
L’hybridation génomique comparative ouvre de nouvelles perspectives dans l’exploration des...
11
L'hybridation g( nomique comparative ouvre de nouvelles perspectives dans I'exploration des modifications qui accompagnent les I( sions (pr( -)malignes du tube digestif G. A MEIJER, M. AJA HERMSEN, S. G. MEUWISSEN, J. PA BAAK Department of Pathology and Gastroenterology, Free University Hospital, Amsterdam Comparative Genomic Hybridization opens up new perspectives in the research of genetic changes underlying (pre-)malignant lesions of the gastrointestinal tract RI~SUME L'hybridation g~nomique compar6e (HGC) est une technique developp4e r6cemment qui permet, en une seule analyse, la d6tection d'aberrations g6n6tiques pour la totalit6 du g6nome de la tumeur sans n6cessiter de culture cellulaire; elle peut s'appliquer/~ du mat6riel conserve pour archive. Elle procure une vision globale du gain ou de la perte d'ADN dans une tumeur en comparaison ~ de I'ADN eontr61e. Les pertes peuvent correspondre ~ des gOnes suppresseurs de tumeurs connus ou non et les gains ~ l'amplification d'oncogOnes, les uns et les autres impliqu6s dans la pathog~n~se du cancer colorectal. Au cours de cette proc6dure. I'ADN test obtenu ~ partir d'6chantillons de tumeurs est marqu6 avec un fluorochrome vert et m61ang6 avec de I'ADN contr61e marqu6 en rouge et obtenu '~ partir de cellules avec un contenu chromosomique diplo~de normal. Cette mixture est hybrid6e ~ des pr6parations m6taphasiques normales. L'hybridation des s6quenees d'ADN test ou contr61e, qui rentrent en comp6tition lors de l'appariement, est reconnue grace aux diff6rents fluoroehromes. Le ratio de fluorescence verte par rapport ~ rouge pour chaque r6gion chromosomique, obtenu par analyse d'image digitale, est la mesure de la sous-ou sur-repr6sentation de mat6riel g6nOtique pour eette r6gion de la tumeur 6tudi6e. Compar6e ~ d'autres techniques, I'HGC parvient ~ une information plus sp6cifique que, par exemple, l'analyse de I'ADN en cytom& trie de flux, et/~ une meilleure vision globale des modifications g6n6tiques au sein d'une tumeur qu'avec I'hybridation in situ ou Fanalyse de perte d'h6t6rozygotie. Les applications de l'hybridation g6nomique comparative sont multiples, par exemple pour le criblage d'aberrations chromosomiques dans une large sOrie de tumeurs destinO "~ identifier des loci chromosomiques qui renferment possiblement de nouveaux g~nes suppresseurs de tumeurs ou de nouveaux oncogbnes, pour l'analyse g6n6tique des modifications au tours des stades successifs de la carcinogOn~se, ou pour 61ucider les bases g6n6tiques des caract6ristiques cliniques et ph6notypiques de certaines tumeurs. SUMMARY Comparative Genomic Hybridization (CGH) is a recently developed technique that allows to detect in one analysis genetic aberrations in the complete tumour genome without the need of cell culturing, and it can be applied to archival material. B pro rides an overview of chromo- somal losses and gains of DNA in a tumour as compared to control DNA. Losses can point to known and putative tumour suppressor genes and gains to amplification of oncogenes, involved in the pathogenesis of colorectal cancer. In this procedure, test DNA obtained from tumour speci- mens is labelled with a green fluorochrome and mixed with red labelled control DNA obtained from cells with a normal diploid chromosome complement. This mixture is hybridized to normal metaphase preparations. Hybridized test and control DNA sequences, which compete in the annealing process, are recognized via the different fluorochromes. The ratio of green to red fluorescence for each chromosomal region, analysed by digital image processing, is a measure of the under- or over-representation of genetic material f~r that region in the tumour studied. Compared to other techniques, CGH yields more specific information than e.g. DNA cytometry, and a better overview of genetic changes in a tumour than is possible with in situ hybridization or loss of heterozygosity analysis. The applications of comparative genomic hybridization are multiple, e.g. to screen for chromosomal aberrations in large series of tumours aiming to identify chromosomal loci that possibly harbour new tumour suppressor genes or oncogenes, to analyse genetic changes in subsequent stages of carcinogenesis, or to elucidate the genetic basis of clinical and phenotypical tumour characteristics. INTROD UCTION Le d6veloppement de tumeurs solides est associ6 l'acquisition et h l'accumulation de modifications g6n6tiques [1-3]. L'analyse des anomalies gdn6tiques dans des 6chantillons de tumeurs devient un nouvel aspect important de la pathologie tumorale, qui aura une influence croissante sur la prise de d6cision m6di- cale [4, 5]. La tendance en pathologie est de viser le plus haut degr6 d'information diagnostique et pro- nostique, aussi t6t que possible pendant la pdriode de prise de d4cision diagnostique, et d'accomplir cette tSche avec un minimum de m6thodes invasives. L'analyse gdn6tique peut se faire de multiples fa~ons, partir de techniques relativement simples comme l'analyse de I'ADN en cytom6trie de flux jusqu'h des TirEs ~ part: Corresponding author: Prof.Dr. J.P.A. Baak, Dept. of Pathology, Frec University Hospital, P.O. Box 7057, 1007 MB Amsterdam. Mots-clOs : chromosomes, Hybridation g6nomique comparative, hybridation in situ. Key-words: chromosomes, Comparative genomic hybridization, in situ hybridization. Acta Endoscopica Volume 27 - N ~2 - 1997 89
Transcript of L’hybridation génomique comparative ouvre de nouvelles perspectives dans l’exploration des...

L'hybridation g( nomique comparative ouvre de nouvelles perspectives dans I'exploration des modifications
qui accompagnent les I( sions (pr( -)malignes du tube digestif
G. A MEIJER, M. AJA HERMSEN, S. G. MEUWISSEN, J. PA BAAK
Department of Pathology and Gastroenterology, Free University Hospital, Amsterdam
Comparative Genomic Hybridization opens up new perspectives in the research of genetic changes underlying (pre-)malignant lesions of the gastrointestinal tract
RI~SUME
L'hybridation g~nomique compar6e (HGC) est une technique developp4e r6cemment qui permet, en une seule analyse, la d6tection d'aberrations g6n6tiques pour la totalit6 du g6nome de la tumeur sans n6cessiter de culture cellulaire; elle peut s'appliquer/~ du mat6riel conserve pour archive. Elle procure une vision globale du gain ou de la perte d 'ADN dans une tumeur en comparaison ~ de I 'ADN eontr61e. Les pertes peuvent correspondre ~ des gOnes suppresseurs de tumeurs connus ou non et les gains ~ l'amplification d'oncogOnes, les uns et les autres impliqu6s dans la pathog~n~se du cancer colorectal. Au cours de cette proc6dure. I 'ADN test obtenu ~ partir d'6chantillons de tumeurs est marqu6 avec un fluorochrome vert et m61ang6 avec de I 'ADN contr61e marqu6 en rouge et obtenu '~ partir de cellules avec un contenu chromosomique diplo~de normal. Cette mixture est hybrid6e ~ des pr6parations m6taphasiques normales. L'hybridation des s6quenees d 'ADN test ou contr61e, qui rentrent en comp6tition lors de l'appariement, est reconnue grace aux diff6rents fluoroehromes. Le ratio de fluorescence verte par rapport ~ rouge pour chaque r6gion chromosomique, obtenu par analyse d'image digitale, est la mesure de la sous-ou sur-repr6sentation de mat6riel g6nOtique pour eette r6gion de la tumeur 6tudi6e.
Compar6e ~ d'autres techniques, I 'HGC parvient ~ une information plus sp6cifique que, par exemple, l'analyse de I'ADN en cytom& trie de flux, et/~ une meilleure vision globale des modifications g6n6tiques au sein d'une tumeur qu'avec I'hybridation in situ ou Fanalyse de perte d'h6t6rozygotie. Les applications de l'hybridation g6nomique comparative sont multiples, par exemple pour le criblage d'aberrations chromosomiques dans une large sOrie de tumeurs destinO "~ identifier des loci chromosomiques qui renferment possiblement de nouveaux g~nes suppresseurs de tumeurs ou de nouveaux oncogbnes, pour l'analyse g6n6tique des modifications au tours des stades successifs de la carcinogOn~se, ou pour 61ucider les bases g6n6tiques des caract6ristiques cliniques et ph6notypiques de certaines tumeurs.
S U M M A R Y
Comparative Genomic Hybridization (CGH) is a recently developed technique that allows to detect in one analysis genetic aberrations in the complete tumour genome without the need o f cell culturing, and it can be applied to archival material. B pro rides an overview of chromo- somal losses and gains o f DNA in a tumour as compared to control DNA. Losses can point to known and putative tumour suppressor genes and gains to amplification o f oncogenes, involved in the pathogenesis o f colorectal cancer. In this procedure, test DNA obtained from tumour speci- mens is labelled with a green fluorochrome and mixed with red labelled control DNA obtained from cells with a normal diploid chromosome complement. This mixture is hybridized to normal metaphase preparations. Hybridized test and control DNA sequences, which compete in the annealing process, are recognized via the different fluorochromes. The ratio o f green to red fluorescence for each chromosomal region, analysed by digital image processing, is a measure o f the under- or over-representation o f genetic material f~r that region in the tumour studied.
Compared to other techniques, CGH yields more specific information than e.g. DNA cytometry, and a better overview of genetic changes in a tumour than is possible with in situ hybridization or loss o f heterozygosity analysis. The applications o f comparative genomic hybridization are multiple, e.g. to screen for chromosomal aberrations in large series o f tumours aiming to identify chromosomal loci that possibly harbour new tumour suppressor genes or oncogenes, to analyse genetic changes in subsequent stages o f carcinogenesis, or to elucidate the genetic basis o f clinical and phenotypical tumour characteristics.
INTROD UCTION
Le d6veloppement de tumeurs solides est associ6 l'acquisition et h l'accumulation de modifications g6n6tiques [1-3]. L'analyse des anomalies gdn6tiques dans des 6chantillons de tumeurs devient un nouvel aspect important de la pathologie tumorale, qui aura une influence croissante sur la prise de d6cision m6di-
cale [4, 5]. La tendance en pathologie est de viser le plus haut degr6 d'information diagnostique et pro- nostique, aussi t6t que possible pendant la pdriode de prise de d4cision diagnostique, et d'accomplir cette tSche avec un minimum de m6thodes invasives. L'analyse gdn6tique peut se faire de multiples fa~ons,
partir de techniques relativement simples comme l'analyse de I'ADN en cytom6trie de flux jusqu'h des
TirEs ~ part: Corresponding author: Prof.Dr. J.P.A. Baak, Dept. of Pathology, Frec University Hospital, P.O. Box 7057, 1007 MB Amsterdam.
Mots-clOs : chromosomes, Hybridation g6nomique comparative, hybridation in situ.
Key-words: chromosomes, Comparative genomic hybridization, in situ hybridization.
Acta Endoscopica Volume 27 - N ~ 2 - 1997 89

techniques tr6s d6taill6es comme la cytog6n6tique des tumeurs, l'hybridation in situ (HIS) ou l'6tude de perte d'h6t6rozygotie (PH) par PCR.
Rdcemment, une nouvelle m6thode d'hybridation g6nomique comparative (HGC) a 6t6 d6crite, qui est capable d'une analyse h la lois d6taill6e et extensive des changements g6n6tiques pour l 'ensemble du g6nome de tumeurs [6, 7]. Elle procure une informa- tion sur les pertes et gains ou amplifications chromo- somiques en une seule expdrience. Dans ce qui suit, nous allons pr6senter une description d6taill6e de toutes les 6tapes de la technique d 'HGC et discuter les avantages et inconv6nients de la m6thode elle- m6me puis montrer plusieurs de ses applications.
D~tails techniques de I'HGC
Une vue sch6matique de la technique est montr6e en figure 1. Premi6rement, I 'ADN de la tumeur est marqu6 avec un fluorochrome vert et de I 'ADN contrfle normal (diplmde, ne provenant pas n6cessai- rement du m~me patient) est marqu6 avec un fluoro- chrome rouge. Ces ADN marqu6s sont m61ang6s et hybrid6s h des pr6parations de m6taphases humaines normales. Les ADN marqu6s en rouge et en vert ren- trent en comp6tition pour s'hybrider h ces chromo- somes. En utilisant la microscopie fluorescente et l'analyse d'image digitale, le ratio de fluorescence verte sur rouge est calcul6 pour chaque chromosome. Ce ratio est une mesure de la sous- ou sur-repr6sen- tation de mat6riel g6n6tique pour cette r6gion de la tumeur 6tudi6e. Le processus complet d'analyse d'une tumeur par HGC suit les 6tapes suivantes.
PREPARA TION D'UNE M[~TA PHASE NORMAL E
Des lames de m6taphases normales sont pr6par6es partir de cultures de lymphocytes circulants d'un
donneur sain stimul6es ?a la phytoh6maglutinine. Les cellules sont arr~t6es en mitose avec de la colchicine, r6colt6es et trait6es au KC1 hypotonique puis fix6es au m6thanol/acide ac6tique. Les cellules sont ensuite mont6es sur des lames de mani~re ?ace que les chro- mosomes des cellules mitotiques soient convenable- ment 6tal6s.
ISOLATION D'ADN DE TUMEUR
A partir d'un minimum d'environ 200 000 cellules ou approximativement 1 mm 3 de tissu, I 'ADN de tumeur peut ~tre obtenu par n'importe quelle tech- nique d'isolation d 'ADN qui permet d'obtenir de I'ADN de haut poids moleculaire, requis pour l'utilisa- tion en HGC. Aussi I'ADN extrait de mat6riel inclus en paraffine, qui peut ~tre partiellement degrad6, sera utilis6 avec des ajustements au cours du marquage. De tr~s petites quantit6s d 'ADN peuvent ~tre utilis6es en combinaison avec une r6action de PCR amorc6e avec des oligonucl6otides d6g6n6r6s (PCR-OD). L 'ADN normal utilis6 en r6f6rence peut 6tre obtenu ~a partir des lymphocytes circulants d'un individu sain.
Figure 1 Vue schematique de la technique d'HGC. L'ADN de la tumeur et celui du
contr61e sont marques avec des fluorochromes vert et rouge, respectivement, et hybrid~s & un etalement de metaphases normales
(voir aussi fig. 2b). Les images correspondant aux signaux fluorescents sont acquises et les ratios du signal vert sur rouge sont quantifies digitalement le long de I'axe du chromosome, et ce pour chaque
chromosome (reproduit a partir de la reference 5).
MARQUAGE DE L 'ADN
Une m6thode standard de marquage (par nick- translation) est utilis6e pour marquer I 'ADN puis pour dig6rer I 'ADN en fragments d'une taille opti- male de 600-2000 paires de bases. Une petite quantit6 d 'ADN - - entre 0,5 et 1 lag - - est suffisante. Dans le cas de la PCR-OD, le marquage peut se faire directe- ment pendant l'6tape d'amplification. Jusqu'h pr6- sent, on incorporait surtout des deoxynucl6otides conjugu6s h la biotine ou h la digoxygenine au cours du marquage, ce qui n6cessite une 6tape de d6tection apr6s l'hybridation ~ l'aide de ligands fluorescents (avidin-FITC et anticorps de mouton anti-digoxyge- nine-TRITC, respectivement). I1 est aussi possible d'utiliser des deoxynucl6otides directement conju- gu6s hun fluorochrome, ce qui donne un signal d'hy- bridation plus net - - mais aussi plus faible - - sur les chromosomes. I1 est important que les fragments d 'ADN marqu6s provenant de la tumeur et du contr61e aient la m6me taille et soient dans l'inter- valle 600-2000 paires de bases.
HYBRIDA TION
Approximativement 150-200 ng D 'ADN marqu6 de la tumeur et du contr61e (sondes) sont m61ang6s avec une quantit6 100 fois sup6rieure d 'ADN Cot-1 non marqu6, pr6cipit6 et resuspendu dans 10 ml du m61ange d'hybridation contenant 50 % de formamide et 10 % de dextrane sulfate. L 'ADN Cot-1 consiste en des s6quences r6p6titives purifi6es et est ajout6 pour bloquer les s6quences r6p6titives hautement polymorphiques de I'ADN. La sonde et les lames de m6taphases normales sont d6natur6es ~ part. L'hybri- dation se d6roule dans un incubateur humide ~ 37 ~ pour 2-4 jours. Apr~s l'hybridation, les lames sont lav6es et color6es au 4,6-diamidino-2-phenylindole (DAPI) pour faire apparaitre les bandes chromo-
90 Volume 27- N~ 2 - 1997 Acta Endoscopica
somiques qui permettent l'identification des chromo- somes et l'6tablissement du caryotype standard.
A CQ UISITION D'IMA GE E T A N A L YSE
En microscopie fluorescente, en utilisant trois pas- sages successifs avec des filtres diffdrents (pour DAPI, FITC et TRITC, respectivement) en combi- naison avec une cam6ra fi enregistrement digital, trois images digitales en noir et blanc sont acquises et chaque mdtaphase est reconstitude sur un 6cran d'or- dinateur (fig. 2b). L'analyse d'image pour I'HGC a 6t6 revue en ddtail ailleurs [8, 9]. Un programme sp6- cial permet le caryotypage interactif des chromo- somes et le calcul du ratio de fluorescence verte sur rouge pour chaque chromosome. Lc ratio moyen pour plusieurs mdtaphases s61ectionndes pour leur qualit6, de m~me que l'intervalle de confiance fi 95 %, sont indiquds ~ c8t6 des iddogrammes repr6sentant
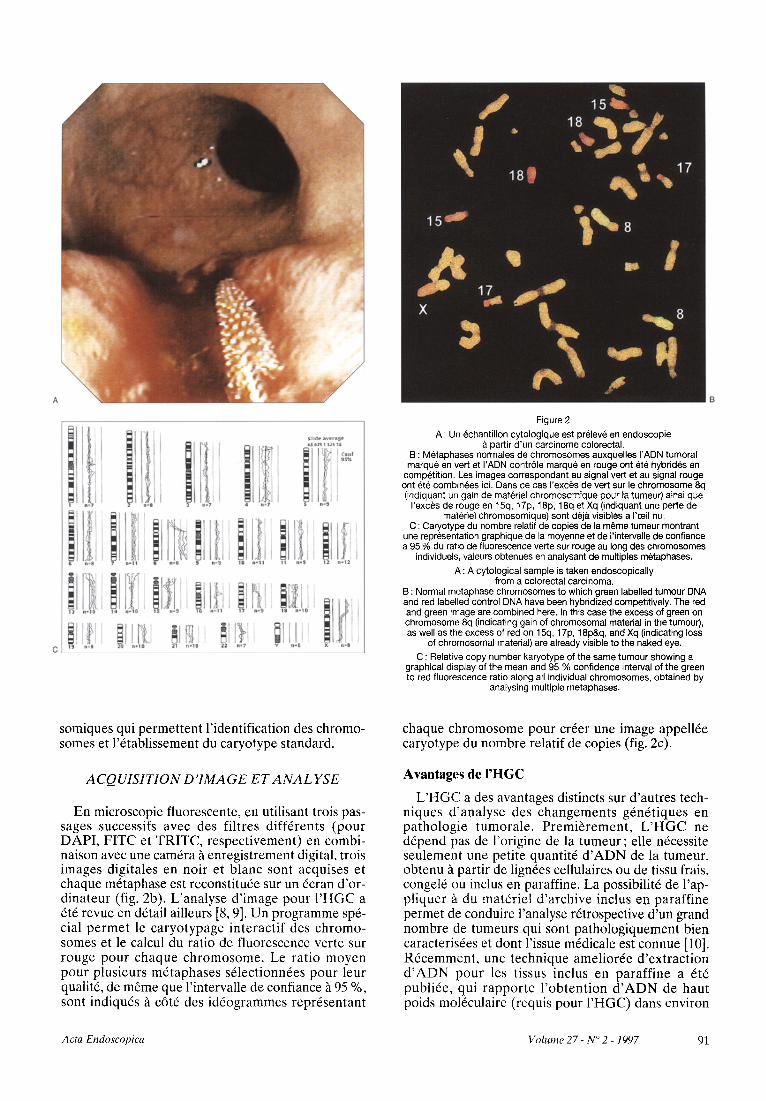
Figure 2
A : Un echantillon cytologique est prelev~ en endoscopie partir d'un carcinome colorectal.
B : Metaphases normales de chromosomes auxquelles I'ADN tumoral marque en vert et I'ADN contrSle marqu~ en rouge ont ere hybrid6s en
competition. Les images correspondant au signal vert et au signal rouge ont ete combinees icL Dans ce cas I'exces de vert sur le chromosome 8q (indiquant un gair~ de materiel chromosomique pour 1a tumeur) ainsi que
I'exces de rouge en 15q, 17p, 18p, 18q et Xq (indiquant une perte de materiel chromosomique) sont d6ja visibles a I'aeil nu.
C : Caryotype du nombre relatif de copies de la m~me tumeur montrant une representation graphique de la moyenne et de I'intervalle de confiance a 95 % du ratio de fluorescence verte sur rouge au long des chromosomes
individuels, valeurs obtenues en analysant de multiples metaphases.
A : A cytological sample is taken endoscopically from a colorectal carcinoma.
B : Normal metaphase chromosomes to which green labelled tumour DNA and red labelled control DNA have been hybridized competitively. The red and green image are combined here. In this case the excess of green on chromosome 8q (indicating gain of chromosomal material in the tumour), as well as the excess of red on 15q, 17p, 18p&q, and Xq (indicating loss
of chromosomal material) are already visible to the naked eye.
C : Relative copy number karyotype of the same tumour showing a graphical display of the mean and 95 % confidence interval of the green to red fluorescence ratio along all individual chromosomes, obtained by
analysing multiple metaphases.
chaque chromosome pour crder une image appell6e caryotype du nombre relatif de copies (fig. 2c).
Avantages de I'HGC
L'HGC a des avantages distincts sur d'autres tech- niques d'analyse des changements g6n6tiques en pathologie tumorale. Premi6rement, L 'HGC ne d6pend pas de l'origine de la tumeur; elle n6cessite seulement une petite quantit6 d 'ADN de la tumeur, obtenu ~ partir de ligndes cellulaires ou de tissu frais, congel6 ou inclus en paraffine. La possibilit6 de l'ap- pliquer ~t du mat6riel d'archive inclus en paraffine permet de conduire l'analyse r6trospective d'un grand nombre de tumeurs qui sont pathologiquement bien caracteris6es et dont l'issue m6dicale est connue [10]. R6cemment, une technique amelior6e d'extraction d 'ADN pour les tissus inclus en paraffine a 6t6 publi6e, qui rapporte l 'obtention d 'ADN de haut poids mol~culaire (requis pour I'HGC) dans environ
A c t a E n d o s c o p i c a V o l u m e 2 7 - N ~ 2 - 1997 91

80 % des cas [11]. Dans les cas ou la quantit6 d 'ADN obtenue/~ partir de tissus inclus en paraffine est trop faible (moins que 1-2 pg), I 'HGC est encore possible grhce ~ une 6tape d'amplification globale par PCR (PCR-OD). Avec cette modification, un minimum d'environ 2 000 cellules est suffisant. Ce nombre de 2 000 cellules peut 6tre facilement obtenu juste en grattant la section tissulaire. Si le mat6riel a 6t6 fix6 darts de la formaline tamponn6e, un nombre encore inf6rieur (10-30 cellules) peut ~tre suffisant [12]. L 'HGC ?a partir d 'ADN amplifi6 par PCR s'est av6r6 ~tre une m6thode reproductible e t a montr6 les m6mes anomalies cytog6n6tiques qu'avec de I 'ADN de la m6me tumeur, isol6 ~ partir d'un 6chantillon congel6 [10]. Deuxi6mement, I 'HGC est capable de fournir une information d6taill6e sur les gains et pertes chromosomiques et de le faire pour tout le g6nome en une seule exp6rience. La technique d 'HGC peut 6tre sp6cialement indiqu6e dans l'ana- lyse des changements g6n6tiques qui accompagnent les tumeurs solides, qui sont relativement inacces- sibles aux techniques conventionnelles d'analyse cytog6n6tique parce qu'il est tr6s difficile d'obtenir des cultures cellulaires a partir de ces tumeurs.
Limitations de I'HGC
L 'HGC ne peut pas ddtecter des translocations chromosomiques 6quilibrdes, qui sont fr6quemment retrouv6es dans des tumeurs h6matologiques et m6senchymateuses et ~un bien moindre degr6 dans des tumeurs solides 6galement. Le criblage de ces anomalies ne peut 6tre r6alis6 que par caryotypage (spectral) apres culture des cellules tumorales. De plus, les d616tions submicroscopiques ne seront pas apparentes en HGC. Th6oriquement, la limite infd- rieure de d6tection est d6termin6e par le produit du nombre de copies d'un g6ne par la taille du fragment de g6ne concern6. On estime qu'une amplification du nombre de copies de 50 % doit 6tre d6tectable si la r6gion mesure 10 Mbase ou plus, et qu'une r6gion de 250 Kbase sera d6tectde si le nombre de copies est augment6 de 400 % [13]. La limite de d6tection pour les d616tions est sup6rieure, car la perte maximum ne peut exc6der 100 %. Dans notre exp6rience, une perte de 10Mbase 6tait encore ddtectable [14]. La contami- nation d'un 6chantillon tumoral par des cellules nor- males (i.e. cellules stromales, lymphocytes) dimi- nuera la sensibilit6 de l 'analyse par HGC. Kallioniemi et al. et d'autres ont trouv6 que 50 % de cellules non tumorales dans l'6chantillon test consti- tuait la limite de d6tection des monosomies ou triso- mies [10, 15]. La contamination de l'6chantillon test par des cellules non tumorales peut 6tre r6duite par microdissection des tissus histologiquement concer- n6s. En outre, les r6gions p6ricentrom6riques et t61o- m6riques sont exclues de l'analyse. Ces r6gions contiennent des s6quences rdpdtitives polymor- phiques et sont donc bloqu6es par I 'ADN Cot-1 non marqu6. Finalement, les profils de ratio pour les rdgions correspondant/a l'extr6mit6 du chromosome 1 et le bras court des chromosomes 16, 19 et 22 peuvent varier et doivent donc ~tre interpr6t6s avec pr6cau- tion [8, 15].
Validation de la technique d'HGC
La technique d 'HGC a 6t6 valid6e de plusieurs faqons. Premi6rement, par l'analyse d 'ADN normal et d 'ADN tumoral pr6sentant des gains ou des pertes connus. Deuxi6mement en comparant les r6sultats de I 'HGC avec d'autres techniques comme les analyses cytog6n6tiques classiques et l'analyse de perte d'h6t6rozygotie. A partir de ces exp6riences, on peut conclure que I 'HGC ne produit pas de faux n6gatifs ou positifs.
L 'HGC comparant deux 6chantillons d 'ADN nor- mal a montr6 des profils un6quivoques avec des inter- valles de confiance qui contenaient le ratio 1 pour tousles chromosomes, h l'exception des r6gions cen- trom6riques et t61om6riques [8, 15]. L'augmentation du nombre de copies de chromosome s'est r6vel6e corr61er le ratio moyen vert sur rouge [6]. Cependant, I 'HGC ne devrait pas ~tre utilis6e pour quantifier le nombre de copies en gain ou en perte car cette corr6- lation n'est pas lin6aire.
Une 6tude par notre groupe, destin6e a comparer les r6sultats de l'analyse HGC sur six cultures cellulaires de carcinomes oraux a cellules squameuses par rapport
ceux obtenus par l'analyse caryotypique, a montr6 que les modifications g6n6tiques, en particulier les pertes, 6taient plus fid~lement d6tect6es par HGC que par caryotypage [16]. Dans les cas de diff6rences notoires entre I'HGC et l'analyse cytog6n6tique, l'ana- lyse par hybridiation in situ fluorescente (FISH) en uti- lisant des chromosomes entiers a confirm6 les r6sultats de I'HGC. En g6n6ral, l'analyse par FISH montrait que la pr6sence de chromosomes inidentifiables en cytog6n6tique, et appel6s marqueurs, dans les caryo- types de cellules cultiv6es 6tait la cause la plus pro- bable de ces diff6rences. Cela signifie que, si il est fond6 sur l'analyse cytog6n6tique seule, le nombre de pertes chromosomiques est g6n6ralement surestim6. Les r6sultats de I'HGC en ce qui concerne les pertes de mat6riel chromosomique ont aussi 6t6 compar6s aux r6sultats obtenus par l'analyse de perte d'h6t6rozygo- tie (PH). Bien que la PH d6tecte la perte d'un all61e alors que I'HGC d6tecte la perte relative pour un chro- mosome compar6 au contenu d 'ADN total ou plo~die, les deux techniques produisent des r6sultats remarqua- blement similaires. Voorter et al. ont rapport6 100 % de PH h des loci oil I 'HGC avait d6tect6 des pertes chromosomiques dans des tumeurs de la vessie [17]. D'un autre c6t6, seulement 50 % des loci PH 6taient identifi6s par HGC. I1 existe deux explications pos- sibles pour ce manque apparent de sensibilit6. Soit la d616tion est trop petite pour que I 'HGC puisse la d6tecter, soit une disomie uniparentale (c'est-a-dire la perte d'un chromosome suivi par la duplication du chromosome restant) s'est produite dans cette r6gion. Dans une autre 6tude, l'analyse par PH et HGC de quatre lign6es de k6ratinocytes s6par6ment transfec- t6es avec HPV a montr6 que seulement environ 50 % des pertes d6tect6es par HGC 6taient aussi d6tect6es par PH. Ceci pourrait ~tre dfi h la perte de seulement un des trois chromosomes dans ces lign6es cellulaires triploides, laissant deux h6t6rozygotes homologues. D'un autre c6t6, 100 % des loci qui montraient une PH 6taient aussi identifi6s en perte par l'analyse HGC
92 Volume 27- N~ 2 - 1997 Acta Endoscopica

(Steenbergen, RDM et al., r6sultats non publi6s). Kal- lioniemi et al. et Sakamoto et a/.ont rapport6 une concordance entre HGC et PH pour environ 80 % des cancers du sein et de l'ovaire, respectivement [18, 19]. En conclusion, PH et HGC peuvent ~tre consid6r6es comme des techniques compl6mentaires, ayant cha- curie leurs propres possibilit6s d'application.
Applications de I'HGC
Les applications de I'HGC sont multiples. L'HGC a 6t6 utilis6e pour cribler des nombres 61ev6s de g6nomes tumoraux afin de d6tecter des r6gions chro- mosomiques pr6sentant r6guli6rement une amplifica- tion ou une d616tion de g6nes dans des types de tumeurs spdcifiques. D6j~, le nombre de tumeurs solides analys6es par HGC a d6pass6 celui des tumeurs solides analys6es en cytogdn6tique classique. Au cours des trois dernibres ann6es, un vaste assortiment de types tumoraux, comprenant m61anomes, cancers du poumon, du colon, de la t6te et du cou, de la prostate, de l'ovaire, du rein et de la vessie, ost6osarcomes, gliomes, tumeurs hdmatologiques et des tissus mous, ont 6t6 cribl6s par HGC pour la recherche de perte ou gain chromosomiques [17, 19-30].
La base g6n6tique du d6veloppement tumoral peut ~tre recherch6e en analysant diffdrentes 6tapes de la maladie ( par exemple les 16sions pr6-malignes, le cancer invasif et les mdtastases). Dans cette voie, I 'HGC peut servir de premi6re ligne d'analyse en g6n6tique des tumeurs pour d6tecter globalement des r6gions chromosomiques intdressantes qui peuvent 6tre ensuite etudi6es plus en ddtail grfice ~ d'autres techniques de g6n6tique mol6culaire. Dans le cas du cancer du sein, l'analyse par HGC a conduit ~a l'iden- tification de nouvelles r6gions chromosomiques qui sont possiblement impliqu6es dans la pathog6n6se du cancer du sein et le ddveloppement de ses mdtastases. Dans une s6rie de 33 tumeurs primaires invasives, Kallioniemi et al. ont trouv6, en plus des gains chro- mosomiques en lq et 8q d6j~ d6crits, des gains rdcur- rents en 17q et 20q [28]. La position du gain en 20q a 6t6 d6finie plus pr6cis6ment au niveau de la bande 20q13 ; cependant, la recherche de possibles onco- g6nes ~ cette position par des techniques de gdnd- tique mol6culaire n'a pas encore abouti [31]. Tou- jours dans le cancer du seim en comparant des tumeurs primaires et leurs distantes mdtastases, Kuu- kasjg~rvi et al. ont trouv6 qu'un gain en Xq dtait une anomalie acquise au niveau des mdtastases, indiquant un possible r61e pour cette r6gion dans le d6veloppe- ment des m6tastases [32].
En outre, I 'HGC peut aider ~ rdv61er des m6ca- nismes biologiques responsables de certaines caract6- ristiques d6veloppdes in v i vo comme la non-rdponse
la thdrapie et l'h6t6rog6ndite gdn6tique. L'analyse par HGC de tumeurs prostatiques r6currentes chez des patients qui 6taient trait6s ~ l'aide d'une th6rapie androgdno-suppressive a r6v616 l'amplification de la r6gion chromosomique Xqll-12 dans 55 % des cas, alors qu'aucune amplification n'6tait d6tect6e dans les tumeurs primaires. La bande chromosomique Xql l contient le gbne codant pour le r6cepteur pour l'androgbne (RA). En utilisant RA comme sonde en
FISH et en hybridation in si tu d'ARNm, il a 6t6 mon- tr6 que RA 6tait amplifi6 et surexprim6 dans 28 % des cas. Ces r6sultats indiquent que l'6chec de la th6rapie androg6no-suppressive est dfi h l'expansion clonale de cellules tumorales capables d 'augmenter leur expression du r6cepteur pour l'androg6ne [33].
L'existence d'une h6t6rog6n6it6 intra-tumorale au niveau ph6notypique 6tait d6j~a bien connue des patho- logistes, et ce ph6nom6ne a 6t6 d6montr6 par analyse de I'ADN en cytom6trie de flux [34-36]. Nous avons pu d6montrer cette h6t6rog6n6it6 au niveau chromoso- mique par analyse HGC d'6chantillons pr61ev6s/~ dif- f6rents endroits du m6me carcinome colorectal. Nous avons trouv6 que certaines pertes et gains 6taient pr6- sents de m6me mani6re dans les deux 6chantillons alors que dans d'autres cas ces anomalies 6taient bien moins apparentes voire m~me insignifiantes dans un des 6chantillons compar6 h l'autre [30]. Ceci indique peut-6tre que les gains et pertes communs aux deux 6chantillons repr6sentent des changements pr6coces alors que les diff6rences entre les 6chantillons t6moi- gnent de changements tardifs. Ceci ouvre la possibilit6 d'analyser la base g6n6tique des diff6rences ph6noty- piques au sein d'une m6me tumeur.
Finalement, la prise de d6cision clinique pourrait tirer b6n6fice de la recherche par HGC. La possibilit6 d'utiliser I'HGC pour du mat6riel d'archive inclus en paraffine (si besoin en combinaison avec la PCR- OD) augmentera le nombre d'analyses importantes pour l'aspect clinique. A partir de l'id6e que des modifications g6n6tiques influencent le comporte- ment des tumeurs, certains pathologistes et cliniciens sont d6j~ habitu6s ~a prendre des d6cisions en s'ap- puyant sur des histogrammes d 'ADN dans le cas de certains types de tumeurs. Ces histogrammes r6v61ent s'il y a ou non une modification du contenu d'ADN par cellule dans une tumeur. C'est bien stir une m6thode plut6t grossi6re d'analyser des modifica- tions de I'ADN. L'HGC peut montrer sur quels chro- mosomes, et ou sur ces chromosomes, des pertes ou gains d 'ADN sont pr6sents et pourrait donc 6tre d'une utilit6 clinique bien plus importante que l'ana- lyse de ploi'die de I'ADN. On a m6me montr6 qu'une augmentation de l'instabilit6 g6n6tique, illustr6e par le nombre d'alt6rations g6n6tiques, 6tait en corr61a- tion avec un pronostic n6gatif et/ou avec certaines caract6ristiques clinicopathologiques des tumeurs, comme par exemple dans le cancer du colon et du sein [37, 38]. L'utilisation de I'HGC pour du mat6riel d'archive inclus en paraffine pour lequel un suivi long terme est disponible permet une large 6tude l'6chelle du g6nome de l'importance clinique d'aber- rations chromosomiques sp6cifiques [39]. En 6va- luant les changements qui sont associ6s, par exemple,
la transition du stade pr6-malin au stade malin, et en corr61ant les r6sultats de I 'HGC au pronostic, I 'HGC peut servir de base ~ la classification de la maladie et ?a la pr6vision de son issue clinique [4, 5].
Utilisation de I'HGC pour les tumeurs du tube digestif
Le nombre des 6tudes appliquant la technique d 'HGC aux cancers et aux lesions pr6-malignes du
A c t a E n d o s c o p i c a V o l u m e 2 7 - N ~ 2 - 1997 93

tube digestif ne cesse d'augmenter. Dans une 6tude r6cente, nous avons analys6 des 6couvillonages pris ?a la surface de 16 tumeurs colorectales, soit ?a partir d'6chantillons frais obtenus apr6s ressection du tissu canc6reux, soit in situ au cours de l'endoscopie (Fig. 2a) [30]. Dans les quatre ad6nomes analys6s, une moyenne de 6,8 (entre 4 et 10) modifications g6n6tiques furent identifi6es. Dans les 12 carcinomes une moyenne de 11,1 (entre 5 et 17) 6v6nements furent trouvds par tumeur. A la fois dans les ad6- nomes et les carcinomes, les gains 6taient plus frd- quents que les pertes. Les gains les plus frdquents cor- respondaient ~ 13q, 7p et 7q, 8q et 20q, alors que les pertes se produisaient surtout en 18q et aussi 4q, 8p et 17p. L 'HGC pratiqu6e sur les 6chantillons obtenus en endoscopie ou ~ partir de resections donnaient d'aussi bons resultats. Le nombre total d'6v6nements par tumeur d6tect6s par HGC montraient une corr6- lation positive avec l'index en ADN (IA) determin6 en cytom6trie de flux, r = 0,67, p = 0,004. Ces r6sultats confirment les donn6es de Ried et al. [40].
L 'HGC a ddmontr6 que de multiples aberrations chromosomiques 6taient constamment associ6es avec la carcinog6n6se colorectale. Le gain sur le chromo- some 7 apparait comme un 6v6nement pr6coce dans le d6veloppement de l'ad6nome alors qu'un gain en 13q ou 20q pourrait ~tre associ6 ~ la transition entre ad6nome et carcinome. De plus, L'HGC r6alis6e sur des frottis de tumeurs colorectales obtenus en endo- scopie permet la detection de gains ou de pertes pour le g6nome complet de la tumeur en une seule analyse rapide.
Un sous-groupe particulier de cancers colorectaux comprend ceux pr6sentent une instabilit6 des micro- satellites ou des erreurs de r6plication (RER+) dus un dysfonctionnement des m6canismes de r6paration de I'ADN. L'analyse en HGC des carcinomes colo- rectaux avec des instabilit6s des microsatellites a r6v616 qu'ils 6taient en g6n6ral diplo~des et sans ano- malies chromosomiques. Ceci est en contraste avec les tumeurs qui ne montrent pas d'instabilit6 des microsatellites et qui forment la majorit6 des carci- nomes colorectaux [41]. Ceci pourrait signifier que dans les carcinomes colorectaux RER+ l'accumula- tion de changements au niveau des g~nes suppres- seurs de tumeurs et des oncog6nes qui induisent le cancer est caus6e par des mutations de I'ADN pro- prement dit, alors que pour les tumeurs RER- des aberrations au niveau de l'organisation des chromo- somes ou des bras chromosomiques pourraient alt6-
rer la fonction des g6nes suppresseurs de tumeurs et des oncog~nes.
Dans des spdcimens de colectomie provenant de patients souffrant de colite ulc6rative avec un haut degr6 de dysplasie ou un carcinome, L 'HGC a pu d6tecter des aberrations chromosomiques dans des portions de muqueuse qui ne montraient pas ou tr6s peu de signes de dysplasie, indiquant la possibilit6 d'6v6nements pr6coces [18].
Dans une premi6re 6tude des cancers de l'estomac, des gains sur le chromosome 7 et en 13q sont apparus comme les 6v6nements les plus fr6quemment retrou- v6s [19]. D'une manibre intdressante, ces 6v6nements sont aussi fr6quemment retrouvds dans les cancers colorectaux [30, 40].
Implications diniques Actuellement, la tendance pour I 'HGC est d'dtu-
dier un grand nombre de tumeurs afin de trouver des corrdlations avec l'issue clinique. En outre, nous avons montr6 que des 6chantillons obtenus en endo- scopie permettaient une 6tude des anomalies chromo- somiques pour le g6nome complet de la tumeur. Un avantage suppldmentaire de cette approche tient au fait que, en frottant la surface de la tumeur, une forte proportion de cellules 6pithdliales tumorales peut 6tre obtenue alors que la contamination par des cellules stromales, qui diminue le pouvoir de r6solution de I'HGC, est minimale. De ce fait, il n'y a pas de limita- tions pratiques a l'application de I'HGC, ce qui ouvre la voie a de nombreuses applications cliniques pos- sibles pour cette nouvelle et puissante technique. Cette approche ne devrait passe limiter aux tumeurs colorectales. L'HGC sur frottis pourrait aussi s'appli- quer par exemple aux cancers de l'aesophage et de l'estomac et peut-Stre aux 16sions pr6-malignes comme l'cesophage de Barrett, la colite ulcdrative et la gastrite atrophique. I1 est clair que, dans le court laps de temps qui a suivi son introduction, I 'HGC s'est taill6e une part pro6minente dans l'arsenal des tech- niques d'analyse g6n6tique des tumeurs non seule- ment pour les biologistes mol6culaires et les g6n6ti- ciens, mais aussi pour les gastroent6rologistes et les pathologistes. De nouvelles innovations, comme I'HGC rdalis6e sur de I'ADN dispos6 sur des ~< puces ,> semblables h celles des micro-processeurs, destin6es accroitre le pouvoir de r6solution et la sensibilit6 de I'HGC contribueront certainement fi son d6veloppe- ment [42].
R E F E R E N C E S
1. BISHOP J.M. - - Molecular themes in oncogenes. Cell,, 1991, 64, 235-248.
2. B O O N E C.W., K E LLOFF G.J. - - Development of surrogate endpoint biomarkers for clinical trials of cancer chemopreven- tive agents: relationships to fundamental properties of prein- vasive (intraepithelial) neoplasia. J. Cell. Biochem. Suppl. , 1994, 19, 10-22.
3. S I D R A N S K Y D. - - Molecular genetics of head and neck cancer. Curr. Opin. Oncol., 1995, 7, 229-233.
4. H O U L D S W O R T H J., C H A G A N T I R.S.K. - - Comparative genomic hybridization: An overview. Am. J. Pathol., 1994, 145, 1253-1260.
5. H E R M S E N M.A.J .A. , M E I J E R G.A., B A A K J.P.A., JOENJE H., W A L B O O M E R S J.J.M. - - Comparative geno-
94 Volume 27 - N ~ 2 - 1997 Acta Endoscopica

mic hybridization: a new tool in cancer pathology. H u m . Pathol. , 1996, 27, 342-349.
6. KALLIONIEMI A., KALLIONIEMI O.P., SUDAR D. et aL - - Comparative genomic hybridization for molecular cyto- genetic analysis of solid tumors. Science., 1992, 258, 818-821.
7. MANOIR S.DU, SPEICHER M.R., JOOS S. et al. - - Detec- tion of complete and partial chromosome gains and losses by comparative genomic in situ hybridization. Hum. Genet., 1993, 90, 590-610.
8. MANOIR S.DU, SCHROCK E., BENTZ M. et al. - - Quanti- tative analysis of comparative genomic hyrbidization. Cytome- try., 1995,19, 27-4l.
9. RAAP T. - - Cytometry for CGH. Cytometry . , 1995, 19, 1-3. 10. SPEICHER M.R., DU-MANOIR S., SCHROCK E. et al. - -
Molecular cytogenetic analysis of formalin-fixed, paraffin- embedded solid tumors by comparative genomic hybridization after universal DNA-amplification. Hum. Mol. Genet., 1993, 2, 1907-1914.
11. ISOLA J., DEVRIES S., CHU L., GHAZVINI S., WALD- MAN F. - - Analysis of changes in DNA sequence copy num- ber by comparative genomic hybridization in archival paraf- fin-embedded tumor samples. A m . J. Pa tho l . , 1994, 145, 1301-1308.
12. JOOS S., OTANO-JOOS M.I., ZIEGLER S. et aL - - Primary mediastinal (thymic) B-cell lymphoma is characterized by gains of chromosomal material including 9p and amplification of the REL gene. Blood. , 1996, 87, 1571-1578.
13. PIPER J., RUTOVITZ D., SUDAR D. et al. - - Computer image analysis of comparative genomic hybridization. Cyto- metry. , 1995,19, 10-26.
14. MEULEMEESTER M.DE, VINK A., JAKOBS M. et al. - - The application of microwave denaturation in comparative genomic hybridization. Genet ic Ana lys i s (B iomed ica l Enginee- ring), 1996, (in press).
15. KALLIONIEMI O.P., KALLIONIEMI A., PIPER J. et al. - - Optimizing comparative genomic hybridization for analysis of DNA sequence copy number changes in solid tumors. Genes. Chromosom. Cancer., 1994, 10, 231-243.
16. HERMSE'N M.A.J.A., JOENJE H., ARWERT F. et al. - - Assessment of genomic imbalances in oral squamous cell car- cinoma: CGH versus cytogenetic analysis. European confe- rence on comparative genomic hybridization, Semmering, Austria, February 29 - March 2.
17. VOORTER C., JOOS S., BRINGUIER P.P. et al. - - Detec- tion of chromosomal imbalances in transitional cell carcinoma of the bladder by comparative genomic hybridization. A m . J. PathoL, 1995, 146, 1341-1354.
18. WILLENBUCHER R.F., RABINOVITCH P.S., BRENT- NALL T.A. et al. - - Comparative genomic hybridization (CGH) identifies altered DNA sequence copy number in early ulcerative colitis-related neoplasia. Gastroenterol. , 1996, 110, A614.
19. KOKKOLA A., MONNI O., PUOLAKKAINEN P., HAA- PIAINEN R., KIVILAAKSO E., KNUUTILA S. - - DNA copy number changes in gastric carcinoma (GC) as evaluated by comparative genomic hybridization (CGH). Gastroenterol. , 1996, 110, A554.
20. VISAKORPI T., KALLIONIEMI A.H., SYVANEN A.C. et aL - - Genetic changes in primary and recurrent prostate can- cer by comparative genomic hybridization. Cancer. Res., 1995, 55, 342-347.
21. TARKKANEN M., KARHU R., KALLIONIEMI A. et aL - - Gains and losses of DNA sequences in osteosarcomas by comparative genomic hybridization. Cancer. Res. , 1995, 55, 1334-1338.
22. SUIJKERBUIJK R.F., OLDE WEGHUIS D.E., VAN DEN BERG M. et al. - - Comparative genomic hybridization as a tool to define two distinct chromosome 12-derived amplifica- tion units in well-differentiated liposarcomas. Genes. Chromo- som. Cancer., 1994, 9, 292-295.
23. SPEICHER M.R., PRESCHER G., DU MANOIR S. et al. - - Chromosomal gains and losses in uveal melanomas detected by comparative genomic hybridization. Cancer. Res., 1994, 54, 3817-3823.
24. SPEICHER M.R., SCHOELL B., DU MANOIR S. et al. - - Specific loss of chromosomes 1, 2, 6, 10, 13, 17, and 21 in chromophobe renal cell carcinomas revealed by comparative genomic hybridization. A m . J. Pathol . , 1994,145, 356-364.
25. SCHROCK E., THIEL G., LOZANOVA T. et al. - - Compa- rative genomic hybridization of human malignant gliomas reveals multiple amplification sites and nonrandom chromo- somal gains and losses. A m . J. Pathol . , 1994, 144, 1203-1218.
26. RIED T., PETERSEN I., HOLTGREVE-GREZ H. et al. - - Mapping of multiple DNA gains and losses in primary small cell lung carcinomas by comparative genomic hybridization. Cancer. Res., 1994, 54, 1801-1806.
27. RIED T., KNUTZEN R., STEINBECK R. et al. - - Compara- tive genomic hybridization reveals a specific pattern of chro- mosomal gains and losses during the genesis of colorectal tumors. Genes, C h r o m o s o m e s & Cancer, 1996, 15, 234-245.
28. KALLIONIEMI A., KALLIONIEMI O.P., PIPER J. etal. - - Detection and mapping of amplified DNA sequences in breast cancer by comparative genomic hybridization. Proc. Natl . Acad. Sci. U. S. A . , 1994, 91, 2156-2160.
29. BENTZ M., HUCK K., DU MANOIR S. et aL - - Compara- tive genomic hybridization in chronic B-cell leukemias shows a high incidence of chromosomal gains and losses. Blood. , 1995, 85, 3610-3618.
30. MEIJER G.A., HERMSEN M.A.J.A., DIEST P.J.VAN, et al. - - Survey of genetic changes in colorectal tumours by comparative genomic hybridization. Gastroenterol. , 1996, 110, A556.
31. TANNER M.M., TIRKKONEN M., KALLIONIEMI A. et al. - - Increased copy number at 20q13 in breast cancer: defi- ning the critical region and exclusion of candidate genes. Can- cer. Res., 1994, 54, 4257-4260.
32. KUUKASJJkRVI T., KARHU R., TANNER M., KALLIO- NIEMI O.P., ISOLA J. - - Analysis of the genetic evolution of breast cancer from primary tumors to distant metastases by comparative genomic hybridization (CGH). European confe- rence on comparative genomic hybridization, Semmering, Austria, February 29 - March 2, 1996.
33. VISAKORPI T., HYYTINEN E., KOIVISTO P. et al. - - In vivo amplification of the androgen receptor gene and progres- sion of human prostate cancer. Nat. Genet., 1995, 9, 401-406.
34. QUIRKE P., DYSON J.E., DIXON M.F., BIRD C.C., JOS- LIN C . A . - - Heterogeneity of colorectal adenocarcinomas evaluated by flow cytometry and histopathology. Br. J. Cancer., 1985, 51, 99-106.
35. HIDDEMANN W., VON-BASSEWITZ D.B., KLEINE- MEIER H.J. et al. - - DNA stemline heterogeneity in colorec- tal cancer. Cancer., 1986, 58, 258-263.
36. BERGERS E., DIEST P.J.VAN, BAAK J.P.A. - - Tumour heterogeneity of DNA cell cycle variables in breast cancer measured by flow cytometry. J. Clin. Pathol. , 1996, 49, 931-937.
37. KERN S.E., FEARON E.R., TERSMETTE K.W. et al. - - Allelic loss in colorectal carcinoma. Jama. , 1989, 261, 3099- 3103.
38. HARADA Y., KATAGIRI T., ITO I. et al. - - Genetic studies of 457 breast cancers. Clinicopathologic parameters compared with genetic alterations. Cancer., 1994, 74, 2281-2286.
39. ISOLA J.J., KALLIONIEMI O.P., CHU L.W. et al. - - Gene- tic aberrations detected by comparative genomic hybridiza- tion predict outcome in node-negative breast cancer. A m . J. PathoL, 1995, 147, 905-911.
40. RIED T., KNUTZEN R., STEINBECK R. et al. - - Compara- tive genomic hybridization reveals a specific pattern of chro- mosomal gains and losses during the genesis of colorectal tumors. Genes, C h r o m o s o m e s & Cancer, 1996,15, 234-245.
41. SCHLEGEL J., STUMM G., SCHERTHAN H. et al. - - Comparative genomic in situ hybridization of colon carcino- mas with replication error. Cancer. Research. , 1995, 55, 6002- 6005.
42. M0LLER U., PIPER J., ALUL R., RUFFALO T., CAO Q. - - Genosensor based comparative genomic hybridization. European conference on comparative genomic hybridization, Semmering, Austria, February 29 - March 2, 1996.
Acta Endoscopica V o l u m e 2 7 - N ~ 2 - 1997 95

INTRODUCTION
The development of solid tumours is associated with the acquisition and accumulation of genetic changes [1-3]. Analysis of genetic abnormalities in tumour spe- cimens is evolving as an important new aspect o f tumour pathology, which will have increasing influence on clinical decision making. [4, 5] The trend in pathology is to aim for the highest degree of dia- gnostic and prognostic information, as early as pos- sible in the stage o f diagnostic decision making, and achieved with minimally invasive methods. Genetic analyses can be performed in many different ways, ranging from somewhat crude techniques like DNA cytometry to very detailed methods like tumour cyto- genetics, in situ hybridization (ISH) or PCR based study of loss of heterozygosity (L OH).
Recently, a new method, comparative genomic hybridization (CGH) has been described that is capable of both detailed and extensive evaluation of the genetic changes throughout the whole genome of tumours. [6, 7] It provides information on chromoso- mal losses and gains or amplifications in a single expe- riment. In the following, we will present a detailed des- cription o f all the steps of the CGH technique and discuss the advantages, disadvantages of the method itself and show a number of different applications.
Technical details of CGH
A schematic overview of the technique is shown in figure 1. First, tumour DNA is labelled with a green fluorochrome, and normal (diploid) control DNA (not necessarily from the same patient) is labelled with a red fluorochrome. These labelled DNAs are mixed and hybridized to normal human metaphase prepara- tions. The green and the red labelled DNA compete for hybridization to the chromosomes L~ing fluorescence microscopy and digital image analysis, the ratio of the green and red colour along all the chromosomes is quantified. The ratio is a measure of under- or over- representation of genetic material of the tumour for each particular chromosomal region. The whole pro- cess of analysing a tumour with CGH takes the follo- wing steps:
N O R M A L M E T A P H A S E PREPARATION. Normal meta- phase slides are prepared from phytohaematoglutinine stimulated peripheral blood lymphocyte cultures from a healthy individual. Cells are arrested in mitosis by colchicine, harvested, treated with hypotonic KCl and fixed in methanol~acetic acid. The cells are mounted onto slides in such a way that chromosomes from mitotic cells are nicely spread.
TUMOUR DNA ISOLATION. Starting with a minimum of + 200,000 cells or approximately 1 mm 3 of tissue, tumour DNA can be obtained by any type o f DNA isolation which yields high molecular DNA, required for use in CGH. Also DNA extracted from paraffin embedded material, which may be partly degraded, can be used with adjustments in the labelling. Very small amounts of DNA may be used in combination with degenerate oligo-primed PCR amplification (DOP-PCR). Normal DNA for use as a reference can
be taken from blood lymphocytes from any healthy individual
DNA LABELLING. A standard labelling (nick trans- lation) is used to label the DNA and to digest the DNA to the optimal fragment length of 600-2000 base pairs. A small amount ofO.5 - 1.0 tag DNA is sufficient. In the case of DOP-PCR, labelling may take place directly in the amplification process. Up until now, mostly biotinylated and digoxygenin-conjugated deoxynucleotides are incorporated in the DNA label- ling, which necessitates a detection step after hybridi- zation with fluorochrome labelled antibodies (respec- tively Avidin-FITC and Sheep-anti-Digoxygenin- TRITC). It is also possible to use directly fluoro- chrome-conjugated deoxynucleotides, which render a smoother, but weaker, hybridization signal along the chromosomes. It is important that the labelled DNA fragments of both tumour and reference DNA have the same length and are within the 600-2000 base pairs range.
HYBRIDIZATION. Approximately 150-200 ng of both tumour and reference labelled DNA (the probes) are mixed with 100 times the same amount of unlabelled human Cot-1 DNA, precipitated and resolved in 10 ml hybridization mix containing 50 % formamid and 10 % dextran sulphate. Cot-1 DNA consists of purified repetitive sequences and is added to block the highly polymorphic repeat sequences in the DNA. The probe and the normal metaphase slides are denatured sepa- rately. The hybridization takes place in a humid incu- bator at 37~ for 2-4 days. After hybridization, the slides are washed and counterstained with 4,6-diami- dino-2-phenylindole (DAPI) to produce a banding pattern, that enables chromosome identification and standard karyotyping.
I M A G E A C Q U I S I T I O N A N D A N A L Y S I S . With fluores- cence microscopy, using three separate band pass fil- ters (for DAPI, FITC and TRITC respectively) in combination with a CCD camera, three digital black
Figure 1 Schematic overview of the CGH technique. Tumour and reference DNA
are labelled with a green and red fluorochrome, respectively and hybridized to normal metaphase spreads (see also fig. 2b). Images of
the fluorescent signals are captured and the green to red signal ratios are digitally quantified along the chromosomal axis for each chromosome.
(Reprinted from reference 5).
96 Volume 27 - N ~ 2 - 1997 Acta Endoscopica

Figure 2
C : Relative copy number karyotype of the same tumour showing a graphical display of the mean and 95 % confidence interval of the green to red fluorescence ratio along all individual chromosomes, obtained by
analysing multiple metaphases.
and white images are captured of each metaphase and reconstituted on a computer screen (Figure 2b). The image processing for CGH has been reviewed extensi- vely. [8, 9] Special computer software enables interac- tive karyotyping of the chromosomes and the calcula- tion of the green to red fluorescence ratio o f each chromosome. The averaged ratios of several well- selected metaphases, together with the 95 % confidence interval, are plotted along ideograms of the correspon- ding chromosomes in a so-called relative copy number karyotype (Figure 2c).
Advantages of CGH
CGH has distinct advantages over other techniques for analysis of genetic changes in tumour pathology. First, CGH is not dependent on the tumour source; it requires only a small amount of tumour DNA, obtai- ned either from cell lines or from fresh, frozen or paraffin embedded tissue. The applicability to archival paraffin embedded material allows retrospective ana- lysis of large numbers of tumours that are pathologi- cally well-characterized and of which the clinical out- come is known [10]. Recently, an improved DNA extraction technique for paraffin embedded tissue has been published which yields high molecular DNA (needed for CGH) in about 80 % of all cases [11]. In cases where the DNA yield from paraffin embedded tissue is too low (less than 1-2 pg), CGH may still be possible after universal PCR amplification (DOP- PCR). With this modification, a minimum of some 2,000 cells is sufficient. A number of 2,000 cells can easily be scraped from the tissue sections. I f the mate- rial has been fixed in buffered formalin, an even lower number (10- 30 cells) can be enough [12]. CGH ana- lysis with PCR amplified DNA was found to be repro- ducible and showed the same cytogenetic abnormali- ties as with DNA from the same tumour isolated from frozen tissue [10]. Secondly, it is able to produce detai- led information on gains and losses and to do that throughout the whole genome in only a single experi-
ment. The CGH assay can be especially helpful in the analysis of the genetic changes underlying solid tumours, which are relatively inaccessible to conven- tional cytogenetic analysis because cell cultures o f these tumours are very difficult to obtain.
Limitations of CGH
CGH cannot detect balanced chromosomal translo- cations, which are frequently found in haematological and mesenchymal tumours and to a much lesser extent, also in solid tumours. Screening for these abnormali- ties can only be performed with (spectral) karyoty- ping, after culturing of tumour cells. In addition, sub- microscopical deletions will be missed by CGH. Theoretically, the lower limit of detectability is deter- mined by the product of the excess copy number and the size of the amplicon. It is estimated that a copy number increase of 50 % should be detectable if the region is about 10 Mbase or larger, and an amplified region of 250 kbase if the copy number is 400 % increa- sed [13]. The detection limits for deletions are higher, because the maximum loss can only be 100 %. In our experience a 10 Mbase loss could still be detected [14]. Contamination of tumour with normal cells (i.e. stro- mal cells, lymphocytes) will lower the sensitivity of the CGH analysis. Kallioniemi et al., and others have sta- ted that 50 % non-tumour cells in the test sample is the limit for the detection o f monosomies or trisomies [10, 15]. The admixture of non-tumour cells in the test sample can be reduced using microdissection of the histologically relevant tissue parts. Furthermore, peri- centromeric and telomeric regions are excluded from analysis. These regions contain polymorphic repetitive sequences and therefore are blocked by unlabelled Cot-1 DNA in the hybridization. In addition, ratio profiles from regions at the tip of chromosome 1, the short arm of chromosome 16, chromosome 19 and 22 can vary and should therefore be interpreted with cau- tion [8, 15].
Validation of the CGH technique
The CGH technique has been validated in many ways. First, by analysing normal DNA and by analy- sing tumour DNA with known gains and deletions. Secondly, by comparing CGH results to other tech- niques such as classical cytogenetics and LOH analy- sis. From these experiments it can be concluded that CGH does not produce false positive or negative results.
CGH with normal versus normal DNA has been shown to produce straight profiles with confidence intervals that contain the ratio 1.0 on all chromosomes, with the exception o f centromeric and telomeric regions [8, 15]. The increase o f chromosome copy number has been shown to correlate with the average green to red ratio [6]. However, CGH should not be used to quantify the copy number of gains and losses, because the correlation is not linear.
A study of our group, designed to compare the results of CGH analysis on six oral squamous cell car-
A c t a E n d o s c o p i c a V o l u m e 2 7 - N ~ 2 - 1 9 9 7 97

cinoma cell cultures to those revealed by karyotypic analysis, showed that genetical changes, particularly losses, could be more reliably detected by CGH analy- sis than by karyotyping [16]. In cases of marked diffe- rent findings between CGH and cytogenetic analysis, FISH analysis using whole chromosome paints confir- med the CGH results. In general, FISH analysis sho- wed that the presence of cytogenetically unidentifiable chromosomes, so called markers, in the karyotypes of the cell cultures was the most likely cause of the discre- pancies. This means that when based on the cytogene- tical analysis alone, the number of chromosomal losses is generally overestimated.
The CGH results concerning loss of chromosomal material have also been compared to the results of loss o f heterozygosity (LOH) analysis. Although L O H analysis detects the loss of an allele, and CGH the rela- tive loss of a chromosome compared to the total DNA content or ploidy, the two techniques produce remar- kably similar results. Voorter et al., reported 100 % L O H at loci where CGH detected losses in bladder tumours [17]. On the other hand, only 50 % of LOH loci were identified by CGH. There are two possible explanations for this seemingly lack o f sensitivity. Either the deletion is too small for CGH to detect, or uniparental disomy (i.e. loss o f one chromosome, fol- lowed by duplication of the remaining chromosome) had occurred in this region. In another study, analy- sing LOH in four separately HPV transfected kerati- nocyte cell lines, only about 50 % of the losses detected by CGH were also detected by L O H analysis. This could be due to loss of only one of three chromosomes in these triploid cell lines, leaving two heterozygous homologues. On the other hand, I00 % of the loci that displayed L O H were also identified as lost by CGH analysis (Steenbergen, RDM et al., unpublished results). Kallioniemi, OP et al., and Sakamoto et al. reported a concordance between CGH and LOH for about 80 % for breast and ovarian cancer, respecti- vely. [18, 19] In conclusion, L O H and CGH can be regarded as supplementary techniques, each having their own specific applicability.
Applications of CGH
The applications of CGH can be multiple. CGH has been used to screen large numbers of tumour genomes for consistent chromosomal regions with amplification or deletion o f genes in specific types of tumours. Already has the number of solid tumours analysed by CGH outranked the total number o f solid tumours that have been analysed by classical cytogenetics. In the past three years, a wide range of tumour types, ran- ging from melanoma, carcinoma of lung, colon, head and neck, prostate, ovary, renal and bladder cancer, osteosarcoma, glioma, soft tissue and haematological tumours, have been screened for chromosomal gains and losses by CGH. [17,19-30]
The genetic basis o f tumour development can be investigated by analysing different stages of disease (i.e. premalignant lesions, invasive cancer and metas- tases). In this way, CGH may serve as a first line ana- lysis in tumour genetics, globally pointing out chromo-
somal regions of interest that subsequently can be stu- died in more detail by other molecular genetic tech- niques. In the case of breast cancer, CGH analysis has led to the identification of new chromosomal regions that are possibly involved in the pathogenesis of breast cancer and in the metastatic process. In a series of 33 primary invasive tumours, Kallioniemi and coworkers found, besides the already known chromosomal gains at lq and 8q, recurrent gains at 17q and 20q [28]. The gain at 20q was further localized at band 20q13, howe- ver the search for putative oncogenes with molecular genetic techniques has not yet been successful [31]. Also in breast cancer, by comparing primary tumours and their distant metastases, Kuukasji~rvi et al., found gain of Xq to be a newly acquired abnormality in the metastases, indicating a possible role for this region in the process of metastasis [32].
Furthermore, CGH can help unravel cell biological mechanisms which can explain in vivo characteristics such as unresponsiveness to therapy and genetic hete- rogeneity. CG H analysis of recurrent prostate tumours from patients which were treated with androgen depri- vation therapy revealed amplification of chromosome region Xqll-12 in 55 % of all cases, while in the pri- mary tumours no amplification was detected at all. Chromosome band X q l l harbours the gene for androgen receptor (AR). Using A R as a probe for FISH and for mRNA in situ hybridization it was shown that AR was amplified and over-expressed in 28 % of all cases. These results indicate that failure of androgen deprivation therapy is caused by clonal expansion of tumour cells that are able to up-regulate the expression of the androgen receptor [33].
The existence of intra-tumour heterogeneity at the phenotypical level was already well known to the pathologist, and also by DNA cytometry this pheno- menon has been demonstrated [34-36]. We could demonstrate this heterogeneity at the chromosomal level by CGH analysis of samples, taken from two dif- ferent spots o f the same colorectal carcinomas. We found that some gains or losses were present to the same extend in both samples, whereas other findings were much less outspoken or even no longer signifi- cant in one sample compared to the other [30]. This could indicate that gains and losses common to both samples represent early changes whereas the diffe- rences between the samples may be late events. This opens up the possibility to analyse the genetic basis for phenotypical differences within one tumour.
Finally, clinical decision making could benefit from CGH research. The applicability of CGH to archival paraffin embedded material (if needed in combination with DOP-PCR) will further increase the amount of clinically relevant analyses. Based on the idea that changes in the genome influence the behaviour o f tumours, pathologists and clinicians already are used to make decisions on the basis of DNA histograms in the case of certain types of tumours. These histograms tell whether or not in a tumour there is a change in the amount of DNA per cell. This of course is a rather crude way of analysing changes in DNA. CGH can tell on which chromosomes, and where on these chromo-
98 V o l u m e 2 7 - N ~ 2 - 1997 Ac ta Endoscop ica

somes losses or gains of DNA are present and it could therefore be o f even greater clinical importance than DNA ploidy. Indeed it has been shown that an increa- sed genetic instability, as illustrated by the number of genetic alterations, correlates with adverse prognosis and / or clinicopathological tumour characteristics in e.g. colorectal carcinomas and breast cancer [37, 38]. Application of CGH to archival, paraffin embedded material of which a long-term follow up is available, enables a genome wide survey of the clinical relevance of specific chromosomal aberrations [39]. By investi- gating changes that are associated with the transition from e.g. premalignant to a malignant stage, and by correlating CGH results to prognosis, CGH may serve as a basis for classification of disease and prediction o f clinical outcome [4, 5].
CGH in tumours of the gastrointestinal tract
The number o f studies applying CGH to cancers and pre-malignant lesions of the gastrointestinal tract is growing. In a recent study, we analysed brushings taken from the surface of16 colorectal tumours, either from fresh resection specimens, or in situ during endo- scopy (Fig. 2a) [30]. In the four adenomas analysed, on average 6.8 (range 4 - 10) genetic changes were identified. In the 12 carcinomas on average 11.1 (range 5 - 17) events were found per tumour. Both in adeno- mas and carcinomas, gains occurred more frequently than losses. Frequent gains concerned 13q, 7p & q, 8q and 20q, whereas losses most frequently occurred at 18q, and also 4q, 8p, and 17p. CGH on endoscopically obtained samples and samples taken from fresh resec- tion specimens yielded equally good results. The total number of events per tumour detected by CGH sho- wed a positive correlation with DNA index (DI) asses- sed by DNA cytometry, r = 0.67, p = 0.004. These fin- dings confirmed the results o f Ried et al. [40].
CGH demonstrated multiple chromosomal aberra- tions to be consistently associated with colorectal carci- nogenesis. Gain of chromosome 7 appears to be an early event in adenoma development while gain of 13q and 20q could be associated with the transition of adenoma to carcinoma. Moreover, CGH on endoscopically obtai- ned brushings from colorectal tumours, allows for the detection of chromosomal gains and losses in the com- plete tumour genome in one rapid analysis.
A special subgroup of colorectal cancers are those with microsatellite instability or replication errors (RER+) due to a disorder in the DNA mismatch repair system, which often is HNPCC. CGH analysis revea-
led that colorectal carcinomas which showed microsa- tellite instability were in general diploid and harboured no chromosomal abnormalities. This in contrast with tumours that did not show microsatellite instability, which form the majority o f colorectal carcinomas [41]. This could mean that in RER+ colorectal carcinomas, the accumulation of changes in tumour suppressor genes and oncogenes that induce cancer is caused by mutations on the D N A level, whereas for RER- tumours aberrations caused at the level of the chromo- somes or chromosome arms might affect the function of tumour suppressor genes and oncogenes.
In colectomy specimens from patients with ulcera- tive colitis with high grade dysplasia or carcinoma, CGH could detect chromosomal aberrations already in background mucosa negative for dysplasia or sho- wing only low grade dysplasia, pointing at possible early events [18].
In a first study of gastric cancers, gains of chromo- somes 7 and 13q appeared to be the most frequent fin- dings [19]. Interestingly, these events were also fre- quently found in colorectal cancers [30, 40].
Clinical implications
At the present stage, the emphasis in CGH research is on investigating large numbers of tumours in order to find correlations with clinical outcome. Further- more, we have shown that endoscopically obtained samples allow for a survey of chromosomal aberra- tions in the complete tumour genome. An extra advan- tage of this approach is the fact that, by brushing the surface of the tumour, a high yield of epithelial tumour cells can be obtained, while admixture of stroma cells, which lowers the resolution of CGH, is minimal. In this respect there are no practical limitations to the application of CGH, opening the way for many pos- sible clinical applications o f this new and powerful technique. This approach does not need to be limited to colorectal tumours. CGH on brushings could also be applied to e.g. cancers of the oesophagus and sto- mach, and perhaps to premalignant lesions like Bar- rett' s oesophagus, ulcerative colitis and atrophic gastri- tis. It is clear that in the short period of time since its introduction, CGH has claimed a prominent position among the arsenal o f tools for genetic analysis o f tumours available not only to molecular biologists and geneticists, but also to gastroenterologists and patholo- gists. New innovations like CGH on DNA micro- arrays, aimed at increasing the resolution and sensiti- vity o f C G H will certainly contribute to this development [42].
Acta Endoscopica Volume 27- N~ 2 - 1997 99




















