Localizing the movement proteins of Abutilon mosaic geminivirus in yeast by subcellular...
15
Arch Virol (2002) 147: 103–117 Localizing the movement proteins of Abutilon mosaic geminivirus in yeast by subcellular fractionation and freeze-fracture immuno-labelling H.-J. Aberle 1,∗ , M.-L. Rütz 2,∗ , M. Karayavuz 1 , S. Frischmuth 1 , C. Wege 1 , D. Hülser 2 , and H. Jeske 1 1 Department of Molecular Biology and Plant Virology, University of Stuttgart, Stuttgart, Germany 2 Department of Biophysics, University of Stuttgart, Stuttgart, Germany Accepted August 1, 2001 Summary. The movement proteins BC1 and BV1 of Abutilon mosaic gemi- nivirus fused to glutathion-S-transferase (GST) and Flag-peptide were expressed in fission yeast (Schizosaccharomyces pombe) cells to analyse the fundamental intracellular distribution of these proteins in an eukaryotic cell in the absence of plant-specific factors. Most of BC1 protein sedimented rapidly after cell lysis and differential centrifugation. Using freeze-fracture immuno-labelling, the protein was detected in situ predominantly at plasma membranes and to a lower extent at cytoplasmic vesicles but not in the cytoplasm, the nuclei, or the mitochon- dria. Anti-BC1, anti-GST, and anti-Flag antibodies tagged smooth flecks only at the protoplasmic faces of the plasma membrane. The consequences of the BC1 behaviour for its use in two-hybrid analysis in yeast are discussed. In contrast, BV1 was detected mainly in the nucleus and partially in the cytoplasm but never associated with membranes. Introduction Plant viral movement proteins have attracted attention not only because they transport viral RNA or DNA from cell to cell [4] but also because they provide models for the spread mechanism of host mRNA as a key step of genetic cross talk between adjacent cells [25, 26]. Both aspects have been reviewed recently [3, 5, 6, 27]. Two modes of viral transport have been discriminated. One group ∗ H.-J. A. (expression) and M-L. R. (electron microscopy) contributed equally to this work.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Localizing the movement proteins of Abutilon mosaic geminivirus in yeast by subcellular...

Arch Virol (2002) 147: 103–117
Localizing the movement proteins of Abutilon mosaicgeminivirus in yeast by subcellular fractionation
and freeze-fracture immuno-labelling
H.-J. Aberle1,∗, M.-L. Rütz 2,∗, M. Karayavuz1, S. Frischmuth1,C. Wege1, D. Hülser2, and H. Jeske1
1Department of Molecular Biology and Plant Virology,University of Stuttgart, Stuttgart, Germany
2Department of Biophysics, University of Stuttgart, Stuttgart, Germany
Accepted August 1, 2001
Summary. The movement proteins BC1 and BV1 of Abutilon mosaic gemi-nivirus fused to glutathion-S-transferase (GST) and Flag-peptide were expressedin fission yeast (Schizosaccharomyces pombe) cells to analyse the fundamentalintracellular distribution of these proteins in an eukaryotic cell in the absence ofplant-specific factors. Most of BC1 protein sedimented rapidly after cell lysis anddifferential centrifugation. Using freeze-fracture immuno-labelling, the proteinwas detected in situ predominantly at plasma membranes and to a lower extentat cytoplasmic vesicles but not in the cytoplasm, the nuclei, or the mitochon-dria. Anti-BC1, anti-GST, and anti-Flag antibodies tagged smooth flecks only atthe protoplasmic faces of the plasma membrane. The consequences of the BC1behaviour for its use in two-hybrid analysis in yeast are discussed. In contrast,BV1 was detected mainly in the nucleus and partially in the cytoplasm but neverassociated with membranes.
Introduction
Plant viral movement proteins have attracted attention not only because theytransport viral RNA or DNA from cell to cell [4] but also because they providemodels for the spread mechanism of host mRNA as a key step of genetic crosstalk between adjacent cells [25, 26]. Both aspects have been reviewed recently[3, 5, 6, 27]. Two modes of viral transport have been discriminated. One group
∗H.-J. A. (expression) and M-L. R. (electron microscopy) contributed equally to thiswork.

104 H.-J. Aberle et al.
of viruses, typified by Tobacco mosaic virus, encodes a movement protein whichbinds to plasmodesmata, changes their size exclusion limit, and binds to RNAin order to thread the RNA-protein complex through the symplastic bridges be-tween adjacent plant cells [2, 13]. The second group, typified by Cowpea mosaicvirus, produces tubular extensions of the plasma membrane for the virions’ traf-fic [22, 38, 40, 48]. In the case of DNA-containing geminiviruses the detailsof cell-to-cell transport are less well understood. As geminiviruses multiply inthe nuclei, viral DNA has to cross two distinct barriers, the nuclear envelopeand the plasmodesmata [30, 34–36, 44, 45]. The genome of geminiviruses con-sists of circular single-stranded DNA and is either monopartite (genus Mastre,Curtovirus) or bipartite (most members of the genus Begomovirus) [32]. Forbegomoviruses two movement protein genes (bc1 and bv1) have been identifiedon the DNA B component [7, 9, 42]. BC1 is thought to be responsible for thetransfer of viral DNA across the cell wall whereas BV1 is involved in the ex-port of viral DNA from the nucleus [28, 36]. How the proteins interact is still amatter of debate [5] and several techniques have been developed during the lastdecade to study such interactions in yeast cells [41]. Besides the role of BC1in transport it is well-known as a major determinant of symptom formation butthe underlying mechanism is unknown [7, 18, 29, 37, 43]. The divergence inthe sequences of BC1 proteins from distinct viruses might reflect adaptations todifferent hosts. Indeed, first evidence hints at a function of BC1 in host range de-termination [16, 18]. It might be possible that interactions between viral and hostproteins at the plasmodesmata govern the selectivity of the transport. BC1 hasbeen detected close to the plasmamembrane of host cells [35] and in the so-called“cell wall-enriched fraction” during differential centrifugation [42]. Using elec-tron microscopic immuno-labelling on ultrathin sectionsWard et al. [45] detectedBC1 (called BL1) of Squash leaf curl virus (SLCV) in endoplasmic reticulum-derived tubules, but exclusively in special cells, namely immature phloem cells.In fusions with green fluorescent protein (GFP) BC1 of AbMV targets the cellperiphery or cap-like structures around the nucleus in onion (nonhost) and tobacco(host) cells [50]. Fluorescence appeared most frequently as small punctateflecks close to the cell wall in source cells of host and nonhost plants, but asdisc-like structures in sink cells of host plants. To examine which aspect of theintracellular routing of BC1 is contributed by a general interaction with mem-branes or by plant-specific factors we chose fission yeast (Schizosaccharomyespombe) for heterologous expression.Moreover, yeasts are very common in study-ing protein-protein interactions using two-hybrid systems [41]. For the classicalversion of this approach it is necessary to know whether the protein of interestenters the nucleus. Therefore it is of crucial importance to test whether gem-iniviral movement proteins target the cell periphery in yeast as in plants. Theresults of freeze-fracture immuno-labelling show that most of the expressed BC1is also targeted to the plasma membrane in fission yeast but was exclusivelylocalized to its protoplasmic face. Therefore, this binding is very general, butnevertheless host specific factors might be responsible for the transport from cellto cell.

Movement proteins of Abutilon mosaic geminivirus 105
Materials and methods
Virus
Abutilon mosaic virus (AbMV) has been described [19, 47] and its DNA sequenced [9];minor errors in the sequence of BC1 have been corrected recently, EMBL accession numbersX15983; X15984). AbMV BC1 and BV1 have been identified as movement proteins [7].
Antibodies
Primary polyclonal antisera against AbMV BC1 or BV1 expressed in E. coli were producedin mice, and their selectivity was shown [46]. The following antibodies were obtained fromcommercial suppliers: Goat polyclonal antiserum against glutathion-S-transferase (GST)(Amersham Pharmacia Biotech, #27-4577-01), mouse antibodies against Flag (Stratagene,#200471), secondary 15 nmgold-labelled goat-anti-mouse IgG (H+L) antibodies (BioTrend,Köln, Germany, #115.022) and 15 nm gold-labelled rabbit-anti-goat IgG (H+L) (Bio Trend,#815.077).
Cloning and expression
Standard molecular techniques were performed according to Sambrook et al. [33]. Thebc1 and the bv1 genes were amplified by PCR from pBIN19-AbMV.DNA B [9] usingPfuTurbo polymerase (Stratagene) and the following primer pairs: 5′-TACGGATCCATGGATTCTCAGTTAGTA-3′ and 5′-TACTAGGGCCCTTATTTCAATGATTTGGC-3′ for bc1;5′-TACAGATCTATGTACCCGTCTAGGAATAAA-3′ and 5′-GATCTGCCCGGGTTAACCAATATAGTCAAGGTC-3′ for bv1. The products were inserted into the expression plas-mid pESP-1 to obtain N-terminal fusions with GST and the Flag peptide (DYKDDDDK,Immunex Corporation). The constructs were verified by sequencing [LiCor 4000L, MWG].The recombinant plasmids pESP1:BC1, pESP1:BV1 as well as a control plasmid pESP1:KI(GST fusion with a 12 kDa control protein; provided with the Stratagene kit) were trans-formed into competent yeast cells (Schizosaccharomyces pombe). Cells were selected onEMM-thiamine (Edinburgh Minimal Medium including 5�M thiamine) plates at 30 ◦C for4–6 days. Frozen stocks for further use were prepared by streaking colonies directly into20% aqueous glycerol and storing at −80 ◦C.
For expression, cells were inoculated into 25ml ofYES (yeast extract plus supplements)medium, cultured overnight at 30 ◦C, transferred to new YES medium for 5 h and inducedby exchanging the medium to minimal medium (EMM without the repressor thiamine). Af-ter 18–20 h of induction, cells were harvested by centrifugation (1000 × g, 5min, roomtemperature), washed with PBS (140mM NaCl, 2.7mM KCl, 10mM Na2HPO4, 1.8mMKH2PO4), resuspended in PBST-PI (including 1% Triton X-100 and protease inhibitors:1mM PMSF, 1mg/ml aprotinin, 1�MpepstatinA, 100�M leupeptin, 1�g/ml chymostatin)and disrupted by glass beads [1]. Aliquots were centrifuged (Eppendorf, rotor F45-30-11,12.000 × g, 5min, 4 ◦C), pellets (P) and supernatants (S) were collected separately andsubjected to sodium dodecyl sulfate (SDS) polyacrylamide gel electrophoresis (PAGE) ac-cording to [24]. Gels were stained with Serva violet, marker proteins were from PharmaciaBiotech (LMW electrophoresis-calibration kit).
Freeze-fracture immuno-labelling (FreeFI)
The principle of FreeFI is shown in Fig. 1 and was developed by Gruijters et al. [15] andFujimoto [11]. Yeast cells harbouring the control plasmid (KI), pESP1:BC1 or pESP1:BV1were grown as described above and sedimented (1000× g, 5min, room temperature, HeraeusVarifuge). Cells were washed once with 10ml of cold PBS and sedimented as described

106 H.-J. Aberle et al.
Fig. 1. Schematic view of freeze-fracture immuno-labelling. Nomenclature according toRoberts et al. [31]: external surface (ES), external face (EF), protoplasmic face (PF) andprotoplasmic surface (PS). During freeze-fracture, membranes are preferentially cleavedalong the hydrophobic inner plane. Some proteins stick to one half of the membrane leavingpits in the other (a/a′; b/b′), others are broken (d/d′) or remain attached only to one half of themembrane (c). After platinum/carbon (Pt/C) evaporation the cellular material is removed bySDS treatment. Only proteins in close contact to the Pt/C layer remain at the fractured leafletand can be detected by a specific primary antibody and a gold-labelled secondary antibody.Note that the shapes of PF and EF are visualized by Pt shading whereas labelling of proteins
occurs at PS and ES
above, placed between two gold specimen carriers and immediately frozen in liquid propane.Samples were stored in liquid nitrogen before fracturing. Freeze-fracturing was performedand the replica were processed as described [11, 17]. Briefly, cells were fractured under highvacuum, shadowed with platinum (45 ◦) and stabilized by evaporating carbon (90 ◦). Replicawere floated on PBS and left overnight in a solution of 2.5% SDS in 10mM Tris-HCl,

Movement proteins of Abutilon mosaic geminivirus 107
pH 8.3, and 30mM sucrose to remove most of the cell debris. After washing ten times inPBS, non-specific reactions were blocked with 2% bovine serum albumin (BSA) in PBS.Replica were placed on droplets of primary antibodies (diluted 1:1000 for mouse-anti-BC1,1:100 for goat-anti-GST, 1:100 for mouse-anti-Flag) for 60min at room temperature, washedten times in PBS, blocked in 2% BSA, incubated on a droplet of secondary antibody (diluted1:30) for 60min at room temperature, washed ten times in PBS, twice in distilled H2O beforebeing fished on copper grids.
Electron microscopy
Specimens were examined in a Zeiss electron microscope [EM 10A, Zeiss, Oberkochen,Germany] at 60 kV. At magnifications from 10,000 to 40,000, photographs were taken onKodak film (Kodak Electron Microscope Film, ESTAR Thick Base 4489, Cat #1613108,Eastman Kodak Company, Rochester, NewYork, USA).
Results
Several transformed yeast clones containing either the control plasmid or pESP1:BC1 were analysed by SDS-PAGE (Fig. 2). All clones expressed the foreign pro-teins with the expected mobility as a double band.After low speed centrifugationof the lysatemost of the GST:BC1 fusion protein was pelleted whereas the controlprotein (GST:KI) remained in the supernatant. This behaviour could either resultfrom aggregation of BC1, as it has been shown for bacterially expressedBC1 [46],or from an attachment ofBC1 to cell walls or other rapidly sedimenting complexesof the yeast. To discriminate between these possibilities yeast cells expressing thecontrol protein or GST:BC1 fusion proteins were analysed by FreeFI. Figure 3shows an overview of two single GST:BC1-expressing cells. Substantial antibodylabel of BC1 is found on the plasma membranes (pm). In fission yeast, as in most
Fig. 2. Expression of GST:BC1 fusion protein (arrow heads) in fission yeast and E. coli.a.Molecular weight marker (sizes in kDa indicated at the left side). b. GST:BC1 expressedin E. coli for comparison. Induced (c, d) and repressed (e, f) protein expression patterns inthe pellet fractions (c, e) or in the supernatants (d, f) after low speed centrifugation. Notethe double band which is only present in the induced-pellet fraction in c. The lower band
comigrates with the bacterially expressed protein. Stain: Serva violet

108 H.-J. Aberle et al.
Fig. 3. Freeze-fractured fission yeast cells expressing GST:BC1. The replica were incubatedwith anti-BC1 antibody as described in Materials and methods. Gold-labelled antibodies ac-cumulated mainly on plasma membranes (pm) and in some cases on endomembrane vesicles(v) but not in the nucleus (n), the mitochondria (m) or the cytoplasm. Note that the protoplas-mic faces (PF) of the plasma membranes are exhibited with the characteristic invaginationscaused by protruding cell wall. Helpful for interpretation is to imagine that the antibodiesbind to the antigen from below the picture plane (protoplasmic surface, PS). Bar: 500 nm;
arrowheads indicate direction of platinum evaporation
other organisms, the protoplasmic face (PF; compare Fig. 1) contains a higherdensity of particles than the external face (EF). Moreover, the surface sculpture ofthe plasma membrane allows a reliable identification of the two faces. In yeasts,cell walls typically protrude into the protoplasts [12] leaving invaginations in PFand protrusions in EF after internal cleavage of the plasma membrane.
To interpret FreeFI images it is important to note that the view is differentfrom that in ultrathin immunohistochemistry. As freeze-fracture proceeds alongthe center ofmembranes, between the two lipid layers, the topographies of external
Movement proteins of Abutilon mosaic geminivirus 109
(EF) and protoplasmic faces (PF) are represented by platinum evaporation [31]whereas antibodies recognize proteins only at the corresponding surfaces (ES andPS; see Fig. 1).
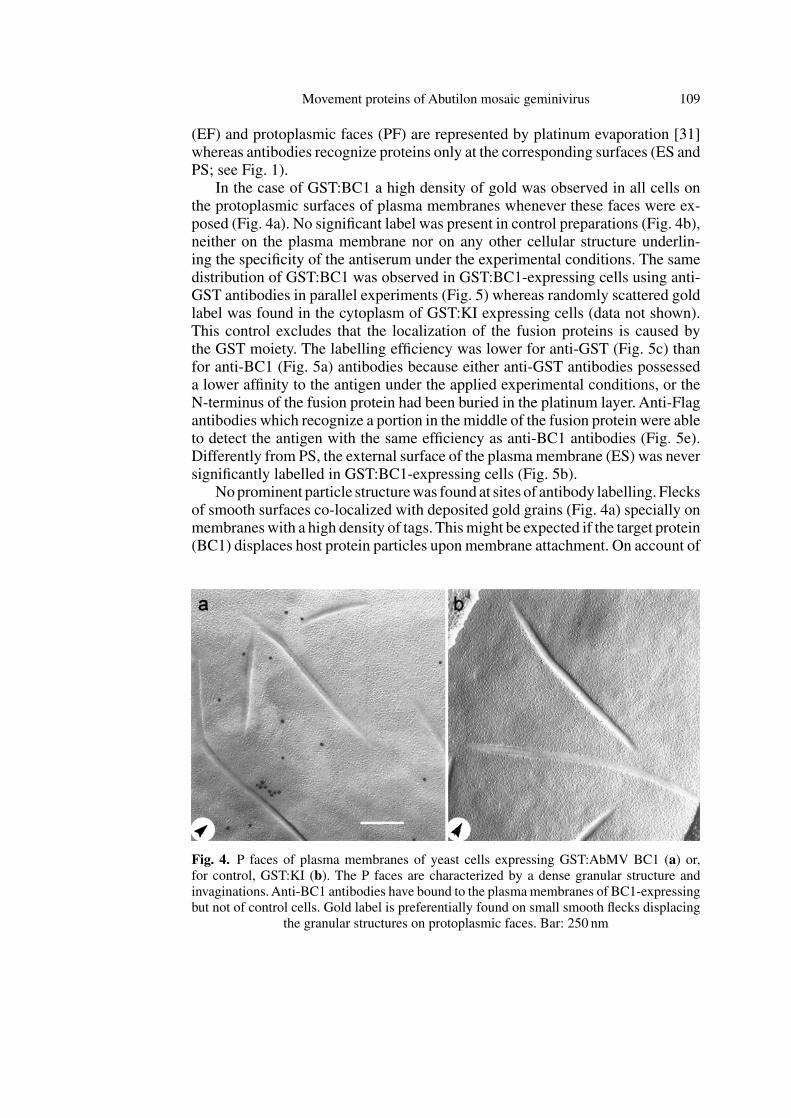
In the case of GST:BC1 a high density of gold was observed in all cells onthe protoplasmic surfaces of plasma membranes whenever these faces were ex-posed (Fig. 4a). No significant label was present in control preparations (Fig. 4b),neither on the plasma membrane nor on any other cellular structure underlin-ing the specificity of the antiserum under the experimental conditions. The samedistribution of GST:BC1 was observed in GST:BC1-expressing cells using anti-GST antibodies in parallel experiments (Fig. 5) whereas randomly scattered goldlabel was found in the cytoplasm of GST:KI expressing cells (data not shown).This control excludes that the localization of the fusion proteins is caused bythe GST moiety. The labelling efficiency was lower for anti-GST (Fig. 5c) thanfor anti-BC1 (Fig. 5a) antibodies because either anti-GST antibodies possesseda lower affinity to the antigen under the applied experimental conditions, or theN-terminus of the fusion protein had been buried in the platinum layer. Anti-Flagantibodies which recognize a portion in the middle of the fusion protein were ableto detect the antigen with the same efficiency as anti-BC1 antibodies (Fig. 5e).Differently from PS, the external surface of the plasmamembrane (ES) was neversignificantly labelled in GST:BC1-expressing cells (Fig. 5b).
Noprominent particle structurewas found at sites of antibody labelling. Flecksof smooth surfaces co-localized with deposited gold grains (Fig. 4a) specially onmembraneswith a high density of tags. Thismight be expected if the target protein(BC1) displaces host protein particles upon membrane attachment. On account of
Fig. 4. P faces of plasma membranes of yeast cells expressing GST:AbMV BC1 (a) or,for control, GST:KI (b). The P faces are characterized by a dense granular structure andinvaginations.Anti-BC1 antibodies have bound to the plasmamembranes of BC1-expressingbut not of control cells. Gold label is preferentially found on small smooth flecks displacing
the granular structures on protoplasmic faces. Bar: 250 nm

110 H.-J. Aberle et al.
Fig. 5. Views of plasma membranes after antibody tagging of yeast cells expressingGST:Flag:AbMV BC1- (a, b, c, e) or, for control, GST:Flag:KI- (d, f). Replica were incu-bated with anti-BC1 (a, b), anti-GST (c, d) or anti-Flag antibodies (e, f). Whereas anti-BC1has tagged the P surface (a) it did not bind to the E surface (b) of plasma membranes. TheE faces are distinguishable from P faces by their less dense granular substructure and pro-trusions. Frequently the granules form typical strings on the E faces, irrespective of whetherBC1 or KI fusions were expressed. Anti-BC1 (a) and anti-Flag (e) antibodies have reactedmore intensely than anti-GST antibodies (c). All three have tagged exclusively the P surfaces
of cells expressing BC1 but not KI fusion protein. Bar: 250 nm

Movement proteins of Abutilon mosaic geminivirus 111
their small size (33 kDa) single BC1moleculeswould be visible only if assembledinto larger complexes.
Besides the predominant label on plasma membranes, microsomal vesicleswere tagged by anti-BC1 antibodies (Fig. 3a; Fig. 6) indicating that BC1 couldbe transported to the plasma membrane via such vesicles. This type of label wascompletely absent in control cells. No other organelles (nuclei, mitochondria, orGolgi apparatus) were found to be significantly labelled in these experiments. Inspite of an extensive search we did not find any inclusion bodies in the cytoplasmas they were readily formed in bacteria [46].
The second movement protein ofAbMV (BV1) was mainly detected in nucleiand to a lower extent in the cytoplasm (Fig. 7). No relevant label was found onmembranes. In nuclei and in the cytoplasm the goldgrains were clustered ratherthan randomly distributed.Rarely, nuclear poreswere labelled (seeFig. 7b, insert).
Fig. 6. BC1 antigen detected on various vesicles (a–d) at their protoplasmic surfaces, in-cluding those adjacent to the plasma membrane (pm in a, arrows). m Mitochondrium. Bar:
1�m

Fig. 7. S. pombe cells expressing GST:Flag:BV1 protein labelled by anti-BV1 antibodies.Gold grains are predominantly deposited on the cross-fracture of the nuclei (n in a, b), to alower extent on the cytoplasm (cy in b) and rarely close to nuclear pores (np in b, insert).The nuclear envelope (ne) and the white box of the enlarged insert are marked in (b). Bars
represent 2�m in a and 1�m in b

H.-J. Aberle et al.: Movement proteins of Abutilon mosaic geminivirus 113
The transport through this structure might be too fast to be detected with theapplied technique.
Discussion
Upon heterologous expression in fission yeast AbMV BC1 is found in the pelletfraction after low speed differential centrifugation. Such behaviour was previ-ously found in E. coli [46]. The reason for rapid sedimentation in E. coli wasthe formation of large inclusion bodies. Such aggregates were extremely stableand only dissolved under high urea concentrations (>6M; [46]; Karayavuz, un-published data). They re-appeared at once if urea was removed indicating thatself-aggregation is an intrinsic property of overexpressedAbMVBC1 in aqueoussolutions or that chaperones might be necessary for correct folding. AbMV BC1remained in the supernatant during subcellular fractionation of plant sap [50] butthis difference might have resulted from the low concentration of AbMV BC1 ininfected plants. Therefore it is interesting thatAbMVBC1 also sediments rapidlywhen expressed under a strong promotor in yeast. In contrast to the expressionin E. coliAbMV BC1 never formed inclusion bodies in the cytoplasm but accu-mulated at the plasma membrane in yeast (Fig. 3). Therefore it is probable thatAbMV BC1 pellets together with the cell wall during differential centrifugationin homogenates from yeasts rather than due to self-aggregation as in E. coli.
The amino acid sequence of BC1 harbours several putative phosphorylationsites. In vivo phosphorylation of BC1 was shown in insect cells [30] and inE. coli [46]. The appearance of a double band in SDS-PAGE (Fig. 2) might reflectsuch a modification also in yeast. Phosphorylation is frequently involved in thecellular site changes of various proteins [10, 14, 20, 21, 23, 49]. Therefore differ-ent loci ofAbMVBC1 accumulationmight be expected in the cell. The intensitiesof the two bands in SDS-PAGE are nearly the same. If each of the bands representsBC1 from a different cellular site two loci of equal immunolabel density will bepostulated. Indeed, fusions of AbMV BC1 and GFP protein were localized at thecell periphery as well as in the vicinity of the nucleus in plant cells [50]. Thelatter occurred, however, only in a minority of cells. No indication of a secondsite of accumulation was found in yeast so far. Therefore it is unlikely that one ofthe two bands seen after expression in yeast (Fig. 2, lane b) results from a secondintracellular location.
The presence of some antibody label on vesicles in yeast cells indicates thatmicrosomal membranes might be involved in the transport of BC1 to the plasmamembrane. The signal density on the vesicles is expected to be lower during therapid transport than on the plasmamembranewhereBC1 accumulates presumablyto steady state levels. These findings correspond to observations ofWard et al. [45]for SLCV which induced tubular structures containing BC1 and an ER marker(binding protein, BIP) in plants.
In comparison to ultrathin sections, the great advantage of freeze-fracture im-ages is the presentation of a three-dimensional impression of the membrane witha considerably higher resolution of membrane-associated complexes. Moreover,

114 H.-J. Aberle et al.
it is sometimes possible to assign a protein complex to a certain face of themembrane. The interpretation is, however, complicated by the fact that duringfreeze-fracture the membranes are split along the inner hydrophobic plane andthat integral proteins are thereby pulled out to one of the membrane leaflets leav-ing a hole in the other (Fig. 1, marked a/a′). In the case of plasmamembranesmostproteins remain anchored in the protoplasmic face (see Fig. 5a) whereas only alimited amount of granular material, presumably those proteins that are fixed tothe cell wall, was seen on the external face (Fig. 1, marked b/b′; Fig. 5b; for dis-cussion of this general context see [31, 39], and of the special structures in yeastssee [12]). Consequently, not only peripheral but also integral membrane proteinsare regularly found on only one side of the plasma membrane. True peripheralproteins, however, are washed away by the technique applied due to the action ofSDS. Only proteins which are anchored to one half of the membrane (partiallyintegrated) (Fig. 1, marked c), and thereby become attached to the platinum orcarbon layer, remain fixed. In consequence, AbMV BC1 could be both, a trans-membrane protein or a protein which is fixed in the protoplasmic leaflet of themembrane. In any case it must possess a cytoplasmic domain detected by BC1-and Flag-specific antibodies. The appearance of smooth flecks on the protoplas-mic face rather than sculptured granules tagged by the antibodies might be anargument in favour of a partial integration of BC1 as indicated in Fig. 1 marked c.The small size of the protein, however, prohibits a firm conclusion. Further bio-chemical experiments have to answer this question unequivocally. In any case,BC1 was never detected free in the cytoplasm or as luminal protein in vesicles asit had been proposed for the ER-derived tubules of SLCV in planta [45].
The results shown above exclude that host-specific proteins are necessaryfor this fundamental distribution of AbMV BC1 within the cell. If such specificfactors are involved, they must be responsible for different processes in plants,e.g. gating of specialized plasmodesmata. Host specificitymight also be conferredindirectly via differential velocities of viral movement in the respective hosts.
In contrast to BC1, BV1 was localized in the nucleoplasm and the cytoplasmbut never attached to membranes. A nuclear localisation of BV1 was also foundin plants by in situ immuno-labelling and GFP-tagged BV1 [50]. This is in accor-dance with findings for other begomoviruses [36]. After translation, monomericBV1 would be small enough to diffuse passively into the nucleus where it couldaccumulate due to its DNA-binding properties. Nevertheless, nuclear localisa-tion as well as nuclear export signals have been defined for SLCV [44] makingBV1 (synonymBR1) a nuclear shuttle protein. Such signal recognition-dependenttransport would be necessary for multimeric BV1, especially if the protein is as-sociated with the viral DNA. On this background the clustering of BV1, whichis shown here for the the first time in the cytoplasm as well as in the nucleus(Fig. 7b), is consistent with multimerization.
The targeting of AbMV BC1 to plasma membranes might have general con-sequences for the use of the yeast two-hybrid system with geminivirus movementproteins. Preliminary experiments (unpublished data) indicated that substantialamounts of BC1 occurred at the Saccharomyces cerevisiae plasma membrane,

Movement proteins of Abutilon mosaic geminivirus 115
even if the proteinwas fused to theDNA-binding domain of theGal4 transcriptionfactor which provides a strong nuclear localization signal. Therefore difficultiesmay arise if protein-protein interactions are to be verified by the classical versionof the two-hybrid system [41]. Correspondingly, we never detected an interactionof BV1 and BC1 in two-hybrid experiments (unpublished data) although such aninteraction was evident in planta after coexpression of GFP-fused BV1 and un-fused BC1 [50]. In plants, BC1 mobilized BV1 not only to the plasma membranebut also to the neighbour cell [50].
Acknowledgements
This work was supported by a grant of the Deutsche Forschungsgemeinschaft (Je116/5-3).We thank PD Dr. T. Frischmuth and Prof. Dr. H. Wolf for helpful discussions andMrs. C. Kocher and Mrs. B. Rehkopf for excellent technical assistance.
References
1. Ausubel FM, Brent R, Ekingston R, Moore DD, Smith JA, Seidman JG, Sruhl K (1990)Current protocols in molecular biology. Green Publishing Associates, John Wiley andSons – Interscience, NewYork
2. Citovsky V (1993) Probing plasmodesmal transport with plant viruses. Plant Physiol102: 1071–1076
3. Crawford KM, Zambryski PC (1999) Phloem transport: Are you chaperoned? Curr Biol9: 281–285
4. Deom CM, Oliver MJ, Beachy RN (1987) The 30- kilodalton gene product of tobaccomosaic virus potentiates virus movement. Science 237: 389–394
5. Ding B (1998) Intercellular trafficking through plasmodesmata. Plant Mol Biol 38:279–310
6. Ding B, Itaya A, Woo Y-M (1999) Plasmodesmata and cell-to-cell communication inplants. Int Rev Cytol 190: 251–316
7. Evans D, JeskeH (1993) DNAB facilitates, but is not essential for, the spread ofAbutilonmosaic virus in agroinoculated Nicotiana benthamiana. Virology 194: 752–757
8. Frischmuth T, Roberts S, von Arnim A, Stanley J (1993) Specificity of bipartite gemi-nivirus movement proteins. Virology 196: 666–673
9. Frischmuth T, Zimmat G, Jeske H (1990) The nucleotide sequence of abutilon mosaicvirus reveals prokaryotic as well as eukaryotic features. Virology 178: 461–468
10. FrumanDA, Rameh LE, Cantley LC (1999) Phosphoinositide binding domains: embrac-ing 3-phosphate. Cell 97: 817–820
11. Fujimoto K (1995) Freeze-fracture replica electron microscopy combined with SDSdigestion for cytochemical labeling of integral membrane proteins. J Cell Sci 108:3443–3449
12. Garrison RG (1981) Vegetative ultrastructure. In: Arnold WN (ed)Yeast cell envelopes:Biochemistry, biophysics, and ultrastructureVol II. CRC Press, Boca Raton, pp 139–160
13. Ghoshory S, Lartey R, Sheng J, Citovsky V (1997) Transport of proteins and nucleicacids through plasmodesmata. Annu Rev Plant Physiol Plant Mol Biol 48: 27–50
14. Gillham H, Golding MC, Pepperkok R, Gullick WJ (1999) Intracellular movement ofgreen fluorescent protein-tagged phosphatidylinositol 3-kinase in response to growthfactor receptor signaling. J Cell Biol 146: 869–880
15. Gruijters WTM, Kistler J, Bullivant S, Goodenough A (1987) Immunolocalization ofMP70 in lens fiber 16–17-nm intercellular junctions. Cell Biol 104: 565–572

116 H.-J. Aberle et al.
16. Hou Y-M, Paplomatas EJ, Gilbertson RL (1998) Host adaptation and replication prop-erties of two bipartite geminiviruses and their pseudorecombinants. Mol Plant MicrobeInteract 11: 208–217
17. Hülser DF, Rehkopf B, Traub O (1997) Dispersed and aggregated gap junction channelsidentified by immunogold labeling of freeze-fractured membranes. Exp Cell Res 233:240–251
18. Ingham DJ, Pascal E, Lazarowitz SG (1995) Both bipartite geminivirus movement pro-teins define viral host range, but only BL1 determines viral pathogenicity. Virology 207:191–204
19. Jeske H, Menzel D, Werz G (1977) Electron microscopic studies on intranuclearvirus-like inclusions in mosaic-diseased Abutilon sellowianum Reg. Phytopathol Z 89:289–295
20. Kann M, Sodeik B, Vlachou A, Gerlich WH, Helenius A (1999) Phosphorylation-dependent binding of hepatitis B virus core particles to the nuclear pore complex.J Cell Biol 145: 45–55
21. Karpova OV, Rodionova NP, Ivanov KI, Kozlovsky SV, Dorokhov YL, Atabekov JG(1999) Phosphorylation of tobacco mosaic virus movement protein abolishes its trans-lation repressing ability. Virology 261: 20–24
22. Kasteel DTJ, van derWelNN, JansenKAJ, GoldbachRW, van Lent JWM(1997)Tubule-forming capacity of the movement proteins of alfalfa mosaic virus and brome mosaicvirus. J Gen Virol 78: 2089–2093
23. Kawakami S, Padgett HS, Hosokawa D, Okada Y, Beachy RN, Watanabe Y (1999)Phosphorylation and/or presence of serine 37 in the movement protein of tomato mosaictobamovirus is essential for intracellular localization and stability in vivo. J Virol 73:6831–6840
24. Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature 227: 680–685
25. Lazarowitz SG, Beachy RN (1999) Viral movement proteins as probes for intracellularand intercellular trafficking in plants. Plant Cell 11: 535–548
26. Lucas WJ, Bouche-Pillon S, Jackson DP, Nguyen L, Baker L, Ding B, Hake S (1995)Selective trafficking of KNOTTED1 homeodomain protein and its mRNA through plas-modesmata. Science 270: 1980–1983
27. Nelson RS, van Bel AJE (1998) The mystery of virus trafficking into, through and outof vascular tissue. Progr Botany 59: 476–533
28. Noueiry AO, Lucas WJ, Gilbertson RL (1994) Two proteins of a plant DNA virus coor-dinate nuclear and plasmodesmatal transport. Cell 76: 925–932
29. Pascal E, Goodlove PE, Wu LC, Lazarowitz SG (1993) Transgenic tobacco plants ex-pressing the geminivirus BL1 protein exhibit symptoms of viral disease. Plant Cell 5:795–807
30. Pascal E, SanderfootAA,Ward BM, Medville R, Turgeon R, Lazarowitz SG (1994) Thegeminivirus BR1 movement protein binds single-stranded DNA and localizes to the cellnucleus. Plant Cell 6: 995–1006
31. Roberts RL, Kessel RG, Tung H-N (1991) Freeze fracture images of cells and tissues.Oxford University Press, NewYork Oxford
32. Rybicki EP, Briddon RW, Brown JK, Fauquet CM, Maxwell DP, Harrison BD,Markham PG, Bisaro DM, Robinson D, Stanley J (2000) Family Geminiviridae.In: Van Regenmortel MHV, Fauquet CM, Bishop DHL et al. (eds) Virus Taxonomy.Classification and Nomenclature of Viruses. Academic Press, San Diego, pp 285–297;
33. Sambrook J, Fritsch EF,Maniatis T (1989)Molecular cloning: a laboratorymanual. ColdSpring Laboratory Harbor Press, NewYork

Movement proteins of Abutilon mosaic geminivirus 117
34. Sanderfoot AA, Ingham DJ, Lazarowitz SG (1996) A viral movement protein as anuclear shuttle. The geminivirus BR1 movement protein contains domains essential forinteraction with BL1 and nuclear localization. Plant Physiol 110: 23–33
35. Sanderfoot AA, Lazarowitz SG (1995) Cooperation in viral movement: the geminivirusBL1 movement protein interacts with BR1 and redirects it from the nucleus to the cellperiphery. Plant Cell 7: 1185–1194
36. Sanderfoot AA, Lazarowitz SG (1996) Getting it together in plant virus movement:cooperative interactions between bipartite geminivirus movement proteins. Trends CellBiol 6: 353–358
37. Saunders K, Wege C, Veluthambi K, Jeske H, Stanley J (2001) The distinct diseasephenotypes of the common and yellow vein strains of Tomato golden mosaic virus aredetermined by nucleotide differences in the 3′-terminal region of the gene encoding themovement protein. J Gen Virol 82: 45–51
38. SijenT,GoldbachRW,vanKammenA (1993)The cowpeamosaic virusMRNA-encoded48-kilodalton protein is responsible for induction of tubular structures in protoplasts.J Virol 67: 3660–3664
39. Stolinski C, BreathnachAS (1975) Freeze-fracture replication of biological tissues.Aca-demic Press London NewYork
40. StormsMMH, Kormelink R, Peters D, Lent JWMv, Goldbach RW (1995) The nonstruc-tural NSm protein of tomato spotted wilt virus induces tubular structures in plant andinsect cells. Virology 214: 485–493
41. Vidal M, Legrain P (1999) Survey and summary. Yeast forward and reverse ‘n’-hybridsystems. Nucleic Acids Res 27: 919–929
42. vonArnimA, Frischmuth T, Stanley J (1993) Detection and possible functions ofAfricancassava mosaic virus DNA B gene products. Virology 192: 264–272
43. von Arnim A, Stanley J (1992) Determinants of tomato golden mosaic virus symptomdevelopment located on DNA B. Virology 186: 286–293
44. WardBM, Lazarowitz SG (1999)Nuclear export in plants: Use of geminivirusmovementproteins for a cell-based export assay. Plant Cell 11: 1267–1276
45. Ward BM, Medville R, Lazarowitz SG, Turgeon R (1997) The geminivirus BL1 move-ment protein is associated with endoplasmic reticulum-derived tubules in developingphloem cells. J Virol 71: 3726–3733
46. Wege C, Jeske H (1998) Abutilon mosaic geminivirus proteins expressed and phospho-rylated in Escherichia coli. J Phytopathol 146: 613–621
47. Wege C, Gotthardt R-D, Jeske H (2000) Fulfilling Koch’s postulates forAbutilon mosaicvirus. Arch Virol 145: 2217–2225
48. Wellink J, van Lent JW, Verver J, Sijen T, Goldbach RW, van Kammen A (1993) Thecowpea mosaic virusMRNA-encoded 48-kilodalton protein is responsible for inductionof tubular structures in protoplasts. J Virol 67: 3660–3664
49. Yeakley JM, Tronchere H, Olesen J, Dyck JA,Wang HY, Fu XD (1999) Phosphorylationregulates in vivo interaction and molecular targeting of serine/arginine-rich pre-mRNAsplicing factors. J Cell Biol 145: 447–455
50. Zhang SC, Wege C, Jeske H (2001) Movement proteins (BC1 and BV1) of Abutilonmosaic virus are cotransported in and between cells of sink but not of source leaves asdetected by green fluorescent protein tagging. Virology 290 (in press)
Author’s address: Dr. H. Jeske, University of Stuttgart, Biological Institute,Pfaffenwaldring 57, D-70550 Stuttgart, Germany; e-mail: [email protected]
Received April 23, 2001