
Leaf-litter quality effects on stream ecosystem functioning: a comparison among five species
10
Leaf-litter quality effects on stream ecosystem functioning: a comparison among five species Aingeru Martínez*, Aitor Larrañaga, Javier Pérez, Ana Basaguren and Jesús Pozo 1 With 5 figures and 3 tables Abstract: The replacement of native vegetation by exotic species has the potential to impact detritus based stream ecosystems. To test the possible consequences of vegetation alteration on stream ecosystem functioning we com- pared the processing rate and invertebrate colonisation of leaves from five different tree species, two natives (Alnus glutinosa and Quercus pyrenaica) and three exotics (Populus × canadensis, Platanus hispanica and Pinus radiata) in three unaltered forested streams in the Atlantic region of northern Spain. Breakdown rates, nitrogen concentra- tion and C:N ratio of incubated leaves, and invertebrate communities in the benthos and bags were analysed. Break- down rates differed among leaf species and were positively related to initial nitrogen concentration and negatively to the C:N ratio. After the incubation period (6 –7 weeks), the differences in nutritional quality among leaf species and the relationship between breakdown rates and litter quality persisted. The density and biomass of macroinverte- brates and shredders associated with litter bags correlated with leaf-nitrogen concentration and explained the differ- ences in breakdown rates. The results indicate that the replacement of native deciduous forest by plant commercial species with leaf-litter of low quality such as plane and pine can affect in-stream organic matter processing and the subsequent transference of energy throughout the food web. Key words: vegetation alteration, headwater streams, leaf-litter decomposition, nitrogen concentration, C:N ratio, macroinvertebrates. Fundam. Appl. Limnol. Vol. 183/3 (2013), 239–248 Article Stuttgart, October 2013 © 2013 E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart, Germany www.schweizerbart.de DOI: 10.1127/1863-9135/2013/0514 1863 - 9135/13/0514 $ 2.50 Introduction Detrital processing plays a key role in the function- ing of any ecosystem (Moore et al. 2004). This fact is particularly evident in low order forested streams, where the inputs of dead organic matter from the sur- rounding forest, mainly in the form of leaves (Abelho 2001), represent the principal source of matter and energy (Webster et al. 1999). The organisms respon- sible for allochthonous detritus processing, microbial decomposers, primarily aquatic hyphomycete fungi (Gulis & Suberkropp 2003), and invertebrate detriti- vores (Hieber & Gessner 2002), are decisive for the transfer of energy to higher trophic levels (Wallace et al. 1997). Thus, the detrital pathway has been the focus of functional studies in these systems over the last three decades (Tank et al. 2010). Several studies focus on the quality of the detritus, which mostly de- pends on the species of the surrounding forest (Swan & Palmer 2006), among other factors (i.e. quantity of the inputs or their timing), and can limit the per- formance of consumers (Tibbets & Molles 2005, Re- inhart & VandeVoort 2006). The physico-chemical characteristics of leaves such as toughness, roughness, secondary compounds or nutrient content determine decomposer colonisation and activity (Bärlocher & Oertli 1978, Dang et al. 2007, Kearns & Bärlocher 2008) and its consumption by detritivores (Haapala et al. 2001, Kominoski & Pringle 2009). Among these intrinsic characteristics, nutrient content plays a key Authors’ address: 1 Laboratory of Stream Ecology, Department of Plant Biology and Ecology, University of the Basque Country, P.O. Box 644, 48080 Bilbao, Spain * Author for correspondence; [email protected] E
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Leaf-litter quality effects on stream ecosystem functioning: a comparison among five species

Leaf-litter quality effects on stream ecosystem functioning: a comparison among fi ve species
Aingeru Martínez*, Aitor Larrañaga, Javier Pérez, Ana Basaguren and Jesús Pozo 1
With 5 fi gures and 3 tables
Abstract: The replacement of native vegetation by exotic species has the potential to impact detritus based stream ecosystems. To test the possible consequences of vegetation alteration on stream ecosystem functioning we com-pared the processing rate and invertebrate colonisation of leaves from fi ve different tree species, two natives (Alnus glutinosa and Quercus pyrenaica) and three exotics (Populus × canadensis, Platanus hispanica and Pinus radiata) in three unaltered forested streams in the Atlantic region of northern Spain. Breakdown rates, nitrogen concentra-tion and C:N ratio of incubated leaves, and invertebrate communities in the benthos and bags were analysed. Break-down rates differed among leaf species and were positively related to initial nitrogen concentration and negatively to the C:N ratio. After the incubation period (6 –7 weeks), the differences in nutritional quality among leaf species and the relationship between breakdown rates and litter quality persisted. The density and biomass of macroinverte-brates and shredders associated with litter bags correlated with leaf-nitrogen concentration and explained the differ-ences in breakdown rates. The results indicate that the replacement of native deciduous forest by plant commercial species with leaf-litter of low quality such as plane and pine can affect in-stream organic matter processing and the subsequent transference of energy throughout the food web.
Key words: vegetation alteration, headwater streams, leaf-litter decomposition, nitrogen concentration, C:N ratio, macroinvertebrates.
Fundam. Appl. Limnol. Vol. 183/3 (2013), 239–248 ArticleStuttgart, October 2013
© 2013 E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart, Germany www.schweizerbart.deDOI: 10.1127/1863-9135/2013/0514 1863 - 9135/13/0514 $ 2.50
Introduction
Detrital processing plays a key role in the function-ing of any ecosystem (Moore et al. 2004). This fact is particularly evident in low order forested streams, where the inputs of dead organic matter from the sur-rounding forest, mainly in the form of leaves (Abelho 2001), represent the principal source of matter and energy (Webster et al. 1999). The organisms respon-sible for allochthonous detritus processing, microbial decomposers, primarily aquatic hyphomycete fungi (Gulis & Suberkropp 2003), and invertebrate detriti-vores (Hieber & Gessner 2002), are decisive for the transfer of energy to higher trophic levels (Wallace et al. 1997). Thus, the detrital pathway has been the
focus of functional studies in these systems over the last three decades (Tank et al. 2010). Several studies focus on the quality of the detritus, which mostly de-pends on the species of the surrounding forest (Swan & Palmer 2006), among other factors (i.e. quantity of the inputs or their timing), and can limit the per-formance of consumers (Tibbets & Molles 2005, Re-inhart & VandeVoort 2006). The physico-chemical characteristics of leaves such as toughness, roughness, secondary compounds or nutrient content determine decomposer colonisation and activity (Bärlocher & Oertli 1978, Dang et al. 2007, Kearns & Bärlocher 2008) and its consumption by detritivores (Haapala et al. 2001, Kominoski & Pringle 2009). Among these intrinsic characteristics, nutrient content plays a key
Authors’ address:1 Laboratory of Stream Ecology, Department of Plant Biology and Ecology, University of the Basque Country, P.O. Box 644,
48080 Bilbao, Spain* Author for correspondence; [email protected]
E

240 Aingeru Martínez, Aitor Larrañaga, Javier Pérez, Ana Basaguren and Jesús Pozo
role determining leaf-litter processing (Enriquez et al. 1993, Richardson et al. 2004, Muto et al. 2011) since invertebrate detritivores prefer leaf species with high nutrient concentrations to supply their stoichiometric demands (Sterner & Elser 2002, Woodward 2009). A replacement of the surrounding vegetation can alter the quantity, quality and timing of nutrients that enter in the form of leaf-litter to streams (Pozo et al. 1997, Molinero & Pozo 2006), which might alter biotic com-munities (Weijters et al. 2009) and ecosystem func-tioning (Martínez et al. 2013, Menéndez et al. 2013).
Spain is a European country with a large surface dedicated to forest plantations (2.7 Mha, FAO 2010). Therefore, it is important to gain an insight into the consequences of this practice on ecosystems, streams included. The most commonly planted tree species in Spain, such as pine, eucalypt, spruce or hybrids of poplar (FAO 2010), normally deliver a lower quality detritus than the native forest tree species they are re-placing. This reduction of the detritus quality is par-ticularly strong in areas, such as the Atlantic region of northern Spain, where the native riparian forests are constituted by plant species that offer high qual-ity, nutrient-rich, detritus such as Fraxinus excelsior L., Corylus avellana L. and primarily Alnus glutinosa (L) Gaertner (see quality of these species in Hladyz et al. 2009). In this region, headwater streams can be especially sensitive to the vegetation substitution, as has been reported previously (e. g. Molinero et al. 1996, Pozo et al. 1997, Larrañaga et al. 2009a,b). In the present study we focus on three exotic tree spe-cies in the Iberian Peninsula, namely poplar (Popu-lus × canadensis Moench), plane (Platanus hispanica Mill. ex Münchh.) and Monterey pine (Pinus radiata D. Don), and compare the processing of their leaf-lit-ter to that of two native species, common alder (A. glu-tinosa) and Pyrenean oak (Quercus pyrenaica Willd.). Poplar and plane are very common trees in the ripar-ian corridors, either planted or naturalized (Herrera & Campos 2010), while Monterey pine plantations can cover large areas in northern Spain (e.g. 62 % of surface in Biscay province, www.bizkaia.net). On the other hand, while alder is a nitrogen fixing species that dominates the natural riparian forests of the area, the Pyrenean oak forms natural forests in siliceous soils. Together the five species span a wide range of nitro-gen concentration in natural and exotic leaf-litter of the common tree species in the area (Molinero & Pozo 2006, Arroita et al. 2012, Casas et al. 2013, Martínez et al. 2013).
Our goal in this work was to gain insight into the role of the quality of detritus, measured in terms of
nitrogen content, on stream ecosystem functioning. To this end, we studied the breakdown rates and colo-nizing macroinvertebrates of five leaf species in three undisturbed streams in the Atlantic region of northern Spain. Thus, the overall results of this work could be used to ascertain the implications of natural vegetation replacement on the conservation status of headwater stream ecosystems.
Material and methods
Study area
The study was carried out in three low-order (2nd– 3rd) streams (S1, S2 and S3) in catchments over siliceous substrata, flowing into the Atlantic Ocean (Cordillera Cantábrica, northern Spain). The streams drain native deciduous forests, the dominant tree species being common hawthorn (Crataegus monogyna (Jacqs.)) (range of trees per 100 m2: 2 –18), alder (0 – 23), beech (0 –16) and hazel (0 –16) in the riparian corridor and English oak (being Quercus robur L.) and beech (Fagus sylvatica L.) in the rest of the catchment. Anthropic impacts in the basins are negligible. During the study period (November 2009 to January 2010), water temperature was continuously monitored (every hour) with SmartButton data loggers (ACR Systems Inc., Sur-rey, BC, Canada). At each field visit (n = 5 – 6), conductivity, pH, dissolved oxygen (WTW multiparametric sensor) and river flow (Martin Marten Z30, Current-meter) were measured; wa-ter samples were collected and filtered in the laboratory (0.7 µm pore size glass fiber filter, Whatman GF/F). An aliquot of the filtered water was used to determine alkalinity by titration to an end pH of 4.5 (APHA 2005) and the rest was stored (at – 20 °C) for subsequent analyses. Nutrient analyses of the water were conducted using the capillary ion electrophoresis for nitrate (Agilent CE), the manual salicylate method for ammonium, the sulphanilamide method for nitrite and the molybdate method for soluble reactive phosphorus (SRP) (APHA 2005).
To characterise the macroinvertebrate community of each stream, five benthic samples (Surber 0.09 m2, 0.5 mm mesh size) were taken from randomly chosen riffles in January 2010. Fauna was preserved in 70 % ethanol until further identifica-tion. In the laboratory, macroinvertebrates were identified to family level (Oligochaeta to order, Chironomidae to tribe) us-ing Tachet et al. (2002). All individuals were counted and sorted into shredders and other macroinvertebrates (after Tachet et al. 2002, Merritt & Cummins 2007). The dry mass of shredders and of the rest of invertebrates was determined by oven-drying (70 °C; 72 h). For each macroinvertebrate sample, taxa richness and Shannon diversity index were calculated.
Decomposition experiment and colonizing macroinvertebrates
Leaves of the selected five tree species have a different initial quality and nitrogen content (see Casas et al. 2013). In Octo-ber – November 2009, leaves were collected from the floor immediately after natural abscission except for pine needles, which were collected from dry branches. Leaf-litter was air-dried, and approximately 5 g (± 0.25) of each species were placed separately into mesh bags (15 × 20 cm, 5 mm mesh size

241Leaf-litter quality effects on stream ecosystem functioning
to allow stream biota access). Pine needles for each bag were grouped into five packs tied with a bridle (ca. 1 g each), to avoid whole needles washing out of the bag but without limit-ing biota access. At each site, five iron bars were anchored to the streambed. Five bags, one of each species, were fastened with nylon lines to each bar (25 bags per site). Five extra bags of each species were used to estimate initial ash-free dry mass (AFDM); an additional bar in one of the streams (S2) was used to tie five extra bags of each species to determine loss by leach-ing of initial material. Bags were placed between November 23 and November 26. After 24 h, bags for the leaching estimation were collected. The remaining bags were retrieved at the same time from each stream, after 45 days in S1 and 53 days in the other two streams, coinciding with the estimated 50 % of mass loss for alder according to a parallel study, and were transported refrigerated to the laboratory for subsequent processing.
Bag contents were rinsed with filtered (0.2 mm mesh size) stream water on a 0.5 mm sieve and macroinvertebrates were separated from leaf-litter and preserved in 70 % ethanol until further identification. Leaf material was oven-dried to obtain dry mass (70 °C, 72 h); sub-samples of each bag were used for nutrient analyses (C and N) and the rest was ashed (500 °C, 4 h) to determine the remaining AFDM. The leaf nutrient con-tent analyses were carried out on initial material (post-leaching) and on incubated material. Leaf C and N concentrations were determined using a Perkin Elmer series II CHNS/O elemental analyser. Results were expressed as percentage of elemental concentrations (C and N) of leaf-litter dry mass and in molar ratio (C:N). Macroinvertebrates colonizing leaf-litter bags were studied using the same methodologies used for benthic fauna.
Statistical analyses
The leaf-mass losses were transformed into linear rates con-sidering the incubation period, (M0 – Mt)/t, where M0 is the
initial mass after leaching and Mt the remaining mass at time t. As there were slight temperature differences between streams, processing rates were also calculated using degree-days (dd, the sum of mean daily temperatures during the incubation time) to standardize the effect of temperature on the process. Analyses of differences in mass loss rates, leaf elemental ratios and asso-ciated macroinvertebrate variables (density, biomass, richness and diversity of total invertebrates and shredders) were per-formed with two-way ANOVA (stream; leaf species) followed by post-hoc testing using the Tukey test (Zar 2010). Bivariate relationships were tested by ordinary least-square linear regres-sions. When necessary, data were transformed (log (x +1)) to meet the requirements for parametric analyses. All statistical analyses were conducted using R statistical program (version 2.11.1; R Development Core Team 2010).
Results
Stream water characteristics
The water of the three streams was near saturation, with neutral pH and low nutrient concentrations. The only remarkable difference in the water chemistry was the higher nitrate concentration in S1 than in the other two streams (Table 1; ANOVA: F2,14 = 85.75, p < 0.001).
Mass loss rates and resource quality
The mass loss rates differed among leaf species (Fig. 1; ANOVA: F4,60 = 205.57, p < 0.001). Alder leaves showed the fastest rates (mean AFDM remaining of
Table 1. Location, reach characterisation and water physico-chemical features of the studied streams (mean ± SE; n = 5 – 6). For water temperature, daily mean values and their range are shown.
S1 S2 S3Latitude 43º 12′ 32″ N 42º 59′ 48″ N 42º 59′ 59″ NLongitude 3º 16′ 03″ W 2º 52′ 47″ W 2º 53′ 00″ WBasin (ha) 225 361 357Altitude (m a.s.l) 315 420 400Reach slope (%) 20.5 14.2 13.3Width (m) 3.52 3.27 3.73Tree canopy cover (%) 42.2 50.5 55.05Flow (l s–1) 72.7 ± 15.5 46.1 ± 10.4 98.2 ± 32.6Water temperature (°C) 6.92 6.39 5.96
(2.85 –10.45) (2.06 – 9.62) (1.35 – 9.46)SRP (µg P l–1) 14.4 ± 1.2 16.9 ± 1.3 15.8 ± 0.8Nitrate (µg N l–1) 600.1 ± 59.2 33.1 ± 6.8 35.3 ± 8.3Nitrite (µg N l–1) 1.1 ± 0.1 0.8 ± 0.1 0.8 ± 0.1Ammonium (µg N l–1) 34.0 ± 5.4 34.1 ± 7.7 32.9 ± 5.7pH 7.10 ± 0.12 7.45 ± 0.13 7.58 ± 0.08Alkalinity (meq l–1) 0.43 ± 0.04 1.23 ± 0.09 0.69 ± 0.10Conductivity (µS cm–1) 134.0 ± 7.7 171.2 ± 7.3 120.4 ± 10.2FPIM (mg l–1) 1.22 ± 0.38 0.92 ± 0.21 1.35 ± 0.38FPOM (mg l–1) 1.21 ± 0.15 0.51 ± 0.13 0.50 ± 0.14% O2 saturation 103.5 ± 4.2 99.9 ± 3.9 107.2 ± 2.4

242 Aingeru Martínez, Aitor Larrañaga, Javier Pérez, Ana Basaguren and Jesús Pozo
the three streams 43.7 %), followed by poplar (mean AFDM remaining 56.4 %), oak (mean AFDM remain-ing 65.8 %) and finally plane (mean AFDM remaining 88.1 %) and pine (mean AFDM remaining 89.9 %). Only rates for alder and oak showed differences among streams (ANOVAAld: F2,12 = 12.41, p = 0.001; ANOVAOak: F2,12 = 6.35, p = 0.013). The leaves of
the five species showed differences in their initial C and N concentrations (ANOVAC: F4,20 = 12.72, p < 0.001; ANOVAN: F4,20 = 453.98, p < 0.001), C:N ratio (ANOVA: F4,20 = 64.05, p < 0.001) and percent-age of ash (ANOVA: F4,20 = 94.58, p < 0.001), with alder showing the highest and plane and pine the low-est overall quality (Table 2). Mass loss rate positively correlated with initial %N and negatively with the C:N ratio of material (Fig. 2). After in-stream incubation, the %C diminished, especially in oak. The incubated leaves showed differences in %C content (Fig. 3; ANOVA: F4,60 = 51.08, p < 0.001), being lowest in oak and highest in pine. Leaf N concentrations after incu-bation were higher than initial ones, particularly for alder, and maintained the initial observed differences among leaf species, with alder showing the highest concentration and pine the lowest (Fig. 3; ANOVA: F4,60 = 630.09, p < 0.001). The increase in leaf %N was more evident in the stream with more dissolved nitro-gen availability (S1). After incubation, leaves showed differences in the C:N ratio among species (ANOVA: F4,60 = 318.75, p < 0.001; Pine > Plane > Poplar = Oak > Alder) and among streams (Fig. 3; ANOVA: F2,60 = 19.41, p < 0.001; S2 = S3 > S1). Ash content also
Fig. 1. Mass loss rates (%AFDM dd–1; mean ± SE) of the dif-ferent leaf species in the three streams. Statistical differences (ANOVA and Tukey post-hoc) among leaf species (letters) and among streams (* and +) are shown.
Fig. 2. Mass loss rate vs. initial leaf %N or C:N ratio (mean values).
Table 2. Initial percentage of C, N, ash and atomic C:N ratio of leaf species. Letters denote statistical differences (ANOVA and Tukey post-hoc; n = 5).
Leaf %C %N C:N % ash contentAlder 51.2a 3.05a 19.66c 4.54c
Poplar 43.9c 0.74b 69.25b 4.56bc
Oak 46.0bc 0.79b 68.61b 10.02a
Plane 48.8ab 0.68bc 84.45ab 6.21b
Pine 49.5a 0.59c 99.93a 2.53d
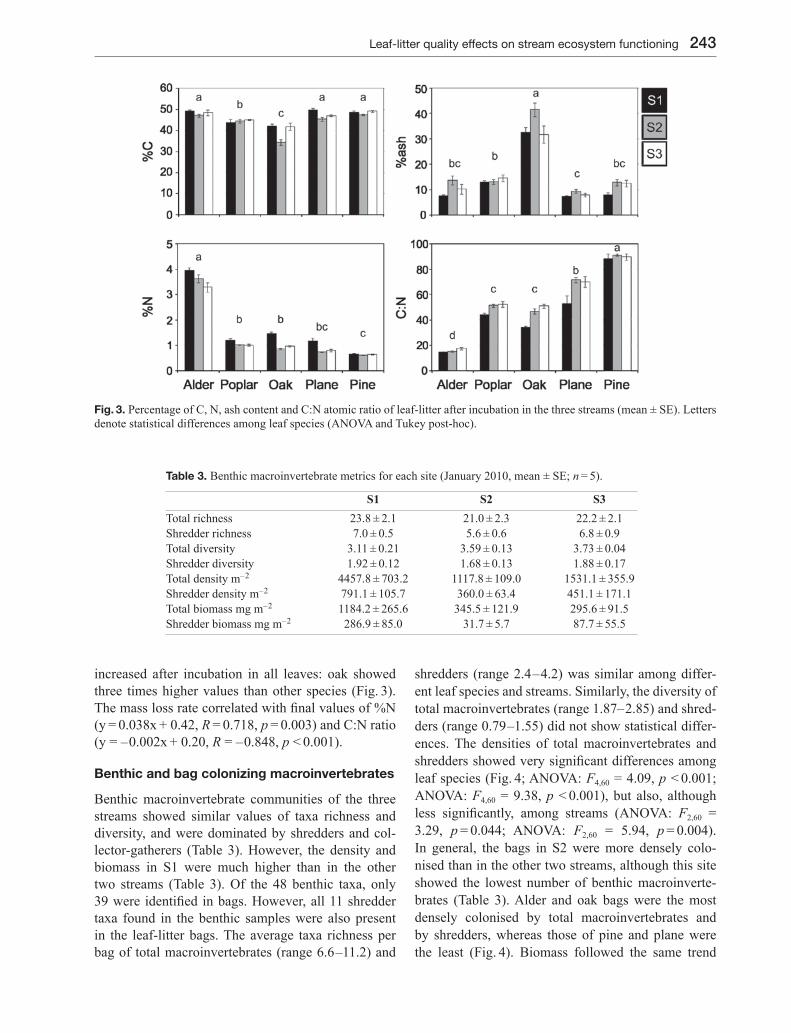
243Leaf-litter quality effects on stream ecosystem functioning
increased after incubation in all leaves: oak showed three times higher values than other species (Fig. 3). The mass loss rate correlated with final values of %N (y = 0.038x + 0.42, R = 0.718, p = 0.003) and C:N ratio (y = – 0.002x + 0.20, R = – 0.848, p < 0.001).
Benthic and bag colonizing macroinvertebrates
Benthic macroinvertebrate communities of the three streams showed similar values of taxa richness and diversity, and were dominated by shredders and col-lector-gatherers (Table 3). However, the density and biomass in S1 were much higher than in the other two streams (Table 3). Of the 48 benthic taxa, only 39 were identified in bags. However, all 11 shredder taxa found in the benthic samples were also present in the leaf-litter bags. The average taxa richness per bag of total macroinvertebrates (range 6.6 –11.2) and
shredders (range 2.4 – 4.2) was similar among differ-ent leaf species and streams. Similarly, the diversity of total macroinvertebrates (range 1.87– 2.85) and shred-ders (range 0.79 –1.55) did not show statistical differ-ences. The densities of total macroinvertebrates and shredders showed very significant differences among leaf species (Fig. 4; ANOVA: F4,60 = 4.09, p < 0.001; ANOVA: F4,60 = 9.38, p < 0.001), but also, although less significantly, among streams (ANOVA: F2,60 = 3.29, p = 0.044; ANOVA: F2,60 = 5.94, p = 0.004). In general, the bags in S2 were more densely colo-nised than in the other two streams, although this site showed the lowest number of benthic macroinverte-brates (Table 3). Alder and oak bags were the most densely colonised by total macroinvertebrates and by shredders, whereas those of pine and plane were the least (Fig. 4). Biomass followed the same trend
Fig. 3. Percentage of C, N, ash content and C:N atomic ratio of leaf-litter after incubation in the three streams (mean ± SE). Letters denote statistical differences among leaf species (ANOVA and Tukey post-hoc).
Table 3. Benthic macroinvertebrate metrics for each site (January 2010, mean ± SE; n = 5).
S1 S2 S3Total richness 23.8 ± 2.1 21.0 ± 2.3 22.2 ± 2.1Shredder richness 7.0 ± 0.5 5.6 ± 0.6 6.8 ± 0.9Total diversity 3.11 ± 0.21 3.59 ± 0.13 3.73 ± 0.04Shredder diversity 1.92 ± 0.12 1.68 ± 0.13 1.88 ± 0.17Total density m– 2 4457.8 ± 703.2 1117.8 ± 109.0 1531.1 ± 355.9Shredder density m– 2 791.1 ± 105.7 360.0 ± 63.4 451.1 ± 171.1Total biomass mg m– 2 1184.2 ± 265.6 345.5 ± 121.9 295.6 ± 91.5Shredder biomass mg m– 2 286.9 ± 85.0 31.7 ± 5.7 87.7 ± 55.5

244 Aingeru Martínez, Aitor Larrañaga, Javier Pérez, Ana Basaguren and Jesús Pozo
of density (Fig. 4), and showed differences for total macroinvertebrates and shredders among leaf spe-cies (ANOVA: F4,60 = 6.01, p < 0.001; ANOVA: F4,60 = 4.41, p = 0.003) and streams (ANOVA: F2,60 = 5.82, p = 0.005; ANOVA: F2,60 = 6.11, p = 0.004). Communi-
ties on leaf-litter showed a higher relative abundance and biomass of shredders than benthic ones. Among leaf species, alder bags showed higher relative abun-dances of shredders than plane (ANOVA: F4,60 = 2.75, p = 0.036). Shredders correlated negatively with initial
Fig. 4. Density and biomass of total macroinvertebrates and shredders per bag (mean ± SE). Results of the Tukey post-hoc test among leaves are shown. Note the different scales. Biomass in pine bags of stream S1 is strongly influenced by a single large T ipulidae that appeared in one of the five replicates (cross lined bars). In order to avoid its effect, the mean biomass value consid-ering the other four replicas is displayed (black bars). Statistical results do not change when considering four or five replicates.
Fig. 5. Mass loss rates vs. log10 shredder density or biomass. Shredder biomass in pine bags of stream S1 is bracketed as it was strongly influenced by a single large Tipulidae that appeared in one of the five replicates. In order to avoid its effect, the mean bio-mass value considering the other four replicates is displayed and was used for the regressions

245Leaf-litter quality effects on stream ecosystem functioning
C:N ratio, in terms of density (y = – 0.006×+1.66, R2 = 0.32, p = 0.027) and biomass (y = – 0.009×+1.19, R2 = 0.39, p = 0.013), and helped to explain the observed mass loss rate variability (Fig. 5).
Discussion
Mass loss rates differed among leaf species indepen-dently of the stream and species ranking remained constant in the three streams. Only alder and oak pro-cessing rates showed significant differences among streams, which were unrelated to the availability of nutrients in the water and the abundance of consum-ers. Thus, leaf identity appears to be more important than inter-stream differences in determining process-ing rates, as has been noted previously (Leroy & Matks 2006, Arroita et al. 2012). Among the numerous characteristics of the leaves that modulate breakdown rate, previous studies have demonstrated that initial ni-trogen concentration and/or the C:N ratio can become main drivers of decomposition (e.g. Enriquez et al. 1993, Richardson et al. 2004, Hladyz et al. 2009, Muto et al. 2011). Indeed, in the present study, leaves with higher nitrogen content and lower C:N ratio broke down faster than those with a lower nitrogen content and higher ratio. The nitrogen concentration increased and the C:N ratio decreased with the incubation in the five leaf litter species. This is in accordance with pre-vious studies in the area that observe an increment of nitrogen and a reduction of C:N until the middle of the process , i.e. 50 % of decomposition (Pozo 1993, Pérez et al. 2012), a threshold only slightly surpassed by alder in our experiment. This quality enhancement has been often attributed to microbial growth (Can-hoto & Graça, 2008, Webster et al. 2009) that removes dissolved nutrients from the water column and incor-porates them into microbial biomass (Gusewell & Gessner 2009). Among the leaf species, alder showed the highest absolute increase in nitrogen concentra-tion, probably due to a higher fungal colonisation. This species has characteristics that favour microbial colonisation, such as non-cereous soft cuticles and a low concentration of inhibitory compounds (Canhoto & Graça 1999). In pine needles, contrarily, the tiny changes in nitrogen concentration suggest a lower fun-gal colonisation due to the hard cuticle and antifungal agents present in conifer needles (Bärlocher & Oertli 1978). Despite these changes in N concentration and C:N ratio during incubation, the differences in quality among leaf species and the relationship between leaf quality and processing rates remained. Nevertheless,
some leaf-specific response of the processing rate was also observed: despite the similar nutritional quality between poplar and oak, decomposition was slower in oak. Other studies observed no relationship between breakdown rate and nitrogen concentration (Gessner & Chauvet 1994, Royer & Minshall 2001, Lecerf & Chauvet 2008), attributing the variability of process-ing to other leaf characteristics such as toughness, lignin or phosphorous concentration that modulate the consumption rate of leaf litter by consumers. We lack information about microbial assemblages on leaves in the present study, but the higher leaf nitrogen enrich-ment occurring in the stream with the highest dissolved nitrogen (S1) might be attributed to a higher immobi-lization of nutrients by microorganisms. Nevertheless, mass loss rates in this stream were not stimulated in comparison to the other two streams, contrary to what is usually observed (Suberkropp et al. 2010, Pérez et al. 2012). This suggests that although leaf nitrogen concentration could be a successful descriptor of the breakdown rate, a combination of factors, including the biotic community, are needed to better explain it, as found by other authors (Schindler & Gessner 2009, Ferreira et al. 2012).
The total invertebrate and shredder assemblages in bags represented a subset of the benthic community, although the relative contribution of shredders to the macroinvertebrate assemblage was higher in bags, re-gardless of the leaf species, than in the benthos. This concentration of shredders in bags has a fundamen-tally methodological explanation, i.e. the incubation of litter in bags promotes shredder colonisation due to food resource concentration (Tiegs et al. 2008). The density and biomass of this feeding group were positively related to nitrogen content, as reported in other studies (Haapala et al. 2001, Hladyz et al. 2009, Kominoski & Pringle 2009), and this resource selec-tion helps to explain the observed breakdown rates. Nevertheless, the similar density and biomass of in-vertebrates (total and shredders) between alder and oak were not in accordance with their differences in quality and mass loss rates. It is often unclear whether invertebrates found in litter bags are consuming leaves or seeking refuge (Hofer & Richardson 2007). In this context, the stiffness of the Pyrenean oak leaves, that create interstices when packed, and their hairy under-side, that accumulate sediments, make this species the best candidate for a high quality habitat for macroin-vertebrates among the five materials considered in the present study. Actually, the higher ash content in oak leaves is a sign of the higher sediment accumula-tion onto this material. This sedimentation might limit

246 Aingeru Martínez, Aitor Larrañaga, Javier Pérez, Ana Basaguren and Jesús Pozo
macroinvertebrate activity and slow down leaf pro-cessing rates, as other authors note (Niyogi et al. 2003, Rabeni et al. 2005). On the other hand, the large C:N imbalance between shredders and leaf resources (see Cross et al. 2005, Hladyz et al. 2009) might explain the shredder preference for high quality leaf species, such as alder, that more easily fulfil their nutritional demands (Sterner & Elser 2002, Woodward 2009). In essence, the preference of macroinvertebrates to-wards alder and oak leaves might indicate that these two species would be the ones that will support highly abundant macroinvertebrate assemblages in nature. The substitution of native alder by plane or poplar in riparian corridors, and the replacement of native oak forests by pine plantations in the rest of the basins can be viewed as detrimental for macroinvertebrates and, by extension, to other biotic assemblages and overall ecosystem functioning. A recent study relates pine monocultures with a reduction of the processing capacity of leaf litter and a change of the descriptors of the macroinvertebrate communities (Martínez et al. 2013). Similarly, functional impairment has been at-tributed to the presence of plane in the riparian cor-ridor (Menéndez et al. 2013), although effects at the community level are still unknown. We think that more studies that focus on the effects of specific tree species on freshwater systems are needed. The present and other similar studies should become basic refer-ences for natural resources conservation managers when projecting land use changes to minimize their impact on stream ecosystems.
Acknowledgements
This study was funded by the Spanish Ministry of Science and Innovation (project CGL2007– 66664-C04 – 01) and by the Basque Government (Research grants IT-422 – 07 and IT-302 –10). The authors thank the technicians of SGIker’s SCAB Service, of the University of The Basque Country, UPV/EHU, for the nitrate measurements. Grants were awarded to A. Mar-tínez by the Basque Government and to J. Pérez by the Univer-sity of the Basque Country. We are grateful to Ana Caballero and Liseth Castellanos for field and laboratory assistance. We thank the Gorbeia Natural Park for the sampling permits. The authors want to thank anonymous reviewers for their help in improving the manuscript.
References
Abelho, M., 2001: From litterfall to breakdown in streams: A review. – The Scientific World 1: 656 – 680.
APHA (American Public Health Association), 2005: Standard Methods for the Examination of Water and Wastewater, 21st ed. – American Public Health Association, American Wa-ter Works Association, and Water Environment Federation, Washington.
Arroita, M., Aristi, I., Flores, L., Larrañaga, A., Díez, J., Mora, J., Romaní, A. M. & Elosegi, A., 2012: The use of wooden sticks to assess stream ecosystem functioning: Compari-son with leaf breakdown rates. – Sci. Total Environ. 440: 115 –122.
Bärlocher, F. & Oertli, J. J., 1978: Inhibitors of aquatic hy-phomycetes in dead conifer needles. – Arch. Hydrobiol. 81: 462 – 474.
Canhoto, C. & Graça, M. A. S., 1999: Leaf barriers to fungal colonization and shredders (Tipula lateralis) consumption of decomposing Eucalyptus globulus. – Microb. Ecol. 37: 163 –172.
Canhoto, C. & Graça, M. A. S., 2008: Interactions between fungi and stream invertebrates: back to the future. – In: Srid-har, S., Bärlocher, F. & Hyde, K. D. (eds): Novel Techniques and Ideas in Mycology. – Fungal Diversity Research Series 20: 305 – 325, Hong Kong University Press, Yunnan.
Casas, J. J., Larrañaga, A., Menéndez, M., Pozo, J., Basaguren, A., Martínez, A., Pérez, J., González, J. M., Mollá, S., Cas-ado, C., Descals, E., Roblas, N., López-González, J. A. & Va-lenzuela, J. L., 2013: Leaf litter decomposition of native and introduced tree species of contrasting quality in headwater streams: How does the regional setting matter? – Sci. Total Environ. 458 – 460: 197– 208.
Cross, W. F., Benstead, J. P., Frost, P. C. & Thomas, S. A., 2005: Ecological stoichiometry in freshwater benthic sys-tems: recent progress and perspectives. – Freshw. Biol. 50: 1895 –1912.
Dang, C. K., Gessner, M. O. & Chauvet, E., 2007: Influence of conidial traits and leaf structure on attachment success of aquatic hyphomycetes on leaf litter. – Mycologia 99: 24 – 32.
Enriquez, S., Duarte, C. M. & Sand-Jensen, K., 1993: Patterns in decomposition rates among photosynthetic organisms: the importance of detritus C:N:P content. – Oecologia 94: 457– 471.
FAO, 2010: Global forest resources assessment. Main report, no 163. – Food and Agriculture Organization of the United Nations, Rome 20101– 340.
Ferreira, V., Encalada, A. C. & Graça, M. A. S., 2012: Effects of litter diversity on decomposition and biological coloniza-tion of submerged litter in temperate and tropical streams. – Freshw. Sci. 31: 945 – 962.
Gessner, M. O. & Chauvet, E., 1994: Importance of stream mi-crofungi in controlling breakdown rates of leaf-litter. – Ecol-ogy 75: 1807–1817.
Gulis, V. & Suberkropp, K., 2003: Leaf litter decomposition and microbial activity in nutrient enriched and unaltered reaches of a headwater stream. – Freshw. Biol. 48: 123 –134.
Güsewell, S. & Gessner, M. O., 2009: N:P ratios influence lit-ter decomposition and colonization by fungi and bacteria in microcosms. – Funct. Ecol. 23: 211– 219.
Haapala, A., Muotka, T. & Markkola, A., 2001: Breakdown and macroinvertebrate and fungal colonization of alder, birch, and willow leaves in a boreal forest stream. – J. N. Am. Benthol. Soc. 20: 395 – 407.
Herrera, M. & Campos, J. A., 2010: Flora alóctona invasora en Bizkaia. Instituto para la Sostenibilidad de Bizkaia. – Diputación Foral de Bizkaia, pp. 1–196.
Hieber, M. & Gessner, M. O., 2002: Contribution of stream det-rivores, fungi, and bacteria to leaf breakdown based on bio-mass estimates. – Ecology 84: 1026 –1038.
Hladyz, S., Gessner, M. O., Giller, P. S., Pozo, J. & Woodward, G., 2009: Resource quality and stoichiometric constraints on stream ecosystem functioning. – Freshw. Biol. 54: 957– 970.

247Leaf-litter quality effects on stream ecosystem functioning
Hofer, N. & Richardson, J. S., 2007: Comparisons of the colo-nisation by invertebrates of three species of wood, alder leaves, and plastic “leaves” in a temperate stream. – Int. Rev. Hydrobiol. 6: 647– 655.
Kearns, S. G. & Bärlocher, F., 2008: Leaf surface roughness influences colonization success of aquatic hyphomycete co-nidia. – Fungal Ecol. 1: 13 –18.
Kominoski, J. S. & Pringle, C. M., 2009: Resource–consumer diversity: testing the effects of leaf litter species diversity on stream macroinvertebrate communities. – Freshw. Biol. 54: 1461–1473.
Larrañaga, A., Basaguren, A. & Pozo, J., 2009: Impacts of Eu-calyptus globulus plantations on physiology and population densities of invertebrates inhabiting Iberian Atlantic streams. – Int. Rev. Hydrobiol. 94: 497– 511.
Larrañaga, A., Basaguren, A., Elosegi, A. & Pozo, J., 2009: Im-pacts of Eucalyptus globulus plantations on Atlantic streams: changes in invertebrate density and shredder traits. – Fun-dam. Appl. Limnol. 175: 151–160.
Lecerf, A. & Chauvet, E., 2008: Intra-specific variability in leaf traits strongly affects alder leaf decomposition in a stream. – Basic Appl. Ecol. 9: 598 – 607.
Leroy, C. J. & Marks, J. C., 2006: Litter quality, stream char-acteristics and litter diversity influence decomposition rates and macroinvertebrates. – Freshw. Biol. 51: 605 – 617.
Martínez, A., Larrañaga, A., Pérez, J., Descals, E., Basaguren, A. & Pozo, J., 2013: Effects of pine plantations on structural and functional attributes of forested streams. – Forest Ecol. Manage. 310: 147–155.
Menéndez, M., Descals, E., Riera, T. & Moya, O., 2013: Do non-native Platanus hybrida riparian plantations affect leaf litter decomposition in streams? – Hydrobiologia DOI 10.1007/s10750 – 013 –1539 – 0.
Merritt, R. W. & Cummins, K. W., 2007: An Introduction to the Aquatic Insects of North America. – Kendall/Hunt Publish-ing Company, Dubuque.
Molinero, J. & Pozo, J., 2006: Organic matter, nitrogen and phosphorus fluxes associated with leaf litter in two small streams with different riparian vegetation: a budget ap-proach. – Arch. Hydrobiol. 166: 143 –165.
Molinero, J., Pozo, J. & González, E., 1996: Litter breakdown in streams of the Agüera catchment: influence of dissolved nutrients and land use. – Freshw. Biol. 36: 745 –756.
Moore, J. C., Berlow, E. L., Coleman, D. C., de Ruiter, P. C., Dong, Q., Hastings, A., Collins, Johnson, N., MacCann, K. S., Melville, K., Morin, P. J., Nadelhoffer, K., Rosemond, A. D., Post, D. M., Sabo, J. L., Scow, K. M., Vanni, M. J. & Wall, D. H., 2004: Detritus, trophic dynamics and biodiver-sity. – Ecol Lett 7: 584 – 600.
Muto, E. A., Kreutzweiser, D. P. & Sibley, P. K., 2011: Over-winter decomposition and associated macroinvertebrate com-munities of three deciduous leaf species in forest streams on the Canadian Boreal Shield. – Hydrobiologia 658: 111–126.
Niyogi, D. K., Simon, K. S. & Townsend, C. R., 2003: Break-down of tussock grass in streams along a gradient of agri-cultural development in New Zealand. – Freshw. Biol. 48: 1698 –1708.
Pérez, J., Descals, E. & Pozo, J., 2012: Aquatic hyphomycete communities associated with decomposing alder leaf litter in reference headwater streams of the Basque Country (north-ern Spain). – Microb. Ecol. 64: 279 – 290.
Pozo, J., 1993: Leaf litter processing of alder and eucalyp-tus in the Agüera stream system (North Spain) I. Chemical changes. – Arch. Hydrobiol. 127: 299 – 317.
Pozo, J., González, E., Díez, J. R., Molinero, J. & Elósegui, A., 1997: Inputs of particulate organic matter to streams with different riparian vegetation. – J. N. Am. Benthol. Soc. 16: 602 – 611.
R Development Core Team, 2010: R: A Language and Environ-ment for Statistical Computing. – R Foundation for Statisti-cal Computing: Vienna.
Rabeni, C. F., Doisy, K. E. & Zweig, L. D., 2005: Stream inver-tebrate community functional responses to deposited sedi-ment. – Aquat. Sci. 67: 395 – 402.
Reinhart, K. O. & VandeVoort, R., 2006: Effect of native and exotic leaf litter on macroinvertebrate communities and de-composition in a western Montana stream. – Divers Distrib. 12: 776 –781.
Richardson, J. S., Shaughnessy, C. R. & Harrison, P. G., 2004: Litter breakdown and invertebrate association with three types of leaves in a temperate rainforest stream. – Arch. Hy-drobiol. 159: 309 – 325.
Royer, T. V. & Minshall, G. W., 2001: Effects of nutrient en-richment and leaf quality on the breakdown of leaves in a hardwater stream. – Freshw. Biol. 46: 603 – 610.
Schindler, M. H. & Gessner, M. O., 2009: Functional leaf traits and biodiversity effects on litter decomposition in a stream. – Ecology 90: 1641–1649.
Sterner, R. W. & Elser, J. J., 2002: Ecological Stoichiometry: The Biology of Elements From Molecules to the Biosphere. – Princeton University Press, Princeton.
Suberkropp, K., Gulis, V., Rosemond, A. D. & Benstead, J. P., 2010: Ecosystem and physiological scales of microbial re-sponses to nutrients in a detritus-based stream: results of a 5-year continuous enrichment. – Limnol. Oceanog. 55: 149 –160.
Swan, C. M. & Palmer, M. A., 2006: Composition of speciose leaf litter alters stream detritivore growth, feeding activity and leaf breakdown. – Oecologia 147: 469 – 478.
Tachet, H., Richoux, P., Bournaud, M. & Usseglio-Polatera, P., 2002: Invertébrés d’eau Douce: Systématique, Biologie et Écologie. – CNRS, Paris.
Tank, J. L., Rosi-Marshall, E. J., Griffiths, N. A., Entrekin, S. A. & Stephen, M. L., 2010: A review of allochthonous organic matter dynamics and metabolism in streams. – J. N. Am. Benthol. Soc. 29: 118 –146.
Tibbets, T. M. & Molles, M. C., 2005: C:N:P stoichiometry of dominant riparian trees and arthropods along the Middle Rio Grande. – Freshw. Biol. 50: 1882 –1894.
Tiegs, S. D., Peter, F. D., Robinson, C. T., Uehlinger, U. & Gess-ner, M. O., 2008: Leaf decomposition and invertebrate colo-nization responses to manipulated litter quantity in streams. – J. N. Am. Benthol. Soc. 27: 321– 331.
Wallace, J. B., Eggert, S. L., Meyer, J. L. & Webster, J. R., 1997: Multiple trophic levels of a forest stream linked to terrestrial litter inputs. – Science 277: 102 –104.
Webster, J. R., Benfield, E. F., Ehrman, T. P., Schaeffer, M. A., Tank, J. L., Hutchens, J. J. & D’Angelo, D. J., 1999: What happens to allochthonous material that falls into streams? A synthesis of new and published information from Coweeta. – Freshw. Biol. 41: 687–705.

248 Aingeru Martínez, Aitor Larrañaga, Javier Pérez, Ana Basaguren and Jesús Pozo
Webster, J. R., Newbold, J. D., Thomas, S. A., Valett, H. M. & Mulholland, P. J., 2009: Nutrient uptake and mineralization during leaf decay in streams – a model simulation. – Int. Rev. Hydrobiol. 94: 372 – 390.
Weijters, M. J., Janse, J. H., Alkemade, R. & Verhoeven, T. A., 2009: Quantifying the effect of catchment land use and wa-ter nutrient concentrations on freshwater river and stream
biodiversity. – Aquat. Conserv.: Mar. Freshw. Ecosyst. 19: 104 –112.
Woodward, G., 2009: Biodiversity, ecosystem functioning and freshwater food webs: assembling the jigsaw puzzle. – Freshw. Biol. 54: 2171– 2187.
Zar, J. H., 2010: Biostatistical Analysis, 5th ed. – Pearson Pren-tice Hall, Upper Saddle River.
Submitted: 11 June 2013; accepted: 18 October 2013.




















