
LIGHT AND TEMPERATURE EFFECT ON THE GERMINATION OF Leandra breviflora COGN., Tibouchina benthamiana...
12
EFEITO DA LUZ E DA TEMPERATURA NA GERMINAÇÃO DE Leandra breviflora COGN., Tibouchina benthamiana COGN., Tibouchina grandifolia COGN. E Tibouchina moricandiana (DC.) BAILL. (MELASTOMATACEAE) 1 ANTONIO CARLOS SILVA DE ANDRADE 2 Revista Brasileira de Sementes, vol. 17, n o 1, p. 29-35, 1995 RESUMO - O objetivo deste trabalho foi estudar a germinação de sementes de Leandra breviflora, Tibouchina benthamiana, Tibouchina grandifolia e Tibouchina moricandiana (Melastomataceae) sob diferentes condições de luz e temperatura, utilizando-se como parâmetros a porcentagem e a velocidade de germinação. Os resultados demonstraram que a temperatura ótima para a germinação de sementes de todas as espécies foi de 30°C; verificou-se a redução ou a ausência de germinação nas temperaturas de 15 e 35°C. Todas as espécies estudadas são fotoblásticas positivas em todas as temperaturas testadas; a exposição ao fotoperíodo de 24 horas causou a redução nos valores de porcentagem e de velocidade de germinação, o que sugere que as sementes das espécies estudadas apresentam um comportamento fotoperiódico de dias curtos. Termos para indexação: fotoperíodo, temperatura, Melastomataceae, germinação LIGHT AND TEMPERATURE EFFECT ON THE GERMINATION OF Leandra breviflora COGN., Tibouchina benthamiana COGN., Tibouchina grandifolia COGN. AND Tibouchina moricandiana (DC.) BAILL. (MELASTOMATACEAE) ABSTRACT - The purpose of this study was to investigate seed germination of Leandra breviflora, Tibouchina benthamiana, Tibouchina grandifolia and Tibouchina moricandiana under different light and temperature conditions, by assessing germination percentage and speed. The optimum temperature for seed germination of all species was 30°C; reduction or absence of germination took place at 15 and 35°C. All species studied are fotoblastic positive for all temperatures tested; 24 hours photoperiod caused a reduction in germination percentage and speed, which suggests that the seeds of the species studied have a photoperiodic behaviour of short days. Index terms: photoperiod, temperature, Melastomataceae, germination INTRODUÇÃO A família Melastomataceae é representada na flora brasileira por cerca de 69 gêneros e aproximadamente 1.500 espécies (Baumgratz, 1983-1985). 1 Aceito para publicação em 28.12.94 2 Eng. Agr., M.Sc., Pesquisador do Programa Mata Atlântica - FBMM/JBRJ - Laboratório de Sementes do Jardim Botânico do Rio de Janeiro, Rua Pacheco Leão, 915 - CEP 22.460-030 - Rio de Janeiro-RJ
Transcript of LIGHT AND TEMPERATURE EFFECT ON THE GERMINATION OF Leandra breviflora COGN., Tibouchina benthamiana...

EFEITO DA LUZ E DA TEMPERATURA NA GERMINAÇÃO DE Leandra breviflora COGN., Tibouchina benthamiana COGN., Tibouchina grandifolia COGN. E Tibouchina moricandiana (DC.) BAILL. (MELASTOMATACEAE)1
ANTONIO CARLOS SILVA DE ANDRADE2
Revista Brasileira de Sementes, vol. 17, no 1, p. 29-35, 1995
RESUMO - O objetivo deste trabalho foi estudar a germinação de sementes de Leandra breviflora, Tibouchina benthamiana, Tibouchina grandifolia e Tibouchina moricandiana (Melastomataceae) sob diferentes condições de luz e temperatura, utilizando-se como parâmetros a porcentagem e a velocidade de germinação. Os resultados demonstraram que a temperatura ótima para a germinação de sementes de todas as espécies foi de 30°C; verificou-se a redução ou a ausência de germinação nas temperaturas de 15 e 35°C. Todas as espécies estudadas são fotoblásticas positivas em todas as temperaturas testadas; a exposição ao fotoperíodo de 24 horas causou a redução nos valores de porcentagem e de velocidade de germinação, o que sugere que as sementes das espécies estudadas apresentam um comportamento fotoperiódico de dias curtos. Termos para indexação: fotoperíodo, temperatura, Melastomataceae, germinação
LIGHT AND TEMPERATURE EFFECT ON THE GERMINATION OF Leandra breviflora COGN., Tibouchina benthamiana COGN., Tibouchina grandifolia
COGN. AND Tibouchina moricandiana (DC.) BAILL. (MELASTOMATACEAE)
ABSTRACT - The purpose of this study was to investigate seed germination of Leandra breviflora, Tibouchina benthamiana, Tibouchina grandifolia and Tibouchina moricandiana under different light and temperature conditions, by assessing germination percentage and speed. The optimum temperature for seed germination of all species was 30°C; reduction or absence of germination took place at 15 and 35°C. All species studied are fotoblastic positive for all temperatures tested; 24 hours photoperiod caused a reduction in germination percentage and speed, which suggests that the seeds of the species studied have a photoperiodic behaviour of short days. Index terms: photoperiod, temperature, Melastomataceae, germination
INTRODUÇÃO
A família Melastomataceae é representada na flora brasileira por cerca de 69 gêneros e aproximadamente 1.500 espécies (Baumgratz, 1983-1985). 1 Aceito para publicação em 28.12.94 2 Eng. Agr., M.Sc., Pesquisador do Programa Mata Atlântica - FBMM/JBRJ - Laboratório de
Sementes do Jardim Botânico do Rio de Janeiro, Rua Pacheco Leão, 915 - CEP 22.460-030 - Rio de Janeiro-RJ

Dentre as espécies conhecidas, muitas apresentam grande valor ornamental, tais como a quaresmeira (Tibouchina granulosa Cogn.) e o manacá-da-serra (Tibouchina mutabilis Cogn.). Além disso, muitas espécies de melastomataceas são rústicas e heliófitas, sendo utilizadas com freqüência na recuperação e reflorestamento de áreas degradadas (Lorenzi, 1992; Pompéia et al., 1989). Outras espécies, tais como jacatirão (Miconia cinnamomifolia (DC.)) apresentam potencial econômico na sua exploração seletiva e manejo de sua regeneração natural, pelas características de sua madeira (Kageyama & Reis, 1993).
A espécie Leandra breviflora é endêmica da região de Friburgo-RJ, enquanto que Tibouchina benthamiana, Tibouchina grandifolia e Tibouchina moricandiana apresentam ampla distribuição pelo estado do Rio de Janeiro. Das quatro espécies citadas, somente Leandra breviflora ocorre em áreas de sub-bosque; as demais aparecem com bastante freqüência nas bordas das clareiras e beiras de estradas. Tibouchina benthamiana e Tibouchina grandifolia apresentam hábito arbustivo, enquanto que Leandra breviflora e Tibouchina moricandiana possuem porte arbóreo (Programa Mata Atlântica, 1990; Andrade, Com. Pessoal).
A temperatura e a luz são os principais fatores ambientais que promovem a germinação de sementes em solos úmidos. Para muitas espécies, se fornecidas as condições ideais de luz e umidade, a temperatura predominante do solo determina tanto a fração de sementes germinadas de uma amostra como a sua velocidade de germinação. A fração de sementes que germina, frequentemente permanece constante dentro de uma grande amplitude de temperaturas e decresce rapidamente para valores abaixo e acima dessa mesma amplitude (Heydecker, 1977).
Conforme Dau & Labouriau (1974) e Labouriau & Pacheco (1978), os limites extremos de temperatura de germinação fornecem informações de interesse biológico e ecológico, onde sementes de diferentes espécies apresentam faixas distintas de temperatura para a germinação. Dentro dessas faixas, pode ser considerada como temperatura ótima aquela na qual a mais alta porcentagem de germinação é obtida dentro do menor espaço de tempo. Seriam consideradas ainda a mínima e a máxima, respectivamente, como a mais baixa e a mais alta temperatura onde a germinação ocorre (Mayer & Poljakoff-Mayber, 1989).
Em relação ao fator luz, observou-se que as sementes de muitas espécies cultivadas apresentam alta germinação, tanto em presença como em ausência de luz. Entretanto, o estímulo luminoso à germinação é bastante variável para sementes de muitas espécies selvagens (Mayer & Poljakoff-Mayber, 1989). Assim, as sementes foram classificadas em 3 grandes grupos, com relação a sua resposta de germinação ao estímulo luminoso: fotoblásticas positivas, que não germinam no escuro e são produzidas principalmente por plantas heliófitas (as quais requerem luz solar intensa para crescer); fotoblásticas negativas, cuja germinação é inibida pela luz; e indiferentes à luz, produzidas principalmente por árvores de sub-bosques e plantas de sombra (Côme, 1970, apud Orozco-Segovia & Vázquez-Yanes, 1992).
A fisiologia de fotocontrole é complexa e bastante variada. Sementes de algumas espécies têm sua germinação promovida por curtas exposições à luz

branca, com baixo nível de energia; outras requerem irradiação intermitente; algumas precisam de iluminação diária, com certo número de horas de luz (fotoperíodo); outras são estimuladas por curtos períodos de exposição à luz e inibidas por longos períodos de exposição à luz; e quase todas estão sujeitas a interações aparentemente complexas entre a temperatura e as condições de iluminação (Bewley & Black, 1985).
A germinação das espécies pertencentes à família Melastomataceae tem sido pouco estudada. Queiroz (1982) analisou a importância da maturidade dos frutos e da influência da luz na germinação de sementes de Miconia cinnamomifolia, constatando que a germinação de sementes provenientes de frutos imaturos foi próxima àquela encontrada para frutos maturos. Além disso, suas sementes apresentaram fotoblastismo acentuado, germinando somente quando expostas à luz. Em Tibouchina fothergillae, Pinto et al. (1983) verificaram que, apesar do fotoblastismo positivo em suas sementes, a germinação sob ausência de luz atingiu valores próximos a 35%.
Face à existência de poucos estudos enfocando a germinação em espécies da família Melastomataceae, este trabalho teve como objetivo estudar os efeitos de diferentes condições de luz e temperatura na germinação de sementes de Leandra breviflora, Tibouchina benthamiana, Tibouchina grandifolia e Tibouchina moricandiana. MATERIAL E MÉTODOS
Sementes de Leandra breviflora, Tibouchina benthamiana, Tibouchina grandifolia e Tibouchina moricandiana foram obtidas a partir de frutos maduros, coletados de pelo menos dois indivíduos na Reserva Ecológica de Macaé-de-Cima (RJ), sendo armazenadas durante 15 dias sob ausência de luz, a 10°C e 20% U.R., no Laboratório de Sementes do Jardim Botânico do Rio de Janeiro.
No primeiro ensaio, estudou-se o efeito de cinco diferentes temperaturas constantes (15, 20, 25, 30 e 35 0,5 C), sob fotoperíodo de 8 horas, na porcentagem final e velocidade de germinação de sementes. No segundo ensaio, estudou-se o número de dias de exposição à luz, sob fotoperíodo de 8 horas, necessários à máxima germinação de sementes. Finalmente, avaliou-se a influência de diferentes fotoperíodos (ausência de luz, 8 e 24 horas de luz) na porcentagem final e velocidade de germinação de sementes. Para os dois últimos ensaios, utilizou-se a melhor temperatura de germinação para as espécies, conforme resultados obtidos no primeiro ensaio (30°C).
Para todos os experimentos, os testes de germinação foram conduzidos em placas de Petri, com seis repetições contendo 25 sementes por placa, as quais foram semeadas sobre duas folhas de papel de filtro umedecidas com água destilada. Os experimentos foram conduzidos em câmaras de germinação do tipo B.O.D., iluminadas com lâmpadas fluorescentes do tipo “luz do dia” (4 x 20w), com densidade do fluxo radiante na altura das placas de 15µmol m-2 s-1, medidas com quantum radiômetroLi-Cor.
O critério utilizado para avaliar a germinação foi o aparecimento das estruturas essenciais da plântula em perfeito estágio de desenvolvimento (germinação normal), sendo verificada diariamente para todos os tratamentos com luz. Na condição de escuro contínuo, as placas de Petri foram inseridas

em embalagens de polietileno preta com camada interna de papel aluminizado, com a germinação avaliada somente ao final do experimento. A velocidade de germinação (v) foi determinada de acordo com o critério estabelecido por Labouriau & Valadares (1976), obtida pela fórmula: v = ni/(ni.ti), onde ni é o número de sementes germinadas dentro do intervalo de tempo ti, sendo os resultados expressos em dias-1.
Os valores em porcentagem foram transformados em arco seno (%/100)1/2 para normalização de sua distribuição (Bartlett, 1947). Os resultados foram avaliados estatiticamente, onde foi adotado o delineamento inteiramente casualizado. Foi utilizado o teste de Tukey, ao nível de 5% de probabilidade para comparação entre as médias, quando houve significância pelo teste F (Steel & Torne, 1980).
RESULTADOS E DISCUSSÃO Influência da luz e da temperatura
Em todos os experimentos, as sementes foram submetidas a períodos de 24 horas de embebição. Esse período de embebição pode ser necessário para que o fitocromo seja transformado em sua forma ativa Fve, já que em tecidos vegetais desidratados, a fototransformação pode não ocorrer (Wulff & Medina, 1969; Takaki et al., 1981).
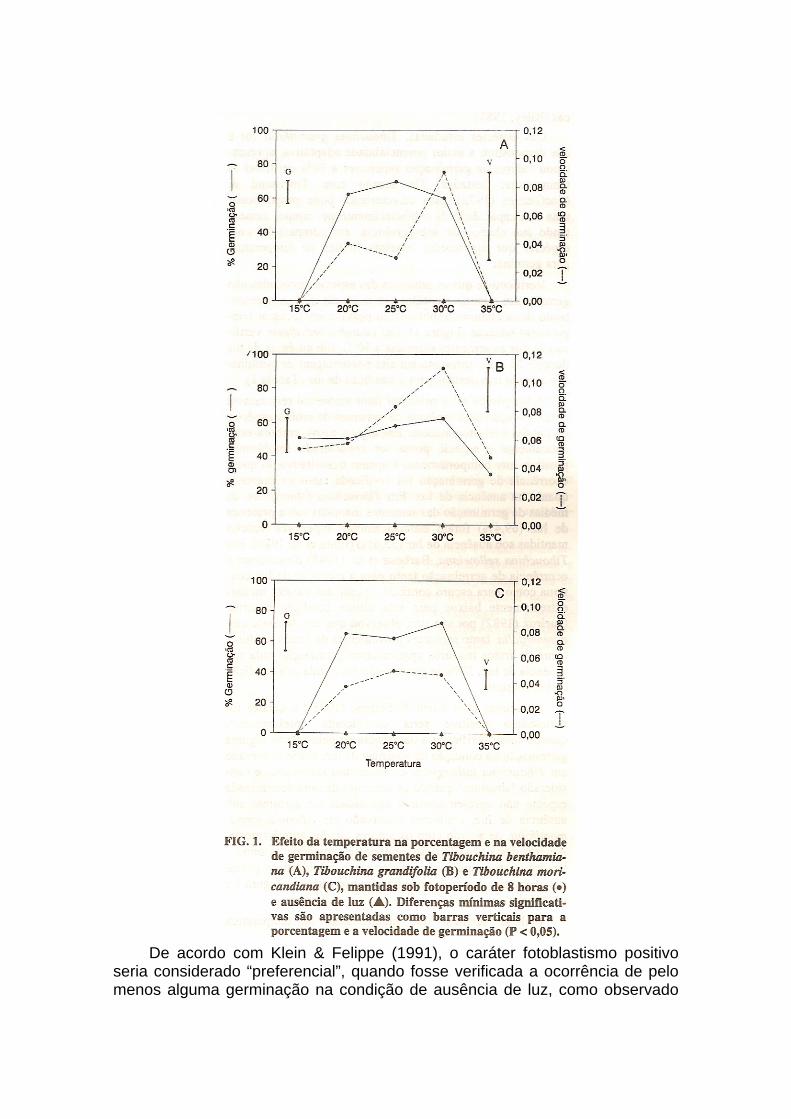
Com relação aos valores de porcentagem de germinação, verificou-se que as temperaturas de 20, 25 e 30°C foram estatisticamente superiores às demais temperaturas testadas, em todas espécies estudadas, exceto para Tibouchina grandifolia, onde a temperatura de 15°C não apresentou valores de germinação estatisticamente diferentes às melhores temperaturas.
Para as médias de velocidade de germinação, verificou-se que, independente da espécie estudada, os maiores valores foram obtidos na temperatura de 30°C, com exceção para Tibouchina moricandiana, onde a temperatura de 25°C produziu os maiores valores de velocidade de germinação. Em Tibouchina benthamiana e Tibouchina moricandiana, os valores de velocidade de germinação não apresentaram diferenças estatísticas significativas nas temperaturas de 20, 25 e 30°C; em Tibouchina grandifolia, a velocidade de germinação não apresentou diferenças significativas nas temperaturas de 25 e 30°C.
A temperatura de 15°C não permitiu o desenvolvimento do processo germinativo em Tibouchina benthamiana e Tibouchina moricandiana. Todavia, quando transferidas à temperatura de 30°C, os valores de germinação foram semelhantes aos obtidos para as temperaturas de 25 e 30°C (resultados não demonstrados). Em Tibouchina grandifolia, verificou-se que, apesar dos valores de germinação a 15°C, não apresentarem diferenças estatísticas em relação às melhores temperaturas, a sua velocidade de germinação foi estatisticamente inferior à temperatura de 25 e 30°C. De acordo com Hendricks & Taylorson (1976), baixas temperaturas podem reduzir as taxas metabólicas até que as vias essenciais ao início da germinação não possam mais operar. Em sementes fotoblásticas, a temperatura ainda pode afetar a germinação através

dos processos de reidratação, síntese do fitocromo e da interconversão de suas formas (Toole, 1973).
Na temperatura de 35°C, apesar de iniciado o processo de germinação através da protusão da radícula, verificou-se que o subseqüente desenvolvimento do processo germinativo não ocorreu, resultando no aparecimento de poucas plântulas normais, como em Tibouchina grandifolia (30%) ou mesmo a sua ausência, como em Tibouchina benthamiana e Tibouchina moricandiana (Figura 1). Quando transferidas para a temperatura de 30°C, verificou-se a ausência de germinação para as três espécies, o que permite supor que o tratamento 35°C induziu as sementes à dormência secundária ou provocou a perda da sua viabilidade. Segundo Vidaver & Hsiao (1975), altas temperaturas produzem dormência térmica ou até mesmo a perda da viabilidade das sementes através do estresse térmico. Na maioria dos casos, o estresse térmico retarda o desenvolvimento do processo germinativo, podendo suprimí-lo em sementes quiescentes ou para as que já haviam iniciado sua germinação (Pollock & Ross, 1972).
Em geral, altas temperaturas provocam uma diminuição do suprimento de aminoácidos livres, da síntese de RNA e de proteínas e o decréscimo da velocidade de reações metabólicas (Riley, 1981).
Das espécies estudadas, Tibouchina grandifolia foi a que demonstrou a maior potencialidade adaptativa, apresentando valores de germinação superiores a 30% em todas as temperaturas testadas. De acordo com Townsend & MacGinnies (1972), essa característica pode proporcionar uma alta capacidade de estabelecimento no campo, aumentando sua chance de sobrevivência, em comparação com espécies que apresentam estreitos limites de temperatura para germinar.
Verificou-se que as sementes das espécies estudadas não germinaram quando mantidas sob ausência de luz, apresentando desta maneira fotoblastismo positivo em todas as temperaturas testadas (Figura 1). Em Leandra breviflora, verificou-se que as sementes mantidas a 30°C, sob ausência de luz durante 25 dias, apresentaram alta porcentagem de germinação quando transferidas para a condição de luz (Tabela 1).
A luz parece ser o principal fator ambiental responsável pela superação da dormência de sementes de muitas espécies da família Melastomataceae. Em alguns casos, embora estatisticamente a espécie possa ser considerada fotoblástica positiva, este comportamento é apenas quantitativo, já que a ocorrência de germinação foi verificada tanto na presença quanto na ausência de luz. Em Tibouchina fothergillae, as médias de germinação das sementes mantidas sob a presença de luz (69,4%) foram estatisticamente superiores àquelas mantidas sob ausência de luz (35,6%) (Pinto et al., 1983). Em Tibouchina sellowiana, Barbosa et al. (1988) observaram a ocorrência de germinação tanto para a condição de luz contínua como para escuro contínuo, apesar dos valores médios extremamente baixos para esta última condição descrita. Queiroz (1982), por sua vez, observou que em Miconia cinnamomifolia, tanto sementes provenientes de frutos imaturos como de frutos maduros, apresentaram germinação nula sob ausência de luz; 50% de germinação foi obtida sob condição de luz contínua.
De acordo com Klein & Felippe (1991), o caráter fotoblastismo positivo
seria considerado “preferencial”, quando fosse verificada a ocorrência de pelo menos alguma germinação na condição de ausência de luz, como observado

em Tibouchina fothergillae e Tibouchina sellowiana e considerado “absoluto” quando as sementes de uma determinada espécie não apresentassem a capacidade de germinar sob ausência de luz, conforme observado em Miconia cinnamomifolia, de acordo com os autores citados anteriormente. A partir desta classificação, poder-se-ia considerar as sementes das quatro espécies estudadas como fotoblásticas positivas absolutas, diante dos resultados observados na Figura 1 e na Tabela 1.
Pode-se verificar que, independente da temperatura testada, as sementes
das espécies estudadas não germinaram quando submetidas à condição de ausência de luz; o processo de germinação foi iniciado somente após exposição à luz durante 8 horas por dia. Para muitas espécies estudadas, o comportamento de fotoblastismo de suas sementes pode estar diretamente relacionado com a temperatura testada (Bewley & Black, 1985). Para as sementes de Hyoscyamus desertorum, Roth-Bejerano et al. (1971) observaram que a ocorrência de fotoblastismo foi dependente da temperatura testada. A germinação sob ausência de luz alcançou valores acima de 20% nas temperaturas de 25, 30 e 35°C. Entretanto, nas temperaturas de 10, 15 e 20°C, verificou-se a ausência de germinação para a mesma condição de luz. Semelhante ao comportamento observado para as Melastomataceae estudadas no presente trabalho, Rollin & Bidault (1963) verificaram que a germinação de sementes de Lythrum salicaria ocorreu somente quando expostas à luz (fotoperíodo de 24 horas); na ausência de luz, os valores de germinação foram nulos para todas as temperaturas testadas.
Neste experimento, as sementes das duas espécies foram submetidas aos tratamentos 3, 6, 9 e 12 dias de luz (fotoperíodo de 8 horas), sendo mantidas posterionnente sob 20 dias de escuro contínuo.
Os resultados demonstraram que o número mínimo de dias necessários para alcançar a máxima germinação foi distinto para as espécies estudadas (Figura 2). Sementes de Tibouchina moricandiana apresentaram máxima germinação aos doze dias de luz consecutivos, não apresentando diferenças
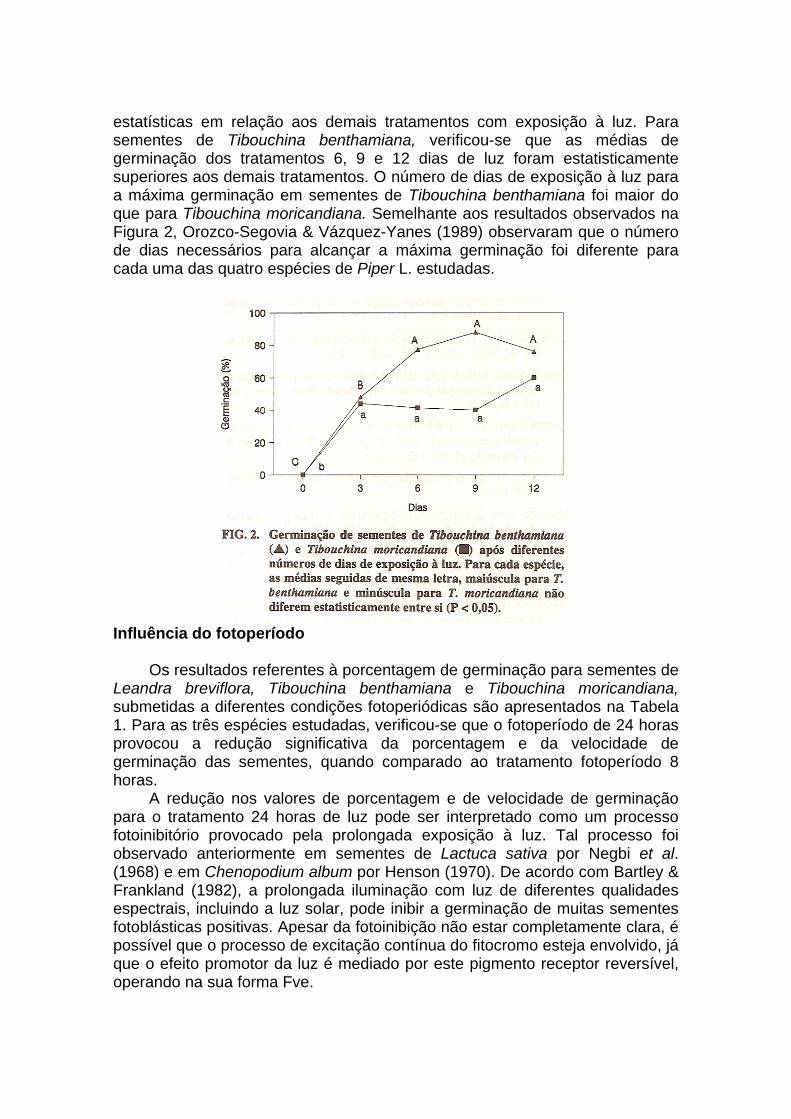
estatísticas em relação aos demais tratamentos com exposição à luz. Para sementes de Tibouchina benthamiana, verificou-se que as médias de germinação dos tratamentos 6, 9 e 12 dias de luz foram estatisticamente superiores aos demais tratamentos. O número de dias de exposição à luz para a máxima germinação em sementes de Tibouchina benthamiana foi maior do que para Tibouchina moricandiana. Semelhante aos resultados observados na Figura 2, Orozco-Segovia & Vázquez-Yanes (1989) observaram que o número de dias necessários para alcançar a máxima germinação foi diferente para cada uma das quatro espécies de Piper L. estudadas.
Influência do fotoperíodo
Os resultados referentes à porcentagem de germinação para sementes de Leandra breviflora, Tibouchina benthamiana e Tibouchina moricandiana, submetidas a diferentes condições fotoperiódicas são apresentados na Tabela 1. Para as três espécies estudadas, verificou-se que o fotoperíodo de 24 horas provocou a redução significativa da porcentagem e da velocidade de germinação das sementes, quando comparado ao tratamento fotoperíodo 8 horas.
A redução nos valores de porcentagem e de velocidade de germinação para o tratamento 24 horas de luz pode ser interpretado como um processo fotoinibitório provocado pela prolongada exposição à luz. Tal processo foi observado anteriormente em sementes de Lactuca sativa por Negbi et al. (1968) e em Chenopodium album por Henson (1970). De acordo com Bartley & Frankland (1982), a prolongada iluminação com luz de diferentes qualidades espectrais, incluindo a luz solar, pode inibir a germinação de muitas sementes fotoblásticas positivas. Apesar da fotoinibição não estar completamente clara, é possível que o processo de excitação contínua do fitocromo esteja envolvido, já que o efeito promotor da luz é mediado por este pigmento receptor reversível, operando na sua forma Fve.

Toole (1973) observou que a provável causa da fotoinibição estaria relacionada à “reação de alta energia”, a qual anularia os efeitos reversíveis do fitocromo. Essa inibição poderia ocorrer através da destruição do Fve abaixo de um nível crítico ou pela contínua remoção do Fve do seu sítio de ação. Górska & Górska (1979), interpretaram a fotoinibição como resultado de interconversões entre Fv e Fve, levando ao acúmulo de formas intermediárias fisiologicamente inativas.
O efeito dos sucessivos ciclos de irradiações diárias foi considerado por Evenari (1965, apud Frankland & Taylorson, 1980) como uma evidência do controle fotoperiódico na germinação de sementes de muitas espécies. Segundo o autor, a germinação pode ser promovida até certo número máximo de horas de luz. Em sementes de Atriplex demarphostegia, altos valores de germinação foram obtidos para fotoperíodos entre 1 e 8 horas, havendo uma redução progressiva naqueles valores com o aumento do fotoperíodo (Koller, 1970). Em contraste, Fujii (1962) observou que a porcentagem de germinação para sementes de Eragrostis ferruginea, submetidas a condições fotoperiódicas, foi inferior à encontrada na condição de luz contínua. De acordo com Labouriau (1983), os padrões das reações descritas para as duas espécies citadas poderiam ser descritos como germinação fotoperiódica de dias curtos e de dias longos, respectivamente.
Desta forma, os resultados da Tabela 1, obtidos para sementes de Leandra breviflora, Tibouchina benthamiana, e Tibouchina moricandiana podem ser interpretados como de um comportamento de germinação fotoperiódica de dias curtos, já que tanto os valores de porcentagem de germinação como os de velocidade de germinação foram superiores para o fotoperíodo de 8 horas, em comparação à condição de luz contínua.
CONCLUSÕES
De acordo com os resultados, foi possível concluir que: - O melhor desempenho germinativo das sementes para as espécies
estudadas foi obtido sob a temperatura de 30°C; em todas as temperaturas testadas, as sementes não germinaram quando mantidas sob ausência de luz, apresentando portanto fotoblastismo positivo.
- O número mínimo de dias de luz para a máxima germinação foi de 12 e 9 dias para Tibouchina moricandiana e Tibouchina benthamiana, respectivaente.
- O fotoperfodo de 24 horas reduziu o desempenho germinativo das sementes de Leandra breviflora, Tibouchina benthamiana e Tibouchina moricandiana, quando comparado ao fotoperíodo de 8 horas, sugerindo a existência de um comportamento fotoperiódico de dias curtos para as espécies estudadas.
AGRADECIMENTOS
O autor agradece à técnica de laboratório Ana Paula Martins Cruz, pelo auxílio na condução dos experimentos, à equipe do Centro de Informações e

Serviços do P.M.A., pela colaboração na confecção dos gráficos, e à SHELL do Brasil S.A., MacArthur Foundation e ao CNPq, pelo apoio financeiro.
REFERÊNCIAS BARROSA, J.M.; BARROSA, L.M.; PINTO, M.M. & AGUIAR, I.B. Efeito do
substrato, temperatura e luminosidade na gerininação de sementes de quaresmeira. Revista Brasileira de Sementes, 10(3):69-77. 1988.
BARTLETT, M.S. The use of transformations. Biometrics, 3:39-52. 1947.
BARTLEY, M.R. & FRANKLAND, B. Analysis of the dual role of phytochrome in the photoinhibition of seed germination. Nature, 300:750-752, 1982.
BAUMGRATZ, J.F.A. Morfologia dos frutos e sementes de Melastomataceas brasileiras. Arquivos do Jardim Botânico do Rio de Janeiro, 27:113-155, 1983-1985.
BEWLEY, J.D. & BLACK, M. Seeds: Phisiology of development and germination. New York: Plenum Press, 1985. 367p.
DAU, L. & LABOURIAU, L.G. Temperature control of seed germination in Pereskia aculeata Mill. An. Acad. Bras. Cien., 46:311-322, 1974.
FRANKLAND, B. & TAYLORSON, R.B. Light control of seed germination. In: SHROPSHIRE, Jr., & MOHR, H. (eds.). Encyclopedia of plant physiology. New Series. Springer-Verlag, 1983. v.16a, p.428-456.
FUJII, T. Studies on photoperiodic responses involved in the germination of Eragrostis seeds. Botanic Magazine, 75:56-62, 1962.
GÓRSKI, T. & GÓRSKA, K. Inhibitory effects of full daylight the germination of Lactuca sativa L. Planta, 144:121-124, 1979.
HEIDECKER, W. Stress and seed germination: as agronomic view. In: KHAN, A. (Eds.). The physiology and biochemistry of seed dormancy and germination. Amsterdam: Elsevier, 1977. p.237-282.
HENDRICKS, S.B. & TAYLORSON, R.B. Variation in the germination and amino acid leakage of seeds with temperature related to membrane phase change. Plant Physiol., 58:7-11, 1976.
HENSON, I.E. The effects of light, potassium nitrate and temperature on the germination of Chenopodium album L. Weed Res., 10:27-39, 1970.
KAGEYAMA, P. & REIS, A. Areas of secondary vegetation in the Itajai Valley Santa Catarina, Brasil: Perspectives for management and conservation. Forest Genetic Resources Information, 21:37-41, 1993.

KLEIN, A. & FELIPPE, G.M. Efeito da luz na germinação de sementes de ervas invasoras. Pesq. Agropec. Bras., 26(7):955-966, 1991.
KOLLER, D. Analysis of the dual action of white light on germination of Atriplex dimorphostegia (Chenopo-diaceae). Israel Journal of Botany, 19:499-516, 1970.
LABOURIAU, L.G. A germinação das sementes. Mongr. Cientif. O.E.A., Washington, U.S.A., 1983. 170p.
LABOURIAU, L.G. & VALADARES, M.B. On the germination of seeds of Calotropis procera. An. Acad. Bras. Cienc., 48:174-186, 1976.
LABOURIAU, L.G. & PACHECO, A. On the frequency of isothermal germination in seeds of Dolichos bilorus L. Plant & Cell. Physiol., 19:507-512, 1978.
LORENZI, H. Árvores brasileiras. São Paulo: Plantarum, 1992. 368p.
MAYER, A.M. & POLJAKOFF-MAYBER, A. The germination of seeds. Oxford: Pergamon Press, 1989. 270p.
NEGBI, M.; BLACK, M. & BEWLEY, J.D. Far-red sensitive dark processes essential for light-and gibberelin-induced germination of lettuce seed. Plant Physiol., 43:35-40, 1968.
OROZCO-SEGOVIA, A. & VAZQUEZ-YANES, C. Light effect on seed germination in Piper L. Oecologia Plantarum, 10(2):123-146, 1989.
OROZCO-SEGOVIA, A. & VAZQUEZ-YANES, C. Los sentidos de las plantas: La sensibilidad de las semillas a la luz. Ciencia, 43:399-411, 1992.
PINTO, M.M.; BARBOSA, J.M. & SILVEIRA, R.B.A. 1983. Contribuição à caracterização cultural de Tibouchina fothergillae Cogn. In: CONGRESSO DA SOCIEDADE BRASILEIRA DE FLORICULTURA E PLANTAS ORNAMENTAIS, 40, Anais... Rio de Janeiro, RJ, p.179-186.
POLLOCK, B.M. & ROSS, E.E. Seed and seedling vigor. In: KOZLOWSKY, T.T., (Ed), Seed biology. New York: Academic Press, 1972, p. 313-387.
POMPEIA, S.L.; PRADELLA, D.Z.A.; MARTINS, S.E.; SANTOS, R.C. & DINIZ, K.M. A semeadura aérea na serra do Mar em Cubatão. Ambiente, 3(1):13-19, 1989.
PROGRAMA MATA ATLÂNTICA, 1990. Relatório final. Rio de Janeiro: Jardim Botânico do Rio de Janeiro. 1990. 220p.
QUEIROZ, M.H. Aspectos preliminares de beneficiamento e germinação de Miconia cinnamomifolia. Silvicultura em São Paulo, 16A(pt.1):318-321, 1982.

RILEY, G.J.P. Effects of high temperature on protein synthesis during germination of maize (Zea mays L.). Planta, 151:75-80, 1981.
ROLLIN, P. & BIDAULT, Y. Etude des facteurs determinant la germination des graines de Lythrum salicaria. Photochem. Photobiol., 2:59-71, 1963.
ROTH-BEJERANO, N.; KOLLER, D. & NEGBI, M. Photocontrol of germination in Hyoscyamus desertorium, a kinetic analysis. Israel Journal of Botany, 20:28-40, 1971.
STEEL, R.G.D. & TORRIE, J.H. Principles and procedures of statistics. A biometrical approach. New York: McGraw Hill, 1980. 633p.
TAKAKI, M.; KENDRIK, R.E. & DIETRICH, S.M.C. Interaction light and temperature on germination of Rumex obtusifolius L. Planta, 152:209-214, 1981.
TOOLE, V.K. Effects of light, temperature and their interactions on. the germination of seeds. Seed Sci. & Technol., 1:339-396, 1973.
TOWNSEND, C.E. & McGINNIES, W.J. Mechanical scarification of Cicer milkvetch (Astragalus cicer L.) seed. Crop Science, 12:392-394, 1972.
VIDAVER, W. & HSIAO, A,I. Secondary dormancy in light sensitive lettuce seeds incubated anaerobically or at elevated temperature. Can. J. Bot., 53: 2557-2560, 1975.
WULFF, R. & MEDINA, E. Germination of seeds in Jussiaea suffruticosa. Plant Cell & Physiology, 10:503-511, 1969.




















