually more attractive?
So if my time association measure of female choice is inaccurate and the latency to mating test is
more accurate, how does this change the interpretation of the results? Females did not spawn sooner with
larger males so male size does not appear to be a cue of choice. Females may, however, still base their
mating preferences on parental care or a trait correlated with parental care: females spawned sooner with
better fathers (i.e. males that showed higher quality of parental care). Having discounted body size as a
cue, what other cues could females be basing their choice on?
Fanning is the most obvious cue as females spawned sooner with non-preferred males who also
had a higher fan beat rate. Fanning itself has been shown to be a trait for mate choice; sand gobies fan
longer and more frequently in the presence of females (Pampoulie et al. 2004) suggesting the this trait
could also be under sexual selection. Males from many species fan before eggs are in a nest, e.g.,
displacement or ‘courtship fanning’ in sticklebacks (Sevenster 1961). Male Florida flagfish Jordanella
floridae also show fanning behaviour prior to the presence of eggs, and male mating success has been
correlated with both prespawning and early postspawning fanning (St Mary et al. 2001). Fanning care
behaviours can also be an honest signal of male quality. Östlund and Ahnesjö (1998) found that female
fifteen-spined sticklebacks preferred males that court more (which is positively correlated to fan beat rate)
and that males who increased their fin beat rate to a high level lost more weight than males who did not.
Courtship and fan beat rate are therefore condition-dependent traits and appear to provide honest cues
for female choice (Zahavi 1975). Courtship can also signal parental care quality (e.g., Knapp and Kovach
1991; Karino 1995; Östlund and Ahnesjö 1998; Wong 2004).
1 12.

References
Ahnejsö I, Kvarnemo C, Merilaita S (2001) Using potential reproductive rates to predict mating competition among individuals qualified to mate. Behavioral Ecology 12:397-401
Amundsen T, Forsgren E (2001) Male mate choice selects for female coloration in a fish. Proceedings of the National Academy of Sciences, U.S.A. 98:13155-13160
Balshine-Earn S (1995) The costs of parental care in Galilee St Peter's fish, Sarotherodon galilaeus. Animal Behaviour 50:1-7
Bateman AJ (1948) Intra-Sexual Selection In Drosophila. Heredity 2:349-368
Belles-lsles JC, FitzGerald GJ (1991) Filial cannibalism in sticklebacks - a reproductive management strategy. Ethology Ecology & Evolution 3:49-62
Berglund A, Bisazza A, Pilastro A (1996) Armaments and ornaments: An evolutionary explanation of traits of dual utility. Biological Journal of the Linnean Society 58:385-399
Bisazza A, Marconato A (1988) Female mate choice, male-male competition and parental care in the river bullhead, Cottus gobio L. (Pisces, Cottidae). Animal Behaviour 36:1352-1360
Blumer LS (1979) Male parental care in the bony fishes. Quarterly Review of Biology 54:149-161
Blumstein DT, Evans CS, Daniel JC (2000) JWatcher. In, 0.9 edn. Animal Behaviour Laboratory,Macquarie University, Sydney, pp A powerful tool for the quantitative analysis of behaviour.
Brown JL (1997) A theory of mate choice based on heterozygosity. Behavioral Ecology 8:60-65
Buchanan KL, Catchpole CK (2000) Song as an indicator of male parental effort in the sedge warbler. Proceedings of the Royal Society of London Series B-Biological Sciences 267:321-326
Burley N (1986) Sexual selection for aesthetic traits in species with biparental care. American Naturalist 127:415-445
Burley N (1988) The differential-allocation hypothesis: an experimental test. American Naturalist 132:611- 628
Burley NT, Parker PG, Lundy K (1996) Sexual selection and extrapair fertilization in a sociallymonogamous passerine, the zebra finch (Taeniopygia guttata). Behavioral Ecology 7:218-226
Candolin U (1999a) Male-male competition facilitates female choice in sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 266:785-789
Candolin U (1999b) The relationship between signal quality and physical condition: is sexual signalling honest in the three spined stickleback? Animal Behaviour 58:1261-1267
Chellappa S, Huntingford SA (1989) Depletion of energetic reserves during reproductive aggression in male three spined stickleback, Gasterosteus aculeatus. Journal of Fish Biology 35:275-286
1 1 3 .

Clark DL, Biesiadecki B (2002) Mating success and alternative reproductive strategies of the dimorphic jumping spider, Maevia inclemens (Aaneae, Salticidae). Journal of Arachnology 30:511-518
Clutton-Brock TH (1991) The Evolution of Parental Care. Princeton University Press, Princeton
Clutton-Brock TH, Parker GA (1992) Potential reproductive rates and the operation of sexual selection. Quarterly Review of Biology 67:437-456
Coleman RM, Fischer RU (1991) Brood size, male fanning effort and the energetics of a nonshareable parental investment in bluegill sunfish, Lepomis-macrochirus (teleostei, centrarchidae). Ethology87:177-188
Coleman RM, Gross MR, Sargent RC (1985) Parental investment decision rules - a test in bluegill sunfish. Behavioral Ecology and Sociobiology 18:59-66
Cooke SJ, Schreer JF, Philipp DP, Weatherhead PJ (2003) Nesting activity, parental care behavior, and reproductive success of smallmouth bass, Micropterus dolomieu, in an unstable thermal environment. Journal of Thermal Biology 28:445-456
Cordero C, Eberhard WG (2003) Female choice of sexually antagonistic male adaptations: a critical review of some current research. Journal of Evolutionary Biology 16:1-6
Cöte IM, Hunte W (1989) Male and female mate choice in the redlip blenny - why bigger is better. Animal Behaviour 38:78-88
Cotton S, Rogers DW, smallJ, Pomiankowski A, Fowler K (2006) Variation in preference for a maleornament is positively associated with female eyespan in the stalk-eyed fly Diasemopsis meigenii. Proceedings of the Royal Society of London Series B-Biological Sciences 273:1287-1292
Cunninqham EJA, Russell AF (2000) Eqq investment is influenced by male attractiveness in the mallard. Nature 404:74-77
Dawkins R, Carlisle TR (1976) Parental investment, mate desertion and a fallacy. Nature 262:131-133
de Lope F, Möller AP (1993) Female reproductive effort depends on the degree of ornamentation of their mates. Evolution 47:1152-1160
Demartini EE (1987) Paternal defense, cannibalism and polygamy - factors influencing the reproductive success of painted greenling (pisces, hexagrammidae). Animal Behaviour 35:1145-1158
Downhower JF, Brown L (1980) Mate preferences of female mottled sculpins, Cottus bairdi. Animal Behaviour 28:728-734
Emlen ST, Oring LW (1977) Ecology, sexual selection, and the evolution of mating systems. Science 197:215-223
FitzGerald GJ (1992) Filial cannibalism in fishes: Why do parents eat their offspring? Trends in Ecology & Evolution 7:7-10
Fitzpatrick S, Berglund A, Rosenqvist G (1995) Ornaments or offspring: costs to reproductive success restrict sexual selection processes. Biological Journal of the Linnean Society 55:251-260
114.

Forsgren E (1997) Female sand gobies prefer good fathers over dominant males. Proceedings of the Royal Society of London Series B-Biological Sciences 264:1283-1286
Forsgren E, Karlsson A, Kvarnemo C (1996) Female sand gobies gain direct benefits by choosing males with eggs in their nests. Behavioral Ecology and Sociobiology 39:91-96
Friberg U, Arnqvist G (2003) Fitness effects of female mate choice: preferred males are detrimental for Drosophila melanogasterfemales. Journal of Evolutionary Biology 16:797-811
Garant D, Dodson JD, Bernatchez L (2005) Offspring genetic diversity increases fitness of female Atlantic salmon (Salmo salar). Behavioral Ecology and Sociobiology 57:240-244
Goto A (1987) Polygyny in the river sculpin, Cottus hangiongensis (Pisces: Cottidae), with special reference to male mating success. Copeia 1:32-40
Goulet D (1998) Spawning success in the damselfish Amblyglyphidodon leucogaster: the influence of eggs in the nest. Animal Behaviour 55:651-664
Grafen A (1990) Biological signals as handicaps. Journal of Theoretical Biology 144:517-546
Green BS, McCormick Ml (2005) 02 replenishment to fish nests: males adjust brood care to ambient conditions and brood development. Behavioral Ecology and Sociobiology 16:389-397
Griffth SC, Owens IPF, Burke T (1998) Sexual selection in the house sparrow: the best males are the worst fathers. (In preparation.)
Gross MR, Sargent RC (1985) The evolution of male and female parental care in fishes. American Zoologist 25:807-822
Guderley H, Couture P (2005) Stickleback fights: why do winners win? Influence of metabolic and morphometric parameters. Physiological and Biochemical Zoology 78:173-181
Gwynne DT (1991) Sexual competition among females: what causes courtship-role reversal? . Trends in Ecology & Evolution 6:118-121
Hagelin JC (2002) The kinds of traits involved in male-male competition: a comparison of plumage, behavior, and body size in quail. Behavioral Ecology 13:32-41
Hale RE, St Mary CM, Lindström K (2003) Parental responses to changes in costs and benefits along an environmental gradient. Environmental Biology of Fishes 67:107-116
Hastings PA (1988) Female choice and male reproductive success in the angel blenny, Coralliozetus- angelica (teleostei, chaenopsidae). Animal Behaviour 36:115-124
Heywood JS (1989) Sexual selection by the handicap mechanism. Evolution 43:1387-1397
Hoelzer G (1988) Filial cannibalism in a non-brood cycling marine fish. Environmental Biology of Fishes 21:309-313
Hoelzer GA (1989) The good parent process of sexual selection. Animal Behaviour 38:1067-1078
1 1 5 .

Hoelzer GA (1992) The ecology and evolution of partial clutch cannibalism by paternal Cortez Damselfish. Oikos 65:113-120
Hoffman Jl, Forcada J, Trathan PN, Amos W (2007) Female fur seals show active choice for males that are heterozygous and unrelated. Nature 445:912-914
Houde AE, Endler JA (1990) Correlated evolution of female mating preferences and male color patterns in the guppy Poecilia reticulata. Science 248:1405-1408
Houle D, Kondrashov AS (2002) Coevolution of costly mate choice and condition-dependent display of good genes. Proceedings of the Royal Society of London Series B-Biological Sciences 269:97- 104
Houston AI, Szekely T, McNamara JM (2005) Conflict between parents over care. Trends in Ecology & Evolution 20:33-38
Hunt J, Brooks R, Jennions MD, Smith MJ, Bentsen CL, Bussiere LF (2004) High-quality male field crickets invest heavily in sexual display but die young. Nature 432:1024-1027
Itzkowitz M, Makie D (1986) Food provisioning and foraging behavior of the snowy egret, Egretta-thula. Biology of Behaviour 11:111-115
Iwasa Y, Pomiankowski A, Nee S (1991) The evolution of costly mate preferences. II. The ‘handicap’ principle. Evolution 45:1431-1442
Jennions MD, Polakow DA (2001) The effect of partial brood loss on male desertion in a cichlid fish: an experimental test. Behavioral Ecology 12:84-92
Johnstone RA, Reynolds JD, Deutsch JC (1996) Mutual mate choice and sex differences in choosiness. Evolution 50:1382-1391
Jones JC, Reynolds JD (1999) Costs of egg ventilation for male common gobies breeding in conditions of low dissolved oxygen. Animal Behaviour 57:181-188
Kamler E (1992) Early life histories of fish: an energetics approach. Chapman & Hall, London
Karino K (1995) Male-male competition and female mate choice through courtship display in the territorial damselfish Stegastes nigricans. Ethology 100:126-138
Karino K (1997) Influence of brood size and offspring size on parental investment in a biparental cichlid fish, Neolamprologus moorii. Journal of Ethology 15:39-43
Karino K, Arai R (2006) Effect of clutch size on male egg-fanning behavior and hatching success in thegoby, Eviota prasina (Klunzinger). Journal of Experimental Marine Biology and Ecology 334:43-50
Keenleyside MHA, Rangeley RW, Küppers BU (1985) Female mate choice and male parental defense behaviour in the cichlid fish Cichlasoma nigrofasciatum. Canadian Journal of Zoology 63:2489- 2493
Kirkpatrick M (1985) Evolution of female choice and male parental investment in polygynous species: the demise of the “sexy son”. American Naturalist 125:788-810
116.

Kirkpatrick M (1996) Good genes and direct selection in evolution of mating preferences. Evolution 50:2125-2140
Kirkpatrick M, Barton NH (1997) The strength of indirect selection on female mating preferences.Proceedings of the National Academy of Sciences of the United States of America 94:1282-1286
Klug H, Chin A, St Mary CM (2005) The net effects of guarding on egg survivorship in the flagfish, Jordanella floridae. Animal Behaviour 69:661-668
Knapp RA, Kovach JT (1991) Courtship as an honest indicator of male parental quality in the bicolor damselfish, Stegastes partitus. Behavioral Ecology 2:295-300
Knapp RA, Warner RR (1991) Male parental care and female choice in the bicolor damselfish, Stegastes partitus: bigger is not always better. Animal Behaviour 41:747-756
Kokko H (1998) Should advertising parental care be honest? Proceedings of the Royal Society of London Series B-Biological Sciences 265:1871-1878
Kokko H (2001) Fisherian and "good genes" benefits of mate choice: how (not) to distinguish between them. Ecology Letters 4:322-326
Kokko H, Brooks R, Jennions MD, Morley J (2003) The evolution of mate choice and mating biases. Proceedings of the Royal Society of London Series B-Biological Sciences 270:653-664
Kokko H, Monaghan P (2001) Predicting the direction of sexual selection. Ecology Letters 4:159-165
Kotiaho J, Alatalo RV, Mappes J, Parri S (1997) Fighting success in relation to body mass and drumming activity in the male wolf spider Hygrolycosa rubrofasciata. Canadian Journal of Zoology-Revue Canadienne De Zoologie 75:1532-1535
Kotiaho JS (2001) Costs of sexual traits: a mismatch between theoretical considerations and empirical evidence. Biological Reviews 76:365-376
Kraak SBM (1996) Female preference and filial cannibalism in Aidablennius sphynx (Teleostei, Blenniidae); A combined field and laboratory study. Behavioural Processes 36:85-97
Kraak SBM, Groothuis TGG (1994) Female preference for nests with eggs is based on the presence of the eggs themselves. Behaviour 131:189-206
Kraak SBM, Videler JJ (1991) Mate choice in aidablennius-sphynx (teleostei, blenniidae) - females prefer nests containing more eggs. Behaviour 119:243-266
Kramer DL (1987) Dissolved oxygen and fish behavior. Environmental Biology of Fishes 18:81-92
Kunzler R, Bakker TCM (2000) Pectoral fins and paternal quality in sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 267:999-1004
Kvarnemo C (1997) Food affects the potential reproductive rates of sand goby females, but not of males. Behavioral Ecology 8:605-611
117.

Kvarnemo C, Ahnejsö I (1996) The dynamics of operational sex ratios and competition for mates. Trends in Ecology & Evolution 11:404-408
Kvarnemo C, Svensson O, Forsgren E (1998) Parental behaviour in relation to food availability in the common goby. Animal Behaviour 56:1285-1290
Lavery RJ, Keenleyside MHA (1990) Parental investment of a biparental cichlid fish, cichlasoma-nigrofasciatum, in relation to brood size and past investment. Animal Behaviour 40:1128-1137
Lindström K (1988) Male-male competition for nest sites in the sand goby, Pomatoschistus-minutus. Oikos 53:67-73
Lindström K (1998) Effects of costs and benefits of brood care on filial cannibalism in the sand goby. Behavioral Ecology and Sociobiology 42:101-106
Lindström K, Sargent RC (1997) Food access, brood size and filial cannibalism in the fantail darter, Etheostoma flabellare. Behavioral Ecology and Sociobiology 40:107-110
Linville SU, Breitwisch R, Schilling AJ (1998) Plumage brightness as an indicator of parental care in northern cardinals. Animal Behaviour 55:119-127
Lissaker M, Kvarnemo C, Svensson O (2003) Effects of a low oxygen environment on parental effort and filial cannibalism in the male sand goby, Pomatoschistus minutus. Behavioral Ecology 14:374-381
Lopez P, Munoz A, Martin J (2002) Symmetry, male dominance and female mate preferences in the Iberian rock lizard, Lacerta monticola. Behavioral Ecology and Sociobiology 52:342-347
Mackereth RW, Noakes DLG, Ridgway MS (1999) Size-based variation in somatic energy reserves and parental expenditure by male smallmouth bass, Micropterus dolomieu. Environmental Biology of Fishes 56:263-275
Maekawa K, Iguchi K, Katano O (1996) Reproductive success in male Japanese minnows,Pseudorasbora parva: Observations under experimental conditions. Ichthyological Research 43:257-266
Manica A (2002a) Alternative strategies for a father with a smallbrood: mate, cannibalise or care. Behavioral Ecology and Sociobiology 51:319-323
Manica A (2002b) Filial cannibalism in teleostfish. Biological Reviews 77:261-277
Manica A (2003) The effect of brood size and age on partial filial cannibalism in the scissortail sergeant. Journal of Fish Biology 63:37-47
Mank JE, Promislow DEL, Avise JC (2005) Phylogenetic perspectives in the evolution of parental care in ray-finned fishes. Evolution 59:1570-1578
Marconato A, Bisazza A (1986) MALES WHOSE NESTS CONTAIN EGGS ARE PREFERRED BY FEMALE COTTUS-GOBIO L (PISCES, COTTIDAE). Animal Behaviour 34:1580-1582
Marconato A, Bisazza A, Fabris M (1993) The cost of parental care and egg cannibalism in the river bullhead, cottus-gobio I (pisces, cottidae). Behavioral Ecology and Sociobiology 32:229-237
118.

Markman S, Yomtov Y, Wright J (1995) Male parental care in the orange-tufted sunbird - behavioral- adjustments in provisioning and nest guarding effort. Animal Behaviour 50:655-669
Marques PAM (2003) Male mate desertion in the Spanish sparrow Passer hispaniolensis. Ardea 91:245- 250
Marques PAM (2004) Parental care, male desertion, and reproductive success in the Spanish sparrow, Passer hispaniolensis. Zoological Studies 43:123-131
McNamara JM, Szekely T, Webb JN, Houston AI (2000) A dynamic game-theoretic model of parental care. Journal of Theoretical Biology 205:605-623
Möller AP, Thornhill R (1998) Male parental care, differential parental investment by females and sexual selection. Animal Behaviour 55:1507-1515
Neff BD, Gross MR (2001) Dynamic adjustment of parental care in response to perceived paternity. Proceedings of the Royal Society of London Series B-Biological Sciences 268:1559-1565
Nilsson SO, Nilsson GE (2000) Free choice by female sticklebacks: lack of preference for maledominance traits. Canadian Journal of Zoology-Revue Canadienne De Zoologie 78:1251-1258
Noonan K (1983) Female mate choice in a cichlid fish Cichlasoma nigrofasciatum. Animal Behaviour 31:1005-1010
Okuda N (1999) Sex roles are not always reversed when the potential reproductive rates is higher in females. American Naturalist 153:540-548
Okuda N, Ito S, Iwao H (2002) Female spawning strategy in Rhinogobius sp OR: how do females deposit their eggs in the nest? Ichthyological Research 49:371-379
Okuda N, Yanagisawa Y (1996) Filial cannibalism by mouthbrooding males of the cardinal fish, Apogon doederleini, in relation to their physical condition. Environmental Biology of Fishes 45:397-404
Osorno JL, Szekely T (2004) Sexual conflict and parental care in magnificent frigatebirds: full compensation by deserted females. Animal Behaviour 68:337-342
Östlund S, Ahnesjö I (1998) Female fifteen-spined sticklebacks prefer better fathers. Animal Behaviour 56:1177-1183
Pampoulie C, Lindstrom K, St Mary CM (2004) Have your cake and eat it too: male sand gobies show more parental care in the presence of female partners. Behavioral Ecology 15:199-204
Parker GA, Royle NJ, Hartley IR (2002) Intrafamilial conflict and parental investment: a synthesis.Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 357:295- 307
Parker GA, Simmons LW (1996) Parental investment and the control of sexual selection: predicting the direction of sexual competition. Proceedings of the Royal Society of London Series B-Biological Sciences 263:315-321
119.

Payne AG, Smith C, Campbell AC (2002) Filial cannibalism improves survival and development of beaugregory damselfish embryos. Proceedings of the Royal Society of London Series B- Biological Sciences 269:2095-2102
Payne AG, Smith C, Campbell AC (2004) A model of oxygen-mediated filial cannibalism in fishes. Ecological Modelling 174:253-266
Petersen CW (1990) The occurrence and dynamics of clutch loss and filial cannibalism in two Caribbean damselfishes. Journal of Experimental Marine Biology and Ecology 135:117-133
Pilastro A, Evans JP, Sartorelli S, Bisazza A (2002) Male phenotype predicts insemination success in guppies. Proceedings of the Royal Society of London Series B-Biological Sciences 269:1325- 1330
Polis GA (1981) The evolution and dynamics of intraspecific predation. Annual Review of Ecology and Systematics 12:225-251
Pomiankowski A (1987) The costs of choice in sexual selection. Journal of Theoretical Biology 128:195- 218
Pomiankowski A, Iwasa Y, Nee S (1991) The evolution of costly mate preferences. I. Fisher and biased mutation. Evolution 45:1422-1430
Queller DC (1997) Why do females care more than males? Proceedings of the Royal Society of London Series B-Biological Sciences 264:1555-1557
Qvarnström A (1997) Experimentally increased badge size increases male competition and reduces male parental care in the collared flycatcher. Proceedings of the Royal Society of London Series B- Biological Sciences 264:1225-1231
Raouf SA, Parker PG, Ketterson ED, Nolan V, Ziegenfus C (1997) Testosterone affects reproductivesuccess by influencing extra-pair fertilizations in male dark-eyed juncos (Aves: Junco hyemalis). Proceedings of the Royal Society of London Series B-Biological Sciences 264:1599-1603
Reebs SG, Whoriskey FG, FitzGerald GJ (1984) Diel patterns of fanning activity, egg respiration, and the nocturnal behavior of male 3-spined sticklebacks, Gasterosteus-aculeatus I (f-trachurus). Canadian Journal of Zoology-Revue Canadienne De Zoologie 62:329-334
Reichard M, Bryja J, Ondrackova M, Davidova M, Kaniewska P, Smith C (2005) Sexual selection for male dominance reduces opportunities for female mate choice in the European bitterling (Rhodeus sericeus). Molecular Ecology 14:1533-1542
Reimchen TE (1989) Loss of nuptial color in threespine sticklebacks (Gasterosteus aculeatus). Evolution 43:450-460
Reynolds JD, Jones JC (1999) Female preference for preferred males is reversed under low oxygen conditions in the common goby (Pomatoschistus microps). Behavioral Ecology 10:149-154
Rogers W, Barlow GW (1991) Sex differences in mate choice in a monogamous biparental fish, the midas cichlid (Cichlasoma citrinellum). Ethology 87:249-261
120.

Rohwer S (1978) Parent cannibalism of offspring and egg raiding as a courtship strategy. American Naturalist 112:429-440
Sanz JJ, Kranenbarg S, Tinbergen JM (2000) Differential response by males and females to manipulation of partner contribution in the great tit (Parus major). Journal of Animal Ecology 69:74-84
Sargent RC (1985) Territoriality and reproductive tradeoffs in the 3-spine stickleback, gasterosteus- aculeatus. Behaviour 93:217-226
Sargent RC (1988) Paternal care and egg survival both increase with clutch size in the fathead minnow, pimephales-promelas. Behavioral Ecology and Sociobiology 23:33-37
Sargent RC (1992) Ecology of filial cannibalism in fish: theoretical perspectives. In: Elgar MAC, B. J. (ed) Cannibalism: ecology and evolution among diverse taxa. Oxford University Press, pp 38-62
Sargent RC (1997) Parental care. In: Godin JGJ (ed) Behavioural Ecology of Teleost Fishes. Oxford University Press, Oxford, pp 292-315
Sargent RC, Crowley PH, Huang C, Lauer M, Neergaard D, Schmoetzer L (1995) A dynamic program for male parental care in fishes: Brood cycling and filial cannibalism. Behaviour 132:1059-1078
Sargent RC, Gebier JB (1980) Effects of nest site concealment on hatching success, reproductivesuccess, and paternal behavior of the threespine stickleback, Gasterosteus-aculeatus. Behavioral Ecology and Sociobiology 7:137-142
Sargent RC, Gross MR (1985) Parental investment decision rules and the concorde fallacy. Behavioral Ecology and Sociobiology 17:43-45
Schaffer NM (1974) Optimal reproductive effort in fluctuating environments. American Naturalist 108:783- 798
Schwagmeyer PL, Mock DW, Parker GA (2002) Biparental care in house sparrows: negotiation or sealed bid? Behavioral Ecology 13:713-721
Sevenster P (1961) A causal analysis of a displacement activity (fanning in Gasterosteus aculeatus L.). Behaviour 9 (Suppl.)
Shackleton MA, Jennions MD, Hunt J (2005) Fighting success and attractiveness as predictors of male mating success in the black field cricket, Teleogryllus commodus\ the effectiveness of no-choice tests. Behavioral Ecology and Sociobiology 58:1-8
Sheldon BC (2000) Differential allocation: tests, mechanisms and implications. Trends in Ecology and Evolution 15:397-402
Skolbekken R, Utne-Palm AC (2001) Parental investment of male two-spotted goby, Gobiusculus flavescens (Fabricius). Journal of Experimental Marine Biology and Ecology 261:137-157
Slagsvold T, Lifjeld JT (1988) Plumage color and sexual selection in the pied flycatcher Ficedula- hypoleuca. Animal Behaviour 36:395-407
121.

Smiseth PT, Amundsen T (2000) Does female plumage coloration signal parental quality? A male removal experiment with the bluethroat (Luscinia s. svecica). Behavioral Ecology and Sociobiology 47:205- 212
Smiseth PT, Ornborg J, Andersson S, Amundsen T (2001) Is male plumage reflectance correlated with paternal care in bluethroats? Behavioral Ecology 12:164-170
Smith C (1992) Filial cannibalism as a reproductive strategy in care-giving teleosts. Netherlands Journal of Zoology 42:607-613
Smith C, Wootton RJ (1995) The costs of parental care in teleost fishes. Reviews in Fish Biology and Fisheries 5:7-22
Soler JJ, de Neve L, Martinez JG, Soler M (2001) Nest size affects clutch size and the start of incubation in magpies: an experimental study. Behavioral Ecology 12:301-307
St Mary CM, Noureddine CG, Lindström K (2001) Environmental effects on male reproductive success and parental care in the Florida flagfish Jordanella floridae. Ethology 107:1035-1052
Stott MK, Poulin R (1996) Parasites and parental care in male upland bullies (Eleotridae). Journal of Fish Biology 48:283-291
Suk HY, Choe JC (2002) The presence of eggs in the nest and female choice in common freshwater gobies (Rhinogobius brunneus). Behavioral Ecology and Sociobiology 52:211-215
Svensson OLA, Magnhagen C, Forsgren E, Kvarnemo C (1998) Parental behaviour in relation to the occurrence of sneaking in the common goby. Animal Behaviour 56:175-179
Takegaki T, Nakazono A (1999) Responses of the egg-tending gobiid fish Valenciennes longipinnis to the fluctuation of dissolved oxygen in the burrow. Bulletin of Marine Science 65:815-823
Tomkins JL, Radwan J, Kotiaho JS, Tregenza T (2004) Genic capture and resolving the lek paradox. Trends in Ecology & Evolution 19:323-328
Torricelli P, Lugli M, Gandolfi G (1985) A quantitative-analysis of the fanning activity in the male Padogobius martensi (pisces, gobiidae). Behaviour 92:288-301
Tregenza T, Wedell N (2000) Genetic compatibility, mate choice and patterns of parentage: Invited review. Molecular Ecology 9:1013-1027
Trivers R (1985) Social Evolution. The Benjamin/Cummings Publishing Company, New York
Trivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Sexual Selection and the Descent of man. Aldine Publishing Company, London, pp 136-179
Unger LM, Sargent RC (1988) Allopaternal care in the fathead minnow, Pimephales promelas - Females prefer males with eggs. Behavioral Ecology and Sociobiology 23:27-32
van lersel JJA (1953) An analysis of parental behaviour of the male three spined stickleback (Gasterosteus aculeatus L.). Behaviour 3 (Suppl.):1-159
122.

Vincent A, Ahnejsö I, Berglund A, Rosenqvist G (1992) Pipefishes and seahorses: are they all sex role reversed? Trends in Ecology & Evolution 7:237-241
Vinyoles D, Cöte IM, de Sostoa A (1999) Egg cannibalism in river blennies: the role of natural prey availability. Journal of Fish Biology 55:1223-1232
Wagner WEJ (1998) Measuring female mating preferences. Animal Behaviour 55:1029-1042
Webb JN, Houston AI, McNamara JM, Szekely T (1999) Multiple patterns of parental care. Animal Behaviour 58:983-993
Whittingham LA, Dunn PO, Robertson RJ (1994) Female response to reduced male parental care in birds - an experiment in tree swallows. Ethology 96:260-269
Williams GC (1966) Natural selection costs of reproduction and a refinement of Lacks principle. American Naturalist 100:687-&
Winkler DW (1987) A general model for parental care. American Naturalist 130:526-543
Wong BBM (2004) Superior fighters make mediocre fathers in the Pacific blue-eye fish. Animal Behaviour 67:583-590
Wong BBM, Candolin U (2005) How is female choice affected by male competition? Biological Review 80:559-571
Wootton RJ (1998) Ecology of teleost fishes. Kluwer, Dordrecht
Wright J, Cuthill I (1989) MANIPULATION OF SEX-DIFFERENCES IN PARENTAL CARE. Behavioral Ecology and Sociobiology 25:171-181
Zeh JA, Zeh DW (1996) The evolution of polyandry I: Intragenomic conflict and genetic incompatibility. Proceedings of the Royal Society of London Series B-Biological Sciences 263:1711-1717
Zeh JA, Zeh DW (1997) The evolution of polyandry 2: Post-copulatory defences against geneticincompatibility. Proceedings of the Royal Society of London Series B-Biological Sciences 264:69- 75
Zoran MJ, Ward JA (1983) Parental egg care behavior and fanning activity for the orange chromide, Etroplus maculatus. Environmental Biology of Fishes 8:301-310
123.

CHAPTER 4 Foraging behaviour and substrate
discrimination in the northern trout
gudgeon (Mogurnda mogurnda)
4.1 Introduction
Optimal foraging models have proven useful in predicting prey and habitat use by foragers
(Mittelbach 1981; Werner and Mittelbach 1981; Werner et al. 1983; Benkman 1987; Osenberg and
Mittelbach 1989). Optimal foraging theory is devoted to four main types of questions (and the
associated behaviour that animals exhibit): which food types an animal should eat; which patch types
an animal should feed in; how much time an animal should devote to feeding in different patches; and
what pattern or speed of movements an animal should engage in while foraging (reviewed in Pyke et
al. 1977).
The choice of foraging habitat encompasses both the question of which patches an animal
should feed in and of how long they should spend there. Habitats vary in food density and microhabitat
complexity, which will affect the optimal search times (and ultimately the food values per unit of time).
As well as theses specific factors, it is important to remember that many other internal and external
factors can affect habitat choice such as predation risk, social interactions, hunger levels, habitat
structure, reproductive state, dietary constraints and even phylogenetic factors (reviewed in Perry and
Pianka 1997).
4.1.1 Food density and distribution
Optimal foraging theory predicts the movement and behaviour of foragers in relation to food
densities. The ideal free distribution (IFD) states that if all individuals are equally “free” to occupy any
space in the habitat, then in environments where resources have patchy distributions relatively high
quality areas are expected to contain more individuals than relatively low quality areas. This theory
assumes, however, that there are no competitive asymmetries among individual so that all individuals
gain equal access to resources (Fretwell and Lucas 1970). Although most wild populations do not
conform to the assumptions of the IFD model (Inman 1990) the theory has still been very useful for
predicting patterns of foraging behaviour. For example, Milinski (1979) demonstrated that by adding a
food source at different rates on two sides of an aquarium, he could generate an ideal free distribution
among three spined sticklebacks Gasterosteus aculeatus. More recently, Collins et al. (2005) found
124.

that the mean size of marine scavenging fish increased significantly with ocean depth while in non
scavengers there was a significant decline in size, explaining this dissimilarity in distribution with
simple energetic models that matched the distribution of fish to different patterns of food distribution
for each group.
The assumption that there is no variation in competitive abilities is clearly an over
simplification. Since animals do differ in their competitive abilities, territory distributions are expected to
become ideal despotic, with dominant individuals securing the highest quality territories while less able
competitors are forced into unfavourable habitats. Schneider (1984) also showed that dominance
affected foraging strategies; less dominant individuals suffered more aggression while feeding than
more dominant individuals in while-throated sparrows Zonotrichia albicollis. White-throated sparrows
also show a marked preference for habitat in dense shrubbery, arguably because it provides greater
protection from predators (Grubb and Greenwald 1982). In an experimental study, Schneider (1984)
showed that dominant individuals fed from sites nearer to shelter than less dominant individuals as
their rank allowed them greater access to preferred sites. Interestingly, Schneider (1984) also showed
that predation risk was a greater contribution factor to foraging behaviour than optimal energy intake:
individuals depleted the resources close to the shelter before moving to feeding at sites further from
cover even when such sites provided higher energy intake. This is one example of how maximising
the amount of energy intake per unit time may not be an optimal solution for all animals (Milinski and
Heller 1978; Caraco et al. 1980; Krebs 1980; Sih 1980; van den Berghe el al. 1989).
Milinski (1982) showed the effect of different competitive ability on foraging in three spined
sticklebacks. First, individuals were rated in terms of their relative competitive ability in pairwise
foraging tests. Fish were then presented with large and small Daphnia in such a way that handling and
searching times for different sized prey was the same. Larger prey represented greater energetic gain
per unit time and were more profitable, but they required better foraging skills to catch. The relative
capture proportion was noted for each individual and used as a determinant of competitive ability. Fish
where then allowed to forage in the absence of a competitor. Surprisingly, individuals that were more
competitive still successfully incorporated a higher proportion of large prey in their diet. Less
competitive animals had a similar attack rate on larger prey but were less successful at capturing them
and therefore had a higher proportion of small prey in their diet. Milinkski’s (1982) study therefore
neatly demonstrates the importance of considering individual variation in competitive ability in foraging
models.
125.

4.1.2 Food density and individual variation
Though both optimal foraging theory and IFD have their flaws, it is still reasonable to predict
that individuals forage selectively to maximise their energetic gains within certain constraints. Having
established that variation in behaviour between animals has important effects on foraging success
when animals interact, the ability to recognise and subsequently discriminate between variation in
environments could provide additional selective advantages to more competitive foragers or a means
of compensation for more passive foragers. For example, weaker competitors could avoid areas that
are more likely to attract strong competitors.
The amount of food available in a foraging patch is one factor that determines the benefit that
can be obtained for a given level of foraging cost. Animals that adjust their behaviour accordingly can
maximise their competitive ability and energetic gains. So how does food density affect a forager’s
behaviour? A study on the aquatic insect backswimmer Notonecta hoffmanni showed that foragers
adapted their responses to the spatial distribution of prey (Sih 1982). Juvenile N. hoffmanni spent
more time foraging in patches of higher food density and this increased time in better patches
translated into a higher feeding rate. It is interesting to note, however, that Sih (1982) also showed that
N. hoffmanni did not show “optimal” choice of foraging patches or time allocation and that higher
feeding rate did not translate into higher fitness. Also noteworthy was the fact that older starved
foragers were more efficient foragers than their younger or better-fed counterparts. Hunger has been
shown to be a driving force of foraging effort (Beukema 1968) and this highlights another important
interaction that should be taken into account; between life history (age and previous feeding) and
foraging behaviour. Studies have shown a wide range of responses to variation in resource
abundance in fish. Some studies show no effect of food density on foraging time or risk taking
behaviour (Cerri and Fraser 1983; Fraser and Huntingford 1986; Holbrook and Schmitt 1988). In
contrast, Gilliam and Fraser (1987) showed a positive correlation between risk taking and resource
abundance, while Abrahams and Dill (1989) reported a whole variety of responses in guppies
depending on their sex and the level of predation risk.
Variation in prey size is a different form of food density, as optimal allocation time to be spent
searching for and handling prey will change with the energy return from different sized food items
(Werner and Hall 1974). As mentioned above, in Milinski’s (1982) study on three spined sticklebacks,
larger prey provided a higher net energetic profit when searching and handling times were equal for all
size prey. Several species of generalist feeding fish have been shown to selectively choose food
based on their size (e.g., Ivlev 1961; Galbrait 1967; Brooks 1968; Hall et al. 1970; Munk 1992;
Forrester et al. 1994; Luo et al. 1996; Graeb et al. 2006; Hyvarinen and Huusko 2006). Food size is
126.

important to fish growth and may studies demonstrate a positive relationship between the two (Leeren
1958; Parker and Larkin 1959; Paloheim and Dickie 1966; Graeb et al. 2004; Harter and Heck 2006;
Persson and De Roos 2006). Another example of prey size choice can be found in smallmouth bass
Micropterus doloieuf (Stein 1977). Bass select the smallest size class of crayfish when these are
presented on sand, but intermediate sized crayfish on a pebble substrate. Stein (1977) explained that
this pattern occurred because smaller prey are better hidden on larger substrates and the associated
increase in search time meant a decrease in their value relative to more exposed intermediate sized
prey. This is one of the earliest studies to note that microhabitat variation can effect the distribution
and profitability of different prey.
4.1.3 Microhabitat complexity
The complexity of a microhabitat or substrate has been shown to influence both the density
and detection rate of prey (Eklov and Diehl 1994; Merilaita 2003) and the focus so far has been on
patch choice based on prior experience of food abundance. Is there evidence of a patch choice
preference based on potential food abundance? Prey attempt to minimise mortality by adjusting their
use of microhabitats and behaviour in the presence of a predator (Charnov 1976; Stein and Magnuson
1976), so there will often be a higher density of prey around structurally complex habitats. Can
predators discriminate between microhabitats and do they adjust their behaviour accordingly?
Individuals able to discriminate between substrates should select to forage in areas where prey
encounter rates are higher (Krebs et al. 1981).
The structural composition of foraging grounds affects foraging behaviour. Co-existing fish
species in reef communities have been shown to differ in their substrate preferences and foraging
behaviours (e.g., Clarke 1977; Robertson et al. 1979; Robertson and Lässig 1980; Anderson et al.
1981; Robertson 1984; Ormond and Edwards 1987; Krajewski et al. 2006) with the suggestion that
differences in habitat use between species are important in sustaining biodiversity in these
communities. Different microhabitat structure and substrate have certainly been shown to effect
foraging behaviour and growth rate of juvenile fish (Chan et al. 1997; Peake 1999; Nguyen and
Crocker 2006; Scharf et al. 2006).
Increased macroinvertebrate density is related to increased physical complexity at the
microhabitat level, such as increased abundance of vegetation cover (Rabe and Gibson 1984; Gregg
and Rose 1985; Lodge 1985; Chick and Mclvor 1994; Kurashov et al. 1996). Increased prey density
can also be found around more topographical complexity substrates (Rempel et al. 2000; Cole et al.
2003; Macia et al. 2003; Ellis and Bell 2004) as more structurally and physically complex habitats
127.

provide greater refuge spaces for prey (Kelly 1996; Scharf et al. 2006). Increased prey density around
complex microhabitat may, however, also be the result of a reduction in the predator’s ability to
capture prey by decreasing manoeuvrability or visibility (Werner et al. 1983; Babbitt and Jordan 1996;
Babbitt and Tanner 1998; Baber and Babbitt 2004). Studies in the laboratory have shown that complex
habitats contribute to lower predation vulnerability by interfering with predator search and pursuit
(Nelson and Bonsdorff 1990; Gotceitas and Brown 1993; Gotceitas et al. 1995; Fraser et al. 1996;
Lindholm et al. 1999; Bartholomew et al. 2000) and these findings are supported by studies in the field
(Tupper and Boutilier 1995; Beukers and Jones 1998; Heck et al. 2003). As such, prey encounter and
capture rate are usually negatively related to the increase in structural complexity of the microhabitat
(Savino and Stein 1982; Russo 1987; Diehl 1988; Tatrai and Herzig 1995; Mayer et al. 2001). The
degree to which habitat complexity mediates predation may depend on prey anti-predator responses,
the predator posing the threat, and the quality and quantity of the habitat structure (Diehl 1992; Eklov
1997).
4.1.4 Discriminating between microhabitats
Studies show that prey can discriminate between microhabitats and foraging substrates.
Cryptic colouration is an important anti-predatory trait for many aquatic preys (Endler 1978). In some
studies, stream insects prefer cryptic habitat patches (Clifford et al. 1989; Feltmate and Williams 1991;
Feltmate et al. 1992). Stream bottoms are composition of different rock types, overturned rocks and
fallen leaves with stream insects often being uniform dark in colour so that they are well camouflage
against dark backgrounds. Insects are able to distinguish between differently coloured substrate
patches and some species have been shown to chose darker substrates e.g., the lotic mayfly larvae
(Tikkanen et al. 2000), and tide pool sculpins (Houtman and Dill 1994). Macroinvertebrates are not the
only animals to show substrate discrimination as a mechanism of predator avoidance. Differential
habitat use to avoid predators is common (Fraser and Cerri 1982; Holomuzki 1986; Mittelbach 1986;
Ryer et al. 2004; Scharf et al. 2006). Even prey such as juvenile lemon sharks have been shown to
exhibit differential substrate preference, again probably for predator avoidance (Morrissey and Gruber
1993).
If prey chose habitat patches to maximise camouflage and avoid predators, given that more
complex foraging sites potentially have greater prey density, fish should preferentially forage at sites
that are more physically intricate to maximise their energy gain. So, do predators discriminate between
foraging grounds with difference microhabitat characteristics? If so, do predators choose to forage in
128.

areas with high prey density but lower capture rate? Or do they choose to forage in areas with fewer
potential prey but a higher probability of successful capture?
Fish represent a model system for studying foraging behaviour, particularly patch choice and
habitat experiments, because they are easily acclimatised to laboratory experiments, and their
habitats are straightforward to replicate. In a recent study, Webster and Hart (2004) showed that three
spined sticklebacks Gasterosteus aculeatus can not only discriminate between substrates of differing
structural complexity, but also show a strong preference for more complex substrates. Both
smallmouth bass Micropterus doloieuf and pumpkinseed sunfish Lepomis gibbosus have also been
shown to select different prey depending on the substrate on which they are presented and the levels
of topographical complexity (Stein 1977; Macchiusi and Baker 1991).
Webster and Hart (2004) presented three spined sticklebacks Gasterosteus aculeatus
simultaneously with food items on simple and complex substrates, repeating the experiment with
different densities of food items and used fish with differing hunger levels. The sticklebacks showed a
marked preference for complex microhabitats when food densities were equal between substrates,
and when density was greater on the complex substrate. Webster and Hart (2004) explained the result
as adaptive because increased substrate complexity is associated with higher macroinvertebrate
densities. The opposite preference could have been expected, however, as the increased handling
and searching times of smaller prey in more complex structures could have decreased the associated
value of that food item. The result could lie anywhere along a continuum from net energy being higher
due to increased food density (preference for complex substrate) to higher energy gain from prey that
is easier to see and catch (preference for simpler substrate). I wanted to conduct a similar study to see
if this result was repeatable, and where along the continuum Mogurnda mogurnda would lie.
129.

4.1.5 Aims of this study
Using Mogurnda mogurnda, I investigated whether fish can discriminate between substrates
of differing structural complexity when foraging. M mogurnda are a generalist predator of
macroinvertebrates, feeding on insects, crustaceans, worms, molluscs, fishes and plant materials.
They live in slow moving waters in rivers, creeks and billabongs, congregating around vegetation or
rocks where natural differences in habitat structure occur, which make them an excellent candidate for
this study.
I tested whether individuals preferred foraging grounds with more complex structures that
potentially provide refuge to more prey items. I investigated whether fish showed a preference for
foraging grounds of different structural complexity by manipulating actual prey densities in the two
substrates. Hunger is a driving force of foraging effort (Beukema 1968) and had significant effects on
foraging behaviour in the three spined stickleback (Webster and Hart 2004) so I also investigated
whether an individual’s hunger level affected foraging ground choice. Finally, since I manipulated the
amount and proportion of food available in each substrate I looked at which substrate fish consumed
the most food from (i.e. did they choose the substrate that led to the higher energy gain).
130.

4.2 Methods
As in previous chapters, fish were kept in aquaria at 26° Celsius in climate controlled rooms
on a 14:10 hr day: night illumination cycle. The holding tanks (6’ x 2’ x 2’) where fish were kept
between experiments were connected to sump filters filled with bioballs containing bacteria that break
down waste products in the water. Four holding tanks were used, two for fish before experiments and
two for after experiments. The sexes were housed separately in these holding tanks.
4.2.1 Binary Substrate Choice
I presented focal fish with the choice of a “simple” or “complex” substrate. The “simple”
substrate consisted of fine sand (< 3mm) in a tray on one side of the tank. This presented the fish with
a site that was homogeneous for structural complexity and coloration. The “complex” substrate
consisted of large gravel (longest axis range 5-10mm) and a scattering of large rocks (40mm > 80mm)
which created a site that was heterogeneous for structural complexity and coloration with many
interstitial spaces. The experimental tank had three marked zones similar to the experimental set up in
Chapters 2 and 3 with two “preference” zones (marked by the simple and complex substrate trays)
and a non-preference zone (the area in between the trays). The two trays (each measuring 25cm x 21
cm x 1.5cm) were placed on opposite ends of the tank (measuring 2’ x 1’ x 1’). The central “non
preference“ area was filled with gravel identical to the gravel used to cover the bottom of the holding
tanks (approximately 7mm) and was not treated as a foraging ground.
I had eight treatment groups each with twelve trials (total n = 96). I caught fish from the stock
tanks and allocated them randomly into one of eight holding tanks, trying to avoid individuals that were
at the extreme ends of the size scale. The eight fish were randomly assigned into two groups and then
to one of four prey density: (1) 0:0, complex: simple (no food on either substrate); (2) 1:3,
complex : simple (6 food items in the simple tray vs. 18 food items in the complex tray); (3) 3:1,
complex : simple (18 items in the simple tray vs. 6 items in the complex tray); and (4) 1:1,
complex : simple (6 items in the simple tray vs. 6 items in the complex tray). Hunger levels were also
manipulated within treatments. For each treatment, one group was fed to satiation 72 hours before the
trial and the other group was fed to satiation one hour before the trial.
The food used in the experiment was the protein pellets that were the fish’s sole diet so that
they were familiar with the food being offered. I set the trays in the tank, added the food at random
points on the tray then carefully filled the tank with water to begin the experiment. Each trial consisted
of placed one fish under a clear container in the centre of the non-preference zone between the two
131 .

trays for a five minutes acclimatisation period to ensure that it could see both substrates. After five
minutes, I carefully lifted the container covering the fish and started recording their position in the tank
for ten minutes. Using JWatcher™ (Blumstein et al. 2000), I recorded how often each fish entered a
zone, how long each fish was in each zone and how many feeding strikes each fish made in each
zone. After ten minutes, I caught the fish and housed them in a used stock tank. I then used a vacuum
gravel cleaner to remove at least half a tank of water, and if the treatment included food items I
removed any uneaten food particles. I then added food at random points which would sink to the
bottom for the next trial, and carefully refilled the tank.
I swapped the trays between the opposite ends of the tank after half of the trials (n = 6) within
each treatment. I tried to reduce the number of times I shifted the trays because it was difficult to do
without disturbing the substrate topography and I wanted to keep the presentation of the substrates as
similar as possible throughout the experiments.
4.2.2 Statistical Analyses - Foraging Analysis
In the analysis of substrate preference, I considered the following variables:
• The number of visits was the number of times a fish entered a substrate preference zone.
• The total duration was the total amount of time a fish spent in a substrate preference zone.
• The mean duration was the average duration per visit a fish spent in a substrate preference
zone.
• The duration proportion was the proportion of time a fish spent in a substrate preference
zone relative to the total duration of the observation period (10 minutes).
• The relative proportion was the proportion of time a fish spent in a substrate preference zone
relative to the time a female spent in both preference zones (excludes the time in the non
preference zone).
I used one-sample t-tests to quantify the effect of substrate on the response variables. If there
was no significant difference in the response variables between the substrates, a t-test would show
that the difference between the values for the two substrates (e.g., complex - simple substrate) did not
differ significantly from zero. I tested for a difference between substrates in the number of visits, total
duration of time spent on each substrate and the mean duration of each visit. In my analyses, I used
the difference in values between the substrates because this provides a simple way to analyse and
present the data. This method of analysis takes into account that the use of substrates is mutually
exclusive, and the aim of the experiment is to detect a difference in usage between the two substrates.
132.

I also used general linear models with sex, feeding treatment and hunger level as fixed factors
to test for their effect on the number of visits, total duration of time spent on each substrate and the
mean duration of each visit. For these models, I again used the difference between substrates for
each variable (e.g., difference in number of visits to the complex substrate minus the number of visits
to the simple substrate). I used the difference in values rather than including substrate as a fixed factor
in the models because this makes it easier to detect the main effect of the treatment rather than
having to test for interactions (e.g., a treatment*substrate interaction because the food density on
substrate types differs between treatments so the preferred substrate should vary between
treatments). I also tested for the effect of sex, feeding treatment and hunger level on the relative
proportion of time spent in one substrate preference zone compared to the total time spent in both
preference zones (i.e. total time excludes time spent in the non-preference zone). I used the
proportion of time on the complex substrate. Assumptions of normality were tested for, and satisfied,
in all the models.
I also repeated the models but used only data from treatments 2 (food ratio of 1:3 for
complex : simple) and treatment 3 (food ratio of 3:1 complex : simple). I repeated the model (with
feeding treatment and hunger level as fixed factors) because these two treatments showed the
greatest extremes in the difference in food ratio between the two substrates. It was therefore more
likely than I would detect a treatment effect than in the case where I included the two treatments with
equal food ratio per substrate (as these two treatments should not differ). I tested for sources of
variation in the number of visits, difference in total time spent on a substrate and the mean duration of
each visit as well as the relative proportion of time spent on each substrate. I did not include sex in the
models with the reduced data set because the analysis on the full data set indicated that there was no
effect of sex on substrate-based foraging decisions.
I also conducted separate one-sample t-tests on the relative proportion of time spent on the
complex substrate for each treatment and hunger level. Finally, I used Mann-Whitney U tests to look
for an effect of treatment and hunger level on the total number of feeding strikes on each substrate.
133.

4.3 Results
There was no evidence of a foraging preference for either the complex or the simple substrate
(Table 4.3.1). The t-tests showed there was no significant difference in the use of complex and simple
substrates. There was no difference in the number of visits, total time on each substrate or the mean
duration of each visit. There was also no effect of treatment or hunger level on the number of visits,
total time or the mean duration of visits to each substrate. There was, however, a significant difference
between the sexes for the mean duration of visits to complex versus simple substrates (Table 4.3.2,
Fig 4.3.1: F = 9.849, df = 1, p = 0.002). Females preferred the complex substrate (Fig. 4.3.2; t = -
2.989, df = 47, p = 0.004) while males showed no preference for either substrate (Fig. 4.3.3; t = 1.566,
df = 47, p = .124).
There was no significant difference between the sexes in the difference in the number of visits
or total time spent on each substrate, but looking at the difference in the mean total time spent on
each substrate among the treatments does suggest a treatment effect (Fig. 4.3.4). Small sample sizes
make it difficult to detect a statistically significance result so this could be worthy of future
investigation. There was no difference in the relative proportion of time spent on each substrate
between the sex, among treatments or between hunger levels.
Analysing only treatments 2 and 3, individuals still showed no foraging preference for either
the complex or the simple substrate (Table 4.3.3). There was no effect of treatment or hunger level on
the number of visits, total time or mean duration of visits to each substrate type. There was also no
effect of treatment or hunger level on the relative proportion of time spent on each substrate (Table
4.3.4).
There was, however, a difference in the number of feeding strikes between hunger levels
across treatments (Table 4.3.5). There were no feeding strikes for treatment 1 (as there was no food
present) but there were also no strikes for fish fed to satiation one hour before the trial (Fig. 4.3.5).
The treatment by hunger interaction effect was due to difference between treatment 1 and the other
treatments, as there were large numbers of feeding strikes in fish fed 72 hours before the trial in
treatments 2, 3 and 4 but not in treatment 1.
134.

Table 4.3.1 The difference between the two substrates for the listed response variables measured as
response variable for complex substrate minus response variable for simple complex (n = 96 for all
variables). The response variables are the number of visits, total time spent on each substrate and the
mean duration of each visit. Difference is measured as response variable for complex substrate minus
response variable for simple complex. A significant bias towards the use of a substrate occurs if the
mean difference is greater or less than zero (one-sample t-tests).
Response Variable Mean ± SD t df Sig.
Difference in Number of Visits 0.20 ± 2.25 0.863 95 0.390
Difference in Total Time Duration (sec) -19.19 ±206.25 -0.912 95 0.364
Difference in Mean Duration (sec) -5.11 ±604.00 -0.829 95 0.409
135.
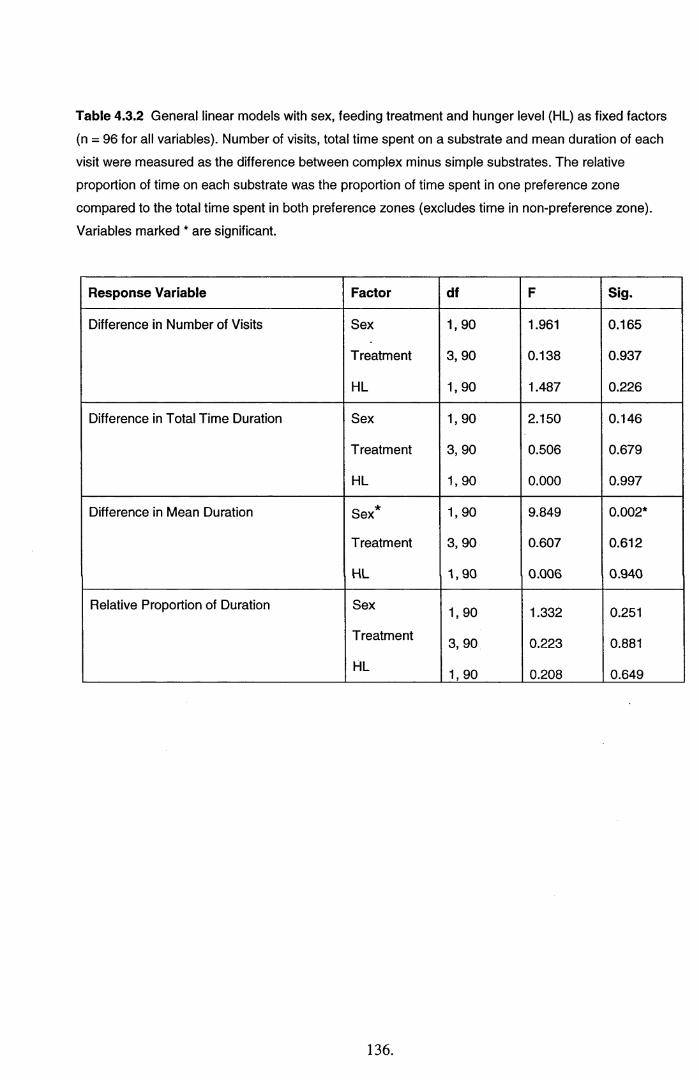
Table 4.3.2 General linear models with sex, feeding treatment and hunger level (HL) as fixed factors
(n = 96 for all variables). Number of visits, total time spent on a substrate and mean duration of each
visit were measured as the difference between complex minus simple substrates. The relative
proportion of time on each substrate was the proportion of time spent in one preference zone
compared to the total time spent in both preference zones (excludes time in non-preference zone).
Variables marked * are significant.
Response Variable Factor df F Sig.
Difference in Number of Visits Sex 1,90 1.961 0.165
Treatment 3, 90 0.138 0.937
HL 1,90 1.487 0.226
Difference in Total Time Duration Sex 1,90 2.150 0.146
T reatment 3, 90 0.506 0.679
HL 1,90 0.000 0.997
Difference in Mean Duration Sex* 1,90 9.849 0.002*
T reatment 3, 90 0.607 0.612
HL 1,90 0.006 0.940
Relative Proportion of Duration Sex 1,90 1.332 0.251
T reatment 3, 90 0.223 0.881
HL 1,90 0.208 0.649
136.

Table 4.3.3 General linear models with feeding treatment and hunger level (HL) as fixed factors only
for treatments 2 (food ratio of 1:3 complex : simple) and 3 (food ratio of 3:1 complex : simple). N = 96
for all variables). Number of visits, total time spent on a substrate and mean duration of each visit were
measured as the difference between complex minus simple substrates. The relative proportion of time
on each substrate was the proportion of time spent in one preference zone compared to the total time
spent in both preference zones (excludes time in non-preference zone).
Response Variable Factor df F Sig.
Difference in Number of Visits Treatment 1 0.060 0.808
HL 1 1.207 0.278
Difference in Total Time Duration T reatment 1 0.452 0.505
HL 1 0.004 0.951
Difference in Mean Duration T reatment 1 0.832 0.367
HL 1 0.151 0.699
Relative Proportion of Duration T reatment 1 0.148 0.702
HL 1 0.063 0.804
137.

Table 4.3.4 General linear models split by feeding treatment and hunger level (HL), n = 96 for all
variables). The relative proportion of time on each substrate was the proportion of time spent in one
preference zone compared to the total time spent in both preference zones (excludes time in non
preference zone). Feeding treatments were: (1) 0:0, complex : simple (no food on either substrate); (2)
1:3, complex : simple (6 food items in the simple tray vs. 18 food items in the complex tray; (3) 3:1
complex : simple (18 food items in the simple tray vs. 6 food items in the complex tray); (4) 1:1,
complex : simple (6 food items in the simple tray vs. 6 food items in the complex tray). Hunger level 1
was feeding fish to satiation 72 hours before the trial and hunger level 2 is feeding fish to satiation one
hour before the trial.
Response Variable T reatment HL t df Sig.
Relative Proportion of Duration 0:0 1 -0.219 11 0.830
2 -0.187 11 0.855
1:3 1 0.787 11 0.448
2 -0.829 11 0.425
3:1 1 -0.102 11 0.921
2 0.790 11 0.446
V.1 1 -0.187 11 0.855
2 -8.290 11 0.425
138.

Table 4.3.5 The comparison of the total number of feeding strikes during the 10 minute observation
period for hunger levels within each treatment (Mann-Whitney) (n = 96 for all variables). Feeding
treatments were: (1) 0:0, complex : simple (no food on either substrate); (2) 1:3, complex : simple (6
food items in the simple tray vs. 18 food items in the complex tray; (3) 3:1 complex : simple (18 food
items in the simple tray vs. 6 food items in the complex tray); (4) 1:1, complex : simple (6 food items in
the simple tray vs. 6 food items in the complex tray).Treatments marked * are significant. Hunger level
1 was feeding fish to satiation 72 hours before the trial and hunger level 2 is feeding fish to satiation
one hour before the trial.
Response Variable Treatment Mean ± SD U Z Sig.
Total Feeding Strikes 0:0, complex : simple
HL1 - 0.00 ± 0.00
HL2 - 0.00 ± 0.00 72.00 0.000 1.000
1:3, complex : simple:*
HL1 - 0.00 ± 0.00
HL2 - 2.75 ± 2.05 96.00 -3.599 0.000*
3:1, complex : simple*
HL1 -0 .0 0 ±0.00
HL2 - 4.58 ±2.31 78.00 -4.459 0.000*
1:1, complex: simple*
HL1 - 0.00 ± 0.00
HL2 - 2.92 ± 2.39 96.00 -3.591 0.000*
139.
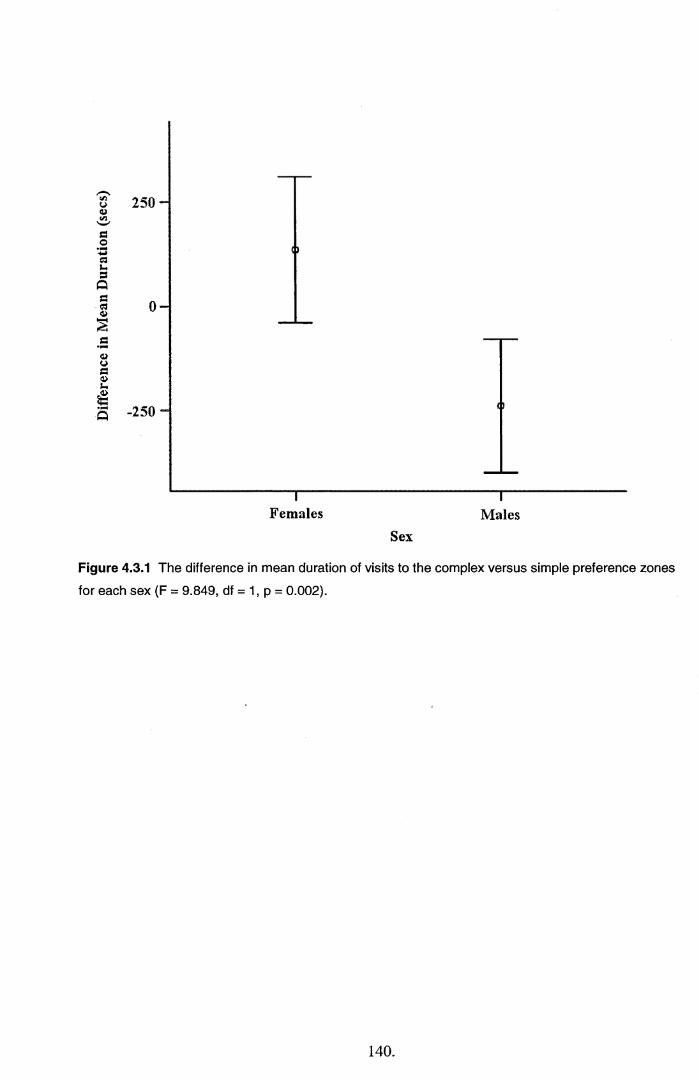
Diff
eren
ce i
250 -
ao
<rsu3Q
4>
a
0 “
o
-250 -o
jFemales
.I...Males
Sex
Figure 4.3.1 The difference in mean duration of visits to the complex versus simple preference zones
for each sex (F = 9.849, df = 1, p = 0.002).
140.
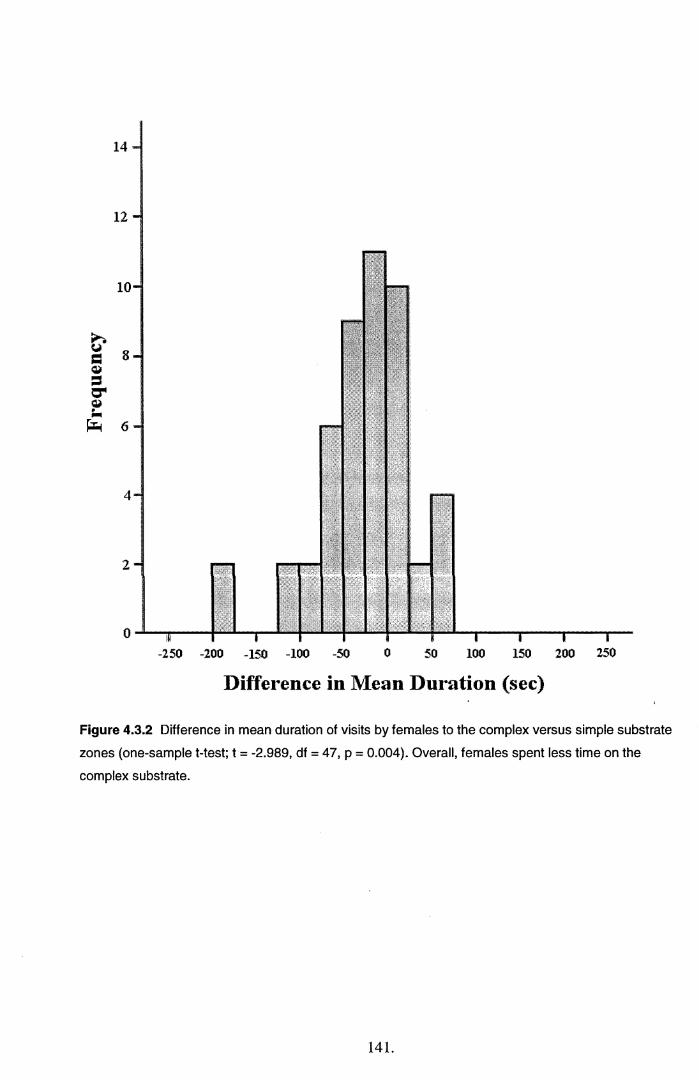
Freq
uenc
y
14 H
12 - j
-250 -200 -150 -100 -50 0 50 100 150 200 250
Difference in Mean Duration (sec)
Figure 4.3.2 Difference in mean duration of visits by females to the complex versus simple substrate
zones (one-sample t-test; t = -2.989, df = 47, p = 0.004). Overall, females spent less time on the
complex substrate.
141.
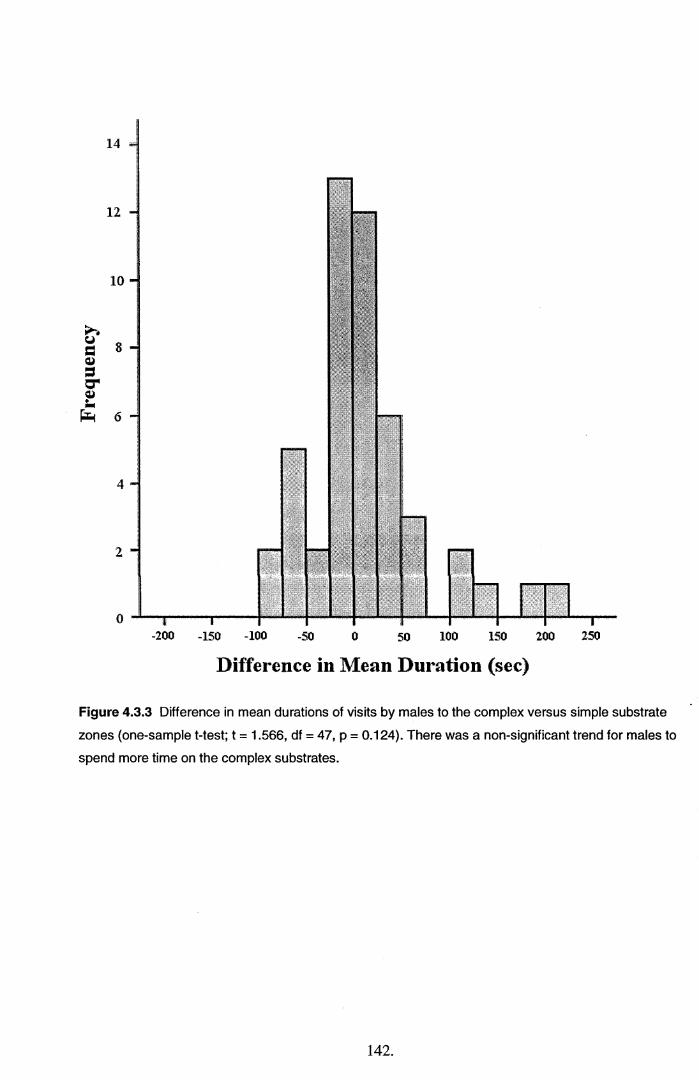
Freq
uenc
y
Difference in Mean Duration (sec)
Figure 4.3.3 Difference in mean durations of visits by males to the complex versus simple substrate
zones (one-sample t-test; t = 1.566, df = 47, p = 0.124). There was a non-significant trend for males to
spend more time on the complex substrates.
142.

-20 -
-40 -
-60 -
Feeding Treatment
Figure 4.3.4 The means for the difference in total time spent on the complex versus simple. Feeding
treatments are: 0:0, complex = simple (no food on either substrate); 1:1, complex = simple (6 food
items in the simple tray vs. 6 food items in the complex tray); (3) 1:3, complex = simple (18 food items
in the simple tray vs. 6 food items in the complex tray); (4) 3:1, complex = simple (6 food items in the
simple tray vs. 18 food items in the complex tray).
143.

5 * r __<b J t
♦ P N
o 4 Hasc3 1 _£
oH 1 —
1 -
0 - e © © ©
0:0 1:3 3:1Feeding Treatment
1:1
Figure 4.3.5 The mean ± SE for total number of feeding strikes per individual in the four feeding
treatment clustered by hunger level. Circles designate huger level 1 where fish were fed one hour
before the trial, and triangles designate hunger level 2 where fish that were only fed three days before
the trial. Feeding treatments were: (1) 0:0, complex : simple (no food on either substrate); (2) 1:3,
complex : simple (6 food items in the simple tray vs. 18 food items in the complex tray; (3) 3:1,
complex : simple (18 food items in the simple tray vs. 6 food items in the complex tray); (4) 1:1,
complex : simple (6 food items in the simple tray vs. 6 food items in the complex tray).
144.

4.4 Discussion
In the previous chapters, I discussed the costs of reproduction: the costs of mating (Daly
1978; Pomiankowski 1988; Iwasa et al. 1991), of mate choice (Hedrick and Dill 1993; Rowe 1994;
Grafe 1997; Watson et al. 1998) and of parental care (e.g., Stanley 2002). In light of this, the
processes whereby animals obtain energy for these activities becomes of interest, especially when
discussing traits that are condition-dependent (Zahavi 1975; Getty 1998). Both optimal foraging
models (reviewed in Pyke et al. 1977; and Schoener 1986) and ideal free distribution theory (Fretwell
and Lucas 1970) discuss individual foraging behaviour in terms of food density. Though these theories
are somewhat oversimplified, it is still reasonable to follow their predictions that individuals will forage
selectively to maximise their energetic gains.
Different habitat structures present animals with differences in food density and detection rate
of prey (Eklov and Diehl 1994; Merilaita 2003). There will often be a higher abundance of prey around
structurally complex habitats (Charnov 1976; Stein and Magnuson 1976) so predators might be
expected to prefer foraging in patches of greater structural complexity. For example, both smallmouth
bass Micropterus doloieuf and pumpkinseed sunfish Lepomis gibbosus select different prey depending
on the substrate on which they are presented and the differing levels of topographical complexity
(Stein 1977; Macchiusi and Baker 1991). Different microhabitat structure and substrate have been
shown to affect the foraging behaviour and growth rate of juvenile fish (Chan et al. 1997; Peake 1999;
Nguyen and Crocker 2006; Scharf et al. 2006).
There was no overall effect of habitat complexity (substrate) on foraging preference in
Mogurnda mogurnda. Females, however, showed a preference for simple substrates while males
showed none. There was no interaction effect of sex on treatment or hunger level, indicating this was
a general trend across the various experimental groups. It is unclear why females should display this
preference but males should not. (Abrahams and Dill 1989) reported a variety of responses in guppies
depending on their sex and the predation risk level, and other studies have shown a wide range of
responses to variation in resource abundance from no effect of food density (Cerri and Fraser 1983;
Fraser and Huntingford 1986; Holbrook and Schmitt 1988) to a positive correlation (Gilliam and Fraser
1987). It possible that females could have been at a stage in their reproductive cycle where they were
producing eggs and the higher energetic expenditure led them to a foraging strategy that maximised
the number of food items. Food items were easier to find in the simple habitat, thus lowering searching
and handling time and increased the number of successful food strikes. As the preference was not
detectable across all individuals, I consider this a weak preference trend but this results highlight the
145.

fact that individual variation should be taken into account in such studies; foraging behaviour can be
influenced by many factors such as predation risk, social interactions, reproductive state, and even
phylogenetic factors (reviewed in Perry and Pianka 1997).
There was a clear interaction between the effect of hunger level and food density on the
number of food strikes between substrates. There were no feeding strikes for treatment 1 (as there
was no food present) but there were also no strikes for fish fed to satiation one hour before the trial.
The difference lay between treatment 1 and the other treatments, as there were large numbers of
feeding strikes in fish that had not been fed for three days before the trial in treatments where food
was present. Hunger has been shown to be a driving force of foraging effort (Beukema 1968; Sih
1982). My result supports the fact that M. mogurnda showed no substrate preference rather than they
were simply non-responsive to the experiment and disinterested in feeding.
In a recent study, Webster and Hart (2004) showed that hunger-motivated three spined
sticklebacks Gasterosteus aculeatus prefer to forage in complex substrates. My results are not
equivalent to their findings, as hungry sticklebacks showed a marked preference for complex
microhabitats when food densities were equal between substrates, and when density was greater on
the complex substrate. Consistent with my results, however, satiated fish showed no preference, nor
was there a preference for complex substrates when food density was higher on simple substrates.
Webster and Hart (2004) explained their results as adaptive because increased substrate complexity
provides more refuges for prey, and is associated with higher macroinvertebrate densities (Kelly 1996;
Scharf et al. 2006). Increased prey density around complex microhabitat has, however, also been
explained by a reduction in predator’s ability to capture prey by decreasing manoeuvrability or the
visibility (Babbitt and Jordan 1996; Babbitt and Tanner 1997; Baber and Babbitt 2004). This has been
supported by studies in the laboratory (Nelson and Bonsdorff 1990; Gotceitas and Brown 1993;
Gotceitas et al. 1995; Fraser et al. 1996; Lindholm et al. 1999; Bartholomew et al. 2000) and in the
field (Tupper and Boutilier 1995; 1997; Beukers and Jones 1997; Heck et al. 2003). As such, prey
encounter and capture rates are usually negatively correlated to the increase in structural complexity
of the microhabitat (Savino and Stein 1982; Russo; 1987; Diehl 1988; Tatrai and Herzig; 1995 Mayer
et al. 2001).
A reverse preference or no effect of substrate could therefore also be predicted as the
increased handling and searching times of smaller prey in more complex structures could decrease
the associated value of a food item. Optimal foraging theory predicts that allocation time spent
searching for and handling prey will change the energy return for different sized food items (Werner
and Hall 1974) with larger prey presenting a higher net energetic profit when searching and handling
146.

times are equal for all sizes (Milinski 1982). Searching and handling times are rarely equal, however,
and are certainly not in structurally different microhabitats. In addition, prey employing cryptic
colouration have been shown to prefer darker substrates e.g., the lotic mayfly larvae (Tikkanen et al.
2000), and tide pool sculpins (Houtman and Dill 1994). If the diet of M. mogurnda includes cryptic
species of prey that choose habitat patches to maximise camouflage and avoid predators, the further
increase in searching and handling time may reduce a preference for complex substrates.
Habitat aside, several species of generalist feeding fish have been shown to selectively
choose food based on their size (e.g., Ivlev 1961; Galbrait 1967; Brooks 1968; Hall et al 1970; Werner
and Hall 1974; Munk 1992; Forrester et al. 1994; Luo et al. 1996; Hyvarinen and Huusko 2006).
Recent foraging models are more comprehensive, and awareness of the complexities of the demands
of an organism has increased. Empirical studies are developing to further explore foraging theory.
Ultimately, the degree to which habitat complexity mediates predation may depend on prey
antipredator responses, the predator posing the threat, and the quality and quantity of the habitat
structure (Diehl 1992, Eklöv 1997).
147.

References
(2004) Fishbase (www.fishbase.org). In: Froese R, Pauly D (eds)
Abrahams MV, Dill LM (1989) A DETERMINATION OF THE ENERGETIC EQUIVALENCE OF THE RISK OF PREDATION. Ecology 70:999-1007
Ah-King M, Kvarnemo C, Tullberg BS (2005) The influence of territoriality and mating system on the evolution of male care: a phylogenetic study on fish. Journal of Evolutionary Biology 18:371- 382
Ahnejsö I, Kvarnemo C, Merilaita S (2001) Using potential reproductive rates to predict mating competition among individuals qualified to mate. Behavioral Ecology 12:397-401
Alatalo RV, Carlson A, Lundberg A (1988) The search cost in mate choice of the Pied Flycatcher. Animal Behaviour 36:289-291
Allen GR, Midgley SH, Allen M (2002) Field Guide to the Freshwater Fishes of Australia. Western Australian Museum, Perth
Amundsen T (2003) Fishes as models in studies of sexual selection and parental care. Journal of Fish Biology 63:17-52
Anderson GRV, Ehrlich AH, Ehrlich PR, Roughgarden JD, Russell BC, Talbot FH (1981) The community structure of coral reef fishes. American Naturalist 117:476-495
Andersson M (1994) Sexual Selection. Princeton University Press, Princeton, New Jersey
Andersson S, Pryke SR, Ornborg J, Lawes MJ, Andersson M (2002) Multiple receivers, multiple ornaments, and a trade-off between agonistic and epigamic signaling in a widowbird. American Naturalist 160:683-691
Arnqvist G, Nilsson T (2000) The evolution of polyandry: multiple mating and female fitness in insects. Animal Behaviour 60:145-164
Arnqvist G, Rowe L (1995) Sexual Conflict and Arms Races between the Sexes - a Morphological Adaptation for Control of Mating in a Female Insect. Proceedings of the Royal Society of London Series B-Biological Sciences 261:123-127
Arnqvist G, Rowe L (2005) Sexual Conflict. Princeton University Press, Princeton, NJ
Arnqvist G, Rowe L, Krupa J J, Sih A (1996) Assortative mating by size: a meta-analysis of mating patterns in water striders. Evolutionary Ecology 10:265-284
Babbitt KJ, Jordan F (1996) Predation on Bufo terrestris tadpoles: Effects of cover and predator identity. Copeia:485-488
Babbitt KJ, Tanner GW (1998) Effects of cover and predator size on survival and development of Rana utricularia tadpoles. Oecologia 114:258-262
Baber MJ, Babbitt KJ (2004) Influence of habitat complexity on predator-prey interactions between the fish (Gambusia holbrooki) and tadpoles of Hyla squirella and Gastrophryne carolinensis. Copeia:173-177
148.

Bakker TCM (1993) Positive correlation between female preference and preferred male ornament in sticklebacks. Nature 363
Bakker TCM, Kunzler R, Mazzi D (1999) Condition-related mate choice in sticklebacks. Nature 401:234
Bakker TCM, Mundwiler B (1999) Pectoral fin size in a fish species with paternal care: a condition- dependent sexual trait revealing infection status. Freshwater Biology 41:543-551
Balshine-Earn S (1995) The costs of parental care in Galilee St Peter's fish, Sarotherodon galilaeus. Animal Behaviour 50:1-7
Barlow G, Francis RC, Baumgartner FV (1990) Do the colours of parents, companions and self influence assortative mating in the polychromatic Midas cichlid? Animal Behaviour 40:713-722
Bartholomew A, Diaz RJ, Cicchetti G (2000) New dimensionless indices of structural habitat complexity: predicted and actual effects on a predator's foraging success. Marine Ecology- Progress Series 206:45-58
Bateman AJ (1948) Intra-Sexual Selection In Drosophila. Heredity 2:349-368
Baylis JR (1981) The evolution of parental care in fishes, with reference to Darwins rule of male sexual selection. Environmental Biology of Fishes 6:223-251
Belles-lsles JC, Fitzgerald GJ (1991) Filial cannibalism in sticklebacks - a reproductive management strategy. Ethology Ecology & Evolution 3:49-62
Benkman CW (1987) Food profitability and the foraging ecology of crossbills. Ecological Monographs 57:251-267
Berejikian BA, Tezak EP, Larae AL (2000) Female mate choice and spawning behaviour of Chinook salmon under experimental conditions. Journal of Fish Biology 57:647-661
Berglund A, Bisazza A, Pilastro A (1996) Armaments and ornaments: An evolutionary explanation of traits of dual utility. Biological Journal of the Linnean Society 58:385-399
Beukema JJ (1968) Predation by 3-spined stickleback (gasterosteus aculeatus I) - influence of hunger and experience. Behaviour 31:1-&
Beukers JS, Jones GP (1998) Habitat complexity modifies the impact of piscivores on a coral reef fish population. Oecologia 114:50-59
Birkhead TR, Möller AP (1992) Sperm Competition in Birds. Academic Press, New York.
Bisazza A, Marconato A (1988) Female mate choice, male-male competition and parental care in the river bullhead, Cottus gobio L. (Pisces, Cottidae). Animal Behaviour 36:1352-1360
Blanchfield PJ, Ridgway MS (1997) Reproductive timing and use of redd sites by lake-spawning brook trout (Salvelinus fontinalis). Canadian Journal of Fisheries and Aquatic Sciences 54:747-756
Blanckenhorn WU (2002) The costs of copulating in the dung fly Sepsis cynipsea. Behavioral Ecology and Sociobiology 13
149.

Blanckenhorn WU, Kraushaar U, Reim C (2003) Sexual selection on morphological and physiological traits and fluctuating asymmetry in the yellow dung fly. Journal of Evolutionary Biology 16:903- 913
Blows MW (1999) Evolution of the genetic covariance between male and female components of mate recognition: an experimental test. Proceedings of the Royal Society B: Biological Sciences 266:2169-2174
Blumer LS (1979) Male parental care in the bony fishes. Quarterly Review of Biology 54:149-161
Blumer LS (1982) A bibliography and categorization of bony fish exhibiting parental care. Zoological Journal of the Linnean Society 76:1-22
Blumstein DT, Evans CS, Daniel JC (2000) JWatcher. In, 0.9 edn. Animal Behaviour Laboratory, Maquarie University, Sydney, pp A powerful tool for the quantitative analysis of behaviour.
Bonduriansky R, Rowe L (2005) Sexual selection, genetic architecture, and the condition dependence of body shape in the sexually dimorphic fly Prochyliza xanthostoma (Piophilidae). Evolution 59:138-151
Borgia G (1979) Sexual selection and the evolution of mating systems. In: Blum MS, Blum NA (eds) Sexual selection: testing the alternatives. Academic Press, New York, pp 19-80
Brooks JL (1968) Effects of prey size selection by lake planktivores. Systematic Zoology 17:273-&
Brown WD (1990) Size-assortative mating in the blister beetle Lytta magister (Coleoptera: Meloidae) is due to male and female preference for larger mates. Animal Behaviour 40:901-909
Brown WD (1993) The cause of size-assortative mating in the leaf beetle Trirhabda canadensis (Coleoptera: Chrysomelidae). Behavioral Ecology and Sociobiology 33:151-157
Buchanan KL, Catchpole CK (2000) Song as an indicator of male parental effort in the sedge warbler. Proceedings of the Royal Society of London Series B-Biological Sciences 267:321-326
Budaev SV, Zworykin DD, Mochek AD (1999) Individual differences in parental care and behaviour profile in the convict cichlid: a correlation study. Animal Behaviour 58:195-202
Bukowski TC, Linn CD, Christenson TE (2001) Copulation and sperm release in Gasteracantha cancriformis (Araneae : Araneidae): differential male behaviour based on female mating history. Animal Behaviour 62:887-895
Bulmer M (1989) Structural instability of models of sexual selection. Theoretical Population Biology 35:195-206
Burley N (1986) Sexual selection for aesthetic traits in species with biparental care. American Naturalist 127:415-445
Burley N (1988) The differential-allocation hypothesis: an experimental test. American Naturalist 132:611-628
Burley NT, Parker PG, Lundy K (1996) Sexual selection and extrapair fertilization in a socially monogamous passerine, the zebra finch (Taeniopygia guttata). Behavioral Ecology 7:218-226
Burt A (1995) Perspective: the evolution of fitness. Evolution 49:1-8
150.

Burt A (2000) Natural selection in the wild. Trends in Ecology & Evolution 15:306-307
Candolin U (1999a) Male-male competition facilitates female choice in sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 266:785-789
Candolin U (1999b) The relationship between signal quality and physical condition: is sexual signalling honest in the three spined stickleback? Animal Behaviour 58:1261-1267
Candolin U (2000) Changes in expression and honesty of sexual signalling over the reproductive lifetime of sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 267:2425-2430
Candolin U (2003) The use of multiple cues in mate choice. Biological Review 78:575-595
Caraco T, Martindale S, Whittam TS (1980) An empirical demonstration of risk-sensitive foraging preferences. Animal Behaviour 28:820-830
Carlisle TR (1982) Brood success in variable environments - Implications for parental care allocation. Animal Behaviour 30:824-836
Cerri RD, Fraser DF (1983) Predation and risk in foraging minnows - balancing conflicting demands. American Naturalist 121:552-561
Chan MD, Dibble ED, Kilgore KJ (1997) Laboratory examination of water velocity and substrate preference by age in Gulf Sturgeons. Transactions of the American Fisheries Society 126:330-333
Charnov EL (1976) Optimal Foraging, Marginal Value Theorem. Theoretical Population Biology 9:129- 136
Chellappa S, Huntingford SA (1989) Depletion of energetic reserves during reproductive aggression in male three spined stickleback, Gasterosteus aculeatus. Journal of Fish Biology 35:275-286
Chippindale AK, Gibson JR, Rice WR (2001) Negative genetic correlation for adult fitness between sexes reveals ontogenetic conflict in Drosophila. Proceedings of the National Academy of Sciences of the United States of America 98:1671-1675
Choe JC, Crespi BJ (1997) Mating systems in insects and arachnids. Cambridge University Press, Cambridge
Clark DL, Biesiadecki B (2002) Mating success and alternative reproductive strategies of the dimorphic jumping spider, Maevia inclemens (Aaneae, Salticidae). Journal of Arachnology 30:511-518
Clarke RD (1977) Habitat distribution and species diversity of chaetodontid and pomacentrid fish near Bimini, Bahamas. Marine Biology 40:277-289
Clement TS, Grens KE, Fernald RD (2005) Female affiliative preference depends on reproductive state in the African cichlid fish, Astatotilapia burtoni. Behavioral Ecology 16:83-88
Clifford HF, Gotceitas V, Casey RJ (1989) Roughness and color of artificial substratum particles as possible factors in colonization of stream invertebrates. Hydrobiologia 175:89-95
Clutton-Brock TH (1991) The Evolution of Parental Care. Princeton University Press, Princeton
151 .

Clutton-Brock TH, Parker GA (1992) Potential reproductive rates and the operation of sexual selection. Quarterly Review of Biology 67:437-456
Coleman RM, Fischer RU (1991) Brood size, male fanning effort and the energetics of a nonshareable parental investment in bluegill sunfish, Lepomis-macrochirus (Teleostei, Centrarchidae). Ethology 87:177-188
Coleman RM, Gross MR, Sargent RC (1985) Parental investment decision rules - a test in bluegill sunfish. Behavioral Ecology and Sociobiology 18:59-66
Collins MA, Bailey DM, Ruxton GD, Priede IG (2005) Trends in body size across an environmental gradient: A differential response in scavenging and non-scavenging demersal deep-sea fish. Proceedings of the Royal Society B-Biological Sciences 272:2051-2057
Cordero C, Eberhard WG (2003) Female choice of sexually antagonistic male adaptations: a critical review of some current research. Journal of Evolutionary Biology 16:1-6
Cote IM, Hunte W (1989) Male and female mate choice in the redlip blenny - why bigger is better. Animal Behaviour 38:78-88
Cotton S, Rogers DW, smallJ, Pomiankowski A, Fowler K (2006a) Variation in preference for a male ornament is positively associated with female eyespan in the stalk-eyed fly Diasemopsis meigenii. Proceedings of the Royal Society of London Series B-Biological Sciences 273:1287- 1292
Cotton S, smallJ, Pomiankowski A (2006b) Sexual Selection and Review Condition-Dependent Mate Preferences. Current Biology 16:755-765
Crudgington HS, Siva-Jothy MT (2000) Genital damage, kicking and early death - The battle of the sexes takes a sinister turn in the bean weevil. Nature 407:855-856
Cunningham EJA, Russell AF (2000) Egg investment is influenced by male attractiveness in the mallard. Nature 404:74-77
Curio E (1988) Relative realized life span and delayed cost of parental care. American Naturalist 131:825-836
Daly M (1978) Cost of mating. American Naturalist 112:771-774
Davis JC (1975) Exchange of oxygen at gills of fish in response to oxygen availability. Journal of the Electrochemical Society 122:C101-C101
Dawkins R, Carlisle TR (1976) Parental investment, mate desertion and a fallacy. Nature 262:131-133
de Lope F, Möller AP (1993) Female reproductive effort depends on the degree of ornamentation of their mates. Evolution 47:1152-1160
del Hoyo J, Elliott A, Sargatal J (1992-2003) Handbook of the Birds of the World. Lynx Edicions, Barcelona
Demartini EE (1987) Paternal defense, cannibalism and polygamy - factors influencing the reproductive success of painted greenling (pisces, hexagrammidae). Animal Behaviour 35:1145-1158
152.

DeWoody JA, Fletcher DE, Wilkins SD, Avise JC (2001) Genetic documentation of filial cannibalism in nature. Proceedings of the National Academy of Sciences of the United States of America 98:5090-5092
Diehl S (1988) Foraging efficiency of 3 fresh-water fishes - effects of structural complexity and light. Oikos 53:207-214
Diehl S (1992) Fish predation and benthic community structure - the role of omnivory and habitat complexity. Ecology 73:1646-1661
Downhower JF, Brown L (1980) Mate preferences of female mottled sculpins, Cottus bairdi. Animal Behaviour 28:728-734
Drent RH, Daan S (1980a) The prudent parent: Energetic adjustments in avian breeding. Ardea 68:225-252
Drent RH, Daan S (1980b) The prudent parent: Energetic adjustments in avian breeding. Ardea 68:225-252
Dudley RG, Eipper AW (1975) Survival of largemouth bass embryos at low dissolved oxygen concentrations. Transactions of the American Fisheries Society 104:122-128
Eberhard WG (1996) Female control: sexual selection by cryptic female choice. Princeton University Press, Princeton, New Jersey
Eberhard WG, Cordero C (1995) Sexual selection by cryptic female choice on male seminal products - a new bridge between sexual selection and reproductive physiology. Trends in Ecology & Evolution 10:493-496
Eggert AK, Reinking M, Muller JK (1998) Parental care improves offspring survival and growth in burying beetles. Animal Behaviour 55:97-107
Eklov P (1997) Effects of habitat complexity and prey abundance on the spatial and temporal distributions of perch (Perea fluviatilis) and pike (Esox lucius). Canadian Journal of Fisheries and Aquatic Sciences 54:1520-1531
Eklov P, Diehl S (1994) Piscivore efficiency and refuging prey - the importance of predator search mode. Oecologia 98:344-353
Eklöv P, Diehl S (1994) Piscivore efficiency and refuging prey - the importance of predator search mode. Oecologia 98:344-353
Emlen ST, Oring LW (1977) Ecology, sexual selection, and the evolution of mating systems. Science 197:215-223
Endler JA (1980) Natural selection on color in Poecilia reticulata. Evolution 34:76-91
Endler JA (1983) Natural and sexual selection on color patterns in Poeciliid fishes. Environmental Biology of Fishes 9:173-190
Endler JA (1988) Sexual selection and predation risk in Guppies. Nature 332:593-594
Endler JA, Basolo AL (1998) Sensory ecology, receiver biases and sexual selection. Trends in Ecology & Evolution 13:415-420
153.

Evans JP, Kelly JL, Bisazza A, Finazzo E, Pilastro A (2004) Sire attractiveness influences offspring performance in guppies. Proceedings of the Royal Society of London Series B-Biological Sciences 271:2035-2042
Fedorka KM, Mousseau TA (2004) Female mating bias results in conflicting sex-specific offspring fitness. Nature 429:65-67
Feltmate BW, Williams DD (1991) EVALUATION OF PREDATOR-INDUCED STRESS ON FIELD POPULATIONS OF STONEFLIES (PLECOPTERA). Ecology 72:1800-1806
Feltmate BW, Williams DD, Montgomerie A (1992) Relationship between diurnal activity patterns, cryptic coloration, and subsequent avoidance of predaceous fish by perlid stoneflies. Canadian Journal of Fisheries and Aquatic Sciences 49:2630-2634
Fetherston IA, Scott MP, Traniello JFA (1990) Parental Care in Burying Beetles - the Organization of Male and Female Brood-Care Behavior. Ethology 85:177-190
Fisher RA (1930) The Genetical Theory of Natural Selection. Clarendon Press, Oxford
Fisher RA (1958) The Genetical Theory of Natural Selection. Dover, New York
Fitzgerald GJ (1992a) Filial cannibalism in fishes - why do parents eat their offspring. Trends in Ecology & Evolution 7:7-10
FitzGerald GJ (1992b) Filial cannibalism in fishes: Why do parents eat their offspring? Trends in Ecology & Evolution 7:7-10
Fitzpatrick S, Berglund A, Rosenqvist G (1995) Ornaments or offspring: costs to reproductive success restrict sexual selection processes. Biological Journal of the Linnean Society 55:251-260
Forrester GE, Chace JG, McCarthy W (1994) Diel and density-related change in food consumption and prey selection by Brook Charr in a New Hampshire stream. Environmental Biology of Fishes 39:301-311
Forsgren E (1997) Female sand gobies prefer good fathers over dominant males. Proceedings of the Royal Society of London Series B-Biological Sciences 264:1283-1286
Forsgren E, Karlsson A, Kvarnemo C (1996) Female sand gobies gain direct benefits by choosing males with eggs in their nests. Behavioral Ecology and Sociobiology 39:91-96
Fraser DF, Cerri RD (1982) Experimental evaluation of predator-prey relationships in a patchy environment - consequences for habitat use patterns in minnows. Ecology 63:307-313
Fraser DF, Huntingford FA (1986) Feeding and avoiding predation hazard - the behavioral-response of the prey. Ethology 73:56-68
Fraser S, Gotceitas V, Brown JA (1996) Interactions between age-classes of Atlantic cod and their distribution among bottom substrates. Canadian Journal of Fisheries and Aquatic Sciences 53:305-314
Freeland WJ (1981) Parasitism and behavioral dominance among male-mice. Science 213:461-462
Fretwell S, Lucas HL (1970) On territorial behavior and other factors influencing habitat distribution in birds. Acta Biotheoretica 19:16-36
154.

Friberg U, Arnqvist G (2003) Fitness effects of female mate choice: preferred males are detrimental for Drosophila melanogaster females. Journal of Evolutionary Biology 16:797-811
Fuller RC (2003) Disentangling female mate choice and male competition in the rainbow darter, Etheostoma caeruleum. Copeia 1:138-148
Fuller RC, Houle D, Travis J (2005) Sensory bias as an explanation for the evolution of mate preferences. American Naturalist 166:437-446
Gabor C (1999) Association patterns of sailfin mollies (Poecilia latipinna): alternative hypotheses. Behavioral Ecology and Sociobiology 46:333-340
Galbrait MG (1967) Size-selective predation on daphnia by rainbow trout and yellow perch. Transactions of the American Fisheries Society 96:1-&
Garant D, Dodson JD, Bernatchez L (2005) Offspring genetic diversity increases fitness of female Atlantic salmon (Salmo salar). Behavioral Ecology and Sociobiology 57:240-244
Gavrilets S, Arnqvist G, Friberg U (2001) The evolution of female mate choice by sexual conflict. Proceedings of the Royal Society of London Series B-Biological Sciences 268:531-539
Gerhardt HC (1994) The evolution of vocalization in frogs and toads. Annual Review of Ecology and Systematics 25:293-324
Getty T (1998) Handicap signalling: when fecundity and viability do not add up. Animal Behaviour 56:127-130
Gibson RM, Bachman GC (1992) The costs of female choice in a lekking bird. Behavioral Ecology 3:300-309
Gilliam JF, Fraser DF (1987) Habitat selection under predation hazard - test of a model with foraging minnows. Ecology 68:1856-1862
Godin JGJ, Briggs SE (1996) Female mate choice under predation risk in the guppy. Animal Behaviour 51:117-130
Goodwin NB, Balshine-Earn S, Reynolds JD (1998) Evolutionary transitions in parental care in cichlid fish. Proceedings of the Royal Society B: Biological Sciences 165
Gotceitas V, Brown JA (1993) Substrate selection by juvenile atlantic cod (gadus-morhua) - effects of predation risk. Oecologia 93:31-37
Gotceitas V, Fraser S, Brown JA (1995) Habitat use by juvenile atlantic cod (gadus-morhua) in the presence of an actively foraging and non-foraging predator. Marine Biology 123:421-430
Goto A (1987) Polygyny in the river sculpin, Cottus hangiongensis (Pisces: Cottidae), with special reference to male mating success. Copeia 1:32-40
Goto A (1993) Male mating success and female mate choice in the river sculpin, Cottus-nozawae (Cottidae). Environmental Biology of Fishes 37:347-353
Goulet D (1998) Spawning success in the damselfish Amblyglyphidodon leucogaster: the influence of eggs in the nest. Animal Behaviour 55:651-664
155.

Gowaty PA, Steinichen R, Anderson WW (2002) Mutual interest between the sexes and reproductive success in Drosophila pseudoobscura. Evolution 56:2537-2540
Graeb BDS, Dettmers JM, Wahl DH, Caceres CE (2004) Fish size and prey availability affect growth, survival, prey selection, and foraging behavior of larval yellow perch. Transactions of the American Fisheries Society 133:504-514
Graeb BDS, Mangan MT, Jolley JC, Wahl DH, Dettmers JM (2006) Ontogenetic changes in prey preference and foraging ability of yellow perch: Insights based on relative energetic return of prey. Transactions of the American Fisheries Society 135:1493-1498
Grafe TU (1997) Costs and benefits of mate choice in the lek-breeding reed frog, Hyperolius marmoratus. Animal Behaviour S3:1103-1117
Grafen A (1990) Biological signals as handicaps. Journal of Theoretical Biology 144:517-546
Gray DA, Cade WH (1999) Quantitative genetics of sexual selection in the field cricket, Gryllus integer. Evolution 53:848-854
Green BS, McCormick Ml (2005) 02 replenishment to fish nests: males adjust brood care to ambient conditions and brood development. Behavioral Ecology and Sociobiology 16:389-397
Griffth SC, Owens IPF, Burke T (1998) Sexual selection in the house sparrow: the best males are the worst fathers. (In preparation.)
Gross MR, Sargent RC (1985) The evolution of male and female parental care in fishes. American Zoologist 25:807-822
Grubb TC, Greenwald L (1982) Sparrows and a brushpile - foraging responses to different combinations of predation risk and energy-cost. Animal Behaviour 30:637-640
Guderley H, Couture P (2005) Stickleback fights: why do winners win? Influence of metabolic and morphometric parameters. Physiological and Biochemical Zoology 78:173-181
Gwynne DT (1991) Sexual competition among females: what causes courtship-role reversal? . Trends in Ecology & Evolution 6:118-121
Hagelin JC (2002) The kinds of traits involved in male-male competition: a comparison of plumage, behavior, and body size in quail. Behavioral Ecology 13:32-41
Hale RE, St Mary CM, Lindström K (2003) Parental responses to changes in costs and benefits along an environmental gradient. Environmental Biology of Fishes 67:107-116
Hall DJ, Cooper WE, Werner EE (1970) An experimental approach to the production dynamics and structure of freshwater animal communities. Limnology and Oceanography 15:829-928
Hall DW, Kirkpatrick M, West B (2000) Runaway sexual selection when female preferences are directly selected. Evolution 54:1862-1869
Halupka K, Borowiec M (2006) Male whitethroats, Sylvia communis, advertise their future contribution to parental care. Behaviour 143:1-14
Hanssen SA, Erikstad KE, Johnsen V, Bustnes JO (2003) Differential investment and costs during avian incubation determined by individual quality: an experimental study of the common eider (Somateria mollissima). Proceedings of the Royal Society of London Series B-Biological Sciences 270:531-537
156.

Harari AR, Handler AM, Landolt PJ (1999) Size-assortative mating, male choice and female choice in the curculionid beetle Diaprepes abbreviatus. Animal Behaviour 58:1191-1200
Harshman LG, Prout T (1994) Sperm displacement without sperm transfer in Drosophila melanogasta. Evolution 48:758-766
Harter SL, Heck KL (2006) Growth rates of juvenile pinfish (Lagodon rhomboides): Effects of habitat and predation risk. Estuaries and Coasts 29:318-327
Hastings PA (1988) Female choice and male reproductive success in the angel blenny, Coralliozetus- angelica (Teleostei, Chaenopsidae). Animal Behaviour 36:115-124
Heck KL, Hays G, Orth RJ (2003) Critical evaluation of the nursery role hypothesis for seagrass meadows. Marine Ecology-Progress Series 253:123-136
Hedrick AV, Dill LM (1993) Mate choice by female crickets is influenced by predation risk. Animal Behaviour 46:193-196
Hegde SN, Krishna MS (1997) Size-assortative mating in Drosophila malerkotliana. Animal Behaviour 54:419-426
Heywood JS (1989) Sexual selection by the handicap mechanism. Evolution 43:1387-1397
Hoelzer G (1988) Filial cannibalism in a non-brood cycling marine fish. Environmental Biology of Fishes 21:309-313
Hoelzer GA (1989) The good parent process of sexual selection. Animal Behaviour 38:1067-1078
Hoelzer GA (1992) The ecology and evolution of partial clutch cannibalism by paternal Cortez Damselfish. Oikos 65:113-120
Hoelzer GA (1995) Filial cannibalism and male parental care in damselfishes. Bulletin of Marine Science 57:663-671
Hoffman Jl, Forcada J, Trathan PN, Amos W (2007) Female fur seals show active choice for males that are heterozygous and unrelated. Nature 445:912-914
Holbrook SJ, Schmitt.RJ (1988) Effects of predation risk on foraging behavior - mechanisms altering patch choice. Journal of Experimental Marine Biology and Ecology 121:151-163
Holland B, Rice WR (1998) Perspective: Chase-away sexual selection: Antagonistic seduction versus resistance. Evolution 52:1-7
Holland B, Rice WR (1999) Experimental removal of sexual selection reverses intersexual antagonistic coevolution and removes a reproductive load. Proceedings of the National Academy of Sciences of the United States of America 96:5083-5088
Holomuzki JR (1986) Predator avoidance and diel patterns of microhabitat use by larval tiger salamanders. Ecology 67:737-748
Houde AE (1994) Effect of artificial selection on male colour patterns on mating preference of female guppies. Proceedings of the Royal Society B: Biological Sciences 256:125-130
Houde AE, Torio AJ (1992) Effect of Parasitic Infection on Male Color Pattern and Female Choice in Guppies. Behavioral Ecology 3:346-351
157.

Houle D (1992) Comparing evolvability and variability of quantitative traits. Genetics 130:195-204
Houston AI, Szekely T, McNamara JM (2005) Conflict between parents over care. Trends in Ecology & Evolution 20:33-38
Houtman R, Dill LM (1994) The influence of substrate color on the alarm response of tidepool sculpins (Oligocottus-maculosus, Pisces, CottidaE). Ethology 96:147-154
Howard DJ, Reece M, Gregory PG, Chu J, Cain ML (1998) The evolution of barriers to fertilization between closely related organisms. In: Howard DJ, Berlocher SH (eds) Endless forms: species and speciation. Oxford University Press, New York, pp 279-288
Hunt J, Brooks R, Jennions MD (2005) Female mate choice as a condition-dependent life-history trait. American Naturalist 166:78-92
Hunt J, Brooks R, Jennions MD, Smith MJ, Bentsen CL, Bussiere LF (2004) High-quality male field crickets invest heavily in sexual display but die young. Nature 432:1024-1027
Hyvarinen P, Huusko A (2006) Diet of brown trout in relation to variation in abundance and size of pelagic fish prey. Journal of Fish Biology 68:87-98
Irschick DJ, Garland TJ (2001) Integrating function and ecology in studies of adaptation: investigations of locomotor capacity as a model system. Annual Review of Ecology and Systematics 32:367- 396
Itzkowitz M, Makie D (1986) Food provisioning and foraging behavior of the snowy egret, Egretta- thula. Biology of Behaviour 11:111-115
Ivlev VS (1961) Experimental ecology of the feeding of fishes. Yale University Press, New Haven, Connecticut
Iwasa Y, Pomiankowski A (1999) Good parent and good genes models of handicap evolution. Journal of Theoretical Biology 200:97-109
Iwasa Y, Pomiankowski A, Nee S (1991) The evolution of costly mate preferences. II. The ‘handicap’ principle. Evolution 45:1431-1442
Iyengar VK, Reeve HK, Eisner T (2002) Paternal inheritance of a female moth’s mating preference: consequences for sexual selection. Nature 419:830-832
Jennions MD, Backwell PRY, Passmore Nl (1995) Repeatability of mate choice: the effect of size in the African painted reed frog, Hyperolius marmoratus. Animal Behaviour 49:181-186
Jennions MD, Petrie M (1997) Variation in mate choice and mating preferences : a review of causes and consequences. Biological Review 72:283-327
Jennions MD, Polakow DA (2001) The effect of partial brood loss on male desertion in a cichlid fish: an experimental test. Behavioral Ecology 12:84-92
Jivoff P (1997) The relative roles of predation and sperm competition on the duration of the post- copulatory association between the sexes in the blue crab, Callinectes sapidus. Behavioral Ecology and Sociobiology 40:175-185
Johnstone RA, Reynolds JD, Deutsch JC (1996) Mutual mate choice and sex differences in choosiness. Evolution 50:1382-1391
158.

Jones JC, Reynolds JD (1999) Costs of egg ventilation for male common gobies breeding in conditions of low dissolved oxygen. Animal Behaviour 57:181-188
Jones TM, Quinnell RJ (2002) Testing predictions for the evolution of lekking in the sandfly, Lutzomyia longipalpis. Animal Behaviour 63:605-612
Kamler E (1992) Early life histories of fish: an energetics approach. Chapman & Hall, London
Karino K (1995) Male-male competition and female mate choice through courtship display in the territorial damselfish Stegastes nigricans. Ethology 100:126-138
Karino K (1997) Influence of brood size and offspring size on parental investment in a biparental cichlid fish, Neolamprologus moorii. Journal of Ethology 15:39-43
Karino K, Arai R (2006) Effect of clutch size on male egg-fanning behavior and hatching success in the goby, Eviota prasina (Klunzinger). Journal of Experimental Marine Biology and Ecology 334:43-50
Keenleyside MHA, Rangeley RW, Küppers BU (1985) Female mate choice and male parental defense behaviour in the cichlid fish Cichlasoma nigrofasciatum. Canadian Journal of Zoology 63:2489-2493
Keller L, Reeve HK (1995) Why do females mate with multiple males - the sexually selected sperm hypothesis. Advances in the Study of Behavior 24:291-315
Kelly JF (1996) Effects of substrate on prey use by belted kingfishers (Ceryle alcyon): A test of the prey abundance - Availability assumption. Canadian Journal of Zoology-Revue Canadienne De Zoologie 74:693-697
Kinne O, Kinne EM (1962) Rates of development in embryos of a cyprinodont fish exposed to different temperature salinity-oxygen combinations. Canadian Journal of Zoology 40:231-&
Kirkpatrick M (1985) Evolution of female choice and male parental investment in polygynous species: the demise of the “sexy son”. American Naturalist 125:788-810
Kirkpatrick M (1987) The evolutionary forces acting on female mating preferences in polygynous animals. In: Bradbury JW, Andersson MB (eds) Sexual selection: testing the alternatives. Wiley, Chichester, UK, pp 67-82
Kirkpatrick M (1996) Good genes and direct selection in evolution of mating preferences. Evolution 50:2125-2140
Klug H, Chin A, St Mary CM (2005) The net effects of guarding on egg survivorship in the flagfish, Jordanella floridae. Animal Behaviour 69:661-668
Knapp RA, Kovach JT (1991) Courtship as an honest indicator of male parental quality in the bicolor damselfish, Stegastes partitus. Behavioral Ecology 2:295-300
Knapp RA, Warner RR (1991) Male parental care and female choice in the bicolor damselfish, Stegastes partitus: bigger is not always better. Animal Behaviour 41:747-756
Kodric-Brown A, Nicoletto P (2005) Courtship behaviour, swimming performance, and microhabitat use of Trinidadian guppies. Environmental Biology of Fishes 73:299-307
Kokko H (1998a) Good genes, old age and life-history trade-offs. Evolutionary Ecology 12:739-750
159.

Kokko H (1998b) Should advertising parental care be honest? Proceedings of the Royal Society of London Series B-Biological Sciences 265:1871-1878
Kokko H (2001) Fisherian and "good genes" benefits of mate choice: how (not) to distinguish between them. Ecology Letters 4:322-326
Kokko H, Brooks R, Jennions MD, Morley J (2003) The evolution of mate choice and mating biases. Proceedings of the Royal Society of London Series B-Biological Sciences 270:653-664
Kokko H, Jennions M (2003) It takes two to tango. Trends in Ecology & Evolution 18:103-104
Kokko H, Jennions MD, Brooks R (2006) Unifying and testing models of sexual selection. Annual Review of Ecology Evolution and Systematics 37:43-66
Kokko H, Monaghan P (2001) Predicting the direction of sexual selection. Ecology Letters 4:159-165
Kolm K (2004) Female courtship in the Banggai cardinalfish: honest signals of egg maturity and reproductive output? Behavioral Ecology and Sociobiology 56:59-64
Kolm N (2001) Females produce larger eggs for large males in a paternal mouthbrooding fish. Proceedings of the Royal Society of London Series B-Biological Sciences 268:2229-2234
Koref-Santibanez S (2001) Effects of age and experience on mating activity in the sibling species Drosophila pavani and Drosophila gaucha. Behavior Genetics 31:287-297
Kotiaho J, Alatalo RV, Mappes J, Parri S (1997) Fighting success in relation to body mass and drumming activity in the male wolf spider Hygrolycosa rubrofasciata. Canadian Journal of Zoology-Revue Canadienne De Zoologie 75:1532-1535
Kotiaho JS (2001) Costs of sexual traits: a mismatch between theoretical considerations and empirical evidence. Biological Reviews 76:365-376
Kraak SBM (1996) Female preference and filial cannibalism in Aidablennius sphynx (Teleostei, Blenniidae); A combined field and laboratory study. Behavioural Processes 36:85-97
Kraak SBM, Groothuis TGG (1994) Female preference for nests with eggs is based on the presence of the eggs themselves. Behaviour 131:189-206
Kraak SBM, Videler JJ (1991) Mate choice in aidablennius-sphynx (teleostei, blenniidae) - females prefer nests containing more eggs. Behaviour 119:243-266
Krajewski JP, Bonaldo RM, Sazima C, Sazima I (2006) Foraging activity and behaviour of two goatfish species (Perciformes: Mullidae) at Fernando de Noronha Archipelago, tropical West Atlantic. Environmental Biology of Fishes 77:1-8
Kramer DL (1987) Dissolved oxygen and fish behavior. Environmental Biology of Fishes 18:81-92
Krebs JR (1980) Optimal foraging, predation risk and territory defense. Ardea 68:83-90
Krebs JR, Houston JA, Charnov EL (1981) Some recent developments in optimal forgaing. In: Kamil AC, Sargent TD (eds) Foraging Behaviour: Ecological, Ethological and Physiological Approaches. Garland STPM Press, New York, pp 3-17
Kunzler R, Bakker TCM (2000) Pectoral fins and paternal quality in sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 267:999-1004
160.

Kuwamura T, Yugo Y, Nakakshima Y (1993) Size-assortative monogamy and paternal egg care in a coral goby Paragobioon echinocephalus. Ethology 95:65-75
Kvarnemo C (1997) Food affects the potential reproductive rates of sand goby females, but not of males. Behavioral Ecology 8:605-611
Kvarnemo C, Ahnejsö I (1996) The dynamics of operational sex ratios and competition for mates. Trends in Ecology & Evolution 11:404-408
Kvarnemo C, Svensson O, Forsgren E (1998) Parental behaviour in relation to food availability in the common goby. Animal Behaviour 56:1285-1290
Lailvaux SP, Irschick DJ (2006) No evidence for female association with high-performance males in the green anole lizard, Anolis carolinensis. Ethology 112:707-715
Lande R (1980) Sexual dimorphism, sexual selection, and adaptation in polygenic characters. Evolution 34
Langmore NE, Davies NB, Hatchwell BJ, Hartley IR (1996) Female song attracts males in the alpine accentor Prunella collaris. Proceedings of the Royal Society of London Series B-Biological Sciences 263:141-146
Lavery RJ, Keenleyside MHA (1990) Parental investment of a biparental cichlid fish, cichlasoma- nigrofasciatum, in relation to brood size and past investment. Animal Behaviour 40:1128-1137
Leboeuf BJ, Mesnick S (1991) Sexual behavior of male Northern Elephant Seals 1. Lethal injuries to adult females. Behaviour 116:143-162
Leeren ED (1958) Observations on the growth of perch (perca-fluviatilis i) over 22 years with special reference to the effects of temperature and changes in population-density. Journal of Animal Ecology 27:287-334
Lessells CM (1999) Sexual conflicts in animals. In: Keller L (ed) Levels of Selection. Princeton University Press, Princeton, New Jersey, pp 75-99
Lifjeld JT, Dunn PO, Westneat DF (1994) Sexual selection by sperm competition in birds: male-male competition or female choice? Journal of Avian Biology 25:244-50
Lindholm JB, Auster PJ, Kaufman LS (1999) Habitat-mediated survivorship of juvenile (0-year) Atlantic cod Gadus morhua. Marine Ecology-Progress Series 180:247-255
Lindström K (1988) Male-male competition for nest sites in the sand goby, pomatoschistus-minutus. Oikos 53:67-73
Lindström K (1998) Effects of costs and benefits of brood care on filial cannibalism in the sand goby. Behavioral Ecology and Sociobiology 42:101-106
Lindstrom K, Sargent RC (1997) Food access, brood size and filial cannibalism in the fantail darter, Etheostoma flabellare. Behavioral Ecology and Sociobiology 40:107-110
Lindström K, Sargent RC (1997) Food access, brood size and filial cannibalism in the fantail darter, Etheostoma flabellare. Behavioral Ecology and Sociobiology 40:107-110
Linville SU, Breitwisch R, Schilling AJ (1998) Plumage brightness as an indicator of parental care in northern cardinals. Animal Behaviour 55:119-127
161.

Lissaker M, Kvarnemo C (2006) Ventilation or nest defense - parental care trade-offs in a fish with male care. Behavioral Ecology and Sociobiology 60:864-873
Lissaker M, Kvarnemo C, Svensson O (2003) Effects of a low oxygen environment on parental effort and filial cannibalism in the male sand goby, Pomatoschistus minutus. Behavioral Ecology 14:374-381
Lopez P, Munoz A, Martin J (2002) Symmetry, male dominance and female mate preferences in the Iberian rock lizard, Lacerta monticola. Behavioral Ecology and Sociobiology 52:342-347
Lopez S (1999) Parasitized female guppies do not prefer showy males. Animal Behaviour 57:1129- 1134
Luo JG, Brandt SB, Klebasko MJ (1996) Virtual reality of planktivores: A fish's perspective of prey size selection. Marine Ecology - Progresses Series 140:271-283
Macchiusi F, Baker RL (1991) Prey behavior and size-selective predation by fish. Freshwater Biology 25:533-538
Mackereth RW, Noakes DLG, Ridgway MS (1999) Size-based variation in somatic energy reserves and parental expenditure by male smallmouth bass, Micropterus dolomieu. Environmental Biology of Fishes 56:263-275
Maekawa K, Iguchi K, Katano O (1996) Reproductive success in male Japanese minnows, Pseudorasbora parva: Observations under experimental conditions. Ichthyological Research43:257-266
Magnhagen C (1991) Predation risk as a cost of reproduction. Trends in Ecology & Evolution 6:183- 185
Manica A (2002) Filial cannibalism in teleost fish. Biological Reviews 77:261-277
Manica A (2003) The effect of brood size and age on partial filial cannibalism in the scissortail sergeant. Journal of Fish Biology 63:37-47
Manica A (2004) Parental fish change their cannibalistic behaviour in response to the cost-to-benefit ratio of parental care. Animal Behaviour 67:1015-1021
Mank JE, Promislow DEL, Avise JC (2005) Phylogenetic perspectives in the evolution of parental care in ray-finned fishes. Evolution 59:1570-1578
Marconato A, Bisazza A (1986) Males whose nests contain eggs are preferred by female Cottus-gobio L (Pisces, Cottidae). Animal Behaviour 34:1580-1582
Marconato A, Bisazza A, Fabris M (1993) The cost of parental care and egg cannibalism in the river bullhead, Cottus-gobio L (Pisces, Cottidae). Behavioral Ecology and Sociobiology 32:229-237
Markman S, Yomtov Y, Wright J (1995) Male parental care in the orange-tufted sunbird - behavioral- adjustments in provisioning and nest guarding effort. Animal Behaviour 50:655-669
Marques PAM (2003) Male mate desertion in the Spanish sparrow Passer hispaniolensis. Ardea 91:245-250
Mayer CM, Rudstam LG, Mills EL, Cardiff SG, Bloom CA (2001) Zebra mussels (Dreissena polymorpha), habitat alteration, and yellow perch (Perea flavescens) foraging: system-wide
162.

effects and behavioural mechanisms. Canadian Journal of Fisheries and Aquatic Sciences 58:2459-2467
Mazzi D (2004) Parasites make male pipefish careless. Journal of Evolutionary Biology 17:519-527
McLister JD (2003) The metabolic cost of amplexus in the gray tree frog (Hyla versicolor): assessing the energetics of male mating success. Canadian Journal of Zoology 81:388-394
McNamara JM, Szekely T, Webb JN, Houston AI (2000) A dynamic game-theoretic model of parental care. Journal of Theoretical Biology 205:605-623
Mehla J, Sheldon BC (2000) Lifetime reproductive success and heritability in nature. American Naturalist 155:301-310
Merilaita S (2003) Visual background complexity facilitates the evolution of camouflage. Evolution 57:1248-1254
Mihelakakis A, Kitajima C (1994) Effects of salinity and temperature on incubation period, hatching rate, and morphogenesis of the silver sea bream, Sparus sarba (Forskal, 1775). Aquaculture 126:361-371
Milinski M (1979) Evolutionarily stable feeding strategy in sticklebacks. Zeitschrift Fur Tierpsychologie- Journal of Comparative Ethology 51:36-40
Milinski M (1982) Optimal foraging - the influence of intraspecific competition on diet selection. Behavioral Ecology and Sociobiology 11:109-115
Milinski M, Heller R (1978) Influence Of A Predator On Optimal Foraging Behavior Of Sticklebacks (Gasterosteus-Aculeatus L). Nature 275:642-644
Mitchell DP, Dunn PO, Whittingham LA, Freeman-Gallant CR (2007) Attractive males provide less parental care in two populations of the common yellowthroat. Animal Behaviour 73:165-170
Mittelbach G (1986) Predator-mediated habitat use - some consequences for species interactions. Environmental Biology of Fishes 16:159-169
Mittelbach GG (1981) Patterns of invertebrate size and abundance in aquatic habitats. Canadian Journal of Zoology 38:896-904
Möller AP (1993) Female preference for apparently symmetrical male sexual ornaments in the barn swallow Hirundo-rustica. Behavioral Ecology and Sociobiology 32:371-376
Möller AP, Alatalo RV (1999) Good-genes effect in sexual selection. Proceedings of the Royal Society of London Series B-Biological Sciences 266:85-91
Möller AP, Thornhill R (1998) Male parental care, differential parental investment by females and sexual selection. Animal Behaviour 55:1507-1515
Montgomerie RD, Weatherhead PJ (1988) Risks and rewards of nest defense by parent birds. Quarterly Review of Biology 63:167-187
Moore AJ, Gowaty PA, Wallin WG, Moore PJ (2001) Sexual conflict and the evolution of female mate choice and male social dominance. Proceedings of the Royal Society of London Series B- Biological Sciences 268:517-523
163.

Moore AJ, Moore PJ (1999) Balancing sexual selection through opposing mate choice and male competition. Proceedings of the Royal Society of London Series B-Biological Sciences 266:711-716
Morrison C, Hero JM, Smith WP (2001) Mate selection in Litoria chloris and Litoria xanthomera: Females prefer smaller males. Austral Ecology 26:223-232
Morrissey JF, Gruber SH (1993) Habitat selection by juvenile lemon sharks, Negaprion brevirostris. Environmental Biology of Fishes 38:311 -319
Mountjoy DJ, Lemon RE (1997) Male song complexity and parental care in the European starling. Behaviour 134:661-675
Munk P (1992) Foraging Behaviour and prey size spectra of larval herring Clupea harengus. Marine Ecology - Progresses Series 80:149-158
Neat FC, Taylor AC, Huntingford FA (1998) Proximate costs of fighting in male cichlid fish: the role of injuries and energy metabolism. Animal Behaviour 55:875-882
Neff BD (2003) Decisions about parental care in response to perceived paternity. Nature 422:716
Neff BD, Gross MR (2001) Dynamic adjustment of parental care in response to perceived paternity. Proceedings of the Royal Society of London Series B-Biological Sciences 268:1559-1565
Nelson CM, Nolen TG (1997) Courtship song, male agonistic encounters, and female mate choice in the house cricket, Acheta domesticus (Orthoptera: Gryllidae). Journal of Insect Behavior 10:557-570
Nelson WG, Bonsdorff E (1990) Fish predation and habitat complexity - are complexity thresholds real. Journal of Experimental Marine Biology and Ecology 141:183-194
Nguyen RM, Crocker CE (2006) The effects of substrate composition on foraging behavior and growth rate of larval green sturgeon, Acipenser medirostris. Environmental Biology of Fishes 76:129- 138
Nicoletto PF (1995) Offspring quality and female choice in the guppy, Poecillia reticulata. Animal Behaviour 49:377-387
Nilsson SO, Nilsson GE (2000) Free choice by female sticklebacks: lack of preference for male dominance traits. Canadian Journal of Zoology-Revue Canadienne De Zoologie 78:1251-1258
Noonan K (1983) Female mate choice in a cichlid fish Cichlasoma nigrofasciatum. Animal Behaviour 31:1005-1010
Nyman KJ (1953) Observations on the behaviour of Gobius microps. Acta Societatis po Fauna et Flora Fennica 69:1-11
Okuda N (1999) Sex roles are not always reversed when the potential reproductive rates is higher in females. American Naturalist 153:540-548
Okuda N, Ito S, Iwao H (2002) Female spawning strategy in Rhinogobius sp OR: how do females deposit their eggs in the nest? Ichthyological Research 49:371-379
Okuda N, Yanagisawa Y (1996) Filial cannibalism by mouthbrooding males of the cardinal fish, Apogon doederleini, in relation to their physical condition. Environmental Biology of Fishes 45:397-404
164.

Oppenheimer JR (1970) Mouthbrooding in fishes. Anima! Behaviour 52:307-314
Ormond RFG, Edwards AJ (1987) Red Sea fishes. In: Edwards AJ, Head SM (eds) Red Sea (key environments). Pergammon Press, Oxford, pp 22-44
Osenberg CW, Mittelbach GG (1989) Effects of body size on the predator prey interaction between pumpkinseed sunfish and gastropods. Ecological Monographs 59:405-432
Osorno JL, Szekely T (2004) Sexual conflict and parental care in magnificent frigatebirds: full compensation by deserted females. Animal Behaviour 68:337-342
Östlund-Nilsson S, Nilsson GE (2004) Breathing with a mouth full of eggs: respiratory consequences of mouthbrooding in cardinalfish. Proceedings of the Royal Society of London Series B- Biological Sciences 271:1015-1022
Östlund S, Ahnesjö I (1998) Female fifteen-spined sticklebacks prefer better fathers. Animal Behaviour 56:1177-1183
Paloheim JE, Dickie LM (1966) Food and growth of fishes .3. relations among food body size and growth efficiency. Journal of the Fisheries Research Board of Canada 23:1209-&
Pampoulie C, Lindstrom K, St Mary CM (2004) Have your cake and eat it too: male sand gobies show more parental care in the presence of female partners. Behavioral Ecology 15:199-204
Parker GA (1974) Assessment strategy and the evolution of fighting behaviour. Journal of Theoretical Biology 47:223-243
Parker GA, Simmons LW (1996) Parental investment and the control of sexual selection: predicting the direction of sexual competition. Proceedings of the Royal Society of London Series B- Biological Sciences 263:315-321
Parker RR, Larkin PA (1959) A concept of growth in fishes. Journal of the Fisheries Research Board of Canada 16:721-745
Parker TH, Garant D (2004) Quantitative genetics of sexually dimorphic traits and capture of genetic variance by a sexually-selected condition-dependent ornament in red junglefowl (Gallus gallus). Journal of Evolutionary Biology 17:1277-1285
Parker TH, Ligon JD (2003) Female mating preferences in red junglefowl: a meta-analysis. Ethology Ecology & Evolution 15:63-72
Parri S, Alatalo RV, Mappes J (1998) Do female leaf beetles Galerucella nymphaeae choose their mates and does it matter? Oecologia 114:127-132
Partridge L, Fowler K (1990) Non-mating costs of exposure to males in female Drosophila melanogaster Journal of Insect Physiology 36:419-425
Payne AG, Smith C, Campbell AC (2002) Filial cannibalism improves survival and development of beaugregory damselfish embryos. Proceedings of the Royal Society of London Series B- Biological Sciences 269:2095-2102
Payne AG, Smith C, Campbell AC (2004) A model of oxygen-mediated filial cannibalism in fishes. Ecological Modelling 174:253-266
Peake S (1999) Substrate preferences of juvenile hatcheryreared Lake sturgeon, Acipenser fulvescens. Environmental Biology of Fishes 56:367-374
165.

Perry G, Levering K, Girard I, Garland TJ (2004) Locomotor performance and dominance in male Anolis cristatellus. Animal Behaviour 67:37-47
Perry G, Pianka ER (1997) Animal foraging: past, present and future. Trends in Ecology & Evolution 12:360-364
Persson L, De Roos AM (2006) Food-dependent individual growth and population dynamics in fishes. Journal of Fish Biology 69:1-20
Peters A, Michiels NK (1996) Evidence for lack of inbreeding avoidance by selective simultaneous hermaphrodite. Invertebrate Biology 115:99-103
Petersen CW (1990) The occurrence and dynamics of clutch loss and filial cannibalism in two Caribbean damselfishes. Journal of Experimental Marine Biology and Ecology 135:117-133
Petrie M (1983) Female moorhens compete for smallfat males. Science 220:413-415
Pianka ER (1976) Natural selection of optimal reproductive tactics. American Zoologist 16:775-784
Pilastro A, Evans JP, Sartorelli S, Bisazza A (2002) Male phenotype predicts insemination success in guppies. Proceedings of the Royal Society of London Series B-Biological Sciences 269:1325- 1330
Pitnick S, Markow TA (1994) Male gametic strategies - Sperm size, testes size, and the allocation of ejaculate among successive mates by the sperm-limited fly Drosophila-Pachea and its relatives. American Naturalist 143:785-819
Pizzari T (2003) Food, vigilance, and sperm: the role of male direct benefits in the evolution of female preference in a polygamous bird. Behavioral Ecology 14:593-601
Polak M, Starmer WT (1998) Parasite-induced risk of mortality elevates reproductive effort in male Drosophila. Proceedings of the Royal Society of London Series B-Biological Sciences 265:2197-2201
Polis GA (1981) The evolution and dynamics of intraspecific predation. Annual Review of Ecology and Systematics 12:225-251
Pomfret J, Knell R (2006) Sexual selection and horn allometry in the dung beetle Euoniticellus intermedius. Animal Behaviour 71:567-576
Pomiankowski A (1987) The costs of choice in sexual selection. Journal of Theoretical Biology 128:195-218
Pomiankowski A (1988) The evolution of female mating preferences for male genetic quality. Oxford Surveys in Evolutionary Biology 5:136-184
Pomiankowski A, Iwasa Y, Nee S (1991) The evolution of costly mate preferences. I. Fisher and biased mutation. Evolution 45:1422-1430
Pomiankowski A, Möller AP (1995 ) A resolution of the lek paradox. Proceedings of the Royal Society B: Biological Sciences 260:21-29
Poulin R (1994) Mate choice decisions by parasitized female upland bullies, Gobiomorphus breviceps. Proceedings of the Royal Society of London Series B-Biological Sciences 256:183-187
166.

Prasad NG, Bedhomme S, Day T, Chippindale AK (2007) An evolutionary cost of separate genders revealed by male-limited evolution American Naturalist 169:29-37
Preston BT, Stevenson IR, Pemberton JM, Wilson K (2001) Dominant rams lose out by sperm depletion - A waning success in siring counters a ram's high score in competition for ewes. Nature 409:681-682
Price T, Schlüter D, Heckman NE (1993) Sexual selection when the female directly benefits. Biological Journal of the Linnean Society 48:187-211
Pyke GH, Pulliam HR, Charnov EL (1977) Optimal Foraging - Selective Review Of Theory And Tests. Quarterly Review of Biology 52:137-154
Queller DC (1997) Why do females care more than males? Proceedings of the Royal Society of London Series B-Biological Sciences 264:1555-1557
Qvarnström A (1997) Experimentally increased badge size increases male competition and reduces male parental care in the collared flycatcher. Proceedings of the Royal Society of London Series B-Biological Sciences 264:1225-1231
Qvarnstrom A, Forsgren E (1998) Should females prefer dominant males? Trends in Ecology & Evolution 13:498-501
Radesater T, Halldorsdottir H (1993) 2 Male types of the common earwig - male-male competition and mating success. Ethology 95:89-96
Raouf SA, Parker PG, Ketterson ED, Nolan V, Ziegenfus C (1997) Testosterone affects reproductive success by influencing extra-pair fertilizations in male dark-eyed juncos (Aves: Junco hyemalis). Proceedings of the Royal Society of London Series B-Biological Sciences 264:1599-1603
Reebs SG, Whoriskey FG, FitzGerald GJ (1984) Die! patterns of fanning activity, egg respiration, and the nocturnal behavior of male 3-spined sticklebacks, gasterosteus-aculeatus I (f-trachurus). Canadian Journal of Zoology-Revue Canadienne De Zoologie 62:329-334
Reichard M, Bryja J, Ondrackova M, Davidova M, Kaniewska P, Smith C (2005) Sexual selection for male dominance reduces opportunities for female mate choice in the European bitterling (Rhodeus sericeus). Molecular Ecology 14:1533-1542
Reynolds JD, Goodwin NB, Freckleton RP (2002) Evolutionary transitions in parental care and live bearing in vertebrates. Philosophical Transactions of the Royal Society of London Series B- Biological Sciences 357:269-281
Reynolds JD, Jones JC (1999) Female preference for preferred males is reversed under low oxygen conditions in the common goby (Pomatoschistus microps). Behavioral Ecology 10:149-154
Rice WR (1984) Sex chromosomes and the evolution of sexual dimorphism. Evolution 38:735-742
Rice WR, Chippindale AK (2001) Intersexual ontogenetic conflict. Journal of Evolutionary Biology 14:685-693
Rice WR, Holland B (1997) The enemies within: intergenomic conflict, interlocus contest evolution (ICE), and the intraspecific Red Queen. Behavioral Ecology and Sociobiology 41:1-10
Rintämaeki PT, Lundberg A, Alatalo RV, Höeglund J (1998) Assortative mating and female clutch investment in black grouse. Animal Behaviour 56:1399-1403
167.

Robertson DR (1984) Cohabitation of competing territorial damselfishes on a Caribbean coral reef. Ecology 65:1121-1135
Robertson DR, Lässig B (1980) Spatial distribution patterns and distribution of a group of territorial damselfishes from the Great Barrier Reef. Bulletin of Marine Science 30:187-203
Robertson DR, Polunin NVC, Leighton K (1979) The behavioural ecology of three Indian Ocean surgeonfishes (Acanthurus linearus, A. leucosternon and Zebrasoma ‘scopas): their feeding strategies and social and mating systems. Environmental Biology of Fishes 4:125- 170
Robson MA, Miles DB (2000) Locomotor performance and dominance in tree lizards, Urosaurus ornatus. Functional Ecology 14:338-344
Rodd FH, Hughes KA, Grether GF, Baril CT (2002) A possible non-sexual origin of mate preference: are male guppies mimicking fruit? Proceedings of the Royal Society of London Series B- Biological Sciences 269:475-481
Rogers W, Barlow GW (1991) Sex differences in mate choice in a monogamous biparental fish, the midas cichlid (Cichlasoma citrinellum). Ethology 87:249-261
Rohwer S (1978) Parent cannibalism of offspring and egg raiding as a courtship strategy. American Naturalist 112:429-440
Rowe L (1994) The costs of mating and mate choice in water striders. Animal Behaviour 48:1049- 1056
Rowe L, Arnqvist G (1996) Analysis of the causal components of assortative mating in water striders. Behavioural Ecology and Sociobiology 38:279-286
Rowe L, Cameron E, Day T (2005) Escalation, retreat, and female indifference as alternative outcomes of sexually antagonistic coevolution. American Naturalist 165:S5-18
Rowe L, Houle D (1996) The lek paradox and the capture of genetic variance by condition dependent traits. Proceedings of the Royal Society B: Biological Sciences 263:1415-1421
Russo AR (1987) ROLE OF HABITAT COMPLEXITY IN MEDIATING PREDATION BY THE GRAY DAMSELFISH ABUDEFDUF-SORDIDUS ON EPIPHYTAL AMPHIPODS. Marine Ecology- Progress Series 36:101-105
Ryan MJ, Rand AS (1990) The sensory basis of sexual selection for complex calls in the tungara frog, physalaemus-pustulosus (sexual selection for sensory exploitation). Evolution 44:305-314
Ryer CH, Stoner AW, Titgen RH (2004) Behavioral mechanisms underlying the refuge value of benthic habitat structure for two flatfishes with differing anti-predator strategies. Marine Ecology - Progresses Series 268:231-243
Sakaluk SK, Belwood JJ (1984) Gecko phonotaxis to cricket calling song: a case of satellite predation. Animal Behaviour 32:659-662
Sanz JJ, Kranenbarg S, Tinbergen JM (2000) Differential response by males and females to manipulation of partner contribution in the great tit (Parus major). Journal of Animal Ecology 69:74-84
Sargent RC (1985) Territoriality and reproductive tradeoffs in the 3-spine stickleback, gasterosteus- aculeatus. Behaviour 93:217-226
168.

Sargent RC (1988) Paternal care and egg survival both increase with clutch size in the fathead minnow, pimephales-promelas. Behavioral Ecology and Sociobiology 23:33-37
Sargent RC (1992) Ecology of filial cannibalism in fish: theoretical perspectives. In: Elgar MAC, B. J. (ed) Cannibalism: ecology and evolution among diverse taxa. Oxford University Press, pp 38- 62
Sargent RC, Crowley PH, Huang C, Lauer M, Neergaard D, Schmoetzer L (1995) A dynamic program for male parental care in fishes: Brood cycling and filial cannibalism. Behaviour 132:1059- 1078
Sargent RC, Gebier JB (1980) Effects of nest site concealment on hatching success, reproductive success, and paternal behavior of the threespine stickleback, gasterosteus-aculeatus. Behavioral Ecology and Sociobiology 7:137-142
Sargent RC, Gross MR (1985) Parental investment decision rules and the Concorde fallacy. Behavioral Ecology and Sociobiology 17:43-45
Sargent RC, Gross MR (1986) Williams' principle: An explanation of parental care in teleost fishes. In: Pitcher TJ (ed) The Behaviour of Teleost Fish. Crrom Helm, London, pp 275-293
Savino JF, Stein RA (1982) Predator-prey interaction between largemouth bass and bluegills as influenced by simulated, submersed vegetation. Transactions of the American Fisheries Society 111:255-266
Schaffer NM (1974a) Optimal reproductive effort in fluctuating environments. American Naturalist 108:783-798
Schaffer NM (1974b) Selection for optimal life histories: The effects of age structure. Ecology 55:291- 303
Scharf FS, Manderson JP, Fabrizio MC (2006) The effects of seafloor habitat complexity on survival of juvenile fishes: Species-specific interactions with structural refuge. Journal of Experimental Marine Biology and Ecology 335:167-176
Schneider KJ (1984) Dominance, Predation, And Optimal Foraging In White-Throated Sparrow Flocks. Ecology 65:1820-1827
Schoener TW (1986) A brief history of optimal foraging theory. In: Kamil AC, Krebs JR, Pulliam HR (eds) Foraging Behaviour. Plenium Press
Schwagmeyer PL, Mock DW, Parker GA (2002) Biparental care in house sparrows: negotiation or sealed bid? Behavioral Ecology 13:713-721
Seko T, Nakasuji F (2004) Effect of egg size variation on survival rate, development and fecundity of offspring in a migrant skipper, Parnara guttata guttata (Lepidoptera : Hesperiidae). Applied Entomology and Zoology 39:171-176
Semler DE (1971) Some aspects of adaptation in a polymorphism for breeding colours in Threespine Stickleback (Gasterosteus-Aculeatus). Journal of Zoology 165:291-&
Senar JC, Figuerola J, Pascual J (2002) Brighter yellow blue tits make better parents. Proceedings of the Royal Society of London Series B-Biological Sciences 269:257-261
Sevenster P (1961) A causal analysis of a displacement activity (fanning in Gasterosteus aculeatus L.). Behaviour 9 (Suppl.)
169.

Shackleton MA, Jennions MD, Hunt J (2005) Fighting success and attractiveness as predictors of male mating success in the black field cricket, Teleogryllus commodus: the effectiveness of no-choice tests. Behavioral Ecology and Sociobiology 58:1-8
Sheldon BC (2000) Differential allocation: tests, mechanisms and implications. Trends in Ecology and Evolution 15:397-402
Sih A (1980) Optimal Foraging - Partial Consumption Of Prey. American Naturalist 116:281-290
Sih A (1982) Optimal patch use - variation in selective pressure for efficient foraging. American Naturalist 120:666-685
Simmons LW (1992) Quantification of role reversal in relative parental investment in a bush cricket. Nature 358:61-63
Simmons LW, Parker GA (1989) Nuptial feeding in insects - mating effort versus paternal investment. Ethology 81:332-343
Skolbekken R, Utne-Palm AC (2001) Parental investment of male two-spotted goby, Gobiusculus flavescens (Fabricius). Journal of Experimental Marine Biology and Ecology 261:137-157
Slagsvold T, Lifjeld JT (1988) Plumage color and sexual selection in the pied flycatcher ficedula- hypoleuca. Animal Behaviour 36:395-407
Sloman KA, Motherwell G, O’Connor Kl, Taylor AC (2000) The effect of social stress on the standard metabolic rate (SMR) of brown trout, Salmo trutta. Fish Physiology and Biochemistry 23:49-53
Smiseth PT, Amundsen T (2000) Does female plumage coloration signal parental quality? A male removal experiment with the bluethroat (Luscinia s. svecica). Behavioral Ecology and Sociobiology 47:205-212
Smiseth PT, Ornborg J, Andersson S, Amundsen T (2001) Is male plumage reflectance correlated with paternal care in bluethroats? Behavioral Ecology 12:164-170
Smith C, Wootton RJ (1995) The costs of parental care in teleost fishes. Reviews in Fish Biology and Fisheries 5:7-22
Soler JJ, de Neve L, Martinez JG, Soler M (2001) Nest size affects clutch size and the start of incubation in magpies: an experimental study. Behavioral Ecology 12:301-307
Spence R, Smith C (2006) Mating preference of female zebrafish, Danio rerio, in relation to male dominance. Behavioral Ecology 17:779-783
Spoor WA (1984) Oxygen requirements of larvae of the smallmouth bass, Micropterus dolomieui Lacepede. Journal of Fish Biology 25:587-592
St Mary CM, Noureddine CG, Lindström K (2001) Environmental effects on male reproductive success and parental care in the Florida flagfish Jordanella floridae. Ethology 107:1035-1052
Stanley TR (2002) How many kilojoules does a Black-billed Magpie nest cost? Journal of Field Ornithology 73:292-297
Stein RA (1977) Selective Predation, Optimal Foraging, And Predator-Prey Interaction Between Fish And Crayfish. Ecology 58:1237-1253
170.

Stein RA, Magnuson JJ (1976) Behavioral response of crayfish to a fish predator. Ecology 57:751-761
Stutt AD, Siva-Jothy MT (2001) Traumatic insemination and sexual conflict in the bed bug Cimex lectularius. Proceedings of the National Academy of Sciences of the United States of America 98:5683-5687
Suk HY, Choe JC (2002) The presence of eggs in the nest and female choice in common freshwater gobies (Rhinogobius brunneus). Behavioral Ecology and Sociobiology 52:211-215
Summers K, Weigt LA, Boag P, Bermingham E (1999) The evolution of female parental care in poison frogs of the genus Dendrobates: evidence form mitochondrial DNA. Herpetologica 55:254-270
Svensson PA, Pelabon C, Blount JD, Surai PF, Amundsen T (2006) Does female nuptial coloration reflect egg carotenoids and clutch quality in the Two-Spotted Goby (Gobiusculus flavescens, Gobiidae)? Functional Ecology 20:689-698
Syriatowicz A, Brooks R (2004) Sexual responsiveness is condition-dependent in female guppies, but preference functions are not. BMC Ecology 4
Takahashi D, Kohda M (2004) Courtship in fast water currents by a male stream goby (Rhinogobius brunneus) communicates the parental quality honestly. Behavioral Ecology and Sociobiology 55:431-438
Takegaki T, Nakazono A (1999) Responses of the egg-tending gobiid fish Valenciennea longipinnis to the fluctuation of dissolved oxygen in the burrow. Bulletin of Marine Science 65:815-823
Tallamy DW (2000) Sexual selection and the evolution of exclusive paternal care in arthropods. Animal Behaviour 60:559-567
Tatrai I, Herzig A (1995) Effect of habitat structure on the feeding efficiency of young stages of razor fish (pelecus-cultratus (L)) - An experimental approach. Hydrobiologia 299:75-81
Taylor PD, Williams GC (1982) The lek paradox is not resolved. Theoretical Population Biology 22:392-409
Thresher RE (1984) Reproduction in reef fishes. T. F. H. Publications, Neptuen City, New Jersey
Tikkanen P, Huhta A, Muotka T (2000) Determinants of substrate selection in lotic mayfly larvae: is cryptic coloration important? Archiv Fur Hydrobiologie 148:45-57
Tilson RL (1981) Family formation strategies of Kloss's gibbons. Folia Primatologica 35:259-281
Tinbergen N (1951) The Study of Instinct. Oxford University Press, London
Tokarz RR (2002) An experimental test of the importance of the dewlap in male mating success in the lizard Anolis sagrei. Herpetologica 58:87-94
Tomkins JL, Radwan J, Kotiaho JS, Tregenza T (2004) Genic capture and resolving the lek paradox. Trends in Ecology & Evolution 19:323-328
Torricelli P, Lugli M, Gandolfi G (1985) A quantitative-analysis of the fanning activity in the male padogobius martensi (Pisces, Gobiidae). Behaviour 92:288-301
Tregenza T, Wedelt N (1997) Definitive evidence for cuticular pheromones in a cricket. Animal Behaviour 54:979-984
171.

Trivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Sexual Selection and the Descent of man. Aldine Publishing Company, London, pp 136-179
Tupper M, Boutilier RG (1995) Effects of habitat on settlement, growth, and postsettlement survival of atlantic cod (gadus-morhua). Canadian Journal of Fisheries and Aquatic Sciences 52:1834- 1841
Unger LM, Sargent RC (1988) Allopaternal care in the fathead minnow, Pimephales promelas - Females prefer males with eggs. Behavioral Ecology and Sociobiology 23:27-32
Vahed K (1998) The function of nuptial feeding in insects: a review of empirical studies. Biological Reviews 73:43-78
van den Berghe EP, Wernerus F, Warner RR (1989) Female choice and the mating cost of peripheral males. Animal Behaviour 38:875-884
van lersel JJA (1953) An analysis of parental behaviour of the male three spined stickleback (Gasterosteus aculeatus L). Behaviour 3 (Suppl.):1-159
Vincent A, Ahnejsö I, Berglund A, Rosenqvist G (1992) Pipefishes and seahorses: are they all sex role reversed? Trends in Ecology & Evolution 7:237-241
Vinyoles D, Cöte IM, de Sostoa A (1999) Egg cannibalism in river blennies: the role of natural prey availability. Journal of Fish Biology 55:1223-1232
Voltura KM, Schwagmeyer PL, Mock DW (2002) Parental feeding rates in the house sparrow, Passer domesticus: Are larger-badged males better fathers? Ethology 108:1011-1022
Wagner WE, Harper CJ (2003) Female life span and fertility are increased by the ejaculates of preferred males. Evolution 57:2054-2066
Wagner WEJ (1998) Measuring female mating preferences. Animal Behaviour 55:1029-1042
Watson PJ, Arnqvist G, Stallmann RR (1998) Sexual conflict and the energetic costs of mating and mate choice in water striders. American Naturalist 151:46-58
Webb JN, Houston AI, McNamara JM, Szekely T (1999) Multiple patterns of parental care. Animal Behaviour 58:983-993
Webster MM, Hart PJB (2004) Substrate discrimination and preference in foraging fish. Animal Behaviour 68:1071-1077
Wedell N, Kvarnemo C, Lessells CM, Tregenza T (2006) Sexual conflict and life histories. Animal Behaviour 71:999-1011
Werner EE, Gilliam JF, Hall DJ, Mittelbach GG (1983) An experimental test of the effects of predation risk on habitat use in fish. Ecology 64:1540-1548
Werner EE, Hall DJ (1974) Optimal Foraging and Size Selection of Prey by Bluegill Sunfish (Lepomis macrochirus). Ecology 55:1042-1052
Werner EE, Mittelbach GG (1981) Optimal Foraging - Field-Tests Of Diet Choice And Habitat Switching. American Zoologist 21:813-829
172.

West-Eberhard MJ (1984) Sexual selection, competitive communication and species-specific signals in insects. In: Lewis T (ed) Insect Communication Academic Press, London, pp 283-324
Westneat DF, Sherman PW, Morton ML (1990) The ecology and evolution of extra-pair copulations in birds. Current Ornithology 7:331-369
Weygoldt P (1987) Evolution of parental care in dart poison frogs (Amphibia: Anura: Dendrobatidae). Z. Zool. Syst. Evol. 25:51-67
White DJ, King AP, West MJ (2002) Facultative development of courtship and communication in juvenile male cowbirds (Molothrus ater). Behavioral Ecology 13:487-496
Whittingham LA, Dunn PO, Robertson RJ (1994) Female response to reduced male parental care in birds - an experiment in tree swallows. Ethology 96:260-269
Wiley RH, Poston J (1996) Indirect mate choice, competition for mates, and coevolution of the sexes. Evolution 50:1371-1381
Williams GC (1966a) Adaptation and natural selection. Princeton University Press, Princeton
Williams GC (1966b) Natural selection costs of reproduction and a refinement of Lacks principle. American Naturalist 100:687-&
Winkler DW (1987) A general model for parental care. American Naturalist 130:526-543
Wong BBM (2004) Superior fighters make mediocre fathers in the Pacific blue-eye fish. Animal Behaviour 67:583-590
Wootton RJ (1998) Ecology of teleost fishes. Kluwer, Dordrecht
Wright J, Cuthill I (1989) Manipulation of sex-differences in parental care. Behavioral Ecology and Sociobiology 25:171 -181
Yenisetti SC, Hegde SN (2003) Size-related mating and reproductive success in a drosophilid: Phorticella striata. Zoological Studies 42:203-210
Zahavi A (1975) Mate selection: a selection for a handicap. Journal of Theoretical Biology 53:205-214
Zink AG (2003) Quantifying the costs and benefits of parental care in female treehoppers. Behavioral Ecology 14:687-693
Zoran MJ, Ward JA (1983) Parental egg care behavior and fanning activity for the orange chromide, Etroplus maculatus. Environmental Biology of Fishes 8:301-310
173.

CHAPTER 5 General Discussion
5.1 Aims of this study
In this study, I aimed to measure and quantity what traits female Mogurnda mogurnda
preferred in relation to male size and fighting ability. There is growing acceptance that the previously
assumed positive correlation between attractiveness and male size/dominance is not always correct.
The results of my simultaneous choice test imply, however, that females do in fact prefer larger males.
I feel confident inferring that preferred males are more dominant, as this species is very aggressive
and observational data clearly shows that larger individuals win more fights. Interestingly, I found
evidence of size assortative mating with larger females tending to spend more time with larger than
smaller males, and I also found that that larger females exhibited a stronger preference for the
preferred male than smaller females, implying that female choice in this species is condition-
dependent.
In the recent literature, however, the accuracy of simultaneous choice tests has been called
into question. I aimed to compare the expression of female mate preferences from a traditional
association preference tests to a method of assessing choice that has been gaining ground in recent
years: latency to mating tests. Often when the results of association preference tests are compared to
latency tests, either preferred characteristics are no longer preferred or different characteristics
become indicators of male attractiveness. I found the male trait that was preferred in traditional tests
(male size) were not preferred in latency tests, supporting this view. At this point, I wish to clarify the
terminology I will use in this discussion due to female choice being judged on two different
methodologies and that led to opposing results. I will continue using “preferred” and “non-preferred”
when discussing the males chosen under traditional choice tests in Chapter 2, but I will refer to males
chosen by the latency to mating test as “mated” and “non-mated” males. I chose this terminology
because the trend of females taking longer to spawn with preferred males was largely due to five
preferred males failing to spawn in the allotted time.
Another aim was to identify any traits that honestly signalled male parental quality and
whether females were choosing males based on these characteristics. I found no correlations between
male size and parental care. Fan beat rate was the only parental care behaviour that correlated with a
male trait, and is was negatively correlated with female preference itself, and so was a dishonest
signal of care. In the latency to mating tests, it is unclear what traits females based their preference
174.

response on, but since they spawned sooner with non-preferred males who showed higher quality
parental care than preferred males, females could still be choosing males based on a trait correlated
with quality of care. (Note: in this chapter, I will conceptually treat male attractiveness as a trait
because if it is correlated to one or more variable phenotypic traits that females base their choice on
which is a sound assumption otherwise females would not be discriminating between males). Male
attractiveness could be treated as a trait when looking for effects on care behaviour.
Finally, with the focus on so many energetically expensive activities I decided to examine one
aspect of foraging in M. mogurnda: patch choice and substrate discrimination. I found that there was
no preference for the greater potential for food presented by the higher incidence of interstitial spaces
in “complex” substrates. The only preference I found was by females but it was for “simple” substrates,
which was the opposite result from a recent paper that conducted a similar study. Potential prey
density is not the only factor influencing foraging behaviour, however, and the increased search and
handling time associated with prey in complex microhabitats and the lower energetic award from
smaller prey items may have balanced the value of seeking prey on complex and simple substrates.
5.2 Condition-dependent female choice
The discovery that female choice in this species is potentially condition-dependent is exciting.
This species satisfies two predictions of condition-dependence (stronger preference in larger females
and size assortative mating) while empirical evidence to support this theory is still scarce. Size-
assortative mating itself is common across taxa in fish, such as cichlids (Barlow et al. 1990; Budaev et
al. 1999), and coral gobies Paragobioon echinocephalus (Kuwamura et al. 1993). Condition-
dependent female choice theory predicts that better quality females exhibit stronger preferences for
the same costs as lower quality females and therefore have better mates through either more intense
searching or having a higher rejection rate than lower quality females.
Finding of stronger expression of preference in larger females is not uncommon, however.
Notable examples include guppies Poecilia reticulata (Lopez 1999; Syriatowicz and Brooks 2004),
three spined sticklebacks Gasterosteus aculeatus (Bakker et al. 1999), upland bullies Gobiomorphus
breviceps (Poulin 1994) and deep-snouted pipefish Syngnathus typhie (Mazzi 2004). Mate preference
is predicted to be differentially costly with respect to female quality (Cotton et al. 2006) with higher
quality females suffering lower costs for a given mate preference, or are able to be more discerning for
the same cost, than poorer quality females. By much the same mechanism that secondary sexual
ornaments are predicted to be condition-dependent, female mating preferences are predicted to be
175.

weaker when the costs are higher so the strength of female preference is expected to vary with female
condition (Pomiankowski 1987; Grafen 1990; Iwasa et al. 1991; Iwasa and Pomiankowski 1999).
Interestingly, honest signaling of male secondary sexual characteristics may be partially affected by
female condition-dependence (Cotton et al. 2006). Mate preferences for honest signals should be
more beneficial for high quality females if low quality females have less to gain from discrimination. For
example, in a species where male display rate honestly signals male investment in parental care, high
quality females who can produce more offspring than the average male can care for should prefer
well-ornamented mates, as this will directly increase their fitness. Lower quality females whose
maximal egg production is smallenough that an average male could care for them would have less to
benefit from expressing this mating preference and would more likely to mate randomly to avoid
paying the costs of mate choice (Cotton et al. 2006).
5.3 Parental care behaviours in M. Mogurnda
Individuals are expected to maximise their overall reproductive success by trading off
investment between current and future reproductive events (Winkler 1987). Parents are expected to
adjust parental care in relation to their own fitness, because parental care is demonstrably costly (e.g.,
Drent and Daan 1980a; Stanley 2002; Hanssen et al. 2003). Body condition is an important factor
affecting the quality of parental care an individual can provide, so it is reasonable to expect larger
individuals in better condition to be better quality parents. I did not, however, find this to be the case in
my study of M. mogurnda.
I found no relationship between male size and any of the measured parental care behaviours:
fanning, guarding, filial cannibalism or the resulting hatching time. Fanning is a particularly expensive
care behaviour, especially under sup-optimal condition such as low dissolved oxygen levels (Hale et
al. 2003; Lissaker et al. 2003). There may not have been any difference in fanning behaviours
between males of different sizes because of the favourable conditions in the breeding tanks. Oxygen
levels and water quality remained high at all times, requiring relatively low effort from males to care for
the eggs. The very low proportion of whole filial cannibalism (13.1%) supports this, and these
incidences were almost certainly due to human disturbance as they almost always occurred the day
after I recorded parental care behaviours. Partial filial cannibalism has been suggested to helps
parents recover the energetic costs of parental care (Rohwer 1978) and there were no recorded partial
filial cannibalisms, suggesting that care under these conditions were not particularly expensive. There
176.

also may not have been enough difference in condition between large and smallmales to detect
differences in care, as I fed fish a reasonable amount of food every two days.
I also found no difference in any of the parental care behaviours between preferred and non
preferred, except for fanning rate. It was unexpected to find that preferred males fanned at a lower
rate. As already discussed, this poses a dilemma as it makes female choice maladaptive, with females
actually suffering a cost of decreased male care from their mating preference. This leads to the
question of how female choice is maintained in this species if females are incurring higher costs for
their mating preferences. There are several possible explanations for the discovery of maladaptive
female choice in this study; females could be gaining material benefits unrelated to parental care;
females could be gaining genetic benefits, or there is the alternate interpretation that my method for
discriminating female preference is inaccurate.
5.4 Maintaining a maladaptive preference
I found no difference in any of the parental care behaviours between preferred and non
preferred, except for fanning rate. I did not expect to find that preferred males were lower quality
parents, as the most obvious benefit from female choice in this species would seem to be improved
parental care because females receive little else from males. An increase in female reproductive
output offers the most straightforward explanation as to how mate choice benefits outweighs the costs
of choice (Iwasa and Pomiankowski 1999), which explains why it has been the main focus of research
in many studies (e.g., Pizzari 2003; Wagner and Harper 2003). Female choice is a powerful tool of
sexual selection, having been suggesting as the driver for the evolution of exclusive paternal care in
arthropods (Tallamy 2000) and territorial fish (Ah-King et al. 2005). I discussed other potential material
and genetic benefits in Chapter 3, and females could receive a mixture of direct and indirect benefits
to help compensate for the costs of mate choice (Cordero and Eberhard 2003; Kokko et al. 2003).
There is also the possibility that the sensory bias model of choice may be maintaining female choice
(West-Eberhard 1984; Ryan and Rand 1990; Endler and Basolo 1998; reviewed in Fuller et al. 2005).
It has the benefit of being applicable to instances of maladaptive choice (Rice and Holland 1997;
Holland and Rice 1998) such as this one, and females may still accrue benefits from better-quality
mates. Unfortunately, however, there is no direct evidence from my study to support this model of
female choice.
177.

5.5 Comparing female choice tests
The most obvious answer to the appearance of maladaptive choice in this study is that
female’s may not actually prefer worse fathers because the preferred males in this study may not
actually be more attractive that non-preferred males. There is growing evidence that simultaneous
choice tests are not the most accurate method of discriminating female choice (Wagner 1998;
Shackleton et al. 2005; Cotton et al. 2006). It is possible that the use of female preference responses
from traditional choice tests simply did not tell the whole story. The incongruence of female choice
between the two methods of measuring female choice implies that mating preferences are based on
parameters that are more complex than visual (and possibly chemical) cues (Candolin 1999a; Nilsson
and Nilsson 2000). Choosing who to associate with who to actually mate with could be quite different
decisions, and the expression of choice based on association into actual increased mating success
might reflect this.
5.6 Variation in expression of mating preference
It is quite plausible that females show a context dependent expression of choice. When
restricted from mating cues and interactions, females may chose to associate with the larger male in a
non-reproductive context, or male size may only be one of many preference cues. As already
discussed, M. mogurnda are an aggressive species and males might protect females from other
aggressive individuals during or after spawning. Females might also initially choose larger males if
they provide access to superior resources, such as better foraging grounds or better spawning sites.
In a mating context where females have access to the whole gamut of cues and interactions, they may
select using different criteria. A size-based preference might diminish if females can read cues that
imply larger males provide lower quality parental care (Forsgren 1997; Wong 2004), are likely to
transmit diseases (Freeland 1981) or are sperm depleted (Pitnick and Markow 1994; Preston et al.
2001).
Females were quite young (though all were old enough to be identified as females which
means they are of reproductive age) when the first association trials were conduction. This may have
had an effect. Clement et al. (2005) showed that differences in female reproductive state correspond
to changes in female affiliation with males. In the African cichlid fish Astatotilapia burtoni, gravid and
non-gravid females were given the choice of associating with territorial (reproductively active) or non
territorial males. Gravid females preferentially associated with territorial males, whereas non-gravid
178.

females showed no preference. This study highlights that reproductive state of both males and
females could be an important factor in the expression of mating preferences.
As discussed in Chapter 3, the incongruence between the two results could be due to sex role
reversal. The delay in spawning time (including the five pairs who did not spawn at all) could be
explained as being a consequence of males exerting mate choice and delaying or preventing mating
with unattractive females. Male M. mogurnda invest heavily into offspring by guarding them (possible
to the detriment of foraging which will reduce their condition) while females only provide eggs. If a
restricting factor such as suitable spawning sites is scarce, the operational sex ratio (OSR) may skew
towards females and sex roles are largely determined by OSR (Emlen and Oring 1977; Clutton-Brock
and Parker 1992; Kvarnemo and Ahnejsö 1996). The difference in the cost of breeding between
sexes, however, appears theoretically to be more important than the OSR in determining the sex roles
in many cases (Johnstone et al. 1996; Kokko and Monaghan 2001). The sex differences in this
species in terms of costs of choice are therefore are very likely to have a strong effect on choosiness.
5.7 Cues to mating preference
The only detectable relationship between parental care and male attractiveness (as defined by
association trials) was a negative one. The latency to mating measure of female choice, however, may
be a more accurate measure of choice, which changes the interpretation of my results. Using the
spawning time measure of choice, females showed no preference for male size. Mated males (defined
as the males females spawned with) were also the non-preferred males from the mate choice
experiment and the non-preferred males fanned at a higher rate. So, even though there was no
obvious cues as to why females preferred mated males, this indicates that mated males are better
quality parents. Female may then still be basing their mating preferences on parental care or a trait
correlated with it.
Possible other cues for female choice could be fanning itself, as it has been shown to be a
trait for mate choice (e.g., Sevenster 1961; St Mary et al. 2001; Pampoulie et al. 2004) and could also
be an honest signal of male quality (Östlund and Ahnesjö 1998). It is logical that females base their
preference on a trait that honestly signals condition, as condition will effect the quality of parental care.
There is a great deal of literature on the correlation between sexual displays and parental investment
showing positive (e.g., Linville et al. 1998; Candolin 2000; Senar et al. 2002; Voltura et al. 2002; Kolm
2004; Halupka and Borowiec 2006), negative (e.g., Qvarnström 1997; Wong 2004; Mitchell et al.
2007) and no relationship (e.g., Mountjoy and Lemon 1997; Smiseth et al. 2001). This species
179.

displays no obvious secondary sexual traits, but signals of parental care need not necessarily be
sexually selected traits. For example, male three spined sticklebacks have larger pectoral fins than
females, which females can use as a cue to male parental quality (Bakker and Mundwiler 1999).
Finally, with the focus of so much of this study on energetically expensive activities like mate
searching, mate choice, mating, and parental care I turn to the means of maintaining condition and
obtaining energy for these activities.
5.8 Foraging
The efficiency with which individuals gain energy is of immense importance and affects their
competitiveness in competition (e.g., dominance struggles) and the expression of traits that are
condition-dependent (e.g., honest signals of quality). As discussed earlier, different habitat structures
present animals with differences in food density and associated food values, such as search times and
there will often be a higher abundance of prey around structurally complex habitats (Charnov 1976);
Stein & Magnuson 1976). Predators might therefore be expected to prefer foraging in patches of
greater structural complexity, such as in Webster and Plarts’ (2004) study on three spined sticklebacks
Gasterosteus aculeatus. The sticklebacks spent more time foraging in complex microhabitats when
food densities were equal between substrates, and when density was greater on the complex
substrate. In my study, I did not find a clear preference for either zone in M. mogurnda. Females did
show a preference for simple substrates (based on one of several measure of association) while
males showed no preference at all. Although increased structural complexity hides a potentially higher
prey density, the prey will have an associated increase in handling and searching times and this will
decrease the net energetic value of prey and therefore the net intake of energy for that patch. It could
also be that the fish used in this experiment were quite young when they were caught and may not
have leaned which habitat patches have greater abundances of prey. Ultimately, the degree to which
habitat complexity influences foraging behaviour depends on a complex suite of factors, such as prey
antipredator responses, the foraging ability of the predator, and the quality and quantity of the habitat
structure (Eklöv and Diehl 1994).
180.

5.9 Conclusion
Fish are an under utilised model system for studies of mate choice and parental care. They
present unique opportunities because they are the most numerous and varied of the vertebrates and
show a wide range of often dynamic care patterns ranging from no-care, to uniparental, paternal and
maternal care (Gross and Sargent 1985; Reynolds et al. 2002). If fish biologists are to work on a
species that is not well-known like guppies or sticklebacks, they need to spend a lot of time learning
about the fish they are studying. This study has made some interesting discoveries and posing some
even more interesting questions that I am unable to answer because I do not have sufficient data on
the natural breeding system, detailed information about the mating and care behaviours, and most
importantly the relative costs and benefits of these behaviours. I think Mogurnda mogurnda shows the
potential for being an exciting species to examine in terms of empirical studies on mate choice and
parental care. The first step would be to study this species in the wild to learn how to apply the
theoretical background I have outlined in this thesis. Physiological measures of the costs of care,
particularly along environmental gradients such as those associated with oxygen and turbidity, would
address aspects of the literature that have not yet been comprehensively covered. I think this species
is highly suited to such studies.
181 .

References
(2004) Fishbase (www.fishbase.org). In: Froese R, Pauly D (eds)
Abrahams MV, Dill LM (1989) A determination of the energetic equivalence of the risk of predation. Ecology 70:999-1007
Ah-King M, Kvarnemo C, Tullberg BS (2005) The influence of territoriality and mating system on the evolution of male care: a phylogenetic study on fish. Journal of Evolutionary Biology 18:371- 382
Alatalo RV, Carlson A, Lundberg A (1988) The search cost in mate choice of the Pied Flycatcher. Animal Behaviour 36:289-291
Allen GR, Midgley SH, Allen M (2002) Field Guide to the Freshwater Fishes of Australia. Western Australian Museum, Perth
Amundsen T (2003) Fishes as models in studies of sexual selection and parental care. Journal of Fish Biology 63:17-52
Anderson GRV, Ehrlich AH, Ehrlich PR, Roughgarden JD, Russell BC, Talbot FH (1981) The community structure of coral reef fishes. American Naturalist 117:476-495
Andersson M (1994) Sexual Selection. Princeton University Press, Princeton, New Jersey
Andersson S, Pryke SR, Ornborg J, Lawes MJ, Andersson M (2002) Multiple receivers, multiple ornaments, and a trade-off between agonistic and epigamic signaling in a widowbird. American Naturalist 160:683-691
Arnqvist G, Nilsson T (2000) The evolution of polyandry: multiple mating and female fitness in insects. Animal Behaviour 60:145-164
Arnqvist G, Rowe L (1995) Sexual Conflict and Arms Races between the Sexes - a Morphological Adaptation for Control of Mating in a Female Insect. Proceedings of the Royal Society of London Series B-Biological Sciences 261:123-127
Arnqvist G, Rowe L (2005) Sexual Conflict. Princeton University Press, Princeton, NJ
Arnqvist G, Rowe L, Krupa JJ, Sih A (1996) Assortative mating by size: a meta-analysis of mating patterns in water striders. Evolutionary Ecology 10:265-284
Babbitt KJ, Jordan F (1996) Predation on Bufo terrestris tadpoles: Effects of cover and predator identity. Copeia:485-488
Babbitt KJ, Tanner GW (1998) Effects of cover and predator size on survival and development of Rana utricularia tadpoles. Oecologia 114:258-262
Baber MJ, Babbitt KJ (2004) Influence of habitat complexity on predator-prey interactions between the fish (Gambusia holbrooki) and tadpoles of Hyla squirella and Gastrophryne carolinensis. Copeia:173-177
BakkerTCM (1993) Positive correlation between female preference and preferred male ornament in sticklebacks. Nature 363
182.

Bakker TCM, Kunzler R, Mazzi D (1999) Condition-related mate choice in sticklebacks. Nature 401:234
Bakker TCM, Mundwiler B (1999) Pectoral fin size in a fish species with paternal care: a condition- dependent sexual trait revealing infection status. Freshwater Biology 41:543-551
Balshine-Earn S (1995) The costs of parental care in Galilee St Peter's fish, Sarotherodon galilaeus. Animal Behaviour 50:1-7
Barlow G, Francis RC, Baumgartner FV (1990) Do the colours of parents, companions and selfinfluence assortative mating in the polychromatic Midas cichlid? Animal Behaviour 40:713-722
Bartholomew A, Diaz RJ, Cicchetti G (2000) New dimensionless indices of structural habitatcomplexity: predicted and actual effects on a predator's foraging success. Marine Ecology- Progress Series 206:45-58
Bateman AJ (1948) Intra-Sexual Selection In Drosophila. Heredity 2:349-368
Baylis JR (1981) The evolution of parental care in fishes, with reference to Darwins rule of male sexual selection. Environmental Biology of Fishes 6:223-251
Belles-lsles JC, Fitzgerald GJ (1991) Filial cannibalism in sticklebacks - a reproductive management strategy. Ethology Ecology & Evolution 3:49-62
Benkman CW (1987) Food profitability and the foraging ecology of crossbills. Ecological Monographs 57:251-267
Berejikian BA, Tezak EP, Larae AL (2000) Female mate choice and spawning behaviour of Chinook salmon under experimental conditions. Journal of Fish Biology 57:647-661
Berglund A, Bisazza A, Pilastro A (1996) Armaments and ornaments: An evolutionary explanation of traits of dual utility. Biological Journal of the Linnean Society 58:385-399
Beukema JJ (1968) Predation by 3-spined stickleback (gasterosteus aculeatus I) - influence of hunger and experience. Behaviour 31:1-&
Beukers JS, Jones GP (1998) Habitat complexity modifies the impact of piscivores on a coral reef fish population. Oecologia 114:50-59
Birkhead TR, Möller AP (1992) Sperm Competition in Birds. Academic Press, New York.
Bisazza A, Marconato A (1988) Female mate choice, male-male competition and parental care in the river bullhead, Cottus gobio L. (Pisces, Cottidae). Animal Behaviour 36:1352-1360
Blanchfield PJ, Ridgway MS (1997) Reproductive timing and use of redd sites by lake-spawning brook trout (Salvelinus fontinalis). Canadian Journal of Fisheries and Aquatic Sciences 54:747-756
Blanckenhorn WU (2002) The costs of copulating in the dung fly Sepsis cynipsea. Behavioral Ecology and Sociobiology 13
Blanckenhorn WU, Kraushaar U, Reim C (2003) Sexual selection on morphological and physiological traits and fluctuating asymmetry in the yellow dung fly. Journal of Evolutionary Biology 16:903- 913
183.

Blows MW (1999) Evolution of the genetic covariance between male and female components of mate recognition: an experimental test. Proceedings of the Royal Society B: Biological Sciences 266:2169-2174
Blumer LS (1979) Male parental care in the bony fishes. Quarterly Review of Biology 54:149-161
Blumer LS (1982) A bibliography and categorization of bony fish exhibiting parental care. Zoological Journal of the Linnean Society 76:1-22
Blumstein DT, Evans CS, Daniel JC (2000) JWatcher. In, 0.9 edn. Animal Behaviour Laboratory,Macquarie University, Sydney, pp A powerful tool for the quantitative analysis of behaviour.
Bonduriansky R, Rowe L (2005) Sexual selection, genetic architecture, and the condition dependence of body shape in the sexually dimorphic fly Prochyliza xanthostoma (Piophilidae). Evolution 59:138-151
Borgia G (1979) Sexual selection and the evolution of mating systems. In: Blum MS, Blum NA (eds) Sexual selection: testing the alternatives. Academic Press, New York, pp 19-80
Brooks JL (1968) Effects of prey size selection by lake planktivores. Systematic Zoology 17:273-&
Brown WD (1990) Size-assortative mating in the blister beetle Lytta magister (Coleoptera: Meloidae) is due to male and female preference for larger mates. Animal Behaviour 40:901-909
Brown WD (1993) The cause of size-assortative mating in the leaf beetle Trirhabda canadensis (Coleoptera: Chrysomelidae). Behavioral Ecology and Sociobiology 33:151-157
Buchanan KL, Catchpole CK (2000) Song as an indicator of male parental effort in the sedge warbler. Proceedings of the Royal Society of London Series B-Biological Sciences 267:321-326
Budaev SV, Zworykin DD, Mochek AD (1999) Individual differences in parental care and behaviour profile in the convict cichlid: a correlation study. Animal Behaviour 58:195-202
Bukowski TC, Linn CD, Christenson TE (2001) Copulation and sperm release in Gasteracantha cancriformis (Araneae : Araneidae): differential male behaviour based on female mating history. Animal Behaviour 62:887-895
Bulmer M (1989) Structural instability of models of sexual selection. Theoretical Population Biology 35:195-206
Burley N (1986) Sexual selection for aesthetic traits in species with biparental care. American Naturalist 127:415-445
Burley N (1988) The differential-allocation hypothesis: an experimental test. American Naturalist 132:611-628
Burley NT, Parker PG, Lundy K (1996) Sexual selection and extrapair fertilization in a sociallymonogamous passerine, the zebra finch (Taeniopygia guttata). Behavioral Ecology 7:218-226
Burt A (1995) Perspective: the evolution of fitness. Evolution 49:1-8
Burt A (2000) Natural selection in the wild. Trends in Ecology & Evolution 15:306-307
184.

Candolin U (1999a) Male-male competition facilitates female choice in sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 266:785-789
Candolin U (1999b) The relationship between signal quality and physical condition: is sexual signalling honest in the three-spined stickleback? Animal Behaviour 58:1261-1267
Candolin U (2003) The use of multiple cues in mate choice. Biological Review 78:575-595
Caraco T, Martindale S, Whittam TS (1980) An empirical demonstration of risk-sensitive foraging preferences. Animal Behaviour 28:820-830
Carlisle TR (1982) Brood success in variable environments - Implications for parental care allocation. Animal Behaviour 30:824-83^
Charnov EL (1976) Optimal Foraging, Marginal Value Theorem. Theoretical Population Biology 9:129- 136
Chellappa S, Huntingford SA (1989) Depletion of energetic reserves during reproductive aggression in male three-spined stickleback, Gasterosteus aculeatus. Journal of Fish Biology 35:275-286
Chippindale AK, Gibson JR, Rice WR (2001) Negative genetic correlation for adult fitness between sexes reveals ontogenetic conflict in Drosophila. Proceedings of the National Academy of Sciences of the United States of America 98:1671-1675
Choe JC, Crespi BJ (1997) Mating systems in insects and arachnids. Cambridge University Press, Cambridge
Clark DL, Biesiadecki B (2002) Mating success and alternative reproductive strategies of thedimorphic jumping spider, Maevia inclemens (Aaneae, Salticidae). Journal of Arachnology 30:511-518
Clarke RD (1977) Habitat distribution and species diversity of chaetodontid and pomacentrid fish near Bimini, Bahamas. Marine Biology 40:277-289
Clement TS, Grens KE, Fernald RD (2005) Female affiliative preference depends on reproductive state in the African cichlid fish, Astatotilapia burtoni. Behavioral Ecology 16:83-88
Clifford HF, Gotceitas V, Casey RJ (1989) Roughness and color of artificial substratum particles as possible factors in colonization of stream invertebrates. Hydrobiologia 175:89-95
Clutton-Brock TH (1991) The Evolution of Parental Care. Princeton University Press, Princeton
Clutton-Brock TH, Parker GA (1992) Potential reproductive rates and the operation of sexual selection. Quarterly Review of Biology 67:437-456
Coleman RM, Fischer RU (1991) Brood size, male fanning effort and the energetics of a nonshareable parental investment in bluegill sunfish, lepomis-macrochirus (Teleostei, Centrarchidae). Ethology 87:177-188
Coleman RM, Gross MR, Sargent RC (1985) Parental investment decision rules - a test in bluegill sunfish. Behavioral Ecology and Sociobiology 18:59-66
Collins MA, Bailey DM, Ruxton GD, Priede IG (2005) Trends in body size across an environmental gradient: A differential response in scavenging and non-scavenging demersal deep-sea fish. Proceedings of the Royal Society B-Biological Sciences 272:2051-2057
185.

Cordero C, Eberhard WG (2003) Female choice of sexually antagonistic male adaptations: a critical review of some current research. Journal of Evolutionary Biology 16:1-6
Cöte IM, Hunte W (1989) Male and female mate choice in the redlip blenny - why bigger is better. Animal Behaviour 38:78-88
Cotton S, Rogers DW, Small J, Pomiankowski A, Fowler K (2006a) Variation in preference for a male ornament is positively associated with female eyespan in the stalk-eyed fly Diasemopsis meigenii. Proceedings of the Royal Society of London Series B-Biological Sciences 273:1287- 1292
Cotton S, Small J, Pomiankowski A (2006b) Sexual Selection and Review Condition-Dependent Mate Preferences. Current Biology 16:755-765
Crudgington HS, Siva-Jothy MT (2000) Genital damage, kicking and early death - The battle of the sexes takes a sinister turn in the bean weevil. Nature 407:855-856
Cunningham EJA, Russell AF (2000) Egg investment is influenced by male attractiveness in the mallard. Nature 404:74-77
Curio E (1988) Relative realized life span and delayed cost of parental care. American Naturalist 131:825-836
Daly M (1978) Cost of mating. American Naturalist 112:771-774
Davis JC (1975) Exchange of oxygen at gills of fish in response to oxygen availability. Journal of the Electrochemical Society 122:C101-C101
Dawkins R, Carlisle TR (1976) Parental investment, mate desertion and a fallacy. Nature 262:131-133
de Lope F, Möller AP (1993) Female reproductive effort depends on the degree of ornamentation of their mates. Evolution 47:1152-1160
del Hoyo J, Elliott A, Sargatal J (1992-2003) Handbook of the Birds of the World. Lynx Edicions, Barcelona
DeWoody JA, Fletcher DE, Wilkins SD, Avise JC (2001) Genetic documentation of filial cannibalism in nature. Proceedings of the National Academy of Sciences of the United States of America 98:5090-5092
Diehl S (1988) Foraging efficiency of 3 fresh-water fishes - effects of structural complexity and light. Oikos 53:207-214
Diehl S (1992) Fish predation and benthic community structure - the role of omnivory and habitat complexity. Ecology 73:1646-1661
Drent RH, Daan S (1980a) The prudent parent: Energetic adjustments in avian breeding. Ardea 68:225-252
Drent RH, Daan S (1980b) The prudent parent: Energetic adjustments in avian breeding. Ardea 68:225-252
Dudley RG, Eipper AW (1975) Survival of largemouth bass embryos at low dissolved oxygen concentrations. Transactions of the American Fisheries Society 104:122-128
186.

Eberhard WG (1996) Female control: sexual selection by cryptic female choice. Princeton University Press, Princeton, New Jersey
Eberhard WG, Cordero C (1995) Sexual selection by cryptic female choice on male seminal products - a new bridge between sexual selection and reproductive physiology. Trends in Ecology & Evolution 10:493-496
Eggert AK, Reinking M, Muller JK (1998) Parental care improves offspring survival and growth in burying beetles. Animal Behaviour 55:97-107
Eklov P (1997) Effects of habitat complexity and prey abundance on the spatial and temporaldistributions of perch (Perea fluviatilis) and pike (Esox lucius). Canadian Journal of Fisheries and Aquatic Sciences 54:1520-1531
Eklov P, Diehl S (1994) Piscivore efficiency and refuging prey - the importance of predator search mode. Oecologia 98:344-353
Eklöv P, Diehl S (1994) Piscivore efficiency and refuging prey - the importance of predator search mode. Oecologia 98:344-353
Emlen ST, Oring LW (1977) Ecology, sexual selection, and the evolution of mating systems. Science 197:215-223
Endler JA (1980) Natural selection on color in Poecilia reticulata. Evolution 34:76-91
Endler JA (1983) Natural and sexual selection on color patterns in Poeciliid fishes. Environmental Biology of Fishes 9:173-190
Endler JA (1988) Sexual selection and predation risk in Guppies. Nature 332:593-594
Endler JA, Basolo AL (1998) Sensory ecology, receiver biases and sexual selection. Trends in Ecology & Evolution 13:415-420
Evans JP, Kelly JL, Bisazza A, Finazzo E, Pilastro A (2004) Sire attractiveness influences offspring performance in guppies. Proceedings of the Royal Society of London Series B-Biological Sciences 271:2035-2042
Fedorka KM, Mousseau TA (2004) Female mating bias results in conflicting sex-specific offspring fitness. Nature 429:65-67
Feltmate BW, Williams DD (1991) Evaluation of predator-induced stress on field populations of stoneflies (Plecoptera). Ecology 72:1800-1806
Feltmate BW, Williams DD, Montgomerie A (1992) Relationship between diurnal activity patterns, cryptic coloration, and subsequent avoidance of predaceous fish by perlid stoneflies. Canadian Journal of Fisheries and Aquatic Sciences 49:2630-2634
Fetherston IA, Scott MP, Traniello JFA (1990) Parental Care in Burying Beetles - the Organization of Male and Female Brood-Care Behavior. Ethology 85:177-190
Fisher RA (1930) The Genetical Theory of Natural Selection. Clarendon Press, Oxford
Fisher RA (1958) The Genetical Theory of Natural Selection. Dover, New York
187.

Fitzgerald GJ (1992) Filial cannibalism in fishes - why do parents eat their offspring. Trends in Ecology & Evolution 7:7-10
Forrester GE, Chace JG, McCarthy W (1994) Diel and density-related change in food consumption and prey selection by Brook Charr in a New Hampshire stream. Environmental Biology of Fishes 39:301-311
Forsgren E (1997) Female sand gobies prefer good fathers over dominant males. Proceedings of the Royal Society of London Series B-Biological Sciences 264:1283-1286
Forsgren E, Karlsson A, Kvarnemo C (1996) Female sand gobies gain direct benefits by choosing males with eggs in their nests. Behavioral Ecology and Sociobiology 39:91-96
Fraser DF, Cerri RD (1982) Experimental evaluation of predator-prey relationships in a patchy environment - consequences for habitat use patterns in minnows. Ecology 63:307-313
Fraser S, Gotceitas V, Brown JA (1996) Interactions between age-classes of Atlantic cod and their distribution among bottom substrates. Canadian Journal of Fisheries and Aquatic Sciences 53:305-314
Freeland WJ (1981) Parasitism and behavioral dominance among male-mice. Science 213:461-462
Fretwell S, Lucas HL (1970) On territorial behavior and other factors influencing habitat distribution in birds. Acta Biotheoretica 19:16-36
Friberg U, Arnqvist G (2003) Fitness effects of female mate choice: preferred males are detrimental for Drosophila melanogasterierr\a\es. Journal of Evolutionary Biology 16:797-811
Fuller RC (2003) Disentangling female mate choice and male competition in the rainbow darter, Etheostoma caeruleum. Copeia 1:138-148
Fuller RC, Houle D, Travis J (2005) Sensory bias as an explanation for the evolution of mate preferences. American Naturalist 166:437-446
Gabor C (1999) Association patterns of sailfin mollies (Poecilia latipinna): alternative hypotheses. Behavioral Ecology and Sociobiology 46:333-340
Galbrait MG (1967) Size-selective predation on daphnia by rainbow trout and yellow perch. Transactions of the American Fisheries Society 96:1-&
Garant D, Dodson JD, Bernatchez L (2005) Offspring genetic diversity increases fitness of female Atlantic salmon (Salmo salar). Behavioral Ecology and Sociobiology 57:240-244
Gerhardt HC (1994) The evolution of vocalization in frogs and toads. Annual Review of Ecology and Systematics 25:293-324
Getty T (1998) Handicap signalling: when fecundity and viability do not add up. Animal Behaviour 56:127-130
Gibson RM, Bachman GC (1992) The costs of female choice in a lekking bird. Behavioral Ecology 3:300-309
Gilliam JF, Fraser DF (1987) Habitat selection under predation hazard - test of a model with foraging minnows. Ecology 68:1856-1862
188.

Godin JGJ, Briggs SE (1996) Female mate choice under predation risk in the guppy. Animal Behaviour 51:117-130
Goodwin NB, Balshine-Earn S, Reynolds JD (1998) Evolutionary transitions in parental care in cichlid fish. Proceedings of the Royal Society B: Biological Sciences 165
Gotceitas V, Brown JA (1993) Substrate selection by juvenile atlantic cod (gadus-morhua) - effects of predation risk. Oecologia 93:31-37
Gotceitas V, Fraser S, Brown JA (1995) Habitat use by juvenile atlantic cod (gadus-morhua) in the presence of an actively foraging and non-foraging predator. Marine Biology 123:421-430
Goto A (1993) Male mating success and female mate choice in the river sculpin, cottus-nozawae (Cottidae). Environmental Biology of Fishes 37:347-353
Goulet D (1998) Spawning success in the damselfish Amblyglyphidodon leucogaster: the influence of eggs in the nest. Animal Behaviour 55:651-664
Gowaty PA, Steinichen R, Anderson WW (2002) Mutual interest between the sexes and reproductive success in Drosophila pseudoobscura. Evolution 56:2537-2540
Graeb BDS, Dettmers JM, Wahl DH, Caceres CE (2004) Fish size and prey availability affect growth, survival, prey selection, and foraging behavior of larval yellow perch. Transactions of the American Fisheries Society 133:504-514
Graeb BDS, Mangan MT, Jolley JC, Wahl DH, Dettmers JM (2006) Ontogenetic changes in preypreference and foraging ability of yellow perch: Insights based on relative energetic return of prey. Transactions of the American Fisheries Society 135:1493-1498
Grafen A (1990) Biological Signals As Handicaps. Journal of Theoretical Biology 144:517-546
Gray DA, Cade WH (1999) Quantitative genetics of sexual selection in the field cricket, Gryllus integer. Evolution 53:848-854
Green BS, McCormick Ml (2005) 02 replenishment to fish nests: males adjust brood care to ambient conditions and brood development. Behavioral Ecology and Sociobiology 16:389-397
Griffth SC, Owens IPF, Burke T (1998) Sexual selection in the house sparrow: the best males are the worst fathers. (In preparation.)
Gross MR, Sargent RC (1985) The evolution of male and female parental care in fishes. American Zoologist 25:807-822
Grubb TC, Greenwald L (1982) Sparrows and a brushpile - foraging responses to different combinations of predation risk and energy-cost. Animal Behaviour 30:637-640
Guderley H, Couture P (2005) Stickleback fights: why do winners win? Influence of metabolic and morphometric parameters. Physiological and Biochemical Zoology 78:173-181
Gwynne DT (1991) Sexual competition among females: what causes courtship-role reversal? . Trends in Ecology & Evolution 6:118-121
Hagelin JC (2002) The kinds of traits involved in male-male competition: a comparison of plumage, behavior, and body size in quail. Behavioral Ecology 13:32-41
189.

Hale RE, St Mary CM, Lindström K (2003) Parental responses to changes in costs and benefits along an environmental gradient. Environmental Biology of Fishes 67:107-116
Hall DJ, Cooper WE, Werner EE (1970) An experimental approach to the production dynamics and structure of freshwater animal communities. Limnology and Oceanography 15:829-928
Hall DW, Kirkpatrick M, West B (2000) Runaway sexual selection when female preferences are directly selected. Evolution 54:1862-1869
Hanssen SA, Erikstad KE, Johnsen V, Bustnes JO (2003) Differential investment and costs duringavian incubation determined by individual quality: an experimental study of the common eider (Somateria mollissima). Proceedings of the Royal Society of London Series B-Biological Sciences 270:531-537
Harari AR, Handler AM, Landolt PJ (1999) Size-assortative mating, male choice and female choice in the curculionid beetle Diaprepes abbreviates. Animal Behaviour 58:1191-1200
Harshman LG, ProutT (1994) Sperm displacement without sperm transfer in Drosophila melanogasta. Evolution 48:758-766
Harter SL, Heck KL (2006) Growth rates of juvenile pinfish (Lagodon rhomboides): Effects of habitat and predation risk. Estuaries and Coasts 29:318-327
Hastings PA (1988) Female choice and male reproductive success in the angel blenny, coralliozetus- angelica (Teleostei, Chaenopsidae). Animal Behaviour 36:115-124
Heck KL, Hays G, Orth RJ (2003) Critical evaluation of the nursery role hypothesis for seagrass meadows. Marine Ecology-Progress Series 253:123-136
Hegde SN, Krishna MS (1997) Size-assortative mating in Drosophila malerkotliana. Animal Behaviour 54:419-426
Heywood JS (1989) Sexual selection by the handicap mechanism. Evolution 43:1387-1397
Hoelzer G (1988) Filial cannibalism in a non-brood cycling marine fish. Environmental Biology of Fishes 21:309-313
Hoelzer GA (1989) The good parent process of sexual selection. Animal Behaviour 38:1067-1078
Hoelzer GA (1992) The ecology and evolution of partial clutch cannibalism by paternal Cortez Damselfish. Oikos 65:113-120
Hoelzer GA (1995) Filial cannibalism and male parental care in damselfishes. Bulletin of Marine Science 57:663-671
Hoffman JI, Forcada J, Trathan PN, Amos W (2007) Female fur seals show active choice for males that are heterozygous and unrelated. Nature 445:912-914
Holland B, Rice WR (1998) Perspective: Chase-away sexual selection: Antagonistic seduction versus resistance. Evolution 52:1-7
Holland B, Rice WR (1999) Experimental removal of sexual selection reverses intersexual antagonistic coevolution and removes a reproductive load. Proceedings of the National Academy of Sciences of the United States of America 96:5083-5088
190.

Holomuzki JR (1986) Predator avoidance and diel patterns of microhabitat use by larval tiger salamanders. Ecology 67:737-748
Houde AE (1994) Effect of artificial selection on male colour patterns on mating preference of female guppies. Proceedings of the Royal Society B: Biological Sciences 256:125-130
Houde AE, Torio AJ (1992) Effect of Parasitic Infection on Male Color Pattern and Female Choice in Guppies. Behavioral Ecology 3:346-351
Houle D (1992) Comparing evolvability and variability of quantitative traits. Genetics 130:195-204
Houston AI, Szekely T, McNamara JM (2005) Conflict between parents over care. Trends in Ecology & Evolution 20:33-38
Houtman R, Dill LM (1994) The influence of substrate color on the alarm response of tidepool sculpins (oligocottus-maculosus, Pisces, Cottidae). Ethology 96:147-154
Howard DJ, Reece M, Gregory PG, Chu J, Cain ML (1998) The evolution of barriers to fertilization between closely related organisms. In: Howard DJ, Berlocher SH (eds) Endless forms: species and speciation. Oxford University Press, New York, pp 279-288
Hunt J, Brooks R, Jennions MD (2005) Female mate choice as a condition-dependent life-history trait. American Naturalist 166:78-92
Hunt J, Brooks R, Jennions MD, Smith MJ, Bentsen CL, Bussiere LF (2004) High-quality male field crickets invest heavily in sexual display but die young. Nature 432:1024-1027
Hyvarinen P, Huusko A (2006) Diet of brown trout in relation to variation in abundance and size of pelagic fish prey. Journal of Fish Biology 68:87-98
Irschick DJ, Garland TJ (2001) Integrating function and ecology in studies of adaptation: investigations of locomotor capacity as a model system. Annual Review of Ecology and Systematics 32:367- 396
Ivlev VS (1961) Experimental ecology of the feeding of fishes. Yale University Press, New Haven, Connecticut
Iwasa Y, Pomiankowski A (1999) Good parent and good genes models of handicap evolution. Journal of Theoretical Biology 200:97-109
Iwasa Y, Pomiankowski A, Nee S (1991) The evolution of costly mate preferences. II. The ‘handicap’ principle. Evolution 45:1431-1442
Iyengar VK, Reeve HK, Eisner T (2002) Paternal inheritance of a female moth’s mating preference: consequences for sexual selection. Nature 419:830-832
Jennions MD, Backwell PRY, Passmore Nl (1995) Repeatability of mate choice: the effect of size in the African painted reed frog, Hyperolius marmoratus. Animal Behaviour 49:181-186
Jennions MD, Petrie M (1997) Variation in mate choice and mating preferences : a review of causes and consequences. Biological Review 72:283-327
Jennions MD, Polakow DA (2001) The effect of partial brood loss on male desertion in a cichlid fish: an experimental test. Behavioral Ecology 12:84-92
191.

Jivoff P (1997) The relative roles of predation and sperm competition on the duration of the post-copulatory association between the sexes in the blue crab, Callinectes sapidus. Behavioral Ecology and Sociobiology 40:175-185
Johnstone RA, Reynolds JD, Deutsch JC (1996) Mutual mate choice and sex differences in choosiness. Evolution 50:1382-1391
Jones JC, Reynolds JD (1999) Costs of egg ventilation for male common gobies breeding in conditions of low dissolved oxygen. Animal Behaviour 57:181 -188
Jones TM, Quinnell RJ (2002) Testing predictions for the evolution of lekking in the sandfly, Lutzomyia longipalpis. Animal Behaviour 63:605-612
Kamler E (1992) Early life histories of fish: an energetics approach. Chapman & Hall, London
Karino K (1995) Male-male competition and female mate choice through courtship display in the territorial damselfish Stegastes nigricans. Ethology 100:126-138
Karino K (1997) Influence of brood size and offspring size on parental investment in a biparental cichlid fish, Neolamprologus moorii. Journal of Ethology 15:39-43
Karino K, Arai R (2006) Effect of clutch size on male egg-fanning behavior and hatching success in the goby, Eviota prasina (Klunzinger). Journal of Experimental Marine Biology and Ecology 334:43-50
Keller L, Reeve HK (1995) Why do females mate with multiple males - the sexually selected sperm hypothesis. Advances in the Study of Behavior 24:291 -315
Kelly JF (1996) Effects of substrate on prey use by belted kingfishers (Ceryle alcyon): A test of the prey abundance - Availability assumption. Canadian Journal of Zoology-Revue Canadienne De Zoologie 74:693-697
Kinne O, Kinne EM (1962) Rates of development in embryos of a cyprinodont fish exposed to different temperature salinity-oxygen combinations. Canadian Journal of Zoology 40:231-&
Kirkpatrick M (1985) Evolution of female choice and male parental investment in polygynous species: the demise of the “sexy son”. American Naturalist 125:788-810
Kirkpatrick M (1987) The evolutionary forces acting on female mating preferences in polygynous animals. In: Bradbury JW, Andersson MB (eds) Sexual selection: testing the alternatives. Wiley, Chichester, UK, pp 67-82
Kirkpatrick M (1996) Good genes and direct selection in evolution of mating preferences. Evolution 50:2125-2140
Klug H, Chin A, St Mary CM (2005) The net effects of guarding on egg survivorship in the flagfish, Jordanella floridae. Animal Behaviour 69:661-668
Knapp RA, Kovach JT (1991) Courtship as an honest indicator of male parental quality in the bicolor damselfish, Stegastes partitus. Behavioral Ecology 2:295-300
Knapp RA, Warner RR (1991) Male parental care and female choice in the bicolor damselfish, Stegastes partitus: bigger is not always better. Animal Behaviour 41:747-756
192.

Kodric-Brown A, Nicoletto P (2005) Courtship behaviour, swimming performance, and microhabitat use of Trinidadian guppies. Environmental Biology of Fishes 73:299-307
Kokko H (1998a) Good genes, old age and life-history trade-offs. Evolutionary Ecology 12:739-750
Kokko H (1998b) Should advertising parental care be honest? Proceedings of the Royal Society of London Series B-Biological Sciences 265:1871-1878
Kokko H (2001) Fisherian and "good genes" benefits of mate choice: how (not) to distinguish between them. Ecology Letters 4:322-326
Kokko H, Brooks R, Jennions MD, Morley J (2003) The evolution of mate choice and mating biases. Proceedings of the Royal Society of London Series B-Biological Sciences 270:653-664
Kokko H, Jennions M (2003) It takes two to tango. Trends in Ecology & Evolution 18:103-104
Kokko H, Jennions MD, Brooks R (2006) Unifying and testing models of sexual selection. Annual Review of Ecology Evolution and Systematics 37:43-66
Kokko H, Monaghan P (2001) Predicting the direction of sexual selection. Ecology Letters 4:159-165
Kolm N (2001) Females produce larger eggs for large males in a paternal mouthbrooding fish.Proceedings of the Royal Society of London Series B-Biological Sciences 268:2229-2234
Koref-Santibanez S (2001) Effects of age and experience on mating activity in the sibling species Drosophila pavani and Drosophila gaucha. Behavior Genetics 31:287-297
Kotiaho J, Alatalo RV, Mappes J, Parri S (1997) Fighting success in relation to body mass anddrumming activity in the male wolf spider Hygrolycosa rubrofasciata. Canadian Journal of Zoology-Revue Canadienne De Zoologie 75:1532-1535
Kotiaho JS (2001) Costs of sexual traits: a mismatch between theoretical considerations and empirical evidence. Biological Reviews 76:365-376
Kraak SBM (1996) Female preference and filial cannibalism in Aidablennius sphynx (Teleostei, Blenniidae); A combined field and laboratory study. Behavioural Processes 36:85-97
Kraak SBM, Groothuis TGG (1994) Female preference for nests with eggs is based on the presence of the eggs themselves. Behaviour 131:189-206
Kraak SBM, Videler JJ (1991) Mate choice in aidablennius-sphynx (teleostei, blenniidae) - females prefer nests containing more eggs. Behaviour 119:243-266
Krajewski JP, Bonaldo RM, Sazima C, Sazima I (2006) Foraging activity and behaviour of two goatfish species (Perciformes: Mullidae) at Fernando de Noronha Archipelago, tropical West Atlantic. Environmental Biology of Fishes 77:1-8
Kramer DL (1987) Dissolved oxygen and fish behavior. Environmental Biology of Fishes 18:81-92
Krebs JR (1980) Optimal foraging, predation risk and territory defense. Ardea 68:83-90
Krebs JR, Houston JA, Charnov EL (1981) Some recent developments in optimal forgaing. In: Kamil AC, Sargent TD (eds) Foraging Behaviour: Ecological, Ethological and Physiological Approaches. Garland STPM Press, New York, pp 3-17
193.

Kunzler R, Bakker TCM (2000) Pectoral fins and paternal quality in sticklebacks. Proceedings of the Royal Society of London Series B-Biological Sciences 267:999-1004
Kuwamura T, Yugo Y, Nakakshima Y (1993) Size-assortative monogamy and paternal egg care in a coral goby Paragobioon echinocephalus. Ethology 95:65-75
Kvarnemo C (1997) Food affects the potential reproductive rates of sand goby females, but not of males. Behavioral Ecology 8:605-611
Kvarnemo C, Ahnejsö I (1996) The dynamics of operational sex ratios and competition for mates. Trends in Ecology & Evolution 11:404-408
Kvarnemo C, Svensson O, Forsgren E (1998) Parental behaviour in relation to food availability in the common goby. Animal Behaviour 56:1285-1290
Lailvaux SP, Irschick DJ (2006) No evidence for female association with high-performance males in the green anole lizard, Anolis carolinensis. Ethology 112:707-715
Lande R (1980) Sexual dimorphism, sexual selection, and adaptation in polygenic characters. Evolution 34
Langmore NE, Davies NB, Hatchwell BJ, Hartley IR (1996) Female song attracts males in the alpine accentor Prunella collaris. Proceedings of the Royal Society of London Series B-Biological Sciences 263:141-146
Lavery RJ, Keenleyside MHA (1990) Parental investment of a biparental cichlid fish, cichlasoma-nigrofasciatum, in relation to brood size and past investment. Animal Behaviour 40:1128-1137
Leboeuf BJ, Mesnick S (1991) Sexual behavior of male Northern Elephant Seals 1. Lethal injuries to adult females. Behaviour 116:143-162
Leeren ED (1958) Observations on the growth of perch (perca-fluviatilis i) over 22 years with special reference to the effects of temperature and changes in population-density. Journal of Animal Ecology 27:287-334
Lessells CM (1999) Sexual conflicts in animals. In: Keller L (ed) Levels of Selection. Princeton University Press, Princeton, New Jersey, pp 75-99
Lifjeld JT, Dunn PO, Westneat DF (1994) Sexual selection by sperm competition in birds: male-male competition or female choice? Journal of Avian Biology 25:244-50
Lindholm JB, Auster PJ, Kaufman LS (1999) Habitat-mediated survivorship of juvenile (0-year) Atlantic cod Gadus morhua. Marine Ecology-Progress Series 180:247-255
Lindström K (1988) Male-male competition for nest sites in the sand goby, pomatoschistus-minutus. Oikos 53:67-73
Lindström K (1998) Effects of costs and benefits of brood care on filial cannibalism in the sand goby. Behavioral Ecology and Sociobiology 42:101-106
Lindstrom K, Sargent RC (1997) Food access, brood size and filial cannibalism in the fantail darter, Etheostoma flabellare. Behavioral Ecology and Sociobiology 40:107-110
Lindström K, Sargent RC (1997) Food access, brood size and filial cannibalism in the fantail darter, Etheostoma flabellare. Behavioral Ecology and Sociobiology 40:107-110
194.

Linville SU, Breitwisch R, Schilling AJ (1998) Plumage brightness as an indicator of parental care in northern cardinals. Animal Behaviour 55:119-127
Lissaker M, Kvarnemo C (2006) Ventilation or nest defense - parental care trade-offs in a fish with male care. Behavioral Ecology and Sociobiology 60:864-873
Lissaker M, Kvarnemo C, Svensson O (2003) Effects of a low oxygen environment on parental effort and filial cannibalism in the male sand goby, Pomatoschistus minutus. Behavioral Ecology 14:374-381
Lopez P, Munoz A, Martin J (2002) Symmetry, male dominance and female mate preferences in the Iberian rock lizard, Lacerta monticola. Behavioral Ecology and Sociobiology 52:342-347
Lopez S (1999) Parasitized female guppies do not prefer showy males. Animal Behaviour 57:1129- 1134
Luo JG, Brandt SB, Klebasko MJ (1996) Virtual reality of planktivores: A fish's perspective of prey size selection. Marine Ecology - Progresses Series 140:271-283
Macchiusi F, Baker RL (1991) PREY BEHAVIOR AND SIZE-SELECTIVE PREDATION BY FISH. Freshwater Biology 25:533-538
Mackereth RW, Noakes DLG, Ridgway MS (1999) Size-based variation in somatic energy reserves and parental expenditure by male smallmouth bass, Micropterus dolomieu. Environmental Biology of Fishes 56:263-275
Magnhagen C (1991) Predation risk as a cost of reproduction. Trends in Ecology & Evolution 6:183- 185
Manica A (2002) Filial cannibalism in teleost fish. Biological Reviews 77:261-277
Manica A (2003) The effect of brood size and age on partial filial cannibalism in the scissortail sergeant. Journal of Fish Biology 63:37-47
Manica A (2004) Parental fish change their cannibalistic behaviour in response to the cost-to-benefit ratio of parental care. Animal Behaviour 67:1015-1021
Mank JE, Promislow DEL, Avise JC (2005) Phylogenetic perspectives in the evolution of parental care in ray-finned fishes. Evolution 59:1570-1578
Marconato A, Bisazza A (1986) Males whose nests contain eggs are preferred by female cottus-gobio L (Pisces, Cottidae). Animal Behaviour 34:1580-1582
Markman S, Yomtov Y, Wright J (1995) Male parental care in the orange-tufted sunbird - behavioral- adjustments in provisioning and nest guarding effort. Animal Behaviour 50:655-669
Mayer CM, Rudstam LG, Mills EL, Cardiff SG, Bloom CA (2001) Zebra mussels (Dreissenapolymorpha), habitat alteration, and yellow perch (Perea flavescens) foraging: system-wide effects and behavioural mechanisms. Canadian Journal of Fisheries and Aquatic Sciences 58:2459-2467
Mazzi D (2004) Parasites make male pipefish careless. Journal of Evolutionary Biology 17:519-527
McLister JD (2003) The metabolic cost of amplexus in the gray tree frog (Hyla versicolor): assessing the energetics of male mating success. Canadian Journal of Zoology 81:388-394
195.

McNamara JM, Szekely T, Webb JN, Houston AI (2000) A dynamic game-theoretic model of parental care. Journal of Theoretical Biology 205:605-623
Merila J, Sheldon BC (2000) Lifetime reproductive success and heritability in nature. American Naturalist 155:301-310
Merilaita S (2003) Visual background complexity facilitates the evolution of camouflage. Evolution 57:1248-1254
Mihelakakis A, Kitajima C (1994) Effects of salinity and temperature on incubation period, hatching rate, and morphogenesis of the silver sea bream, Sparus sarba (Forskal, 1775). Aquaculture 126:361-371
Milinski M (1979) Evolutionarily stable feeding strategy in sticklebacks. Zeitschrift Fur Tierpsychologie- Journal of Comparative Ethology 51:36-40
Milinski M (1982) Optimal foraging - the influence of intraspecific competition on diet selection. Behavioral Ecology and Sociobiology 11:109-115
Milinski M, Heller R (1978) Influence Of A Predator On Optimal Foraging Behavior Of Sticklebacks (Gasterosteus-Aculeatus L). Nature 275:642-644
Mitchell DP, Dunn PO, Whittingham LA, Freeman-Gallant CR (2007) Attractive males provide less parental care in two populations of the common yellowthroat. Animal Behaviour 73:165-170
Mittelbach G (1986) Predator-mediated habitat use - some consequences for species interactions. Environmental Biology of Fishes 16:159-169
Mittelbach GG (1981) Patterns of invertebrate size and abundance in aquatic habitats. Canadian Journal of Zoology 38:896-904
Möller AP (1993) Female preference for apparently symmetrical male sexual ornaments in the barn swallow hirundo-rustica. Behavioral Ecology and Sociobiology 32:371-376
Möller AP, Alatalo RV (1999) Good-genes effect in sexual selection. Proceedings of the Royal Society of London Series B-Biological Sciences 266:85-91
Möller AP, Thornhill R (1998) Male parental care, differential parental investment by females and sexual selection. Animal Behaviour 55:1507-1515
Montgomerie RD, Weatherhead PJ (1988) Risks and rewards of nest defense by parent birds. Quarterly Review of Biology 63:167-187
Moore AJ, Gowaty PA, Wallin WG, Moore PJ (2001) Sexual conflict and the evolution of female mate choice and male social dominance. Proceedings of the Royal Society of London Series B- Biological Sciences 268:517-523
Moore AJ, Moore PJ (1999) Balancing sexual selection through opposing mate choice and male competition. Proceedings of the Royal Society of London Series B-Biological Sciences 266:711-716
Morrison C, Hero JM, Smith WP (2001) Mate selection in Litoria chloris and Litoria xanthomera: Females prefer smaller males. Austral Ecology 26:223-232
196.

Morrissey JF, Gruber SH (1993) Habitat selection by juvenile lemon sharks, negaprion brevirostris. Environmental Biology of Fishes 38:311-319
Mountjoy DJ, Lemon RE (1997) Male song complexity and parental care in the European starling. Behaviour 134:661-675
Munk P (1992) Foraging Behaviour and prey size spectra of larval herring Clupea harengus. Marine Ecology - Progresses Series 80:149-158
Neat FC, Taylor AC, Huntingford FA (1998) Proximate costs of fighting in male cichlid fish: the role of injuries and energy metabolism. Animal Behaviour 55:875-882
Neff BD (2003) Decisions about parental care in response to perceived paternity. Nature 422:716
Neff BD, Gross MR (2001) Dynamic adjustment of parental care in response to perceived paternity. Proceedings of the Royal Society of London Series B-Biological Sciences 268:1559-1565
Nelson WG, Bonsdorff E (1990) Fish predation and habitat complexity - are complexity thresholds real. Journal of Experimental Marine Biology and Ecology 141:183-194
Nicoletto PF (1995) Offspring quality and female choice in the guppy, Poecillia reticulata. Animal Behaviour 49:377-387
Nilsson SO, Nilsson GE (2000) Free choice by female sticklebacks: lack of preference for maledominance traits. Canadian Journal of Zoology-Revue Canadienne De Zoologie 78:1251-1258
Nyman KJ (1953) Observations on the behaviour of Gobius microps. Acta Societatis po Fauna et Flora Fennica 69:1-11
Okuda N, Ito S, Iwao H (2002) Female spawning strategy in Rhinogobius sp OR: how do females deposit their eggs in the nest? Ichthyological Research 49:371-379
Okuda N, Yanagisawa Y (1996) Filial cannibalism by mouthbrooding males of the cardinal fish,Apogon doederleini, in relation to their physical condition. Environmental Biology of Fishes 45:397-404
Oppenheimer JR (1970) Mouthbrooding in fishes. Animal Behaviour 52:307-314
Ormond RFG, Edwards AJ (1987) Red Sea fishes. In: Edwards AJ, Head SM (eds) Red Sea (key environments). Pergammon Press, Oxford, pp 22-44
Osenberg CW, Mittelbach GG (1989) Effects of body size on the predator prey interaction between pumpkinseed sunfish and gastropods. Ecological Monographs 59:405-432
Osorno JL, Szekely T (2004) Sexual conflict and parental care in magnificent frigatebirds: full compensation by deserted females. Animal Behaviour 68:337-342
Östlund-Nilsson S, Nilsson GE (2004) Breathing with a mouth full of eggs: respiratory consequences of mouthbrooding in cardinalfish. Proceedings of the Royal Society of London Series B- Biological Sciences 271:1015-1022
Östlund S, Ahnesjö I (1998) Female fifteen-spined sticklebacks prefer better fathers. Animal Behaviour 56:1177-1183
197.

Paloheim JE, Dickie LM (1966) Food and growth of fishes .3. relations among food body size and growth efficiency. Journal of the Fisheries Research Board of Canada 23:1209-&
Pampoulie C, Lindstrom K, St Mary CM (2004) Have your cake and eat it too: male sand gobies show more parental care in the presence of female partners. Behavioral Ecology 15:199-204
Parker GA (1974) Assessment strategy and the evolution of fighting behaviour. Journal of Theoretical Biology 47:223-243
Parker GA, Simmons LW (1996) Parental investment and the control of sexual selection: predicting the direction of sexual competition. Proceedings of the Royal Society of London Series B- Biological Sciences 263:315-321
Parker RR, Larkin PA (1959) A concept of growth in fishes. Journal of the Fisheries Research Board of Canada 16:721-745
Parker TH, Garant D (2004) Quantitative genetics of sexually dimorphic traits and capture of genetic variance by a sexually-selected condition-dependent ornament in red junglefowl (Gallus gallus). Journal of Evolutionary Biology 17:1277-1285
Parker TH, Ligon JD (2003) Female mating preferences in red junglefowl: a meta-analysis. Ethology Ecology & Evolution 15:63-72
Parri S, Alatalo RV, Mappes J (1998) Do female leaf beetles Galerucella nymphaeae choose their mates and does it matter? Oecologia 114:127-132
Partridge L, Fowler K (1990) Non-mating costs of exposure to males in female Drosophila melanogaster Journal of Insect Physiology 36:419-425
Payne AG, Smith C, Campbell AC (2002) Filial cannibalism improves survival and development of beaugregory damselfish embryos. Proceedings of the Royal Society of London Series B- Biological Sciences 269:2095-2102
Payne AG, Smith C, Campbell AC (2004) A model of oxygen-mediated filial cannibalism in fishes. Ecological Modelling 174:253-266
Perry G, Levering K, Girard I, Garland TJ (2004) Locomotor performance and dominance in male Anolis cristatellus. Animal Behaviour 67:37-47
Perry G, Pianka ER (1997) Animal foraging: past, present and future. Trends in Ecology & Evolution 12:360-364
Persson L, De Roos AM (2006) Food-dependent individual growth and population dynamics in fishes. Journal of Fish Biology 69:1-20
Peters A, Michiels NK (1996) Evidence for lack of inbreeding avoidance by selective simultaneous hermaphrodite. Invertebrate Biology 115:99-103
Petersen CW (1990) The occurrence and dynamics of clutch loss and filial cannibalism in twoCaribbean damselfishes. Journal of Experimental Marine Biology and Ecology 135:117-133
Petrie M (1983) Female moorhens compete for small fat males. Science 220:413-415
Pianka ER (1976) Natural selection of optimal reproductive tactics. American Zoologist 16:775-784
198.

Pilastro A, Evans JP, Sartorelli S, Bisazza A (2002) Male phenotype predicts insemination success in guppies. Proceedings of the Royal Society of London Series B-Biological Sciences 269:1325- 1330
Pitnick S, Markow TA (1994) Male gametic strategies - Sperm size, testes size, and the allocation of ejaculate among successive mates by the sperm-limited fly Drosophila-Pachea and its relatives. American Naturalist 143:785-819
Polak M, Starmer WT (1998) Parasite-induced risk of mortality elevates reproductive effort in male Drosophila. Proceedings of the Royal Society of London Series B-Biological Sciences 265:2197-2201
Polis GA (1981) The evolution and dynamics of intraspecific predation. Annual Review of Ecology and Systematics 12:225-251
Pomfret J, Knell R (2006) Sexual selection and horn allometry in the dung beetle Euoniticellus intermedius. Animal Behaviour 71:567-576
Pomiankowski A (1987) The costs of choice in sexual selection. Journal of Theoretical Biology 128:195-218
Pomiankowski A (1988) The evolution of female mating preferences for male genetic quality. Oxford Surveys in Evolutionary Biology 5:136-184
Pomiankowski A, Iwasa Y, Nee S (1991) The evolution of costly mate preferences. I. Fisher and biased mutation. Evolution 45:1422-1430
Pomiankowski A, Möller AP (1995 ) A resolution of the lek paradox. Proceedings of the Royal Society B: Biological Sciences 260:21-29
Poulin R (1994) Mate choice decisions by parasitized female upland bullies, Gobiomorphus breviceps. Proceedings of the Royal Society of London Series B-Biological Sciences 256:183-187
Prasad NG, Bedhomme S, Day T, Chippindale AK (2007) An evolutionary cost of separate genders revealed by male-limited evolution American Naturalist 169:29-37
Preston BT, Stevenson IR, Pemberton JM, Wilson K (2001) Dominant rams lose out by spermdepletion - A waning success in siring counters a ram's high score in competition for ewes. Nature 409:681-682
Price T, Schlüter D, Heckman NE (1993) Sexual selection when the female directly benefits. Biological Journal of the Linnean Society 48:187-211
Pyke GH, Pulliam HR, Charnov EL (1977) Optimal Foraging - Selective Review Of Theory And Tests. Quarterly Review of Biology 52:137-154
Queller DC (1997) Why do females care more than males? Proceedings of the Royal Society of London Series B-Biological Sciences 264:1555-1557
Qvarnström A (1997) Experimentally increased badge size increases male competition and reduces male parental care in the collared flycatcher. Proceedings of the Royal Society of London Series B-Biological Sciences 264:1225-1231
Qvarnstrom A, Forsgren E (1998) Should females prefer dominant males? Trends in Ecology & Evolution 13:498-501
199.

Raouf SA, Parker PG, Ketterson ED, Nolan V, Ziegenfus C (1997) Testosterone affects reproductive success by influencing extra-pair fertilizations in male dark-eyed juncos (Aves: Junco hyemalis). Proceedings of the Royal Society of London Series B-Biological Sciences 264:1599-1603
Reebs SG, Whoriskey FG, FitzGerald GJ (1984) Diel patterns of fanning activity, egg respiration, and the nocturnal behavior of male 3-spined sticklebacks, gasterosteus-aculeatus I (f-trachurus). Canadian Journal of Zoology-Revue Canadienne De Zoologie 62:329-334
Reichard M, Bryja J, Ondrackova M, Davidova M, Kaniewska P, Smith C (2005) Sexual selection for male dominance reduces opportunities for female mate choice in the European bitterling (Rhodeus sericeus). Molecular Ecology 14:1533-1542
Reynolds JD, Goodwin NB, Freckleton RP (2002) Evolutionary transitions in parental care and live bearing in vertebrates. Philosophical Transactions of the Royal Society of London Series B- Biological Sciences 357:269-281
Reynolds JD, Jones JC (1999) Female preference for preferred males is reversed under low oxygen conditions in the common goby (Pomatoschistus microps). Behavioral Ecology 10:149-154
Rice WR (1984) Sex chromosomes and the evolution of sexual dimorphism. Evolution 38:735-742
Rice WR, Chippindale AK (2001) Intersexual ontogenetic conflict. Journal of Evolutionary Biology 14:685-693
Rice WR, Holland B (1997) The enemies within: intergenomic conflict, interlocus contest evolution (ICE), and the intraspecific Red Queen. Behavioral Ecology and Sociobiology 41:1-10
Rintämaeki PT, Lundberg A, Alatalo RV, Höeglund J (1998) Assortative mating and female clutch investment in black grouse. Animal Behaviour 56:1399-1403
Robertson DR (1984) Cohabitation of competing territorial damselfishes on a Caribbean coral reef. Ecology 65:1121-1135
Robertson DR, Lässig B (1980) Spatial distribution patterns and distribution of a group of territorial damselfishes from the Great Barrier Reef. Bulletin of Marine Science 30:187-203
Robertson DR, Polunin NVC, Leighton K (1979) The behavioural ecology of three Indian Oceansurgeonfishes (Acanthurus linearus, A. leucosternon and Zebrasoma ‘scopas): their feeding strategies and social and mating systems. Environmental Biology of Fishes 4:125-170
Robson MA, Miles DB (2000) Locomotor performance and dominance in tree lizards, Urosaurus ornatus. Functional Ecology 14:338-344
Rodd FH, Hughes KA, Grether GF, Baril CT (2002) A possible non-sexual origin of mate preference: are male guppies mimicking fruit? Proceedings of the Royal Society of London Series B- Biological Sciences 269:475-481
Rohwer S (1978) Parent cannibalism of offspring and egg raiding as a courtship strategy. American Naturalist 112:429-440
Rowe L, Arnqvist G (1996) Analysis of the causal components of assortative mating in water striders. Behavioural Ecology and Sociobiology 38:279-286
200.

Rowe L, Houle D (1996) The lek paradox and the capture of genetic variance by condition dependent traits. Proceedings of the Royal Society B: Biological Sciences 263:1415-1421
Russo AR (1987) Role of habitat complexity in mediating predation by the gray damselfish abudefduf- sordidus on epiphytal amphipods. Marine Ecology-Progress Series 36:101-105
Ryan MJ, Rand AS (1990) The sensory basis of sexual selection for complex calls in the tungara frog, physalaemus-pustulosus (sexual selection for sensory exploitation). Evolution 44:305-314
Ryer CH, Stoner AW, Titgen RH (2004) Behavioral mechanisms underlying the refuge value of benthic habitat structure for two flatfishes with differing anti-predator strategies. Marine Ecology - Progresses Series 268:231-243
Sakaluk SK, Belwood JJ (1984) Gecko phonotaxis to cricket calling song: a case of satellite predation. Animal Behaviour 32:659-662
Sanz JJ, Kranenbarg S, Tinbergen JM (2000) Differential response by males and females tomanipulation of partner contribution in the great tit (Parus major). Journal of Animal Ecology 69:74-84
Sargent RC (1985) TErritoriality and reproductive tradeoffs in the 3-spine stickleback, gasterosteus- aculeatus. Behaviour 93:217-226
Sargent RC (1988) Paternal care and egg survival both increase with clutch size in the fathead minnow, pimephales-promelas. Behavioral Ecology and Sociobiology 23:33-37
Sargent RC (1992) Ecology of filial cannibalism in fish: theoretical perspectives. In: Elgar MAC, B. J.(ed) Cannibalism: ecology and evolution among diverse taxa. Oxford University Press, pp 38- 62
Sargent RC, Gebier JB (1980) Effects of nest site concealment on hatching success, reproductive success, and paternal behavior of the threespine stickleback, gasterosteus-aculeatus. Behavioral Ecology and Sociobiology 7:137-142
Sargent RC, Gross MR (1985) Parental investment decision rules and the concorde fallacy. Behavioral Ecology and Sociobiology 17:43-45
Sargent RC, Gross MR (1986) Williams' principle: An explanation of parental care in teleost fishes. In: Pitcher TJ (ed) The Behaviour of Teleost Fish. Crrom Helm, London, pp 275-293
Savino JF, Stein RA (1982) Predator-prey interaction between largemouth bass and bluegills as influenced by simulated, submersed vegetation. Transactions of the American Fisheries Society 111:255-266
Schaffer NM (1974a) Optimal reproductive effort in fluctuating environments. American Naturalist 108:783-798
Schaffer NM (1974b) Selection for optimal life histories: The effects of age structure. Ecology 55:291- 303
Scharf FS, Manderson JP, Fabrizio MC (2006) The effects of seafloor habitat complexity on survival of juvenile fishes: Species-specific interactions with structural refuge. Journal of Experimental Marine Biology and Ecology 335:167-176
201.

Schneider KJ (1984) Dominance, Predation, And Optimal Foraging In White-Throated Sparrow Flocks. Ecology 65:1820-1827
Schoener TW (1986) A brief history of optimal foraging theory. In: Kamil AC, Krebs JR, Pulliam HR (eds) Foraging Behaviour. Plenium Press
Schwagmeyer PL, Mock DW, Parker GA (2002) Biparental care in house sparrows: negotiation or sealed bid? Behavioral Ecology 13:713-721
Seko T, Nakasuji F (2004) Effect of egg size variation on survival rate, development and fecundity of offspring in a migrant skipper, Parnara guttata guttata (Lepidoptera : Hesperiidae). Applied Entomology and Zoology 39:171-176
Semler DE (1971) Some aspects of adaptation in a polymorphism for breeding colours in Threespine Stickleback (Gasterosteus-Aculeatus). Journal of Zoology 165:291-&
Sevenster P (1961) A causal analysis of a displacement activity (fanning in Gasterosteus aculeatus L.). Behaviour 9 (Suppl.)
Shackleton MA, Jennions MD, Hunt J (2005) Fighting success and attractiveness as predictors of male mating success in the black field cricket, Teleogryllus commodus: the effectiveness of no-choice tests. Behavioral Ecology and Sociobiology 58:1-8
Sheldon BC (2000) Differential allocation: tests, mechanisms and implications. Trends in Ecology and Evolution 15:397-402
Sih A (1980) Optimal Foraging - Partial Consumption Of Prey. American Naturalist 116:281-290
Sih A (1982) Optimal patch use - variation in selective pressure for efficient foraging. American Naturalist 120:666-685
Simmons LW (1992) Quantification of role reversal in relative parental investment in a bush cricket. Nature 358:61-63
Simmons LW, Parker GA (1989) Nuptial feeding in insects - mating effort versus paternal investment. Ethology 81:332-343
Skolbekken R, Utne-Palm AC (2001) Parental investment of male two-spotted goby, Gobiusculus flavescens (Fabricius). Journal of Experimental Marine Biology and Ecology 261:137-157
Slagsvold T, Lifjeld JT (1988) Plumage color and sexual selection in the pied flycatcher ficedula- hypoleuca. Animal Behaviour 36:395-407
Sloman KA, Motherwell G, O’Connor Kl, Taylor AC (2000) The effect of social stress on the standard metabolic rate (SMR) of brown trout, Salmo trutta. Fish Physiology and Biochemistry 23:49-53
Smiseth PT, Ornborg J, Andersson S, Amundsen T (2001) Is male plumage reflectance correlated with paternal care in bluethroats? Behavioral Ecology 12:164-170
Smith C, Wootton RJ (1995) THE COSTS OF PARENTAL CARE IN TELEOST FISHES. Reviews in Fish Biology and Fisheries 5:7-22
Soler J J, de Neve L, Martinez JG, Soler M (2001) Nest size affects clutch size and the start of incubation in magpies: an experimental study. Behavioral Ecology 12:301-307
202.

Spence R, Smith C (2006) Mating preference of female zebrafish, Danio rerio, in relation to male dominance. Behavioral Ecology 17:779-783
Spoor WA (1984) Oxygen requirements of larvae of the smallmouth bass, Micropterus dolomieui Lacepede. Journal of Fish Biology 25:587-592
St Mary CM, Noureddine CG, Lindström K (2001) Environmental effects on male reproductive success and parental care in the Florida flagfish Jordanella floridae. Ethology 107:1035-1052
Stanley TR (2002) How many kilojoules does a Black-billed Magpie nest cost? Journal of Field Ornithology 73:292-297
Stein RA (1977) Selective Predation, Optimal Foraging, And Predator-Prey Interaction Between Fish And Crayfish. Ecology 58:1237-1253
Stein RA, Magnuson JJ (1976) BEHAVIORAL RESPONSE OF CRAYFISH TO A FISH PREDATOR. Ecology 57:751-761
Stutt AD, Siva-Jothy MT (2001) Traumatic insemination and sexual conflict in the bed bug Cimexlectularius. Proceedings of the National Academy of Sciences of the United States of America 98:5683-5687
Suk HY, Choe JC (2002) The presence of eggs in the nest and female choice in common freshwater gobies (Rhinogobius brunneus). Behavioral Ecology and Sociobiology 52:211-215
Summers K, Weigt LA, Boag P, Bermingham E (1999) The evolution of female parental care in poison frogs of the genus Dendrobates: evidence form mitochondrial DNA. Herpetologica 55:254-270
Svensson PA, Pelabon C, Blount JD, Surai PF, Amundsen T (2006) Does female nuptial coloration reflect egg carotenoids and clutch quality in the Two-Spotted Goby (Gobiusculus flavescens, Gobiidae)? Functional Ecology 20:689-698
Syriatowicz A, Brooks R (2004) Sexual responsiveness is condition-dependent in female guppies, but preference functions are not. BMC Ecology 4
Takahashi D, Kohda M (2004) Courtship in fast water currents by a male stream goby (Rhinogobius brunneus) communicates the parental quality honestly. Behavioral Ecology and Sociobiology 55:431-438
Takegaki T, Nakazono A (1999) Responses of the egg-tending gobiid fish Valenciennes longipinnis to the fluctuation of dissolved oxygen in the burrow. Bulletin of Marine Science 65:815-823
Tallamy DW (2000) Sexual selection and the evolution of exclusive paternal care in arthropods. Animal Behaviour 60:559-567
Tatrai I, Herzig A (1995) Effect of habitat structure on the feeding efficiency of young stages of razor fish (pelecus-cultratus (I)) - an experimental approach. Hydrobiologia 299:75-81
Taylor PD, Williams GC (1982) The lek paradox is not resolved. Theoretical Population Biology 22:392-409
Thresher RE (1984) Reproduction in reef fishes. T. F. H. Publications, Neptuen City, New Jersey
Tikkanen P, Huhta A, Muotka T (2000) Determinants of substrate selection in lotic mayfly larvae: is cryptic coloration important? Archiv Fur Hydrobiologie 148:45-57
203.

Tilson RL (1981) Family formation strategies of Kloss’s gibbons. Folia Primatologica 35:259-281
Tinbergen N (1951) The Study of Instinct. Oxford University Press, London
Tokarz RR (2002) An experimental test of the importance of the dewlap in male mating success in the lizard Anolis sagrei. Herpetologica 58:87-94
Tomkins JL, Radwan J, Kotiaho JS, Tregenza T (2004) Genic capture and resolving the lek paradox. Trends in Ecology & Evolution 19:323-328
Torricelli P, Lugli M, Gandolfi G (1985) A Quantitative-analysis of the fanning activity in the male padogobius martensi (Pisces, Gobiidae). Behaviour 92:288-301
Tregenza T, Wedell N (1997) Definitive evidence for cuticular pheromones in a cricket. Animal Behaviour 54:979-984
Trivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Sexual Selection and the Descent of man. Aldine Publishing Company, London, pp 136-179
Tupper M, Boutilier RG (1995) Effects of habitat on settlement, growth, and postsettlement survival of atlantic cod (Gadus-Morhua). Canadian Journal of Fisheries and Aquatic Sciences 52:1834- 1841
Unger LM, Sargent RC (1988) Allopaternal care in the fathead minnow, Pimephales promelas - Females prefer males with eggs. Behavioral Ecology and Sociobiology 23:27-32
Vahed K (1998) The function of nuptial feeding in insects: a review of empirical studies. Biological Reviews 73:43-78
van den Berghe EP, Wernerus F, Warner RR (1989) Female choice and the mating cost of peripheral males. Animal Behaviour 38:875-884
van lersel JJA (1953) An analysis of parental behaviour of the male three-spined stickleback (Gasterosteus aculeatus L.). Behaviour 3 (Suppl.):1-159
Vincent A, Ahnejsö I, Berglund A, Rosenqvist G (1992) Pipefishes and seahorses: are they all sex role reversed? Trends in Ecology & Evolution 7:237-241
Vinyoles D, Cöte IM, de Sostoa A (1999) Egg cannibalism in river blennies: the role of natural prey availability. Journal of Fish Biology 55:1223-1232
Wagner WEJ (1998) Measuring female mating preferences. Animal Behaviour 55:1029-1042
Watson PJ, Arnqvist G, Stallmann RR (1998) Sexual conflict and the energetic costs of mating and mate choice in water striders. American Naturalist 151:46-58
Webb JN, Houston AI, McNamara JM, Szekely T (1999) Multiple patterns of parental care. Animal Behaviour 58:983-993
Webster MM, Hart PJB (2004) Substrate discrimination and preference in foraging fish. Animal Behaviour 68:1071-1077
Wedell N, Kvarnemo C, Lessells CM, Tregenza T (2006) Sexual conflict and life histories. Animal Behaviour 71:999-1011
204.

Werner EE, Gilliam JF, Hall DJ, Mittelbach GG (1983) An experimental test of the effects of predation risk on habitat use in fish. Ecology 64:1540-1548
Werner EE, Hall DJ (1974) Optimal Foraging and Size Selection of Prey by Bluegill Sunfish (Lepomis macrochirus). Ecology 55:1042-1052
Werner EE, Mittelbach GG (1981) Optimal Foraging - Field-Tests Of Diet Choice And Habitat Switching. American Zoologist 21:813-829
West-Eberhard MJ (1984) Sexual selection, competitive communication and species-specific signals in insects. In: Lewis T (ed) Insect Communication Academic Press, London, pp 283-324
Westneat DF, Sherman PW, Morton ML (1990) The ecology and evolution of extra-pair copulations in birds. Current Ornithology 7:331-369
Weygoldt P (1987) Evolution of parental care in dart poison frogs (Amphibia: Anura: Dendrobatidae). Z. Zool. Syst. Evol. 25:51-67
White DJ, King AP, West MJ (2002) Facultative development of courtship and communication in juvenile male cowbirds (Molothrus ater). Behavioral Ecology 13:487-496
Whittingham LA, Dunn PO, Robertson RJ (1994) Female response to reduced male parental care in birds - an experiment in tree swallows. Ethology 96:260-269
Wiley RH, Poston J (1996) Indirect mate choice, competition for mates, and coevolution of the sexes. Evolution 50:1371-1381
Williams GC (1966a) Adaptation and natural selection. Princeton University Press, Princeton
Williams GC (1966b) Natural selection costs of reproduction and a refinement of Lacks principle. American Naturalist 100:687-&
Winkler DW (1987) A general model for parental care. American Naturalist 130:526-543
Wong BBM (2004) Superior fighters make mediocre fathers in the Pacific blue-eye fish. Animal Behaviour 67:583-590
Wootton RJ (1998) Ecology of teleost fishes. Kluwer, Dordrecht
Wright J, Cuthill I (1989) Manipulation of sex-differences in parental care. Behavioral Ecology and Sociobiology 25:171 -181
Yenisetti SC, Hegde SN (2003) Size-related mating and reproductive success in a drosophilid: Phorticella striata. Zoological Studies 42:203-210
Zahavi A (1975) Mate selection: a selection for a handicap. Journal of Theoretical Biology 53:205-214
Zink AG (2003) Quantifying the costs and benefits of parental care in female treehoppers. Behavioral Ecology 14:687-693
Zoran MJ, Ward JA (1983) Parental egg care behavior and fanning activity for the orange chromide, Etroplus maculatus. Environmental Biology of Fishes 8:301-310
205.
















![Błąd wychowawczy a stosowana dyrektywność rodzicielska [The parental mistake and applied parental control]](https://static.fdokumen.com/doc/165x107/6331b765f008040551041fa0/blad-wychowawczy-a-stosowana-dyrektywnosc-rodzicielska-the-parental-mistake.jpg)



