
Light-regulated large-scale reorganization of chromatin during the floral transition in Arabidopsis
10
Light-regulated large-scale reorganization of chromatin during the floral transition in Arabidopsis Federico Tessadori, Roeland Kees Schulkes, Roel van Driel and Paul Fransz* Nuclear Organization Group, Swammerdam Institute for Life Sciences, BioCentrum Amsterdam, University of Amsterdam, Kruislaan 318, 1098 SM, Amsterdam, The Netherlands Received 8 November 2006; revised 30 January 2007; accepted 9 February 2007. *For correspondence (fax + 31205257935; e-mail [email protected]). Summary The floral transition marks the switch from vegetative to reproductive growth, and is controlled by different pathways responsive to endogenous and exogenous cues. The developmental switch is accompanied by local changes in chromatin such as histone modifications. In this study we demonstrate large-scale reorganization of chromatin in rosette leaves during the floral transition. An extensive reduction in chromocenters prior to bolting is followed by a recovery of the heterochromatin domains after elongation of the floral stem. The transient reduction in chromocenters is a result of relocation away from chromocenters of methylated DNA sequences, 5S rDNA and interspersed pericentromeric repeats, but not of 45S rDNA or the 180-bp centromere tandem repeats. Moreover, fluorescence in situ hybridization analysis revealed decondensation of chromatin in gene-rich regions. A mutant analysis indicated that the blue-light photoreceptor CRYPTOCHROME 2 is involved in triggering chromatin decondensation, suggesting a light-signaling pathway towards large-scale chromatin modulation. Keywords: Arabidopsis, floral transition, nucleus, heterochromatin. Introduction Floral transition is a fundamental stage in plant develop- ment, during which the shoot apical meristem switches from generating leaf primordia to production of reproductive organs. A pivotal factor for successful plant reproduction is the timing of the floral transition, as favorable environ- mental conditions are essential for fertilization and optimal seed production. Plants have developed various strategies to comply with this aim. For example, although facultative long-day plants, such as Arabidopsis, are induced to flower by a rise in temperature and increasing day length, flowering in maize and rice is promoted by shorter days. In the dicot- yledonous short-day plant Pharbitis nil, even a single exposure to a 14-h night is floral inductive (Liu et al., 2001; Vince-Prue and Gressel, 1985). A number of different pathways have been described in Arabidopsis that induce the floral transition (Boss et al., 2004; He and Amasino, 2005). Two of these, the vernalization and photoperiod pathways, are dependent on environmen- tal factors and promote flowering after a prolonged cold period and a long-day regime, respectively. In contrast, intrinsic control mechanisms, such as the gibberellin path- way and the autonomous pathway, are largely dependent on endogenous developmental signals. The external and endogenous cues converge into a complex interaction between flower-inhibiting and -promoting signals, the bal- ance of which determines the timing of the floral transition. Flower-inducing signals are integrated at the downstream end of the flower-promoting pathways by the MADS-box protein SUPPRESSOR OF OVEREXPRESSION OF CON- STANS 1 (SOC1) and FLOWERING LOCUS T (FT), of which the biochemical function is unknown. These proteins control the expression of floral meristem identity genes such as LEAFY (LFY) and APETALA 1 (AP1; reviewed in Boss et al., 2004; Putterill et al., 2004). SOC1 and FT themselves are under direct control of CONSTANS (CO), a zinc-finger transcription factor, and FLOWERING LOCUS C (FLC), a MADS-box transcription factor. Flowering time is also controlled by the photoreceptors PHYTOCHROME A (PHYA) and CRYPTOCHROME 2 (CRY2) through the photoperiod pathway, by positively regulating CO activity (Valverde et al., 2004; Yanovsky and Kay, 2002). Under long-day light conditions CO exerts a positive control on floral meristem 848 ª 2007 The Authors Journal compilation ª 2007 Blackwell Publishing Ltd The Plant Journal (2007) 50, 848–857 doi: 10.1111/j.1365-313X.2007.03093.x
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Light-regulated large-scale reorganization of chromatin during the floral transition in Arabidopsis

Light-regulated large-scale reorganization of chromatinduring the floral transition in Arabidopsis
Federico Tessadori, Roeland Kees Schulkes, Roel van Driel and Paul Fransz*
Nuclear Organization Group, Swammerdam Institute for Life Sciences, BioCentrum Amsterdam, University of Amsterdam,
Kruislaan 318, 1098 SM, Amsterdam, The Netherlands
Received 8 November 2006; revised 30 January 2007; accepted 9 February 2007.
*For correspondence (fax + 31205257935; e-mail [email protected]).
Summary
The floral transition marks the switch from vegetative to reproductive growth, and is controlled by different
pathways responsive to endogenous and exogenous cues. The developmental switch is accompanied by local
changes in chromatin such as histone modifications. In this study we demonstrate large-scale reorganization
of chromatin in rosette leaves during the floral transition. An extensive reduction in chromocenters prior to
bolting is followed by a recovery of the heterochromatin domains after elongation of the floral stem. The
transient reduction in chromocenters is a result of relocation away from chromocenters of methylated DNA
sequences, 5S rDNA and interspersed pericentromeric repeats, but not of 45S rDNA or the 180-bp centromere
tandem repeats. Moreover, fluorescence in situ hybridization analysis revealed decondensation of chromatin
in gene-rich regions. A mutant analysis indicated that the blue-light photoreceptor CRYPTOCHROME 2 is
involved in triggering chromatin decondensation, suggesting a light-signaling pathway towards large-scale
chromatin modulation.
Keywords: Arabidopsis, floral transition, nucleus, heterochromatin.
Introduction
Floral transition is a fundamental stage in plant develop-
ment, during which the shoot apical meristem switches from
generating leaf primordia to production of reproductive
organs. A pivotal factor for successful plant reproduction is
the timing of the floral transition, as favorable environ-
mental conditions are essential for fertilization and optimal
seed production. Plants have developed various strategies
to comply with this aim. For example, although facultative
long-day plants, such as Arabidopsis, are induced to flower
by a rise in temperature and increasing day length, flowering
in maize and rice is promoted by shorter days. In the dicot-
yledonous short-day plant Pharbitis nil, even a single
exposure to a 14-h night is floral inductive (Liu et al., 2001;
Vince-Prue and Gressel, 1985).
A number of different pathways have been described in
Arabidopsis that induce the floral transition (Boss et al.,
2004; He and Amasino, 2005). Two of these, the vernalization
and photoperiod pathways, are dependent on environmen-
tal factors and promote flowering after a prolonged cold
period and a long-day regime, respectively. In contrast,
intrinsic control mechanisms, such as the gibberellin path-
way and the autonomous pathway, are largely dependent on
endogenous developmental signals. The external and
endogenous cues converge into a complex interaction
between flower-inhibiting and -promoting signals, the bal-
ance of which determines the timing of the floral transition.
Flower-inducing signals are integrated at the downstream
end of the flower-promoting pathways by the MADS-box
protein SUPPRESSOR OF OVEREXPRESSION OF CON-
STANS 1 (SOC1) and FLOWERING LOCUS T (FT), of which
the biochemical function is unknown. These proteins control
the expression of floral meristem identity genes such as
LEAFY (LFY) and APETALA 1 (AP1; reviewed in Boss et al.,
2004; Putterill et al., 2004). SOC1 and FT themselves are
under direct control of CONSTANS (CO), a zinc-finger
transcription factor, and FLOWERING LOCUS C (FLC), a
MADS-box transcription factor. Flowering time is also
controlled by the photoreceptors PHYTOCHROME A (PHYA)
and CRYPTOCHROME 2 (CRY2) through the photoperiod
pathway, by positively regulating CO activity (Valverde
et al., 2004; Yanovsky and Kay, 2002). Under long-day light
conditions CO exerts a positive control on floral meristem
848 ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd
The Plant Journal (2007) 50, 848–857 doi: 10.1111/j.1365-313X.2007.03093.x

identity genes, among others through the production of a
long-distance systemic florigen signal in the leaves (An
et al., 2004; Huang et al., 2005; Martinez-Garcia et al., 2002;
Zeevaart, 1976). It was recently demonstrated that the FT
gene product is an important component of this signal
(Huang et al., 2005). In contrast to CO, FLC has an inhibiting
action on the expression of the SOC1 and FT genes
(Hepworth et al., 2002). FLC is under the control of several
regulators, including the FRIGIDA gene (FRI), which upreg-
ulates FLC expression (Johanson et al., 2000; Sheldon et al.,
1999). The active FLC locus is associated with transcriptional
activation hallmarks, such as methylation at lysine 4 of
histone 3 and acetylation of histones (He et al., 2003, 2004).
FLC repression is achieved by dimethylation of histone H3 at
lysine 9 (H3 K9me2), mediated by VERNALIZATION INSEN-
SITIVE 1 (VIN1), VERNALIZATION 1 (VRN1) and a Polycomb-
like protein VERNALIZATION 2 (VRN2; Bastow et al., 2004;
Gendall et al., 2001; Levy et al., 2002; Sung and Amasino,
2004). Another protein that epigenetically regulates flower-
ing time is the Arabidopsis HP1 homologue LIKE HETERO-
CHROMATIN PROTEIN 1 (LHP1), also called TERMINAL
FLOWER 2 (TFL2) (Gaudin et al., 2001; Kotake et al., 2003;
Larsson et al., 1998). LHP1 may participate in repression of
the FT gene by counteracting CO (Takada and Goto, 2003).
Consequently, lhp1 mutants are early flowering. The mutant
appears to be impaired in FLC repression, possibly through
interaction with VRN1 (Mylne et al., 2006). VRN1 binds to
chromatin, but the binding site is not restricted to the FLC
locus. Similar to LHP1, the VRN1 protein decorates euchro-
matin regions. Whether the function of VRN1 is part of a
machinery that affects more genes apart from FLC, like LHP1,
(Gaudin et al., 2001; Kotake et al., 2003; Libault et al., 2005) is
not known.
Floral transition is a major developmental switch that
involves physiological processes in the apical meristem, but
also in the rosette leaves, which sense the changing
environmental conditions such as the photoperiod. The
accumulation of light signals triggers a response in the
leaves that is transduced to the shoot apical meristem.
Considering that several components of the light-signa-
ling pathway operate in the nucleus, we addressed the
question of whether the floral transition affects nuclear
processes such as chromatin compaction, involving large
regions of the Arabidopsis genome. In the Arabidopsis
nucleus, the pericentric heterochromatin and the inactive
ribosomal genes are generally clustered into compact
chromatin domains, so-called chromocenters (Fransz et al.,
2002), whereas gene-rich regions, including active and
inactive genes, are largely outside the chromocenters
(Fransz et al., 2006). Unless stated otherwise, in this study
we use the term heterochromatin to indicate compact
chromatin or chromocenters. Previous studies have repor-
ted changes in the appearance of chromocenters under
certain physiological, developmental conditions. For exam-
ple, changes in heterochromatin organization have been
observed during seedling differentiation (Mathieu et al.,
2003), ageing of rosette leaves (Tessadori et al., 2004), de-
differentiation to protoplasts (Tessadori et al., 2007) and
pathogen infection (Pavet et al., 2006). In this study we
demonstrate a transient large-scale reduction of compact
chromatin over a short period (2–4 days) that corresponds
specifically with the floral transition. Mutant analysis of
photoperiod pathway components revealed no typical
reduction in compact chromatin in plants lacking the blue-
light photoreceptor CRY2, suggesting that CRY2 mediates
light-signaling to chromatin organization during the floral
transition.
Results
Mesophyll cells of rosette leaves display large-scale reor-
ganization of heterochromatin domains prior to bolting
The floral transition is marked by changes in the expression
of hundreds of genes (Schmid et al., 2003). We addressed the
question of whether this developmental switch involves
major changes in large-scale chromatin organization.
Parenchyma nuclei from inflorescences display 6–10 con-
spicuous chromocenters (Fransz et al., 2002). We observed a
similar phenotype in nuclei from rosette leaves (Figure 1a).
Up to 20% of the nuclei, however, displayed a phenotype
with smaller or elongated chromocenters and more hetero-
geneous euchromatin (Figure 1b). Remarkably, the fraction
of nuclei with this aberrant phenotype increased to 50%
when visible flower buds appeared at the center of the ro-
sette, which corresponds to stage 5.10 (Boyes et al., 2001).
We therefore carried out a quantitative analysis of
heterochromatin at different developmental stages around
the floral transition. We measured the mean relative hetero-
chromatin fraction (RHF), which is determined by the area
and fluorescence intensity of all chromocenters in relation to
the area and fluorescence intensity of the entire nucleus
(Soppe et al., 2002; Tessadori et al., 2004). Four days before
the appearance of the flower buds (stage 1.09) the RHF
showed a sharp reduction in compact chromatin content
(Figure 1c). We then established the percentage of nuclei
with normal chromocenter appearance (i.e. number, shape
and size), here denoted as the heterochromatin index (HX).
The HX decreased from 70% to about 50% during the same
period before bolting (Figure 1d). Apparently, the appear-
ance of flower buds is accompanied by a large-scale
decondensation of chromocenters. Moreover, within
3 days after bolting the RHF and the HX increased to normal
values, indicating that the reduction of heterochromatin is
transient.
We addressed the question if the change in nuclear
organization is dependent on the flowering time, and
examined two accessions that differ from Col-0 in flowering
Chromatin reorganization during floral transition 849
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 50, 848–857

time. Ler flowers earlier (day 17), whereas Cvi-0 flowers
more than 30 days after germination. We monitored the RHF
value of Col-0, Ler and Cvi-0 plants before and during the
floral transition (Figure 2). The results indicate that the
decrease in chromocenters corresponds with the transition
to flowering irrespective of the flowering time. Strikingly,
although Cvi-0 constitutively shows a lower content of
condensed chromatin compared to Col-0 or Ler, it still
Figure 1. Morphological changes of mesophyll
nuclei and reduction in compact chromatin dur-
ing the floral transition.
(a) and (b) 4¢,6-Diamidino-2-phenylindole stained
nuclei before the floral transition (a) and 2-4 days
prior to bolting (b). Note the difference in chro-
mocenter morphology. Scale bar = 5 lm.
(c) and (d) Quantification of compact chromatin
content by means of relative heterochromatin
fraction (c) and heterochromatin index (d). Bars
represent stages before (white) or after (gray)
bolting. For each bar the data are mean � SEM
of more than 30 nuclei (c) or more than 68 nuclei
(d).
850 Federico Tessadori et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 50, 848–857

displays a decrease in heterochromatin during flower trans-
ition (Figure 2).
To investigate if heterochromatin decondensation is cor-
related with the transition to flowering, we delayed the floral
transition in Col-0 plants by growing them under a shorter
day regime (SD, 12-h light/12-h dark). These plants bolted
19 days later than plants grown under long-day conditions
(LD, 16-h light/ 8-h dark).
In leaves of SD-grown plants the RHF value decreased
after 38 days, which is just before bolting, and recovered
after the appearance of the first flower buds (stage 5.10;
Figure 3a). The simultaneous delay in both bolting and
heterochromatin reduction further indicates a strong corre-
lation between heterochromatin reorganization and floral
transition.
We finally tested if chromatin reorganization can be
promoted by transferring plants from a short-day to a
long-day light regime. Such a transfer will induce the
transition from a vegetative state to a reproductive state.
Col-0 plants were grown under short-day conditions for
37 days and subsequently transferred to a long-day regime.
Only 24 h after the transfer the mesophyll nuclei showed a
sharp decrease in RHF value (Figure 3b). By 4 days after the
transfer the plants produced visible flower buds and the
mesophyll nuclei displayed a normal appearance. These
observations confirm that the decrease in heterochromatin
coincides with the period during which bolting is triggered.
Both pericentromeric heterochromatin and gene-rich chro-
matin show large-scale decondensation during the floral
transition
The reduction of chromocenters indicates that certain
chromosomal regions are no longer in heterochromatin
domains. To determine which DNA sequences are involved
in the reduction of heterochromatin, we applied fluores-
cence in situ hybridization (FISH) with probes of dispersed
and tandemly arranged repeat sequences, known to localize
in chromocenters (Fransz et al., 2002): (i) the 180-bp cen-
tromeric tandem repeat, (ii) the pericentromeric BAC F28D6,
which contains numerous interspersed repeats, such as
transposable elements present on all chromosomes, and (iii)
tandemly arranged 5S and 45S rRNA sequences. All repeats
are condensed and reside in chromocenters of mesophyll
nuclei before the floral transition. During the floral transition
the 180-bp tandem repeat and 45S rDNA sequences re-
mained condensed in chromocenters (Figure 4a). In con-
trast, pericentromeric sequences, targeted by a 5S rDNA
probe and BAC F28D6, were decondensed and dispersed.
This decondensation is likely to be responsible for the
reduction in heterochromatin and the decreased RHF.
To find out whether euchromatic regions are also affected
during the floral transition, we monitored the compaction
state of an approximately 0.3-Mb segment in gene-rich chro-
matin. We applied FISH with the BAC contig F28A21, F13C5
and T18B16, which map to the long arm of chromosome 4,
Figure 2. The reduction of heterochromatin in three accessions.
The relative heterochromatin fraction was measured in Col-0, Ler and Cvi-0
plants grown under long-day conditions before (white bars) and during the
floral transition (gray bars). For each bar the data are mean � SEM of more
than 15 nuclei.
Figure 3. Heterochromatin fraction under short-day (SD) conditions.
(a) Relative heterochromatin fraction (RHF) of plants grown in SD conditions
(12-h light/12-h dark). FS: plants have developed a floral stem, but no flower is
yet open.
(b) RHF of plants grown in SD conditions for 37 days and then transferred to
long-day (LD) conditions. Note the decrease in RHF only 24 h after transfer
and the bolting after 96 h. Bars represent stages before (white) or after (gray)
bolting. For each bar the data are mean � SEM of more than 30 nuclei.
Chromatin reorganization during floral transition 851
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 50, 848–857

Figure 4. Cytological localization of repetitive sequences, gene-rich regions and DNA methylation.
(a) Fluorescence in situ hybridization (FISH) localization of repetitive sequences (right) and 4¢,6-diamidino-2-phenylindole (DAPI) counterstaining (left) in mesophyll
nuclei during the floral transition. Note that although the pericentromeric repeat-rich F28D6 BAC (I, green) and 5S rDNA (II, green) are dispersed in nuclei during
floral transition, the centromeric 180-bp (I, II and III, red) and the subtelomeric 45S rDNA (III, green) regions remain compact and colocalize with chromocenters.
(b) FISH localization of the F28A21-F13C5-T18B16 BAC contig, covering approximately 0.3 Mb. F13C5 appears in red; T18B16 and F28A21 appear in green. The high
compaction state prior to floral transition (I; arrows) is lost in nuclei during the 2–4-day period before bolting (II; arrows). Nuclei are counterstained with DAPI.
(c) Fraction of nuclei with normal levels of heterochromatin (gray), and condensed FISH signals of BAC F13C5 (red) and T18B16-F28A21 (green) before and during
the floral transition. Data are mean � SEM of 29 nuclei.
(d) Immunolocalization of 5-methylcytosine (5-mC, right) counterstained with DAPI (left). (I) Prior to floral transition 5-mC signals are clustered at chromocenters. (II)
A dispersed pattern of 5-mC is observed in nuclei with low chromocenter content during floral transition. Scale bars = 5 lm.
852 Federico Tessadori et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 50, 848–857
and measured the frequency of relatively compact FISH
signals before and during the floral transition (Figure 4b).
Concomitant to the decrease in chromocenters, the occur-
rence of condensed BAC signals decreased by about 33%
(Figure 4c), indicating that the decompaction of chromatin is
not restricted to the heterochromatic sections of the genome.
As chromocenters are rich in methylated DNA (Fransz et al.,
2002) and because a 5-methyl-cytosine (5-mC) imprint is
required to maintain a chromocenter state (Soppe et al.,
2002), we examined leaf nuclei for changes in nuclear
distribution of 5-mC. Col-0 nuclei displayed a dispersed
pattern of 5-mC during the floral transition (Figure 4d),
similar to the dispersion of the 5S rDNA and the transpo-
son-rich BAC F28D6. This suggests that heavily methylated
DNA sequences are responsible for the decrease of the
chromocenters. Furthermore, we found no reduction in DNA
methylation at centromeric repeats, indicating that no large-
scale decrease of DNA methylation has occurred during the
floral transition (Figure S1).
CO and FT do not affect heterochromatin reorganization,
whereas the photoreceptor CRY2 does
As the decrease in heterochromatin coincides with the floral
transition and its timing is affected by the day length, we
inferred that the reorganization of heterochromatin is under
control of components of the photoperiod pathway. We
therefore examined the RHF in mutants of CO, FT and CRY2.
In the photoperiod pathway, the upregulation of CO in
rosette leaves stimulates the production of an FT transcript
that moves through the phloem to the shoot apex triggering
flower development (Abe et al., 2005; Wigge et al., 2005).
Both co-2 and ft-1 mutants are in the Ler background and
have a late-flowering phenotype in long-day conditions
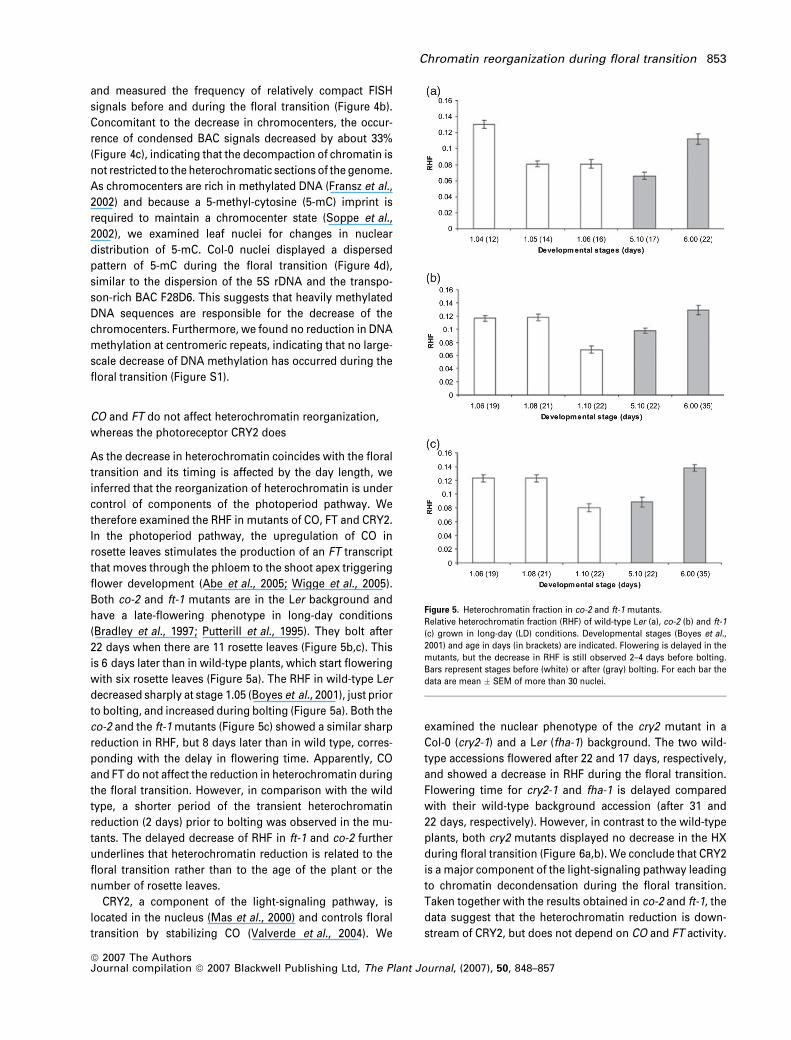
(Bradley et al., 1997; Putterill et al., 1995). They bolt after
22 days when there are 11 rosette leaves (Figure 5b,c). This
is 6 days later than in wild-type plants, which start flowering
with six rosette leaves (Figure 5a). The RHF in wild-type Ler
decreased sharply at stage 1.05 (Boyes et al., 2001), just prior
to bolting, and increased during bolting (Figure 5a). Both the
co-2 and the ft-1 mutants (Figure 5c) showed a similar sharp
reduction in RHF, but 8 days later than in wild type, corres-
ponding with the delay in flowering time. Apparently, CO
and FT do not affect the reduction in heterochromatin during
the floral transition. However, in comparison with the wild
type, a shorter period of the transient heterochromatin
reduction (2 days) prior to bolting was observed in the mu-
tants. The delayed decrease of RHF in ft-1 and co-2 further
underlines that heterochromatin reduction is related to the
floral transition rather than to the age of the plant or the
number of rosette leaves.
CRY2, a component of the light-signaling pathway, is
located in the nucleus (Mas et al., 2000) and controls floral
transition by stabilizing CO (Valverde et al., 2004). We
examined the nuclear phenotype of the cry2 mutant in a
Col-0 (cry2-1) and a Ler (fha-1) background. The two wild-
type accessions flowered after 22 and 17 days, respectively,
and showed a decrease in RHF during the floral transition.
Flowering time for cry2-1 and fha-1 is delayed compared
with their wild-type background accession (after 31 and
22 days, respectively). However, in contrast to the wild-type
plants, both cry2 mutants displayed no decrease in the HX
during floral transition (Figure 6a,b). We conclude that CRY2
is a major component of the light-signaling pathway leading
to chromatin decondensation during the floral transition.
Taken together with the results obtained in co-2 and ft-1, the
data suggest that the heterochromatin reduction is down-
stream of CRY2, but does not depend on CO and FT activity.
Figure 5. Heterochromatin fraction in co-2 and ft-1 mutants.
Relative heterochromatin fraction (RHF) of wild-type Ler (a), co-2 (b) and ft-1
(c) grown in long-day (LD) conditions. Developmental stages (Boyes et al.,
2001) and age in days (in brackets) are indicated. Flowering is delayed in the
mutants, but the decrease in RHF is still observed 2–4 days before bolting.
Bars represent stages before (white) or after (gray) bolting. For each bar the
data are mean � SEM of more than 30 nuclei.
Chromatin reorganization during floral transition 853
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 50, 848–857

Discussion
The floral transition is one of the major developmental
switches in plants. Hundreds of genes are up- or downreg-
ulated when Arabidopsis is induced to flower (Schmid et al.,
2003). Such an extensive change in the gene expression
pattern is likely to involve major changes in chromatin state.
Our study demonstrates a dramatic decondensation of
pericentromeric and gene-rich chromatin in leaf mesophyll
nuclei during the floral transition. The decrease in compact
chromatin content is under control of a light-signaling
pathway that involves the blue-light photoreceptor CRY2.
Considering that, for an annual plant such as Arabidopsis,
the floral transition is a fundamental and irreversible
developmental event, a correctly established transition is
crucial. Hence, all processes during the floral transition
should be properly timed in order to initiate floral meristem
development. The decondensation of large chromatin
regions including gene-rich and pericentric regions may
facilitate this important developmental switch.
Floral transition is referred to as the switch from veget-
ative to reproductive growth. However, although this devel-
opmental switch results in distinctive morphological
features, such as the formation of flower primordia and the
elongation of internodes (Hempel and Feldman, 1994;
Koornneef and Peeters, 1997), its precise temporal bound-
aries have not been clearly defined. During vegetative
growth the shoot apical meristem produces leaf primordia
and associated axillary buds. Under certain environmental
conditions that induce flowering, such as extended photo-
periods, the shoot apical meristem produces floral primor-
dia upon reception of a florigen signal. This signal is
generated in the leaves and transported to the apical
meristem. It is reasonable to define floral transition under
long-day conditions as the lapse of time between the
initiation of the florigen production in the leaves and the
bolting of the immature flower buds. Of all possible com-
ponents that make up the florigen signal so far only the FT
gene product (mRNA and/or protein) has been identified
(Huang et al., 2005). FT transcription is activated in the
vascular tissue of leaves under long-day conditions by the
flowering activator CO (An et al., 2004; Samach et al., 2000;
Suarez-Lopez et al., 2001; Takada and Goto, 2003). The FT
mRNA and/or protein must be transported to induce
flowering. Within 6 h of induction in the leaves, significant
FT mRNA levels are detectable in the shoot meristem
(Huang et al., 2005). The expression of FT mRNA in the
leaves is maintained for up to 3 days and leads to upregu-
lation of floral identity genes such as AP1 and LFY. This time
frame, between initiation of FT transcript production and the
induction of floral identity genes, corresponds remarkably
well with the period in which we observe the reduction of
pericentric heterochromatin. It suggests that the transient
state of reduced heterochromatin may be mechanistically
related to the emission of FT gene product. In this study we
report decondensation of pericentromeric heterochromatin
and gene-rich chromatin during the floral transition. There-
fore, we propose that decondensation of chromatin in
flowering-induced plants may enhance the accessibility to
DNA for the transcription machinery on a genome-wide
scale.
High CO levels necessary for FT stimulation are estab-
lished by the coincidence of clock-driven high CO expression
in conjunction with CO-stabilizing activities of the photore-
ceptors CRY2 and PHYA during the late afternoon (Valverde
et al., 2004). Both PHYA and CRY2 are involved in the
perception of the photoperiod. CRY2 is a nuclear protein that
has been shown to associate with chromosomes (Cutler
et al., 2000). CRY2 shows homology with DNA photolyase,
but does not have photolyase activity. Virtually nothing is
known about a direct functional interaction between CRY2
and chromatin. The cry2 mutant shows delayed flowering in
LD conditions, whereas overexpression of CRY2 accelerates
flowering under SD conditions (Guo et al., 1998). Our study
Figure 6. Nuclear fraction with normal heterochromatin levels in null mutants
of CRY2 during the floral transition.
Heterochromatin index in cry2 mutant plants in a Col-0 background (a) and a
Ler background, fha-1 (b). Plants were grown in long-day (LD) conditions. Bars
represent stages before (white) or after (gray) bolting. For each bar the data
are mean � SEM of more than 75 nuclei. The age of the plant in days is shown
in brackets.
854 Federico Tessadori et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 50, 848–857

has demonstrated that CRY2 is required for chromatin
decondensation during the floral transition. The transient
reduction of heterochromatin coincides with the floral
transition except in the cry2 mutant, where heterochromatin
levels remain stable. Notably, the correlation between
absence of CRY2 and stable levels of heterochromatin
(Fig. 6) is not only observed during the floral transition.
We have found a dramatic reduction of heterochromatin in
low-light treated wild-type plants, whereas cry2 mutants did
not show this reduction (van Zanten et al., unpublished data
). Low light levels trigger a shade-avoidance response,
which activates a number of processes, such as hyponastic
growth (Ballare, 1999; Millenaar et al., 2005), leading to
morphological changes. Apparently, under certain light
conditions that promote changes of the nuclear program,
CRY2 triggers reorganization of chromatin. The deconden-
sation event is not a prerequisite for flowering, as the cry2
mutant shows high levels of heterochromatin during the
floral transition, yet it produces flowerbuds, albeit delayed
compared with the wild type. We therefore propose that
chromatin decondensation is under control of a light-sign-
aling mechanism involving CRY2, and may facilitate CRY2-
mediated responses to environmental changes.
Heterochromatin reduction coincides with the floral trans-
ition in both LD and SD conditions. The perception of LD or
SD involves the activity of the photoreceptor CRY2 (Guo
et al., 1998). CRY2 stabilizes CO under LD, as its level
fluctuates in SD (El-Din El-Assal et al., 2001). Consequently,
the floral transition is delayed and under control of the
gibberellin pathway (Reeves and Coupland, 2001). Coinci-
dently, the heterochromatin level decreases prior to bolting.
The same happens to co and ft plants grown in LD. However,
in the absence of CRY2, the floral transition is delayed and
under control of the gibberellin pathway, but reduction of
heterochromatin does not take place. This implies that
chromatin decondensation depends on CRY2 action, but is
not affected by CO or FT activity. Conversely, the activities of
CO and FT are unlikely to take place downstream of
heterochromatin decondensation, as they are expressed
before the floral transition. Therefore, we propose that CRY2
promotes two independent processes: CO-mediated flower-
ing and reorganization of chromatin. The two events coin-
cide at the transition from vegetative to reproductive
growth. This explains why the reduction in heterochromatin
occurs prior to bolting and is absent in the cry2 mutant. How
CRY2 may regulate these processes is not clear. It is likely
that E3 ligase complexes, such as COP1, are involved. A
reciprocal effect has been reported between CRY2 and
COP1. CRY2 is unstable and rapidly degraded in blue light
by COP1, whereas COP1 activity in turn is negatively
regulated by CRY2 (Shalitin et al., 2002; Wang et al., 2001).
The latter is carried out via interaction between the WD40
domain of COP1 and the C-terminal domain of CRY2 (Wang
et al., 2001). How CO is stabilized by CRY2 is not yet clear.
CO is degraded via a mechanism that involves SPA proteins
(Laubinger et al., 2006). Proteins of the SPA family are
known to act with COP1 in the degradation of transcription
factors of photomorphogenesis. Accordingly, CRY2 might
stabilize CO via negatively regulating its degradation by a
COP1-like complex. Whether COP1 is required to degrade
CO and if the same mechanism controls chromatin organ-
ization remains to be elucidated.
Experimental procedures
Plant material
Arabidopsis accessions Col-0 (N1092); Ler (NW20) and Cvi-0 (N902)were used for this study. The co-2, ft-1, cry2-1 and fha-1 mutantswere kindly provided by M. Koornneef (MPIZ, Cologne, Germany).The co-2, ft-1 and fha-1 mutants are in the Ler background. The cry2-1 mutant is in the Col-0 background. Plants were grown under whitefluorescent light (70–90 lmol m)2 sec)1) in photoperiods of 16-hlight/8-h dark or 12-h light/12-h dark. Temperature (22�C) andhumidity (70%) remained constant. For experiments, tissue fromyoung rosette leaves, 5–15 mm in size, was used. The first twojuvenile leaves were excluded. The developmental stage of theplants was assessed according to the method described by Boyeset al., 2001.
Fluorescence in situ hybridization (FISH)
The DNA fragments used as FISH probes were labeled with eitherbiotin- or digoxigenin-Nick Translation Mixes (Roche, http://www.roche.com). Plasmid pAL1 (Martinez-Zapater et al., 1986) wasused to detect the 180-bp centromeric tandem repeat. BAC F28D6(DDBJ/EMBL/GenBank accession No. AF147262) in pBeloBAC-Kanvector was obtained from the Nottingham Arabidopsis Stock Centre(NASC, http://nasc.nott.ac.uk) . The AGAMOUS locus was localizedusing BAC F13C5 (DDBJ/EMBL/GenBank accession No. AL021711;119.1 kb) and the two adjacent BACs T18B16 (DDBJ/EMBL/GenBankaccession No. AL021687; 96.5 kb) and F28A21 (DDBJ/EMBL/Gen-Bank accession No. AL025526; 94.3 kb).
FISH experiments were carried out as described in Schubert et al.,(2001) and detected with antibodies Avidin-Texas Red (VectorLaboratories, http://www.vectorlabs.com) and goat anti-Avidin(Vector Laboratories) for biotin-labeled probes; mouse anti-digo-xigenin (Roche), rabbit anti-mouse�FITC (Sigma-Aldrich, http://www.sigmaaldrich.com) and goat anti-rabbit�Alexa488 (MolecularProbes, http://www.invitrogen.com) for the detection of digoxige-nin-labeled probes. Prior to observation, slides were counterstainedwith 4¢,6-diamidino-2-phenylindole (DAPI, 2 lg ml)1; Roche) inVectashield (Vector Laboratories).
5-Methylcytosine detection
Preparations were dried at 60�C for 30 min, treated with RNAse(10 lg ml)1 in 2X SSC ; Roche) for 60 min at 37�C, rinsed twice for5 min in 2X SSC and 5 min in PBS (10 mM sodium phosphate,pH 7.0, 143 mM NaCl), fixed for 10 min in 1% formaldehyde in PBSat room temperature, rinsed twice for 5 min in PBS, dehydrated bysuccessive 1-min baths in 70, 90 and 100% ethanol and then airdried. Denaturation was carried out by adding 50 ll HB50 (50%formamide in 1X SSC) and heating at 80�C for 2 min. The slides
Chromatin reorganization during floral transition 855
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 50, 848–857

where then washed in 70% ice-cold ethanol and dehydrated bysuccessive ethanol baths. Blocking of the slides was carried out for1 h in 1% BSA in PBS and, after three 5-min washes in TNT (1 M Tris/HCl, pH 8.0, 1 M NaCl, 0.5% Tween 20), incubation with themouse antibody against 5-methylcytosine (Eurogentec, http://www.eurogentec.be) was carried out at room temperature overnight.Detection of the antibody was carried out with the same antibodiesused for FISH with digoxigenin-labeled probes.
Image acquisition and processing
Image acquisition was carried out on an Olympus BX60 microscope(Olympus, http://www.olympus-global.com) with filters for DAPI,Texas Red and fluorescein isothiocyanate (FITC). Pictures werecaptured with a charge-coupled device (CCD) camera (Coolsnap FX;Photometrics, http://www.photomet.com). The images were digit-ally processed using ADOBE PHOTOSHOP (Adobe, http://www.adobe.-com).
Measurement of RHF and HX
Greyscale images were analyzed with OBJECTIMAGE freeware (http://simon.bio.uva.nl/object-image.html). The RHF was calculated frommesophyll nuclei as described by Soppe et al., 2002; Tessadoriet al., 2004; and is defined as the area and fluorescence intensity ofchromocenters in relation to the area and fluorescence intensity ofthe entire nucleus. The HX is defined as the percentage of normalnuclei showing a relatively high content of compact chromatin,represented by 6–10 round or oval conspicuous chromocenters(Fransz et al., 2002, 2003). Nuclei containing fewer chromocenters,smaller chromocenters or chromocenters with vague contours areconsidered abnormal. Only mesophyll nuclei are used to measureRHF or HX. Nuclei from epidermis, vascular tissue or endoredupli-cated cells are not included in the measurements.
Acknowledgements
We thank Maarten Koornneef (MPIZ, Cologne, Germany) forvaluable discussions and Corrie Hanhart (WVR, Wageningen,Netherlands) for the seeds of the co-2, ft-1, cry2-1 and fha-1 mutantsused in this work.
Supplementary Material
The following supplementary material is available for this articleonline:Figure S1. Southern blot analysis of DNA methylation.
This material is available as part of the online article from http://www.blackwell-synergy.com
References
Abe, M., Kobayashi, Y., Yamamoto, S., Daimon, Y., Yamaguchi, A.,
Ikeda, Y., Ichinoki, H., Notaguchi, M., Goto, K. and Araki, T. (2005)FD, a bZIP protein mediating signals from the floral pathwayintegrator FT at the shoot apex. Science, 309, 1052–1056.
An, H.L., Roussot, C., Suarez-Lopez, P. et al. (2004) CONSTANS actsin the phloem to regulate a systemic signal that induces photo-periodic flowering of Arabidopsis. Development, 131, 3615–3626.
Ballare, C.L. (1999) Keeping up with the neighbours: phytochromesensing and other signalling mechanisms. Trends Plant Sci. 4, 201.
Bastow, R., Mylne, J.S., Lister, C., Lippman, Z., Martienssen, R.A.
and Dean, C. (2004) Vernalization requires epigenetic silencing ofFLC by histone methylation. Nature, 427, 164–167.
Boss, P.K., Bastow, R.M., Mylne, J.S. and Dean, C. (2004) Multiplepathways in the decision to flower: enabling, promoting, andresetting. Plant Cell, 16(Suppl.), S18–S31.
Boyes, D.C., Zayed, A.M., Ascenzi, R., McCaskill, A.J., Hoffman, N.E.,
Davis, K.R. and Gorlach, J. (2001) Growth stage-based phenotypicanalysis of arabidopsis: a model for high throughput functionalgenomics in plants. Plant Cell, 13, 1499–1510.
Bradley, D., Ratcliffe, O., Vincent, C., Carpenter, R. and Coen, E.
(1997) Inflorescence commitment and architecture in Arabidop-sis. Science, 275, 80–83.
Cutler, S.R., Ehrhardt, D.W., Griffitts, J.S. and Somerville, C.R.
(2000) Random GFP::cDNA fusions enable visualization of sub-cellular structures in cells of Arabidopsis at a high frequency.Proc. Natl Acad. Sci. USA, 97, 3718–3723.
El-Din El-Assal, S., Alonso-Blanco, C., Peeters, A.J., Raz, V. and
Koornneef, M. (2001) A QTL for flowering time in Arabidopsisreveals a novel allele of CRY2. Nat. Genet. 29, 435–440.
Fransz, P., De Jong, J.H., Lysak, M., Castiglione, M.R. and Schubert,
I. (2002) Interphase chromosomes in Arabidopsis are organizedas well defined chromocenters from which euchromatin loopsemanate. Proc. Natl Acad. Sci. USA, 99, 14584–14589.
Fransz, P., Soppe, W. and Schubert, I. (2003) Heterochromatin ininterphase nuclei of Arabidopsis thaliana. Chromosome Res. 11,227–240.
Fransz, P., ten Hoopen, R. and Tessadori, F. (2006) Composition andformation of heterochromatin in Arabidopsis thaliana. Chromo-some Res. 14, 71–82.
Gaudin, V., Libault, M., Pouteau, S., Juul, T., Zhao, G., Lefebvre, D.
and Grandjean, O. (2001) Mutations in LIKE HETEROCHROMATINPROTEIN 1 affect flowering time and plant architecture in Ara-bidopsis. Development, 128, 4847–4858.
Gendall, A.R., Levy, Y.Y., Wilson, A. and Dean, C. (2001) The VER-NALIZATION 2 gene mediates the epigenetic regulation of ver-nalization in Arabidopsis. Cell, 107, 525–535.
Guo, H., Yang, H., Mockler, T.C. and Lin, C. (1998) Regulation offlowering time by Arabidopsis photoreceptors. Science, 279,1360–1363.
He, Y. and Amasino, R.M. (2005) Role of chromatin modification inflowering-time control. Trends Plant Sci. 10, 30–35.
He, Y., Michaels, S.D. and Amasino, R.M. (2003) Regulation offlowering time by histone acetylation in Arabidopsis. Science,302, 1751–1754.
He, Y., Doyle, M.R. and Amasino, R.M. (2004) PAF1-complex-medi-ated histone methylation of FLOWERING LOCUS C chromatin isrequired for the vernalization-responsive, winter-annual habit inArabidopsis. Genes Dev. 18, 2774–2784.
Hempel, F.D. and Feldman, L.J. (1994) Bidirectional InflorescenceDevelopment in Arabidopsis thaliana – acropetal Initiation ofFlowers and Basipetal Initiation of Paraclades. Planta, 192, 276–286.
Hepworth, S.R., Valverde, F., Ravenscroft, D., Mouradov, A. and
Coupland, G. (2002) Antagonistic regulation of flowering-timegene SOC1 by CONSTANS and FLC via separate promoter motifs.EMBO J. 21, 4327–4337.
Huang, T., Bohlenius, H., Eriksson, S., Parcy, F. and Nilsson, O.
(2005) The mRNA of the Arabidopsis gene FT moves from leaf toshoot apex and induces flowering. Science, 309, 1694–1696.
Johanson, U., West, J., Lister, C., Michaels, S., Amasino, R. and
Dean, C. (2000) Molecular analysis of FRIGIDA, a major deter-minant of natural variation in Arabidopsis flowering time. Sci-ence, 290, 344–347.
856 Federico Tessadori et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 50, 848–857

Koornneef, M. and Peeters, A.J.M. (1997) Floral transition mutantsin Arabidopsis. Plant Cell Environ. 20, 779–784.
Kotake, T., Takada, S., Nakahigashi, K., Ohto, M. and Goto, K. (2003)Arabidopsis TERMINAL FLOWER 2 gene encodes a heterochro-matin protein 1 homolog and represses both FLOWERING LOCUST to regulate flowering time and several floral homeotic genes.Plant Cell Physiol. 44, 555–564.
Larsson, A.S., Landberg, K. and Meeks-Wagner, D.R. (1998) TheTERMINAL FLOWER2 (TFL2) gene controls the reproductivetransition and meristem identity in Arabidopsis thaliana. Genet-ics, 149, 597–605.
Laubinger, S., Marchal, V., Gentilhomme, J., Wenkel, S., Adrian, J.,
Jang, S., Kulajta, C., Braun, H., Coupland, G. and Hoecker, U.
(2006) Arabidopsis SPA proteins regulate photoperiodic flower-ing and interact with the floral inducer CONSTANS to regulate itsstability. Development, 133, 3213–3222.
Levy, Y.Y., Mesnage, S., Mylne, J.S., Gendall, A.R. and Dean, C.
(2002) Multiple roles of Arabidopsis VRN1 in vernalization andflowering time control. Science, 297, 243–246.
Libault, M., Tessadori, F., Germann, S., Snijder, B., Fransz, P. and
Gaudin, V. (2005) The Arabidopsis LHP1 protein is a componentof euchromatin. Planta, 222, 910–925.
Liu, J., Yu, J., McIntosh, L., Kende, H. and Zeevaart, J.A. (2001)Isolation of a CONSTANS ortholog from Pharbitis nil and its rolein flowering. Plant Physiol. 125, 1821–1830.
Martinez-Garcia, J.F., Virgos-Soler, A. and Prat, S. (2002) Control ofphotoperiod-regulated tuberization in potato by the Arabidopsisflowering-time gene CONSTANS. Proc. Natl Acad. Sci. USA, 99,15211–15216.
Martinez-Zapater, J.M., Estelle, M.A. and Somerville, C.R. (1986) Ahigh repeated DNA sequence in Arabidopsis thaliana. Mol. Gen.Genet. 204, 417–423.
Mas, P., Devlin, P.F., Panda, S. and Kay, S.A. (2000) Functionalinteraction of phytochrome B and cryptochrome 2. Nature, 408,207–211.
Mathieu, O., Jasencakova, Z., Vaillant, I., Gendrel, A.V., Colot, V.,
Schubert, I. and Tourmente, S. (2003) Changes in 5S rDNAchromatin organization and transcription during heterochroma-tin establishment in Arabidopsis. Plant Cell, 15, 2929–2939.
Millenaar, F.F., Cox, M.C., van Berkel, Y.E., Welschen, R.A., Pierik, R.,
Voesenek, L.A. and Peeters, A.J. (2005) Ethylene-induceddifferential growth of petioles in Arabidopsis. Analyzing naturalvariation, response kinetics, and regulation. Plant Physiol. 137,998–1008.
Mylne, J.S., Barrett, L., Tessadori, F., Mesnage, S., Johnson, L.,
Bernatavichute, Y.V., Jacobsen, S.E., Fransz, P. and Dean, C.
(2006) LHP1, the Arabidopsis homologue of HETEROCHROMA-TIN PROTEIN1, is required for epigenetic silencing of FLC. Proc.Natl Acad. Sci. USA, 103, 5012–5017.
Pavet, V., Quintero, C., Cecchini, N.M., Rosa, A.L. and Alvarez, M.E.
(2006) Arabidopsis displays centromeric DNA hypomethylationand cytological alterations of heterochromatin upon attack bypseudomonas syringae. Mol. Plant Microbe Interact. 19, 577–587.
Putterill, J., Robson, F., Lee, K., Simon, R. and Coupland, G. (1995)The CONSTANS gene of Arabidopsis promotes flowering andencodes a protein showing similarities to zinc finger transcriptionfactors. Cell, 80, 847–857.
Putterill, J., Laurie, R. and Macknight, R. (2004) It’s time to flower:the genetic control of flowering time. Bioessays, 26, 363–373.
Reeves, P.H. and Coupland, G. (2001) Analysis of flowering timecontrol in Arabidopsis by comparison of double and triplemutants. Plant Physiol. 126, 1085–1091.
Samach, A., Onouchi, H., Gold, S.E., Ditta, G.S., Schwarz-Sommer,
Z., Yanofsky, M.F. and Coupland, G. (2000) Distinct roles ofCONSTANS target genes in reproductive development ofArabidopsis. Science, 288, 1613–1616.
Schmid, M., Uhlenhaut, N.H., Godard, F., Demar, M., Bressan, R.,
Weigel, D. and Lohmann, J.U. (2003) Dissection of floral inductionpathways using global expression analysis. Development, 130,6001–6012.
Schubert, I., Fransz, P.F., Fuchs, J. and de Jong, J.H. (2001)Chromosome painting in plants. Methods Cell Sci. 23, 57–69.
Shalitin, D., Yang, H., Mockler, T.C., Maymon, M., Guo, H.,
Whitelam, G.C. and Lin, C. (2002) Regulation of Arabidopsiscryptochrome 2 by blue-light-dependent phosphorylation.Nature, 417, 763–767.
Sheldon, C.C., Burn, J.E., Perez, P.P., Metzger, J., Edwards, J.A.,
Peacock, W.J. and Dennis, E.S. (1999) The FLF MADS box gene: arepressor of flowering in Arabidopsis regulated by vernalizationand methylation. Plant Cell, 11, 445–458.
Soppe, W.J., Jasencakova, Z., Houben, A., Kakutani, T., Meister, A.,
Huang, M.S., Jacobsen, S.E., Schubert, I. and Fransz, P.F. (2002)DNA methylation controls histone H3 lysine 9 methylation andheterochromatin assembly in Arabidopsis. EMBO J. 21, 6549–6559.
Suarez-Lopez, P., Wheatley, K., Robson, F., Onouchi, H., Valverde, F.
and Coupland, G. (2001) CONSTANS mediates between the cir-cadian clock and the control of flowering in Arabidopsis. Nature,410, 1116–1120.
Sung, S. and Amasino, R.M. (2004) Vernalization in Arabidopsisthaliana is mediated by the PHD finger protein VIN3. Nature, 427,159–164.
Takada, S. and Goto, K. (2003) Terminal flower2, an Arabidopsishomolog of heterochromatin protein1, counteracts the activationof flowering locus T by constans in the vascular tissues of leavesto regulate flowering time. Plant Cell, 15, 2856–2865.
Tessadori, F., van Driel, R. and Fransz, P. (2004) Cytogenetics as atool to study gene regulation. Trends Plant Sci. 9, 147–153.
Tessadori, F., Chupeau, M.-C., Chupean, Y., Knip, M., Germann, S.,
van Driel, R., Fransz, P. and Gaudin, V. (2007) Large-scale dis-sociation and sequential reassembly of pericentric hetero-chromatin dedifferentiated Arabidopsis cells. J. Cell Sci, in press.
Valverde, F., Mouradov, A., Soppe, W., Ravenscroft, D., Samach, A.
and Coupland, G. (2004) Photoreceptor regulation of CONSTANSprotein in photoperiodic flowering. Science, 303, 1003–1006.
Vince-Prue, D. and Gressel, J. (1985) Pharbitis nil. in Handbook ofFlowering, Vol. IV (Halevy, A., ed.). Boca Raton, FL: CRC Press, pp.47–81.
Wang, H., Ma, L.G., Li, J.M., Zhao, H.Y. and Deng, X.W. (2001) Directinteraction of Arabidopsis cryptochromes with COP1 in lightcontrol development. Science, 294, 154–158.
Wigge, P.A., Kim, M.C., Jaeger, K.E., Busch, W., Schmid, M.,
Lohmann, J.U. and Weigel, D. (2005) Integration of spatial andtemporal information during floral induction in Arabidopsis.Science, 309, 1056–1059.
Yanovsky, M.J. and Kay, S.A. (2002) Molecular basis of seasonaltime measurement in Arabidopsis. Nature, 419, 308–312.
Zeevaart, J.A.D. (1976) Physiology of flower formation. Annu. Rev.Plant Physiol. 27, 321–348.Accession numbers: co-2, AT5 G15840;seed stock, CS55; ft-1, AT1 G65480; seed stock, CS56; cry2-1,AT1 G04400; seed stock, CS3732; fha-1, AT1 G04400; seed stock,CS108.
Chromatin reorganization during floral transition 857
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 50, 848–857




















