
Arrival Cities. Migrating Artists and New ... - Open Access LMU
Upload
independentCategory
view
1download
0
LR
D
D
BcmobpepwotoecwppplCtsttremcTrcsi
Adrmsastvb
BA
SIC–
ALIM
ENTA
RY
TRA
CT
GASTROENTEROLOGY 2009;136:1328–1338
ocalized Release of Serotonin (5-Hydroxytryptamine) by a Fecal Pelletegulates Migrating Motor Complexes in Murine Colon
ANTE J. HEREDIA, EAMONN J. DICKSON, PETER O. BAYGUINOV, GRANT W. HENNIG, and TERENCE K. SMITH
epartment of Physiology & Cell Biology, University of Nevada School of Medicine, Reno, Nevada
tprdcmt
rvscAffpsnws
tdtbttbiote55cpn
ljt
ackground & Aims: The colonic migrating motoromplex (CMMC) is a motor pattern that regulates theovement of fecal matter through a rhythmic sequence
f electrical activity and/or contractions along the largeowel. CMMCs have largely been studied in emptyreparations; we investigated whether local reflexes gen-rated by a fecal pellet modify the CMMC to initiateropulsive activity. Methods: Recordings of CMMCsere made from the isolated murine large bowel, withr without a fecal pellet. Transducers were placed alonghe colon to record muscle tension and propulsive forcen the pellet and microelectrodes were used to recordlectrical activity from either side of a fecal pellet, cir-ular muscle cells oral and anal of a pellet, and in colonsithout the mucosa. Results: Spontaneous CMMCsropagated in both an oral or anal direction. When aellet was inserted, CMMCs increased in frequency andropagated anally, exerting propulsive force on the pel-
et. The amplitude of slow waves increased during theMMC. Localized mucosal stimulation/circumferen-
ial stretch evoked a CMMC, regardless of stimulustrength. The serotonin (5-hydroxytryptamine-3) recep-or antagonist ondansetron reduced the amplitude ofhe CMMC, the propulsive force on the pellet, and theesponse to mucosal stroking, but increased the appar-nt conduction velocity of the CMMC. Removing theucosa abolished spontaneous CMMCs, which still
ould be evoked by electrical stimulation. Conclusions:he fecal pellet activates local mucosal reflexes, which
elease serotonin (5-hydroxytryptamine) from entero-hromaffin cells, and stretch reflexes that determine theite of origin and propagation of the CMMC, facilitat-ng propulsion.
bnormal colonic motor patterns have been impli-cated in the pathogenesis of severe constipation and
iarrhea.1–5 One such major motor pattern involved inegulating the movement of fecal matter is the colonic
igrating motor complex (CMMC).3– 8 The CMMC con-ists of a rhythmically occurring sequence of electricalctivity and/or contractions that can occur as either atationary event or as a propagating wave of contractionshat conduct in either an oral or anal direction overarying lengths of the large bowel. CMMCs are generated
y the enteric nervous system because they are blocked byetrodotoxin and hexamethonium, and reduced by atro-ine.3–7 Although it has been proposed that discrete neu-al pattern generators within the myenteric plexus andistributed along the colon generate the CMMC,8 oururrent studies suggest that serotonin (5-hydroxytrypta-ine [5-HT]) release from the mucosa is required for
riggering the neural circuitry underlying the CMMC.A study of the literature suggests that, similar to heart
ate,9 the frequency of the CMMC may be related in-ersely to body mass, occurring at a higher frequency inmall mammals such as the mouse,4 –7 compared with theolon of larger mammals,3,10 –14 including humans.1–3,13
lthough the CMMC occurs during both the fasted anded state,3,11 they have been observed to occur at a higherrequency in the fed state.11,15 In humans, CMMCs ap-ear to be the events referred to as propagating (pressure)
equences.13 Propagating sequences propagate mainly, butot exclusively, in the anal direction in control subjects,hereas in patients with obstructed defecation there is a
ignificant increase in orally propagating sequences.13
Serotonin (5-HT) plays an essential role in gastrointes-inal motility.16,17 5-HT3–receptor antagonists, which re-uce intestinal transit, are used to treat multiple symp-oms in patients with diarrhea-predominant irritableowel syndrome. However, because of rare, but poten-ially serious, side effects, their use currently is restrictedo patients who are refractory to conventional therapyecause they can predispose some patients to develop
schemic colitis.16 –18 5-HT3–receptor antagonists such asndansetron and alosetron have been shown to reducehe amplitude of spontaneous CMMCs in the isolatedmpty murine large bowel.19,20 In the isolated colon-HT3 antagonists can act only on enteric mechanisms.-HT is known to be secreted by enterochromaffin (EC)ells and acts on 5-HT3 receptors located on intrinsicrimary afferent neurons or in neurons in descendingerve pathways.16,17
Abbreviations used in this paper: CM, circular muscle; CMMC, co-onic migrating motor complex; EC, enterochromaffin; EJP, excitatoryunction potential; 5-HT, 5-hydroxytryptamine (serotonin); IJP, inhibi-ory junction potential; TNS, transmural nerve stimulation.
© 2009 by the AGA Institute0016-5085/09/$36.00
doi:10.1053/j.gastro.2008.12.010

fIgort
kBdor
moemlwatcst1
F(CpiowmstoAhpa(sausfoawTctssopsctttmtccot
BA
SIC–
ALI
MEN
TARY
TRA
CT
April 2009 SYNERGY BETWEEN A FECAL PELLET AND CMMC 1329
We have examined whether CMMCs are essential forecal pellet propulsion in the isolated murine large bowel.mportantly, we found that there appears to be a syner-istic relationship between the CMMC and the presencef a fecal pellet that generates local intrinsic reflexes toegulate the generation and direction of propagation ofhe CMMC.
Materials and Methods
C57BL/6 mice (20 –90 days old) of either sex wereilled humanely by inhalation of anesthetic (Isofluorane;axter, Deerfield, IL) and cervical dislocation, in accor-ance with the animal ethics committee of the Universityf Nevada School of Medicine, and the whole colon was
igure 1. Experimental setups.A) Effect of a fecal pellet on theMMC. An epoxy-coated fecalellet attached to a silk thread was
nserted into an isolated segmentf colon and was fixed in positionith a pin when it reached theiddle of the colon. Isometric ten-
ion transducers monitored CMension before and after insertionf the pellet. (B) Partitioned bath.n epoxy-coated fecal pellet waseld in position between the 2artitions by attaching a thread viapulley to a tension transducer.
C) Mucosal and stretch reflex. Aection of the colon was cut opennd pinned with the mucosappermost, allowing mechanicaltimulation with a brush. Circum-erential stretch was applied tone edge of the opened region byttaching it to a pulley system tohich weights could be added.ension transducers monitoredontractile activity. (D) Microelec-rode recordings made on eitheride of a fecal pellet. Two 10-mmections of colon were cut open,n either side of a fecal pellet, andinned circumferentially, with theerosa uppermost, so that electri-al activity could be recorded fromhe underlying CM. (E) Microelec-rode recordings from prepara-ions without the mucosa. Theucosa was dissected away from
he colon, which was pinned cir-umferentially. Microelectrode re-ordings were made 10-mm analf transmural stimulating elec-rodes (TMS).
emoved.6,7,20 w
PreparationsSeveral preparations were used in this study to
onitor circular muscle (CM) tension, propulsive forcen a pellet, and electrical activity during spontaneous orvoked CMMCs, as well as in preparations devoid of theucosa. The preparations were designed to do the fol-
owing: (1) monitor spontaneous CMMC activity with orithout a fixed pellet (Figure 1A); (2) confine drugs usingpartitioned chamber with a fixed pellet to sites around
he pellet (Figure 1B); (3) evoke intrinsic reflexes by mu-osal stimulation applied with a brush or circumferentialtretch (Figure 1C); (4) record intracellular electrical ac-ivity from CM cells oral and anal to a fixed pellet (FigureD); and (5) record electrical activity in preparations
ithout the mucosa (Figure 1E).
tcit(i
pafia
o
(Ciis
ttPwtstipswtm6amw
Cips
fwl
aa1
wntpb
t3s(vt
tvrsts
pfT11C
mAcntofC1oaBn5
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1330 HEREDIA ET AL GASTROENTEROLOGY Vol. 136, No. 4
The isolated large bowel was attached to the floor ofhe organ bath by pinning the mesentery. An epoxy-oated fecal pellet, with a thread glued to one end, wasnserted into the oral end, allowed to propagate down tohe middle of the colon and held by either a threadFigure 1A), a force transducer (Figure 1B), or by pinningn front and behind a pellet (Figure 1D).
Two greased polycarbonate (Evonik CYRO LLC, Parsi-anny, NJ) partitions were slotted into position (15-mmpart) around the pellet (Figure 1B). The partitions ef-ectively divided the colon, for pharmacological purposes,nto an oral segment, a middle (stimulation) segment,nd an anal segment.21
Each chamber or preparation was perfused continu-usly with Krebs solution at 37.0°C � 0.5°C.20 –22
Recording the Mechanical Activity of theCircular MuscleSuture silk was used to connect force transducers
model TST125C; Biopac Systems Inc., Santa Barbara,A) along the colon (Figure 1A–C). Resting tension was
nitially set at 8 mN, and monitored using an MP100nterface and recorded on a PC running Acqknowledgeoftware 3.2.6 (Biopac Systems, Inc.).20
Evoking Neural ReflexesThe colon was opened along a 10-mm length in
he middle, the oral end, or the anal end of the prepara-ion and pinned with the mucosa uppermost (Figure 1C).ressure or brush strokes were applied to the mucosaith a script liner 5/0 artist’s brush. A rake-holder system
hen was attached to the opposite edge so that it could betretched circumferentially using weights (2–20 g) addedo a pulley system (Figure 1C).23,24 The stretch was mon-tored by viewing the preparation with a video cameraositioned over the stimulation site (see below). When notretch was applied, the opened segment of colon had aidth of 4.20 � 0.01 mm. After the addition of weights
he average width increased as follows: 2 g: 5.42 � 0.01m; 5 g: 6.05 � 0.03 mm; 10 g: 6.40 � 0.01 mm; 15 g:
.70 � 0.03 mm; and 20 g: 6.90 � 0.03 mm (n � 4). Theverage circumference of a fecal pellet was 6.4 � 0.01m, which corresponded to the width applied by a 10-geight.
Intracellular Recording From Whole-ColonPreparations With and Without MucosaDual microelectrode impalements were made into
M cells as described previously.7,22–25 Electrical record-ngs were made from the CM on either side of a fixedellet by advancing the electrode through the serosalurface of the open segment (Figure 1D).
In other experiments, the mucosa was dissected awayrom the whole colon that was pinned circumferentiallyith the serosa uppermost. Platinum transmural stimu-
ating wires (diameter, 0.2 mm), which were connected to b
Grass SD stimulator (West Warwick, RI), were placedbove and below the preparation (1-mm apart and0-mm oral of the recording site).
Video Recording of Colonic Movements andSpatiotemporal Map AnalysisThe propagation of a fecal pellet along the colon
as monitored with a video camera (WV-BP330; Panaso-ic CCTV, Seacaucus, NJ; Powermac G4 Apple, Cuper-ino, CA). Video frames were converted into spatiotem-oral diameter or D maps, from which the velocity coulde calculated.22,25
Analysis of Data and Statistical MethodsThe frequency, duration, and amplitude of con-
ractile complexes were measured using Acqknowledge.2.6 (Biopac Systems, Inc, Goleta, CA) and tests fortatistical significance were made using Sigma Plot 5.0Jandel Scientific, San Rafael, CA).20 The propagationelocity was determined by calculating the delay betweenhe 50% amplitude points of CMMCs.20
Statistical comparisons of data were performed usinghe Student (paired or unpaired) t tests or analysis ofariance, and a minimum level of significance waseached at a P value of less than .05. In the Resultsection, n refers to the number of animals from whichissue was taken. All data are presented as means �tandard error of the mean.
Drugs and SolutionsHexamethonium bromide and ondansetron were
urchased from Sigma (St. Louis, MO). Alosetron was a giftrom Glaxo Pharmaceuticals (Research Triangle Park, NC).he Krebs solution was (in mmol/L) 120.35 NaCl, 5.9 KCl,5.5 NaHCO3, 1.2 NaH2PO4, 1.2 MgSO4, 2.5 CaCl2, and1.5 glucose, gassed continuously with a mixture of 3%O2–97% O2 (vol/vol), to give a final pH of 7.3–7.4.
ResultsSpontaneous Activity Without a Fecal PelletTension recordings of spontaneous CMMCs were
ade along the isolated large bowel (Figures 1A and 2A).t the oral recording site in the proximal colon theseonsisted of irregular low-frequency (0.28 � 0.08 c/min;� 8), low-amplitude (12.0 � 1.3 mN), and long-dura-
ion (150.00 � 14.6 s) CMMCs that consisted of a seriesf phasic contractions that usually failed to propagatearther down the proximal colon (Figure 2A*). TheseMMCs were interspersed with higher-amplitude (18.03 �.27 mN), shorter-duration (61.63 � 7.68 s) CMMCs, whichccurred at a frequency of 0.31 � 0.03 c/min that prop-gated in either an oral or anal direction (Figure 2A#).oth types of spontaneous CMMCs were blocked by theicotinic antagonist hexamethonium (100 �mol/L; n �), suggesting they required fast synaptic transmission to
e generated.
pwwaWCppt
ptepow(C6ampncac
pt
tr
bsgsmentmslTtibslTb
FoCla(icmf(dg(wpTcC(ftt
BA
SIC–
ALI
MEN
TARY
TRA
CT
April 2009 SYNERGY BETWEEN A FECAL PELLET AND CMMC 1331
Activity Induced by a Fecal PelletWhen an epoxy-coated fecal pellet was allowed to
ropagate down the colon and then held in positionhen it reached the middle of the colon, regular CMMCsere generated that were of shorter duration but highermplitude, an example of which is shown in Figure 2B.hen the pellet was present, the overall frequency of
MMCs was faster (control: 0.17 � 0.01 c/min; withellet: 0.31 � 0.03; P � .01; n � 8). The CMMCs ap-eared to originate oral to the pellet and propagate downhe colon in an oral to anal direction (Figure 2B and C).
We also measured the propulsive tension on the fixedellet held in the middle of the large bowel, as well asension of the CM along the colon (Figure 1B). In thesexperiments, CMMCs appeared to originate oral to theellet and propagate anally. The peak propulsive tensionn the pellet, which often outlasted the CMMC, occurredhen the CMMC was detected just anal to the pellet
Figure 2C). The apparent conduction velocity of theMMC over the fixed pellet was 0.80 � 0.10 mm/s (n �), which was similar to the velocity of a pellet that wasllowed to propagate freely down the colon (0.82 � 0.1m/s; n � 6; Figure 2C and D). When the 3-chambered
artitioned bath was used, hexamethonium (100 �mol/L;� 5) applied to the middle chamber containing the pellet
ompletely blocked the CMMC in both the middle andnal chambers and disrupted the CMMC in the oral
igure 2. Effect of a fecal pelletn spontaneous CMMCs. (A)MMC activity in an empty co-
on. *Nonpropagated burst ofctivity. #Propagated CMMCs.B) When an artificial pellet wasntroduced into the middle of theolon, CMMC activity becameore organized and propagated
rom the oral tension transducerTo) to the anal tension trans-ucer (Ta). (C) CMMCs propa-ated in an oral to anal direction
see inset) and were associatedith considerable force on theellet (Tension pellet top trace).he peak tension on the pelletoincided with the peak of theMMC in the middle chamber.
D) The conduction velocity of areely moving fecal pellet downhe colon was measured fromhe spatiotemporal map.
hamber (Figure 3A). Therefore, long cholinergic nerve n
athways are involved in the coordination and propaga-ion of the CMMC.
Pellet Propulsion and Intrinsic ReflexesWe attempted to determine whether a pellet con-
rols the initiation of the CMMC by activating localeflexes.
Stroking the mucosa or applying pressure with therush (1–10 strokes or pressure) or circumferentialtretch (5–20 g) in the middle, or at either end, alwaysenerated a CMMC that was similar if not identical topontaneous CMMCs. Evoked CMMCs were usually
ore synchronized and propagated smoothly along thentire length of the colon (Figure 3B and C). There waso response to circumferential stretch to weights lesshan 5 g, which corresponds to a width of less than 6.05
m (unstretched flap, 4.20 mm), suggesting stretch re-ponses had a threshold for activation that was slightlyess than the circumference of a fecal pellet (6.40 mm).he level of circumferential stretch at 10 g (6.40 mm) was
he same as the circumference of a fecal pellet. Unlikentrinsic nervous reflexes in the guinea pig and rat largeowel, a graded response to stimulation was never ob-erved regardless of the number of brush strokes or theevel of circumferential stretch (Figure 3B and C).21–26
he evoked CMMC responses to either set of stimuli werelocked completely by hexamethonium (100 �mol/L;
� 5; Figure 3B and C).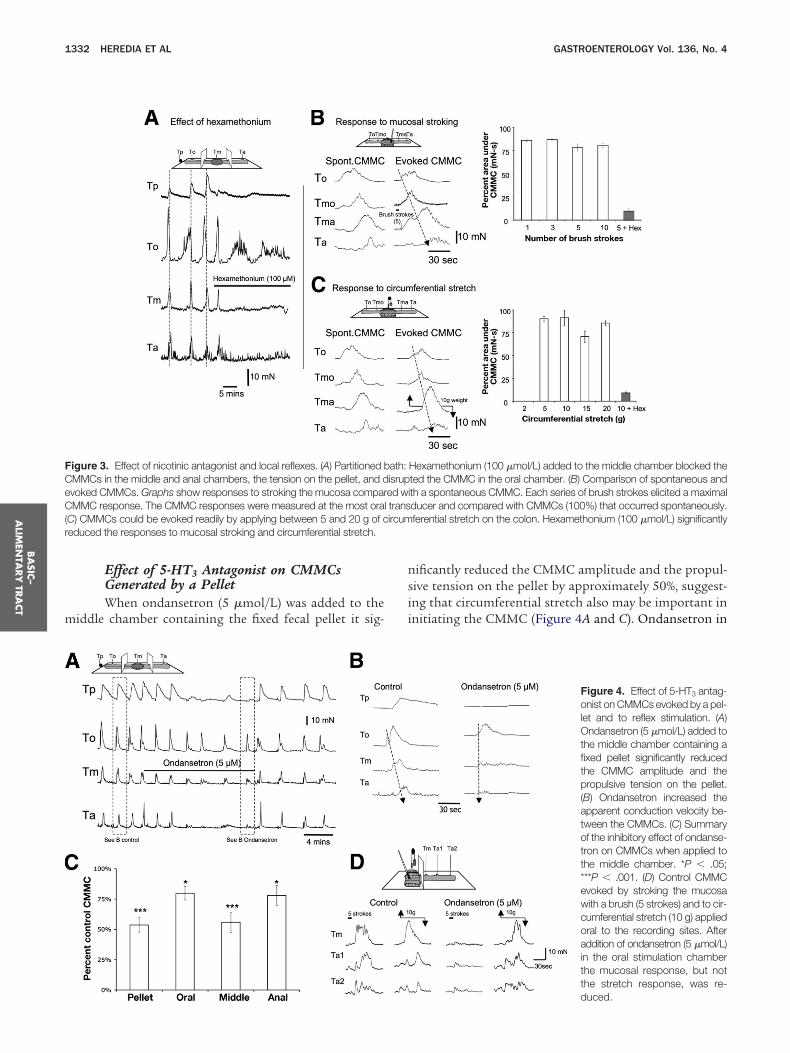
m
nsii
FCeC(r
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1332 HEREDIA ET AL GASTROENTEROLOGY Vol. 136, No. 4
Effect of 5-HT3 Antagonist on CMMCsGenerated by a PelletWhen ondansetron (5 �mol/L) was added to the
iddle chamber containing the fixed fecal pellet it sig-
igure 3. Effect of nicotinic antagonist and local reflexes. (A) Partitioned bMMCs in the middle and anal chambers, the tension on the pellet, and dvoked CMMCs. Graphs show responses to stroking the mucosa comparMMC response. The CMMC responses were measured at the most ora
C) CMMCs could be evoked readily by applying between 5 and 20 g of ceduced the responses to mucosal stroking and circumferential stretch.
ificantly reduced the CMMC amplitude and the propul-ive tension on the pellet by approximately 50%, suggest-ng that circumferential stretch also may be important innitiating the CMMC (Figure 4A and C). Ondansetron in
Figure 4. Effect of 5-HT3 antag-onist on CMMCs evoked by a pel-let and to reflex stimulation. (A)Ondansetron (5 �mol/L) added tothe middle chamber containing afixed pellet significantly reducedthe CMMC amplitude and thepropulsive tension on the pellet.(B) Ondansetron increased theapparent conduction velocity be-tween the CMMCs. (C) Summaryof the inhibitory effect of ondanse-tron on CMMCs when applied tothe middle chamber. *P � .05;***P � .001. (D) Control CMMCevoked by stroking the mucosawith a brush (5 strokes) and to cir-cumferential stretch (10 g) appliedoral to the recording sites. Afteraddition of ondansetron (5 �mol/L)in the oral stimulation chamberthe mucosal response, but notthe stretch response, was re-
Hexamethonium (100 �mol/L) added to the middle chamber blocked theted the CMMC in the oral chamber. (B) Comparison of spontaneous andth a spontaneous CMMC. Each series of brush strokes elicited a maximalducer and compared with CMMCs (100%) that occurred spontaneously.ferential stretch on the colon. Hexamethonium (100 �mol/L) significantly
ath:isruped wil transircum
duced.

ttmF
�sms
ifitpobiSttg
iq
iswa5doadahw
wp�i6p
T
C
C
S
C
1 �
BA
SIC–
ALI
MEN
TARY
TRA
CT
April 2009 SYNERGY BETWEEN A FECAL PELLET AND CMMC 1333
he middle chamber also increased the apparent conduc-ion velocity between CMMCs (control: 0.71 � 0.04
m/s; ondansetron: 3.03 � 0.29 mm/s; P � .01; n � 8;igure 4B).
Effect of 5-HT3 Antagonists on Local ReflexesEither ondansetron (5 �mol/L) or alosetron (5
mol/L) applied specifically to the stimulation chamberignificantly reduced the CMMC response to stroking the
ucosa, but did not significantly affect the CMMC re-ponse to circumferential stretch (Figure 4D; Table 1).
CMMCs Recorded With IntracellularMicroelectrodesSimultaneous intracellular microelectrode record-
ngs were made from CM on the oral and anal side of axed pellet (Figure 1D) in unparalyzed muscle prepara-ions, had a resting membrane potential (between com-lexes) of 62.47 � 2.61 mV (n � 5). Regular CMMCs werebserved that consisted of a hyperpolarization followedy a prolonged depolarization, upon which were super-
mposed excitatory junction potentials (EJPs) (Table 1).7
everal action potentials (2.60 � 0.27) were observed onhe depolarizing phase of the CMMC. It was clear thathe CMMC originated oral to a fixed pellet and propa-
able 1. Effect of 5-HT3 Antagonist on Components of the C
C
MMCs evoked by local reflexesMucosal brush strokes, area under the curve mN-s 343.8Circumferential stretch, area under the curve mN-s 374.8
Mucosal brush strokes, area under the curve mN-s 288Circumferential stretch, area under the curve mN-s 333
MMCs evoked by a fixed pelletResting membrane potential, mV 62.4Interval between CMMCs, s 225.2Duration of CMMCs, s 50.3Amplitude of hyperpolarization, mV 12.1Amplitude of depolarization, mV 15.3Number of action potentials on depolarization 2.6
pontaneous junction potentialsInterval between IJPs, s 2.1Amplitude of IJPs, mV 10.0Duration of IJPs, mV 0.6Duration of EJPs, s 0.6Amplitude of EJPs, mV 8.4
Atropin
MMCs evoked by TNS (no mucosa)Amplitude of depolarization, mV 16.4Duration of depolarization, s 44.8Amplitude of hyperpolarization, mV 13.4Duration of hyperpolarization, s 44.8
ated anally (Figure 5A). Between CMMCs, spontaneous r
nhibitory junction potentials (IJPs) occurred at a fre-uency of 28.57 cpm (n � 5; Table 1).
Furthermore, when the electrode was advanced deepernto the tissue towards the submucosal border electricallow waves were recorded (Figure 5A and B). The slowaves became more periodic and increased in amplitudend elicited action potentials during the CMMC (FiguresB and 6A). During this period, slow waves within theeep CM had a frequency of 17.14 c/min, an amplitudef 20.39 � 1.46 mV (maximum amplitude, 35 mV), andduration of 3.50 � 0.16 s (n � 4). The slow waves wereistinguished easily from EJPs (duration, 0.60 � 0.05 s;mplitude, 8.49 � 0.59 mV; P � .01) because slow wavesad a significantly longer duration and unlike the EJPsere not blocked by atropine (1 �mol/L; n � 3).
Effect of 5-HT3 AntagonistWhen intracellular microelectrode recordings
ere made from unparalyzed CM just anal to a fixed fecalellet (Figure 1D) we found that ondansetron (1–3mol/L; n � 4) abolished the CMMCs without a signif-
cant change in the resting membrane potential (FigureA; Table 1). Ondansetron significantly reduced the am-litude, duration, and frequency of spontaneous IJPs and
l Ondansetron (5 �mol/L) n P value
35.61 65.85 � 20.26 4 �.0146.2 343.72 � 20.26 4 �.05
Alosetron, 5 �mol/L
33.6 45.7 � 20.3 4 �.0146.2 326.7 � 33.6 4 �.05
Ondansetron, 1–3 �mol/L
2.61 60.98 � 3.63 5 �.0539.54 Blocked 4 �.012.46 Blocked 4 �.010.50 Blocked 4 �.011.41 Blocked 4 �.010.27 Blocked 4 �.01
Ondansetron, 1–3 �mol/L
0.15 12.05 � 0.65 5 �.050.57 4.79 � 0.57 5 �.050.03 0.58 � 0.05 5 �.050.05 1.23 � 0.11 5 �.050.59 9.34 � 0.25 5 �.05
�mol/L Ondansetron, 1 �mol/L
1.55 17.19 � 1.55 5 �.015.66 49.7 � 4.34 5 �.012.05 3.74 � 0.36 5 �.015.66 21.85 � 2.48 5 �.01
MMC
ontro
5 �5 �
.7 �
.4 �
7 �2 �9 �8 �5 �0 �
0 �5 �5 �0 �9 �
e, 1
2 �1 �4 �
evealed more ongoing EJPs (Figure 6A; Table 1).

tip1pptrbci
f
ap(tdTs
tTnTfi
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1334 HEREDIA ET AL GASTROENTEROLOGY Vol. 136, No. 4
Effect of Removing the MucosaThe effects of ondansetron on the CMMC suggest
hat 5-HT release from EC cells in the mucosa16,17 may bemportant for generating the CMMC. To test this hy-othesis we removed the mucosa from the colon (FigureE). When impalements were made from CM cells inreparations without the mucosa the resting membraneotential (63.08 � 3.52 mV; P � .01; n � 4) was similaro that in colons with an intact mucosa. In these prepa-ations without the mucosa, CMMCs were not presentut ongoing fast IJPs still occurred at a frequency (21.43pm) similar to those in preparations with the mucosantact (Figure 6B; Table 1).
Transmural nerve stimulation (TNS) (pulses of 10–20 Hz
or 1 s, 0.4 ms, 30 V) evoked a CMMC, consisting of either wdepolarization, or a hyperpolarization followed by a de-olarization (Figure 6C; Table 1). The addition of atropine
1 �mol/L) significantly increased the amplitude and dura-ion of the preceding hyperpolarizion to TNS, whereas theepolarizing response was similar to control (Figure 6D;able 1). After several TNSs, a spontaneous CMMC occa-
ionally was observed (Figure 6E).In the presence of atropine, the addition of ondanse-
ron (1 �mol/L) caused a significant reduction in theNS-evoked hyperpolarizing response, but did not sig-ificantly affect the depolarizing response (Figure 6F;able 1). This suggests that ondansetron was reducing
ast synaptic transmission via 5-HT3 receptors27 to inhib-tory motor neurons. In addition, the spontaneous IJPs
Figure 5. Electrical activity re-corded oral and anal to a fixed pel-let. (A) The top trace (EO) illustratesslow-wave activity in the CM oralto the pellet, and the bottom trace(Ea) shows the electrical activity re-corded from a CM muscle cellanal to the pellet. (B) Slow-waveactivity during the CMMC oral to afecal pellet. Note that before theCMMC occurs there is a reductionin the excitability of the cell,whereas when the CMMC occursthere is an increase in the ampli-tude of slow waves, which elicitaction potentials. Black dots indi-cate the start of the CMMC. Theexpanded trace shows the char-acteristic depolarization during theCMMC.
ere reduced in frequency by ondansetron (3.16 c/min).

lrbisoCacgfl
bcig5t
dii
FtSstdf(bmthq((mss1tawmhpCsssdtoda
BA
SIC–
ALI
MEN
TARY
TRA
CT
April 2009 SYNERGY BETWEEN A FECAL PELLET AND CMMC 1335
DiscussionOur study suggests that there is a synergistic re-
ationship between the spontaneous CMMCs and localeflexes generated by a fecal pellet in the murine largeowel. The propagation of a fecal pellet down the colon
s dependent on the CMMC, which normally occurspontaneously in an empty colon. However, the presencef a fecal pellet determines the site of origin of a colonicMMC and its apparent propagation down the colon byctivating both mucosal reflexes and reflexes activated byircumferential stretch. The neural circuitry primarilyenerates a CMMC in response to input from local re-
igure 6. Effect of 5-HT3 an-agonist on electrical activity. (A)pontaneous CMMCs were ob-erved in a CM cell 10-mm analo a fixed pellet. Between anduring the CMMC, spontaneous
ast IJPs occurred. Ondansetron1 �mol/L) blocked the CMMCsut did not affect the restingembrane potential. The spon-
aneous IJPs (see expanded left-and trace) were reduced in fre-uency and EJPs were revealed
see expanded right-hand trace).B) When the mucosa was re-
oved from the colon onlypontaneous IJPs were ob-erved. (C) TMS (0.4 ms, 30 V,0–20 Hz for 1 s) applied oral tohe recording electrode evoked
CMMC in preparations inhich the mucosa was re-oved. (D) Atropine revealed the
yperpolarization before the de-olarizing phase of the evokedMMC. (E) Occasionally, aftereveral periods of stimulation, apontaneous CMMC was ob-erved. (F) The addition of on-ansetron significantly reducedhe hyperpolarizing componentf the CMMC while leaving theepolarizing phase relatively un-ffected.
exes, rather than peristaltic reflexes. CMMCs initiated g
y mucosal reflexes likely involve 5-HT release from ECells in the mucosa, which activate 5-HT3 receptors onntrinsic sensory neurons.16,17 The neural circuitry forenerating the CMMC is in the myenteric plexus, but-HT release from the mucosa is essential for triggeringhe CMMC.
Propagation of a PelletPreviously it has been assumed that the CMMC
rives pellet propulsion.4 –7 We found that a freely mov-ng pellet propagates down the colon at a velocity that isdentical to the apparent conduction velocity of a CMMC
enerated by a fixed pellet, suggesting the 2 events may be
eTapwpC
srmncbi
ndgearutp
Cs
sostpimcabsmsbtTtgp5pm
Fmad uronc
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1336 HEREDIA ET AL GASTROENTEROLOGY Vol. 136, No. 4
quivalent and that the CMMC drives pellet propulsion.his strongly is supported by the observation that whenpellet is fixed in the colon, the CMMC nearly always
ropagates in an oral to anal direction and is associatedith considerable tension on a pellet. The tension on theellet reached a maximum after the passage of theMMC over the pellet.
Slow Waves and the CMMCSlow waves in the large bowel originate in inter-
titial cells of Cajal at the submucosal border.28,29 Theeason slow waves have not been reported during the
urine CMMC is that they are sensitive to L-type chan-el blockers,29 which are used routinely to prevent mus-le contraction.7 Slow waves near the submucosal borderecame more periodic and elicited action potentials dur-
ng the CMMC.
Local Intrinsic ReflexesThe fact that the presence of a fecal pellet orga-
izes the CMMC suggests that the pellet itself exerts aominant effect on the neural circuitry responsible forenerating CMMCs. We assumed that the pellet wouldither stimulate the mucosa or stretch the bowel toctivate local peristaltic reflexes.24 In the guinea pig andat colon these reflexes are graded according to the stim-lus strength.21,24,26 Therefore, we were surprised to findhat mucosal stimulation and circumferential stretch ap-
igure 7. Possible elements of intrinsic neural circuit underlying the Cucosa, which activates sensory neurons via 5-HT3 receptors. Activati
s the activation of ascending excitatory cholinergic nerve pathways,escending serotonergic nerve pathways that excite inhibitory motor neircumferential stretch.
lied to the murine large bowel elicited a maximal t
MMC response above a threshold regardless of thetrength of stimulation.
Role of 5-HT3 ReceptorsWe found that ondansetron (and alosetron) sub-
tantially reduced the responses to mechanical stimulationf the mucosa but not the responses to circumferentialtretch, and significantly reduced the propulsive tension onhe fixed pellet and the amplitude of the CMMC over theellet when added to the middle chamber. Presumably, the
ncomplete blockade of the CMMCs by odansetron in theiddle chamber may be due to the reflex response to
ircumferential stretch elicited by the pellet. It could bergued that the remaining response in the middle cham-er involved other 5-HT receptors30 –32; however, thiseems unlikely because ondansetron and alosetron al-
ost completely blocked the response to mucosaltimulation. In addition, ondansetron also completelylocked CMMCs recorded with intracellular microelec-rodes without changing resting membrane potential.hese results suggest that 5-HT released from EC cells in
he mucosa is important for initiating mucosal reflexesenerated by stroking the mucosa or from mucosal com-ression by the pellet. Previously, it has been shown that-HT can activate the endings of myenteric intrinsicrimary afferent neurons (or AH neurons) within theucosa of both the guinea pig large and small intes-
. The presence of a pellet increases 5-HT release from EC cells in theintrinsic sensory neurons leads to the generation of the CMMC as wellh activate excitatory motor neurons (EMN) via nicotinic receptors ands (IMN) via 5-HT3 receptors. The interneurons also may be activated by
MMCon ofwhic
ine,33,34 largely by stimulating 5-HT3 receptors.34 In con-

ta
itbm(tsTwC
tcsasnmw
otctosCttnIttgcimvlhapCna
fsas
mb
1
1
1
1
1
1
1
1
1
1
2
2
BA
SIC–
ALI
MEN
TARY
TRA
CT
April 2009 SYNERGY BETWEEN A FECAL PELLET AND CMMC 1337
rast, the mucosal endings of submucosal AH neuronsre activated by 5-HT1P receptors.35
However, ondansetron added to the chamber contain-ng the fecal pellet also increased the apparent conduc-ion velocity of the CMMC, suggesting that it also may belocking 5-HT3 receptors mediating fast synaptic trans-ission27 in descending inhibitory nervous pathways
Figure 7). Likely, 5-HT3 receptors are on inhibitory mo-or neurons because ondansetron reduced the number ofpontaneous IJPs and the hyperpolarization evoked byNS. The activation of descending inhibitory nerve path-ays is essential for the apparent propagation of theMMC.7
Mucosa Necessary for Triggering the CMMCRemoving the mucosa appeared to abolish spon-
aneous CMMCs, suggesting that the mucosa is normallyritical for their generation. However, transmural nervetimulation could elicit a CMMC-like response. Also, intropine, an occasional spontaneous CMMC was ob-erved after several periods of TNS, suggesting that theeural circuitry underlying the CMMC was within theyenteric plexus, but stimuli arising from the mucosaere necessary for triggering the CMMC.The trigger for the CMMC appears to be spontaneous
r evoked (ie, a fecal pellet) release of 5-HT from EC cellso stimulate AH neurons, whereas reflexes activated byircumferential stretch may involve mechanosensitive in-erneurons.23,36 A CMMC can be initiated from any partf the colon; however, fecal pellets focus the initiationite and propagation direction (oral to anal) of theMMC. Presumably, the pressure exerted by a pellet on
he mucosa increases 5-HT release that would increasehe probability that a critical number of myenteric AHeurons are activated for the generation of the CMMC.
n fact, the migrating motor complex in the small intes-ine has been modeled using recurrent connections be-ween AH neurons.37 The concept of discrete patternenerators distributed along the colon8 is unlikely be-ause each ganglia contains AH neurons that are likely tontegrate different levels of sensory information from the
ucosa to form the CMMC, probably through the acti-ation of slow excitatory postsynaptic potentials by re-ease of calcitonin gene-related peptide.30,36 The effects ofexamethonium suggest that the enteric neural pathwayslong the whole length of the colon are involved inropagating a CMMC. The apparent propagation of theMMC involves activation of descending inhibitoryerve pathways, and the sudden coordination of manyscending and descending interneurons (Figure 7).7
In summary, there is a synergistic relationship betweenecal matter and the CMMC. A fecal pellet influences theite of generation and propagation of the CMMC byctivating local reflexes that enhance activity in intrinsic
ensory neurons. Furthermore, the interactions betweenany fecal pellets and the CMMC need further studyecause these are likely to be more complex.22,25
References
1. Bueno L, Fiormonti J, Flexinos J, et al. Colonic myoelectricalactivity in diarrhea and constipation. Hepatogastroenterology1980;27:381–389.
2. Snape WJ Jr, Carlson GM, Matarazzo SA, et al. Evidence thatabnormal myoelectrical activity produces colonic motor dysfunc-tion in the irritable bowel syndrome. Gastroenterology 1977;72:383–387.
3. Sarna SK. Physiology and pathophysiology of colonic motor activ-ity (1). Dig Dis Sci 1991;36:827–862.
4. Brann L, Wood JD. Motility of the large intestine of piebald-lethalmice. Am J Dig Dis 1976;21:633–640.
5. Spencer NJ, Bayguinov P, Hennig GW, et al. Activation of neuralcircuitry and Ca2� waves in longitudinal and circular muscleduring CMMCs and the consequences of rectal aganglionosis inmice. Am J Physiol 2007;292:G546–G555.
6. Bywater RA, Small RC, Taylor GS. Neurogenic slow depolarisa-tions and rapid oscillations in circular muscle of mouse colon.J Physiol 1989;413:505–519.
7. Spencer NJ, Hennig GW, Dickson E, et al. Synchronization ofenteric neuronal firing during the murine colonic MMC. J Physiol2005;564:829–847.
8. Wood JD. Enteric nervous system: reflexes, pattern generatorsand motility. Curr Opin Gastroenterol 2008;24:149–158.
9. West GB, Brown JH, Enquist BJ. The origin of universal scalinglaws in biology. In: Brown JH, West GB, eds. Scaling in biology.Santa Fe Institute Studies in the Sciences of Complexity Proceed-ings. Oxford University Press, Cary, NC: 2000:87–112.
0. Roger T, Ruckebusch Y. Pharmacological modulation of postpran-dial colonic motor activity in the pony. J Vet Pharmacol Ther1987;10:273–282.
1. Cherbut C, Ruckebusch Y. The effect of indigestible particles ondigestive transit time and colonic motility in dogs and pigs. Br JNutr 1985;53:549–557.
2. Ferré JP, Ruckebusch Y. Myoelectrical activity and propulsion in thelarge intestine of fed and fasted rats. J Physiol 1985;362:93–106.
3. Dinning PG, Szczesniak MM, Cook IJ. Twenty-four hour spatiotem-poral mapping of colonic propagating sequences provides patho-physiological insight into constipation. Neurogastroenterol Motil2008;20:1017–1021.
4. Scott SM, Pilot MA, Barnett TG, et al. Prolonged ambulatorycanine colonic motility. Am J Physiol 199;268:G650–G662.
5. Powell AK, Fida R, Bywater RA. Motility in the isolated mousecolon: migrating motor complexes, myoelectric complexes andpressure waves. Neurogastroenterol Motil 2003;15:257–266.
6. Gershon MD, Liu MT. Serotonin and neuroprotection in functionalbowel disorders. Neurogastroenterol Motil 2007;19(Suppl 2):19–24.
7. De Giorgio R, Barbara G, Furness JB, et al. Novel therapeutictargets for enteric nervous system disorders. Trends PharmacolSci 2007;28:473–481.
8. Fayyaz M, Lackner JM. Serotonin receptor modulators in thetreatment of irritable bowel syndrome. Ther Clin Risk Manag2008;4:41–48.
9. Fida R, Bywater RA, Lyster DJ, et al. Chronotropic action of5-hydroxytryptamine (5-HT) on colonic migrating motor complexes(CMMCs) in the isolated mouse colon. J Auton Nerv Syst 2000;80:52–63.
0. Bush TG, Spencer NJ, Watters N, et al. Effects of alosetron onspontaneous migrating motor complexes in murine small andlarge bowel in vitro. Am J Physiol 2001;281:G974–G983.
1. Smith TK, McCarron S. Nitric oxide modulates cholinergic reflexpathways to the longitudinal and circular muscle in the isolated
guinea-pig distal colon. J Physiol 1998;512:893–906.
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
3
R
CN6
A
rr
C
F
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1338 HEREDIA ET AL GASTROENTEROLOGY Vol. 136, No. 4
2. Dickson EJ, Spencer NJ, Hennig GW, et al. An enteric occult reflexunderlies accommodation and slow transit in the distal largebowel. Gastroenterology 2007;132:1912–1924.
3. Spencer NJ, Dickson EJ, Hennig GW, et al. Sensory elementswithin the circular muscle are essential for mechanotransductionof ongoing peristaltic reflex activity in guinea-pig distal colon.J Physiol 2006;576:519–531.
4. Spencer NJ, Smith TK. Simultaneous intracellular recordingsfrom longitudinal and circular muscle during the peristaltic reflexin guinea-pig distal colon. J Physiol 2001;533:787–799.
5. Dickson EJ, Hennig GW, Heredia DJ, et al. Intrinsic neural reflexes inresponse to colonic elongation. J Physiol 2008;86:4225–4240.
6. Grider JR, Makhlouf GM. Regulation of the peristaltic reflex bypeptides of the myenteric plexus. Arch Int Pharmacodyn Ther1990;303:232–251.
7. Galligan JJ. Pharmacology of synaptic transmission in the entericnervous system. Curr Opin Pharmacol 2002;2:623–629.
8. Smith TK, Reed JB, Sanders KM. Origin and propagation ofelectrical slow waves in circular muscle of canine proximal colon.Am J Physiol 1987;252:C215–C224.
9. Yoneda S, Takano H, Takaki M, et al. Properties of spontaneouslyactive cells distributed in the submucosal layer of mouse proxi-mal colon. J Physiol 2002;542:887–897.
0. Grider JR. Neurotransmitters mediating the intestinal peristalticreflex in the mouse. J Pharmacol Exp Ther 2003;307:460–467.
1. Kadowaki M, Wade PR, Gershon MD. Participation of 5-HT3,5-HT4, and nicotinic receptors in the peristaltic reflex of guineapig distal colon. Am J Physiol 1996;271:G849–G857.
2. Jin JG, Foxx-Orenstein AE, Grider JR. Propulsion in guinea pigcolon induced by 5-hydroxytryptamine (HT) via 5-HT4 and 5-HT3receptors. J Pharmacol Exp Ther 1999;288:93–97.
3. Smith TK. Myenteric AH neurons are sensory neurons in theguinea-pig proximal colon: an electrophysiological analysis in
intact preparations. Gastroenterology 1994;862:A-216. D4. Bertrand PP, Kunze WA, Furness JB, et al. The terminals ofmyenteric intrinsic primary afferent neurons of the guinea-pigileum are excited by 5-hydroxytryptamine acting at 5-hydroxytryp-tamine-3 receptors. Neuroscience 2000;101:459–469.
5. Gershon MD, Kirchgessner AL. Identification, characterizationand projections of intrinsic primary afferent neurones of thesubmucosal plexus: activity-induced expression of c-fos immuno-reactivity. J Auton Nerv Syst 1991;33:185–187.
6. Smith TK, Spencer NJ, Hennig GW, et al. Recent advances inenteric neurobiology: mechanosensitive interneurons. Neurogas-troenterol Motil 2007;19:869–878.
7. Thomas EA, Sjövall H, Bornstein JC. Computational model of themigrating motor complex of the small intestine. Am J PhysiolGastrointest Liver Physiol 2004;286:G564–G572.
Received October 8, 2008. Accepted December 4, 2008.
eprint requestsAddress requests for reprints to: Department of Physiology &
ell Biology, University of Nevada School of Medicine, Reno,evada. e-mail: [email protected]; fax: (775) 784-903.
cknowledgmentsD.J.H. and E.J.D. contributed equally to this work and were
esponsible for the tension and electrophysiologic recordings,espectively.
onflicts of interestThe authors disclose no conflicts.
undingThis study was funded by a grant from the National Institute of
iabetes and Digestive and Kidney Diseases (RO1 DK45713).
Copyright © 2022 FDOKUMEN




















