
Local RNA Target Structure Influences siRNA Efficacy: A Systematic Global Analysis
11
Local RNA Target Structure Influences siRNA Efficacy: A Systematic Global Analysis Marita Overhoff 1 †, Martina Alken 2 †, Rosel Kretschmer-Kazemi Far 1 Marc Lemaitre 3 , Bernard Lebleu 2 , Georg Sczakiel 1 * and Ian Robbins 2 1 Universita ¨t zu Lu ¨beck, Institut fu ¨r Molekulare Medizin Ratzeburger Allee 160, D-23538 Lu ¨beck, Germany 2 UMR 5124 CNRS, Laboratoire des De ´fenses Antivirales et Antitumorales, Universite ´ Montpellier II, 34095 Montpellier Cedex 5, France 3 EUROGENTEC S.A., Lie `ge Science Park, B-4102 Seraing Belgium The efficiency with which small interfering RNAs (siRNAs) down-regulate specific gene expression in living cells is variable and a number of sequence-governed, biochemical parameters of the siRNA duplex have been proposed for the design of an efficient siRNA. Some of these parameters have been clearly identified to influence the assembly of the RNA-induced silencing complex (RISC), or to favour the sequence preferences of the RISC endonuclease. For other parameters, it is difficult to ascertain whether the influence is a determinant of the siRNA per se, or a determinant of the target RNA, especially its local structural characteristics. In order to gain an insight into the effects of local target structure on the biological activity of siRNA, we have used large sets of siRNAs directed against local targets of the mRNAs of ICAM-1 and survivin. Target structures were classified as accessible or inaccessible using an original, iterative computational approach and by experimental RNase H mapping. The effectiveness of siRNA was characterized by measuring the IC 50 values in cell culture and the maximal extent of target suppression. Mean IC 50 values were tenfold lower for accessible local target sites, with respect to inaccessible ones. Mean maximal target suppression was improved. These data illustrate that local target structure does, indeed, influence the activity of siRNA. We suggest that local target screening can significantly improve the hit rate in the design of biologically active siRNAs. q 2005 Elsevier Ltd. All rights reserved. Keywords: RNA structure; siRNA, design; siRNA, mechanism *Corresponding author Introduction RNA interference is an evolutionarily conserved process in which short regions of double-stranded RNA are produced endogenously from larger precursors and induce the post-transcriptional inhibition of the expression of specific genes. 1,2 The experimental introduction of such small inter- fering RNA (siRNA) into eukaryotic cells is rapidly becoming one of the most important new tools in functional genomics and holds great potential as a therapeutic strategy. 3 The efficiency with which siRNA down-regulates specific gene expression in living cells is highly variable. Indeed, large-scale screens of a number of mRNAs have highlighted extensive positional effects, and some siRNAs show limited effi- ciency. 4–6 This suggests that the efficacy of a specific siRNA species is influenced by intrinsic properties of the siRNA itself, by intrinsic properties of the target system (such as target RNA structure) or by a combination of both. A number of thermodynamic and structural characteristics of the siRNA moiety have been suggested to contribute to the effective- ness of siRNA. 7,8 Recent studies have led to the establishment of a list of eight such parameters that, when incorporated into a rational design algorithm, permit an increase in the probability of selecting an effective siRNA (i.e. one that is capable of down- regulating gene expression O50%. 6 This leads to the suggestion that siRNA could be effective indepen- dently of its target sequence. However, although it is becoming clear that certain of the biochemical characteristics of the siRNA duplex influence the probability of the antisense siRNA strand (asRNAi) being recruited by the RNA-induced silencing complex (RISC), 7,8 it is not yet possible to establish 0022-2836/$ - see front matter q 2005 Elsevier Ltd. All rights reserved. † M.O. & M.A. contributed equally to this work. Abbreviations used: siRNA, small interfering RNA; RISC, RNA-induced silencing complex; asRNAi, antisense siRNA strand; FCS, fetal calf serum. E-mail address of the corresponding author: [email protected] doi:10.1016/j.jmb.2005.03.012 J. Mol. Biol. (2005) 348, 871–881
-
Upload
independent -
Category
Documents
-
view
7 -
download
0
Transcript of Local RNA Target Structure Influences siRNA Efficacy: A Systematic Global Analysis

doi:10.1016/j.jmb.2005.03.012 J. Mol. Biol. (2005) 348, 871–881
Local RNA Target Structure Influences siRNA Efficacy:A Systematic Global Analysis
Marita Overhoff1†, Martina Alken2†, Rosel Kretschmer-Kazemi Far1
Marc Lemaitre3, Bernard Lebleu2, Georg Sczakiel1* and Ian Robbins2
1Universitat zu Lubeck, Institutfur Molekulare MedizinRatzeburger Allee 160, D-23538Lubeck, Germany
2UMR 5124 CNRS, Laboratoiredes Defenses Antivirales etAntitumorales, UniversiteMontpellier II, 34095Montpellier Cedex 5, France
3EUROGENTEC S.A., LiegeScience Park, B-4102 SeraingBelgium
0022-2836/$ - see front matter q 2005 E
†M.O. & M.A. contributed equalAbbreviations used: siRNA, smal
RISC, RNA-induced silencing compantisense siRNA strand; FCS, fetalE-mail address of the correspond
The efficiency with which small interfering RNAs (siRNAs) down-regulatespecific gene expression in living cells is variable and a number ofsequence-governed, biochemical parameters of the siRNA duplex havebeen proposed for the design of an efficient siRNA. Some of theseparameters have been clearly identified to influence the assembly of theRNA-induced silencing complex (RISC), or to favour the sequencepreferences of the RISC endonuclease. For other parameters, it is difficultto ascertain whether the influence is a determinant of the siRNA per se, or adeterminant of the target RNA, especially its local structural characteristics.In order to gain an insight into the effects of local target structure on thebiological activity of siRNA, we have used large sets of siRNAs directedagainst local targets of the mRNAs of ICAM-1 and survivin. Targetstructures were classified as accessible or inaccessible using an original,iterative computational approach and by experimental RNase H mapping.The effectiveness of siRNAwas characterized by measuring the IC50 valuesin cell culture and the maximal extent of target suppression. Mean IC50
values were tenfold lower for accessible local target sites, with respect toinaccessible ones. Mean maximal target suppression was improved. Thesedata illustrate that local target structure does, indeed, influence the activityof siRNA. We suggest that local target screening can significantly improvethe hit rate in the design of biologically active siRNAs.
q 2005 Elsevier Ltd. All rights reserved.
Keywords: RNA structure; siRNA, design; siRNA, mechanism
*Corresponding authorIntroduction
RNA interference is an evolutionarily conservedprocess in which short regions of double-strandedRNA are produced endogenously from largerprecursors and induce the post-transcriptionalinhibition of the expression of specific genes.1,2
The experimental introduction of such small inter-fering RNA (siRNA) into eukaryotic cells is rapidlybecoming one of the most important new tools infunctional genomics and holds great potential as atherapeutic strategy.3
The efficiency with which siRNA down-regulatesspecific gene expression in living cells is highlyvariable. Indeed, large-scale screens of a number of
lsevier Ltd. All rights reserve
ly to this work.l interfering RNA;lex; asRNAi,calf serum.ing author:
mRNAs have highlighted extensive positionaleffects, and some siRNAs show limited effi-ciency.4–6 This suggests that the efficacy of a specificsiRNA species is influenced by intrinsic propertiesof the siRNA itself, by intrinsic properties of thetarget system (such as target RNA structure) or by acombination of both. A number of thermodynamicand structural characteristics of the siRNA moietyhave been suggested to contribute to the effective-ness of siRNA.7,8 Recent studies have led to theestablishment of a list of eight such parameters that,when incorporated into a rational design algorithm,permit an increase in the probability of selecting aneffective siRNA (i.e. one that is capable of down-regulating gene expressionO50%.6 This leads to thesuggestion that siRNA could be effective indepen-dently of its target sequence. However, although itis becoming clear that certain of the biochemicalcharacteristics of the siRNA duplex influence theprobability of the antisense siRNA strand (asRNAi)being recruited by the RNA-induced silencingcomplex (RISC),7,8 it is not yet possible to establish
d.

872 Target Structure and siRNA Efficacy
for others whether the critical parameter affected issiRNA-dependent (i.e. RISC formation), target-dependent (i.e. sequence- or structure-dependent),or asRNAi/target duplex-dependent (i.e. substratefor endonuclease activity).
A number of reports have suggested that theselection of local target sequences is critical,4,5,9 as isthe local structural target accessibility.4,5,9–11 Thecomplex secondary structure of mRNAs, however,is not amenable to accurate modelling,12–15 makingit difficult to correlate siRNA efficacy to targetstructure. By narrowing the definition of secondarystructure to two broad categories, accessible or non-accessible (vis a vis to Watson–Crick binding) andusing an established, iterative computationalanalysis,16,17 relevant structural information can begleaned from primary sequence. This kind oftheoretical approach has been demonstrated togive results that are highly consistent with struc-tural probing of RNA in the presence of cellularextracts,18 suggesting that it represents a validbasis to study the relationship between targetaccessibility and efficacy of siRNA.
In order to better define the role of local targetstructure in siRNA efficacy, we have estimated IC50
values for siRNAs targeted to regions defined, bythis computational analysis, as structurally acces-sible, or inaccessible, for two biologically distanttarget systems ICAM-1 and survivin. In addition, forthe survivin transcript, we have used combinatorialoligonucleotide–RNase H mapping to confirmcomputed accessibility. We demonstrate thatsiRNA targeting putative accessible sites have ahigher probability of exhibiting low IC50 and showgreater maximal inhibition than those targetingputative inaccessible sites. Our data are in line withthe accompanying study by Schubert et al. (accom-panying paper) using intentionally designed targetstructures to study the relationship between localaccessibility and efficacy.31 This suggests that targetstructure does, indeed, influence siRNA activity.
Results
The ICAM-1 and survivin targets: structuralanalysis and selection of local target sites ofmRNA
The expression of ICAM-1 in ECV304 cells andthe expression of survivin in K562 cells representtwo independent biological systems for this study.For both systems, experimental protocols, includingcell culture conditions and transfection protocols,have been optimized. Target sites along the ICAM-1and survivinmRNAs were analysed by a systematiccomputational calculation of local RNA secondarystructures and classified according to their struc-tural accessibility as described.11,16 Briefly, thetarget sequence is dissected into overlappingwindows of a given length (e.g. 800 nt and overlapsof 100 nt) and secondary structures of thosewindows are calculated. Subsequently, we record
stretches of consecutively unpaired nucleotides(R10 nt) and their abundance in all predictions.We regard as accessible the target segments thatshow the longest and most highly conservedconsecutively unpaired sequence segments. Inprinciple, this computational analysis reveals non-paired segments rather than specific folding units.This means that this study does not rely on therelevance of predicted RNA structures but on themeaning of the statistical prediction of sequencesegments of the target RNA that are not involved inintramolecular folding and, thus are regarded asaccessible in this work. This approach was shownearlier to give rise to results that are consistent witha relevant protocol for the experimental probing oftarget RNA in the presence of cellular extracts.18
Structurally accessible and inaccessible local targetsites were defined according to this concept for thefull-length spliced mRNAs of ICAM-1 and survivinas summarized in Figure 1.
Design of siRNA
The analysis of local structural target accessibilityshown in Figure 1 was used to define the sequencesof siRNA such that the antisense strand of theselected siRNAwas complementary to an accessibleor an inaccessible stretch of target sequences.Further, target sites were distributed along theentire target RNA (Figure 1 and Table 1). In additionto local structural characteristics of the target RNAthe design of siRNA sequences in this work wasbased on criteria related to their sequences includ-ing nucleotide sequence homology with humansequences and, with one exception (see below), theexclusion of specific motifs such as G-quartets. AllsiRNA constructs contained a 21 bp double-stranded portion and were tested for nucleotidesequence homology with unrelated human tran-scribed sequences. The controls with scramblednucleotide sequences (see Table 1) were designedsuch that they cover a range of GC content (9–15 nt)and consequent thermal stability.
Efficacy of ICAM-1 and survivin-directed siRNAin transfected cells
Suppression of ICAM-1 gene expression wasmeasured by transient transfection of ECV304cells with siRNA followed by stimulation ofICAM-1 gene expression by IL-1b.11 In K562 cells,survivin-directed siRNAwas transfected by electro-poration because we observed that these cellsshowed an increase of survivin expression with theuse of most transfecting agents (data not shown).Since the action of siRNA occurs post-transcrip-tionally, siRNA-mediated effects were monitored atthe target RNA level by quantitative RT-PCR. Inorder to compare the efficacy of all siRNA, twoparameters were derived, the calculated half-maximal inhibitory concentration (IC50) and themaximal extent of target suppression withinthe tested range. Values were standardized to the

Figure 1. Accessibility of local target sites of ICAM-1 mRNA (top panel) and survivin mRNA (lower panel). Sitespredicted to be accessible by iterative computational analysis are indicated by dark green arrows, those predicted to beinaccessible are indicated by red arrows. Local target sites identified as accessible by RNase Hmapping are indicated bylight green arrows, those that were found to be inaccessible by this technique are indicated by black arrows. Note that thesite 729 of survivingmRNA is subdivided into 729-1 and 729-3 (see also Figure 3). Abbreviations: 5 0-UTR, 5 0 untranslatedregion; cds, coding sequence; 3 0-UTR, 3 0 untranslated region.
Target Structure and siRNA Efficacy 873
IL-1b-induced state in the case of ICAM-1expression (Table 2) and to the expression ofsurvivin in the presence of an unrelated siRNAagainst EGFP which had no effect on survivinexpression in K562 (cf. non-treated cells; Table 3).
In the context of the IC50 values, in the ICAM-1system there is clearly a more efficient targetsuppression by siRNAs directed against theaccessible target sites, with a mean value of 4 nM(Table 2A). The siRNAs targeted to the inaccessiblesites, on the other hand, were tenfold less effective,with a mean IC50 value of 42 nM (Table 2B). Thismajor difference between the two groups can beaccounted for by the greater abundance of lesseffective (or even non-effective) siRNA specieswithin the group directed against inaccessibletargets, although this group includes species thatexert high efficacy, e.g. si640, si650, or si1437(Table 2B). In contrast, all of the siRNAs directedagainst accessible local target sites were effective, asreflected by their low IC50 values (in the 1–10 nMrange). Some species (si862, si1628, and si1637)showed equally strong inhibition between 1 nMand 6 nM, though with a high error range. Thesedata are, therefore, excluded from Table 2.
A similar relationship between the computationalanalysis of target site accessibility and the effective-ness of siRNA directed against survivin wasobserved in K562 cells (Table 3). Although a smallernumber of cases was evaluated, and generallyhigher IC50 values were calculated, the improve-ment of the efficacy when considering the structuralanalysis of local targets was reflected here by an11-fold improvement of the mean value for IC50 forthe accessible target group.
The major difference between the accessible andinaccessible target groups is the proportion ofsiRNA species with a low IC50. All siRNAs
targeting accessible sites of ICAM-1 and survivinhave an IC50%10 nM, whereas 50% of ICAM-1-targeted siRNAs, and 75% of survivin-targetedsiRNAs, in the inaccessible groups had IC50O10 nM. This dichotomy in the efficacy of siRNAtargeting accessible and inaccessible groups isreflected also by the extent of inhibition at themaximal dose tested (100 nM for ICAM-1 and1000 nM for survivin). However, with only oneexception, all of the siRNAs, whether directedagainst accessible or inaccessible local targets,were capable of inducing at least 50% inhibition(Tables 2 and 3). This would suggest that, inprinciple, the majority of siRNA may be active athigher concentrations. It is noteworthy that a plot ofthe IC50 values of ICAM-1-directed siRNA versusthe position of local targets sites along the ICAM-1mRNA indicates a site-related rather than siRNA-related correlation with biological activity for thethree most upstream located targets (Figure 2).Interestingly, a number of siRNAs behaved
irregularly (Table 4). In most cases we observed alack of dose–response relationship. For example,siRNA1141 (ICAM-1) and si729-1 (survivin)showed an inhibition of 50% and 25%, respectively,at all doses tested (up to 1 mM for si729-1; data notshown) and si2304 showed very strong but dose-independent inhibition. In some cases this may bedue to structural constraints inherent to the siRNAmolecule per se. For instance, si729-1 was directedagainst a sequence within the survivin transcript(729–744) that we identified as an obvious andhighly promising locally accessible target site. ThissiRNA contains a consecutive stretch of six GCbase-pairs (with five consecutive G nucleotides inthe sense strand), which may support the formationof higher-order structures. A stimulation of ICAM-1and survivin expression was observed occasionally

Table 1. Nulceotide sequences and target positions of ICAM-1-directed siRNA and control siRNA used in this study
Nomenclature Nucleotide sequence of the sense strand 5 0/3 0 Target positions
5 0 30
ICAM-1si-424 UCUUGGCAGCCAGUGGGCAAG 424 444si-429 GCAGCCAGUGGGCAAGAACCU 429 459si-640 CAAGGGCUGGAGCUGUUUGAG 640 660si-650 AGCUGUUUGAGAACACCUCGG 650 670si-839 CCACAGUCACCUAUGGCAACG 839 869si-840 CACAGUCACCUAUGGCAACGA 840 860si-841 ACAGUCACCUAUGGCAACGAC 841 861si-842 CAGUCACCUAUGGCAACGACU 842 862si-843 AGUCACCUAUGGCAACGACUC 843 863si-859 GACUCCUUCUCGGCCAAGGCC 859 889si-860 ACUCCUUCUCGGCCAAGGCCU 860 880si-861 CUCCUUCUCGGCCAAGGCCUC 861 881si-862 UCCUUCUCGGCCAAGGCCUCA 862 882si-1141 CCAGAGGACAACGGGCGCAGC 1141 1161si-1153 GGGCGCAGCUUCUCCUGCUCU 1153 1173si-1437 GAGCACUCAAGGGGAGGUCAC 1437 1457si-1452 GGUCACCCGCGAGGUGACCGU 1452 1472si-1546 GGCCUCAGCACGUACCUCUAU 1546 1566si-1566 UAACCGCCAGCGGAAGAUCAA 1566 1586si-1591 UACAGACUACAACAGGCCCAA 1591 1611si-1595 GACUACAACAGGCCCAAAAAG 1595 1615si-1628 AACCGAACACACAAGCCACGC 1628 1648si-1637 CACAAGCCACGCCUCCCUGAA 1637 1657si-2289 GUACACUGCAGGAGAGUGCCU 2289 2319si-2304 GUGCCUGGCAAAAAGAUCAAA 2304 2324
survivinsi260-1 CCAUAGAGGAACAUAAAAAGC 267 287si323-1 GUUUGAAGAAUUAACCCUUGG 325 345si729-1 UUUUUGGGGGCUCAUUUUUGC 716 736si729-3 CUGUUUUGAUUCCCGGGCUUA 736 756si745-2 GUGAGAAGUGAGGGAGGAAGA 761 781si858-2 UCAUGUUGUUGAGGCUGUCAC 858 878si1098-3 CACAGAAUAGCACAAACUACA 1103 1123si1133-2 GCCAUUCUAAGUCAUUGGGGA 1142 1162si1530-2 GAGCUGCAGGGUGGAUUGUUA 1533 1553
Controlssi-sc1 CGAACUCACUGGUCUGACCAU – –si-sc2 CGAACUCACUGGUCUGACCGC – –si-sc3 CGGACGCACUGGUCUGACCGG – –si-sc4 AGAACACACAGGUCUGAACAU – –si-sc5 CGAACUCACUGGUCUGAC – –EGFP siRNA CUACAACAGCCACAACGUCTT – –
874 Target Structure and siRNA Efficacy
after cationic lipid delivery of siRNA (unpublisheddata) in a way that seems to include nucleotidesequence-specificity. Therefore, one might speculateon dual influences: a suppressive effect by siRNAmechanisms and a stimulatory effect due tounknown effects that might lead to the loss ofconcentration dependency, perhaps includingeffects by the short, double-stranded siRNA. Ifstimulation of the target dominates over siRNA-mediated inhibition, one might even observeincreased target expression at increasing doses ofsiRNA.
Relationship between accessible sites asdefined by RNase H mapping or bycomputational analysis and efficacy in livingcells
Occasionally we found biologically effectivesiRNA designed against local target sites that havenot been predicted to be accessible by
computational analysis, e.g. si260-1, si745-2, orsi1530-2 (Table 3B). In order to shed light on thepossibility that this is due to misleading results ofthe computational analysis of target RNA or to acomplex structure–function relationship, we per-formed experimental structural probing of theselocal target sites. Combinatorial in vitro screeninghas proved useful for the design of antisenseoligonucleotides20 but, to our knowledge, thisapproach has not been applied to the design ofsiRNA.
The two sites clearly identified as inaccessible bycomputational analysis for the survivin transcript(323 and 858) were clearly inaccessible in themapping analysis (Figures 1 and 3). Three sitespredicted to be accessible (745, 1098, and 1133) weremapped as accessible using the combinatoriallibraries. Two sites shown to have clear accessibilityin the RNase H mapping (260 and 1530), however,did not show up in the computational analysis. Thesites located between nucleotides 715 and 767 of the
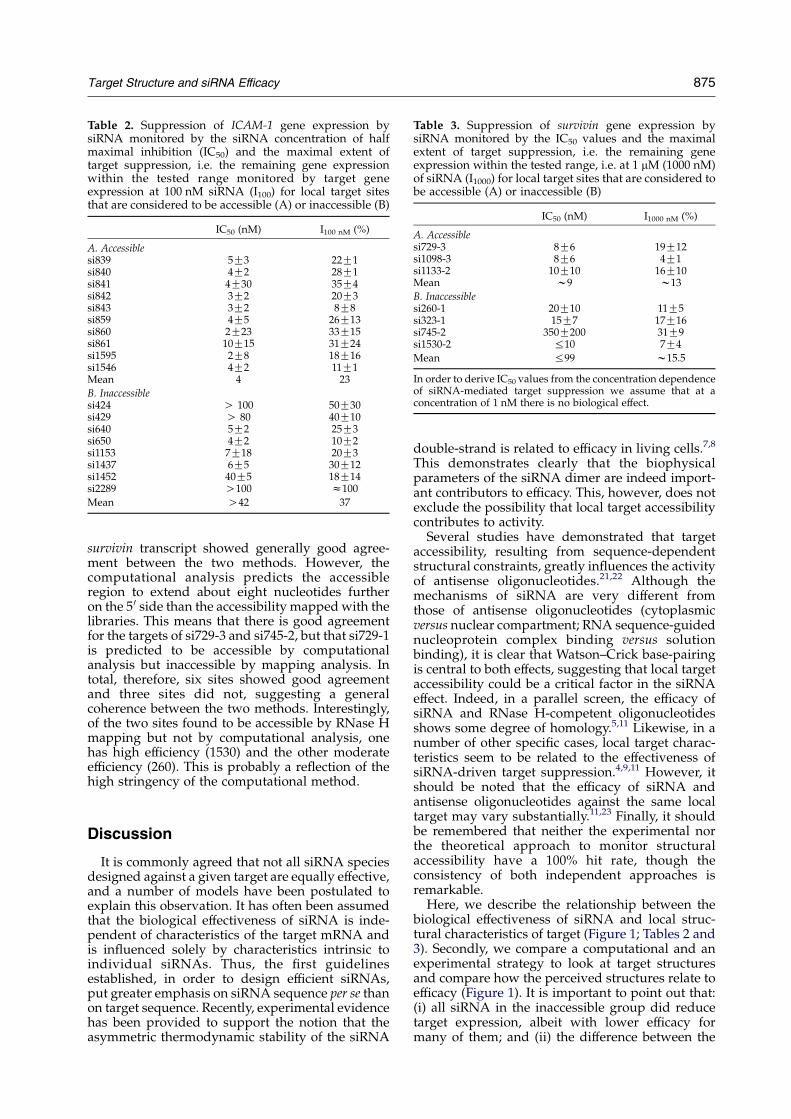
Table 3. Suppression of survivin gene expression bysiRNA monitored by the IC50 values and the maximalextent of target suppression, i.e. the remaining geneexpression within the tested range, i.e. at 1 mM (1000 nM)of siRNA (I1000) for local target sites that are considered tobe accessible (A) or inaccessible (B)
IC50 (nM) I1000 nM (%)
A. Accessiblesi729-3 8G6 19G12si1098-3 8G6 4G1si1133-2 10G10 16G10Mean w9 w13
B. Inaccessiblesi260-1 20G10 11G5si323-1 15G7 17G16si745-2 350G200 31G9si1530-2 %10 7G4
Mean %99 w15.5
In order to derive IC50 values from the concentration dependenceof siRNA-mediated target suppression we assume that at aconcentration of 1 nM there is no biological effect.
Table 2. Suppression of ICAM-1 gene expression bysiRNA monitored by the siRNA concentration of halfmaximal inhibition (IC50) and the maximal extent oftarget suppression, i.e. the remaining gene expressionwithin the tested range monitored by target geneexpression at 100 nM siRNA (I100) for local target sitesthat are considered to be accessible (A) or inaccessible (B)
IC50 (nM) I100 nM (%)
A. Accessiblesi839 5G3 22G1si840 4G2 28G1si841 4G30 35G4si842 3G2 20G3si843 3G2 8G8si859 4G5 26G13si860 2G23 33G15si861 10G15 31G24si1595 2G8 18G16si1546 4G2 11G1Mean 4 23
B. Inaccessiblesi424 O 100 50G30si429 O 80 40G10si640 5G2 25G3si650 4G2 10G2si1153 7G18 20G3si1437 6G5 30G12si1452 40G5 18G14si2289 O100 z100
Mean O42 37
Target Structure and siRNA Efficacy 875
survivin transcript showed generally good agree-ment between the two methods. However, thecomputational analysis predicts the accessibleregion to extend about eight nucleotides furtheron the 5 0 side than the accessibility mappedwith thelibraries. This means that there is good agreementfor the targets of si729-3 and si745-2, but that si729-1is predicted to be accessible by computationalanalysis but inaccessible by mapping analysis. Intotal, therefore, six sites showed good agreementand three sites did not, suggesting a generalcoherence between the two methods. Interestingly,of the two sites found to be accessible by RNase Hmapping but not by computational analysis, onehas high efficiency (1530) and the other moderateefficiency (260). This is probably a reflection of thehigh stringency of the computational method.
Discussion
It is commonly agreed that not all siRNA speciesdesigned against a given target are equally effective,and a number of models have been postulated toexplain this observation. It has often been assumedthat the biological effectiveness of siRNA is inde-pendent of characteristics of the target mRNA andis influenced solely by characteristics intrinsic toindividual siRNAs. Thus, the first guidelinesestablished, in order to design efficient siRNAs,put greater emphasis on siRNA sequence per se thanon target sequence. Recently, experimental evidencehas been provided to support the notion that theasymmetric thermodynamic stability of the siRNA
double-strand is related to efficacy in living cells.7,8
This demonstrates clearly that the biophysicalparameters of the siRNA dimer are indeed import-ant contributors to efficacy. This, however, does notexclude the possibility that local target accessibilitycontributes to activity.Several studies have demonstrated that target
accessibility, resulting from sequence-dependentstructural constraints, greatly influences the activityof antisense oligonucleotides.21,22 Although themechanisms of siRNA are very different fromthose of antisense oligonucleotides (cytoplasmicversus nuclear compartment; RNA sequence-guidednucleoprotein complex binding versus solutionbinding), it is clear that Watson–Crick base-pairingis central to both effects, suggesting that local targetaccessibility could be a critical factor in the siRNAeffect. Indeed, in a parallel screen, the efficacy ofsiRNA and RNase H-competent oligonucleotidesshows some degree of homology.5,11 Likewise, in anumber of other specific cases, local target charac-teristics seem to be related to the effectiveness ofsiRNA-driven target suppression.4,9,11 However, itshould be noted that the efficacy of siRNA andantisense oligonucleotides against the same localtarget may vary substantially.11,23 Finally, it shouldbe remembered that neither the experimental northe theoretical approach to monitor structuralaccessibility have a 100% hit rate, though theconsistency of both independent approaches isremarkable.Here, we describe the relationship between the
biological effectiveness of siRNA and local struc-tural characteristics of target (Figure 1; Tables 2 and3). Secondly, we compare a computational and anexperimental strategy to look at target structuresand compare how the perceived structures relate toefficacy (Figure 1). It is important to point out that:(i) all siRNA in the inaccessible group did reducetarget expression, albeit with lower efficacy formany of them; and (ii) the difference between the
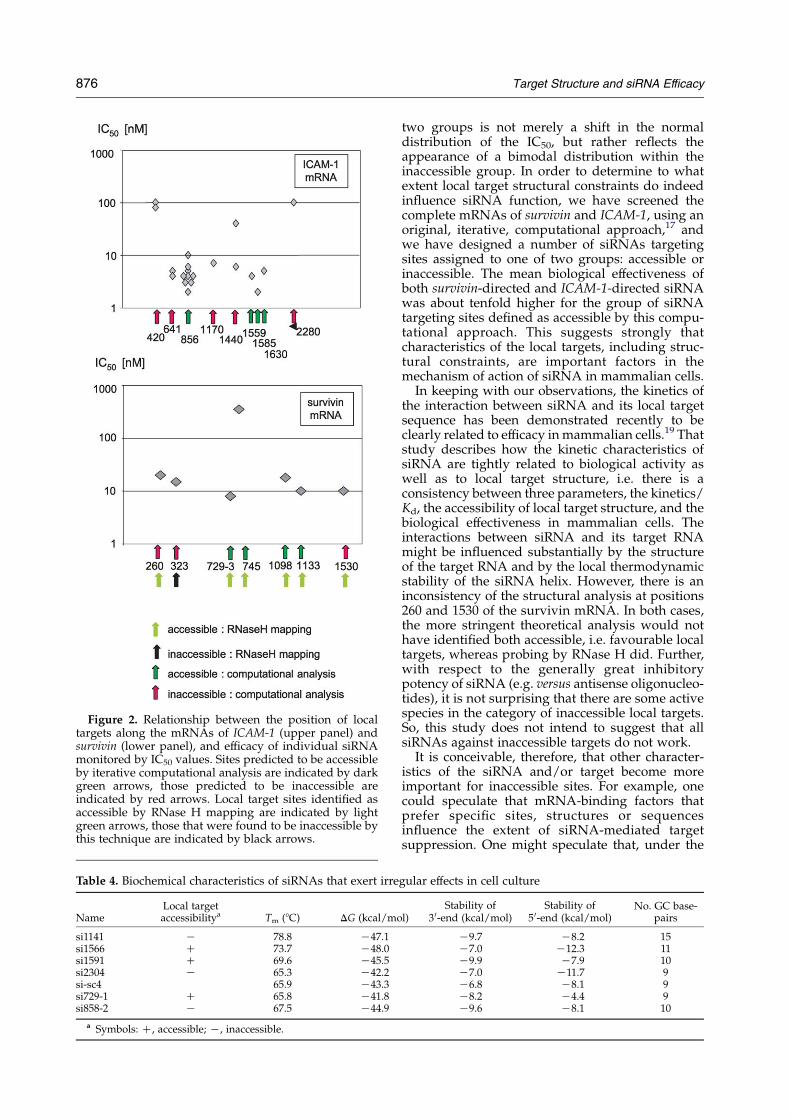
Figure 2. Relationship between the position of localtargets along the mRNAs of ICAM-1 (upper panel) andsurvivin (lower panel), and efficacy of individual siRNAmonitored by IC50 values. Sites predicted to be accessibleby iterative computational analysis are indicated by darkgreen arrows, those predicted to be inaccessible areindicated by red arrows. Local target sites identified asaccessible by RNase H mapping are indicated by lightgreen arrows, those that were found to be inaccessible bythis technique are indicated by black arrows.
Table 4. Biochemical characteristics of siRNAs that exert irre
NameLocal targetaccessibilitya Tm (8C) DG (kcal/mo
si1141 K 78.8 K47.1si1566 C 73.7 K48.0si1591 C 69.6 K45.5si2304 K 65.3 K42.2si-sc4 65.9 K43.3si729-1 C 65.8 K41.8si858-2 K 67.5 K44.9
a Symbols: C, accessible; K, inaccessible.
876 Target Structure and siRNA Efficacy
two groups is not merely a shift in the normaldistribution of the IC50, but rather reflects theappearance of a bimodal distribution within theinaccessible group. In order to determine to whatextent local target structural constraints do indeedinfluence siRNA function, we have screened thecomplete mRNAs of survivin and ICAM-1, using anoriginal, iterative, computational approach,17 andwe have designed a number of siRNAs targetingsites assigned to one of two groups: accessible orinaccessible. The mean biological effectiveness ofboth survivin-directed and ICAM-1-directed siRNAwas about tenfold higher for the group of siRNAtargeting sites defined as accessible by this compu-tational approach. This suggests strongly thatcharacteristics of the local targets, including struc-tural constraints, are important factors in themechanism of action of siRNA in mammalian cells.
In keeping with our observations, the kinetics ofthe interaction between siRNA and its local targetsequence has been demonstrated recently to beclearly related to efficacy inmammalian cells.19 Thatstudy describes how the kinetic characteristics ofsiRNA are tightly related to biological activity aswell as to local target structure, i.e. there is aconsistency between three parameters, the kinetics/Kd, the accessibility of local target structure, and thebiological effectiveness in mammalian cells. Theinteractions between siRNA and its target RNAmight be influenced substantially by the structureof the target RNA and by the local thermodynamicstability of the siRNA helix. However, there is aninconsistency of the structural analysis at positions260 and 1530 of the survivin mRNA. In both cases,the more stringent theoretical analysis would nothave identified both accessible, i.e. favourable localtargets, whereas probing by RNase H did. Further,with respect to the generally great inhibitorypotency of siRNA (e.g. versus antisense oligonucleo-tides), it is not surprising that there are some activespecies in the category of inaccessible local targets.So, this study does not intend to suggest that allsiRNAs against inaccessible targets do not work.
It is conceivable, therefore, that other character-istics of the siRNA and/or target become moreimportant for inaccessible sites. For example, onecould speculate that mRNA-binding factors thatprefer specific sites, structures or sequencesinfluence the extent of siRNA-mediated targetsuppression. One might speculate that, under the
gular effects in cell culture
l)Stability of
3 0-end (kcal/mol)Stability of
5 0-end (kcal/mol)No. GC base-
pairs
K9.7 K8.2 15K7.0 K12.3 11K9.9 K7.9 10K7.0 K11.7 9K6.8 K8.1 9K8.2 K4.4 9K9.6 K8.1 10
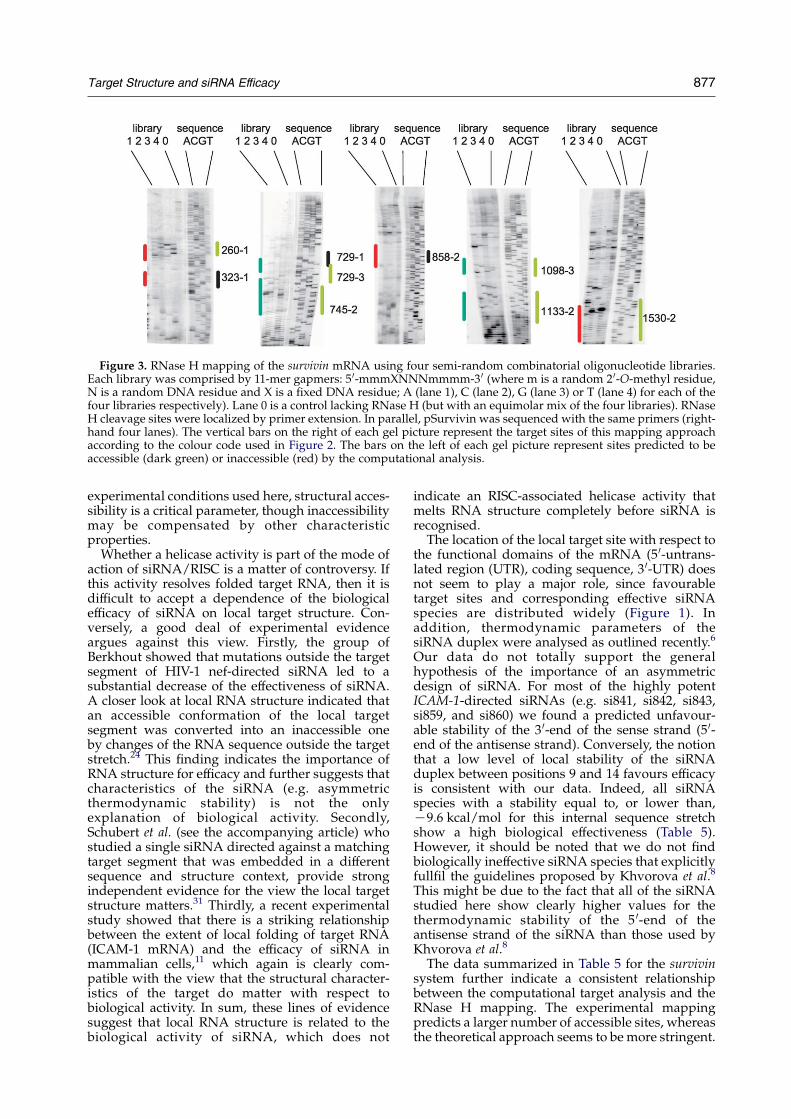
Figure 3. RNase H mapping of the survivin mRNA using four semi-random combinatorial oligonucleotide libraries.Each library was comprised by 11-mer gapmers: 5 0-mmmXNNNmmmm-3 0 (where m is a random 2 0-O-methyl residue,N is a random DNA residue and X is a fixed DNA residue; A (lane 1), C (lane 2), G (lane 3) or T (lane 4) for each of thefour libraries respectively). Lane 0 is a control lacking RNase H (but with an equimolar mix of the four libraries). RNaseH cleavage sites were localized by primer extension. In parallel, pSurvivin was sequenced with the same primers (right-hand four lanes). The vertical bars on the right of each gel picture represent the target sites of this mapping approachaccording to the colour code used in Figure 2. The bars on the left of each gel picture represent sites predicted to beaccessible (dark green) or inaccessible (red) by the computational analysis.
Target Structure and siRNA Efficacy 877
experimental conditions used here, structural acces-sibility is a critical parameter, though inaccessibilitymay be compensated by other characteristicproperties.
Whether a helicase activity is part of the mode ofaction of siRNA/RISC is a matter of controversy. Ifthis activity resolves folded target RNA, then it isdifficult to accept a dependence of the biologicalefficacy of siRNA on local target structure. Con-versely, a good deal of experimental evidenceargues against this view. Firstly, the group ofBerkhout showed that mutations outside the targetsegment of HIV-1 nef-directed siRNA led to asubstantial decrease of the effectiveness of siRNA.A closer look at local RNA structure indicated thatan accessible conformation of the local targetsegment was converted into an inaccessible oneby changes of the RNA sequence outside the targetstretch.24 This finding indicates the importance ofRNA structure for efficacy and further suggests thatcharacteristics of the siRNA (e.g. asymmetricthermodynamic stability) is not the onlyexplanation of biological activity. Secondly,Schubert et al. (see the accompanying article) whostudied a single siRNA directed against a matchingtarget segment that was embedded in a differentsequence and structure context, provide strongindependent evidence for the view the local targetstructure matters.31 Thirdly, a recent experimentalstudy showed that there is a striking relationshipbetween the extent of local folding of target RNA(ICAM-1 mRNA) and the efficacy of siRNA inmammalian cells,11 which again is clearly com-patible with the view that the structural character-istics of the target do matter with respect tobiological activity. In sum, these lines of evidencesuggest that local RNA structure is related to thebiological activity of siRNA, which does not
indicate an RISC-associated helicase activity thatmelts RNA structure completely before siRNA isrecognised.The location of the local target site with respect to
the functional domains of the mRNA (5 0-untrans-lated region (UTR), coding sequence, 3 0-UTR) doesnot seem to play a major role, since favourabletarget sites and corresponding effective siRNAspecies are distributed widely (Figure 1). Inaddition, thermodynamic parameters of thesiRNA duplex were analysed as outlined recently.6
Our data do not totally support the generalhypothesis of the importance of an asymmetricdesign of siRNA. For most of the highly potentICAM-1-directed siRNAs (e.g. si841, si842, si843,si859, and si860) we found a predicted unfavour-able stability of the 3 0-end of the sense strand (5 0-end of the antisense strand). Conversely, the notionthat a low level of local stability of the siRNAduplex between positions 9 and 14 favours efficacyis consistent with our data. Indeed, all siRNAspecies with a stability equal to, or lower than,K9.6 kcal/mol for this internal sequence stretchshow a high biological effectiveness (Table 5).However, it should be noted that we do not findbiologically ineffective siRNA species that explicitlyfullfil the guidelines proposed by Khvorova et al.8
This might be due to the fact that all of the siRNAstudied here show clearly higher values for thethermodynamic stability of the 5 0-end of theantisense strand of the siRNA than those used byKhvorova et al.8
The data summarized in Table 5 for the survivinsystem further indicate a consistent relationshipbetween the computational target analysis and theRNase H mapping. The experimental mappingpredicts a larger number of accessible sites, whereasthe theoretical approach seems to be more stringent.
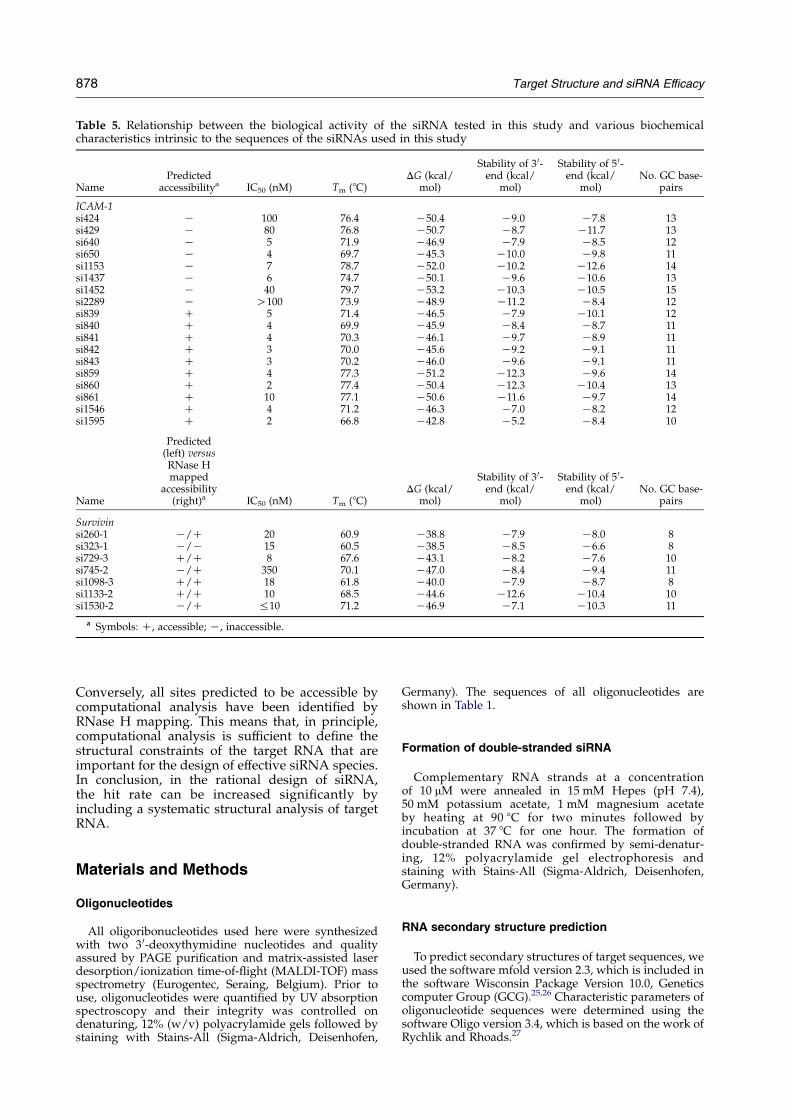
Table 5. Relationship between the biological activity of the siRNA tested in this study and various biochemicalcharacteristics intrinsic to the sequences of the siRNAs used in this study
NamePredicted
accessibilitya IC50 (nM) Tm (8C)DG (kcal/
mol)
Stability of 3 0-end (kcal/
mol)
Stability of 5 0-end (kcal/
mol)No. GC base-
pairs
ICAM-1si424 K 100 76.4 K50.4 K9.0 K7.8 13si429 K 80 76.8 K50.7 K8.7 K11.7 13si640 K 5 71.9 K46.9 K7.9 K8.5 12si650 K 4 69.7 K45.3 K10.0 K9.8 11si1153 K 7 78.7 K52.0 K10.2 K12.6 14si1437 K 6 74.7 K50.1 K9.6 K10.6 13si1452 K 40 79.7 K53.2 K10.3 K10.5 15si2289 K O100 73.9 K48.9 K11.2 K8.4 12si839 C 5 71.4 K46.5 K7.9 K10.1 12si840 C 4 69.9 K45.9 K8.4 K8.7 11si841 C 4 70.3 K46.1 K9.7 K8.9 11si842 C 3 70.0 K45.6 K9.2 K9.1 11si843 C 3 70.2 K46.0 K9.6 K9.1 11si859 C 4 77.3 K51.2 K12.3 K9.6 14si860 C 2 77.4 K50.4 K12.3 K10.4 13si861 C 10 77.1 K50.6 K11.6 K9.7 14si1546 C 4 71.2 K46.3 K7.0 K8.2 12si1595 C 2 66.8 K42.8 K5.2 K8.4 10
Name
Predicted(left) versusRNase Hmapped
accessibility(right)a IC50 (nM) Tm (8C)
DG (kcal/mol)
Stability of 3 0-end (kcal/
mol)
Stability of 5 0-end (kcal/
mol)No. GC base-
pairs
Survivinsi260-1 K/C 20 60.9 K38.8 K7.9 K8.0 8si323-1 K/K 15 60.5 K38.5 K8.5 K6.6 8si729-3 C/C 8 67.6 K43.1 K8.2 K7.6 10si745-2 K/C 350 70.1 K47.0 K8.4 K9.4 11si1098-3 C/C 18 61.8 K40.0 K7.9 K8.7 8si1133-2 C/C 10 68.5 K44.6 K12.6 K10.4 10si1530-2 K/C %10 71.2 K46.9 K7.1 K10.3 11
a Symbols: C, accessible; K, inaccessible.
878 Target Structure and siRNA Efficacy
Conversely, all sites predicted to be accessible bycomputational analysis have been identified byRNase H mapping. This means that, in principle,computational analysis is sufficient to define thestructural constraints of the target RNA that areimportant for the design of effective siRNA species.In conclusion, in the rational design of siRNA,the hit rate can be increased significantly byincluding a systematic structural analysis of targetRNA.
Materials and Methods
Oligonucleotides
All oligoribonucleotides used here were synthesizedwith two 3 0-deoxythymidine nucleotides and qualityassured by PAGE purification and matrix-assisted laserdesorption/ionization time-of-flight (MALDI-TOF) massspectrometry (Eurogentec, Seraing, Belgium). Prior touse, oligonucleotides were quantified by UV absorptionspectroscopy and their integrity was controlled ondenaturing, 12% (w/v) polyacrylamide gels followed bystaining with Stains-All (Sigma-Aldrich, Deisenhofen,
Germany). The sequences of all oligonucleotides areshown in Table 1.
Formation of double-stranded siRNA
Complementary RNA strands at a concentrationof 10 mM were annealed in 15 mM Hepes (pH 7.4),50 mM potassium acetate, 1 mM magnesium acetateby heating at 90 8C for two minutes followed byincubation at 37 8C for one hour. The formation ofdouble-stranded RNA was confirmed by semi-denatur-ing, 12% polyacrylamide gel electrophoresis andstaining with Stains-All (Sigma-Aldrich, Deisenhofen,Germany).
RNA secondary structure prediction
To predict secondary structures of target sequences, weused the software mfold version 2.3, which is included inthe software Wisconsin Package Version 10.0, Geneticscomputer Group (GCG).25,26 Characteristic parameters ofoligonucleotide sequences were determined using thesoftware Oligo version 3.4, which is based on the work ofRychlik and Rhoads.27

Target Structure and siRNA Efficacy 879
RNase H mapping of the survivin transcript
The survivin cDNA was produced from a HeLa celltotal RNA preparation by RT-PCR, using the primers 5 0-TTTGGATCCGCCAGATTTGAATCGCGG-3 0 and 5 0-TTTAAGCTTTTTATTTCTCAGGAAC-3 0, and clonedinto the BamHI/HindIII sites of plasmid pGEM-3Zf(Promega, Mannheim, Germany) to give pSurvivin. Thecloned sequence was verified by sequencing. RNA wastranscribed from pSurvivin using the RiboMAX in vitrotranscription kit (Promega, Mannheim, Germany) andpurified using the RNeasy mini kit (Qiagen, Hilden,Germany).RNase H mapping was performed essentially as
described, using four semi-random nucleotide libraries.28
Each library was comprised by 11-mer gapmers: 5 0-mmmXNNNmmmm-3 0 (where m is a random 2 0-O-methyl residue, N is a random DNA residue and X is afixed DNA residue; A,C,G or T for each of the fourlibraries, respectively). The libraries were synthesizedspecifically by Eurogentec (Seraing, Belgium). In initialexperiments, aimed at optimizing experimental con-ditions, the RNA transcript was 3 0-labelled by enzymaticligation of [32P]pCp (specific activity O3000 Ci/mmol;ICN, Eschwege, Germany) using phage T4 RNA ligase(Amersham, Pharmacia, Biotech). The radio-labelledtranscript (0.3 pmol) was incubated with various concen-trations of each library and Escherichia coli RNase H(Amersham-Pharmacia Biotech, Freiburg, Germany) inmapping buffer (40 mM Tris–HCl (pH 8), 4 mM MgCl2,1 mM DTT) in a final volume of 5 ml for 20 minutes at37 8C. The reaction was stopped by addition of 5 ml of2! sequencing stop buffer, loaded onto a denaturing, 6%(w/v) polyacrylamide sequencing gel, and analyzedusing a PhosphorImager (Molecular Dynamics–Amersham, Freiburg, Germany).Single nucleotide-resolution mapping was performed
as described above, using unlabelled survivin transcriptand optimized concentrations of oligonucleotide libraries(16 mM) and RNase H (0.1 unit) for the reactions with theC and T libraries; 0.2 unit for the reactions with the A andG libraries). RNase H cleavage sites were localized byprimer extension using the AMV Reverse TranscriptasePrimer Extension Kit (Promega, Mannheim, Germany).0.5 pmol of each 20-mer oligonucleotide primer was 5 0-end labelled with 0.25 mCi of [g-32P]ATP (3000 Ci/mmol;ICN, Eschwege, Germany) using one unit of phage T4polynucleotide kinase (Gibco BRL, Eggenstein,Germany). The labelled primer was hybridized to theRNase H-cleaved transcript at 58 8C for 20 minutes andextended according to the manufacturer’s instructions.In parallel, 2 pmol of the radiolabelled primer washybridized to 0.5 pmol of denatured (NaOH) pSurvivinand used for manual sequencing with the T7 Sequenase
v2.0 kit (USB, Cleveland, USA). For each primer, sequenceand mapping reactions were run in parallel on adenaturing, 6% (w/v) polyacrylamide sequencing gel.Data were analysed using a PhosphorImager (MolecularDynamics–Amersham, Freiburg, Germany). Primersused for primer extension in single nucleotide-resolutionmapping experiments were:5 0-GAAGGGCCAGTTCTTGAATG-3 0; 232–251, 50-CCC
AGCCTTCCAGCTCCTTG-3 0; 344–363, 5 0-TCCAGTTTCAAAAATTCACC-3 0; 470–489, 5 0-GGCCAGAGGCCTCAATCCAT-3 0; 608–627, 5 0-AGGAGCACAGTTGAAACATC-3 0; 5 0-TGAGCCCCCAAAAAAGAGAG-3 0; 830–849, 5 0-CACATTCACTGTGGAAGGCT-3 0; 948–967, 5 0-AAACTGTCCTCTGAGGAGGC-3 0; 1045–1064, 5 0-AAACAGTAGAGGAGCCAGGG-3 0; 1179–1188, 5 0-ATTCT
GTCTCCTCATCCACC-3 0; 1305–1324, 5 0-AAAGGATTTAGGCCACTGCC-3 0; 1422–1441, 5 0-GGCGGACTGCGTCTCTCCCC-3 0; 1538–1557, 5 0-GCTGTAACAATCCACCCAGC-3 0; 1590–1609, 5 0-GCTTTTTATTTCTCAGGAAC-3 0.
Cell lines and cell culture
According to the DSMZ-German Collection of Micro-organisms and Cell Cultures (Braunschweig, Germany)the cell-line ECV30429 is a derivative of the humanurinary bladder carcinoma cell line T-24, which expressesICAM-1 in an inducible manner.30 ECV304 cells weremaintained in medium 199 (Sigma-Aldrich, Deisenhofen,Germany) buffered with 25 mM Hepes (pH 7.4) andsupplemented with 0.68 mM L-glutamine and 10% (v/v)fetal calf serum (FCS; Invitrogen, Karlsruhe, Germany).Cells were split routinely two or three times a week aftertrypsinization. For stimulation of ICAM-1 (CD54)200 units/ml of interleukin 1b (IL-1b; PromoCell,Heidelberg, Germany) was added to the medium andcells were incubated overnight (16–18 hours). Thesurvivin-expressing lymphoblast-derived cell line K562was maintained in suspension culture in RPMI-glutamaxmedium (Invitrogen, Karlsruhe, Germany) supplementedwith 10% FCS (Invitrogen, Karlsruhe, Germany). Cellswere passaged every third day (maximum of tenpassages).
Transfection of cells with siRNA
ECV304 cells were seeded in 12-well culture plates at adensity of 1.5!105 cells/well, 15 hours prior to oligo-nucleotide treatment. Then, the cells were washed oncewith prewarmed (37 8C) phosphate-buffered saline (PBS).To deliver siRNA to cells we used Lipofectamine 2000(Invitrogen, Karlsruhe, Germany). The transfection ofsiRNA was performed with 0.4 ml of Opti-MEM Imedium (Invitrogen, Karlsruhe, Germany) containingvarious concentrations of siRNA and 10 mg/ml ofLipofectamine 2000 per well. The cells were incubatedfor four hours at 37 8C under 5% (v/v) CO2. Subsequently,the transfection medium was replaced by medium 199containing 10% FCS. After incubation for four hours at37 8C under 5% CO2, the medium was again substitutedwith medium 199 containing 10% FCS and 200 units/mlof IL-1b to stimulate ICAM-1 expression overnight.K562 cells passaged 24 hours prior to transfection were
recovered by centrifugation, washed in PBS (37 8C) andsuspended in nucleofector (Amaxa: kit V, Koln, Germany)solution (37 8C) at a density of 1!107 cells/ml. Then 2 mlof a 50-fold concentrated siRNA solution was added to100 ml of cell suspension and transfection carried outaccording to the Amaxa electroporation protocol specificfor these cells. Immediately after transfection, 0.5 ml ofpre-warmed complete medium was added to the electro-poration cuvette. The cells in their medium were thentransferred to a well of a six-well culture dish containing1.5 ml of complete medium that had been pre-equili-brated to 37 8C under 5% CO2 for several hours. Cellswere sacrificed 16 hours post transfection.
Quantification of ICAM-1-specific target RNA:quantitative RT-PCR
Total RNA was extracted from transfected cells usingan RNeasy mini kit, including treatment with RNase-freeDNase I (Qiagen, Hilden, Germany). The yield and purityof RNA were determined by spectrophotometry. Syn-thesis of cDNA was carried out using random hexamer

880 Target Structure and siRNA Efficacy
primers and TaqMan reverse transcription reagentsaccording to the manufacturer’s specifications (AppliedBiosystems, Darmstadt, Germany). Quantitative PCRwasperformed using the GeneAmp 5700 sequence detectionsystem (Applied Biosystems, Darmstadt, Germany) andSYBR green PCR core reagents (Eurogentec, Seraing,Belgium). ICAM-1 cDNAwas amplified with the forwardprimer 5 0-GCCACTTCTTCTGTAAGTCTGTGGG-3 0 andreverse primer 5 0-CTACCGGCCCTGGGACG-3 0, result-ing in a fragment of 300 bp. Samples were standardizedusing primers specific to cDNA encoding humanGAPDH(forward primer, 5 0-AACAGCGACACCCACTCCTC-3 0
and reverse primer, 5 0-GGAGGGGAGATTCAGTGTGGT-3 0) resulting in a product of 258 bp. Standard curveswere obtained after amplification of 2.5!10 to 2.5!106
copies of purified plasmid pP5, a derivative of pEGFP-C1carrying the amplicon generated with the ICAM-1 primerset and plasmid pCR-GAPDH, a derivative of pCR 2.1harbouring the GAPDH amplicon sequences.
Quantification of survivin-specific target RNA:quantitative RT-PCR
Total RNA was extracted from transfected cells usingthe RNeasy mini kit, including treatment with RNase-freeDNase I (Qiagen, Hilden, Germany). The yield and purityof RNA were determined by spectrophotometry. cDNAwas synthesized from the survivin-specific primer 5 0-GGAACAGCCGAGATGACCTCC-3 0 and the S26-specificprimer 5 0-TCTCCAGAGAATAGCCTGTCT-3 0 usingSuperscript III reverse transcriptase (Invitrogen,Karlsruhe, Germany) according to the manufacturer’sprotocol. The cDNA products were purified using theQIAquick PCR purification kit (Qiagen, Hilden,Germany) and quantified by real-time PCR using alightcycler (Roche, Mannheim, Germany) and SYBRgreen PCR core reagents (Eurogentec, Seraing, Belgium).The survivin cDNA was amplified with the forwardprimer 5 0-GTCAGCCCAACCTTCACATC-3 0 and reverseprimer 5 0-GAAGCTGTAACAATCCACCCT-3 0, resultingin a fragment of 176 bp. Samples were standardized usingprimers specific to cDNA encoding human S26 (forwardprimer, 5 0-TCTCCGGTCCGTGCCT-3 0 and reverseprimer, 5 0-GCTTCACATACAGCTTGGG-3 0) resulting ina product of 244 bp. Standard curves were obtained afteramplification of 10K6–10K10 dilutions of PCR productsthat had been verified by automated sequencing.
Acknowledgements
We thank the Medical Faculty of the University ofLubeck for financial support within the programFSO to R.K.K.F. and to G.S., and the Associationpour la Recherche sur le Cancer to I.R. and to B.L.
References
1. Hannon, G. J. (2002). RNA interference. Nature, 418,244–251.
2. Mello, C. C. & Conte, D., Jr (2004). Revealing theworld of RNA interference. Nature, 431, 338–342.
3. Dorsett, Y. & Tuschl, T. (2004). siRNAs: applications infunctional genomics and potential as therapeutics.Nature Rev. Drug. Discov. 3, 318–329.
4. Holen, T., Amarzguioui, M., Wiiger, M. T., Babaie, E.& Prydz, H. (2002). Positional effects of shortinterfering RNAs targeting the human coagulationtrigger tissue factor. Nucl. Acids Res. 30, 1757–1766.
5. Vickers, T. A., Koo, S., Bennett, C. T., Crooke, S. T.,Dean, N.M. & Baker, B. F. (2003). Efficient reduction oftarget RNAs by small interfering RNA and RNaseH-dependent antisense agents. A comparativeanalysis. J. Biol. Chem. 278, 7108–7118.
6. Reynolds, A., Leake, D., Boese, Q., Scaringe, S.,Marshall, W. S. & Khvorova, A. (2004). RationalsiRNA design for RNA interference.Nature Biotechnol.22, 326–330.
7. Schwarz, D. S., Hutvagner, G., Du, T., Xu, Z., Aronin,N. & Zamore, P. (2003). Asymmetry in the assembly ofthe RNAi enzyme complex. Cell, 115, 199–208.
8. Khvorova, A., Reynolds, A. & Jayasena, S. D. (2003).Functional siRNAs and miRNAs exhibit strand bias.Cell, 115, 209–216.
9. Bohula, E. A., Salisbury, A. J., Sohail, M., Playford,M. P., Riedemann, J., Southern, E. M. & Macaulay,V. M. (2003). The efficacy of small interfering RNAstargeted to the type 1 IGF receptor is influenced bysecondary structure in the IGF1R transcript. J. Biol.Chem. 278, 15991–15997.
10. Xu, Y., Zhang, H.-Y., Thormeyer, D., Larsson, O., Du,Q., Elmen, J. et al. (2003). Effective small interferingRNAs and phosphorothioate antisense DNAs havedifferent preferences for target sites in the luciferasemRNAs. Biochem. Biophys. Res. Commun. 306, 712–717.
11. Kretschmer-Kazemi Far, R. & Sczakiel, G. (2003). Theactivity of siRNA in mammalian cells is related tostructural target accessibility: a comparison withantisense oligonucleotides. Nucl. Acids Res. 31,4417–4424.
12. Lima, W. F., Monia, B. P., Ecker, D. J. & Freier, S. M.(1992). Implication of RNA structure on antisenseoligonucleotide hybridization kinetics. Biochemistry,31, 12055–12061.
13. Michel, F. & Westhoff, E. (1990). Modelling of thethree-dimensional architecture of group I catalyticintrons based on comparative sequence analysis.J. Mol. Biol. 216, 585–610.
14. Stein, C. A. (2001). The experimental use of antisenseoligonucleotides: a guide for the perplexed. J. Clin.Invest. 108, 641–644.
15. Sohail, M., Akhtar, S. & Southern, E. M. (1999). Thefolding of large RNAs studied by hybridization toarrays of complementary oligonucleotides. RNA, 5,646–655.
16. Patzel, V., Steidl, U., Kronenwett, R., Haas, R. &Sczakiel, G. (1999). A theoretical approach to selecteffective antisense oligonucleotides at high statisticalprobability. Nucl. Acids Res. 27, 4328–4334.
17. Kretschmer-Kazemi Far, R., Nedbal, W. & Sczakiel, G.(2001). Concepts to automate the theoretical design ofeffective antisense oligonucleotides. Bioinformatics, 17,1058–1061.
18. Scherr, M., Rossi, J. J., Sczakiel, G. & Patzel, V. (2000).RNA accessibility prediction: a theoretical approachis consistent with experimental studies in cell extracts.Nucl. Acids Res. 28, 2455–2461.
19. Wunsche, W. & Sczakiel, G. (2005). The biologicalactivity of siRNA is related to the kinetics of siRNA-target RNA recognition in vitro: mechanistic impli-cations. J. Mol. Biol., 345 203–209.
20. Matveeva, O., Felden, B., Tsodikov, A., Johnston, J.,

Target Structure and siRNA Efficacy 881
Monia, B. P., Atkins, J. F. et al. (1998). Prediction ofantisense oligonucleotide efficacy by in vitromethods. Nature Biotechnol. 16, 1374–1375.
21. Kurreck, J. (2003). Antisense technologies. Improve-ment through novel chemical modifications. Eur.J. Biochem. 270, 1628–1644.
22. Sczakiel, G. (2002). Theoretical and experimentalapproaches to design effective antisense oligonucleo-tides. Front. Biosci., 5, 194–201.
23. Grunweller, A., Wyszko, E., Bieber, B., Jahnel, R.,Erdmann, V. A. & Kurreck, J. (2003). Comparison ofdifferent antisense strategies in mammalian cellsusing locked nucleic acids, 2 0-O-methyl RNA, phos-phorothioates and small interfering RNA. Nucl. AcidsRes. 31, 3185–3193.
24. Westerhout, E. M., Ooms, M., Vink, M., Das, A. T. &Berkhout, B. (2005). HIV-1 can escape from RNAinterference by evolving an alternative structure in itsRNA genome. Nucl. Acids Res. 33, 796–804.
25. Zuker, M. (1989). On finding all suboptimal foldingsof an RNA molecule. Science, 244, 48–52.
26. Mathews, D. H., Sabina, J., Zuker, M. & Turner, D. H.(1999). Expanded sequence dependence of thermo-dynamic parameters improves prediction of RNAsecondary structure. J. Mol. Biol. 288, 911–940.
27. Rychlik, W. & Rhoads, R. E. (1989). A computerprogram for choosing optimal oligonucleotides forfilter hybridization, sequencing and in vitro amplifi-cation of DNA. Nucl. Acids Res. 21, 8543–8551.
28. Ho, S. P., Bao, Y., Lesher, T., Malhotra, R., Ma, L. Y.,Fluharty, S. J. & Sakai, R. R. (1998). Mapping ofRNA accessible sites for antisense experimentswith oligonucleotide libraries. Nature Biotechnol. 16,59–63.
29. Takahashi, K., Sawasaki, Y., Hata, J., Mukai, K. &Goto, T. (1990). Spontaneous transformation andimmortalization of human endothelial cells. In VitroCell Dev. Biol. Anim. 26, 265–274.
30. Dirks, W. G., MacLeod, R. A. & Drexler, H. G. (1999).ECV304 (endothelial) is really T24 (bladdercarcinoma): cell line cross-contamination at source.In Vitro Cell Dev. Biol. 35, 558–559.
31. Schubert, S., Grunweller, A., Erdmann, V. A. &Kurreck, J. (2005). Local RNA target structureinfluences SiRNA efficacy: systematic analysis ofintentionally designed binding regions. J. Mol. Biol.(this issue).
Edited by J. Karn
(Received 9 December 2004; received in revised form 2 March 2005; accepted 2 March 2005)




















