
Marginal zone B cell enrichment and strong follicular B cell reduction correlate with a delayed IgG...
12
Marginal zone B cell enrichment and strong follicular B cell reduction correlate with a delayed IgG response in a light chain diversity restricted mouse model Yacine M. Amrani, Danielle Voegtle ´, Eliane Barbier, Adrien Six and Pierre-Andre ´ Cazenave Unite ´ d’Immunophysiopathologie Infectieuse, Institut Pasteur, CNRS URA-1961, Universite ´ Pierre et Marie Curie, Paris-6, Paris, France Recently developed B6.j – k SEG mice (by crossing j – and C57BL/6 mice congenic for the wild Mus spretus SEG strain k locus lacking genes coding for k1 and k3) have a very reduced light chain diversity. B6.j – k SEG mice produce only k2 and kx light chains. Regardless of their Igh haplotype, B6.j – k SEG mice show a restricted B cell distribution by light chain subtype with kx dominance in all peripheral compartments except peritoneal cavity where k2 is dominant. This distribution suggests that selection mechanisms act differently in different B cell compartments on k2 and kx bearing B cells. Sequence analysis before or following immunization did not reveal unusual mechanisms of diversification. B6.j – k SEG mice still respond to various challenging antigens using new Ab patterns. In particular, regardless of Igh a or Igh b haplotypes, the anti-2,4-dinitrophenyl response is characterized by a restricted diversity for both heavy and light chains and a delayed IgG response when compared to B6 and B6.j – mice. We suggest that the delayed IgG response is due to the expansion of marginal zone B cells whereas follicular B cells are strongly reduced. Key words: B lymphocytes / Antibodies / Repertoire / Junctional diversity / Lambda light chains 1 Introduction To study the plasticity of the immune system and its adaptation to some drastic situations of genetic restric- tion of Ab diversity, we’ve characterized a new mouse model (B6.j – k SEG ) presenting an extremely reduced Ab light chain (LC) repertoire. B6.j – k SEG mice are derived from j – mice deficient for j LC expression, representing 95% of mice Ab repertoire, and B6.k SEG , a C57BL/6 (B6) mouse congenic for the Mus spretus SEG/Pas strain k locus [1–3]. This locus is characterized by the absence of the Vk1-Jk3-Ck3-Jk1-Ck1 recombination unit found in laboratory mice. B6.j – k SEG mice produce only k2 and kx and lack k1 and k3 LC [1, 4]. In laboratory mice, k1 represents 60–70% of the peripheral k repertoire at the Ab and B cell levels with the exception of the peritoneal cavity (PeC) where k2 and k3 are dominant among k LC in normal or j – mice, due to a selective process in that compartment [4]. kx represents 8–16% of the k repertoire in the different peripheral organs [2, 4]. Interestingly, j – mice produce normal numbers of PeC B cells in spite of j LC absence, meaning that k B cells are fully able to expand and fill the PeC. In the spleen, B cells are two-to-threefold reduced [4]. These compartments are expansion sites of different B cell subsets. B1 cells expand in the PeC and are long-lived cells with a fetal and neonatal origin [5]. Follicular (FO) B cells, or conventional B2 cells, expand in the spleen and are CD23 + -CD21 + . They are essentially produced during adult life and are responsible for the adaptive humoral immune response. A subset resembling B1 cells represents a small part of spleen B cells, concentrates in the marginal zone (MZ), close to the metallophilic macrophages, is CD23 low/– - CD21 high and is called MZ B cell. It is assumed to be a long-lived B cell subset deriving from early mouse life as suggested by recent data showing only B1 and MZ B cells presence in models of arrested B cell development shortly after birth. It lacks N sequence additions and is strongly diminished in VH81X/TdT double transgenic mice but present in VH81X mice [6–9]. However, [DOI 10.1002/eji.200324396] Received 23/7/03 Revised 19/7/04 Accepted 21/7/04 Abbreviations: B6: C57BL/6 PeC: Peritoneal cavity LC: Ig light chain HC: Ig heavy chain NP: (4-hydroxy-3-nitrophe- nyl)acetyl PC: Phosphorylcholine DNP: 2,4-dinitrophenyl MBP: Myelin basic protein IEF: Isoelectric focusing assay MZ: Marginal zone FO: Follicular 2812 Y. M. Amrani et al. Eur. J. Immunol. 2004. 34: 2812–2823 f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
-
Upload
sorbonne-fr -
Category
Documents
-
view
0 -
download
0
Transcript of Marginal zone B cell enrichment and strong follicular B cell reduction correlate with a delayed IgG...

Marginal zone B cell enrichment and strong follicularB cell reduction correlate with a delayed IgGresponse in a light chain diversity restricted mousemodel
Yacine M. Amrani, Danielle Voegtle, Eliane Barbier, Adrien Six and
Pierre-Andre Cazenave
Unite d’Immunophysiopathologie Infectieuse, Institut Pasteur, CNRS URA-1961, Universite Pierreet Marie Curie, Paris-6, Paris, France
Recently developed B6.j–kSEG mice (by crossing j– and C57BL/6 mice congenic for the wild
Mus spretus SEG strain k locus lacking genes coding for k1 and k3) have a very reduced light
chain diversity. B6.j–kSEG mice produce only k2 and kx light chains. Regardless of their Igh
haplotype, B6.j–kSEG mice show a restricted B cell distribution by light chain subtype with kxdominance in all peripheral compartments except peritoneal cavity where k2 is dominant.
This distribution suggests that selection mechanisms act differently in different B cell
compartments on k2 and kx bearing B cells. Sequence analysis before or following
immunization did not reveal unusual mechanisms of diversification. B6.j–kSEG mice still
respond to various challenging antigens using new Ab patterns. In particular, regardless of
Igha or Ighb haplotypes, the anti-2,4-dinitrophenyl response is characterized by a restricted
diversity for both heavy and light chains and a delayed IgG response when compared to B6
and B6.j– mice. We suggest that the delayed IgG response is due to the expansion of
marginal zone B cells whereas follicular B cells are strongly reduced.
Key words: B lymphocytes / Antibodies / Repertoire / Junctional diversity / Lambda lightchains
1 Introduction
To study the plasticity of the immune system and its
adaptation to some drastic situations of genetic restric-
tion of Ab diversity, we’ve characterized a new mouse
model (B6.j–kSEG) presenting an extremely reduced Ab
light chain (LC) repertoire. B6.j–kSEG mice are derived
from j– mice deficient for j LC expression, representing
95% of mice Ab repertoire, and B6.kSEG, a C57BL/6 (B6)
mouse congenic for the Mus spretus SEG/Pas strain klocus [1–3]. This locus is characterized by the absence of
the Vk1-Jk3-Ck3-Jk1-Ck1 recombination unit found in
laboratory mice. B6.j–kSEG mice produce only k2 and kxand lack k1 and k3 LC [1, 4]. In laboratory mice, k1represents 60–70% of the peripheral k repertoire at the
Ab and B cell levels with the exception of the peritoneal
cavity (PeC) where k2 and k3 are dominant among k LC in
normal or j– mice, due to a selective process in that
compartment [4]. kx represents 8–16% of the k repertoire
in the different peripheral organs [2, 4].
Interestingly, j– mice produce normal numbers of PeC B
cells in spite of j LC absence, meaning that k B cells are
fully able to expand and fill the PeC. In the spleen, B cells
are two-to-threefold reduced [4]. These compartments
are expansion sites of different B cell subsets. B1 cells
expand in the PeC and are long-lived cells with a fetal and
neonatal origin [5]. Follicular (FO) B cells, or conventional
B2 cells, expand in the spleen and are CD23+-CD21+.
They are essentially produced during adult life and are
responsible for the adaptive humoral immune response.
A subset resembling B1 cells represents a small part of
spleen B cells, concentrates in the marginal zone (MZ),
close to the metallophilic macrophages, is CD23low/–-
CD21high and is called MZ B cell. It is assumed to be a
long-lived B cell subset deriving from early mouse life as
suggested by recent data showing only B1 and MZ B
cells presence in models of arrested B cell development
shortly after birth. It lacks N sequence additions and is
strongly diminished in VH81X/TdT double transgenic
mice but present in VH81X mice [6–9]. However,
[DOI 10.1002/eji.200324396]
Received 23/7/03Revised 19/7/04Accepted 21/7/04
Abbreviations: B6: C57BL/6 PeC: Peritoneal cavity LC: Iglight chain HC: Ig heavy chain NP: (4-hydroxy-3-nitrophe-nyl)acetyl PC: Phosphorylcholine DNP: 2,4-dinitrophenylMBP: Myelin basic protein IEF: Isoelectric focusing assayMZ: Marginal zone FO: Follicular
2812 Y. M. Amrani et al. Eur. J. Immunol. 2004. 34: 2812–2823
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

experiments in rats where MZ B cells are an important
part of the spleen B cells (up to 25%) suggest that MZ B
cells originate from recirculating B cells, found only after
the 2nd week of life [10]. This period correlates with the
spleen architecture organization that allows a histological
difference betweenMZ and FOB cells [11, 12]. MZB cells
respond very quickly to stimuli and are the first barrier
against blood-borne Ag, participating, like B1 cells, to the
natural humoral immune response and secreting pre-
ferentially IgM Ab [12]. However, it isn’t known whether
MZB cells are enriched in kB cells like it is the case for B1
cells [2, 13].
B6.j–kSEG (Ighb) and B6.j–kSEG Igha congenic mice are
characterized by the presence of only k2 and kx LC
associated to a complete heavy chain (HC) repertoire
to form Ab molecules. Such situation renders the study
of genetic as well as potential and real repertoires
easier. It allows to investigate for the existence of
unusual LC diversification mechanisms such as gene
conversion, insertion/deletion events [14, 15], increased
junctional diversity by nucleotide deletions or non-
template base pair additions or increased somatic
mutations [14, 16].
In this study, we show that our model confirms the
selective process that occurs in PeC compartment where
k2 B cells are dominant in all B cell lineages. Interestingly,
our model shows a clear dominance of kx cells in all the
other peripheral compartments. However, k2 B cells
contribute more to the immune response after 2,4-
dinitrophenyl (DNP) immunization as suggested by
hybridoma analysis that shows an oligoclonal response
and a restricted CDR3 size for both HC and LC. B6 or
BC8 (B6.Igha congenic) and B6.j–kSEG or B6.j–kSEG Igha
mice immune response against different Ag reveals that
the genetic restriction pushes the immune system toward
the use of novel Ab HC and LC combinations. Strikingly,
this original Ab response isn’t contingent upon increased
or new diversification mechanisms. Finally, we demon-
strate a delayed and diversity-restricted IgG response
following immunization concomitant with an increased
proportion of MZ B cells and a strongly reduced number
of FO B cells. We discuss our findings regarding the
contribution of k LC expressing B cells to the MZ B cell
repertoire and their role during immune responses.
2 Results and discussion
2.1 Expansion of k2 B cells in PeC contrasts withkx B cells dominance in other compartments
By FACS, we analyzed Ig LC expression in the spleen,
mesenteric LN, BM, PBL and PeC of B6, B6.j–kSEG and
B6.j–kSEG Igha 4-month-old mice. All compartments but
PeC show a similar profile of LC sub-type distribution. We
have previously shown a twofold increase of the k/j ratio
in B6 mice PeC, due to a fourfold k2–3 population
increase [2]. k1 and kx proportions decrease in
comparison to k2–3 but are similar to proportions
observed in other compartments (Fig. 1A, B). We
previously described this general picture in j– mice.
Absence of j chains in B6.j– mice doesn’t change kisotype proportions. k1 is dominant in all compartments
but PeC. It is noteworthy that k2–3 and kx populations
are represented equally, except for PeC where k2–3 are
dominant ([2] and data not shown).
As shown previously [1], B6.j–kSEG and B6.j–kSEG Igha
mouse B cells produce only k2 and kx LC recognized by
the 189B3 Ab specific for the shared Ck2 region (Fig. 1A).
Strikingly, the Vk distribution doesn’t differ between
B6.j–kSEG and B6.j–kSEG Igha strains, indicating that the
Igh haplotype doesn’t have a detectable impact on Vk2and Vkx selection. Our results, however, demonstrate a
strict compartmentalized LC distribution. kx cells are
dominant in all compartments representing around 70%
of the B cells with the exception of PeC, where k2dominates (Fig. 1A, B). This result shows that k2 cells are
preferentially selected in PeC as we already described in
j– mice [4]. Additional data strengthen this finding: (i) in
B6.j–kSEG, kx but not k2 fills the gap due to the absence
of k1 and k3 populations in all compartments studied
except PeC; (ii) kx is dominant in BM immature B cells
(B220low-kx/B220low-k2 and data not shown). This last
observation is quite surprising since kx has a “natural”
disadvantage to make a productive rearrangement when
compared to k2 because of a stop codon at the end of the
Vkx germ-line sequence [1, 17]. Additionally, k2 has the
highest probability to perform a productive rearrange-
ment among k chains [18]. Thus, whereas a positive
selective process operates on k2 in PeC, k2 appears
negatively selected in the BM, reflecting the situation in
the peripheral compartments except PeC. Finally, it’s
noteworthy that, whereas the k2 and kx relative
distribution is similar in B6 and B6.j–, k1 and k3 absence
in B6.j–kSEG leads to a different selection of the
remaining k2 and kx (Fig. 1 and data not shown).
2.2 CD5+ B cells in PeC cannot solely account fork2 B cells expansion
To investigate on the reasons leading to an inverted ratio
between k2 and kx in PeC versus other peripheral
compartments, we analyzed B2, B1a and B1b lineage
distribution in B6, B6.j–kSEG and B6.j–kSEG Igha strains.
B2 cells are IgMlow-Mac-1–-CD5–. B1 cells are IgMhigh-
Mac-1+ and can be differentiated as B1a or B1b cells
Eur. J. Immunol. 2004. 34: 2812–2823 MZ B cell expansion in B6.j–kSEG mice 2813
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

through CD5 expression. B1a cells are CD5+ and B1b are
CD5– [5].
B6, B6.j–kSEG and B6.j–kSEG Igha strains don’t exhibit
striking differences in their B2/B1 or B2/B1a/B1b
subpopulation ratios. In all strains, B1a cells dominate
the PeC (Fig. 2A–C). Both B1 and B2 populations account
for the j/k ratio reduction in B6 PeC. k2-k3 increase is
mainly due to B1a cells (Fig. 2A). Similarly, k2 cells are
dominant in B6.j–kSEG and B6.j–kSEG Igha mice,
regardless of the lineage, but more obviously in B1a
cells (Fig. 2B, C). Consequently, our data suggest that
PeC environment rather than intrinsic properties of B1-
cells are responsible for the observed k2/kx ratio
inversion. The same conclusions were drawn for the
k2-k3 increased distribution in B6.j– mice PeC after fetal
liver or adult BM cells transfer into RAG2- mice [4],
strengthening the idea that peritoneal cells are more
prone to selective pressures [19].
2.3 No unusual mechanisms of Igk diversificationare detected in B6.j–kSEG mouse
The analysis of a spleen library of 89 k transcripts
obtained by anchored-PCR doesn’t reveal unusual
mechanisms of diversification (Fig. 3 and data not
shown). kx and k2 LC transcripts still present a restricted
junctional diversity as shown by the dominance of
canonical rearrangements presumably resulting from
short homology recombination mechanism: 17/24 for kxand 47/55 for k2 [18]. Only one P nucleotide was found
and no N addition or nucleotide deletion could be clearly
attributed (Fig. 3).
Fig. 1. B cell distribution by LC isotypes and organs. (A) B6, B6.j–kSEG and B6.j–kSEG Igha mouse spleen and PeC cells were
analyzed for B cells (IgM+) distribution by LC expression with anti-j/k1/k2-3-x and kx Ab. Percentage of cells in the corresponding
gates are indicated. (B) Histograms represent BM, spleen, LN, PBL and PeC j/k1/k2–3 and kx B cell (B220+-CD19+) distribution in
B6 mice (B6); k1/k2–3 and kx B cells in B6 k compartment [B6(k)]; k2 and kx B cells in B6.j–kSEG Igha and B6.j–kSEG mice,
respectively. j, k1 and kx populations are determined directly, k2–3 or k2 populations correspond to k2-3-x or k2-x populations
minus kx population in the histograms. Bars represent standard deviations from results obtained with four 4-month-old animals
per group (B220+-IgM+ staining gives similar results). (A) and (B) are two different experiments.
2814 Y. M. Amrani et al. Eur. J. Immunol. 2004. 34: 2812–2823
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

Therefore, the germinal repertoire reduction isn’t com-
pensated by an increase of k LC junctional diversity. We
didn’t detect clones in the periphery showing N addition
that would come from developing B cells overcoming the
TdT-regulated expression [5], or that would have rear-
ranged their k LC at the pro-B cell stage as observed in
mu-MT and TdT-transgenic mouse pro-B cells [16]. Our
data thus confirm the tight regulation of the TdT enzyme
during B cell development and outline the importance of
the mechanistic constraints during k repertoire establish-
ment [5, 20].
It is noteworthy that no nucleotide deletion is detected.
This observation could be related to a negative selection
of B cells bearing k LC presenting non-canonical CDR3
or to the presence of GC repeats at both Vk2-Vkx and
Jk2 sequences, which have been shown to hamper
nucleotide deletion events [21] (see Fig. 3 for Vk2-Vkxand Jk2 gene segment joining sequences).
Analysis of Vk sequences neither show nucleotide
deletions/insertions, another diversification mechanism
found in HC [15], nor an enhanced rate of somatic
mutations (0.32%). Finally, gene conversion events, that
occur in animals presenting low combinatory diversity
[14], were not detected despite sufficient identity
between Vk2 and Vkx (50%) (data not shown).
Our data thus show that B6.j–kSEG mice naive repertoire
isn’t subject to the increase or the induction of
diversification mechanisms known in vertebrates.
2.4 Expression of a new Ab pattern followingimmunization of B6.j–kSEG mouse
B6.j–kSEG Ab repertoire was compared to either BC8
or B6 repertoires following myelin basic protein (MBP),
Phosphorylcholine-thyroglobulin (PC-Tg) and DNP-OVA
Fig. 2. Analysis of k B cell subsets distribution in PeC. B cells from B6 (A), B6.j–kSEG (B) and B6.j–kSEG Igha (C) mice were divided
into IgM+-Mac-1+ B1 and IgM+-Mac-1– B2 cells or into IgM+-CD5+ B1a, IgMhigh-CD5– B1b and IgMlow-CD5– B2-cells. LC
distribution was determined for all three lineages. Percentage of cells in the corresponding gates or squares are indicated. Data
represent four different experiments. Since both B6.j–kSEG only express kx or k2 LC, kx/B220 double-staining is sufficient to
determine kx and k2 B cell percentages.
Eur. J. Immunol. 2004. 34: 2812–2823 MZ B cell expansion in B6.j–kSEG mice 2815
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
T-dependent immunizations or (4-hydroxy-3-nitro-
phenyl)acetyl-ficoll (NP-ficoll) T-independent immuniza-
tion.
In most laboratory mice, j LC associate with the VHT15
HC and dominate the anti-PC primary response [22, 23].
During the secondary response, another set of Ab
directed against the phenyl bridge between PC and
the carrier protein, using j and k1, appears [23–25]. The
BC8 anti-PC profile in Fig. 4A summarizes this canonical
response. B6.j–kSEG, still respond to PC by using k2 LC
only that don’t associate with VHT15 (Fig. 4A, PC and
data not shown). Surprisingly, k2 LC were also detected
at low titers in BC8 sera.
BC8 and normal mice DNP response, is formed
essentially by Ab bearing j and to a lesser extent k1,k2 and k3 but not kx LC (Fig. 4B) [3, 26]. B6.j–kSEG mice
produce k2 and kx anti-DNP Ab, revealing a new anti-
DNP Ab set characterized by kx expression (Fig. 4B).
In B6 mice, all LC participate to the anti-NP response
except kx ([3, 27], Fig. 4A, NP). B6.j–kSEG mice respond
with both k2 and kx anti-NP Ab. Such Ab are even
present in the pre-immune repertoire when only j LC are
present for B6 (Fig. 4A, NP). Strikingly, B6.j–kSEG have
lower k2 anti-NP titers than B6 in spite of producing only
k2 and kx Ab.
Following immunization with MBP, mice didn’t develop
EAE, consistent with the disease association with few H-
2 haplotypes [28]. It is known that mice produce kx anti-
MBP Ab. These kx LC bear a CDR3 presenting
homology with a TCRb CDR3 of an encephalitogenic T
cell clone [29]. Here, we detect anti-MBP Ab in B6 and
B6.j–kSEG. For B6, all but k1 LC participate to the
response (Fig. 4A, MBP). The primary Ab response is
very discrete when compared with the Ab burst that
follows the second immunization. B6.j–kSEG mice are
characterized by a response involving k2 and kx LC. kxanti-MBP titers are strongly higher following the second
immunization whereas the total k2/kx titers are only
twofold higher, suggesting a preferential selection of kxanti-MBP Ab (Fig. 4A, MBP). Strikingly, naive B6.j–kSEG
have comparable anti-MBP Ab titers to B6 after a first
immunization.
Fig. 4. Immune response to PC/NP/MBP and DNP. (A) Sera
from BC8 (PC) or B6 (NP/MBP) mice in black and B6.j–kSEG
in white were analyzed by RIA for the first (squares) and
second response (circles) and compared to naive sera
(triangles). j/k1/k2-3-x and kx LC levels and VHT15 (only for
PC) usage were determined. (B) The anti-DNP response in
naive or DNP-OVA immunized BC8 and B6.j–kSEG was
quantified for j, k1 and k2-3-x LC. +/++: Magnitude of kxresponse. –: no Ab detection.
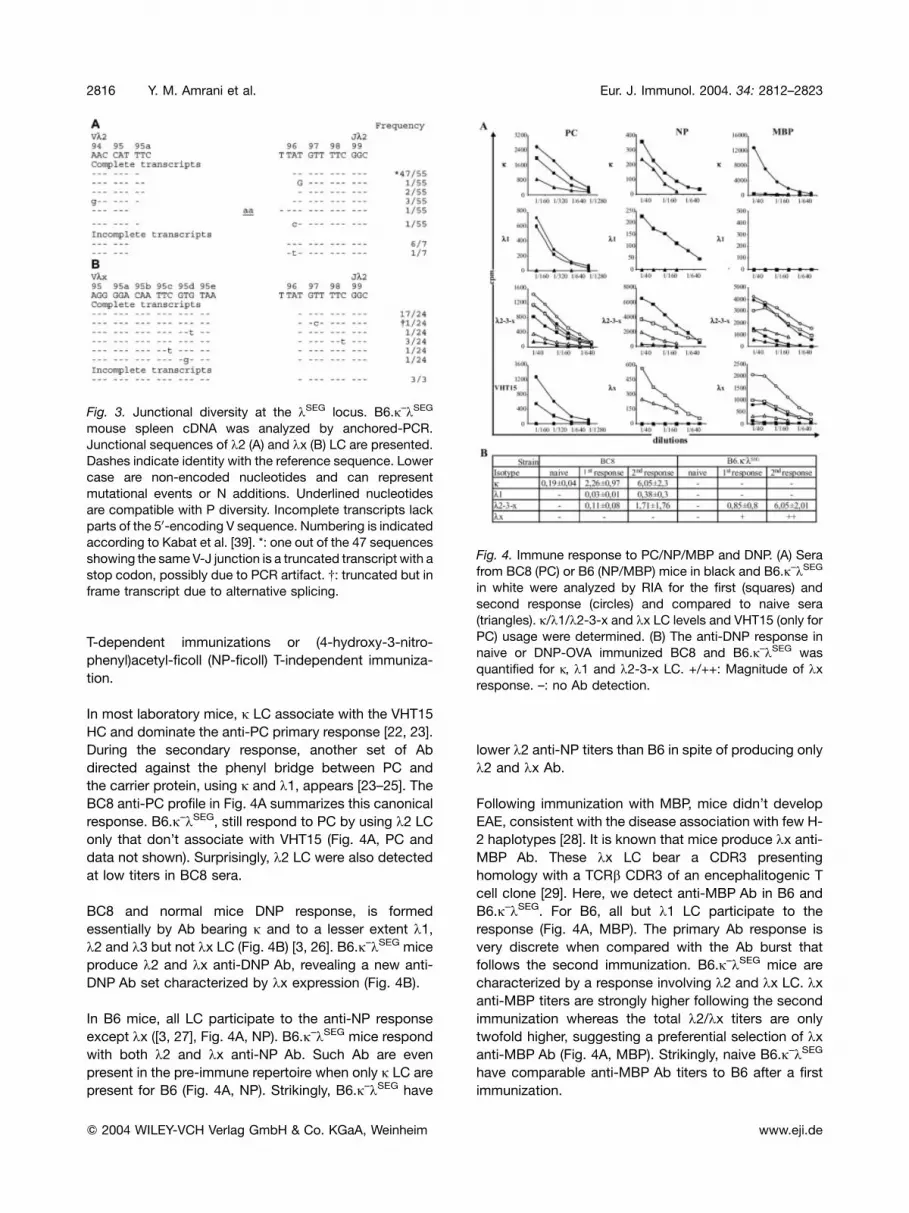
Fig. 3. Junctional diversity at the kSEG locus. B6.j–kSEG
mouse spleen cDNA was analyzed by anchored-PCR.
Junctional sequences of k2 (A) and kx (B) LC are presented.
Dashes indicate identity with the reference sequence. Lower
case are non-encoded nucleotides and can represent
mutational events or N additions. Underlined nucleotides
are compatible with P diversity. Incomplete transcripts lack
parts of the 50-encoding V sequence. Numbering is indicated
according to Kabat et al. [39]. *: one out of the 47 sequences
showing the sameV-J junction is a truncated transcript with a
stop codon, possibly due to PCR artifact. {: truncated but in
frame transcript due to alternative splicing.
2816 Y. M. Amrani et al. Eur. J. Immunol. 2004. 34: 2812–2823
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

2.5 Oligoclonality and restricted CDR3 size foranti-DNP mAb
We produced 13 B6.j–kSEG anti-DNP hybridomas,
including 1 VHQ52/Vkx-IgM and 12 VH36–60-IgG1 in
association with either kx (2/12) or k2 (10/12) LC as
determined by ELISA and PCR (data not shown). One
additional hybridoma (KS-D6) for which we were unable
to determine the HC uses k2.
IgG1 hybridomas HC and LC sequencing showed a nine
amino acids CDR3 length with restricted diversity
following VH36–60 association to JH3 rearranged with
two or three D segments. The IgM hybridoma uses JH4
and a different D segment to produce an eight amino
acids CDR3 (Fig. 5C, D). Similar mutation patterns in
CDR1 and CDR2 together with a restricted CDR3
diversity for HC and LC reflects the oligoclonality of
the anti-DNP response. “KS-D7” and “KS-D4” HC
sharing the same CDR3 pair with kx and k2, respectively.After DNP immunization, one new k2 junction (KS-D8) is
observed when compared to the naive LC repertoire
analyzed by anchored-PCR (Figs 3 and 5A).
Anti-DNP/TNP LC are characterized by a frequent TTT
codon at position 96 rather than the canonical TATcodon
[3, 20]. Here, the canonical junction prevails in 10 of the
11 hybridomas. This is probably due to Vk2SEG, showing
a single Thr>Asn substitution in its CDR3 when
compared to the homologous laboratory mouse Vk2 [1].
2.6 Recurrent characteristics of the anti-DNPresponse in B6.j–kSEG mice
We analyzed by Immunoscope spleen cDNA from
immunized B6, Igha and Ighb B6.j–kSEG mice using
VH36–60 and VHJ558 specific primers with an IgG
reverse primer (Fig. 6). VH36–60 profiles show a
predominant nine amino acids peak in 9/10 B6.j–kSEG
profiles, confirmed by direct PCR sequencing (data not
shown). Neither immunized B6 nor naive profiles show
Fig. 5. Protein sequence comparison of k LC and VH from B6.j–kSEG anti-DNP hybridoma. k2 (A) and kx (B) protein sequences
were compared to k2 and kx LC of anti-DNP hybridomas. (C) KS-D1 VH sequence is compared to B6 VHQ52 and JH4. (D) KS-
D2–5, 7–11 and 13–15 VH sequences are compared to VH36–60B (B6), VH36–60A (A/J strain) and JH3. All protein sequences are
translated from the respective GenBank nucleotide sequences (accession numbers for Vk: AY078484-AY078497; VH: AY078471-AY078483). Dashes: sequence identities. Amino acid substitutions are shown. Bold letters: amino acids probably encoded by D
segments. Italic letters: amino acids probably encoded by N–nucleotides. X: undetermined codons. Strokes set the boundaries of
the different domains. FR, CDR and C regions are indicated according to Kabat et al. [39].
Eur. J. Immunol. 2004. 34: 2812–2823 MZ B cell expansion in B6.j–kSEG mice 2817
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

this predominance. Conversely, VHJ558 profiles don’t
show significant disturbance. This confirms the oligo-
clonality and recurrence of the anti-DNP response in
B6.j–kSEG mice, regardless of the Igh haplotype.
2.7 Delayed IgG response characterizesB6.j-kSEG mice
The heterogeneity of anti-DNP and anti-OVA serum IgG
was characterized by IEF for B6, B6.j–, Igha and Ighb
B6.j–kSEG mice after a first and a second immunization
with DNP-OVA. These assays strengthened our previous
conclusions on the oligoclonal and recurrent anti-DNP
response since a restricted number of HC/LC IgG
associations is observed during the anti-DNP-OVA
immune response in both B6.j–kSEG mice when com-
pared to B6 and B6.j– mice. This result wasn’t expected
for the OVA carrier protein which presents a complex
antigenic structure that could consequently lead to a high
diversity of anti-OVA clones as seen for B6 and to a lesser
extent for B6.j– mice.
Strikingly, independently of the hapten or protein nature
of the Ag, the immune response is delayed in terms of IgG
appearance for both B6.j–kSEG mice since no immune
spectratype is detected following a first immunization in
contrast to B6 and B6.j– mice (Fig. 7A). The IEF revealed
with a polyclonal anti-IgG Ab didn’t detect any difference
between 1st, 2nd response or naive spectratypes in the
four strains (data not shown), confirming our observation.
In a different series of immunizations conducted for all
four strains, we found that anti-DNP and anti-OVA IgG Ab
titers show a picture consistent with that found by IEF.
When control sera showed slight reactivity (DNP) or no
reactivity (OVA) in all four strains, IgG Ab could be
detected at much higher titers in B6 and B6.j– when
compared to Igha or Ighb B6.j–kSEG mice after a first
Fig. 7. IgG spectratypes and ELISA serum titrations of anti-DNP and anti-OVA Ab from B6, B6.j–, B6.j–kSEG and B6.j–kSEG Igha
mice. (A) We compared by IEF, spectratypes of B6, B6.j–, B6.j–kSEG and B6.j–kSEG Igha mice IgG Ab against DNP and OVA in the
sera of naive mice (C), following the first (1), and the second immunization (2). Numbers on the right side indicate the pH gradient.
One representative spectratype is shown in C and 1, and in 2 for OVA/B6. (B) Sera from B6 (squares), B6.j– (circles), B6.j–kSEG
(diamonds) and B6.j–kSEG Igha (triangles) mice were analyzed by ELISA for IgG anti-DNP and anti-OVA responses in naive mice
(C), following a first (1) and a second immunization (2). Bars represent SD.
Fig. 6.Clonal expansion of VH36–60 IgG transcripts following
anti-DNP immunization of B6.j–kSEG and B6.j–kSEG Igha
mice. Representative Immunoscope CDR3 profiles obtained
after VH36–60-IgG amplification and run-off with a nested
IgG primer are compared to VHJ558 profiles for six naive or
five immunized B6, B6.j–kSEG and B6.j–kSEG Igha mice.
Arrows indicate expanded predominant peaks.
2818 Y. M. Amrani et al. Eur. J. Immunol. 2004. 34: 2812–2823
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

response. Following a second immunization, the differ-
ence in the secondary IgG response to both DNP and
OVA appeared less striking between all four strains
(Fig. 7B). This finding strengthens the conclusion that
there is a delayed IgG response rather than a lack or weak
IgG response in both B6.j–kSEG mice. IgM titers for DNP
or OVA responses didn’t show any striking difference
between all four strains (data not shown).
This situation could be related to the involvement of
different clones belonging to different B cell subsets after
a primary or a secondary immunization. MZ B cells have
been shown to respond very strongly and quickly to
blood-borne Agwhile FOB cells are those responsible for
the second wave of Ab or the so-called adapted immune
response [12]. Interestingly, A/J mice anti-phenylarso-
nate cross-reactive idiotype A found at the end of the
primary response and dominating the secondary re-
sponse is preferentially associated with HSAlow-IgDhigh-
IgMlow B cells belonging to FO B cells. Another B cell
subset HSAhigh-IgD–-IgMhigh with a MZ B cell like
phenotype is preferentially associated with the cross
reactive idiotype C found during the early primary
response and persisting throughout the successive
responses in low amounts [12, 30]. As shown in Fig. 5,
the only IgM anti-DNP hybridoma produced in a
B6.j–kSEG mouse uses totally different VH/VL sequences
from the rest of the IgG1 hybridoma, strengthening these
previous observations. It is now to be investigated
whether MZ B cells are involved in the delayed IgG
response in B6.j–kSEG mice.
2.8 MZ B cells are expanding when FO B cells arestrongly decreased in B6.j–kSEG mice
The analysis of BM B cell distribution reveals that newly
produced, immature B cells as well as mature, recirculat-
ing B cells are six-to sevenfold decreased in B6.j–kSEG
and B6.j–kSEG Igha when compared to B6 age-matched
2-month-old mice (Fig. 8A–D). This should be largely
enough to fill the spleen repertoire [11]. However, the
Fig. 8.Marginal zone B cells expand whereas FO B cells are strongly reduced in B6.j–kSEG and B6.j–kSEG Igha mice. (A) BM cells
were analyzed for IgM-B220 expression. B cell progenitors of the A-DHardy fractions (B220low-IgM–), B220low-IgM+ immature and
B220high-IgM+ mature/recirculating B cells are shown [5]. Numbers indicate percentages of cells in corresponding boxes. (B)
CD19+ splenocyte percentages are shown for corresponding mice. (C) Density plots show spleen MZ (CD21high-CD23+/–) and FO
(CD21+-CD23+) B cell distribution. Numbers indicate MZ and FO B cell percentages. (D) Representation of the mean absolute
number in millions � SD of BM immature, total spleen, spleen MZ and FO B cells. Populations percentages in the corresponding
compartments � SD are indicated in parentheses. (E) Ratios were determined from MZ and FO B cell absolute numbers for each
mouse strain, the mean of absolute number of MZ B cells from B6, B6.j–kSEG and B6.j–kSEG Igha mice vs. B6 and the mean of
absolute number of FO B cells from B6, B6.j–kSEG and B6.j–kSEG Igha mice vs. B6. Bars represent SD. Data are representative
(A–C) or are the mean (D–E) of four experiments using 2-month-old mice.
Eur. J. Immunol. 2004. 34: 2812–2823 MZ B cell expansion in B6.j–kSEG mice 2819
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

proportion of splenic B cells is twofold reduced and their
absolute number is fourfold reduced (Fig. 8B, C).
Interestingly, this decrease in B cell number and
distribution affects only FO B cells since their proportion
is reduced by three-to-sixfold when their cell absolute
number is seven-to-tenfold reduced. Conversely, MZ B
cells aren’t only preserved but rather expand in B6.j–kSEG
and B6.j–kSEG Igha mice (Fig. 8C, D). This last finding
appears clearly when one looks at theMZ/FOB cells ratio
for each mouse strain, and the MZ/FO B cells ratio
between B6/B6.j–kSEG/B6.j–kSEG Igha and B6 mice
(Fig. 8E).
Altogether, these data show that B6.j–kSEG and
B6.j–kSEG Igha mice are more able to produce MZ than
FO B cells, suggesting a peculiar feature of k B cells to
more likely differentiate into MZ B cells. A similar relative
expansion of MZ B cells has been already reported in
various mouse models with impaired B cell development
(see for review [12]). However, in B6.j–kSEG and
B6.j–kSEG Igha mouse models, we suggest that there
is no real impaired B cell development since we find an
expansion of k B cells: it’s interesting to point out that k2and kx BM immature B cells represent only 1/50 of the
immature BM B cells in normal mice [2]. Thus, the six-to-
sevenfold decrease in B cell production in B6.j–kSEG and
B6.j–kSEG Igha mice, mentioned above, must result from
a seven-to-eightfold increase in k2 and kx B cell
production by comparison to B6 mice. The decreased
number of produced immature BMB cells by comparison
to B6 mice would be mainly related to a reduced chance
to produce a functional LC since, in our mouse models, jLC production is abolished and k LC production is
theoretically reduced by half due to the absence of one
unit of recombination [1]. Thus, the spleen B cell
repertoire size is reduced due to the reduction of the
Ig LC repertoire when the integrity of the BCR or of the
signaling cascade following BCR activation aren’t
genetically affected. Kearney and al. [31] showed that
in VH81X.Cj– mice that produce only k LC,MZ B cells are
a large part of the B cell compartment; nevertheless, in
these mice the repertoire diversity is strongly affected
and almost totally blocked representing much less than a
tenth of a normal repertoire. Alternatively, there is no
expansion of MZ B cells in this model but rather a
persistence of such cells while B2 cells are dramatically
affected. This isn’t the case in our model because there’s
only 3–4 times less spleen B cells when compared to
control B6 mice. Moreover, the absolute number of MZ B
cells is clearly augmented. This shows that our mice have
expanded rather than maintained MZ B cells, bearing k2or kx LC, as described in other models.
Finally, the delayed IgG response following immunization
showed by IEF/ELISA data, in the light of the B cell
repertoire picture in B6.j–kSEG and B6.j–kSEG Igha mice,
would be due to the largeMZB cell representation. These
compiled observations thus strengthen previous data
establishing MZ B cells as responsible for the very early
immune response [12]. In parallel, a reduced number of
FO B cells decreases the chance to trigger a fast and
quantitative production of switched andmutated Ab. This
explains IgG Ab detection only after repeated immuniza-
tions in B6.j–kSEG mice (Fig. 7).
3 Conclusion
B6.j–kSEG mice, which produce solely k2 and kx LC,
show a preferential selection of k2 B cells only in PeC.
This reduced repertoire neither favors unusual diversifi-
cation mechanisms in mice like gene conversion and
deletion/insertions nor increases rare events such as P/N
additions or deletions at Vk-Jk junctions. Despite the
repertoire reduction, Ab response is obtained against
different Ag characterized by new Ab patterns compared
to conventional Ab responses described in the literature.
However, in contrast to j- mice [32], we show a decrease
of HC diversity during a rather oligoclonal and recurrent
anti-DNP response. The delayed IgG response could
result from the weak affinity of unusual Ab used to
overcome the introduced LC deficiency. Moreover, these
weak Ab seem to originate from expanding MZ B cells
thought to be polyreactive. Due to their origin, they’re in
majority of the IgM isotype. IgG Ab produced especially
by FO B cells are missing in the primary response due to
the strongly decreased number of their producers.
4 Materials and methods
4.1 Mice
C57BL/6N were purchased from Charles-River laboratory
(Saint-Aubin-les Elbeuf/France). B6.j–, B6.j–kSEG and
B6.j–kSEG Igha mice were produced and bred in our
laboratory. BC8 mice were a generous gift from Dr. G.
Bordenave (Immunophysiologie-moleculaire, Institut Pas-
teur).
4.2 FACS
The 4- or 2-month-old B6, B6.j–kSEG Igha and Ighb mouse
single-cell suspensions from BM, spleen, mesenteric LN,
PeC and PBL were incubated with rabbit naive serum,
stained with mAb and analyzed by FACSCalibur (Becton
Dickinson). Used mAb: FITC-conjugated 1D3 (anti-CD19),
AF6–78 (anti-lb), DS-1 (anti- lb/PharMingen, San Diego, CA)
and 7G6 (anti-CD21/PharMingen); biotin/SA-PerCP-conju-
gated 187–1 (anti-j), MS40–13 (anti-k1), 189B3 (anti-k2-k3-
2820 Y. M. Amrani et al. Eur. J. Immunol. 2004. 34: 2812–2823
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

kx) and 10C5 (anti-kx) [2]; PE-conjugated 53–7.3 (anti-CD5/
PharMingen) and B3B4 (anti-CD23/PharMingen); APC-con-
jugated RA3–6B2 (anti-CD45R/B220/PharMingen), M1/70
(anti-Mac-1/PharMingen) and 1D3 (anti-CD19/PharMingen).
4.3 Immunizations
Immunizations were performed s.c. with 100 lg DNP-OVA,
PC-Tg or guinea-pig MBP (SIGMA/St. Louis, MO) in CFA at
day 0 (D0) and in ICF at D12 on five BC8/6 B6.j–kSEG, fiveBC8/8 B6.j–kSEG and three B6/7 B6.j-kSEG mice, respec-
tively. Mice were bled at D11 and D22 for primary and
secondary responses. Three B6 and six B6.j–kSEG Ighb mice
immunized i.p. with 100 lgNP-ficoll in CFAwere bled at D11.
Naive mouse sera were used as controls.
For IEF and Immunoscope assays, five 5–7-week-old DNP-
OVA immunized B6, B6.j–, B6.j–kSEG and B6.j–kSEG Igha
mice were analyzed. After 4 months, B6, Igha and Ighb
B6.j–kSEG mice were immunized i.p. in saline 4 days before
they were killed to prepare spleen cDNA.
For ELISA, on D0 5–7-week-old five B6, five B6.j–, threeB6.j–kSEG, and five B6.j–kSEG Igha mice were bled then
immunized with 100 lg DNP-OVA in CFA, bled on D13,
boosted for the secondary response on D14 and bled on
D23.
4.4 RIA
187–1 (anti-j), TC54 (anti-VHT15) [33], MS40–13 (anti-k1),10C5 (anti-kx) mAb and a polyclonal rabbit anti-Ck2 Ab
which recognizes k2-k3-kx LC (obtained after repeated
immunization with MOPC 315 (IgA, k2), extensive absorption
on HOPC8 (IgA, j) coupled to CnBr-Sepharose column
(Pharmacia, Uppsala, Sweden) and isolation by affinity
chromatography on MG8–47 (IgG1,k3) coupled to CnBr-
Sepharose column were labeled with I125 (NEN) to char-
acterize the immune responses against PC-Tg/DNP-OVA/
NP-ficoll or MBP by solid-phase RIA using PC-BSA/DNP-
BSA/NP-BSA or guinea-pig MBP (SIGMA). MOPC 460 (IgA,
j), TNP5 (IgM, k1), MG8–47 (IgG3, k2) were used as anti-
DNP/TNP standards.
4.5 Hybridomas
Splenocytes from B6.j–kSEG Ighb mouse immunized three
times with DNP-OVA (s.c. at D0/CFA, D30/ICF and i.p. at
D70/saline) were fused to SP2/0 cells at D74. anti-DNP Ab
Hybridomas were selected by Ag-specific ELISA with DNP-
BSA coated microtiter plates (10 lg/ml).
4.6 Anchored-PCR analysis, hybridoma PCR and
sequencing
A 2-month-old B6.j–kSEG spleen Vk-Ck library was con-
structed as described [34] using 30MSCL4DO4 (AC-
CATCTGCCTTCCAGGCCA) and nested 30MSCL4DO3
(AGCTCTTCAGAGGAAGGTGG) Ck primers. The
400–600 bp PCR products were cloned with TOPO-TA
(INVITROGEN,Carlsbad,CA) forsequencingasdescribed [1].
Hybridoma cDNA were prepared as described [34]. PCR
were carried out using different 50VH family specific primers
[35] (a generous gift from Dr.V.Baretto) with 30MMIGM and
MMIGG HC l and c constant region specific primers [36],
respectively. 50Vk2-MSVLSUP1 or Vkx-MMVLXUP2
primers [1] were used with 30Ck2-MMCL2DO1
(CCTCTTTGGTGGGATTTGAA) primer to determine LC
sequence. PCR were sequenced with MMIGM20 (CATGGC-
CACCAGATTCTTAT) or MMIGG20 (ACTGTCACTGGCT-
CAGGGAA) nested primers, B or C VH primers and with
the nested 30MSCL4DO3 or Vk primers as described [1].
4.7 VH36–60 Immunoscope profile
Immunoscope analysis was carried out on spleen cDNA from
six naive 6-month-old B6, Igha and Ighb B6.j–kSEG mice and
five immunized 7-month-old B6, Igha and Ighb B6.j–kSEG
mice. VH36–60-IgG or VHJ558-IgG PCR were carried out
using 50VHC or MVHJ558 and 30MMIGG primers [36] with
4 ll cDNA in PCR conditions described in [1]. Run-off
reactions were carried out and size separated on a 6%
acrylamide gel on an ABI373 automated sequencer (Ap-
pliedBiosystemInc/PE-Biosystems, Warrington, UK) as de-
scribed [36]. CDR3 size distribution and signal intensities
were analyzed with Immunoscope and ISEAPeaks [37, 38].
4.8 Ab isoelectric spectra
IEF analysis was carried out in a 4% polyacrylamide gel
containing 2% ampholytes (Ampholine Preblended pH range
5–8, Amersham Biosciences, France) and 3 M urea.
Samples were obtained by precipitation of 6ll serum at
18% Na2SO4 and dissolution in 20 ll 3 M urea. The gel was
focused at 1,000 V, 2 h and 30 min at 8�C and electroblotted
onto nitrocellulose (Hybond-ECL membrane, Amersham)
15 h at 20 V/cm and 4�C in transfer buffer (10 mM-Trizma-
base/100 mM-glycine/20%-ethanol/0.1%-SDS). The mem-
brane was blocked in PBS/0.1%-Tween20/5%-BSA, probed
with biotin labeled DNP-BSA or OVA. Biotin/SA-HRP
Complexes (Amersham) were revealed by supersignal
chemiluminescent substrate (Pierce-Perbio, France).
Eur. J. Immunol. 2004. 34: 2812–2823 MZ B cell expansion in B6.j–kSEG mice 2821
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

4.9 ELISA
NUNCmicrotiter plates were coatedwith 10 lg/ml DNP-BSA
or 10 lg/ml OVA solutions and BSA saturated. Serial serum
dilutions were incubated overnight at 4�C and washed.
Bound Ab were revealed using HRP-conjugated goat anti-
mouse IgM or IgG Ab (Calbiochem) and development was
conducted with OPD (Sigma).
Acknowledgements: We thank Drs. P. Boudinot, D. Rueff-
Juy and P. Vieira for critical reading of the manuscript. We are
grateful to Dr. A. Cumano, D. Guy-Grand, L. Majlessi and P.
Vieira for their Ab. Y.M.Awas supported by CROUS de Paris-
France, M.E.S.R.S-Algeria, Pfizer-Algeria, Pasteur-Weiz-
mann Foundation and ARC.
References
1 Amrani, Y. M., Voegtle, D., Montagutelli, X., Cazenave, P. A.and Six, A., The Ig light chain restricted B6.j–lSEG mouse strainsuggests that the IGL locus genomic organization is subject toconstant evolution. Immunogenetics 2002. 54: 106–119.
2 Sanchez, P., Drapier, A. M., Cohen-Tannoudji, M., Colucci, E.,Babinet, C. and Cazenave, P. A., Compartmentalization oflambda subtype expression in the B cell repertoire of mice with adisrupted or normal C kappa gene segment. Int. Immunol. 1994.6: 711–9.
3 Eisen, H. N. and Reilly, E. B., Lambda chains and genes in inbredmice. Annu. Rev. Immunol. 1985. 3: 337–65.
4 Sanchez, P., Rueff-Juy, D., Boudinot, P., Hachemi-Rachedi, S.and Cazenave, P. A., The lambda B cell repertoire of kappa-deficient mice. Int. Rev. Immunol. 1996. 13: 357–68.
5 Hardy, R. R. and Hayakawa, K., B cell development pathways.Annu. Rev. Immunol 2001. 19: 595–621.
6 Carvalho, T. L., Mota-Santos, T., Cumano, A., Demengeot, J.and Vieira, P., Arrested B lymphopoiesis and persistence ofactivated B cells in adult interleukin 7–/– mice. J. Exp. Med. 2001.194: 1141–1150.
7 Hao, Z. and Rajewsky, K., Homeostasis of peripheral B cells inthe absence of B cell influx from the bone marrow. J. Exp. Med.2001. 194: 1151–1164.
8 Dammers, P. M., Visser, A., Popa, E. R., Nieuwenhuis, P. andKroese, F. G., Most marginal zone B cells in rat express germlineencoded Ig VH genes and are ligand selected. J. Immunol. 2000.165: 6156–6169.
9 Martin, F. and Kearney, J. F., Positive selection from newlyformed to marginal zone B cells depends on the rate of clonalproduction, CD19, and btk. Immunity 2000. 12: 39–49.
10 Maclennan, I. C. M., Gray, D., Kumararatne, D. S. and Bazin,H., The lymphocytes of splenic marginal zones a distinct B celllineage. Immunol. Today 1982. 3: 3–307.
11 Maclennan, I. C. M., B cell receptor regulation of peripheral Bcells. Curr. Opin. Immunol. 1998. 10: 220–225.
12 Martin,F. and Kearney,J.F., Marginal-zone B cells. NatureReviews.Immunology. 2002. 2: 323–335.
13 Hayakawa, K., Hardy, R. R. and Herzenberg, L. A., PeritonealLy-1 B cells: genetic control, autoantibody production, increasedlambda light chain expression. Eur. J. Immunol. 1986. 16:450–456.
14 Weill,J.C. and Reynaud,C.A., Rearrangement/hypermutation/gene conversion: when, where and why? Immunol.Today 1996.17: 92–97.
15 Wilson, P., Liu, Y. J., Banchereau, J., Capra, J. D. and Pascual,V., Amino acids and deletions contribute to diversify the human Igrepertoire. Immunol. Rev. 1998. 162: 143–151.
16 Bentolila, L. A., Olson, S., Marshall, A., Rougeon, F., Paige, C.J., Doyen, N. and Wu, G. E., Extensive junctional diversity in Iglight chain genes from early B cell progenitors of mu MT mice. J.Immunol. 1999. 162: 2123–2128.
17 Sanchez, P., Marche, P. N., Rueff-Juy, D. and Cazenave, P. A.,Mouse V lambda x gene sequence generates no junctionaldiversity and is conserved in mammalian species. J. Immunol.1990. 144: 2816–2820.
18 Boudinot, P., Rueff-Juy, D., Drapier, A. M., Cazenave, P. A. andSanchez, P., Various V-J rearrangement efficiencies shape themouse lambda B cell repertoire. Eur. J. Immunol. 1995. 25:2499–505.
19 Hardy, R. R., Variable gene usage, physiology and developmentof Ly-1+ (CD5+) B cells. Curr. Opin. Immunol. 1992. 4: 181–185.
20 Boudinot, P., Drapier, A. M., Cazenave, P. A. and Sanchez, P.,Mechanistic and selective constraints act on the establishment ofV lambda J lambda junctions in the B cell repertoire. J. Immunol.1994. 152: 2248–2255.
21 Nadel, B. and Feeney, A. J., Nucleotide deletion and P additionin V(D)J recombination: a determinant role of the coding-endsequence. Mol. Cell Biol. 1997. 17: 3768–3778.
22 Barstad, P., Rudikoff, S., Potter, M., Cohn, M., Konigsberg, W.and Hood, L., Immunoglobulin structure: amino terminalsequences of mouse myeloma proteins that bind phosphorylcho-line. Science 1974. 183: 962–966.
23 Claflin, J. L. and Berry, J., Genetics of the phosphocholine-specific antibody response to Streptococcus pneumoniae.Germ-line but not mutated T15 antibodies are dominantlyselected. J. Immunol. 1988. 141: 4012–4019.
24 Chang, S. P., Brown, M. and Rittenberg, M. B., Immunologicmemory to phosphorylcholine. II. PC-KLH induces two antibodypopulations that dominate different isotypes. J. Immunol. 1982.128: 702–706.
25 Brown, M., Stenzel-Poore, M., Stevens, S., Kondoleon, S. K.,Ng, J., Bachinger, H. P. and Rittenberg, M. B., Immunologicmemory to phosphocholine keyhole limpet hemocyanin. Recur-rent mutations in the lambda 1 light chain increase affinity forantigen. J. Immunol. 1992. 148: 339–346.
26 Pricop, L., Brumeanu, T., Elahi, E., Moran, T., Wang, B. S.,Troustine, M., Huszar, D., Alt, F. and Bona, C., Antibodyresponse elicited by T-dependent and T-independent antigens ingene targeted kappa-deficient mice. Int. Immunol. 1994. 6:1839–1847.
27 Cumano, A. and Rajewsky, K., Structure of primary anti-(4-hydroxy-3-nitrophenyl)acetyl (NP) antibodies in normal andidiotypically suppressed C57BL/6 mice. Eur. J. Immunol. 1985.15: 512–520.
28 Goverman, J., Brabb, T., Paez, A., Harrington, C. andVondassow, P., Initiation and regulation of CNS autoimmunity.Criti. Rev. Immunol. 1997. 17: 469–480.
29 Maier, C. C., Galin, F. S., Jarpe, M. A., Jackson, P., Krishna, N.R., Gautam, A. M., Zhou, S. R., Whitaker, J. N. and Blalock, J.E., A V lambda x-bearing monoclonal antibody with similarspecificity and sequence to encephalitogenic T cell receptors. J.Immunol. 1994. 153: 1132–1140.
30 Luko, C. W., Vansanten, G., Ryelandt, M., Denis, O., Wuilmart,C., Andris, F., Van Acker, A., Brait, M., Cloquet, J. P., Ismaili, N.,Nisol, F., Latinne, D., Brown, A., Leo, O., Bazin, H. and Urbain,
2822 Y. M. Amrani et al. Eur. J. Immunol. 2004. 34: 2812–2823
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

J., Distinct VH repertoires in primary and secondary B celllymphocyte subsets in the preimmune repertoire of A/J mice: theCRI-A idiotype is preferentially associated with the HSA(low) Bcell subset. Eur. J. Immunol. 2000. 30: 2312–2322.
31 Martin, F., Won, W. J. and Kearney, J. F., Generation of thegermline peripheral B cell repertoire: VH81X-lambda B cells areunable to complete all developmental programs. J. Immunol.1998. 160: 3748–3758.
32 Bot, A., Nangpal, A., Pricop, L., Bogen, B., Kaushik, A. andBona, C. A., V lambda-light chain genes reconstitute immuneresponses to defined carbohydrate antigens or haptens byutilizing different VH genes. Mol. Immunol. 1996. 33: 1359–1368.
33 Desaymard, C., Giusti, A. M. and Scharff, M. D., Rat anti-T15monoclonal antibodies with specificity for VH- and VH-VLepitopes. Mol. Immunol. 1984. 21: 961–967.
34 Rueff-Juy, D., Marche, P. N., Drapier, A. M. and Cazenave, P.A., Junctional diversity of H and L chains allows the coexpressionof two mutually exclusive idiotopes (IdI104 and IdI558). J.Immunol. 1991. 146: 4024–4030.
35 Ehlich, A., Martin, V., Muller, W. and Rajewsky, K., Analysis ofthe B cell progenitor compartment at the level of single cells. Curr.Biol. 1994. 4: 573–583.
36 Delassus, S., Gey, A., Darche, S., Cumano, A., Roth, C. andKourilsky, P., PCR-based analysis of the murine immunoglobulinheavy-chain repertoire. J. Immunol. Methods 1995. 184:219–229.
37 Pannetier, C., Cochet, M., Darche, S., Casrouge, A., Zoller, M.and Kourilsky, P., The sizes of the CDR3 hypervariable regions ofthe murine T-cell receptor beta chains vary as a function of therecombined germ-line segments. Proc. Natl. Acad. Sci. USA1993. 90: 4319–4323.
38 Collette, A. and Six, A., ISEApeaks: an Excel platform forGeneScan and Immunoscope data retrieval, management andanalysis. Bioinformatics 2002. 18: 329–330.
39 Kabat, E. A., Wu, T. T., Perry, H. M., Gottesman, K. S. andFoeller, C., Sequences of proteins of immunological interest.Public Health Service, NIH., Washington, DC 1991
Correspondence: Adrien Six, Institut Pasteur, 25 rue du
Dr. Roux, F-75015 Paris, France
Fax: +33-1-4061-3066
e-mail: [email protected]
Eur. J. Immunol. 2004. 34: 2812–2823 MZ B cell expansion in B6.j–kSEG mice 2823
f 2004 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de




















