Looking through the eyes of fungi: molecular genetics of photoreception
11
MicroReview Looking through the eyes of fungi: molecular genetics of photoreception Alfredo Herrera-Estrella 1 and Benjamin A. Horwitz 2 * 1 Laboratorio Nacional de Genómica para la Biodiversidad, Cinvestav Campus Guanajuato, Irapuato 36500, Guanajuato, México. 2 Department of Biology, Technion – Israel Institute of Technology, Haifa 32000, Israel. Summary Filamentous fungi respond to a variety of environ- mental signals. One of them is light, providing critical information about orientation, or impending stress. Cells of filamentous fungi appear to sense blue light through a unique transcription factor that has a flavin chromophore and activates its targets in a light- dependent manner, the white collar (WC) complex. Fungal photophysiology, though, predicted a greater complexity of responses to the whole visible spectrum. The rapidly growing fungal genome data- base provides candidates to explain how fungi see not only blue, but also near-UV, green and red light. At the same time, there are surprises in the genomes, including photoreceptors for which there are no obvious photoresponses. Linking these genes and their functions will help understand how a list of only a few biological chromophores accounts for such a diversity of responses. At the same time, deeper mechanistic understanding of how the WC complex functions will lead to fundamental insights at the point where the environment impinges, in this case in the form of photons, on the transcriptional machinery of the cell. Introduction Filamentous fungi, like other microorganisms, respond to a variety of environmental signals. Information about which way is up or down, for example, is required for spore dispersal. Light also helps a growing fungal cell to orient itself, and might provide a clue to whether the fungus is in the soil, inside a host, or exposed to air and other stresses. A response to light might be very rapid, simply reorienting the direction of growth, or might provide the initial cue to alter transcriptional profiles. These, in turn, provide the enzymes and metabolites needed for protection from damage, as well as programming the dif- ferentiation of new structures (Figs 1 and 2). Photore- sponses are likely to contribute to the survival of the microorganism, its fitness and its ability to compete. Visible light, while not necessarily damaging in itself, can provide early warning of impending UV damage (Gressel and Rau, 1983; Yasuhira and Yasui, 1992; Berrocal-Tito et al., 1999). Keeping UV-A light out of greenhouses, for example, is one way to reduce some plant diseases spread by spores (Sasaki et al., 1985). Such ecological considerations, while useful, are not the only reason to study fungal photoresponses. Photoreceptors are pro- teins or protein complexes that are able to harvest light, generating a signal, which is propagated into the cell to stimulate a cellular response. Their light-absorbing cofactors, known as chromophores, are typically small molecules. Max Delbrück made the point in a 1976 lecture (see Lipson and Horwitz, 1991) that a very short list of chromophores accounts for all of biological photochemistry. The main chromophores are retinal, tet- rapyrroles and flavins. The number of known apoproteins grew rapidly with genome sequencing, and some addi- tions have been made to the chromophore list since 1976. Nevertheless, the list is still short; six main photoreceptor classes are now recognized, three of which detect light by flavin chromophores (van der Horst and Hellingwerf, 2004). Several members of the list are known or predicted to be important for fungi (Table 1; Idnurm and Heitman, 2005a; Corrochano and Galland, 2006; Purschwitz et al., 2006). PAS/LOV domain photosensors: WC1/2 and VIVID The initial breakthrough in molecular understanding of fungal phototransduction occurred when the white collar-1 (wc-1) gene (Fig. 1D and E) was cloned by chromosome walking and complementation of mutants (Ballario et al., 1996). WC-1 is a zinc finger transcription factor that binds the promoters of light-regulated genes. The second white Accepted 20 January, 2007. *For correspondence. E-mail horwitz@ tx.technion.ac.il; Tel. (+972) 4 8293976; Fax (+972) 4 8225153. Molecular Microbiology (2007) 64(1), 5–15 doi:10.1111/j.1365-2958.2007.05632.x © 2007 The Authors Journal compilation © 2007 Blackwell Publishing Ltd
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Looking through the eyes of fungi: molecular genetics of photoreception

MicroReview
Looking through the eyes of fungi: molecular geneticsof photoreception
Alfredo Herrera-Estrella1 and Benjamin A. Horwitz2*1Laboratorio Nacional de Genómica para laBiodiversidad, Cinvestav Campus Guanajuato, Irapuato36500, Guanajuato, México.2Department of Biology, Technion – Israel Institute ofTechnology, Haifa 32000, Israel.
Summary
Filamentous fungi respond to a variety of environ-mental signals. One of them is light, providing criticalinformation about orientation, or impending stress.Cells of filamentous fungi appear to sense blue lightthrough a unique transcription factor that has a flavinchromophore and activates its targets in a light-dependent manner, the white collar (WC) complex.Fungal photophysiology, though, predicted a greatercomplexity of responses to the whole visiblespectrum. The rapidly growing fungal genome data-base provides candidates to explain how fungi seenot only blue, but also near-UV, green and red light. Atthe same time, there are surprises in the genomes,including photoreceptors for which there are noobvious photoresponses. Linking these genes andtheir functions will help understand how a list of onlya few biological chromophores accounts for such adiversity of responses. At the same time, deepermechanistic understanding of how the WC complexfunctions will lead to fundamental insights at thepoint where the environment impinges, in this case inthe form of photons, on the transcriptional machineryof the cell.
Introduction
Filamentous fungi, like other microorganisms, respond toa variety of environmental signals. Information aboutwhich way is up or down, for example, is required forspore dispersal. Light also helps a growing fungal cell toorient itself, and might provide a clue to whether thefungus is in the soil, inside a host, or exposed to air and
other stresses. A response to light might be very rapid,simply reorienting the direction of growth, or might providethe initial cue to alter transcriptional profiles. These, inturn, provide the enzymes and metabolites needed forprotection from damage, as well as programming the dif-ferentiation of new structures (Figs 1 and 2). Photore-sponses are likely to contribute to the survival of themicroorganism, its fitness and its ability to compete.Visible light, while not necessarily damaging in itself, canprovide early warning of impending UV damage (Gresseland Rau, 1983; Yasuhira and Yasui, 1992; Berrocal-Titoet al., 1999). Keeping UV-A light out of greenhouses, forexample, is one way to reduce some plant diseasesspread by spores (Sasaki et al., 1985). Such ecologicalconsiderations, while useful, are not the only reason tostudy fungal photoresponses. Photoreceptors are pro-teins or protein complexes that are able to harvest light,generating a signal, which is propagated into the cellto stimulate a cellular response. Their light-absorbingcofactors, known as chromophores, are typically smallmolecules. Max Delbrück made the point in a 1976lecture (see Lipson and Horwitz, 1991) that a very shortlist of chromophores accounts for all of biologicalphotochemistry. The main chromophores are retinal, tet-rapyrroles and flavins. The number of known apoproteinsgrew rapidly with genome sequencing, and some addi-tions have been made to the chromophore list since 1976.Nevertheless, the list is still short; six main photoreceptorclasses are now recognized, three of which detect light byflavin chromophores (van der Horst and Hellingwerf,2004). Several members of the list are known or predictedto be important for fungi (Table 1; Idnurm and Heitman,2005a; Corrochano and Galland, 2006; Purschwitz et al.,2006).
PAS/LOV domain photosensors: WC1/2 and VIVID
The initial breakthrough in molecular understanding offungal phototransduction occurred when the white collar-1(wc-1) gene (Fig. 1D and E) was cloned by chromosomewalking and complementation of mutants (Ballario et al.,1996). WC-1 is a zinc finger transcription factor that bindsthe promoters of light-regulated genes. The second white
Accepted 20 January, 2007. *For correspondence. E-mail [email protected]; Tel. (+972) 4 8293976; Fax (+972) 4 8225153.
Molecular Microbiology (2007) 64(1), 5–15 doi:10.1111/j.1365-2958.2007.05632.x
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
collar protein, WC-2, is also a GATA-like zinc finger tran-scription factor. A wc-2 mutant that lacks most of theC-terminal part of the protein, including the DNA bindingdomain, is defective in most aspects of the light response(Crosthwaite et al., 1997; Linden and Macino, 1997). Ithas also been found that WC-2 does not directly perceivelight. Both proteins also contain PAS domains (PAS:
Drosophila period, PER – vertebrate aryl hydrocarbonreceptor nuclear translocator, ARNT – Drosophila single-minded, SIM). PAS domains participate in protein–proteininteractions, and have another special property: they maybind cofactors that confer sensory functions (Taylor andZhulin, 1999). The PAS domains are important for theformation of a nuclear complex containing WC-1 and
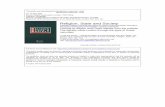
Fig. 1. Fungal photophysiology:morphogenesis and development.A. Fruit body formation in Schizophyllumcommune. Colonies were grown in the dark(left) or exposed to UV-A light (right), andfruiting scored 3 days later. Reprinted fromYli-Mattila (1987) with permission.B. A brief pulse of blue light inducessporulation of Trichoderma atroviride.Colonies were grown on filter paper with liquidmedium; only spores (right) are visible, wherethe colony perimeter was located when itreceived light, 24 h before the photo wastaken. Scanning EM (inserts) shows hyphae(left), and hyphae and conidiophores (right).C. Under appropriate conditions,developmental choice is determined by light inPhycomyces. Left, microphores formed in thedark. Blue light inhibits microphore formation.In both cases, the large sporangiophores(macrophores, see Fig. 2A) were removed.Photos by L. M. Corrochano, reprinted fromCorrochano and Galland (2006) withpermission.D. Continuous blue light promotes carotenoidsynthesis in Neurospora (image: FungalGenetics Stock Center, http://www.fgsc.net/,with thanks to Kevin McCluskey).E. Circadian control of conidiation inNeurospora. The white collar (wc-1 and wc-2)genes, discovered in a screen for defectiveregulation of pigment biosynthesis encode thebest-characterized photoregulator, thewhite-collar complex (WCC). Shown here isphase setting by transfer from light to darkin the band (bd) genetic background,and altered phase in a mutant lacking alight-responsive element (LRE) recognized bythe WCC in the promoter of the frq gene.FRQ is at the core of the circadian clock. Thismutant is deleted for the proximal of the twoLREs in the promoter of FRQ. From(Froehlich et al., 2002), reprinted withpermission from AAAS.
6 A. Herrera-Estrella and B. A. Horwitz
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15
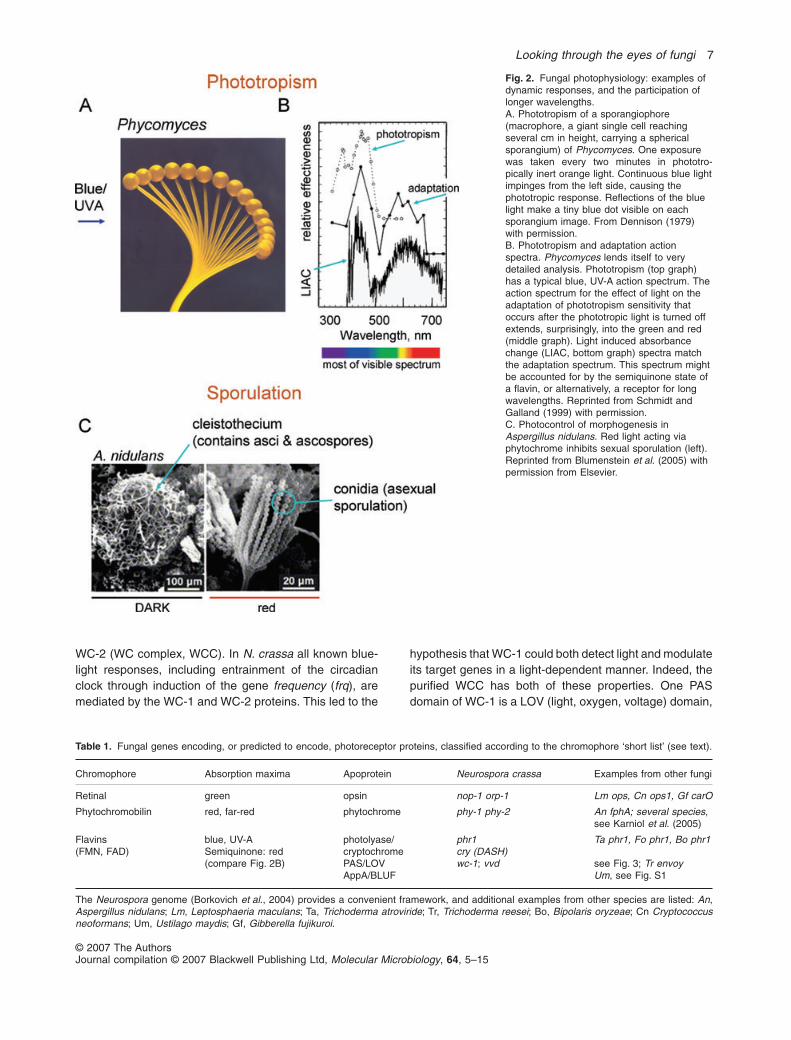
WC-2 (WC complex, WCC). In N. crassa all known blue-light responses, including entrainment of the circadianclock through induction of the gene frequency (frq), aremediated by the WC-1 and WC-2 proteins. This led to the
hypothesis that WC-1 could both detect light and modulateits target genes in a light-dependent manner. Indeed, thepurified WCC has both of these properties. One PASdomain of WC-1 is a LOV (light, oxygen, voltage) domain,
Fig. 2. Fungal photophysiology: examples ofdynamic responses, and the participation oflonger wavelengths.A. Phototropism of a sporangiophore(macrophore, a giant single cell reachingseveral cm in height, carrying a sphericalsporangium) of Phycomyces. One exposurewas taken every two minutes in phototro-pically inert orange light. Continuous blue lightimpinges from the left side, causing thephototropic response. Reflections of the bluelight make a tiny blue dot visible on eachsporangium image. From Dennison (1979)with permission.B. Phototropism and adaptation actionspectra. Phycomyces lends itself to verydetailed analysis. Phototropism (top graph)has a typical blue, UV-A action spectrum. Theaction spectrum for the effect of light on theadaptation of phototropism sensitivity thatoccurs after the phototropic light is turned offextends, surprisingly, into the green and red(middle graph). Light induced absorbancechange (LIAC, bottom graph) spectra matchthe adaptation spectrum. This spectrum mightbe accounted for by the semiquinone state ofa flavin, or alternatively, a receptor for longwavelengths. Reprinted from Schmidt andGalland (1999) with permission.C. Photocontrol of morphogenesis inAspergillus nidulans. Red light acting viaphytochrome inhibits sexual sporulation (left).Reprinted from Blumenstein et al. (2005) withpermission from Elsevier.
Table 1. Fungal genes encoding, or predicted to encode, photoreceptor proteins, classified according to the chromophore ‘short list’ (see text).
Chromophore Absorption maxima Apoprotein Neurospora crassa Examples from other fungi
Retinal green opsin nop-1 orp-1 Lm ops, Cn ops1, Gf carO
Phytochromobilin red, far-red phytochrome phy-1 phy-2 An fphA; several species,see Karniol et al. (2005)
Flavins(FMN, FAD)
blue, UV-ASemiquinone: red(compare Fig. 2B)
photolyase/cryptochromePAS/LOVAppA/BLUF
phr1cry (DASH)wc-1; vvd
Ta phr1, Fo phr1, Bo phr1
see Fig. 3; Tr envoyUm, see Fig. S1
The Neurospora genome (Borkovich et al., 2004) provides a convenient framework, and additional examples from other species are listed: An,Aspergillus nidulans; Lm, Leptosphaeria maculans; Ta, Trichoderma atroviride; Tr, Trichoderma reesei; Bo, Bipolaris oryzeae; Cn Cryptococcusneoformans; Um, Ustilago maydis; Gf, Gibberella fujikuroi.
Looking through the eyes of fungi 7
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15

with a flavin chromophore. In plant phototropins (Briggset al., 2001), LOV domains bind a flavin mononucleotide(FMN) molecule and cyclically modify their structure in alight-dependent manner (undergo photocycles). Thecrystal structure of a LOV-FMN complex (Crosson andMoffat, 2001) revealed a set of residues in the vicinity ofFMN that are highly conserved, including in fungal homo-logues, suggesting that they bind flavin and function asblue-light photoreceptors. Induction of the light responsivegenes frq and al-3 was not restored in Neurospora wc-1null mutants upon introduction of a modified version of theprotein lacking the LOV domain, indicating that the WC-1LOV domain is essential for light-activated transcription.Removal of the predicted flavin-adduct forming cysteine ofWC-1 eliminates light sensitivity. The WC-1 LOV domainbinds FAD, instead of FMN as in phototropin. Comparisonof the LOV domains of WC-1 and phototropin shows alonger linker between a helices A and C, perhaps accom-modating the larger FAD molecule. a helix A contains theflavin-adduct forming cysteine, so WC-1 and phototropinmay share the same photochemistry (Cheng et al., 2003).A cis element (al-3 proximal element, APE) was identifiedin blue-responsive carotenoid biosynthesis genes (Carat-toli et al., 1994). A consensus sequence for a blue-lightresponse element (LRE) has been proposed based on theanalysis of light responsive genes in Neurospora (He andLiu, 2005). These elements have also been found in lightresponsive genes in Trichoderma atroviride (Berrocal-Titoet al., 1999; Rosales-Saavedra et al., 2006). The purifiedWCC recognizes the LREs of its target gene frq even in thedark, activating frq transcription when exposed to light(Froehlich et al., 2002). In short, binding and light-dependent activation are sufficient for the WCC to be aphotoreceptive transcription factor, strong evidence thatthis is indeed what happens in the Neurospora cell.
Variations on this Neurospora paradigm are alreadyapparent. As we shall see below, some wc-1 homologueslack the DNA-binding domain. Furthermore, after bindingof the WCC, transcription needs to be activated. Neuro-spora WC-1 has poly Q tracts that serve as activationdomains near both the N and C termini of the molecule,while those of Tuber borchii and T. atroviride do not(Fig. 3). Thus, a WC-1 activation domain is not alwaysnecessary to couple photoreception to transcription(Ambra et al., 2004). Another activator might be recruitedby these variants of the WCC, or, lacking a conservedsequence signature, though some activation domainsmight escape bioinformatics detection. The T. atrovirideWCC is responsible for light repression as well as induc-tion of target genes, in contrast to Neurospora, where onlylight activation of gene expression has been well-characterized so far (Rosales-Saavedra et al., 2006). Isrepression a direct action of WCC at the promoter, ormediated by induction of a negative-acting factor?
WC-1 and WC-2 are subjected to light-dependent phos-phorylation, with WC-1 being phosphorylated in parallelwith the transient increase in the transcript levels of light-regulated genes (Schwerdtfeger and Linden, 2000).Another element in WC regulation is protein kinase C,which is involved in photoresponses of Neurospora.Constitutive-active and dominant negative PKC alleleshave opposite effects on WC-1 stability and on lightresponses (Franchi et al., 2005). Interaction of PKC withWC-1 in vivo is transiently lost after the start ofillumination. Other kinase activities, responsible for WC-1hyperphosphorylation in the light, remain to be identified.In this context, we note that in T. atroviride overexpressionof the regulatory subunit of protein kinase A blocks theexpression of otherwise blue light induced genes (Casas-Flores et al., 2006). Because PKC regulates WC-1 proteinlevels, phosphorylation by PKC might be relevant to adap-tation, as is a second photoreceptor, VIVID (Table 1),described below. A new player in the action of the WCCwas found by chromatin immunoprecipitation: transientlight-induced histone acetylation at the LRE of the al-3promoter requires WC-1, as well as a histone acetyltrans-ferase, NGF-1 (Grimaldi et al., 2006).
Is the WCC the major photoreceptor throughout thefungi? Classical photophysiology was done in greatestdetail for the zygomycete Phycomyces, resulting in theidentification of several loci including MAD A,B,C, whichwere predicted to encode photoreceptors (Bergman et al.,1969; Cerdá-Olmedo and Lipson, 1987). Phycomyceshas now been shown to have two genes related to wc-1(Fig. 3), probably the result of an ancestral duplication(Idnurm et al., 2006). Sequencing of the wc-1 genes in theoriginal Phycomyces mad mutants showed that madAalleles have mutations in one of the wc-1 genes, makingthis the first Phycomyces photoreceptor to be identified(Idnurm et al., 2006). The function of the second gene,wco1, is not yet known. At least, two photosystems areneeded to cover the 10 orders of magnitude in light inten-sity, from 10-9 to 10 W m-2, to which this organism canrespond (Galland and Lipson, 1987). Another zygo-mycete, Mucor circinelloides, has three genes encodingproteins similar to Neurospora wc-1. One is needed forcarotenogenesis (blue-sensitive) and another for photot-ropism (blue and green). Thus, different wc-1 genes canbe assigned to distinct photoresponses (Silva et al.,2006). There is nothing in their domain structure toprovide a hint to why phototropism has a green compo-nent, while carotenogenesis does not. Turning to thebasidiomycetes, Coprinus cinerea WC-1 has a functionalactivation domain near the C-terminus but lacks a DNA-binding domain (Fig. 3) (Terashima et al., 2005), asdoes the predicted wc-1 homologue in Cryptococcusneoformans. This domain structure, characteristic ofbasidiomycete and some of the zygomycete wc-1 homo-
8 A. Herrera-Estrella and B. A. Horwitz
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15

logues (Fig. 3) is a variation on the Neurospora paradigm.BWC2, which has a Zn finger domain, is apparently suf-ficient to confer DNA binding on the WCC (Idnurm andHeitman, 2005b). Although C. neoformans bwc1 tran-script levels are not altered by light, bwc2 is induced uponexposure to light, except in a bwc1 mutant. bwc2 is thus alight-regulated gene and dependent on the presence ofBWC1 (Idnurm and Heitman, 2005b). The production ofdikaryotic filaments is inhibited by blue light, and bwc1 isrequired for light sensitivity (Lu et al., 2005). In addition,deletion of bwc1 or bwc2 results in hypersensitivity toultraviolet light and reduced virulence of C. neoformans.
A second LOV-domain photoreceptor (vivid, vvd)enables Neurospora to perceive and respond to dailychanges in light intensity. vvd encodes a protein that isessentially just a LOV domain. In constant light mutants inVVD overproduce carotenoids and, thus, are brightorange. This suggested that VVD acts as a general repres-sor of light regulated processes. Indeed VVD defined anew negative feedback loop connecting the output of the
circadian clock with light input (Heintzen et al., 2001;Shrode et al., 2001; Schwerdtfeger and Linden, 2003). Incontrast to the WCC, VVD cannot be an integral part of theclock because rhythmicity itself is not affected in vvdmutants. Loss of vvd results in loss of adaptation underconstant light and insensitivity to a step-up in light intensity,a hallmark of loss of photoadaptation (Schwerdtfeger andLinden, 2001). This was also inferred from the expressionpattern of conidiation-specific genes in vvd mutants(Shrode et al., 2001). Adaptation to continuous light allowsan organism to respond to a change in light intensity. Inother words, adaptation implies that light represses light-induced genes. Because vvd is expressed only in the light,and in Neurospora this expression depends on WCC, anyfunction of VVD will depend on a functional WCC. VVDhomologues might have independent photoreceptor rolesin other species. The first detailed study outside the Neu-rospora model was carried out by Schmoll et al. (2004),who isolated a Trichoderma reesei homologue, ENVOY, byan entirely different route: in a differential screen for early
Fig. 3. White-collar 1 homologues inthree fungal phyla. Domain architectureswere generated by SMART
(http://smart.embl-heidelberg.de/). Nc,Neurospora crassa; Af, Aspergillus fumigatus;Tb, Tuber borchii; Ta, Trichoderma atroviride;Hj, Hypocrea jecorina; Pb, Phycomycesblakesleeanus; Mc, Mucor circinelloides; Cn,Cryptococcus neoformans; Cc, Coprinuscinereus.
Looking through the eyes of fungi 9
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15

cellulose-induced genes. Loss of ENVOY makes thefungus light-sensitive, although growth in the dark is unaf-fected (Schmoll et al., 2005). In the wild type, induction ofcellulase is enhanced by light, but this regulation is defec-tive in the mutant. These phenotypes suggest that, as forNeurospora VVD, ENVOY is needed to detect and respondto constant light. The output, though, seems to be quitedifferent in the two species.
Consideration of the diversity of photoresponses islikely to uncover further diversity in fungal photoreceptors.The genes corresponding to some photoresponsemutants remain to be identified, for example PhycomycesmadB and madC (Bergman et al., 1969), and Tricho-derma dimY (Horwitz et al., 1985). These may prove to bedistant relatives of the wc genes, or belong to completelydifferent classes. Photoactivation of the PhycomycesHSP100 heat shock protein gene hspA requires the WC-1orthologue madA, but also madB, and beta carotene; afterphotoadaptation hspA is still induced by heat shock(Rodriguez-Romero and Corrochano, 2006). The newfungal genome sequences will, no doubt, lead to rapididentification of the other photoreceptors, perhaps relatedto wc, or belonging to some of the other classes dis-cussed in the following sections.
Cryptochrome/photolyase and AppA/BLUF families:non-LOV flavoproteins
The only flavoproteins known to have photosensory rolesin fungi contain PAS/LOV domains. Two more, completelydifferent, photoreceptor classes detect blue light in otherorganisms. First, the cryptochrome/photolyase superfam-ily is indeed represented in fungal genomes (Table 1).Light provides the energy for DNA photorepair, catalysedby DNA photolyases. Fungal photolyase genes are them-selves regulated by blue/UV light (Berrocal-Tito et al.,1999; Alejandre-Duran et al., 2003) or exclusively UV(Kihara et al., 2004), but could cryptochrome/photolyasefamily members have sensory roles? The absorptionspectrum of plant CRY1 measured in its oxidized flavinstate (Zeugner et al., 2005) closely matches the spectralsignature of blue light responses (for example, the actionspectrum of phototropism, Fig. 2B). It might seem ironicthat this family has taken over the original cryptochromename, when PAS/LOV proteins match to the action spec-trum equally well. The genetic evidence, however, forsensory roles in Arabidopsis and Drosophila is very strong(Lin et al., 1998; Ceriani et al., 1999). In fungi, in contrast,no sensory cryptochromes are known. Knockout of theCPD photolyase of Neurospora removed DNA photore-pair but did not confer photosensory phenotypes(Shimura et al., 1999). Overexpression of the Tricho-derma photolyase gene phr1 increased the light sensitiv-ity for induction of phr1 mRNA. Deletion of the gene,
however, also increased light sensitivity (as reported byGFP under control of the phr1 promoter), so PHR1 is notthe photoreceptor for its own induction. Several otherlight-regulated genes showed altered light sensitivity, afirst indication that PHR1 might interact with a transcrip-tional regulatory complex (G. Berrocal-Tito, B.A. Horwitzand A. Herrera-Estrella, unpubl.). A hint to future experi-ments to do in fungi comes from studies on mammaliancryptochromes. In the mammalian circadian clock, cryp-tochromes inhibit CLOCK/BMAL1, a transcription factordimer which acts independently of light to increasetranscription of PER1 (Griffin et al., 1999). A circadianrhythm was found in histone H3 acetylation at theCLOCK/BMAL target genes PER1 and PER2, and intransient expression assays, cryptochromes inhibit theincrease in CLOCK/BMAL activity induced by H3 acetyla-tion (Etchegaray et al., 2003). This cryptochrome functionis light-independent, but suggests the potential useful-ness of looking for histone acetylation as a mediator oflight action through cryptochromes, as reported recentlyfor WC-1 (Grimaldi et al., 2006). Finally, no transcriptionalcomplexes containing photolyases are known. Consider-ing the sequence similarity to cryptochromes, this mightbe a promising direction to pursue.
Second, we should mention the most recent addition tothe eukaryotic photoreceptor families.Anovel flavoprotein,blue light-sensitive adenylate cyclase, was first identified inEuglena (Iseki et al., 2002). Its flavin-binding domainAppA/BLUF (Blue Light Sensing Using Flavin) was knownfrom anoxygenic photosynthetic bacteria, were AppA pro-teins photomodulate the expression of photosynthesisgenes. Strikingly, AppA is a single protein that integratesredox and light signals (Han et al., 2004). A TBLASTNsearch of the fungal genomes using AppA reveals a hypo-thetical protein (E = 2e-8, XP_756335.1) in Ustilagomaydis, but not in other fungal databases. Although theoverall similarity is not very high, all 15 residues identicalbetween prokaryotic AppA and the eukaryotic BluFdomains (Iseki et al., 2002) are also shared by theU. maydis predicted polypeptide (Fig. S1). Has an ances-tral prokaryote photoreceptor appeared only in basidi-omycetes, or do other homologues remain to be identified?Extensive photobiological research is lacking forU. maydis. Fungi, like prokaryotic microbes, must contendwith changing light and redox environments. If the asco-mycetes indeed lack BluF domain sensors, it is tempting tospeculate that PAS/LOV sensors took over this role.
Phytochromes
The fungal kingdom is not blind to light above 600 nm(Fig. 2). Plant phytochromes are photochromic sensorsthat detect light, primarily, in the red and far-red regions.Genetic evidence is very recent, but fungal phytochromes
10 A. Herrera-Estrella and B. A. Horwitz
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15

were proposed as early as 1979 (Valadon et al., 1979).Conidiation of Aspergillus nidulans is sensitive to both redand blue light (Mooney and Yager, 1990; Yager et al.,1998). Light sensitivity depends on velvet A (veA), whichencodes a novel 573 amino acid polypeptide (Kim et al.,2002). Mutation of veA as in standard laboratory strains,favours asexual sporulation, releasing conidiation fromphotocontrol. Deletion of veA in A. nidulans blocks theformation of cleistothecia, while increasing conidial pro-duction (Kim et al., 2002). VeA contains a bipartite nuclearlocalization signal in its N-terminus and in the dark islocated mainly in the nuclei, while in the light it is predomi-nantly found in the cytoplasm, as recently shown by Stin-nett et al. (2006). The nuclear localization of VeA issimilarly affected by white and blue light, with only a smalleffect of red light. The ability to respond to blue light alsodepends on the (yet uncharacterized) bliA locus. A strainwith allele fluG701 in a veA+ (bliA1) background showed adramatic reduction in conidiation in response to red light,as compared with the veA + (bliA1) strain from which it wasderived (Yager et al., 1998). FvVE1, an orthologue of VeAof Fusarium verticillioides, regulates growth and develop-ment (Li et al., 2006). The photobiologists’ guess thatfungi might have phytochrome was confirmed when twophytochrome-like Neurospora genes where identified(Table 1) that, when expressed in vitro, bind the chro-mophores and undergo complete photocycles (Froehlichet al., 2005). Fungal phytochromes, encoded in mostsequenced genomes, are closer to bacterial than to plantphytochromes (Karniol et al., 2005). Mutants of Neuro-spora and Cryptococcus phytochrome genes do not haveany known phenotype (Froehlich et al., 2005; Idnurm andHeitman, 2005b). Neurospora seems to be blind to longwavelengths, and so is not the ideal model to answer thequestion of phytochrome function. Deletion of the singlephytochrome gene of A. nidulans, fphA. resulted in theloss of red light inhibition of cleistothecium formation(Fig. 2C), the first function assigned to a specific fungalphytochrome (Blumenstein et al., 2005). In T. atroviride,red light induces the expression of several genes(Rosales-Saavedra et al., 2006). In addition, there is evi-dence suggesting that the blue-light perception system ofT. atroviride establishes a cross-talk with that involved inred light perception, which is reflected at the level ofmycelial growth and the expression of bld-8, a geneencoding a putative isoamyl alcohol oxidase (Casas-Flores et al., 2004; Rosales-Saavedra et al., 2006). Nev-ertheless, it still remains to be proven if these responsesof T. atroviride are mediated by its phytochrome.
Fungal opsins
Opsins are retinal-binding proteins, belonging to a largeand ancient group including G-protein linked receptors,
capable of absorbing light. Light absorption is in turn usedeither as a signal or for energy-conservation purposes. Asa result of a bioinformatic search, a Neurospora opsin(nop1) was identified, which when expressed and recon-stituted in vitro with retinal, undergoes a photocycle(Bieszke et al., 1999). Phylogenetic analysis could notdistinguish whether NOP1 belongs to the proton pumpingor sensory class. Archaeal sensory rhodopsins usuallyhave slower reaction cycles, and reconstituted NOP1indeed has a slow cycle. A phenotype could not be foundfor nop1 mutants, however, except in the presence of botholigomycin and light. This is a first hint that proton-pumping and light might act together to modulate conidi-ation (Bieszke et al., 1999), but the slow photocyle wouldsuggest a sensory role. One could look at other species insearch of the function(s) of fungal opsins. For example,Leptosphaeria maculans, the agent of black-leg diseaseof Brassica, has a homologue of NOP1 of unknownfunction (Idnurm and Howlett, 2001). A picture oppositeto Neurospora NOP1 emerged upon expression andreconstitution. The Leptosphaeria rhodopsin photocycleis fast, a characteristic of proton-pumping opsins, andits pH dependence and other properties are similarto bacteriorhodopsin. Indeed, the reconstituted Lep-tosphaeria rhodopsin pumps protons (Waschuk et al.,2005). Why Leptosphaeria should pump protons in thelight, and across what membrane, remains a mystery.Absence of the Gibberella or C. neoformans opsinsalso does not produce an obvious phenotype (Pradoet al., 2004; Idnurm and Heitman, 2005b). Neurosporaresponds to blue light, while retinal-reconstituted Nop-1 isa green-absorbing pigment. Fungi that ‘see’ green light(Phycomyces, Fig. 2) might provide new clues. Saranakand Foster (Saranak and Foster, 1997) concluded that arhodopsin is responsible for zoospore phototaxis of thechytrid Allomyces reticulatus. Chytrids diverged beforethe three fungal phyla that lack motile cells. In an attemptto trace the evolution of fungal opsins, it will be helpful toknow whether the Allomyces zoospore phototaxis recep-tor is closer to fungal opsins or to the algal channel-rhodopsins (Nagel et al., 2005). The question of whetherthe fungal opsins are evolutionary ‘relics’ or have a majorfunction that remains to be discovered is still open.
Concluding remarks and questions
The molecular machinery discovered by photobiologistsmight have more general importance. After phototrans-duction, the messenger created in the cell might well be atthe crossroads of different input signals. It should come asno surprise, then, that photoreceptor proteins have light-independent roles. One example is cryptochrome in themammalian circadian clock. In fungi, the much-studiedlight-sensitive transcription factor WC-1 also has light-
Looking through the eyes of fungi 11
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15

independent roles. Light and dark functions in Neurosporacan be uncoupled, to some extent: an allele resulting inlow level production of a truncated protein allows normalphotoinduction of FRQ, yet this strain is deficient in FRQin the dark (Lee et al., 2003). Likewise, a wc-1 alleleisolated in a screen for altered clock function lacks theN-terminal glutamine rich region. This allele permits lightentrainment of conidiation and a small but detectableinduction of carotene synthesis, yet the dark function inthe clock is lost (Toyota et al., 2002). Rhythmic transcrip-tion of frq, at the core of the Neurospora clock, dependson binding of the WCC to the Clock Box, an upstreamelement of frq. This occurs in a circadian time-dependentmanner, even in constant darkness (Froehlich et al.,2003). Light-independent phosphorylation of WC-1occurs at five phosphorylation sites that are not importantfor photoreception, but rather influence circadian param-eters (He et al., 2005). WCC is phosphorylated in anFRQ-dependent manner (at these, or perhaps other,sites), decreasing WCC binding to the Clock Box(Schafmeier et al., 2005). The casein kinase CK-1a asso-ciates with FRQ and mediates WCC phosphorylation, andanother kinase, CKII, may also contribute although it doesnot bind to FRQ or WCC (He et al., 2006). The dynamictranscriptional complexes that make up circadian clocksnot only provide a model for how different kinds of com-plexes might be assembled, but even suggest where tolook for the interacting proteins. Indeed, WC-1 hassequence homology to the mammalian clock transcrip-tional regulator BMAL, and the similarity is not restricted tothe PAS domains, suggesting that that the WCC andBMAL/CLOCK dimers might have parallel functions (Leeet al., 2000). The Trichoderma wc homologues alsoappear to have functions in the dark, as mutants in thecorresponding genes do not conidiate in response tocarbon deprivation in the dark (Casas-Flores et al., 2006).Light-dependent signalling by LOV domain proteins relieson their flavin cofactors. Light-independent signallingcould occur by flavin redox chemistry in the dark, as in theRhodobacter BLUF domain, but there is no direct evi-dence for this so far for WC1. Reactive oxygen speciesenhance light signalling via WC in Neurospora, althoughthe dark levels seem unaffected. It was proposed that theinteraction between redox and light signals occurs at theLOV domain (Yoshida and Hasunuma, 2004), an area forfurther study.
The WC1/2 transcription factor leads the list of well-studied fungal photoreceptors. As more photoreceptorgenes are identified in different species, we can askwhether the short list of chromophores will correspond toan equally short list of apoprotein genes, or whether thesame chromophores will act in different ways in differentspecies. Although the blue region of the spectrum providesthe dominant signal, green and red photoresponses have
often been overlooked. Can the green components of bluelight responses be explained by semiquinone or otherpartially reduced states of flavin photoreceptors? Further-more, some light responses in T. atroviride and A. nidulansappear to be regulated by both the blue and red lightcomponents of the spectrum, with different outcomes. Willthis phenomenon turn out to be the result of the cross-talkbetween a phytochrome and white collar perceptionsystems? Recent work on the classical Phycomycessystem is just on the threshold of explaining the complexityof zygomycete photobiology. Which of the many photore-ceptors encoded by fungal genomes are actually functionalin conveying information about the light environment to thecell? From the evolutionarily ancient and well-studiedopsins, through the cryptochrome/photolyase family, phy-tochromes and LOV-domain proteins, to the AppA/BLUFdomain that might be present in some but not all fungalgenomes, the genetics of fungal photoresponses shouldprovide answers to these questions.
Acknowledgements
We thank Jonathan Gressel for comments on the manuscript,and Tapani Yli-Mattila, David Dennison and Luis Corrochanofor original images.
References
Alejandre-Duran, E., Roldan-Arjona, T., Ariza, R.R., andRuiz-Rubio, M. (2003) The photolyase gene from the plantpathogen Fusarium oxysporum f. sp. lycopersici is inducedby visible light and alpha-tomatine from tomato plant.Fungal Genet Biol 40: 159–165.
Ambra, R., Grimaldi, B., Zamboni, S., Filetici, P., Macino, G.,and Ballario, P. (2004) Photomorphogenesis in the hypo-geous fungus Tuber borchii: isolation and characterizationof Tbwc-1, the homologue of the blue-light photoreceptor ofNeurospora crassa. Fungal Genet Biol 41: 688–697.
Ballario, P., Vittorioso, P., Magrelli, A., Talora, C., Cabibbo,A., and Macino, G. (1996) White collar-1, a central regula-tor of blue light responses in Neurospora, is a zinc fingerprotein. EMBO J 15: 1650–1657.
Bergman, K., Burke, P.V., Cerdá-Olmedo, E., David, C.N.,Delbrück, M., Foster, K.W., and Goodell, E.W. (1969)Phycomyces. Bacteriol Rev 33: 99–157.
Berrocal-Tito, G., Sametz-Baron, L., Eichenberg, K., Horwitz,B.A., and Herrera-Estrella, A. (1999) Rapid light regulationof a Trichoderma harzianum DNA photolyase gene. J BiolChem 274: 14288–14294.
Bieszke, J.A., Spudich, E.N., Scott, K.L., Borkovich, K.A., andSpudich, J.L. (1999) A eukaryotic protein, NOP-1, bindsretinal to form an archaeal rhodopsin-like photochemicallyreactive pigment. Biochemistry 38: 14138–14145.
Blumenstein, A., Vienken, K., Tasler, R., Purschwitz, J., Veith,D., Frankenberg-Dinkel, N., and Fischer, R. (2005) TheAspergillus nidulans phytochrome FphA repressessexual development in red light. Curr Biol 15: 1833–1838.
12 A. Herrera-Estrella and B. A. Horwitz
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15

Borkovich, K.A., Alex, L.A., Yarden, O., Freitag, M., Turner,G.E., Read, N.D., et al. (2004) Lessons from the genomesequence of Neurospora crassa: tracing the path fromgenomic blueprint to multicellular organism. Microbiol MolBiol Rev 68: 1–108.
Briggs, W.R., Christie, J.M., and Salomon, M. (2001) Pho-totropins: a new family of flavin-binding blue light receptorsin plants. Antioxid Redox Signal 3: 775–788.
Carattoli, A., Cogoni, C., Morelli, G., and Macino, G. (1994)Molecular characterization of upstream regulatory se-quences controlling the photoinduced expression of thealbino-3 gene of Neurospora crassa. Mol Microbiol 13:787–795.
Casas-Flores, S., Rios-Momberg, M., Bibbins, M., Ponce-Noyola, P., and Herrera-Estrella, A. (2004) BLR-1 andBLR-2, key regulatory elements of photoconidiation andmycelial growth in Trichoderma atroviride. Microbiology150: 3561–3569.
Casas-Flores, S., Rios-Momberg, M., Rosales-Saavedra, T.,Martinez-Hernandez, P., Olmedo-Monfil, V., and Herrera-Estrella, A. (2006) Cross talk between a fungal blue-lightperception system and the cyclic AMP signaling pathway.Eukaryot Cell 5: 499–506.
Cerdá-Olmedo, E., and Lipson, E.D. (1987) Phycomyces.Cold Spring Harbor, NY: Cold Spring Harbor Laboratory.
Ceriani, M.F., Darlington, T.K., Staknis, D., Mas, P., Petti,A.A., Weitz, C.J., and Kay, S.A. (1999) Light-dependentsequestration of timeless by cryptochrome. Science 285:553–556.
Cheng, P., He, Q., Yang, Y., Wang, L., and Liu, Y. (2003)Functional conservation of light, oxygen, or voltagedomains in light sensing. Proc Natl Acad Sci USA 100:5938–5943.
Corrochano, L.M., and Galland, P. (2006) Photomorphogen-esis and gravitropism in fungi. In: The Mycota I. Growth,Differentiation and Sexuality. Kües, U., and Fischer, R.(eds). Berlin, Heidelberg: Springer-Verlag, pp. 233–259.
Crosson, S., and Moffat, K. (2001) Structure of a flavin-binding plant photoreceptor domain: insights into light-mediated signal transduction. Proc Natl Acad Sci USA 98:2995–3000.
Crosthwaite, S.K., Dunlap, J.C., and Loros, J.J. (1997) Neu-rospora wc-1 and wc-2: transcription, photoresponses, andthe origins of circadian rhythmicity. Science 276: 763–769.
Dennison, D. (1979) Phototropism. In: Encyclopedia of PlantPhysiology, Vol. 7. Haupt, W., and Feinleib, M.E. (eds).Berlin, Heidelberg: Springer-Verlag, pp. 506–566.
Etchegaray, J.P., Lee, C., Wade, P.A., and Reppert, S.M.(2003) Rhythmic histone acetylation underlies transcriptionin the mammalian circadian clock. Nature 421: 177–182.
Franchi, L., Fulci, V., and Macino, G. (2005) Protein kinase Cmodulates light responses in Neurospora by regulating theblue light photoreceptor WC-1. Mol Microbiol 56: 334–345.
Froehlich, A.C., Liu, Y., Loros, J.J., and Dunlap, J.C. (2002)White collar-1, a circadian blue light photoreceptor, bindingto the frequency promoter. Science 297: 815–819.
Froehlich, A.C., Loros, J.J., and Dunlap, J.C. (2003) Rhyth-mic binding of a white collar-containing complex to thefrequency promoter is inhibited by frequency. Proc NatlAcad Sci USA 100: 5914–5919.
Froehlich, A.C., Noh, B., Vierstra, R.D., Loros, J., andDunlap, J.C. (2005) Genetic and molecular analysis ofphytochromes from the filamentous fungus Neurosporacrassa. Eukaryot Cell 4: 2140–2152.
Galland, P., and Lipson, E.D. (1987) Blue-light reception inPhycomyces phototropism: evidence for two photosystemsoperating in low- and high-intensity ranges. Proc Natl AcadSci USA 84: 104–108.
Gressel, J., and Rau, W. (1983) Photocontrol of fungaldevelopment. In: Encyclopedia of Plant Physiology. Shrop-shire W., Jr, and Mohr, H. (eds). Berlin, Heidelberg:Springer-Verlag, pp. 603–639.
Griffin, E.A., Jr, Staknis, D., and Weitz, C.J. (1999) Light-independent role of CRY1 and CRY2 in the mammaliancircadian clock. Science 286: 768–771.
Grimaldi, B., Coiro, P., Filetici, P., Berge, E., Dobosy, J.R.,Freitag, M., et al. (2006) The Neurospora crassa WC-1-dependent blue light response requires acetylation ofhistone H3 lysine 14 by NGF-1. Mol Biol Cell 17: 4576–4583.
Han, Y., Braatsch, S., Osterloh, L., and Klug, G. (2004) Aeukaryotic BLUF domain mediates light-dependent geneexpression in the purple bacterium Rhodobacter Sphaeroi-des 2.4.1. Proc Natl Acad Sci USA 101: 12306–12311.
He, Q., and Liu, Y. (2005) Molecular mechanism of lightresponses in Neurospora: from light-induced transcriptionto photoadaptation. Genes Dev 19: 2888–2899.
He, Q., Shu, H., Cheng, P., Chen, S., Wang, L., and Liu, Y.(2005) Light-independent phosphorylation of white collar-1regulates its function in the Neurospora circadian negativefeedback loop. J Biol Chem 280: 17526–17532.
He, Q., Cha, J., He, Q., Lee, H.C., Yang, Y., and Liu, Y.(2006) CKI and CKII mediate the frequency-dependentphosphorylation of the white collar complex to close theNeurospora circadian negative feedback loop. Genes Dev20: 2552–2565.
Heintzen, C., Loros, J.J., and Dunlap, J.C. (2001) The PASprotein VIVID defines a clock-associated feedback loopthat represses light input, modulates gating, and regulatesclock resetting. Cell 104: 453–464.
van der Horst, M.A., and Hellingwerf, K.J. (2004) Photore-ceptor proteins, ‘star actors of modern times’: a review ofthe functional dynamics in the structure of representativemembers of six different photoreceptor families. Acc ChemRes 37: 13–20.
Horwitz, B.A., Gressel, J., Malkin, S., and Epel, B.L. (1985)Modified cryptochrome in vivo absorption in dim photo-sporulation mutants of Trichoderma. Proc Natl Acad SciUSA 82: 2736–2740.
Idnurm, A., and Heitman, J. (2005a) Photosensing fungi:phytochrome in the spotlight. Curr Biol 15: R829–R832.
Idnurm, A., and Heitman, J. (2005b) Light controls growthand development via a conserved pathway in the fungalkingdom. PLoS Biol 3: e95.
Idnurm, A., and Howlett, B.J. (2001) Characterization of anopsin gene from the ascomycete Leptosphaeria maculans.Genome 44: 167–171.
Idnurm, A., Rodriguez-Romero, J., Corrochano, L.M., Sanz,C., Iturriaga, E.A., Eslava, A.P., and Heitman, J. (2006)The Phycomyces madA gene encodes a blue-light photo-
Looking through the eyes of fungi 13
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15

receptor for phototropism and other light responses. ProcNatl Acad Sci USA 103: 4546–4551.
Iseki, M., Matsunaga, S., Murakami, A., Ohno, K., Shiga, K.,Yoshida, K., et al. (2002) A blue-light-activated adenylylcyclase mediates photoavoidance in Euglena gracilis.Nature 415: 1047–1051.
Karniol, B., Wagner, J.R., Walker, J.M., and Vierstra, R.D.(2005) Phylogenetic analysis of the phytochromesuperfamily reveals distinct microbial subfamilies ofphotoreceptors. Biochem J 392: 103–116.
Kihara, J., Moriwaki, A., Matsuo, N., Arase, S., and Honda, Y.(2004) Cloning, functional characterization, and near-ultraviolet radiation-enhanced expression of a photolyasegene (PHR1) from the phytopathogenic fungus Bipolarisoryzae. Curr Genet 46: 37–46.
Kim, H., Han, K., Kim, K., Han, D., Jahng, K., and Chae, K.(2002) The veA gene activates sexual development inAspergillus nidulans. Fungal Genet Biol 37: 72–80.
Lee, K., Loros, J.J., and Dunlap, J.C. (2000) Interconnectedfeedback loops in the Neurospora circadian system.Science 289: 107–110.
Lee, K., Dunlap, J.C., and Loros, J.J. (2003) Roles for whitecollar-1 in circadian and general photoperception inNeurospora crassa. Genetics 163: 103–114.
Li, S., Myung, K., Guse, D., Donkin, B., Proctor, R.H., Gray-burn, W.S., and Calvo, A.M. (2006) FvVE1 regulates fila-mentous growth, the ratio of microconidia to macroconidiaand cell wall formation in Fusarium verticillioides. MolMicrobiol 62: 1418–1432.
Lin, C., Yang, H., Guo, H., Mockler, T., Chen, J., and Cash-more, A.R. (1998) Enhancement of blue-light sensitivity ofArabidopsis seedlings by a blue light receptor crypto-chrome 2. Proc Natl Acad Sci USA 95: 2686–2690.
Linden, H., and Macino, G. (1997) White collar 2, a partner inblue-light signal transduction, controlling expression oflight-regulated genes in Neurospora crassa. EMBO J 16:98–109.
Lipson, E., and Horwitz, B. (1991) Photosensory receptorsand their transduction. In: Sensory Receptors and SignalTransduction. Modern Cell Biology, Vol. 7. Satir, B. (ed.).New York: Wiley Liss, pp. 1–64.
Lu, Y.K., Sun, K.H., and Shen, W.C. (2005) Blue light nega-tively regulates the sexual filamentation via the Cwc1 andCwc2 proteins in Cryptococcus neoformans. Mol Microbiol56: 480–491.
Mooney, J.L., and Yager, L.N. (1990) Light is required forconidiation in Aspergillus nidulans. Genes Dev 4: 1473–1482.
Nagel, G., Szellas, T., Kateriya, S., Adeishvili, N., Hegemann,P., and Bamberg, E. (2005) Channelrhodopsins: directlylight-gated cation channels. Biochem Soc Trans 33: 863–866.
Prado, M.M., Prado-Cabrero, A., Fernandez-Martin, R., andAvalos, J. (2004) A gene of the opsin family in the caro-tenoid gene cluster of Fusarium fujikuroi. Curr Genet 46:47–58.
Purschwitz, J., Muller, S., Kastner, C., and Fischer, R. (2006)Seeing the rainbow: light sensing in fungi. Curr Opin Micro-biol 9: 566–571.
Rodriguez-Romero, J., and Corrochano, L.M. (2006) Regu-lation by blue light and heat shock of gene transcription in
the fungus Phycomyces: proteins required for photoinduc-tion and mechanism for adaptation to light. Mol Microbiol61: 1049–1059.
Rosales-Saavedra, T., Esquivel-Naranjo, E.U., Casas-Flores, S., Martinez-Hernandez, P., Ibarra-Laclette, E.,Cortes-Penagos, C., and Herrera-Estrella, A. (2006) Novellight-regulated genes in Trichoderma atroviride: a dissec-tion by cDNA microarrays. Microbiology 152: 3305–3317.
Saranak, J., and Foster, K.W. (1997) Rhodopsin guidesfungal phototaxis. Nature 387: 465–466.
Sasaki, T., Honda, Y., Umekawa, M., and Nemoto, M. (1985)Control of certain diseases of greenhouse vegetables withultraviolet-absorbing vinyl film. Plant Dis 69: 530–533.
Schafmeier, T., Haase, A., Kaldi, K., Scholz, J., Fuchs, M.,and Brunner, M. (2005) Transcriptional feedback ofNeurospora circadian clock gene by phosphorylation-dependent inactivation of its transcription factor. Cell 122:235–246.
Schmidt, W., and Galland, P. (1999) Light-induced absor-bance changes in Phycomyces: evidence for cryptochrome-associated flavosemiquinones. Planta 208: 274–282.
Schmoll, M., Zeilinger, S., Mach, R.L., and Kubicek, C.P.(2004) Cloning of genes expressed early during cellulaseinduction in Hypocrea jecorina by a rapid subtractionhybridization approach. Fungal Genet Biol 41: 877–887.
Schmoll, M., Franchi, L., and Kubicek, C.P. (2005) Envoy, aPAS/LOV domain protein of Hypocrea jecorina (AnamorphTrichoderma reesei), modulates cellulase gene transcrip-tion in response to light. Eukaryot Cell 4: 1998–2007.
Schwerdtfeger, C., and Linden, H. (2000) Localization andlight-dependent phosphorylation of white collar 1 and 2, thetwo central components of blue light signaling in Neuro-spora crassa. Eur J Biochem 267: 414–422.
Schwerdtfeger, C., and Linden, H. (2001) Blue light adapta-tion and desensitization of light signal transduction in Neu-rospora crassa. Molec Microbiol 39: 1080–1087.
Schwerdtfeger, C., and Linden, H. (2003) VIVID is a flavopro-tein and serves as a fungal blue light photoreceptor forphotoadaptation. EMBO J 22: 4846–4855.
Shimura, M., Ito, Y., Ishii, C., Yajima, H., Linden, H.,Harashima, T., et al. (1999) Characterization of a Neuro-spora crassa Photolyase-deficient mutant generated byrepeat induced point mutation of the phr gene. FungalGenet Biol 28: 12–20.
Shrode, L.B., Lewis, Z.A., White, L.D., Bell-Pedersen, D., andEbbole, D.J. (2001) vvd is required for light adaptation ofconidiation-specific genes of Neurospora crassa, but notcircadian conidiation. Fungal Genet Biol 32: 169–181.
Silva, F., Torres-Martinez, S., and Garre, V. (2006) Distinctwhite collar-1 genes control specific light responses inMucor circinelloides. Molec Microbiol 61: 1023–1037.
Stinnett, S.M., Espeso, E.A., Cobeno, L., Araujo-Bazan, L.,and Calvo, A.M. (2006) Aspergillus nidulans VeA subcellu-lar localization is dependent on the importin alpha carrierand on light. Molec Microbiol 63: 242–255.
Taylor, B.L., and Zhulin, I.B. (1999) PAS domains: internalsensors of oxygen, redox potential, and light. Microbiol MolBiol Rev 63: 479–506.
Terashima, K., Yuki, K., Muraguchi, H., Akiyama, M., andKamada, T. (2005) The dst1 gene involved in mushroomphotomorphogenesis of Coprinus cinereus encodes a
14 A. Herrera-Estrella and B. A. Horwitz
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15

putative photoreceptor for blue light. Genetics 171: 101–108.
Toyota, K., Onai, K., and Nakashima, H. (2002) A new wc-1mutant of Neurospora crassa shows unique light sensitivityin the circadian conidiation rhythm. Mol Genet Genomics268: 56–61.
Valadon, L., Osman, M., and Mummery, R.S. (1979) Phyto-chrome mediated carotenoid synthesis in the fungus Ver-ticillium agaricinum. Photochem Photobiol 29: 605–607.
Waschuk, S.A., Bezerra, A.G., Jr, Shi, L., and Brown, L.S.(2005) Leptosphaeria rhodopsin: bacteriorhodopsin-likeproton pump from a eukaryote. Proc Natl Acad Sci USA102: 6879–6883.
Yager, L.N., Lee, H.O., Nagle, D.L., and Zimmerman, J.E.(1998) Analysis of fluG mutations that affect light-dependent conidiation in Aspergillus nidulans. Genetics149: 1777–1786.
Yasuhira, S., and Yasui, A. (1992) Visible light-inducible pho-tolyase gene from the goldfish Carassius auratus. J BiolChem 267: 25644–25647.
Yli-Mattila, T. (1987) The effect of UV-A light on cAMP level inthe basidiomycete Schizophyllum commune. Physiol Plant69: 451–455.
Yoshida, Y., and Hasunuma, K. (2004) Reactive oxygen
species affect photomorphogenesis in Neurospora crassa.J Biol Chem 279: 6986–6993.
Zeugner, A., Byrdin, M., Bouly, J.P., Bakrim, N., Giovani, B.,Brettel, K., and Ahmad, M. (2005) Light-induced electrontransfer in Arabidopsis cryptochrome-1 correlates with invivo function. J Biol Chem 280: 19437–19440.
Supplementary material
The following supplementary material is available for thisarticle online:Fig. S1. A hypothetical fungal BIuF domain. AppA, aminoacids 14–110 of the AppA domain of the Rhodobactersphaeroides redox regulator (LA2555) were used to searchthe fungal databases. Um, Ustilago maydis XP_756335.1.Conserved residues (see Iseki et al., 2002) are highlighted.
This material is available as part of the online article fromhttp://www.blackwell-synergy.comPlease note: Blackwell Publishing is not responsible for thecontent or functionality of any supplementary materials sup-plied by the authors. Any queries (other than missing mate-rial) should be directed to the corresponding author for thearticle.
Looking through the eyes of fungi 15
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 64, 5–15