
Le Bars D, Willer JC (2008). Plasticité de la douleur : rôle des contrôles inhibiteurs diffus....
19
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus D. Le Bars, J.-C. Willer La douleur s’élabore au sein d’un cerveau imprégné par son passé. En particulier, les événements somesthésiques antérieurs, qu’ils soient ou non douloureux, sont intégrés dans l’élaboration de la douleur présente. C’est l’ensemble de ces événements qui, de concert avec la proprioception et l’équilibration, permet à chacun de construire progressivement et inconsciemment son schéma corporel, étape essentielle de l’édification biologique du soi. Sa consolidation progressive et son incessante restructuration peuvent cependant être remises en cause par de nombreux facteurs biologiques et psychologiques. On peut faire l’expérience de la distorsion de ce schéma dans certaines situations particulières. Ainsi, les viscères peuvent, du moins pour certains d’entre eux, prendre le pouvoir au sein du schéma corporel. Le besoin pressant d’uriner ou de déféquer, que l’on qualifiera « d’impérieux » lorsque l’on ne peut le satisfaire, distord notre schéma corporel au point que notre être entier se résume à ce besoin physiologique. Le schéma corporel se situe dans un « interland » lâche entre l’inconscient et le conscient. Tapi dans la monotonie du « normal », il se dilue dans l’inconscient, mais il ne demande qu’à se « réveiller » à la moindre occasion, soit vers le plaisir, soit vers la douleur. Les douleurs intenses, les douleurs qui durent, les douleurs qui évoluent vers la chronicité figurent probablement parmi les causes physiques les plus courantes de perturbation du schéma corporel : elles focalisent l’attention sur une partie du corps au détriment des autres, le schéma se déformant par une « sur-représentation » du foyer douloureux. Les neurones à convergence de la corne postérieure de la moelle sont capables de saisir la globalité des informations issues à la fois de l’interface avec le milieu extérieur (la peau) et du milieu intérieur (les viscères, les muscles). Sur la base d’expériences électrophysiologiques effectuées chez l’animal et chez l’homme, nous défendons la thèse suivante : « l’activité somesthésique de base » constituée par l’ensemble de ces informations joue un rôle important dans l’élaboration du schéma corporel, du moins dans sa composante qui complète le schéma postural. Un foyer douloureux, qui active un sous-ensemble segmentaire de neurones à convergence et inhibe le reste de la population, perturbe profondément cette activité. Il en résulte une déformation du schéma corporel au profit du foyer douloureux qui s’extrait de l’activité de fond et devient surdimensionné. © 2008 Elsevier Masson SAS. Tous droits réservés. Mots clés : Douleur ; Corne postérieure ; Neurones à convergence ; Réflexes nociceptifs ; Plasticité ; CIDN ; Schéma corporel Plan ¶ Introduction 1 ¶ Douleur et mémoire du corps 2 ¶ Les nocicepteurs auscultent notre corps sans relâche 2 ¶ Les informations résultant de cette auscultation sont distribuées au cerveau dans son ensemble 2 ¶ Douleur et schéma corporel 3 ¶ Des neurones spinaux au schéma corporel 3 ¶ Plasticité des phénomènes segmentaires 4 ¶ Action perturbatrice d’un foyer douloureux sur l’activité somesthésique de base 6 ¶ Sommation spatiale et temporelle 9 ¶ Influences mutuelles des foyers douloureux 9 ¶ Un modèle chez l’homme 11 ¶ Neurologie des « contrôles inhibiteurs diffus nociceptifs » 14 ¶ Conclusion 15 ■ Introduction Dans l’élaboration d’une perception que nous identifions comme douloureuse, le cerveau entrelace sensation, émotion et cognition. Pour souligner cette complexité, « l’International Association for the Study of Pain » (IASP) reconnaît le rôle central de l’affect dans la définition qu’elle donne de la douleur : « une expérience sensorielle et émotionnelle désagréa- ble, associée à une lésion tissulaire réelle ou potentielle, ou décrite en termes d’une telle lésion ». Cette définition met explicitement l’accent sur la variabilité de la relation entre lésion et douleur et intègre les dimensions sensorielle, affective et cognitive de l’expérience douloureuse. La douleur ne s’exprime pas sur un terrain vierge et dépend de l’état préalable du sujet tant sur les plans biologique et médical que de plans dont la complexité est d’un ordre supérieur : affectif, émotion- nel et motivationnel. Ces derniers, qui appartiennent à la sphère psychologique, voire sociologique, sont une source supplémentaire de difficultés pour quantifier la douleur. Si l’on s’en tient au plan médical, l’influence psychologique de l’observateur, du médecin, de l’acte médical ou du médicament - voire du sorcier, du chaman, de l’envoûteur... - est essentielle ¶ 17-003-G-10 1 Neurologie
-
Upload
sorbonne-fr -
Category
Documents
-
view
0 -
download
0
Transcript of Le Bars D, Willer JC (2008). Plasticité de la douleur : rôle des contrôles inhibiteurs diffus....

Plasticité de la douleur :rôle des contrôles inhibiteurs diffus
D. Le Bars, J.-C. Willer
La douleur s’élabore au sein d’un cerveau imprégné par son passé. En particulier, les événementssomesthésiques antérieurs, qu’ils soient ou non douloureux, sont intégrés dans l’élaboration de la douleurprésente. C’est l’ensemble de ces événements qui, de concert avec la proprioception et l’équilibration,permet à chacun de construire progressivement et inconsciemment son schéma corporel, étapeessentielle de l’édification biologique du soi. Sa consolidation progressive et son incessanterestructuration peuvent cependant être remises en cause par de nombreux facteurs biologiques etpsychologiques. On peut faire l’expérience de la distorsion de ce schéma dans certaines situationsparticulières. Ainsi, les viscères peuvent, du moins pour certains d’entre eux, prendre le pouvoir au sein duschéma corporel. Le besoin pressant d’uriner ou de déféquer, que l’on qualifiera « d’impérieux » lorsquel’on ne peut le satisfaire, distord notre schéma corporel au point que notre être entier se résume à cebesoin physiologique. Le schéma corporel se situe dans un « interland » lâche entre l’inconscient et leconscient. Tapi dans la monotonie du « normal », il se dilue dans l’inconscient, mais il ne demande qu’àse « réveiller » à la moindre occasion, soit vers le plaisir, soit vers la douleur. Les douleurs intenses, lesdouleurs qui durent, les douleurs qui évoluent vers la chronicité figurent probablement parmi les causesphysiques les plus courantes de perturbation du schéma corporel : elles focalisent l’attention sur unepartie du corps au détriment des autres, le schéma se déformant par une « sur-représentation » du foyerdouloureux. Les neurones à convergence de la corne postérieure de la moelle sont capables de saisir laglobalité des informations issues à la fois de l’interface avec le milieu extérieur (la peau) et du milieuintérieur (les viscères, les muscles). Sur la base d’expériences électrophysiologiques effectuées chezl’animal et chez l’homme, nous défendons la thèse suivante : « l’activité somesthésique de base »constituée par l’ensemble de ces informations joue un rôle important dans l’élaboration du schémacorporel, du moins dans sa composante qui complète le schéma postural. Un foyer douloureux, qui activeun sous-ensemble segmentaire de neurones à convergence et inhibe le reste de la population, perturbeprofondément cette activité. Il en résulte une déformation du schéma corporel au profit du foyerdouloureux qui s’extrait de l’activité de fond et devient surdimensionné.© 2008 Elsevier Masson SAS. Tous droits réservés.
Mots clés : Douleur ; Corne postérieure ; Neurones à convergence ; Réflexes nociceptifs ; Plasticité ; CIDN ;Schéma corporel
Plan
¶ Introduction 1
¶ Douleur et mémoire du corps 2
¶ Les nocicepteurs auscultent notre corps sans relâche 2
¶ Les informations résultant de cette auscultation sont distribuéesau cerveau dans son ensemble 2
¶ Douleur et schéma corporel 3
¶ Des neurones spinaux au schéma corporel 3
¶ Plasticité des phénomènes segmentaires 4
¶ Action perturbatrice d’un foyer douloureux sur l’activitésomesthésique de base 6
¶ Sommation spatiale et temporelle 9
¶ Influences mutuelles des foyers douloureux 9
¶ Un modèle chez l’homme 11
¶ Neurologie des « contrôles inhibiteurs diffus nociceptifs » 14
¶ Conclusion 15
■ IntroductionDans l’élaboration d’une perception que nous identifions
comme douloureuse, le cerveau entrelace sensation, émotion etcognition. Pour souligner cette complexité, « l’InternationalAssociation for the Study of Pain » (IASP) reconnaît le rôlecentral de l’affect dans la définition qu’elle donne de ladouleur : « une expérience sensorielle et émotionnelle désagréa-ble, associée à une lésion tissulaire réelle ou potentielle, oudécrite en termes d’une telle lésion ». Cette définition metexplicitement l’accent sur la variabilité de la relation entrelésion et douleur et intègre les dimensions sensorielle, affectiveet cognitive de l’expérience douloureuse. La douleur nes’exprime pas sur un terrain vierge et dépend de l’état préalabledu sujet tant sur les plans biologique et médical que de plansdont la complexité est d’un ordre supérieur : affectif, émotion-nel et motivationnel. Ces derniers, qui appartiennent à lasphère psychologique, voire sociologique, sont une sourcesupplémentaire de difficultés pour quantifier la douleur. Si l’ons’en tient au plan médical, l’influence psychologique del’observateur, du médecin, de l’acte médical ou du médicament- voire du sorcier, du chaman, de l’envoûteur... - est essentielle
¶ 17-003-G-10
1Neurologie

comme en témoigne la puissance de l’effet placebo. En un mot,la douleur ne s’élabore pas au sein d’un cerveau amnésique maisd’un système nerveux central imprégné par son passé, qu’il soitrécent ou plus lointain. La mémoire de ce passé concernel’individu dans son ensemble ainsi que ses relations avec lemonde extérieur, mais c’est la mémoire du corps qui constituele socle sur lequel se bâtit la douleur présente.
■ Douleur et mémoire du corpsD’innombrables facteurs contribuent aux disparités entre
l’étendue apparente d’une lésion et l’intensité de la douleurressentie, mais ils sont à l’évidence gouvernés par la thymie.L’importance de l’anxiété vis-à-vis de l’expérience douloureusea été fréquemment soulignée. L’anesthésiste américain HenryBeecher, qui a participé pendant la Seconde Guerre mondialeaux combats les plus meurtriers de la campagne d’Italie,notamment au débarquement et à la bataille d’Anzio en janvier1944, avait été frappé d’accueillir dans son hôpital de campagnedes soldats profondément blessés, polytraumatisés, qui nesemblaient guère souffrir. [1] Revenu à la vie civile, il a puconstater que des blessures comparables, mais provoquées dansun contexte bien différent, étaient ressenties de façon atroce-ment douloureuse. [2] Alors que la survenue d’un accident estune terrible catastrophe qui peut faire basculer une vie, l’éva-cuation vers un hôpital de campagne, à l’arrière de l’enfer descombats, délivre le soldat d’une terrible menace. Sa douleurpeut alors s’estomper au point de ne pas nécessiter de mor-phine. Le contexte cognitif peut être la source de contrôlesphysiologiques très puissants de la douleur dont nous commen-çons à entrevoir les mécanismes [3, 4]. La part biologique de ceséléments cognitifs est constituée par la mémoire du corps. Lesévénements somesthésiques antérieurs, qu’ils soient douloureux(mémoire de la douleur, anticipation de la douleur probable,etc.) ou non, sont intégrés dans l’élaboration de la douleurprésente. Bien que l’ensemble de la sensibilité corporelle -extéroceptive, intéroceptive et proprioceptive - soit concerné àcet égard, nous privilégierons le rôle particulier qu’y jouent lesnocicepteurs.
■ Les nocicepteurs auscultent notrecorps sans relâche
La douleur se différencie fondamentalement des autressystèmes sensoriels car on ne peut caractériser, sur le planstructural, de récepteurs spécialisés dans la détection desstimulus nociceptifs. Les messages nociceptifs naissent auniveau de terminaisons libres amyéliniques, constituant desarborisations dans les tissus. Ils sont ensuite transmis par desfibres nerveuses périphériques vers la moelle où s’effectue lepremier relais. Les axones de ces nocicepteurs sont faiblement -fibres Ad - ou ne sont pas - fibres C - entourés d’une gaine demyéline.
Parmi les fibres C, qui constituent 60 à 90 % de l’ensembledes fibres afférentes cutanées et la quasi-totalité des fibresafférentes viscérales, le groupe le plus important est sansconteste celui des nocicepteurs polymodaux [5]. On les désigneainsi car ils répondent à des stimulus nociceptifs de différentesnatures, thermique, mécanique et chimique. En réalité lecaractère « polymodal » de l’immense majorité de ces nocicep-teurs est plus étendu car ils sont également activés par desstimulations thermiques ou mécaniques qui ne provoquent pasde douleur [6]. Leur seuil thermique moyen est d’environ 40 °Cet des stimulus puissants quoique non douloureux, comme lefrottement d’une corde de guitare ou la friction avec un gant decrin, les activent énergiquement. Le caractère primitif des fibresC polymodales, peu spécialisées et totipotentes, a été souligné [7]
et l’on peut remarquer que de tels récepteurs existent déjà chezles invertébrés comme l’aplysie ou la sangsue [8]. Des « nocicep-teurs » peu spécifiques ont donc subi l’évolution des espècessans perdre leurs principaux caractères, probablement parce queces derniers leur confèrent une fonction indispensable à la
survie des individus. On doit sans doute les considérer dans leurensemble comme un organe sensoriel qui, sans relâche, « aus-culte » l’ensemble de notre corps. Cette auscultation estméticuleuse puisque la densité moyenne de terminaisons libresdans notre peau n’est jamais inférieure à 600/cm2. Elle est aussitrès redondante car les champs périphériques des fibres Cpolymodales se recouvrent très largement les uns les autres, cequi signifie que le moindre stimulus en active une vastepopulation. En résumé, si leur rôle est prépondérant pourdéclencher la douleur, ce n’est vraisemblablement pas leur seulefonction.
Quant aux fibres nociceptives Ad, elles sont bien moinsnombreuses et, dans l’ensemble, on peut aussi les considérercomme polymodales. Leur seuil d’activation est cependant plusélevé que leurs homologues appartenant aux fibres C (seuilthermique moyen ~ 45 °C) [9]. En bon ingénieur, la sélectionnaturelle a privilégié, en combinant les nocicepteurs C et Ad, unsystème redondant « à double détente » et à « double niveau » :si l’agression physique s’installe lentement, il est lent à réagir,mais le fait pour des températures relativement basses. Lorsquel’agression physique s’installe brusquement, il est beaucoup plusrapide à réagir, mais pour des stimulus plus intenses.
D’ordinaire à l’état de « veille physiologique », cet organed’auscultation peut se mobiliser localement pour alerter avecefficacité et précision le système nerveux central d’une menaceplanant sur l’organisme. Son fonctionnement change alorsradicalement de configuration dans l’aire concernée. Lesnocicepteurs réagissent en effet à de nombreuses substanceschimiques : ce sont des chémorécepteurs. Cette fonctionchémoréceptrice est déterminante pour provoquer les douleursinflammatoires : l’inflammation prolonge l’activation desnocicepteurs et surtout les transforme pour les rendre plussensibles [10]. On peut à cet égard évoquer l’existence d’unsystème d’alarme secondaire, chargé de nous informer de l’étatde dégradation d’une partie de notre corps en la rendant plussensible ; c’est ce qu’on dénomme « hyperalgésie ». Sa constantede temps est supérieure à celle du système d’alarme primaireactivé par l’agression initiale. Ces fonctions d’alarme sontprofondément perturbées lors de neuropathies périphériques.
■ Les informations résultantde cette auscultationsont distribuées au cerveaudans son ensemble
À la différence du système lemniscal qui implique des fibresmyélinisées rapides (fibres Ab) puis seulement deux synapsespour atteindre le cortex, le système à l’origine de la douleur estdiversifié et divergent. Dans les nerfs, le message est acheminéplus lentement, puis il musarde dans la moelle épinière avecquelques relais dans la corne postérieure avant de se distribuerdans de nombreuses structures cérébrales, notamment laformation réticulée, le thalamus et plusieurs aires corticales. Encouplant les techniques d’imagerie fonctionnelle par résonancemagnétique nucléaire et de tomographie par émission depositons [11-14], on a pu montrer que les cortex somesthésiquesprimaire et secondaire étaient bien activés par des stimulationsnociceptives mais dans une moindre mesure que les cortexcingulaire antérieur et insulaire qui, appartenant au systèmelimbique, jouent un rôle primordial dans la genèse des émo-tions. C’est en fin de compte un ensemble d’aires cérébrales quiconcourent probablement à l’élaboration de la perception de ladouleur. La notion de « centre » de la douleur est d’ailleursabandonnée depuis longtemps, le « cerveau douloureux », refletdu « corps douloureux », tendant de plus en plus à envahir deszones qui, bien que circonscrites, sont disséminées dans lecerveau tout entier.
Elles incluent notamment des régions cérébrales plus archaï-ques au niveau du tronc cérébral et de l’hypothalamus. Leuractivation donne lieu à une série de réactions et de contrôles enretour vers la moelle dont le résultat peut se schématiser enréactions végétatives liées au stress. Il convient de souligner ici
17-003-G-10 ¶ Plasticité de la douleur : rôle des contrôles inhibiteurs diffus
2 Neurologie

la forte intrication du « système de la douleur », qu’on pourraitcroire plus ou moins spécifique et spécialisé, avec le systèmevégétatif [15-18]. Ainsi, la douleur aiguë est-elle associée àl’augmentation du rythme cardiaque, de la pression artérielle, àune vasoconstriction périphérique, etc. Ces mécanismes sonttrès fortement contrôlés par l’amygdale, ce qui signifie qu’ilssont influencés par les émotions et les fonctions les plus élevéesdu cerveau. Au travers de ces voies, il existe des connexionsanatomiques précises pour que « le mental » influence lesfonctions végétatives liées à la douleur, voire la douleur elle-même. Au total, sensation et émotion sont intimement liéesl’une à l’autre ainsi qu’aux processus végétatifs, même s’il estdes cas particuliers pathologiques qui permettent de lesdissocier.
■ Douleur et schéma corporelLes récepteurs affectés à cette « auscultation » permanente
envoient donc sans cesse à notre cerveau des informationsprécises concernant notre corps. De concert avec le systèmevestibulaire, l’ensemble de ces informations corporelles estintégré pour synthétiser en permanence des représentationsmentales inconscientes de la réalité physiologique du soi. Laneuropsychologie moderne y adjoint les autres sens, notam-ment la vision, pour définir la notion d’intégration multisenso-rielle [19]. C’est le neurologue Henry Head qui, dès l’aube dusiècle dernier, avait postulé l’existence au sein de notre cerveaud’un schéma corporel, ou plus exactement d’une série d’aumoins deux schémas dont le plus connu est le schéma posturalqui, en quelque sorte, en constitue le « squelette » [20]. Leschéma postural est un modèle global de la position relative desdifférentes parties de notre corps en perpétuel réajustement parrapport aux postures précédentes. Le second qui, en quelquesorte, « habille » le premier des informations extéroceptives, esttout aussi essentiel car il permet notamment de localiser avecprécision sur notre corps une zone touchée, piquée, lésée...Henry Head l’avait décrit comme un modèle de la surface denotre corps, sans doute parce que cela correspondait à sapratique de neurologue, mais on ne doit pas le réduire à unmodèle de l’interface avec le milieu extérieur, la peau ; lesorganes internes participent sans doute aussi à sa construction.À la suite de Schilder [21], on regroupe souvent sous le terme deschéma corporel l’ensemble des deux modèles évoqués ci-dessus.Bien que les deux termes soient parfois utilisés indifféremment,il convient de distinguer le « schéma corporel », modèle cérébralinconscient de notre corps, de « l’image corporelle » qui en estune représentation mentale consciente [22]. Ces notions intéres-sent une large palette de disciplines, de la neurologie à lapsychanalyse [23-31].
Le processus de maturation du schéma corporel, très lent aucours des premiers mois de la vie pendant lesquels la confusionentre le monde intérieur et le monde extérieur est totale, maisqui s’accélère dans la petite enfance pour se stabiliser ultérieu-rement, concourt à la construction d’une « mémoire » du moiphysique. C’est par l’intermédiaire du pouvoir intégrateur dutemps qu’il faut peut-être concevoir les subtiles passerelles quirelient le moi « physiologique » au moi « psychologique » [32], leschéma corporel à l’image corporelle. Les expériences antérieu-res neutres, agréables (les caresses, ...) et désagréables (les bobos,...) concourent ensemble à bâtir cette mémoire. Sa consolidationprogressive peut cependant être remise en cause par de nom-breux facteurs biologiques et psychologiques. Des altérations dece schéma peuvent ainsi être déclenchées en quelques instantspar des situations expérimentales ingénieuses [33], mais lesdistorsions les plus spectaculaires sont d’origine neurologique :membre fantôme des amputés, syndrome d’Anton-Babinski(hémiasomatognosies), syndrome de Gerstmann (agnosiesdigitales), etc. [34].
De façon bien plus ordinaire, on peut faire l’expérience de ladistorsion de ce schéma dans certaines situations particulières.Ainsi, le prurit peut-il focaliser notre attention sur une zonecutanée restreinte qui devient alors la seule partie de notre corps
qui nous importe : on le dira « impérieux », ce qui résume bienson statut dictatorial. Les viscères, on le sait, n’évoquent jamaisde prurit mais peuvent aussi, du moins pour certains d’entreeux, prendre le pouvoir au sein du schéma corporel. Le besoinpressant d’uriner ou de déféquer, que l’on qualifiera d’ailleursaussi « d’impérieux » lorsque l’on ne peut le satisfaire, distordnotre schéma corporel au point que notre être entier se résumeà ce besoin physiologique. Sa satisfaction, les retrouvailles avecnotre schéma corporel d’ordinaire si discret, est vécue nonseulement comme une délivrance mais bien souvent comme unvéritable plaisir.
Certaines interventions médicales peuvent brouiller notreschéma corporel. C’est ce que font quotidiennement les dentis-tes lorsqu’ils anesthésient une partie de notre mâchoire : cetteanesthésie locale ne provoque pas la sensation d’un « trou »dans la mâchoire comme on pourrait s’y attendre puisque c’estl’ensemble des fibres afférentes qui est bloqué. Bien qu’insensi-ble, le territoire anesthésié est perçu comme une masse amor-phe beaucoup plus volumineuse qu’en réalité, bien plus« présente » que le reste de notre corps [35, 36]. Cette situationrévèle à notre conscience un schéma corporel « illusoire ».
Ces quelques exemples permettent une conclusion provi-soire : le schéma corporel se situe dans un « interland » lâcheentre l’inconscient et le conscient. Tapi dans la monotonie du« normal », il se dilue dans l’inconscient, mais il ne demandequ’à se « réveiller » à la moindre occasion, soit vers le plaisir,soit vers la douleur. Les douleurs intenses, les douleurs quidurent, les douleurs qui évoluent vers la chronicité figurent eneffet probablement parmi les causes physiques les plus couran-tes de perturbation du schéma corporel : elles construisent le« corps douloureux » en focalisant l’attention sur une partie ducorps au détriment des autres [37, 38]. Le schéma corporel sedégage alors de l’inconscience relative en se déformant par une« sur-représentation » du foyer douloureux. Les expériencessexuelles malheureuses, notamment lorsqu’elles sont doulou-reuses, leur disputent sans doute la priorité. Nous laisserons auxpsychologues le soin de décider lequel de ces facteurs de ladouleur, du sexe ou de toutes autres causes psychologiquesprédomine dans cette incessante reconstruction du schémacorporel, pour aborder ce problème du point de vue réducteurdu physiologiste.
■ Des neurones spinaux au schémacorporel
Des données expérimentales obtenues chez l’animal suggèrentque la sensation douloureuse pourrait ne pas seulement résulterde la simple augmentation du niveau d’activité neuronale ausein d’un système câblé, unidirectionnel et spécifique, maisqu’elle serait plutôt générée par des gradients d’activité au seinde populations neuronales. C’est à l’étage médullaire que cesgradients ont été le plus étudiés. Des mécanismes périphériques,spinaux et supraspinaux concourent à leur organisation. C’estprobablement une transmission en volume qu’il faut considérercomme le message significatif destiné in fine aux centrescérébraux responsables de l’élaboration de la sensation doulou-reuse. Ce volume s’inscrit dans le cadre plus général d’unereprésentation du corps qui n’est ni une simple « photogra-phie » dont chaque pixel correspondrait à une surface corporelleélémentaire, ni même une « photographie déformée » dont lataille des pixels varierait, à l’instar de « l’homonculus » dePenfield, avec la localisation des territoires corporels représentés.
Les propriétés des « neurones à convergence » de la cornepostérieure de la moelle illustrent bien cette notion [39, 40]. Leurchamp récepteur cutané excitateur, c’est-à-dire la zone cutanéequi les active, présente un gradient de sensibilité (Fig. 1, 2A) :dans leur partie centrale, toute stimulation active le neurone,qu’elle soit nociceptive ou non (mise en jeu de fibres Ab, Adet/ou C) ; dans une zone plus périphérique, seules les stimula-tions nociceptives mettant en jeu des fibres Ad et/ou C déclen-chent une activité neuronale [42]. Compte tenu du recouvrementdes champs excitateurs, l’organisation spatiale de la convergencejoue un rôle essentiel dans l’élaboration du message issu de
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus ¶ 17-003-G-10
3Neurologie

cette classe de neurones (Fig. 2B). En effet, appliqué sur unterritoire donné, un stimulus tactile n’activera qu’un nombrerestreint de neurones, ceux dont le centre du champ excitateurest situé sur ce territoire. En revanche, appliqué exactement aumême endroit, un stimulus nociceptif activera non seulementces mêmes neurones, mais également ceux dont les marges duchamp excitateur incluent ce territoire. Ce n’est donc passeulement en simples termes d’activité qu’il faut raisonner pourtenter de comprendre le rôle de ces neurones, mais aussi entermes de volume global d’information qu’une populationneuronale émet vers les structures cibles, notamment cérébra-les [43, 44]. En outre, certains de ces neurones sont activés depuisdes territoires corporels de natures différentes, par exemplecutané et viscéral ou cutané et musculaire [45-48]. C’est par cetteconvergence qu’on explique le phénomène de douleur référée,comme l’irradiation douloureuse vers le membre supérieurgauche dans l’angine de poitrine. Elle concerne également lesecond type de neurones médullaires impliqués dans la trans-mission des messages nociceptifs, les neurones nociceptifsspécifiques, qui sont essentiellement localisés dans la couche Ide la moelle et ne répondent qu’à l’activation des fibres Adet C.
La signification de ces phénomènes de convergence pourraitdépasser ces curiosités cliniques, par ailleurs parfois bien utilesau diagnostic. On peut en effet souligner cette capacité desneurones à convergence de saisir la globalité des informationsissues de l’interface avec le milieu extérieur (la peau) et dumilieu intérieur (les viscères, les muscles). L’ensemble de cesinformations constitue une « activité somesthésique de base »,dont le rôle fonctionnel reste à déterminer. On peut imaginerque ces neurones ne sont pas seulement impliqués dans lesprocessus douloureux mais que, en transmettant au cerveaucette « activité somesthésique de base », ils signalent qu’aucuneperturbation particulière de l’organisme n’est apportée par lemilieu extérieur ou intérieur. Dans cette conception, l’activitésomesthésique de base n’est plus alors un simple « bruit », maisassiste le système lemniscal dans l’élaboration du « schémacorporel », du moins pour la part qui complète le schémapostural.
■ Plasticité des phénomènessegmentaires
Concevoir ce système en termes de réseaux dynamiquessemble encore plus impératif lorsque l’on tient compte du faitque la taille des champs excitateurs de ces neurones est suscep-tible d’être modifiée. La convergence anatomique des influx surun même neurone est en réalité beaucoup plus importante quecelle que l’on constate sur le plan fonctionnel dans les condi-tions normales. En effet, aux marges du champ excitateur d’unneurone, il existe une zone dont la stimulation entraîne bienune activation cellulaire sous la forme d’une dépolarisation,mais cette dernière est insuffisante pour déclencher des poten-tiels d’action qui, rappelons-le, sont les seuls messages adressésaux centres supérieurs. Le concept de « synapses quiescentes »s’est progressivement développé [49-51] sur la base de l’idée quecette frange de neurones activés de façon sous-liminairepouvait, du moins dans certaines conditions, constituer unmécanisme déterminant de la fonction de transfert de l’infor-mation vers les centres supérieurs. Lorsque l’on utilise lastimulation électrique qui synchronise les influx afférents, onconstate par exemple que le champ excitateur de ces neuronesest plus vaste que celui que l’on observe lorsqu’on applique unstimulus naturel thermique ou mécanique [52]. On peut aussiagrandir cette surface par des stimulations préalables de forteintensité ou un traitement pharmacologique dépolarisant lamembrane des neurones, en appliquant par exemple sur samembrane un acide aminé excitateur ou un antagoniste d’acideaminé inhibiteur [53, 54]. Enfin, et cela est essentiel sur le planclinique, la taille des champs augmente au cours de l’inflamma-tion, qu’elle soit aiguë ou chronique [55]. Sous le terme de« sensibilisation centrale », on regroupe ces phénomènes desensibilisation, que l’on peut également déclencher par somma-tion temporelle [56-58]. Au total, la surface potentielle deschamps excitateurs est plus vaste que celle que l’on observedans les conditions physiologiques normales (Fig. 2C), et leurrecouvrement potentiel en est augmenté d’autant (Fig. 2D).Comme ce dernier concerne aussi leur partie centrale, les
Champ cutané : - champ excitateur (CE) - champ inhibiteur (CI)
CerveauChamp viscéral
Champ musculaire
Coupsde pinceau
41 °C
200 Hz
1 min
43 °C 46 °C 48 °C
fibres Aβ et Aδfibres Aβ
fibres Aδ et C
CE CI
(-)(+)(+)(+)
Figure 1. Organisation schématique des influences périphériques s’exerçant sur les neurones à convergence. Leur champ périphérique est complexe. Ilcomprend toujours une partie cutanée, elle-même composée : d’un champ excitateur (CE) dont le centre (zone rouge et verte) est activé par des stimulusnociceptifs et non nociceptifs et la périphérie (zone rouge) n’est activée que par des stimulations intenses ; et très souvent d’un champ inhibiteur (CI, zonejaune) essentiellement activé que par des stimulus non nociceptifs, surtout s’ils sont appliqués de façon répétitive et rapide (frottements, vibrations, ...). Ilcomprend aussi fréquemment une partie viscérale et parfois musculaire qui ne sont activées que par des stimulus à caractère nociceptif. On constate unesingulière convergence d’informations sur un même neurone. On peut soulager la douleur en renforçant ces contrôles inhibiteurs par des méthodes physiquesde stimulation : frottements, stimulation électrique, etc. D’instinct, on sait mettre en route ces mécanismes lorsque par exemple on se frotte la peau avecénergie pour soulager la douleur déclenchée par une piqûre d’insecte. Ces méthodes font partie de l’arsenal thérapeutique des médecines populairestraditionnelles. Un enregistrement de l’activité d’un neurone à convergence de la moelle lombaire de rat est représenté dans le cartouche. Il montre desréponses à des stimulus appliqués au centre du champ excitateur. La stimulation non nociceptive réitérée (coups de pinceau) active de façon importante leneurone tandis que les applications de chaleur radiante (indiquées par des barres) évoquent des réponses croissantes quand la température augmente. Modifiéd’après [39].
17-003-G-10 ¶ Plasticité de la douleur : rôle des contrôles inhibiteurs diffus
4 Neurologie
A
Cerveau
B
Cerveau
C
Cerveau
D
Cerveau
E
Cerveau
F
Cerveau
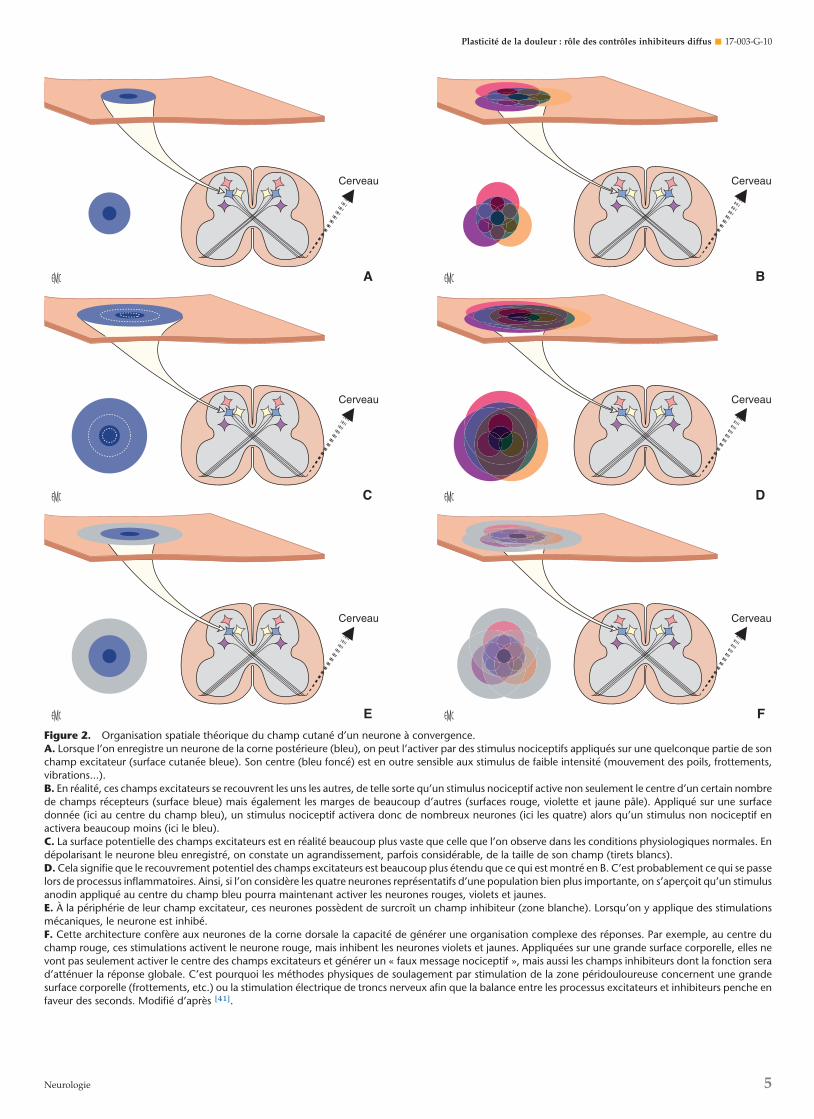
Figure 2. Organisation spatiale théorique du champ cutané d’un neurone à convergence.A. Lorsque l’on enregistre un neurone de la corne postérieure (bleu), on peut l’activer par des stimulus nociceptifs appliqués sur une quelconque partie de sonchamp excitateur (surface cutanée bleue). Son centre (bleu foncé) est en outre sensible aux stimulus de faible intensité (mouvement des poils, frottements,vibrations...).B. En réalité, ces champs excitateurs se recouvrent les uns les autres, de telle sorte qu’un stimulus nociceptif active non seulement le centre d’un certain nombrede champs récepteurs (surface bleue) mais également les marges de beaucoup d’autres (surfaces rouge, violette et jaune pâle). Appliqué sur une surfacedonnée (ici au centre du champ bleu), un stimulus nociceptif activera donc de nombreux neurones (ici les quatre) alors qu’un stimulus non nociceptif enactivera beaucoup moins (ici le bleu).C. La surface potentielle des champs excitateurs est en réalité beaucoup plus vaste que celle que l’on observe dans les conditions physiologiques normales. Endépolarisant le neurone bleu enregistré, on constate un agrandissement, parfois considérable, de la taille de son champ (tirets blancs).D. Cela signifie que le recouvrement potentiel des champs excitateurs est beaucoup plus étendu que ce qui est montré en B. C’est probablement ce qui se passelors de processus inflammatoires. Ainsi, si l’on considère les quatre neurones représentatifs d’une population bien plus importante, on s’aperçoit qu’un stimulusanodin appliqué au centre du champ bleu pourra maintenant activer les neurones rouges, violets et jaunes.E. À la périphérie de leur champ excitateur, ces neurones possèdent de surcroît un champ inhibiteur (zone blanche). Lorsqu’on y applique des stimulationsmécaniques, le neurone est inhibé.F. Cette architecture confère aux neurones de la corne dorsale la capacité de générer une organisation complexe des réponses. Par exemple, au centre duchamp rouge, ces stimulations activent le neurone rouge, mais inhibent les neurones violets et jaunes. Appliquées sur une grande surface corporelle, elles nevont pas seulement activer le centre des champs excitateurs et générer un « faux message nociceptif », mais aussi les champs inhibiteurs dont la fonction serad’atténuer la réponse globale. C’est pourquoi les méthodes physiques de soulagement par stimulation de la zone péridouloureuse concernent une grandesurface corporelle (frottements, etc.) ou la stimulation électrique de troncs nerveux afin que la balance entre les processus excitateurs et inhibiteurs penche enfaveur des seconds. Modifié d’après [41].
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus ¶ 17-003-G-10
5Neurologie

stimulus non nociceptifs deviennent particulièrement efficaceset l’activité engendrée par cette population neuronale devientsource d’allodynie.
En outre, les neurones à convergence ne sont pas sous laseule dépendance d’influences excitatrices issues de la périphé-rie. On peut en effet déclencher des effets inhibiteurs puissantssur les neurones de la corne postérieure en appliquant desstimulations mécaniques sur une zone particulière, adjacente àla partie excitatrice de leur champ récepteur (Fig. 2E). Cettezone inhibitrice doit être considérée comme une composanteimportante du champ récepteur global du neurone [42]. Elle estactivée par des fibres périphériques Ab et Ad. Si l’on raisonne entermes de population neuronale (Fig. 2F), il faudra tenir comptedu recouvrement mutuel des champs excitateurs et inhibiteursdes neurones pour concevoir le rôle de l’organisation spatiale dela convergence dans l’élaboration des messages issus de cetteclasse de neurones [41, 43].
Cette organisation signifie que la stimulation d’un contingentde fibres active une population de neurones, ceux dont leschamps excitateurs sont situés dans le secteur d’où proviennentces fibres, et inhibe une autre population de neurones, ceuxdont les champs inhibiteurs se situent dans ce même secteur. Ceprocessus d’inhibition latérale confère à l’activité des neuronesà convergence la capacité d’envoyer une image contrastée auxstructures cibles. C’est ainsi que les réflexes de retrait sontorganisés de façon « modulaire ». Le muscle d’un membre - qu’ilsoit fléchisseur, extenseur ou autre (supinateur, pronateur) - secontracte lorsque la stimulation est appliquée sur une régionbien définie de la peau afin de l’éloigner de la source nocicep-tive. Ce même stimulus inhibe les muscles dont la contractionaurait pour conséquence de provoquer un mouvement inadé-quat du membre qui rapprocherait cette région de la sourcenociceptive [59].
L’organisation des champs inhibiteurs permet en outred’expliquer la raison pour laquelle l’application de multiplesstimulations non nociceptives sur une grande surface corporelle(incorporant par conséquent de nombreux « centres » dechamps excitateurs) ne génère pas un « faux message nocicep-tif », du moins dans des conditions non pathologiques. Cetteéventualité est prévenue chez l’organisme sain par la stimula-tion concomitante de nombreux champs inhibiteurs dontl’action concourt à une atténuation de la réponse globale de lapopulation. En un sens, il s’agit d’une forme de « lissage » del’activité somesthésique de base.
Au total, l’organisation spatiale de la convergence desinfluences excitatrices et inhibitrices joue probablement un rôleessentiel dans l’élaboration physiologique des messages issus decette classe de neurones. La plasticité potentielle de ces systèmespeut se révéler au cours de processus pathologiques capables dedésorganiser complètement cet équilibre entre processusexcitateurs et inhibiteurs. Une sensibilisation des mécanismesexcitateurs ou un déficit des mécanismes inhibiteurs se tradui-ront à la fois par une augmentation de l’activité d’une popula-tion de neurones concernés par le foyer douloureux et par unaccroissement de cette population. Cette information élaboréedans la moelle est ensuite transmise au cerveau, où elle estdécodée sous la forme d’une hyperalgésie. Un déficit desmécanismes inhibiteurs pourra se traduire également par ledéclenchement d’une importante activité neuronale par desstimulus anodins. Cette information, élaborée dans la moelle,puis transmise au cerveau, peut alors y être décodée sous laforme d’une « allodynie » (douleur déclenchée par un stimulusnon nociceptif), notamment par exemple sous sa forme dyna-mique déclenchée par le frôlement d’un bout de coton, unsymptôme caractéristique de certaines neuropathies.
■ Action perturbatrice d’un foyerdouloureux sur l’activitésomesthésique de base
La description de l’organisation segmentaire de l’informationnociceptive est cependant très insuffisante pour rendre compte
de l’ensemble des phénomènes qui envahissent la moelle avantd’atteindre le cerveau pour déclencher la douleur. On constate,en effet, qu’un stimulus nociceptif met également en jeu descontrôles inhibiteurs descendants bulbospinaux, qui vontprofondément modifier l’activité des neurones de la cornepostérieure médullaire et trigéminale pris dans leur globalité.Lorsqu’on applique un tel stimulus sur un territoire corporel, ondéclenche non seulement les mécanismes segmentaires quenous avons brièvement décrits ci-dessus, mais aussi des phéno-mènes inhibiteurs très puissants qui s’exercent sur les neuronesde la corne postérieure appartenant aux métamères qui, a priori,n’étaient pas concernés.
On constate en effet chez plusieurs espèces - rat, souris, chat,singe - que la très grande majorité des neurones à convergence(et quelques neurones spécifiquement nociceptifs) est trèsprofondément inhibée par des stimulations nociceptives appli-quées sur un quelconque territoire corporel, dans la mesure oùce dernier est distinct de leur champ récepteur excitateur [39]. LaFigure 3 montre [60, 61] des exemples d’enregistrements deneurones à convergence lombaires chez le rat. L’activité desneurones est évoquée, soit par pincement de leur champrécepteur, soit en appliquant un acide aminé excitateur parmicroélectrophorèse sur leur membrane. Ces activités sontfortement inhibées par stimulation nociceptive hétérotopiquemécanique - pincement du museau, de la patte postérieurecontrolatérale ou de la queue - ou thermique - immersion de laqueue sur un bain d’eau à 52 °C - (Fig. 3A). Ces exemplesmontrent que chez le rat, c’est le corps entier qui constitue unchamp récepteur inhibiteur des neurones à convergence. Cespropriétés ont été observées lors d’enregistrements de neuronesà convergence effectués à divers niveaux médullaires et dans lesnoyaux caudalis ou oralis du système trigéminal, ce qui suggèreun principe général d’organisation. On observe toujours unerelation claire entre l’intensité du stimulus conditionnant et lapuissance des inhibitions. Pour les stimulus intenses, les effetsinhibiteurs sont très puissants et persistent bien au-delà de leurdurée d’application. Dans certains cas, on peut observer uneabolition complète d’activité pendant une longue périodesuivant la fin du stimulus conditionnant (« switch-off ») ;l’activité initiale ne peut être rétablie que par une nouvellemanipulation du champ récepteur excitateur (« switch-on ») [62].Les stimulus non nociceptifs sont généralement inefficaces, maisune particularité doit être notée à propos des viscères : certainsstimulus, comme la distension du côlon ou de la vessie,produisent des inhibitions d’installation lente mais dont le seuilest inférieur aux intensités douloureuses [63, 64].
De façon réciproque, un stimulus nociceptif appliqué sur unterritoire corporel donné inhibe les neurones à convergence detous les segments de la corne postérieure et des noyaux caudaliset oralis du système trigéminal. La population de neuronesinhibés inclut des neurones de projection vers le thalamus etdes neurones participant aux réflexes polysynaptiques spinaux.Comme ils ne sont pas organisés de façon somatotopique, maisconcernent l’ensemble du corps, ces phénomènes ont étédénommés « contrôles inhibiteurs diffus nociceptifs » - CIDN- [39]. En résumé, les CIDN sont déclenchés par toutes lesstimulations nociceptives quelle qu’en soit la nature, le degréd’inhibition étant déterminé par son intensité. Lorsque lestimulus est puissant, les effets inhibiteurs peuvent perdurerau-delà de sa période d’application. On peut souligner enfin queles CIDN sont exacerbés lors de douleurs cliniques, par exemplechez un animal souffrant de monoarthrite, de polyarthrite oud’une mononeuropathie périphérique [65, 66] (Fig. 4).
Les CIDN sont déclenchés par l’activation des nocicepteurspériphériques reliés aux fibres de petit diamètre (fibres Ad et C)(Fig. 3B) et s’exercent au travers d’une boucle spino-bulbo-spinale dont les branches ascendantes et descendantes sontconfinées aux quadrants antérolatéraux et aux funiculuspostérolatéraux, respectivement (Fig. 5A). Le sommet de cetteboucle est confiné dans la partie la plus caudale du bulbe quiinclut notamment un noyau particulier, le subnucleus reticularisdorsalis (SRD), situé ventralement sous le nucleus cuneatusentre le noyau caudal du trijumeau et le noyau du tractussolitaire, dont les neurones pourraient jouer un rôle majeur
17-003-G-10 ¶ Plasticité de la douleur : rôle des contrôles inhibiteurs diffus
6 Neurologie
dans les processus qui régissent l’information nociceptive [67].Les CIDN s’exercent in fine par une dépolarisation de lamembrane des neurones à convergence. C’est la raison pourlaquelle ils affectent toutes les activités des neurones à conver-gence, qu’elles soient évoquées depuis la périphérie par desstimulus nociceptifs ou non nociceptifs, naturels ou non, oubien encore directement par application électrophorétiqued’acides aminés excitateurs sur leur membrane (Fig. 3).
Au total, un foyer nociceptif active un sous-ensemble segmen-taire de neurones à convergence de la corne postérieure de lamoelle et inhibe le reste de la population. Ce mécanisme amé-liore le rapport signal/bruit en augmentant le contraste entre lesactivités du foyer segmentaire de neurones activés et le silence dela population résiduelle (Fig. 5B). Tout se passe comme dans uneassemblée bruyante au sein de laquelle un intervenant ne se feraentendre qu’après avoir obtenu le silence. Les CIDN permettent
Stimulation du champ Électrophorèsede glutamate
Électrophorèse
Enregistrement
Champpériphérique
excitateur
Museau (pincement)
Patte(pincement) 10 Hz
1 min
Queue(pincement)
Queue (52 °C)
A
Électrophorèse
Enregistrement
100 mm
Champpériphérique
excitateur
(a)
(b)
(c)
B
20
800 ms6004002000-200
N
10
0
20
800 ms6004002000-200
N
10
0
20
800 ms6004002000-200
N
10
0
210 ms7 ms 330 ms25 ms
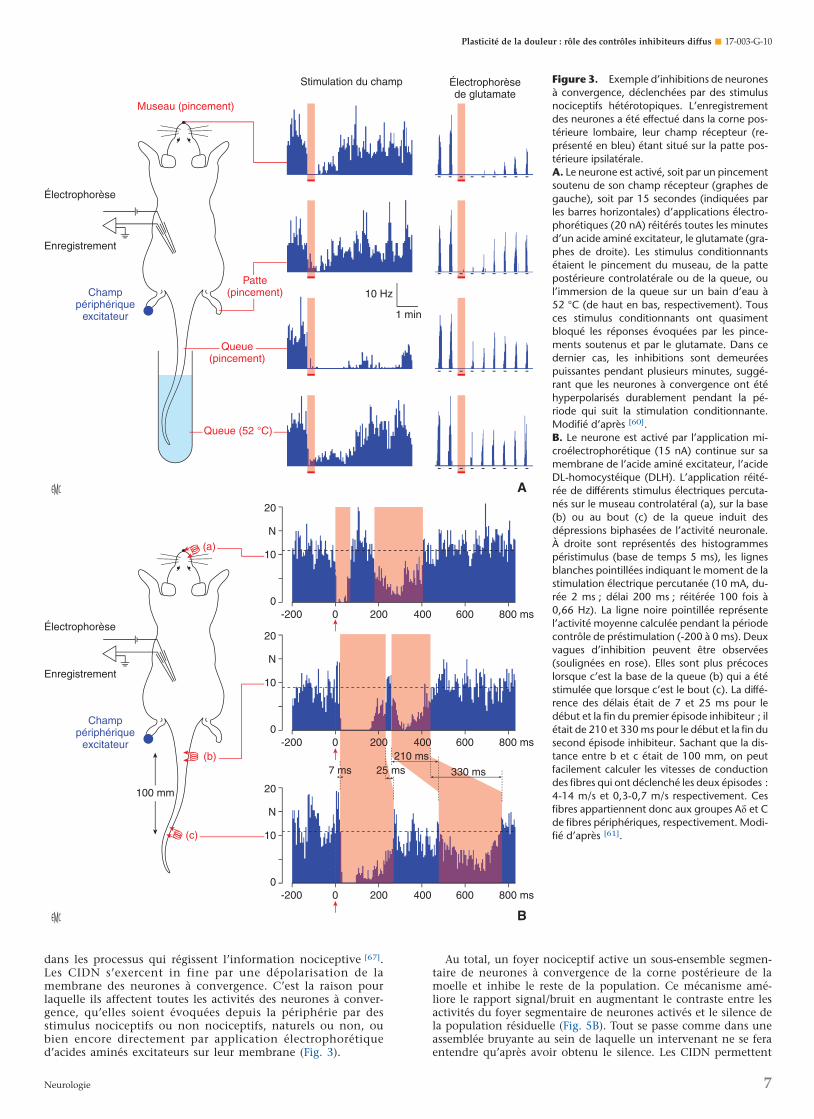
Figure 3. Exemple d’inhibitions de neuronesà convergence, déclenchées par des stimulusnociceptifs hétérotopiques. L’enregistrementdes neurones a été effectué dans la corne pos-térieure lombaire, leur champ récepteur (re-présenté en bleu) étant situé sur la patte pos-térieure ipsilatérale.A. Le neurone est activé, soit par un pincementsoutenu de son champ récepteur (graphes degauche), soit par 15 secondes (indiquées parles barres horizontales) d’applications électro-phorétiques (20 nA) réitérés toutes les minutesd’un acide aminé excitateur, le glutamate (gra-phes de droite). Les stimulus conditionnantsétaient le pincement du museau, de la pattepostérieure controlatérale ou de la queue, oul’immersion de la queue sur un bain d’eau à52 °C (de haut en bas, respectivement). Tousces stimulus conditionnants ont quasimentbloqué les réponses évoquées par les pince-ments soutenus et par le glutamate. Dans cedernier cas, les inhibitions sont demeuréespuissantes pendant plusieurs minutes, suggé-rant que les neurones à convergence ont étéhyperpolarisés durablement pendant la pé-riode qui suit la stimulation conditionnante.Modifié d’après [60].B. Le neurone est activé par l’application mi-croélectrophorétique (15 nA) continue sur samembrane de l’acide aminé excitateur, l’acideDL-homocystéique (DLH). L’application réité-rée de différents stimulus électriques percuta-nés sur le museau controlatéral (a), sur la base(b) ou au bout (c) de la queue induit desdépressions biphasées de l’activité neuronale.À droite sont représentés des histogrammespéristimulus (base de temps 5 ms), les lignesblanches pointillées indiquant le moment de lastimulation électrique percutanée (10 mA, du-rée 2 ms ; délai 200 ms ; réitérée 100 fois à0,66 Hz). La ligne noire pointillée représentel’activité moyenne calculée pendant la périodecontrôle de préstimulation (-200 à 0 ms). Deuxvagues d’inhibition peuvent être observées(soulignées en rose). Elles sont plus précoceslorsque c’est la base de la queue (b) qui a étéstimulée que lorsque c’est le bout (c). La diffé-rence des délais était de 7 et 25 ms pour ledébut et la fin du premier épisode inhibiteur ; ilétait de 210 et 330 ms pour le début et la fin dusecond épisode inhibiteur. Sachant que la dis-tance entre b et c était de 100 mm, on peutfacilement calculer les vitesses de conductiondes fibres qui ont déclenché les deux épisodes :4-14 m/s et 0,3-0,7 m/s respectivement. Cesfibres appartiennent donc aux groupes Ad et Cde fibres périphériques, respectivement. Modi-fié d’après [61].
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus ¶ 17-003-G-10
7Neurologie
d’obtenir un tel silence parmi les neurones spinaux. Inversement,le brouhaha ou le chahut ne permettront pas de réduire l’inter-venant au silence, mais brouilleront suffisamment son messagepour ne plus l’entendre. C’est ce que fait la morphine : les CIDNsont extrêmement sensibles à l’administration de faibles doses de
morphine [68]. Si « l’activité somesthésique de base » est bien àl’origine de certaines composantes du schéma corporel, onconçoit alors que ce dernier soit profondément perturbé par unfoyer douloureux, ce dernier occupant par rapport aux autres undomaine surdimensionné et prééminent.
Enregistrementdans le noyau
caudalisdu système trigéminal
Articulationsaine
Flexionde l’articulation
saine
Phase aiguë (48 h)
Phase chronique (25 j)
Flexionde l’articulation
arthritique
Arthritetibio-
métatarsienne
1 min
5 sp
ikes
Champ périphériqueexcitateur sur le museau
1 min
10 s
pike
s
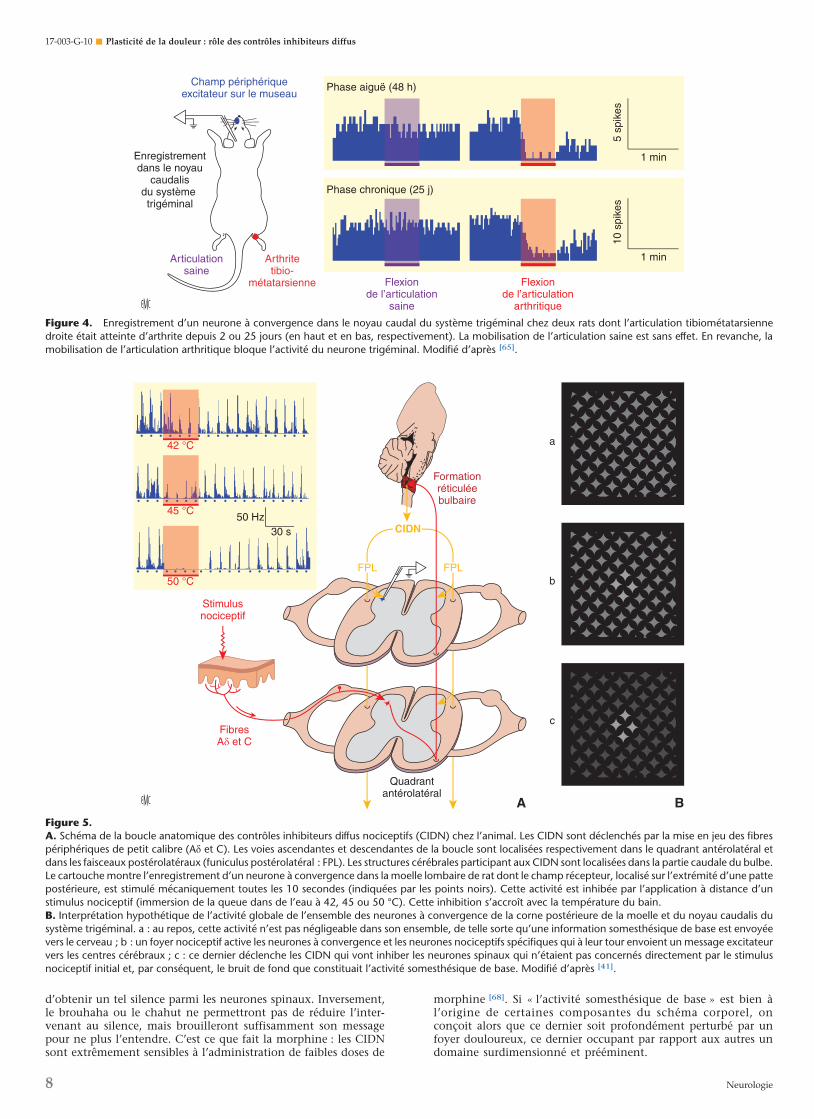
Figure 4. Enregistrement d’un neurone à convergence dans le noyau caudal du système trigéminal chez deux rats dont l’articulation tibiométatarsiennedroite était atteinte d’arthrite depuis 2 ou 25 jours (en haut et en bas, respectivement). La mobilisation de l’articulation saine est sans effet. En revanche, lamobilisation de l’articulation arthritique bloque l’activité du neurone trigéminal. Modifié d’après [65].
Formationréticuléebulbaire
a
b
c
Quadrantantérolatéral
42 °C
45 °C
50 °C
Stimulusnociceptif
FibresAδ et C
CIDN50 Hz
30 s
A B
FPL FPL
Figure 5.A. Schéma de la boucle anatomique des contrôles inhibiteurs diffus nociceptifs (CIDN) chez l’animal. Les CIDN sont déclenchés par la mise en jeu des fibrespériphériques de petit calibre (Ad et C). Les voies ascendantes et descendantes de la boucle sont localisées respectivement dans le quadrant antérolatéral etdans les faisceaux postérolatéraux (funiculus postérolatéral : FPL). Les structures cérébrales participant aux CIDN sont localisées dans la partie caudale du bulbe.Le cartouche montre l’enregistrement d’un neurone à convergence dans la moelle lombaire de rat dont le champ récepteur, localisé sur l’extrémité d’une pattepostérieure, est stimulé mécaniquement toutes les 10 secondes (indiquées par les points noirs). Cette activité est inhibée par l’application à distance d’unstimulus nociceptif (immersion de la queue dans de l’eau à 42, 45 ou 50 °C). Cette inhibition s’accroît avec la température du bain.B. Interprétation hypothétique de l’activité globale de l’ensemble des neurones à convergence de la corne postérieure de la moelle et du noyau caudalis dusystème trigéminal. a : au repos, cette activité n’est pas négligeable dans son ensemble, de telle sorte qu’une information somesthésique de base est envoyéevers le cerveau ; b : un foyer nociceptif active les neurones à convergence et les neurones nociceptifs spécifiques qui à leur tour envoient un message excitateurvers les centres cérébraux ; c : ce dernier déclenche les CIDN qui vont inhiber les neurones spinaux qui n’étaient pas concernés directement par le stimulusnociceptif initial et, par conséquent, le bruit de fond que constituait l’activité somesthésique de base. Modifié d’après [41].
17-003-G-10 ¶ Plasticité de la douleur : rôle des contrôles inhibiteurs diffus
8 Neurologie
■ Sommation spatialeet temporelle
On peut se poser la question de savoir si les CIDN sontcapables d’influencer les réponses neuronales lorsque le foyernociceptif est unique. La taille de ce foyer dépendant despathologies qui lui ont donné naissance, le « volume globald’information nociceptive » primitivement adressé au systèmenerveux central varie en conséquence. Ce « volume d’informa-tion nociceptive » est a priori déterminé par l’intensité, lasurface et la durée d’application du stimulus.
Si les questions de l’intensité et de la durée d’application dustimulus ne posent pas de problèmes théoriques, en revanche,celles de la sommation spatiale méritent d’être examinées avecattention : comment se comportent les neurones de la cornepostérieure lorsque des stimulus nociceptifs de surface croissantesont appliqués ? Une telle application provoque deux effetsopposés chez l’animal : pour des surfaces restreintes de stimula-tion, inférieures à deux fois environ celles du champ cutanéexcitateur des neurones, la fonction de transfert est accéléra-trice, mais au-delà, les réponses sont progressivement contrecar-rées [69]. Comme les CIDN, ce phénomène est observé chezl’animal « entier », mais disparaît chez l’animal « spinal »(Fig. 6) [70]. Cela signifie qu’au-delà d’une surface critique destimulation, la réponse des neurones à convergence est inhibéepar la mise en jeu d’une boucle de rétroaction négative qui meten jeu les structures supraspinales.
Ces observations permettent d’expliquer le fait que chezl’homme, une stimulation thermique nociceptive appliquée surune surface croissante ne provoque une augmentation de lasensation douloureuse que dans une plage restreinte de surfaces.On sait que la sommation spatiale joue un rôle important dansl’élaboration des messages sensoriels. Ainsi en est-il de laperception de chaleur qui augmente de façon proportionnelle àla surface exposée [71, 72]. C’est aussi le cas de la douleur, dumoins pour des stimulations thermiques intéressant de petitessurfaces (≤ 15 cm2) [73-77]. Mais ce n’est plus le cas lorsque dessurfaces plus élevées sont concernées : on constate alors unesaturation des phénomènes de sommation spatiale [78-81]. D’unefaçon plus générale, ces observations pourraient constituer le
reflet expérimental de la difficulté clinique à établir un parallèleentre l’étendue apparente d’une lésion et l’intensité de ladouleur ressentie par le patient [2, 82].
On peut conclure que la survenue d’un foyer douloureux nemet en jeu une boucle de rétroaction négative fonctionnelle-ment opérante que lorsqu’un volume suffisant d’informationnociceptive lui a donné naissance. Ce volume est certes déter-miné par l’intensité et la surface du stimulus, mais il dépendaussi de sa durée. À cet égard, la question de la sommationtemporelle mérite aussi d’être posée et, de fait, la sommationtemporelle des influx nociceptifs est, elle aussi, capable dedéclencher des mécanismes inhibiteurs bulbospinaux [83]. Sur leplan clinique, il importe alors de déterminer la part relative desphénomènes purement spinaux d’hyperexcitabilité [84], notam-ment lorsqu’ils sont déclenchés par l’activation des récepteursN-méthyl-D-aspartate (NMDA) et celle des phénomènes inhibi-teurs qui mettent en jeu des structures supraspinales.
■ Influences mutuelles des foyersdouloureux
Si l’on admet l’importance du rôle des neurones à conver-gence dans la douleur, une seconde implication directe dumodèle est l’existence de phénomènes interactifs entre desmessages nociceptifs issus de territoires corporels éloignés et,partant, d’interactions entre douleurs d’origine topographiquedistincte (Fig. 7) [61]. On sait qu’un stimulus douloureux estcapable de diminuer, voire de masquer la douleur issue d’unfoyer situé sur une autre partie du corps. Ce phénomène estconnu depuis l’Antiquité comme en témoigne l’aphorismed’Hippocrate selon lequel « De deux souffrances survenant enmême temps, mais sur des points différents, la plus forte faittaire la plus faible ».
C’est sur la base de cette observation clinique simple, redé-couverte de façon cyclique à différentes époques [85], parfoisspontanément mise à profit par les patients, qu’ont été déve-loppées, au sein des médecines populaires ou à la marge de la« médecine officielle », de nombreuses méthodes thérapeutiquesà visée antalgique que l’on peut regrouper sous le terme anglo-saxon de « contre-stimulation » [86, 87]. Ainsi en Kabylie, les
Bainthermostaté(48 °C, 15 s)
1,9 cm2
4,8 cm2
7,5 cm2
18 cm2
Rat« intact »
Rat« spinal »
Rats « intacts »
Rats « spinaux »
1,9 cm2 4,8 cm2 7,5 cm2 18 cm2
125
15 s50 Hz
Hz
Surface (cm2)
*
**
100
75
500 5 10 15 20
A B C
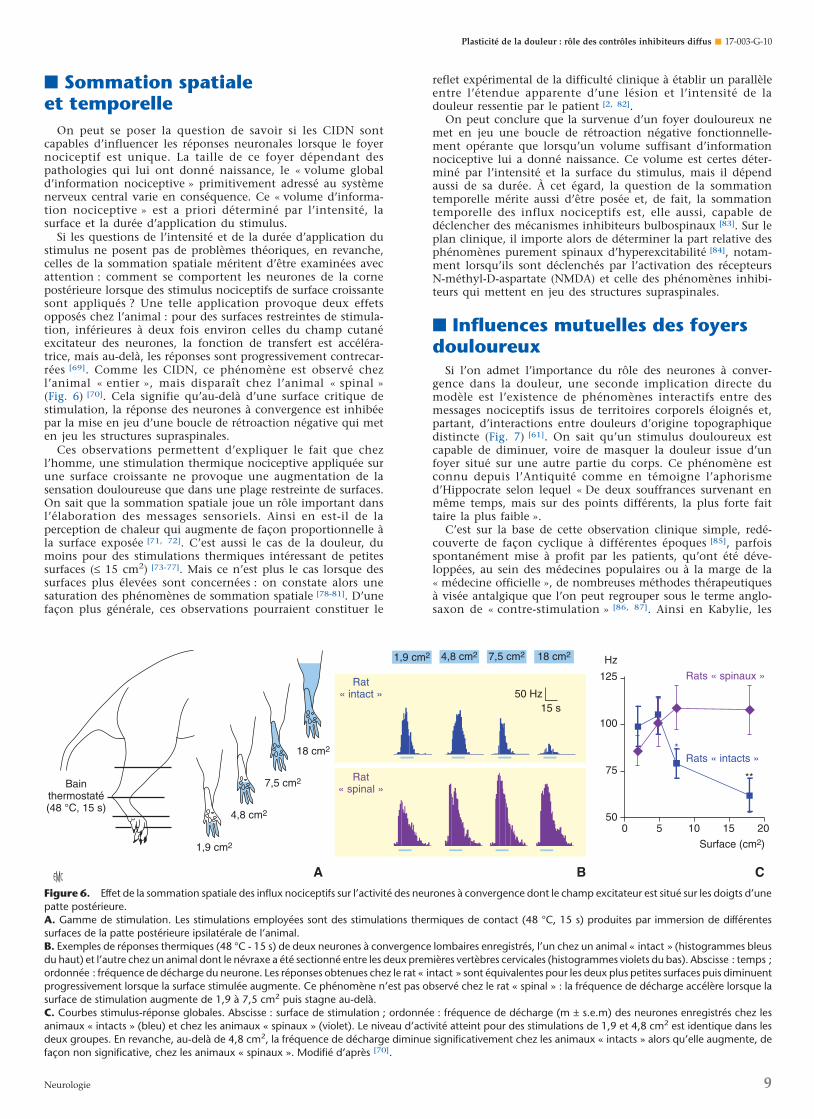
Figure 6. Effet de la sommation spatiale des influx nociceptifs sur l’activité des neurones à convergence dont le champ excitateur est situé sur les doigts d’unepatte postérieure.A. Gamme de stimulation. Les stimulations employées sont des stimulations thermiques de contact (48 °C, 15 s) produites par immersion de différentessurfaces de la patte postérieure ipsilatérale de l’animal.B. Exemples de réponses thermiques (48 °C - 15 s) de deux neurones à convergence lombaires enregistrés, l’un chez un animal « intact » (histogrammes bleusdu haut) et l’autre chez un animal dont le névraxe a été sectionné entre les deux premières vertèbres cervicales (histogrammes violets du bas). Abscisse : temps ;ordonnée : fréquence de décharge du neurone. Les réponses obtenues chez le rat « intact » sont équivalentes pour les deux plus petites surfaces puis diminuentprogressivement lorsque la surface stimulée augmente. Ce phénomène n’est pas observé chez le rat « spinal » : la fréquence de décharge accélère lorsque lasurface de stimulation augmente de 1,9 à 7,5 cm2 puis stagne au-delà.C. Courbes stimulus-réponse globales. Abscisse : surface de stimulation ; ordonnée : fréquence de décharge (m ± s.e.m) des neurones enregistrés chez lesanimaux « intacts » (bleu) et chez les animaux « spinaux » (violet). Le niveau d’activité atteint pour des stimulations de 1,9 et 4,8 cm2 est identique dans lesdeux groupes. En revanche, au-delà de 4,8 cm2, la fréquence de décharge diminue significativement chez les animaux « intacts » alors qu’elle augmente, defaçon non significative, chez les animaux « spinaux ». Modifié d’après [70].
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus ¶ 17-003-G-10
9Neurologie
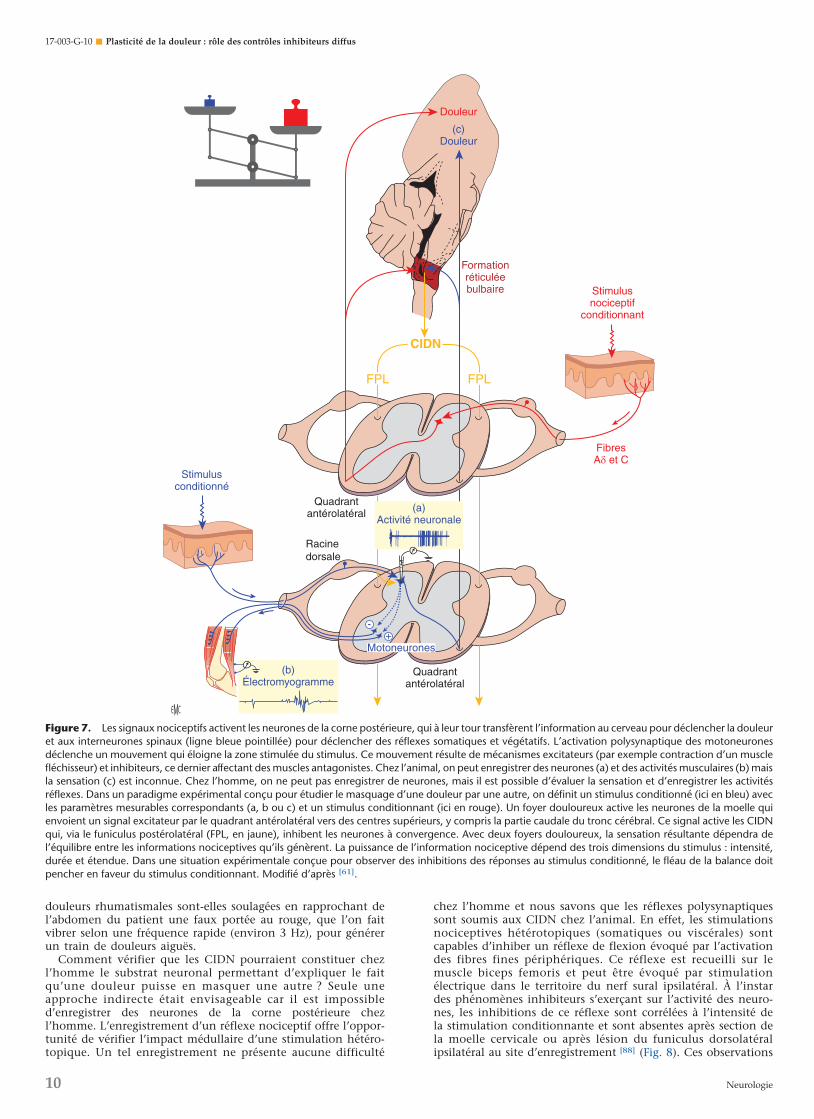
douleurs rhumatismales sont-elles soulagées en rapprochant del’abdomen du patient une faux portée au rouge, que l’on faitvibrer selon une fréquence rapide (environ 3 Hz), pour générerun train de douleurs aiguës.
Comment vérifier que les CIDN pourraient constituer chezl’homme le substrat neuronal permettant d’expliquer le faitqu’une douleur puisse en masquer une autre ? Seule uneapproche indirecte était envisageable car il est impossibled’enregistrer des neurones de la corne postérieure chezl’homme. L’enregistrement d’un réflexe nociceptif offre l’oppor-tunité de vérifier l’impact médullaire d’une stimulation hétéro-topique. Un tel enregistrement ne présente aucune difficulté
chez l’homme et nous savons que les réflexes polysynaptiquessont soumis aux CIDN chez l’animal. En effet, les stimulationsnociceptives hétérotopiques (somatiques ou viscérales) sontcapables d’inhiber un réflexe de flexion évoqué par l’activationdes fibres fines périphériques. Ce réflexe est recueilli sur lemuscle biceps femoris et peut être évoqué par stimulationélectrique dans le territoire du nerf sural ipsilatéral. À l’instardes phénomènes inhibiteurs s’exerçant sur l’activité des neuro-nes, les inhibitions de ce réflexe sont corrélées à l’intensité dela stimulation conditionnante et sont absentes après section dela moelle cervicale ou après lésion du funiculus dorsolatéralipsilatéral au site d’enregistrement [88] (Fig. 8). Ces observations
Formationréticuléebulbaire
CIDN
FPL FPL
Motoneurones
-+
(b)Électromyogramme
Quadrantantérolatéral
Quadrantantérolatéral (a)
Activité neuronale
Stimulusconditionné
Stimulusnociceptif
conditionnant
FibresAδ et C
Douleur
(c)Douleur
Racinedorsale
Figure 7. Les signaux nociceptifs activent les neurones de la corne postérieure, qui à leur tour transfèrent l’information au cerveau pour déclencher la douleuret aux interneurones spinaux (ligne bleue pointillée) pour déclencher des réflexes somatiques et végétatifs. L’activation polysynaptique des motoneuronesdéclenche un mouvement qui éloigne la zone stimulée du stimulus. Ce mouvement résulte de mécanismes excitateurs (par exemple contraction d’un musclefléchisseur) et inhibiteurs, ce dernier affectant des muscles antagonistes. Chez l’animal, on peut enregistrer des neurones (a) et des activités musculaires (b) maisla sensation (c) est inconnue. Chez l’homme, on ne peut pas enregistrer de neurones, mais il est possible d’évaluer la sensation et d’enregistrer les activitésréflexes. Dans un paradigme expérimental conçu pour étudier le masquage d’une douleur par une autre, on définit un stimulus conditionné (ici en bleu) avecles paramètres mesurables correspondants (a, b ou c) et un stimulus conditionnant (ici en rouge). Un foyer douloureux active les neurones de la moelle quienvoient un signal excitateur par le quadrant antérolatéral vers des centres supérieurs, y compris la partie caudale du tronc cérébral. Ce signal active les CIDNqui, via le funiculus postérolatéral (FPL, en jaune), inhibent les neurones à convergence. Avec deux foyers douloureux, la sensation résultante dépendra del’équilibre entre les informations nociceptives qu’ils génèrent. La puissance de l’information nociceptive dépend des trois dimensions du stimulus : intensité,durée et étendue. Dans une situation expérimentale conçue pour observer des inhibitions des réponses au stimulus conditionné, le fléau de la balance doitpencher en faveur du stimulus conditionnant. Modifié d’après [61].
17-003-G-10 ¶ Plasticité de la douleur : rôle des contrôles inhibiteurs diffus
10 Neurologie

confirment plusieurs études effectuées chez l’animal montrantl’inhibition de réflexes nociceptifs ou l’augmentation de seuilsnociceptifs par une stimulation nociceptive hétérotopique [86].Elles assurent le lien avec les études effectuées chezl’homme.
■ Un modèle chez l’hommeLe premier point à vérifier était l’impact médullaire des
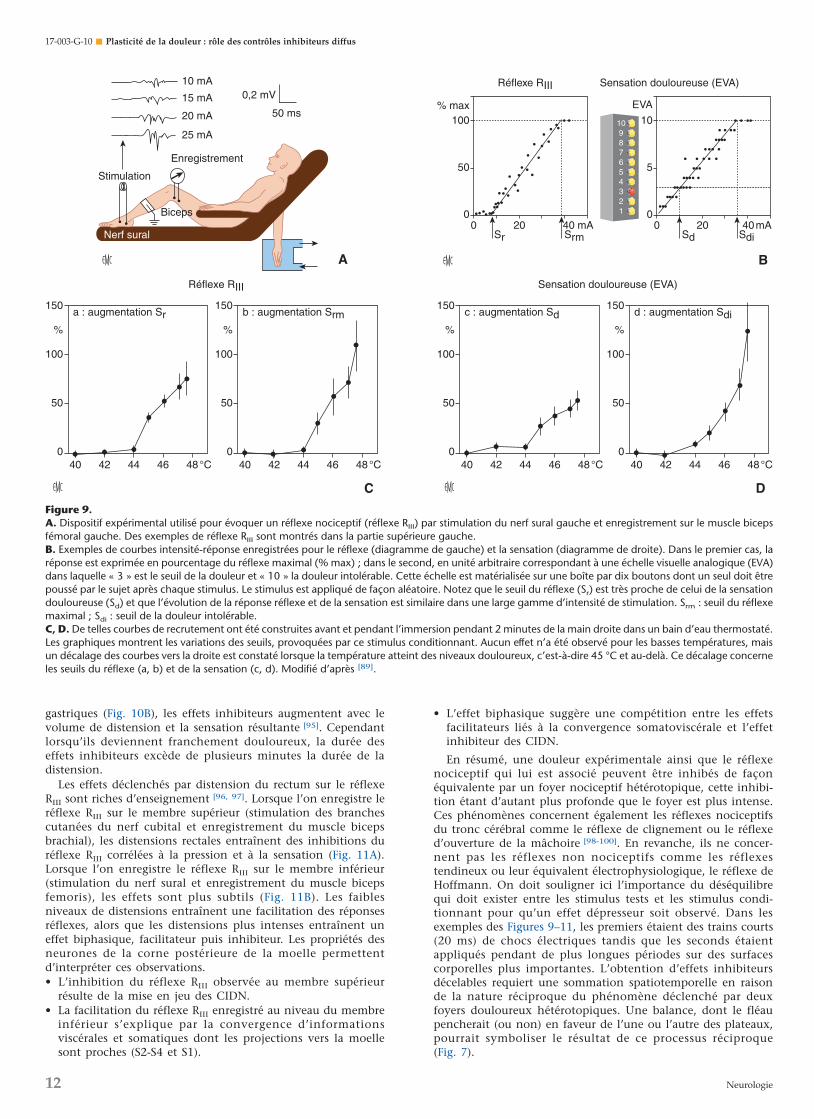
phénomènes de « contre-stimulation ». Dans ce but, le nerfsural de volontaires sains a été stimulé, et un réflexe nociceptifmédullaire (réflexe RIII) enregistré sur le biceps femoris ainsi quela sensation correspondante ont été étudiés simultanémentselon un protocole illustré par la Figure 9. Ce réflexe a été choisicar le seuil de son obtention est quasiment identique au seuilde la sensation douloureuse concomitamment évoquée [90].
Le nerf sural était stimulé régulièrement et la réponseélectromyographique était enregistrée sur le biceps femorisipsilatéral (Fig. 9A). La qualité et l’intensité de la sensationdéclenchées par le stimulus étaient estimées par les sujets surune échelle visuelle analogique à dix niveaux constitués de dixboutons, eux-mêmes reliés à des potentiomètres, le seuildouloureux étant défini par le niveau 3 et le seuil de toléranceà la douleur par le niveau 10. Les intensités de stimulationétaient aléatoires, le réflexe et la sensation convenablementnumérisés étaient exprimés sur un graphe en fonction del’intensité de stimulation. Comme on peut le voir sur la Figure9B, le réflexe et la sensation sont, dans une plage bien précise,une fonction linéaire de l’intensité du courant délivré. C’estainsi que l’on peut définir aisément les seuils du réflexe (Sr) etde la sensation (Sd) ainsi que le seuil du réflexe maximal (Srm)
et le seuil de la douleur intolérable (Sdi). L’évolution de laréponse réflexe est parallèle à celle de la sensation, les seuils duréflexe étant très proches de ceux de la sensation douloureuse.
Ce type de protocole a été réitéré avant et pendant l’applica-tion de stimulus hétérotopiques comme l’immersion de la maincontrolatérale dans de l’eau (Fig. 9C, D) [89]. Lorsque la tempé-rature du bain était inférieure à 45 °C, l’immersion de la mainn’a pas provoqué de variation des courbes intensité-réflexe ouintensité-sensation. En revanche, à partir de 45 °C et enfonction directe de la température (45-47,5 °C), ces courbes ontété décalées vers la droite. Le même type de résultats a étéobtenu lors de l’immersion de la main dans de l’eau à 6 °C(froid douloureux), lors de douleur musculaire provoquée parun exercice de l’avant-bras sous ischémie et par une pressionnociceptive appliquée sur le septum nasal. Si l’on considère cesrésultats dans leur ensemble, on note l’existence d’une très fortecorrélation entre les effets inhibiteurs concernant le réflexe etceux qui concernent la sensation.
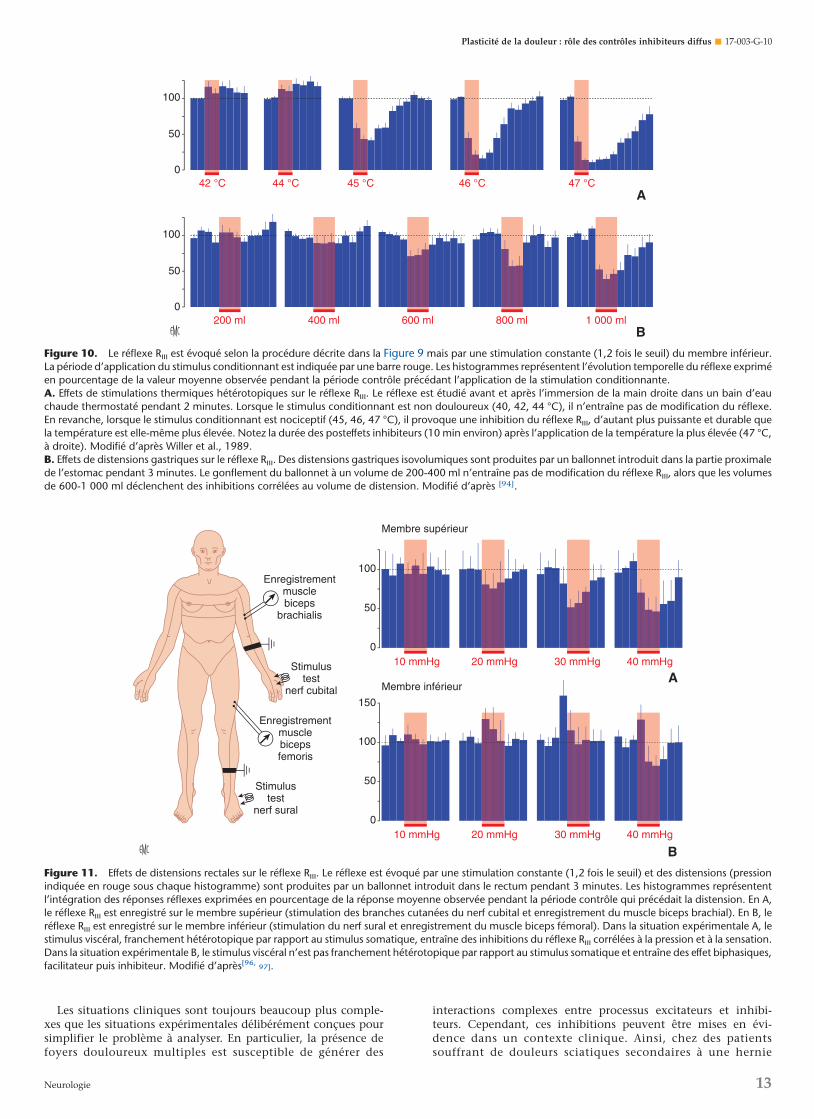
Une autre façon d’étudier ces phénomènes est de stimuler lenerf sural à courant constant et de suivre l’évolution temporelledu réflexe RIII. Les mêmes phénomènes d’inhibition ont ainsipu être mis en évidence [91-94] : ils perdurent bien au-delà de lapériode de stimulation conditionnante (Fig. 10A). Les stimulusconditionnant efficaces n’ont en revanche aucun effet sur unréflexe monosynaptique (réflexe de Hoffmann), qui exclut uneaction directe sur la membrane des motoneurones.
Les stimulations viscérales, gastriques ou rectales sontégalement capables de provoquer des inhibitions du réflexe RIII
chez le volontaire sain. Ces inhibitions sont aussi corrélées àl’intensité du stimulus conditionnant, mais sont d’ores et déjàdéclenchées par des stimulus qui certes sont peu agréables, maisne sont pourtant pas encore douloureux. Lors de distensions
Museau
Patteantérieureipsilatérale
Patteantérieure
controlatérale
« Intacts » « Spinaux »
Pattepostérieure
controlatérale
Queue
1 mV
100 ms
100
0
100
0
100
0
100
0
100
0
100
0
100
0
100
0
100
0
100
0
Figure 8. Effets de stimulations nociceptives hétérotopiques sur le réflexe aux fibres C chez un rat dont le névraxe est intact (graphes de gauche) ou dontla moelle a été sectionnée 2 jours avant au niveau cervical (graphes de droite). L’activité électromyographique (cartouche) évoquée par stimulation électriquedu territoire du nerf sural est enregistrée dans le muscle biceps femoris. La stimulation est appliquée toutes les 6 s. Les stimulations nociceptives hétérotopiquesinhibent le réflexe chez l’animal « intact » mais pas chez l’animal « spinal ». Chez ce dernier cependant, une inhibition faible mais significative, témoin del’intervention de mécanismes propriospinaux, est évoquée en stimulant le membre controlatéral. Modifié d’après [88].
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus ¶ 17-003-G-10
11Neurologie
gastriques (Fig. 10B), les effets inhibiteurs augmentent avec levolume de distension et la sensation résultante [95]. Cependantlorsqu’ils deviennent franchement douloureux, la durée deseffets inhibiteurs excède de plusieurs minutes la durée de ladistension.
Les effets déclenchés par distension du rectum sur le réflexeRIII sont riches d’enseignement [96, 97]. Lorsque l’on enregistre leréflexe RIII sur le membre supérieur (stimulation des branchescutanées du nerf cubital et enregistrement du muscle bicepsbrachial), les distensions rectales entraînent des inhibitions duréflexe RIII corrélées à la pression et à la sensation (Fig. 11A).Lorsque l’on enregistre le réflexe RIII sur le membre inférieur(stimulation du nerf sural et enregistrement du muscle bicepsfemoris), les effets sont plus subtils (Fig. 11B). Les faiblesniveaux de distensions entraînent une facilitation des réponsesréflexes, alors que les distensions plus intenses entraînent uneffet biphasique, facilitateur puis inhibiteur. Les propriétés desneurones de la corne postérieure de la moelle permettentd’interpréter ces observations.• L’inhibition du réflexe RIII observée au membre supérieur
résulte de la mise en jeu des CIDN.• La facilitation du réflexe RIII enregistré au niveau du membre
inférieur s’explique par la convergence d’informationsviscérales et somatiques dont les projections vers la moellesont proches (S2-S4 et S1).
• L’effet biphasique suggère une compétition entre les effetsfacilitateurs liés à la convergence somatoviscérale et l’effetinhibiteur des CIDN.
En résumé, une douleur expérimentale ainsi que le réflexenociceptif qui lui est associé peuvent être inhibés de façonéquivalente par un foyer nociceptif hétérotopique, cette inhibi-tion étant d’autant plus profonde que le foyer est plus intense.Ces phénomènes concernent également les réflexes nociceptifsdu tronc cérébral comme le réflexe de clignement ou le réflexed’ouverture de la mâchoire [98-100]. En revanche, ils ne concer-nent pas les réflexes non nociceptifs comme les réflexestendineux ou leur équivalent électrophysiologique, le réflexe deHoffmann. On doit souligner ici l’importance du déséquilibrequi doit exister entre les stimulus tests et les stimulus condi-tionnant pour qu’un effet dépresseur soit observé. Dans lesexemples des Figures 9–11, les premiers étaient des trains courts(20 ms) de chocs électriques tandis que les seconds étaientappliqués pendant de plus longues périodes sur des surfacescorporelles plus importantes. L’obtention d’effets inhibiteursdécelables requiert une sommation spatiotemporelle en raisonde la nature réciproque du phénomène déclenché par deuxfoyers douloureux hétérotopiques. Une balance, dont le fléaupencherait (ou non) en faveur de l’une ou l’autre des plateaux,pourrait symboliser le résultat de ce processus réciproque(Fig. 7).
Enregistrement
Nerf sural
Stimulation
0,2 mV
50 ms
A
10 mA
15 mA
20 mA
25 mA
Biceps
Réflexe RIII
% max100
50
00 20 40 mA mA
Sr Srm
Sensation douloureuse (EVA)
EVA
10
5
00 20 40
Sd Sdi
10987654321
B
150a : augmentation Sr
%
100
50
040 42 44 46 48°C
C
Réflexe RIII
150b : augmentation Srm
%
100
50
040 42 44 46 48°C
150c : augmentation Sd
%
100
50
040 42 44 46 48°C
D
Sensation douloureuse (EVA)
150d : augmentation Sdi
%
100
50
040 42 44 46 48°C
Figure 9.A. Dispositif expérimental utilisé pour évoquer un réflexe nociceptif (réflexe RIII) par stimulation du nerf sural gauche et enregistrement sur le muscle bicepsfémoral gauche. Des exemples de réflexe RIII sont montrés dans la partie supérieure gauche.B. Exemples de courbes intensité-réponse enregistrées pour le réflexe (diagramme de gauche) et la sensation (diagramme de droite). Dans le premier cas, laréponse est exprimée en pourcentage du réflexe maximal (% max) ; dans le second, en unité arbitraire correspondant à une échelle visuelle analogique (EVA)dans laquelle « 3 » est le seuil de la douleur et « 10 » la douleur intolérable. Cette échelle est matérialisée sur une boîte par dix boutons dont un seul doit êtrepoussé par le sujet après chaque stimulus. Le stimulus est appliqué de façon aléatoire. Notez que le seuil du réflexe (Sr) est très proche de celui de la sensationdouloureuse (Sd) et que l’évolution de la réponse réflexe et de la sensation est similaire dans une large gamme d’intensité de stimulation. Srm : seuil du réflexemaximal ; Sdi : seuil de la douleur intolérable.C, D. De telles courbes de recrutement ont été construites avant et pendant l’immersion pendant 2 minutes de la main droite dans un bain d’eau thermostaté.Les graphiques montrent les variations des seuils, provoquées par ce stimulus conditionnant. Aucun effet n’a été observé pour les basses températures, maisun décalage des courbes vers la droite est constaté lorsque la température atteint des niveaux douloureux, c’est-à-dire 45 °C et au-delà. Ce décalage concerneles seuils du réflexe (a, b) et de la sensation (c, d). Modifié d’après [89].
17-003-G-10 ¶ Plasticité de la douleur : rôle des contrôles inhibiteurs diffus
12 Neurologie
Les situations cliniques sont toujours beaucoup plus comple-xes que les situations expérimentales délibérément conçues poursimplifier le problème à analyser. En particulier, la présence defoyers douloureux multiples est susceptible de générer des
interactions complexes entre processus excitateurs et inhibi-teurs. Cependant, ces inhibitions peuvent être mises en évi-dence dans un contexte clinique. Ainsi, chez des patientssouffrant de douleurs sciatiques secondaires à une hernie
Membre supérieur
100
50
0
Enregistrementmusclebiceps
brachialis
Enregistrementmusclebicepsfemoris
Stimulustest
nerf cubital
Stimulustest
nerf sural
Membre inférieur
10 mmHg
100
150
50
010 mmHg 20 mmHg 30 mmHg 40 mmHg
20 mmHg 30 mmHg 40 mmHg
A
B
Figure 11. Effets de distensions rectales sur le réflexe RIII. Le réflexe est évoqué par une stimulation constante (1,2 fois le seuil) et des distensions (pressionindiquée en rouge sous chaque histogramme) sont produites par un ballonnet introduit dans le rectum pendant 3 minutes. Les histogrammes représententl’intégration des réponses réflexes exprimées en pourcentage de la réponse moyenne observée pendant la période contrôle qui précédait la distension. En A,le réflexe RIII est enregistré sur le membre supérieur (stimulation des branches cutanées du nerf cubital et enregistrement du muscle biceps brachial). En B, leréflexe RIII est enregistré sur le membre inférieur (stimulation du nerf sural et enregistrement du muscle biceps fémoral). Dans la situation expérimentale A, lestimulus viscéral, franchement hétérotopique par rapport au stimulus somatique, entraîne des inhibitions du réflexe RIII corrélées à la pression et à la sensation.Dans la situation expérimentale B, le stimulus viscéral n’est pas franchement hétérotopique par rapport au stimulus somatique et entraîne des effet biphasiques,facilitateur puis inhibiteur. Modifié d’après[96,
97].
100
50
0
A
100
50
0
42 °C 44 °C 45 °C 46 °C 47 °C
200 ml 400 ml 600 ml 800 ml 1 000 mlB
Figure 10. Le réflexe RIII est évoqué selon la procédure décrite dans la Figure 9 mais par une stimulation constante (1,2 fois le seuil) du membre inférieur.La période d’application du stimulus conditionnant est indiquée par une barre rouge. Les histogrammes représentent l’évolution temporelle du réflexe expriméen pourcentage de la valeur moyenne observée pendant la période contrôle précédant l’application de la stimulation conditionnante.A. Effets de stimulations thermiques hétérotopiques sur le réflexe RIII. Le réflexe est étudié avant et après l’immersion de la main droite dans un bain d’eauchaude thermostaté pendant 2 minutes. Lorsque le stimulus conditionnant est non douloureux (40, 42, 44 °C), il n’entraîne pas de modification du réflexe.En revanche, lorsque le stimulus conditionnant est nociceptif (45, 46, 47 °C), il provoque une inhibition du réflexe RIII, d’autant plus puissante et durable quela température est elle-même plus élevée. Notez la durée des posteffets inhibiteurs (10 min environ) après l’application de la température la plus élevée (47 °C,à droite). Modifié d’après Willer et al., 1989.B. Effets de distensions gastriques sur le réflexe RIII. Des distensions gastriques isovolumiques sont produites par un ballonnet introduit dans la partie proximalede l’estomac pendant 3 minutes. Le gonflement du ballonnet à un volume de 200-400 ml n’entraîne pas de modification du réflexe RIII, alors que les volumesde 600-1 000 ml déclenchent des inhibitions corrélées au volume de distension. Modifié d’après [94].
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus ¶ 17-003-G-10
13Neurologie

discale, la manœuvre de Lasègue de la jambe affectée, quidéclenche une douleur radiculaire typique, bloque le réflexenociceptif RIII, que ce dernier soit enregistré sur la jambeaffectée ou non affectée [101]. Effectuée sur le membre sain, lamanœuvre de Lasègue est indolore et n’a aucun effet sur cesréflexes (Fig. 12). Chez des patients souffrant de douleursneuropathiques, les inhibitions du réflexe nociceptif RIII sontobservées lorsque la douleur est déclenchée par une pression(« allodynie statique »), mais pas lorsqu’elle est déclenchée parun effleurement (« allodynie dynamique ») [102]. Rappelons icique les auteurs s’accordent à attribuer la première à l’activationdu système nociceptif, notamment au travers des fibres Ad et C,et la seconde à l’activation de fibres Ab.
■ Neurologie des « contrôlesinhibiteurs diffus nociceptifs »
L’origine médullaire, partielle pour le moins, des phénomènesde contre-stimulation est très probable. Comme il existe unebonne corrélation entre les effets observés sur le réflexe et surla sensation correspondante, on peut penser que les mécanismesinhibiteurs s’expriment in fine sur une population d’interneu-rones, commune aux voies ascendantes de la douleur et auxréflexes nociceptifs. Quelle (s) structure (s) participent audéclenchement de telles inhibitions ?
La participation de structures cérébrales dans ces processus aété démontrée par leur disparition chez des patients tétraplégi-ques présentant une section complète de la moelle d’origine
traumatique [103] (Fig. 13). La lésion médullaire intéressait lesniveaux C5, C6 ou C7 alors que les segments inférieurs,notamment C8 et T1, vers lesquels projettent les branchesdigitales du nerf cubital, étaient indemnes. Les effets d’unstimulus nociceptif appliqué sur les 4e et 5e doigts de la mainont été étudiés chez ces patients et chez des sujets sains. Lastimulation nociceptive des 4e et 5e doigts de la main provoque,chez les sujets sains, une inhibition totale du réflexe RIII suiviede posteffets durant 5 minutes environ. Chez les patients, leréflexe RIII ne différait pas notablement de celui enregistré chezles volontaires sains, mais n’a jamais été inhibé par le stimulusnociceptif conditionnant.
Il est donc probable que les effets inhibiteurs déclenchés parstimulation nociceptive hétérotopique chez l’homme sain sontengendrés par une boucle complexe faisant intervenir desstructures cérébrales. Pour aller plus avant dans cette investiga-tion, il était nécessaire d’étudier ces phénomènes chez despatients porteurs de lésions cérébrales bien circonscrites [104]. Lepremier groupe était porteur d’une lésion thalamique bienvisible au scanner dont une des conséquences neurologiquesétait une hémianesthésie controlatérale à la lésion. L’applicationd’un stimulus nociceptif sur les 4e et 5e doigts de la mainanesthésiée a provoqué une profonde inhibition du réflexe Rlll
enregistré sur le membre inférieur, inhibition en tout pointcomparable à celle observée chez les volontaires sains (Fig. 13C).Ces observations excluent clairement la participation dufaisceau spinothalamique aux phénomènes de contre-irritation.Par ailleurs, ces observations démontrent sans ambiguïté que detels phénomènes peuvent être déclenchés en l’absence de
Jambe sciatalgiqueStimulation (nerf sural)
Enregistrement du réflexe RIII(biceps femoris)
50 mV
50 msJambe saine
Figure 12. Inhibition du réflexe RIII par la manœuvre de Lassègue d’un membre sciatalgique. Le réflexe RIII est enregistré du côté sain (schémas du haut) oudu côté atteint (schémas du bas) chez un patient souffrant de sciatalgie résultant d’une protrusion unilatérale du disque intervertébral. Le réflexe évoqué parune stimulation supraliminaire (1,2 fois le seuil) est étudié avant, pendant et après la manœuvre de Lasègue. Tracés du haut, enregistrement du membre sain :la manœuvre de Lasègue du membre sciatalgique bloque le réflexe enregistré du côté sain ; effectuée sur le membre sain, elle n’affecte pas le réflexe enregistrédu même côté. Tracés du bas, enregistrement du membre sciatalgique : la manœuvre de Lasègue du membre sciatalgique bloque le réflexe enregistré dumême côté ; effectuée sur le membre sain, elle n’affecte pas le réflexe du membre sciatalgique. Modifié d’après [101].
17-003-G-10 ¶ Plasticité de la douleur : rôle des contrôles inhibiteurs diffus
14 Neurologie

sensation douloureuse. Cette observation suggère que le phéno-mène observé ne résulte pas de la compétition entre les foyersattentionnels que représentent chez le volontaire sain la douleurconditionnée et la douleur conditionnante. On sait en effet quela perception de la douleur est sensible aux processus attention-nels [105, 106].
Le second groupe de patients présentait le tableau typique dusyndrome de Wallenberg lié à une lésion ischémique unilatéralede la partie rétro-olivaire du bulbe (Fig. 13D). Ce tableaucomportait notamment une anesthésie thermoalgique controla-térale à la lésion, intéressant les membres supérieurs et infé-rieurs. Chez ces patients, l’application d’un stimulus nociceptifconditionnant sur les 4e et 5e doigts de la main anesthésiée n’aprovoqué aucun phénomène inhibiteur sur le réflexe RIIIenregistré sur le membre inférieur. En revanche, le mêmestimulus appliqué sur la main normale a déclenché des inhibi-tions identiques à celles observées chez les volontaires sains. Onpeut conclure sur la base de ces résultats que la partie caudaleet latérale du bulbe est impliquée chez l’homme dans la boucledes CIDN. C’est très vraisemblablement la lésion du faisceauspinoréticulaire qui était responsable des effets observés. Onpeut donc envisager que, comme chez l’animal, le substratanatomique des CIDN chez l’homme est constitué par uneboucle spino-réticulo-spinale.
Les résultats d’une étude complémentaire tendent à conforterl’idée que la boucle sous-tendant les CIDN est organisée defaçon similaire chez l’homme et chez l’animal. Cette étude a étéréalisée chez un patient présentant un syndrome de Brown-Séquard lié à une hémisection traumatique de la moelle dor-sale [107]. Le réflexe RIII a été enregistré sur chacun des membres.Dans chaque cas, des stimulations nociceptives conditionnantesont été successivement appliquées sur les trois autres membres(Fig. 14). On peut résumer les résultats de cette étude de lamanière suivante :• quel que soit le site d’enregistrement, aucune inhibition du
réflexe RIII n’a été obtenue lorsque la stimulation nociceptiveconditionnante a été appliquée sur le membre inférieurcontrolatéral à la lésion ;
• le réflexe RIII enregistré sur le membre inférieur ipsilatéral àla lésion n’a été inhibé par aucune des stimulations nocicep-tives conditionnantes.En d’autres termes, ces résultats suggèrent que la branche
ascendante de la boucle des CIDN est complètement croisée auniveau médullaire et que la partie descendante de cette boucleest confinée dans la moelle ipsilatérale au lieu d’enregistrement.Ces résultats soulignent les analogies de l’organisation des CIDNchez l’animal et chez l’homme.
■ ConclusionNous sommes en présence d’un faisceau d’arguments, issu
d’expériences effectuées chez l’animal et chez l’homme, quisuggère que les phénomènes de « contre-stimulation » sontsous-tendus par un substrat neurologique bien défini mais dontl’exploration n’est pas terminée. Les effets inhibiteurs déclen-chés par stimulation nociceptive hétérotopique chez l’homme etles CIDN primitivement définis chez l’animal partagent l’essen-tiel des mécanismes qui les sous-tendent. Ces mécanismes sontprobablement mis en route dès l’apparition d’un foyer nocicep-tif, ce qui conduit à envisager leur intervention dans le traite-ment de l’information qui engendre la douleur. On peut eneffet considérer les CIDN comme un système particulier et« généralisé » d’inhibition latérale. Il semble raisonnable de leuraccorder une fonction équivalente ou apparentée. Bien qu’ini-tialement découverts au travers de certaines illusions, lesphénomènes d’inhibition latérale jouent un rôle important enphysiologie sensorielle. Il est très probable que les CIDN, révéléspar l’observation empirique du masquage d’une douleur par uneautre douleur, jouent également un rôle important dans lesprocessus douloureux, probablement en focalisant le réseauneuronal sur une partie particulière du corps au détriment desautres. Les CIDN pourraient ainsi constituer le filtre permettantd’extraire un signal signifiant d’une activité somesthésique debase transmise au cerveau par les neurones à convergence. Si telest bien le cas, réduire ce filtre devrait soulager la douleur. C’estce que fait la morphine qui, comme chez l’animal, bloque lesCIDN chez l’homme (Fig. 15) [108]. La singulière capacité desstimulations viscérales à déclencher les CIDN nous semble aussitrès significative à cet égard.
La contribution de « l’activité somesthésique de base » àl’élaboration du « schéma corporel » reste une hypothèse detravail à démontrer. Paul Schilder (1950) avait affirmé que « leproblème de la douleur ne peut être compris que dans sonrapport avec le schéma corporel » et même, plus radicalement,que « deux facteurs, apparemment, jouent un rôle particulierdans la création de l’image du corps ; l’un est la douleur, l’autreest le contrôle moteur des membres ». La connaissance des« nocicepteurs polymodaux » et des « neurones à convergence »lui aurait sans doute permis de développer ces notions. Laprésente contribution n’a comme objectif que de proposer despistes de réflexion dans ce sens. Les nouvelles approches de laneuropsychologie seront sans doute en mesure d’étayer oud’infirmer cette façon d’envisager les rapports du corps à ladouleur. On sait déjà que le schéma corporel est profondémentaffligé par un foyer douloureux [109, 110].
Sain Tétraplégique Thalamique Wallenberg
150
100
50
0
A B C DFigure 13. Le réflexe RIII est évoqué selon la procédure décrite dans la Figure 9 mais par une stimulation constante (1,2 fois le seuil) du membre inférieur.La période d’application du stimulus conditionnant est indiquée par une barre rouge. Les histogrammes représentent l’évolution temporelle du réflexe expriméen pourcentage de la valeur moyenne observée pendant la période contrôle précédant l’application de la stimulation électrique conditionnante sur le membresupérieur.A. Sujets normaux : la stimulation conditionnante entraîne une inhibition importante du réflexe RIII.B. Patients tétraplégiques : absence d’inhibition.C. Patients présentant une lésion thalamique : comme chez les sujets normaux, la stimulation conditionnante entraîne une inhibition importante du réflexe RIII.D. Patients présentant un syndrome de Wallenberg : la stimulation conditionnante inhibe le réflexe RIII lorsqu’elle est appliquée sur la main normale (ipsilatéraleà la lésion, représentée à gauche) mais ne l’inhibe pas lorsqu’elle est appliquée sur la main analgésique (controlatérale à la lésion, représentée à droite). Modifiéd’après [103, 104].
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus ¶ 17-003-G-10
15Neurologie
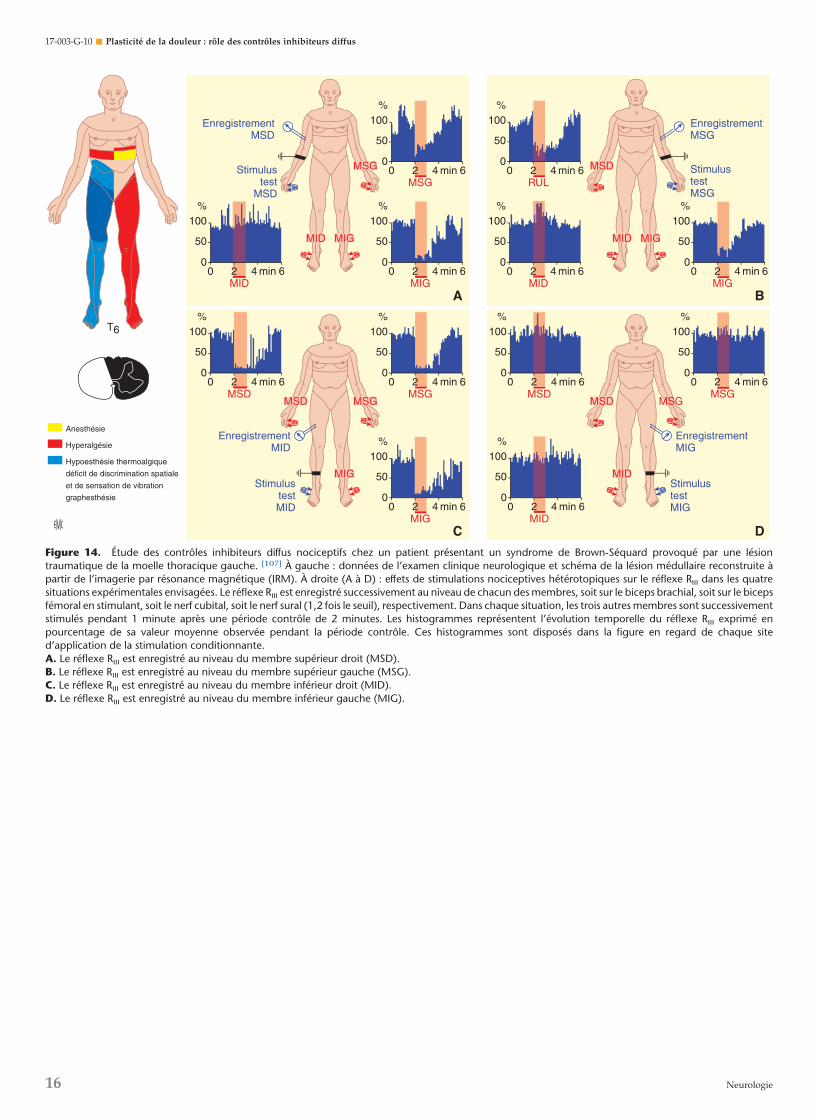
EnregistrementMSD
Stimulustest
MSD
Anesthésie
T6
A B
Hyperalgésie
Hypoesthésie thermoalgique
déficit de discrimination spatiale
et de sensation de vibration
graphesthésie
100%
50
00 2 4 6min
MID
100%
50
00 2 4 6min
MSG
100%
50
00 2 4 6min
RUL
100%
50
00 2 4 6min
MID
100%
50
00 2 4 6min
MIG
100%
50
00 2 4 6min
MIG
MID MIG
MSG
EnregistrementMSG
StimulustestMSG
MID MIG
MSD
EnregistrementMID
StimulustestMID
C D
100%
50
00 2 4 6min
MSD
100%
50
00 2 4 6min
MSG
100%
50
00 2 4 6min
MSD
100%
50
00 2 4 6min
MID
100%
50
00 2 4 6min
MSG
100%
50
00 2 4 6min
MIG
MIG
MSGMSD
EnregistrementMIG
StimulustestMIG
MID
MSD MSG
Figure 14. Étude des contrôles inhibiteurs diffus nociceptifs chez un patient présentant un syndrome de Brown-Séquard provoqué par une lésiontraumatique de la moelle thoracique gauche. [107] À gauche : données de l’examen clinique neurologique et schéma de la lésion médullaire reconstruite àpartir de l’imagerie par résonance magnétique (IRM). À droite (A à D) : effets de stimulations nociceptives hétérotopiques sur le réflexe RIII dans les quatresituations expérimentales envisagées. Le réflexe RIII est enregistré successivement au niveau de chacun des membres, soit sur le biceps brachial, soit sur le bicepsfémoral en stimulant, soit le nerf cubital, soit le nerf sural (1,2 fois le seuil), respectivement. Dans chaque situation, les trois autres membres sont successivementstimulés pendant 1 minute après une période contrôle de 2 minutes. Les histogrammes représentent l’évolution temporelle du réflexe RIII exprimé enpourcentage de sa valeur moyenne observée pendant la période contrôle. Ces histogrammes sont disposés dans la figure en regard de chaque sited’application de la stimulation conditionnante.A. Le réflexe RIII est enregistré au niveau du membre supérieur droit (MSD).B. Le réflexe RIII est enregistré au niveau du membre supérieur gauche (MSG).C. Le réflexe RIII est enregistré au niveau du membre inférieur droit (MID).D. Le réflexe RIII est enregistré au niveau du membre inférieur gauche (MIG).
17-003-G-10 ¶ Plasticité de la douleur : rôle des contrôles inhibiteurs diffus
16 Neurologie
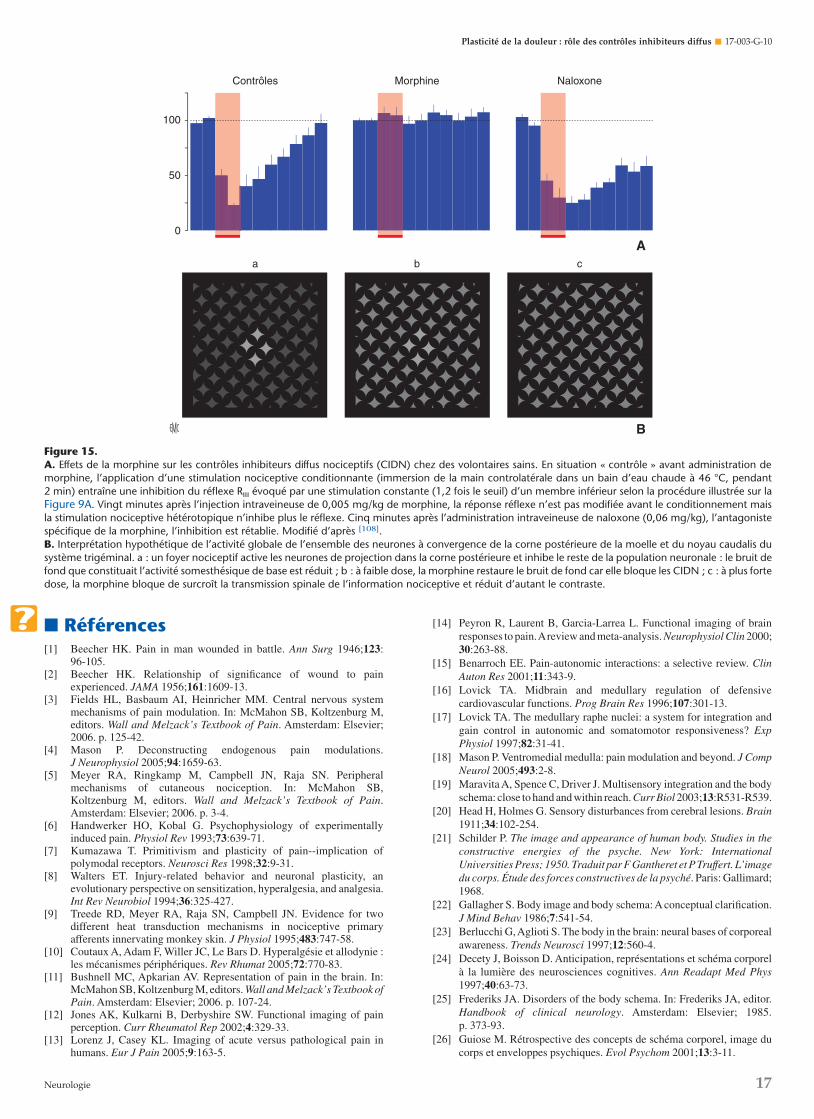
■ Références[1] Beecher HK. Pain in man wounded in battle. Ann Surg 1946;123:
96-105.[2] Beecher HK. Relationship of significance of wound to pain
experienced. JAMA 1956;161:1609-13.[3] Fields HL, Basbaum AI, Heinricher MM. Central nervous system
mechanisms of pain modulation. In: McMahon SB, Koltzenburg M,editors. Wall and Melzack’s Textbook of Pain. Amsterdam: Elsevier;2006. p. 125-42.
[4] Mason P. Deconstructing endogenous pain modulations.J Neurophysiol 2005;94:1659-63.
[5] Meyer RA, Ringkamp M, Campbell JN, Raja SN. Peripheralmechanisms of cutaneous nociception. In: McMahon SB,Koltzenburg M, editors. Wall and Melzack’s Textbook of Pain.Amsterdam: Elsevier; 2006. p. 3-4.
[6] Handwerker HO, Kobal G. Psychophysiology of experimentallyinduced pain. Physiol Rev 1993;73:639-71.
[7] Kumazawa T. Primitivism and plasticity of pain--implication ofpolymodal receptors. Neurosci Res 1998;32:9-31.
[8] Walters ET. Injury-related behavior and neuronal plasticity, anevolutionary perspective on sensitization, hyperalgesia, and analgesia.Int Rev Neurobiol 1994;36:325-427.
[9] Treede RD, Meyer RA, Raja SN, Campbell JN. Evidence for twodifferent heat transduction mechanisms in nociceptive primaryafferents innervating monkey skin. J Physiol 1995;483:747-58.
[10] Coutaux A, Adam F, Willer JC, Le Bars D. Hyperalgésie et allodynie :les mécanismes périphériques. Rev Rhumat 2005;72:770-83.
[11] Bushnell MC, Apkarian AV. Representation of pain in the brain. In:McMahon SB, Koltzenburg M, editors. Wall and Melzack’s Textbook ofPain. Amsterdam: Elsevier; 2006. p. 107-24.
[12] Jones AK, Kulkarni B, Derbyshire SW. Functional imaging of painperception. Curr Rheumatol Rep 2002;4:329-33.
[13] Lorenz J, Casey KL. Imaging of acute versus pathological pain inhumans. Eur J Pain 2005;9:163-5.
[14] Peyron R, Laurent B, Garcia-Larrea L. Functional imaging of brainresponses to pain.Areview and meta-analysis. Neurophysiol Clin 2000;30:263-88.
[15] Benarroch EE. Pain-autonomic interactions: a selective review. ClinAuton Res 2001;11:343-9.
[16] Lovick TA. Midbrain and medullary regulation of defensivecardiovascular functions. Prog Brain Res 1996;107:301-13.
[17] Lovick TA. The medullary raphe nuclei: a system for integration andgain control in autonomic and somatomotor responsiveness? ExpPhysiol 1997;82:31-41.
[18] Mason P. Ventromedial medulla: pain modulation and beyond. J CompNeurol 2005;493:2-8.
[19] Maravita A, Spence C, Driver J. Multisensory integration and the bodyschema: close to hand and within reach. Curr Biol 2003;13:R531-R539.
[20] Head H, Holmes G. Sensory disturbances from cerebral lesions. Brain1911;34:102-254.
[21] Schilder P. The image and appearance of human body. Studies in theconstructive energies of the psyche. New York: InternationalUniversities Press; 1950. Traduit par F Gantheret et PTruffert. L’imagedu corps. Étude des forces constructives de la psyché. Paris: Gallimard;1968.
[22] Gallagher S. Body image and body schema: A conceptual clarification.J Mind Behav 1986;7:541-54.
[23] Berlucchi G, Aglioti S. The body in the brain: neural bases of corporealawareness. Trends Neurosci 1997;12:560-4.
[24] Decety J, Boisson D. Anticipation, représentations et schéma corporelà la lumière des neurosciences cognitives. Ann Readapt Med Phys1997;40:63-73.
[25] Frederiks JA. Disorders of the body schema. In: Frederiks JA, editor.Handbook of clinical neurology. Amsterdam: Elsevier; 1985.p. 373-93.
[26] Guiose M. Rétrospective des concepts de schéma corporel, image ducorps et enveloppes psychiques. Evol Psychom 2001;13:3-11.
Contrôles
100
50
0
Morphine Naloxone
A
B
a b c
Figure 15.A. Effets de la morphine sur les contrôles inhibiteurs diffus nociceptifs (CIDN) chez des volontaires sains. En situation « contrôle » avant administration demorphine, l’application d’une stimulation nociceptive conditionnante (immersion de la main controlatérale dans un bain d’eau chaude à 46 °C, pendant2 min) entraîne une inhibition du réflexe RIII évoqué par une stimulation constante (1,2 fois le seuil) d’un membre inférieur selon la procédure illustrée sur laFigure 9A. Vingt minutes après l’injection intraveineuse de 0,005 mg/kg de morphine, la réponse réflexe n’est pas modifiée avant le conditionnement maisla stimulation nociceptive hétérotopique n’inhibe plus le réflexe. Cinq minutes après l’administration intraveineuse de naloxone (0,06 mg/kg), l’antagonistespécifique de la morphine, l’inhibition est rétablie. Modifié d’après [108].B. Interprétation hypothétique de l’activité globale de l’ensemble des neurones à convergence de la corne postérieure de la moelle et du noyau caudalis dusystème trigéminal. a : un foyer nociceptif active les neurones de projection dans la corne postérieure et inhibe le reste de la population neuronale : le bruit defond que constituait l’activité somesthésique de base est réduit ; b : à faible dose, la morphine restaure le bruit de fond car elle bloque les CIDN ; c : à plus fortedose, la morphine bloque de surcroît la transmission spinale de l’information nociceptive et réduit d’autant le contraste.
.
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus ¶ 17-003-G-10
17Neurologie

[27] Lautenbacher S, Roscher S, Strian F, Pirke KM, Krieg JC. Theoreticaland empirical considerations on the relation between ’body image’,body scheme and somatosensation. J Psychosom Res 1993;37:447-54.
[28] Morin C, Thibierge S. L’image du corps en neurologie : de lacénesthésie à l’image spéculaire. Apports cliniques et théoriques de lapsychanalyse. Evol Psychiatr (Paris) 2004;69:417-30.
[29] Paillard J. Le corps situé et le corps identifié. Une approchepsychophysiologique de la notion de schéma corporel. Rev Med SuisseRomande 1980;100:129-41.
[30] Roll JP, Roll R. Le sentiment d’incarnation : argumentsneurobiologiques. Rev Méd Psychosomat 1993;33:75-90.
[31] Shontz FC. Body image and its disorders. Int J Psychiatry Med 1974;4:461-72.
[32] Hécaen H, de Ajuriaguerra J. Asymbolie à la douleur. Étude anatomo-clinique. Ses rapports avec l’hémiagnosie douloureuse. Sem Hop Paris1959;26:4660-4.
[33] Ramachandran VS. Consciousness and body image: lessons fromphantom limbs, Capgras syndrome and pain asymbolia. Philos Trans RSoc Lond B Biol Sci 1998;353:1851-9.
[34] Lhermitte F, Cambier J. Les perturbations somatognosiques en patho-logie nerveuse. Paris: Masson; 1960.
[35] Gandevia SC, Phegan CM. Perceptual distortions of the human bodyimage produced by local anaesthesia, pain and cutaneous stimulation.J Physiol 1999;514:609-16.
[36] Paqueron X, Leguen M, Rosenthal D, Coriat P, Willer JC, Danziger N.The phenomenology of body image distortions induced by regionalanaesthesia. Brain 2003;126:702-12.
[37] Cambier J, Bovier P. Corps et douleur. In: Jeddi E, editor. Le corps enpsychiatrie. Paris: Masson; 1982. p. 78-86.
[38] Defontaine-Catteau MC. Corps et organe douloureux dans le dessin dela douleur. Doul Analg 1990;3:83-7.
[39] Le Bars D.The whole body receptive field of dorsal horn multireceptiveneurones. Brain Res Rev 2002;40:29-44.
[40] Willis WD, Coggeshall RE. Sensory mechanisms of the spinal cord.New York: Plenum Press; 1991.
[41] Le Bars D, Cadden SW. What is a wide-dynamic-range cell? In:Basbaum AI, Hoy RR, Kaneko A, Shepherd G, Westheimer G, editors.The senses. A comprehensive reference. Amsterdam: Elsevier; 2007(sous presse).
[42] Hillman P, Wall PD. Inhibitory and excitatory factors influencing thereceptive fields of lamina 5 spinal cord cells. Exp Brain Res 1969;9:284-306.
[43] Le Bars D, Chitour D. Do convergent neurons in the spinal corddiscriminate nociceptive from non-nociceptive information? Pain1983;17:1-9.
[44] Price DD, Hayes RL, Ruda MA, Dubner R. Spatial and temporal trans-formations of input to spinothalamic tract neurons and their relation tosomatic sensations. J Neurophysiol 1978;41:933-47.
[45] Arendt-Nielsen L, Svensson P. Referred muscle pain: basic and clinicalfindings. Clin J Pain 2001;17:11-9.
[46] Cervero F, Laird JM. Visceral pain. Lancet 1999;353:2145-8.[47] Foreman RD. Integration of viscerosomatic sensory input at the spinal
level. Prog Brain Res 2000;122:209-21.[48] Joshi SK, Gebhart GF. Visceral pain. Curr Rev Pain 2000;4:499-506.[49] Pubols LM. Electrical stimulation reveals relatively ineffective sural
nerve projections to dorsal horn neurons in the cat. Brain Res 1986;371:109-22.
[50] Wall PD. The presence of ineffective synapses and the circumstanceswhich unmask them. Philos Trans R Soc London Ser B 1977;278:361-72.
[51] Woolf CJ, King AE. Subthreshold components of the cutaneousmechanoreceptive fields of dorsal horn neurons in the rat lumbar spinalcord. J Neurophysiol 1989;62:907-16.
[52] Mendell LM. Modifiability of spinal synapses. Physiol Rev 1984;64:260-324.
[53] Yokota T, Nishikawa Y. Actions of picrotoxin upon trigeminalsubnucleus caudalis neurons in the monkey. Brain Res 1979;171:369-73.
[54] Zieglgänsberger W, Herz A. Changes of cutaneous receptive fields ofspino-cervical tract neurones and other dorsal horn neurones bymicroelectrophoretically administered aminoacids. Exp Brain Res1971;13:111-26.
[55] Schaible HG. Spinal mechanisms contributing to joint pain. NovartisFound Symp 2004;260:4-22.
[56] Herrero JF, Laird JM, Lopez-Garcia JA. Wind-up of spinal cordneurones and pain sensation: much ado about something? ProgNeurobiol 2000;61:169-203.
[57] Ji RR, Woolf CJ. Neuronal plasticity and signal transduction innociceptive neurons: implications for the initiation and maintenance ofpathological pain. Neurobiol Dis 2001;8:1-0.
[58] Mendell LM. Physiological properties of unmyelinated fiber projectionto the spinal cord. Exp Neurol 1966;16:316-32.
[59] Schouenborg J. Modular organisation and spinal somatosensoryimprinting. Brain Res Rev 2002;40:80-91.
[60] Villanueva L, Cadden SW, Le Bars D. Evidence that diffuse noxiousinhibitory controls (DNIC) are mediated by a final post-synapticinhibitory mechanism. Brain Res 1984;298:67-74.
[61] Le Bars D, Willer JC. Diffuse Noxious Inhibitory Controls (DNIC). In:Basbaum AI, Hoy RR, Kaneko A, Shepherd G, Westheimer G, editors.The senses. A comprehensive reference. Amsterdam: Elsevier; 2007(sous presse).
[62] Cadden SW. The ability of inhibitory controls to ’switch-off’ activity indorsal horn convergent neurones in the rat. Brain Res 1993;628:65-71.
[63] Cadden SW, Morrison JF. Effects of visceral distension on the activitiesof neurones receiving cutaneous inputs in the rat lumbar dorsal horn:comparison with effects of remote noxious somatic stimuli. Brain Res1991;558:63-74.
[64] Hobbs SF, Oh UT, Brennan TJ, Chandler MJ, Kim KS, Foreman RD.Urinary bladder and hindlimb stimuli inhibit T1-T6 spinal andspinoreticular cells. Am J Physiol 1990;258:R10-R20.
[65] Danziger N, Le Bars D, Bouhassira D. Diffuse noxious inhibitorycontrols and arthritis in the rat. In: Saadé NR, Apkarian AV, Jabbur SJ,editors. Pain and neuroimmune interactions. Amsterdam: KluwerAcademic Press; 2000. p. 69-78.
[66] Danziger N, Gautron M, Le Bars D, Bouhassira D.Activation of diffusenoxious inhibitory controls (DNIC) in rats with an experimentalperipheral mononeuropathy. Pain 2001;91:287-96.
[67] Villanueva L, Bouhassira D, Le Bars D. The medullary subnucleusreticularis dorsalis (SRD) as a key link in both the transmission andmodulation of pain signals. Pain 1996;67:231-40.
[68] Le Bars D, Bouhassira D, Villanueva L. Opioids and diffuse noxiousinhibitory controls in the rat. Adv Pain Res Ther 1995;22:517-39.
[69] Gall O, Bouhassira D, Chitour D, Le Bars D. Sommation spatiale etcodage médullaire de l’information nociceptive. Doul Analg 1999;2:161-7.
[70] Bouhassira D, Gall O, Chitour D, Le Bars D. Dorsal horn convergentneurones: negative feed-back triggered by spatial summation ofnociceptive afferents. Pain 1995;62:195-200.
[71] Marks LE, Stevens JC. Spatial summation of warmth: influence ofduration and configuration of the stimulus. Am J Psychol 1973;86:251-67.
[72] Stevens JC, Marks LE, Simonson DC. Regional sensitivity and spatialsummation in the warmth sense. Physiol Behav 1974;13:825-36.
[73] Defrin R, Urca G. Spatial summation of heat pain: a reassessment. Pain1996;66:23-9.
[74] Douglass DK, Carstens E, Watkins LR. Spatial summation of humanthermal pain perception: comparison within and between dermatomes.Pain 1992;50:197-202.
[75] Kojo I, Pertovaara A. The effects of stimulus area and adaptationtemperature on warm and heat pain thresholds in man. Intern J Neurosci1987;32:875-80.
[76] Machet-Pietropaoli H, Chery-Croze S. Spatial summation of thermalpain in human beings. Sens Process 1979;3:183-7.
[77] Price DD, McHaffie JG, Larson MA. Spatial summation of heat-induced pain: influenced of stimulus area and spatial separation ofstimuli on perceived pain sensation intensity and unpleasantness.J Neurophysiol 1989;62:1270-9.
[78] Greene LC, Hardy JD. Spatial summation of pain. J Appl Physiol 1958;13:457-64.
[79] Hardy JD, Woolf HG, Goodel H. Studies on pain. A new method formeasuring pain threshold: observation on the spatial summation ofpain. J Clin Invest 1940;16:649-57.
[80] Hardy JD, Woolf HG, Goodel H. Pain sensations and reactions.Baltimore: William and Wilkins; 1952.
[81] Melzack R, Rose G, McGinty D. Skin sensitivity to thermal stimuli.Exp Neurol 1962;6:300-14.
[82] Wall PD. On the relation of injury to pain. The John J. Bonica lecture.Pain 1979;6:253-64.
[83] Gozariu M, Bragard D, Willer JC, Le Bars D. Temporal summation ofC-fibre afferent inputs: competition between facilitatory and inhibitoryeffects on a C-fibre reflex in the rat. J Neurophysiol 1997;78:3165-79.
17-003-G-10 ¶ Plasticité de la douleur : rôle des contrôles inhibiteurs diffus
18 Neurologie

[84] Woolf CJ, Salter MW. Plasticity and pain: role of the dorsal horn. In:McMahon SB, Koltzenburg M, editors. Wall and Melzack’s Textbook ofPain. Amsterdam: Elsevier; 2006.
[85] Rey R. Histoire de la douleur. Paris: La découverte; 1993.[86] Le Bars D, Willer JC. Pain modulation triggered by high-intensity
stimulation: implication for acupuncture analgesia. In: Sato A, Li P,Campbell J, editors. Acupuncture: is there a physiological basis? Inter-national Congress Serie 1238. Amsterdam: Elsevier; 2002. p. 11-29.
[87] Melzack R. Folk medicine and the sensory modulation of pain. In:Wall PD, Melzack R, editors. Textbook of pain. Edinburgh: ChurchillLivingstone; 1989. p. 897-905.
[88] Falinower S, Willer C, Junien JL, Le Bars D.AC-fibre reflex modulatedby heterotopic somatic stimuli in the rat. J Neurophysiol 1994;72:194-213.
[89] Willer JC, RobyA, Le Bars D. Psychophysical and electrophysiologicalapproaches to the pain relieving effect of heterotopic nociceptivestimuli. Brain 1984;107:1095-112.
[90] Willer JC. Comparative study of perceived pain and nociceptive flexionreflex in man. Pain 1977;3:69-80.
[91] Danziger N, Rozenberg S, Bourgeois P, Charpentier G, Willer JC.Depressive effects of segmental and heterotopic application oftranscutaneous electrical nerve stimulation and piezo-electric currenton lower limb nociceptive flexion reflex in human subjects. Arch PhysMed Rehabil 1998;79:191-200.
[92] France CR, Suchowiecki S.Acomparison of diffuse noxious inhibitorycontrols in men and women. Pain 1999;81:77-84.
[93] Sandrini G, Milanov I, Malaguti S, Nigrelli MP, Moglia A, Nappi G.Effects of hypnosis on diffuse noxious inhibitory controls. PhysiolBehav 2000;69:295-300.
[94] Willer JC, De Broucker T, Le Bars D. Encoding of nociceptive thermalstimuli by Diffuse Noxious Inhibitory Controls in humans.J Neurophysiol 1989;62:1028-38.
[95] Bouhassira D, Chollet R, Coffin B, Lemann M, Le Bars D, Willer JC,et al. Inhibition of a somatic nociceptive reflex by gastric distension inhumans. Gastroenterology 1994;107:985-92.
[96] Bouhassira D, Sabate JM, Coffin B, Le Bars D, Willer JC, Jian R.Effects of rectal distensions on nociceptive flexion reflexes in humans.Am J Physiol 1998;275:G410-G417.
[97] Sabate JM, Coffin B, Jian R, Le Bars D, Bouhassira D. Rectal sensitivityassessed by a reflexologic technique: further evidence for two types ofmechanoreceptors. Am J Physiol 2000;279:G692-G699.
[98] Ellrich J, Treede RD. Characterization of blink reflex interneurons byactivation of diffuse noxious inhibitory controls in man. Brain Res1998;803:161-8.
[99] Maillou P, Cadden SW. Effects of remote deep somatic noxious stimulion a jaw reflex in man. Arch Oral Biol 1997;42:323-7.
[100] Pantaleo T, Duranti R, Bellini F. Effects of heterotopic ischemic pain onmuscular pain threshold and blink reflex in humans. Neurosci Lett1988;85:56-60.
[101] Willer JC, Barranquero A, Kahn MF, Sallière D. Pain in sciaticadepresses lower limb nociceptive reflexes to sural nerve stimulation.J Neurol Neurosurg Psychiatry 1987;50:1-5.
[102] Bouhassira D, Danziger N, Guirimand F, Attal N. Comparison of thepain suppressive effects of clinical and experimental painfulconditioning stimuli. Brain 2003;126:1068-78.
[103] Roby-Brami A, Bussel B, Willer JC, Le Bars D. Anelectrophysiological investigation into the pain-relieving effects ofheterotopic nociceptive stimuli: probable involvement of a supraspinalloop. Brain 1987;110:1497-508.
[104] De Broucker T, Cesaro P, Willer JC, Le Bars D. Diffuse NoxiousInhibitory Controls (DNIC) in man: involvement of the spino-reticulartract. Brain 1990;113:1223-34.
[105] Bushnell MC, Duncan GH, Dubner R, Jones RL, Maixner W.Attentional influences on noxious and innocuous cutaneous heatdetection in humans and monkeys. J Neurosci 1985;5:1103-10.
[106] Willer JC, Boureau F, Albe-Fessard D. Supraspinal influences onnociceptive flexion reflex and pain sensation in man. Brain Res 1979;179:61-8.
[107] Bouhassira D, Le Bars D, Bolgert F, Laplane D, Willer JC. Diffusenoxious inhibitory controls (DNIC) in man. A neurophysiologicalinvestigation of a patient with a form of Brown-Séquard syndrome. AnnNeurol 1993;34:536-43.
[108] Le Bars D, Willer JC, De Broucker T. Morphine blocks descending paininhibitory controls in humans. Pain 1992;48:13-20.
[109] Schwoebel J, Coslett HB, Bradt J, Friedman R, Dileo C. Pain and thebody schema: effects of pain severity on mental representations ofmovement. Neurology 2002;59:775-7.
[110] Schwoebel J, Friedman R, Duda N, Coslett HB. Pain and the bodyschema: evidence for peripheral effects on mental representations ofmovement. Brain 2001;124(Pt10):2098-104.
D. Le Bars ([email protected]).Inserm U-713 (« Douleurs et stress »).
J.-C. Willer ([email protected]).Laboratoire de neurophysiologie et Inserm U-731, Faculté de médecine Pitié-Salpêtrière, 91, boulevard de l’Hôpital, 75013 Paris, France.
Toute référence à cet article doit porter la mention : Le Bars D., Willer J.-C. Plasticité de la douleur : rôle des contrôles inhibiteurs diffus. EMC (Elsevier MassonSAS, Paris), Neurologie, 17-003-G-10, 2008.
Disponibles sur www.emc-consulte.com
Arbresdécisionnels
Iconographiessupplémentaires
Vidéos /Animations
Documentslégaux
Informationau patient
Informationssupplémentaires
Auto-évaluations
Plasticité de la douleur : rôle des contrôles inhibiteurs diffus ¶ 17-003-G-10
19Neurologie












