
Long-Term Blockade of Cholecystokinin (CCK): Effects of L 364,718 (a CCK Receptor Antagonist) on...
9
Pnncrcus Vol. 15, No. 3, pp. 3 14-322 0 1997 Lippincott-Raven Publishers, Philadelphia L Long-Term Blockade of Cholecystokinin (CCK): Effects of 364,7 18 (a CCK Receptor Antagonist) on Pancreatic Enzyme Storage and Secretion Ana I. Rodriguez, Manuel A. Manso, Andres C. Garcia-Montero, *Albert0 Orfao, and Isabel de Dios Department of Physiology and Pharmacology, Facultad de Biologia, Edifcio Departamental, and *Flow Cytometry Service, University of Salamanca, Salamanca, Spain Summary: To determine the effect of long-term blockade of cholecystokinin (CCK) on both pancreatic storage and secre- tion processes, L 364,718 (a CCK receptor antagonist) was administered to rats at 0.1 mg/kg/day for 3, 7, and 15 days. Zymogen granules were analyzed by flow cytometry to deter- mine their light scattering properties-forward scatter and side scatter-as well as their amylase content measured by a spe- cific antiserum. The mean number of zymogen granules per cell was counted on pancreatic sections using electron microscopy. DNA content, pancreatic weight, and enzyme secretion were also studied under both basal conditions and CCK infusion at a dose of 1.25 p g k g h , which is able to displace the CCK re- ceptor antagonist. Two subpopulations of zymogen granules (Z, and Z,) were identified on the basis of their light scattering parameters, in both control and L 364,718-treated rats. L 364,718 administered for 3, 7, and 15 days induced a signifi- cant reduction in the amylase content of individual zymogen granules, for both 2, and Z, zymogen granule subsets. In con- trast, the number of zymogen granules per cell increased from It is well established that long-term administration of exogenous cholecystokinin (CCK) or its analogue ceru- lein increases the pancreatic content of both DNA and protein (1-3). Similar effects have been observed in ex- perimental situations in which endogenous CCK was in- creased such as feeding with trypsin inhibitors (4) or bile-pancreatic juice diversion (5). Different CCK receptor antagonists have been used to determine specifically the physiological actions of CCK. L 364,718 has been found to be the most effective CCK receptor blocker in different experimental models (6). Manuscript received August 6, 1996; revised manuscript accepted January 22, 1997. Address correspondence and reprint requests to Dra. 1. de Dios, Dep. Fisiologia y Farmacologia, Edificio Departmental, Campus Miguel de Unamuno, 37007 Salamanca, Spain. day +3 of treatment onward, the highest values being detected at day +7. Hyperplasia was observed only at day + 15. Basal enzyme secretion decreased significantly in rats treated with L 364,718 for 3 and 7 days but recovered to control values after 15 days of treatment. No significant differences in CCK- stimulated amylase secretion were observed between control and L 364,7 18-treated rats. At day + 15 of L 364,7 18 treatment a significant increase in enzyme secretion was observed with respect to shorter treatment periods; this was associated with a significant increase in both the number of cells and the number of zymogen granules per cell. Our results indicate that chronic administration of L 364,7 I8 induces a biphasic effect on pan- creatic function. Interestingly, although enzyme secretion reached recovery after long-term treatment (15 days), the stor- age process is altered since the mean enzyme content in each individual zymogen granule remains significantly reduced. Key Words: Cholecystokinin (CCK) receptor antagonist-CCK- Exocrine pancreas-Zymogen granules-Rat. Accordingly, it has been reported that acute administra- tion of L 364,718 is able to inhibit basal (7) and CCK- stimulated pancreatic secretion (8,9). CCK receptor an- tagonists administered for long periods have been used mainly to study the role of endogenous CCK in the de- velopment and growth of the pancreas. In this sense it has been observed that administration of L 364,718, alone, for 14 days either decreased (10) or failed to alter the normal growth of the pancreas (1 1,12). However, few studies have been devoted to analyzing the effect of chronic administration of L 364,718 on other CCK- related functions, such as pancreatic enzyme storage and secretion. Previously, it has been reported that basal but not CCK-stimulated pancreatic secretion was blocked af- ter 7 days of L 364,718 administration in rats (13,14). However, there are no specific references on its effects 314
Transcript of Long-Term Blockade of Cholecystokinin (CCK): Effects of L 364,718 (a CCK Receptor Antagonist) on...

Pnncrcus Vol. 15, No. 3, pp. 3 14-322 0 1997 Lippincott-Raven Publishers, Philadelphia
L Long-Term Blockade of Cholecystokinin (CCK): Effects of 364,7 18 (a CCK Receptor Antagonist) on Pancreatic Enzyme
Storage and Secretion
Ana I. Rodriguez, Manuel A. Manso, Andres C. Garcia-Montero, *Albert0 Orfao, and Isabel de Dios
Department of Physiology and Pharmacology, Facultad de Biologia, Edifcio Departamental, and *Flow Cytometry Service, University of Salamanca, Salamanca, Spain
Summary: To determine the effect of long-term blockade of cholecystokinin (CCK) on both pancreatic storage and secre- tion processes, L 364,718 (a CCK receptor antagonist) was administered to rats at 0.1 mg/kg/day for 3, 7, and 15 days. Zymogen granules were analyzed by flow cytometry to deter- mine their light scattering properties-forward scatter and side scatter-as well as their amylase content measured by a spe- cific antiserum. The mean number of zymogen granules per cell was counted on pancreatic sections using electron microscopy. DNA content, pancreatic weight, and enzyme secretion were also studied under both basal conditions and CCK infusion at a dose of 1.25 p g k g h , which is able to displace the CCK re- ceptor antagonist. Two subpopulations of zymogen granules (Z, and Z,) were identified on the basis of their light scattering parameters, in both control and L 364,718-treated rats. L 364,718 administered for 3, 7, and 15 days induced a signifi- cant reduction in the amylase content of individual zymogen granules, for both 2, and Z, zymogen granule subsets. In con- trast, the number of zymogen granules per cell increased from
It is well established that long-term administration of exogenous cholecystokinin (CCK) or its analogue ceru- lein increases the pancreatic content of both DNA and protein (1-3). Similar effects have been observed in ex- perimental situations in which endogenous CCK was in- creased such as feeding with trypsin inhibitors (4) or bile-pancreatic juice diversion (5).
Different CCK receptor antagonists have been used to determine specifically the physiological actions of CCK. L 364,718 has been found to be the most effective CCK receptor blocker in different experimental models (6).
Manuscript received August 6, 1996; revised manuscript accepted January 22, 1997.
Address correspondence and reprint requests to Dra. 1. de Dios, Dep. Fisiologia y Farmacologia, Edificio Departmental, Campus Miguel de Unamuno, 37007 Salamanca, Spain.
day +3 of treatment onward, the highest values being detected at day +7. Hyperplasia was observed only at day + 15. Basal enzyme secretion decreased significantly in rats treated with L 364,718 for 3 and 7 days but recovered to control values after 15 days of treatment. No significant differences in CCK- stimulated amylase secretion were observed between control and L 364,7 18-treated rats. At day + 15 of L 364,7 18 treatment a significant increase in enzyme secretion was observed with respect to shorter treatment periods; this was associated with a significant increase in both the number of cells and the number of zymogen granules per cell. Our results indicate that chronic administration of L 364,7 I8 induces a biphasic effect on pan- creatic function. Interestingly, although enzyme secretion reached recovery after long-term treatment (15 days), the stor- age process is altered since the mean enzyme content in each individual zymogen granule remains significantly reduced. Key Words: Cholecystokinin (CCK) receptor antagonist-CCK- Exocrine pancreas-Zymogen granules-Rat.
Accordingly, it has been reported that acute administra- tion of L 364,718 is able to inhibit basal (7) and CCK- stimulated pancreatic secretion (8,9). CCK receptor an- tagonists administered for long periods have been used mainly to study the role of endogenous CCK in the de- velopment and growth of the pancreas. In this sense it has been observed that administration of L 364,718, alone, for 14 days either decreased (10) or failed to alter the normal growth of the pancreas (1 1,12). However, few studies have been devoted to analyzing the effect of chronic administration of L 364,718 on other CCK- related functions, such as pancreatic enzyme storage and secretion. Previously, it has been reported that basal but not CCK-stimulated pancreatic secretion was blocked af- ter 7 days of L 364,718 administration in rats (13,14). However, there are no specific references on its effects
314

BLOCKADE OF CHOLECYSTOKININ IN EXOCRINE PANCREAS 315
on zymogen granules, although the antagonist’s blocking action could have an important influence on this cellular compartment. Recent studies carried out in control (15) and adrenalectomized (16) rats have shown that flow cytometry is a sensitive and useful technique for study- ing the physical characteristics of zymogen granules and their enzymatic content. Flow cytometric analysis of the intragranular enzyme content has been validated after comparison with biochemical methods (1 7).
The aim of the current investigation was to determine if prolonged blockade of endogenous CCK by continu- ous administration of L 364,718 would alter the storage and secretion processes in the rat pancreas. For that pur- pose both the number and the enzymatic content of in- dividual zymogen granules, as well as pancreatic growth and secretion under basal and CCK-stimulated condi- tions, were sequentially analyzed.
MATERIALS AND METHODS
Products and companies L 364,718, a CCK receptor antagonist, was kindly
donated by Merck Sharp and Dohme (Madrid, Spain). Sucrose (Merck), CCK-8 sulfated (fragment 26-33 am- ide, 99% purity), dimethylsulfoxide (DMSO), ethyl- eneglycol-bis-(@-amhoethylether)-N, N, N’, N’-tet- raacetic acid (EGTA), 3-(N-morpholino)propanesulfonic acid (MOPS), phenylmethylsulfonyl fluoride (PMSF), 2-(N-morpholino)ethanesulfonic acid (MES), Percoll, rabbit antiamylase antiserum, goat anti-rabbit IgG immu- noglobulin labeled with phycoerythrin, diphenylamine, deoxyribonucleic acid from thymus, bovine serum albu- min (BSA), and maltose were purchased from Sigma Chemical Co (St. Louis, MO, U.S.A.).
Animals and treatments Male Wistar rats weighing about 250 g were used.
Rats were fed ad libitum with a standard laboratory diet (Panlab rat chow) and had free access to water; they were maintained at 24°C on a 12-h light-dark cycle. Subcu- taneous injections of 0. I mg/kg/day of L 364,718 in 10% DMSO as depot carrier or 10% DMSO in 0.15 M NaCl solution (controls) were administered for 3, 7, and 15 days. Each treatment was applied to two groups of a n - mals simultaneously to analyze pancreatic secretion and overall DNA content in one and pancreatic tissue sec- tions and zymogen granules in the other. Body weight was monitored before treatment and at days 3 ,7 , and 15. Twelve hours after the last injection, and while fasting, the animals used to study pancreatic secretion were sur- gically prepared for the collection of pancreatic juice as detailed below and the pancreata were removed to ana- lyze the DNA content. The rest of the animals were
killed by decapitation and their pancreata were quickly removed, freed from fat and lymph nodes, and weighed. A portion of pancreatic tissue was prepared for electron microscopy studies and the rest was used to study zy- mogen granules by flow cytometry.
Isolation of zymogen granules Pancreata from five animals were pooled and put on
ice-cold homogenization medium containing 250 mM sucrose, 5 mM MOPS (pH 7.0), 0.1 mM MgSO,, and 0.1 mM PMSF at 40 ml/g wet weight pancreas. They were finely minced with scissors and homogenized for 30 s in an electric homogenizer (Omni 2000 International) at low speed. Heavy components were removed by cen- trifugation at low speed. Zymogen granules were isolated according to the method of De Lisle et al. (18) by cen- trifugation at 100,000 g for 20 min in a 40% Percoll gradient composed of a homogenization buffer, pH 5.5 (250 mM sucrose, 50 mM MES, 0.1 mM MgSO,, 0.1 mM PMSF, and 2 mM EGTA) mixed with Percoll.
Staining of zymogen granules with antiamylase antiserum
Specific intragranular staining was carried out ac- cording to a previously reported method (15). Briefly, zymogen granules were incubated for 15 min with a rabbit antiamylase antiserum at a saturation concentra- tion (1.40 pglpl). The unbound antiserum was removed by washing twice (2 x 1,300 g for 15 min) in buffer pH 6.5 (0.3 M sucrose, 0.1 mM MgSO,, 2 mM MOPS, 1 mM EGTA), the pellet was resuspended and incubated in the dark for 15 Inin with a goat anti-rabbit immunoglobulin antiserum labeled with phycoerythrin at a concentration (1 x lo-* pg/pl) able to saturate the rabbit antiserum. To remove the unbound antiserum, washing medium was added and zymogen granules were washed twice (2 x 1,300 g for 15 min). Previous studies (15) have demon- strated that the amount of fluorescence is significantly reduced when zymogen granules are either fixed and permeabilized or permeabilized without fixation. The highest staining level was obtained with intact zymogen granules in accordance with findings reported by Cabana et al. (19). No significant loss of amylase was observed in the supernatants of the centrifugation steps during sample preparation for flow cytometry.
Flow cytometric analysis of zymogen granules All flow cytometry measurements of zymogen gran-
ules were performed for at least 10,000 eventshest of a pooled pancreas of five animals per experiment. A FACScan flow cytometer (BectodDickinson, San Jose, CA, U.S.A.) equipped with an argon ion laser tuned at 488 nm and 15 mW was used, as described previously
Paiicreus, Vol. 15, No. 3, 1997
316 A. I. RODRIGUEZ ET AL.
(1 5,16). Sample (medium washing)/sheath (Facs Flow, BectodDickinson) differential pressure was set at low speed to obtain a minimum diameter of the sample part of the flow at the intersection of the laser beam. During the whole measurement process the zymogen granules remained resuspended in the washing buffer, which was surrounded by the sheath fluid, and no admixture be- tween the two fluids occurred, in accordance with the principle of analysis by FACScan flow cytometer (20). Calibration of the instrument was performed on a daily basis using Calibrite beads (BectodDickinson). Fine ad- justments were performed using unstained zymogen granules. Each day zymogen granules from control and L 364,7 1 8-treated rats were measured in parallel. The Ly- sis I1 software program was used for data acquisition and analysis. Results are expressed as the mean value of at least six experiments. In each experiment the mean value per individual zymogen granule was calculated using Ly- sis 11 software.
Electron microscopic studies For transmission electron microscopic examinations, a
portion of pancreatic tissue of the different experimental groups was prefixed with 0.1 M phosphate buffer (pH 7.4) containing 2% glutaraldehyde at 4°C for 2 h. Pieces were postfixed with 1.5% OsO, in the same buffer, de- hydrated in ethanol, and finally embedded in epon. The ultrathin sections were stained with uranyl acetate and lead citrate. The samples were examined with a Zeis electron microscope (EM 900; Germany). At least 1,000 granules were counted and the number of granules per cell was considered from the number of zymogen gran- uleshumber of nuclei ratio.
Collection of pancreatic juice Rats were anesthetized with sodium pentobarbital (i.p.
injection of 3 mg/100 g body weight). After trache- otomy, a median laparotomy was performed, and the bile duct was cannulated at its exit into the duodenum to collect pure pancreatic juice. The jugular vein was cath- eterized for infusion of 0.15 M NaCl containing 0.2% BSA over 30 min (basal period) and for infusion of 1.25 pg/kg/h of CCK-8 resuspended in 0.15 M NaCl contain- ing 0.2% BSA over 30 min thereafter. Infusion was car- ried out with a peristaltic pump (LKB Model 2132) at a flow rate of 0.5 mlh . The animals were unconscious during the entire juice collection period and the body temperature was kept at 37 & 1°C by placing the rats on a heating pad and monitoring rectal temperature with a thermometer. Pancreatic juice was collected in pre- weighed Eppendorf tubes on ice to preserve the enzyme content. Finally, the pancreas was removed for measure- ment of its DNA content.
Biochemical assays Pancreatic juice volume was estimated by direct weigh-
ing on an electronic balance assuming the density of the juice to be 1 .0 g/ml. Total protein concentrations were de- termined by the Bradford method (21). Amylase and tryp- sin activity in pancreatic juice was measured immediately according to the methods of Noelting and Bernfeld (22) and de Dios and Manso (13), respectively. Pancreatic DNA content was determined by the Burton method (23).
Statistical analysis Results are expressed as means f SEM. Statistical
analysis of the results of different experimental groups was carried out using the analysis of variance (ANOVA) followed by the Scheffk test in those cases where sig- nificant differences were found by ANOVA. An un- paired Student's t test was applied to the results obtained in the two subpopulations of zymogen granules in each experimental group and in those related to body weight gain. In all cases p values <0.05 were considered to be statistically significant.
RESULTS
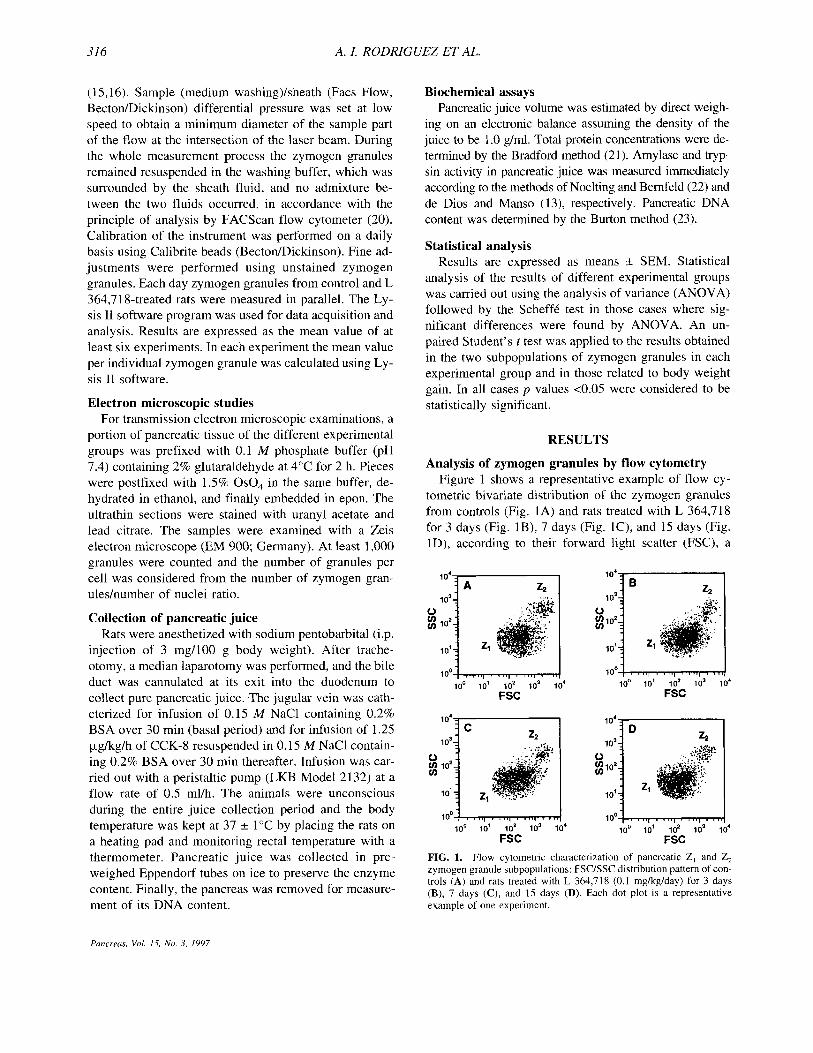
Analysis of zymogen granules by flow cytometry Figure 1 shows a representative example of flow cy-
tometric bivariate distribution of the zymogen granules from controls (Fig. 1A) and rats treated with L 364,718 for 3 days (Fig. IS), 7 days (Fig. lC), and 15 days (Fig. lD), according to their forward light scatter (FSC), a
loo 10' lo2 lo3 10' FSC
1 0 0 1 - I . . . . . 10" 10' lo2 10' 10'
FSC
loo 10' lo2 10' 10' FSC
loo 10' lo2 10' 10' FSC
FIG. 1. Flow cytometric characterization of pancreatic 2, and Z, zymogen granule subpopulations: FSC/SSC distribution pattern of con- trols (A) and rats treated with L 364,718 (0.1 mgkg/day) for 3 days (B), 7 days (C), and 15 days (D). Each dot plot is a representative example of one experiment.
Pancrrus, Vol. 15, N o . 3, 1997

BLOCKADE OF CHOLECYSTOKININ IN EXOCRINE PANCREAS 317
parameter related to the size of the zymogen granule (24), and their side light scatter (SSC), which increases according to internal complexity (25). On the basis of both parameters, two well-differentiated populations of zymogen granules were identified in control and L 364,718-treated rats: a major (Z,) and a minor (2,) sub- set. Detailed analysis of both the Z, and the Z, zymogen granule subsets (Fig. 2) showed that in all experimental groups the two subpopulations were significantly differ- ent according to both FSC and SSC parameters. The mean FSC and SSC values of each individual zymogen granule of the Z, zymogen granule subset are signifi- cantly higher (p < 0.001) than those obtained for the Z, subpopulation. A significant (p < 0.001) decrease in the FSC value of Z, zymogen granules was observed in L 364,718-treated rats for 15 days, while the FSC of Z, zymogen granules was significantly increased (p < 0.05) at day +3 of treatment. In both zymogen granule subsets, the mean SSC value was significantly increased (p < 0.01) after 7 days of L 364,718 administration (Fig. 2). Analysis of the amylase content using a specific antise-
A 1000
-. U k? 500
0 ZI 22 1. CONTROLS I
3 days L364,718
r&p'o.ool
(3 500 N , 400
300
v, 200
100
0 21 22
FIG. 2. FSC and SSC of zymogen granules (ZG) in the Z, and Z, subsets from pancreata of control rats (n = 12) and rats treated with L 364,718 (0.1 mg/kg/day) for 3 days ( n = 6), 7 days ( n = 6), and 15 days ( n = 6). n = number of experiments (five pooled pancreata per each experiment). Values are expressed as mean channels f SEM (ar- bitrary units scaled from 0 to 10,000). ANOVA followed by Scheffe test was applied in each ZG subset to compare the experimental groups, resulting in significant differences: (+ ) p < 0.05, (+ +) p < 0.01, and ( + + +) p < 0.001 cornpared with controls; (+) p < 0.01, (++) p < 0.01, and (+++) p < 0.001 compared with 3-day L 364,718-treated rats; and (**) p < 0.01 compared with 7-day L 364,718-treated rats. An unpaired Student 1 test was applied in each experimental group to compare the Z , and Z, subsets.
700 7 I pco-ool I LD 600
500 -. W 400 v) 5 300 >
200 4
100
0
3 days L364,718
0 15 days L364,718 % 600
x 500
U v) 400 LL
300
200
c
W
5 loo 0
21 22 FIG. 3. Flow cytometrjc analysis of amylase content per zymogen granule (ZG) and amylase contenWFSC ratio in Z, and Z, subsets from pancreata of control rats (n = 12) and rats treated with L 364,718 (0.1 mgkg/day) for 3 days (n = 6), 7 days (n = 6), and 15 days (n = 6). n = number of experiments (five pooled pancreata per each experi- ment). Values are means f SEM (arbitrary units scaled from 0 to 10,000). ANOVA followed by Scheff6 test was applied in each ZG population to compare the experimental groups, resulting in significant differences: (+ +) p < 0.01 and (+ + +) p < 0.001 compared with control rats. An unpaired Student t test was applied in each experimen- tal group to compare the Z, and Z , subsets.
rum (Fig. 3) revealed a significant decrease (p < 0.001) in the amount of this enzyme per individual zymogen granule from day +3 onward of treatment with L 364,718 both the Z, and the Z, subsets. Similar results were ob- tained when the amylase content per granule/FSC ratio was considered. In all experimental groups the mean amy- lase content per individual zymogen granule proved to be significantly higher in the Z, than in the Z, granules.
Analysis of zymogen granules by electron microscopy
Figure 4 shows pancreatic sections of the animal groups analyzed by electron microscopy. The mean num- ber of zymogen granules per cell increased in rats treated with L 364,7 18 from the third day of treatment, reaching significantly (p < 0.001) higher values at days +7 and +15 (Fig. 5).
Analysis of body weight gain, pancreatic weight, DNA content, and pancreatic secretion
Body weight gain was similar in control and L 364,718-treated rats. L 364,718 treatment did not signifi- cantly alter the pancreatic weight. In contrast, a signifi- cant increase (p < 0.01) in pancreatic DNA content was
Pancreas, Vol. 15, N o . 3, 1997

318 A. 1. RODRIGUEZ ET AL.
FIG. 4. Original magnification, 9,000~.
Pancreatic sections from control rats (A) and rats treated with L 364,718 (0.1 mglkglday) for 3 days (B), 7 days (C), and 15 days (D).
observed in rats treated with L 364,718 for 15 days in comparison to controls, as well as to rats treated for 3 and 7 days (Table 1).
Figure 6 shows the results obtained from the analysis of the basal pancreatic secretion. A progressive decrease in protein, amylase, and trypsin output was observed after 3 and 7 days of L 364,718 administration, but the secretion returned to control values after 15 days of treat- ment. Similar results were observed when protein and amylase output/DNA ratio were taken into account.
Upon analyzing the pancreatic secretion under stimu- lation with an intravenous infusion of CCK-8 (Fig. 7), a significant decrease 0, < 0.05) in the overall protein out- put was observed in rats treated with L 364,718 for 7
days. Interestingly, both total protein and amylase out- puts were significantly higher (p < 0.05) in rats treated with L 364,7 I8 for 15 days than in those treated for 3 and 7 days. The latter effect was not observed when the en- zyme secretionDNA ratio was considered.
DISCUSSION
L 364,7 18 is the most potent CCK antagonist capable of selectively blocking pancreatic CCK receptors, with no appreciable affinity for brain CCK receptors and gas- tric gland gastrin receptors, that is devoid of CCK-like agonist activity (26). The dose of L 364,718 used in the present study, administered for 7 days, is able to block
Pancreu\. Vol. IS, No. 3, 1997

BLOCKADE OF CHOLECYSTOKININ IN EXOCRINE PANCREAS 319
c c
0 0 \
m N c 0 L 0 n E 3 L
20
0
3 days L364,718
[3 7 days L364,718
0 15 days L364,718
FIG. 5. Number of zymogen granules per cell in control rats (n = 30) and in rats treated with L 364,7 18 (0.1 mgikglday) for 3 days (n = 25), 7 days (n = 26), and 15 days (n = 20). n = number of cells counted. Values are means 2 SEM. ANOVA followed by Schefft! test revealed statistically significant differences: ( + + +) p < 0.001 com- pared with controls and (999) p < 0.001 compared with 3-day L 364.71 8-treated rats.
the effects of endogenous CCK on the exocrine pancreas, a reduction of enzyme secretion being observed 26 h after administration (13). The use of 10% DMSO as a vehicle prolongs the action of L 364,718 since DMSO protects the free terminals, avoiding the oxidation pro- cesses that could alter its effectiveness (27). This dose can also prevent the edematous acute pancreatitis in- duced by supramaximal doses of cerulein (28). More- over, the previously described competitive nature of L 364,718 against CCK for binding with the CCK recep- tors in the membrane of the acini (29) was also observed after 26 h, since the submaximal dose of CCK used in this study (1.25 kg/kg/h) is able to displace this antago- nist (15). In line with these findings, Louie et al. (30) have shown that no significant differences in the reduc- tion of amylase secretion were observed when doses of 0.1 and 1 mg/kg/day of L 364,718 were administered.
The results obtained in this study regarding changes in pancreatic weight, DNA content, and enzyme secretion indicate that L 364,718 has a biphasic effect on the pan- creas throughout a 15-day treatment. The CCK receptor
antagonist, administered continuously for 7 days, blocks the native CCK bioactivity, but at longer periods (15 days) a recovery in pancreatic function was observed.
At present, the stimulatory effect of exogenous CCK administration on pancreatic growth is well established (1,2). However there is no direct evidence that, at normal levels, endogenous CCK is the only trophic factor for the maintenance of pancreatic growth. Our results show that while native CCK is blocked (3 and 7 days of treatment), L 364,718 fails to inhibit normal pancreatic growth in accordance with previous studies (1 1,12), a notion com- patible with recent reports demonstrating that pancreatic growth is not fully dependent on CCK-A receptors (31). Therefore trophic factors other than CCK can maintain the growth of the gland when normal levels of plasma CCK are blocked. In fact, several factors, such as secre- tin, gastrin, insulin, neurotensin, prostaglandins, and bombesin, have been shown to stimulate pancreatic growth (3,32-34). During 7 days of L 364,718 adminis- tration a progressive inhibition of endogenous CCK was clearly observed in the exocrine pancreatic function. De- creased basal secretion throughout this period reflects a low paragranular enzymatic content, the cellular com- partment mainly involved in nonstimulated secretion (35). In addition, the enzymatic content of individual zymogen granules was also significantly reduced. How- ever, no significant differences in CCK-stimulated amy- lase secretion between control and L 364,7 18-treated rats were found. These findings suggest that continuous ad- ministration of L 364,718 over 7 days blocks the exocy- tosis, leading to a significant increase in both the number of zymogen granules per cell and their SSC properties. Under these conditions, when CCK is infused at a dose capable of displacing the CCK receptor antagonist, a massive release of zymogen granules has to occur to maintain the secretion at control values, despite the fact
TABLE 1. Body weight gain, pancreatic weight, pancreatichody weight ratio, and DNA content in rat pancreas after long-term administration of L 364,718
Body wt gain (g)
Pancreatic wt (g) Pancreatic wt (g/IOO g body wt) DNA (mg)
Controls (n = 6)
L 364,718
3 days 7 days (n = 6) (n = 6)
15 days (n = 6)
23 f 3 (+3 days) 43 & 4 (+7 days) 80 f 4 (+15 days)
1.308 ? 0.02 0.375 f 0.02 5.879 ? 0.53
22 f 3 40 f 3 75 f 4
1.232e0.11 1.128 f 0.02d 1.466 f 0.06 0.358 k 0.01 0.337 & O.Old 0.416 f 0.01 5.282 & 0.39 5.633 f 0.31 8.619 f 0.67”’
Values are means k SEM. n = number of animals. wt, weight. “ Significant difference with respect to controls 0, < 0.01). ’ Significant difference with respect to 3-day L 364,718-treated rats (p < 0.01). ‘ Significant difference with respect to 7-day L 364,718-treated rats (p < 0.01). “Significant difference with respect to 15-day L 364,718-treated rats (p < 0.01).
Pancreas, Vol. 15. No. 3, 1997

320 A. I . RODRIGUEZ ET AL.
4 0 1 T 200 1 T
~~~ ~
CONTROLS
3 days L364,718
7 days L364,718
[7 15 days L364,718
1
FIG. 6. Basal pancreatic secretion (total protein, amylase, and trypsin output) per pancreas and per DNA in control rats (n = 6) and in rats treated with L 364,718 (0.1 mg/kg/day) for 3 days (n = 6), 7 days (n = 6), and 15 days (n = 6). n = number of animals. Values are means * SEM. ANOVA followed by Scheffk test revealed statistically significant differences: ( + ) p < 0.05, ( + + ) p < 0.01, and ( + +) p < 0.001 compared with controls; (++) p < 0.01 compared with 3-day L 364,718-treated rats; and (0) p < 0.05 and ( 0 0 0 ) p < 0.001 compared with 15-day L 364,718-treated rats.
400 A E .- E 300
3 200 n a c 100
2 L O
9 c,
0
0) .- c,
T ?l
"1 T
3000 n C
\ 'S
2
%
E Q o
9 2000
a 1000
c,
r
m - x
400 1 a z * 300 \
2 40 0 \ r 30 s n - 200
20
*.'
2 3
C $
f 2 10 2 0. a
r
d 100 .-
0 0
6o 1 t - 1 'S
3 days L364,718
(3 7 days L364,718
0 15daysL364,718
10
2 8 0 \
a
S 6 a 4
n c,
C
'% 2 P !-
0 FIG. 7. Total protein. amylase, and trypsin output per pancreas and per DNA under CCK-8 infusion (1.25 pg/kg/h) in control rats (n = 6) and rats treated with L 364,718 (0.1 mgkglday) for 3 days (n = 6), 7 days (n = 6), and 15 days (n = 6). n = number of animals. Values are means 5 SEM. ANOVA followed by Scheffk test showed statistically significant differences: (+) p < 0.05 compared with controls; ( 0 ) p < 0.05 and (00) p < 0.01 compared with 15-day L 364,718-treated rats.
Pu'uncreus, Vol. 15. No. 3, 1997

BLOCKADE OF CHOLECYSTOKININ IN EXOCRINE PANCREAS 321
that the mean enzyme content of each zymogen granule is actually reduced.
After 15 days of treatment, L 364,718 lost its ability to inhibit endogenous CCK. At this time both an increase in the overall DNA content of the pancreas and a recovery of basal and CCK-stimulated secretion were observed, which again suggests that enzyme synthesis is being ex- erted by endogenous CCK. However, the packaging of the enzymes in the individual zymogen granules remains altered. The increase in the number of zymogen granules observed in the pancreata of these rats might not be caused by a blockade of exocytosis but, rather, by the formation of granules, with a significantly smaller size (supported by the decreased FSC observed under these conditions), which also indicates that they are newly formed (36). Interestingly, our results suggest that the rate of formation of zymogen granules would exceed that of enzymatic synthesis since the concentration of amy- lase as reflected by the amylase content/FSC ratio is significantly lower than in control rats. This would sup- port the hypothesis that pancreatic granules are not equally filled and their enzyme concentration is physi- ologically regulated rather than determined by physical limits such as the space available for protein packing, which is in line with the observations of Goncz et al. (37). Hyperplasia was observed after 15 days of L 364,7 I 8 administration. A trophic effect has also been reported in response to chronic administration of prog- lumide (38) and low doses of lorglumide (39). Taking into account the significant decrease in trypsin activity in pancreatic juice of L 364,7 IS-treated rats for 7 days, our results could be explained by an increase in plasma CCK levels as a consequence of the prolonged privation of luminal proteases induced by the continuous secretion blockade during treatment. In fact, bile-pancreatic juice diversion has been established as the most potent stimu- lant for pancreatic exocrine secretion and CCK release in conscious rats (40). This notion is supported by previous works in which an elevation of plasma CCK levels after lorglumide infusion in rats (41) and after L 364,718 ad- ministration in dogs (42) and humans (7) has been re- ported. The finding that after 15 days the DNA pancre- atic content of L 364,7 18-treated rats increased signifi- cantly, with the absence of a parallel increase in pancreatic secretion, could be explained by the fact that plasma CCK would show only a slight increase, since the concentration of circulating CCK required for the induc- tion of hyperplasia is lower than that required for the induction of hypertrophy (43). Therefore under these ex- perimental conditions, the increase in the number of cells would be the main factor responsible for the recovery of pancreatic secretion.
Acknowledgment: This work was supported by a grant from DGICYT, Spain (PB-930638).
REFERENCES
1. Mainz DL, Black 0, Webster PO. Hormonal control of pancreatic growth. J Clin invest 1973;52:2300-4.
2. Solomon TE, Vanier M, Morisset J. Cell site and time course of DNA synthesis in pancreas after caerulein and secretin. Am J Phys- iol 1983;245:G99-105.
3. Morisset J. Stimulation of pancreatic growth by secretin and cae- rulein in suckling rats. Biomed Res 1980; 1:405-9.
4. Douglas BR, Woutersen R, Jamsen JB, Verspaget HW, Lamers CB. Comparison of the trophic effects of FOY-305 on the pancreas in hamsters and rats. Digestion 1987;38: 15-6.
5. Gasslander T, Axelson J , Hakanson I, Ihse I, Lilja I, Rehfeld JF. Cholecystokinin is responsible for growth of the pancreas after pancreaticobiliary diversion in rats. Scand J Gastroenterol 1990; 25: 1060-5.
6. Lotti VJ, Cevino DJ, Kling PJ, Chang RS. A new simple mouse model for the in vivo evaluation of cholecystokinin (CCK) antago- nists: comparative potencies and durations of action of nonpeptide antagonists. Lije Sci 1986;39:1631-8.
7. Cantor P, Mortensen PE, Myhre J, et al. The effect of the chole- cystokinin receptor antagonist MK-329 on meal-stimulated pan- creaticobiliary output in humans. Gastroenterology 1992; 102: 1742-5 1.
8. Hosotani R, Chowdhury P, McKay D, Rayford PL. Effect of L- 364,718, a new CCK receptor antagonist on pancreatic secretion in canscions dogs. Gastroenterology 1987;92: 1441.
9. O’Rourke M, Reidelberger D, Solomon TE. Effect of CCK an- tagonist L-364,718 on meal-induced pancreatic secretion in rats. Am J Physiol 1990;258:G179-84.
0. Wisner JR, McLaughlin RE, Rich KA, Ozawa S, Renner IG. Ef- fects of L-364,718, a new cholecystokinin receptor antagonist, on camostate-induced growth of the rat pancreas. Gastroenterology
1. Zucker KA, Adrian TE, Bilchik AJ, Modlin IM. Effects of the CCK receptor antagonist L-364,718 on pancreatic growth in adult and developing animals. Am J Physiol 1989;257:G51 1-6.
2. Wisner JR, Ozawa S, Xue BG, Renner IG. Chronic administration of a potent cholecystokinin receptor antagonist, L-364.7 18, fails to inhibit pancreatic growth in preweanling rats. Pancreas 19903: 434-8.
13. de Dios I , Manso MA. Effect of L-364,718 (CCK receptor an- tagonist) on exocrine pancreatic secretion of hydrocortisone- treated rats. Pancreas 1994;9:212-8.
14. Manso MA, De Dios I. Effect of L-364,718 (CCK receptor an- tagonist) on exocrine pancreatic secretion of adrenalectomized rats. Pancreas 1993;8:362-7.
15. de Dios I, Orfao A, Garcia AC, Rodriguez AI, Manso MA. Char- acterization of zymogen granules from rat pancreas using flow cytometry. Anal Cell Pathol 1995;9:215-28.
16. Garcia-Montero AC, De Dios I, Rodriguez AI, Orfao A, Manso MA. Adrenalectomy induces a decrease in the light scatter prop- erties and amylase content of isolated zymogen granules from rat pancreas as analyzed by flow cytometry. J Endocrinol 1995;147: 43 1-40.
17. de Dios I, Rodriguez A, Garcia-Montero A, Orfao A, Manso MA. Enzyme changes in zymogen granules and in pancreatic secretion throughout long-term CCK treatment. Peptides (in press).
18. De Lisle RC, Schulz I, Tyrakowski T, Haase W, Hopfer U. Isola- tion of stable pancreatic zymogen granules. Am J Physiol 1984; 246:G411-8.
19. Cabana C, Magny P, Nadeau D, Grondin G, Beaudoin A. Freeze- fracture study of the zymogen granule membrane of pancreas: two novel types of intramembrane particles. Eur J Cell Biol 1987;45: 246-55.
1988;94: 109-13.
Pancrea.F, Vol. 15, No. 3. 1997

322 A. 1. RODRIGUEZ ET AL.
20. Kachel V, Fellner-Feldegg H, Menke E. In: Melamed MR, Lindmo T, Mendelsohn ML, eds. Flow cytometry and sorring. New York: J . Wiley & Liss, 1990:27-45.
21. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye-binding. Anal Biochem 1976;72:248-54.
22. Noelting G, Bernfeld P. Sur les enzymes amylolitiques: I’alfa amy- lase: dosage d’activit6 et controle de l’absence de beta-amylase. Helv Chim Acta 1948;31:286-90.
23. Burton K. A study of the conditions and mechanism of the diphe- nylamine reaction for the colorimetric estimation of deoxyribonu- cleic acid. Biochemistry 1956;62:3 15-26.
24. Salzman GC, Singham SB, Jonston RG, Bohren CF. In: Melamed MR, Lindmo T, Mendelsohn ML, eds. Flow cytometry and Jorting. New York: J. Wiley & Liss, 199O:Xl-107.
25. Muirhead KA, Horan PK, Poste G. Flow cytometry: present and future. Biotechnology 1985;3:337-58.
26. Chang RSL, Lotti VJ. Biochemical and pharmacological charac- terization of an extremely potent and selective non-peptide chole- cystokinin antagonist. Proc Nut1 Acud Sci USA 1986;83:4923-6.
27. Makoto I, Guth PH. Role of oxygen derived free radicals in hem- orrhagic shock induced gastric lesions in the rat. Gastroenterology 1985;88:1162-7.
28. Garcia-Montero AC, Manso MA, Rodriguez AI, de Dios I. Thera- peutic and protective effect of subcutaneous injections of L- 364,718 on caerulein-induced acute pancreatitis. Pancreas 1994; 9309-15.
29. Hosotani R, Chowdhury P, McKay D, Rayford PL. Effect of L- 364,718, a new CCK antagonist on amylase secretion in isolated rat pancreatic acini. Pancreas 1988;3:95-8.
30. Louie DS, Liang JP, Owyang Ch. Characterization of a new CCK antagonist, L-364,718: in vitro and in vivo studies. Am J Physiol 1988;255:G261-6.
31. Miyasaka K, Otha M, Kanai S, Sato Y, Masuda M, Fukanoshi A. Role of CCK-A receptor for pancreatic growth after weaning. A study in a new rat model without gene expression. Pancreas 1996; 12:35 1-6.
32. Dembiski A, Konturek SJ. Effects of E, F and I series prostaglan- dins and analogues on growth of gastroduodenal mucosa and pan- creas. Am J Physiol 1985;248:G170-5.
33. Feurle GE, Muller 8 , Ohnheiser BI. Action of neurotensin on 5ize, composition and growth of pancreas and stomach in the rat. Regul Pept 1985;13:53-62.
34. Mossner J, Logsdon CD, Williams JA, Goldfine ID. Insulin, via its own receptor, regulates growth and amylase synthesis in pancreatic acinar AR 42J cells. Diabetes 1985;34:891-7.
35. Rothman SS, Liebow C, Grendell J. Nonparallel transport and mechanisms of secretion. Biochinz Biciphys Actu 1991;107 1: 159- 73.
36. Cope GH. Exocrine glands and protein secretion a stereological view point. J Microsc 1982;131:187-202.
37. Goncz KK, Behrsing R, Rothman SS. The protein content and morphogenesis of zymogen granules. Cell Tissue Res 199S;280: 5 19-30.
38. Yamaguchi T, Tabatata K, Johnson LR. Effect of proglumide on rat pancreatic growth. Ain J Physinl 1985;249:G294-8.
39. Glasbrenner B, Malfertheiner P, Biichler M, Brandle F, Ditschuneit H. Dose dependent effects of chronic CR 1409 administration on the rat exocrine pancreas. Digestion 1988;40:82.
40. Folsch UR, Cantor P, Wilms HM, Schafmayer A, Becker HD, Creutzfeldt W. Role of cholecystokinin in the negative feedback control of pancreatic enzyme secretion in conscious rats. Castro- enterology 1987;92:449-58.
41, Jansen JB, De Jong AJ, Lamers CB. The cholecystokinin receptor antagonist CR 1409 increases plasma cholecystokinin in rats. Regul Pept 1989;24:209-13.
42. Hosotani R, Chowdhury D, McKay D, Rayford PL. Mechanism by which L-364,7 18 regulates biological actions of CCK on pancreas. Gastroenterology 1988;94: 193.
43. Nakano S, Tachibana I, Otsuki M. Effect of cholecystokinin re- ceptor antagonist loxiglumide on rat exocrine pancreas. Pancreas 1994;9:425-33.
Puncruu.). Vol IS, No. 3, 1997




















