
Late Triassic (Norian–Rhaetian) brackish to fresh water habitats at a fluvial–dominated delta...
36
1 23 Palaeobiodiversity and Palaeoenvironments ISSN 1867-1594 Volume 94 Number 4 Palaeobio Palaeoenv (2014) 94:495-528 DOI 10.1007/s12549-014-0168-6 Late Triassic (Norian-Rhaetian) brackish to freshwater habitats at a fluvial- dominated delta plain (Seinstedt, Lower Saxony, Germany) Gregor Barth, Matthias Franz, Carmen Heunisch, Evelyn Kustatscher, Detlev Thies, Jürgen Vespermann & Markus Wolfgramm
-
Upload
natrmuseum -
Category
Documents
-
view
1 -
download
0
Transcript of Late Triassic (Norian–Rhaetian) brackish to fresh water habitats at a fluvial–dominated delta...

1 23
Palaeobiodiversity andPalaeoenvironments ISSN 1867-1594Volume 94Number 4 Palaeobio Palaeoenv (2014) 94:495-528DOI 10.1007/s12549-014-0168-6
Late Triassic (Norian-Rhaetian) brackishto freshwater habitats at a fluvial-dominated delta plain (Seinstedt, LowerSaxony, Germany)
Gregor Barth, Matthias Franz, CarmenHeunisch, Evelyn Kustatscher, DetlevThies, Jürgen Vespermann & MarkusWolfgramm

1 23
Your article is protected by copyright and all
rights are held exclusively by Senckenberg
Gesellschaft für Naturforschung and Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL PAPER
Late Triassic (Norian-Rhaetian) brackish to freshwaterhabitats at a fluvial-dominated delta plain (Seinstedt, LowerSaxony, Germany)
Gregor Barth & Matthias Franz & Carmen Heunisch &
Evelyn Kustatscher &Detlev Thies & Jürgen Vespermann &
Markus Wolfgramm
Received: 13 December 2013 /Revised: 14 March 2014 /Accepted: 6 May 2014 /Published online: 25 June 2014# Senckenberg Gesellschaft für Naturforschung and Springer-Verlag Berlin Heidelberg 2014
Abstract The historic Late Triassic outcrops at Fuchsbergand Langenberg near Seinstedt (Lower Saxony, Germany)are constrained to the Norian/Rhaetian boundary interval bymeans of conchostracan and palynomorph biostratigraphy. Acomprehensive revision revealed a fluvial-dominated delta
plain that formed in response to the successive transgressionof the ‘Rhaetian Sea’ and received siliciclastics from southernsource areas. At Fuchsberg and Langenberg, the distal lowerdelta plain is exposed and brackish subaqueous delta plainwetlands, mouthbar/distributary channel complexes andinterdistributary bay subenvironments are reconstructed.Delta formation was controlled by bifurcation of distributarychannels and avulsion of delta lobes. A diverse ecosystem isdocumented: a rich invertebrate fauna of limulids (1 taxon),insects (at least 20 taxa of 9 orders), malacostracans andconchostracans (several taxa) and a vertebrate fauna of am-phibians (at least 1 taxon), sharks (9 taxa) and osteichthyanfishes (at least 6 taxa). In particular, fossiliferousinterdistributary bay lithologies detail trophic systems of au-tochthonous subaqueous and parautochthonous riparian hab-itats. Abundant remnants of cycadophytes, ferns, horsetailsand large vertebrates from Fuchsberg and adjacent outcrops ofthe Seinstedt area enable the reconstruction of vegetated up-stream environments at the upper delta plain and floodplain.
Keywords Distributary channel .Mouth bar .
Interdistributary bay . Tempestite .Hirmeriella muensteri .
Riparian habitat
Introduction
Upper Keuper sandstones have been exploited in numerousquarries in SE Lower Saxony (Jüngst 1928, 1929). Amongthem, the nowadays abandoned quarry at the so-called‘Fuchsberg’ close to the village of Seinstedt is most prominentbecause of fossiliferous layers that have been subject to de-tailed investigations since the 1850s. Von Strombeck (1852)was the first to refer to a number of quarries around Seinstedt.Schlönbach (1861, 1862), Pflücker y Rico (1868) and Roemer
G. Barth (*) :M. FranzTU Bergakademie Freiberg, Bernard-von-Cotta-Str. 2,09599 Freiberg, Germanye-mail: [email protected]
M. Franze-mail: [email protected]
C. HeunischLandesamt für Bergbau, Energie und Geologie (LBEG), Stilleweg 2,30655 Hannover, Germanye-mail: [email protected]
E. KustatscherNaturmuseum Südtirol, Bindergasse 1, 39100 Bolzano, Italye-mail: [email protected]
E. KustatscherDepartment für Geo- und Umweltwissenschaften, Paläontologie undGeobiologie, Ludwig-Maximilians-Universität und BayerischeStaatssammlung für Paläontologie und Geologie,Richard-Wagner-Straße 10, 80333 München, Germany
D. ThiesInstitut für Geologie, Leibnitz Universität Hannover, Callinstr. 30,30167 Hannover, Germanye-mail: [email protected]
J. VespermannRoemer- und Pelizaeus-Museum, Am Steine 1-2, 31134 Hildesheim,Germanye-mail: [email protected]
M. WolfgrammGTN Geothermie Neubrandenburg GmbH, Seestraße 7A,17033 Neubrandenburg, Germanye-mail: [email protected]
Palaeobio Palaeoenv (2014) 94:495–528DOI 10.1007/s12549-014-0168-6
Author's personal copy

(1874) described fish remains and ganoid scales from differentbonebeds. Coquina beds composed of bivalves (Modiolaminuta, Gervilleia inflata, Protocardia praecursor) were de-scribed by Schlönbach (1862) from an abandoned sandstonequarry near Seinstedt (most probably not the quarry atFuchsberg described here). A bone fragment of the prosauro-pod Plateosaurus found at Fuchsberg (Fig. 6e, f) wasmentioned by Fraas (1897). Brauns (1862, 1866) was thefirst who described a floral assemblage from a quarry thatmost probably refers to the Fuchsberg quarry and proposed aRhaetian age for the exposed succession. Jüngst (1928) andvon Engelhardt (1942) incorporated the Fuchsberg intosedimentological and petrological studies of more regionalscales. Benda (1960, 1963) noted that one of the bonebedsat Fuchsberg contains higher values of uranium. Based onregional mapping, he provided a number of measured sectionsand interpreted the bonebeds in the classical sense as trans-gressive lag deposits of the ‘Rhaetian’ transgression. Appel(1981) introduced the first depositional model based onlithofacies and reconstructions of transport directions andconsidered the Fuchsberg part of a fluvial floodplain.Hauschke and Wilde (1987) described Paleolimulusfuchsbergensis (Figs. 8c, 12g) as well as limulidtracks (Hauschke and Wilde 1996). From a systematic exca-vation of a dark shaly horizon (upper part of the Main FossilLayer -MFL, ‘FossilführendeWechselfolge’, sensu Hauschkeand Wilde 1987, 1996) (Fig. 2) , Hauschke andWilde (1996) reported insect remains assigned to severalorders (e.g. Blattodea, Coleoptera, Hemiptera) (Fig. 7),articulated skeletons of Semionotus-like fishes, coelacanthidscales and several moderate preserved and not exactly speci-fied imprints of Malacostracans assigned to at least two taxa(Fig. 8a, b, e). Hauschke and Wilde (1987, 1996) consideredthe dark shales as fill of either an oxbow lake (resulting frommeander cut-off) or a floodplain lake, respectively. Furthersupport of this interpretation was provided by Seeling (1999).Recently a systematic excavation that included also a smallabandoned quarry at the so-called ‘Langenberg’ close to theFuchsberg provided a great number of arthropods (includinglimulids, conchostracans and insects) and plants (Barth 2011;Barth and Kozur 2011).
The broad evidence of a shallow subaquatic habitat chal-lenged us to revise the depositional model of Appel (1981)and prove the note of Hauschke and Wilde (1996) that theFossillagerstaette at Fuchsberg resembles some features of thefluvio-deltatic Voltziensandstein. Based on analyses oflithofacies and sedimentary architecture, we demonstrate thedeposition of sandstones within distributary channels andmouth bars of a fluvial-dominated delta that tributed into alarger freshwater to slightly brackish interdistributary bay.
Accordingly, we simplify the formation of bonebeds asstorm event layers (tempestites). We consider the deltaic en-vironment as habitat and provide a synopsis of reported taxa
from Fuchsberg/Langenberg including a tentative revision ofthe Rhaetian flora of the Seinstedt area.
Geological setting
In Triassic times, SE Lower Saxony was situated in the south-ern part of the North German Basin, the largest subbasin of theCentral European Basin (CEB), formerly referred to as theGermanic Basin. The epicontinental CEB extended from east-ern onshore United Kingdom to eastern Poland and fromsouthern Scandinavia to northern Switzerland. The CEB wassituated at palaeolatitudes between 35° and 50°N (Stampfli andKozur 2006) and was bordered by the Fennoscandian Highand British-Scandinavian Caledonides to the north and severalremnants of the Varican orogen to the south (Ziegler 1990).
After the Variscan orogeny, the separated North and SouthPermian Basins formed in Late Carboniferous–Early Permiantimes and received considerable amounts of subsidence fromthermal cooling and crustal stretching (Bachmann and Grosse1989; Littke et al. 2008). The stage of basin initiation wasfollowed by a longer stage of basin fill extending from Permianto Early Cretaceous times. Due to successive growth, the Northand South Permian Basins melted together and formed the CEB(Stollhofen et al. 2008). In the basin centre, the Permian-Cretaceous basin fill can reach up to 12 km in thicknesses(Benek et al. 1996; Scheck et al. 1996; Scheck and Bayer 1999).
Lithologies of the typical Late Triassic Keuper basin fill aredominated by variegated shales of terrestrial Playa-like toSabkha-like environments (Nitsch 1996; Reinhardt andRicken 2000). Evaporites occur frequently and modificationby pedogenic processes is a common feature. The sedimenta-tion of coarse-grained siliciclastics was limited to the marginsof the CEB (Franz 2008). Several short-term ingressions fromthe Tethyanwaters reached the CEB through gates to the southand southeast but were mainly limited to the southern parts ofthe basin (Bachmann et al. 2010).
In Latest Norian–Earliest Rhaetian times, the break-upof Pangaea opened new gates at the western margins of theCEB and the ‘Rhaetian Transgression’ triggered the fun-damental change in palaeogeography. The ‘Rhaetian Sea’covered western and central parts of the CEB. Largeamounts of siliciclastics were supplied from northern andsouthern source areas and formed large fluvial-dominateddeltaic plains in NE, Central and South Germany, whereasin eastern parts of the CEB, terrestrial fluvial to Playa-likedepositional environments were maintained (Franz 2008;Pieńkowski et al. 2012). The strata exposed at Fuchsbergand Langenberg are of latest Norian to earliest Rhaetianage (Barth and Kozur 2011; Hauschke and Kozur 2011)and originate from southern source areas (Appel 1981;Lewandowski 1988; Franz 2008). The exposures formeddue to salt tectonics that uplifted Late Triassic strata
496 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

around the so-called ‘Fallstein’ and other salt structures(Röhling 2002).
Localities and methods
Fuchsberg and Langenberg are small hills in the vicinity of thevillage Seinstedt located in SE Lower Saxony (Fig. 1). At bothlocalities, Upper Keuper sandstones have been quarried butboth quarries are now abandoned. The quarry at Fuchsbergexposes an about 150 m long and up to 13 m high quarry face,and the quarry at Langenberg exposes an about 20 m long andup to 5 m high quarry face. The exposed successions in bothquarries dip gently with 2–5° towards the NNW.
As facies varies slightly from east to west, the eastern partof the Fuchsberg quarry exposes a more sandy succession thatbecomes dissected by intercalated shaly and heterolithic layersin the upper part. Towards the western part, thicknesses ofintercalated layers increase up to some decimetres contribut-ing to an overall upward decrease in sand/silt ratio. The furtherdecrease of sand/silt ratio at Langenberg contributes to thisoverall trend (Figs. 2 and 10).
The outcrops at Fuchsberg and Langenberg were analysedto characterise their depositional environment in terms oflithofacies (Table 1) and biofacies. Based on grain-sizes andphysical bedding structures, 9 lithofacies types are identified.For detailed grain-size analyses, 16 sandstone samples weretaken from Fuchsberg and sieved with standard mesh sievesafter disintegration (Table 2). The granulometric values of
LowerSaxony
Berlin
0 200 km
d
c
ba
Germany
Saxony-Anhalt
Seinstedt
Achim
Börßum
Hedeper
Wetzleben
B82
FuchsbergLangenberg
0 1 2 km
Timmern
130
100
120120
110
120
A
A'Langenberg
Fuchsberg
F1 F2F8
L1
F9 F10F11F12
F5F4F3
F13F6 F7
5770
500
5770
800
4408000 4408500
N
0 100 200 300 400 m
KeyF1 number of section
track
contour line120
120
110
FuchsbergLangenbergWSW ENE
coveredoutcrop
outcrop
A A'
L1
F1
F2F3 F4 F5 F6 F7
100 m
15 m
elev.[m]
Palaeobio Palaeoenv (2014) 94:495–528 497
Fig. 1 Locations and outcrops. a, b Overview and local map of theSeinstedt area (Lower Saxony), modified according to Benda (1963). cDetailed contour line map and measured sections at Fuchsberg (F1–F13)and Langenberg (L1). Contour lines based on NIBIS-Map Server (http://
www.lbeg.niedersachsen.de/portal/live.php). d Cross-section fromLangenberg to Fuchsberg (A to A'), light shading outcrops, darkshading covered, overgrown
Author's personal copy

S. o
lsen
i
Tria
ssic
Rha
etia
nN
oria
nLo
wer
Rha
etia
nS
evat
ian
Gre
goriu
sella
pol
onic
aG
rego
riuse
lla p
olon
ica
- S
hipi
ngia
ger
bach
man
ni
GT
r 18
GT
r 17
Enz
onal
aspo
r. -
Con
bacu
latis
p.G
ranu
lope
rcul
atip
ollis
Cor
ollin
a-P
orce
llisp
ora
Cor
ollin
a-E
nzon
alas
porit
es
A B1B2 B3B
“GrünerGrenzhorizont“
clay silt fs
MFL MFL
P8
Corollina-Enzonalasporites
Shipingia gerbachm.Shipingia etzoldiEuestheria sp.
Shipingia gerbachm.Shipingia etzoldiEuestheria sp.
Granuloperculati-pollis rudisEnzonalasp. spp.Classopollis spp.
Classopollis spp. Granulopercul. rudisEnzonalasp. vigens
Classopollis spp. Granulopercul. rudisEnzonalasp. vigens
Classopollis spp. Granulopercul. rudis
Granulopercul. rudisClassopollis spp.
Shipingiagerbachmanni
Shipingia gerbachmanni
Shipingia gerbachm.Shipingia etzoldiEuestheria sp.
Gregoriusella pol.Shipingia sp.
Gregoriusella pol.
0
3
m
2
1
4
5
6
7
8
360 m
15 m
grain sizeclay silt fs ms
LangenbergL1
FuchsbergF1
FuchsbergF2
tempestite
76263
76262
76255
Classopollis spp.Granulopercul. rudisEnzonalasp. spp.
76186
Classopollis spp.Granulopercul. rudisEnzonalasp. spp.
76185
76256
76187
65027
65028
65029
65030
a
8
9
10
11
12
b
0 m
2
1
FuchsbergF 13
sandstone, silty
siltstone, clayely
claystone
(through)cross-bedded
Key
ripple cross-bedded
conchostracans
fishes
phosphorite
B1
B2
B3
B Biostratigraphy
A Chronostratigraphy
plant detrituslimulids
insectscrustaceans
MFL = main fossil layersample conchostracans
sample palynology(No. BGR)
low-anglecross-bedded
horizontalbedding
hummockycross-strat.
wavy bedding
lenticularbedding
heterolithes
glauconite
shark teeth
Kozur and Weems 2010/11 Barth and Kozur 2011
Conchostracans
Heunisch 1999
Lund 1977
Palynomorphs
Palynomorphs
lithology bedding
storm eventlayer(tempestite)
scales
conglomerate
amphibianbones
bone
bed
II
(bonebed I)
Fig. 2 a Representative lithologs of the successions exposed atFuchsberg and Langenberg with sample points. The correlation betweenboth localities is based on the occurrences of Shipingia gerbachmanni(Hauschke and Kozur 2011), the comparable order of strata and the ‘Main
Fossil Layer’ (Benda 1963, Hauschke and Wilde 1996, Barth 2011). P8reconstructed according to Hauschke and Wilde (1996). b Lithology ofsection F13, a small outcrop north of the main quarry at Fuchsberg(Fig. 1c) that is most probably slightly younger
498 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

Tab
le1
Lith
ofaciestypes
LFT
Lith
ofacies
Descriptio
nContacts
Interpretatio
nLocality
IMassive
mudstone(“fluidmud“)
Apparently
unstratified,dark
grey
toblack,up
to3cm
thick,lateralextension
limited
tosomemeters;plantd
ebris
andinvertebrateremains
Sharp
base
andtop
Rapid
depositio
nof
suspended
load
from
buoyantp
lumes,fluid
mud
Langenberg
IILam
inated
claystone/siltstone
Horizontallam
inated,light
todark
grey,partly
brow
nish,lam
inae
upto
1mm
thick,units
upto
5cm
thick,plantd
ebrisand
invertebrateremains
Bases
mostly
sharp,top
gradationaltoLFT
2and4
Suspendedload,settling
from
suspension
Langenberg,Fu
chsberg;
Fig.
4c,d
III
Sandyheterolithics
(flaserb
edded)
Veryfine
tofine
grainedsandstone
andirregularclay
flasersform
upto
4cm
thicksets,sandstone
ripple
cross-bedded
tohorizontal
laminated,plant
debris
BaseandtopgradationaltoLFT
3and4
Repeatedchangesof
sedimentinput
andmodulationby
waves
andcurrents,
bedload
predom
inates
Langenberg,Fu
chsberg;
Fig.
4a
IVClayeyheterolithics
(lenticular
bedded)
Lam
inated
claystone-siltstone
andlenticuls
ofvery
fine
tofine
grainedsandstone
form
upto
5cm
thicksets,partly
soft-sedim
entd
eformations,current
ripplesandwaveripples,partly
moderatebioturbatio
n,plantd
ebris
Bases
mostly
sharpsm
all-scale
fining-upw
ardtrends;g
radatio
nal
toLFT
2and4
Repeatedchangesof
sedimentinput
andmodulationby
waves
andcurrents,
suspendedload
predom
inates
Langenberg,Fu
chsberg;Fig.
4a,b,d
VRipplecross-bedded
sandstone
Veryfine
tofine,rarelymedium
grained
sandstone,lig
htgrey,asymetriccurrent
ripples,individualsetsup
to1cm
high,
stackedsetsup
to2.5cm
thick;
partly
with
scoursurfaces,plant
debris
Sharpto
gradationalb
aseandtop
Low
erplanebedmigratin
gripples,bedload
Langenberg,Fu
chsberg;
Fig.
4a,b
VI
Horizontallam
inated
sandstone
Fine
tomedium
grained,lig
htgrey
towhitish;
setsup
to2cm
thick,
nograinsize
trend
Sharpbase
andgradationaltop
Planebedflow
(criticalflow
)Fu
chsberg;
Fig.
3c
VII
Low
-anglecross-bedded
sandstone
Medium
tocoarse
grained,partly
pebbly
inchaotic
texture,light
grey
towhitish,
toe-setd
ip<10°,individualsetsup
to10
cmthickoccuras
eventlayers,partly
ripup
clasts(upto
4cm
)atbase
consistingof
underlying
heterolithic
background
facies,accum
ulationof
teethandsm
allb
ones
from
aquatic
anim
als(m
ainlyfishes)in
upto
20cm
thickbonebedlayers
Sharpanderosionalb
ase,
sharptop,laterally
bounded
Planebedflow
(criticalflow
),back
flow
ofsuspension
currentstriggered
bystorm
events(tem
pestite),
conglomeraticpartsdepositedfrom
avarietyof
processesin
shallow
water
andassociated
with
bonebeds
Fuchsberg,Langenberg;
Fig.
4d
VIII
Troughcross-bedded
sandstone
Fine
tomedium
grained,lig
htgrey
towhitish,
toe-setstangentialtoconcaveshaped,trough
size
upto
6m
wideand1m
thick,foreset
laminae
upto
1cm
thick
Sharpanderosionalb
ase,
topgradationaltoLFT
5Low
erplanebed,migrating3-Ddunes,
tangentialtoconcaveshaped
toe-sets:
increasing
velocity
andsuspension
Fuchsberg;
Fig.
3e
IXHum
mocky
cross-stratified
sandstone
Fine
tomedium
grained,lig
htgrey
towhitish,
3-Dripplesup
to5cm
,wavelengthup
to15
cm,primarytextureoftennotrecognisable
Mostly
sharpatbase
tovarious
LFT
s;mostly
gradationalat
topto
variousLFT
s
Waveactio
nbelowfairweatherwavebase
Fuchsberg;F
ig.3g
Palaeobio Palaeoenv (2014) 94:495–528 499
Author's personal copy
mean grain-size and sorting were calculated according to Folkand Ward (1957), and the skewness calculated according toWarren (1974). According to its size and good exposure, theoutcrop at Fuchsberg was chosen to document the overallfacies architecture, stacking pattern and transport directions.In combination of depositional mode, geometry of sedimen-tary units and palaeocurrent measurements, 3 lithofacies as-sociations of a lower delta plain are concluded. In order tocharacterise lithofacies associations in terms of geochemistryand reconstruct palaeoredox conditions bulk rock sampleswere taken at both localities every 10–15 cm. FromFuchsberg, 50 samples and, from Langenberg, 46 sampleswere analysed at BGR Hannover, Germany, using X-rayfluorescence spectroscopy (XRF) for measuring major ele-ments and a number of trace elements. In addition, manualgamma-ray measurements were carried out at both sectionsusing a ‘Heger-Breitband’-γ-scintillometer.
For palynological analyses, 34 samples from Fuchsbergand 2 samples from Langenberg were treated with hydrochlo-ric (10%) and hydrofluoric acid to remove carbonates andsilicates, respectively. Following this, insoluble residues wereultrasonically sieved (mesh size: 10 μm) and organic residueswere mounted on slides with glycerin jelly. The slides wereexamined carefully and palynostratigraphically classifiedaccording to Heunisch (1999).
Stratigraphy
The two outcrops lie some 400 m apart and show some minorvariations in terms of lithofacies. Despite this, they can bereliably correlated due to conchostracan biostratigraphy andtheir corresponding order of strata: the overall upward de-crease of sand/silt ratio and the occurrence of prominent
marker beds. These are the fossiliferous layer so-called‘Main Fossil Layer, MFL’ (Hauschke and Wilde 1996) and atempestite bed (Fig. 2).
In terms of biostratigraphy, the succession exposed atFuchsberg was considered to be of Rhaetian age since thenineteenth century. This was based on (1) plant remainsthought to represent a typical Rhaetian assemblage (Brauns1862, 1866) and (2) regional mapping placed the Fuchsbergstrata slightly below the so-called Contorta beds that have beenexposed in the ‘Große Sandgrube’ north of Seinstedt andcomprised the Rhaetian guide form Rhaetavicula contorta(Behrend 1927; Jüngst 1928, 1929; Benda 1963). Later, thesuggested Early Rhaetian age was adopted by Appel (1981),Look (1984), Koch and Schneider (1985), and Hauschke andWilde (1987, 1996). Palynostratigraphic data seem to confirman Early Rhaetian age of the complete succession at Fuchsberg(Heunisch 1998; Seeling 1999; Heunisch 2011a, b). Thesporomorph assemblages of 9 out of 36 investigated samplescomprise Granuloperculatipollis rudis, Enzonalasporites
Table 2 Granulometrical dataset for samples from Fuchsberg
No. LFT Cumulative weight % for sieve mesh [Φ] Cumulative weight % for class interval [Φ] Size [Φ] Sorting Skewness
0.0 1.0 1.5 2.0 2.3 3.0 3.3 4.0 5.0 5.6 Q5 Q16 Q25 Q50 Q75 Q84 Q95 after Folk and Ward 1957
P1 8 0.0 0.0 0.0 1.3 11.8 81.4 91.8 97.9 99.3 100.0 2.11 2.35 2.46 2.70 2.93 3.08 3.65 2.71 0.42 0.14
P2 8 0.0 0.0 0.0 0.0 0.9 69.0 90.8 99.3 99.9 100.0 2.35 2.47 2.58 2.82 3.08 3.22 3.65 2.84 0.38 0.18
P4 8 0.0 0.0 0.0 31.5 60.5 94.7 97.4 99.4 99.8 100.0 1.58 1.75 1.90 2.21 2.61 2.80 3.01 2.25 0.48 0.12
P5 8 0.0 0.0 0.0 0.5 5.8 92.0 98.5 99.7 99.9 100.0 2.26 2.39 2.47 2.68 2.85 2.93 3.15 2.67 0.27 0.00
P6 8 0.0 0.1 0.1 3.5 27.8 83.4 87.5 95.3 99.3 100.0 2.02 2.15 2.29 2.59 2.89 3.04 3.96 2.59 0.52 0.21
P7 8 0.0 0.0 0.0 4.3 23.1 66.0 74.6 92.4 98.7 100.0 2.00 2.19 2.35 2.75 3.32 3.68 4.38 2.87 0.73 0.31
P8 5 0.0 0.0 0.0 0.1 0.2 3.8 17.6 87.7 98.8 100.0 3.02 3.29 3.40 3.62 3.86 3.96 4.65 3.62 0.41 0.14
P9 8 0.0 0.1 0.1 52.8 76.5 92.4 95.7 98.9 99.7 100.0 1.55 1.65 1.73 1.98 2.30 2.65 3.28 2.09 0.51 0.42
P11 5 0.0 0.0 0.0 0.3 0.5 6.5 22.4 96.8 96.8 100.0 2.81 3.19 3.35 3.58 3.79 3.87 3.98 3.54 0.35 −0.23P12 5 0.0 0.0 0.0 0.1 0.2 6.5 50.8 96.8 99.6 100.0 2.82 3.08 3.15 3.32 3.68 3.80 3.97 3.40 0.35 0.24
P13 5 0.0 0.0 0.0 0.0 0.3 20.9 67.4 97.9 99.7 100.0 2.48 2.82 3.03 3.21 3.48 3.68 3.92 3.24 0.43 0.04
P16 8 0.0 0.7 0.7 54.6 76.1 92.5 98.0 99.8 99.9 100.0 1.54 1.62 1.72 1.95 2.31 2.65 3.14 2.07 0.50 0.43
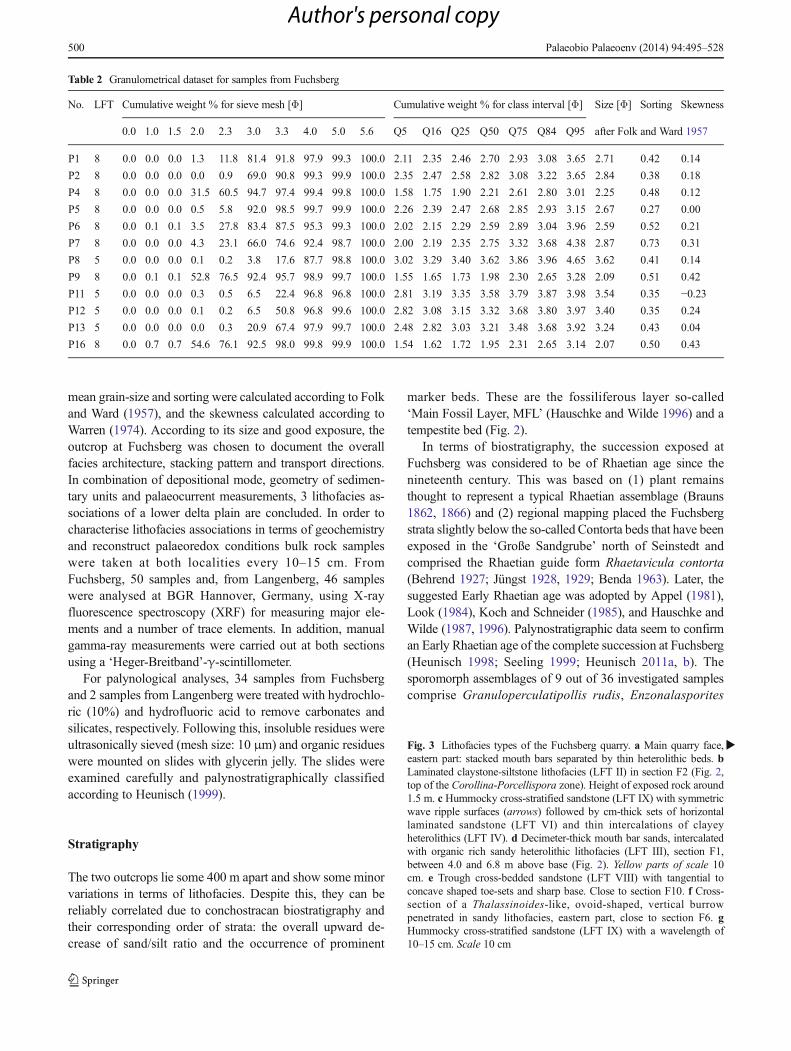
�Fig. 3 Lithofacies types of the Fuchsberg quarry. a Main quarry face,eastern part: stacked mouth bars separated by thin heterolithic beds. bLaminated claystone-siltstone lithofacies (LFT II) in section F2 (Fig. 2,top of the Corollina-Porcellispora zone). Height of exposed rock around1.5 m. c Hummocky cross-stratified sandstone (LFT IX) with symmetricwave ripple surfaces (arrows) followed by cm-thick sets of horizontallaminated sandstone (LFT VI) and thin intercalations of clayeyheterolithics (LFT IV). d Decimeter-thick mouth bar sands, intercalatedwith organic rich sandy heterolithic lithofacies (LFT III), section F1,between 4.0 and 6.8 m above base (Fig. 2). Yellow parts of scale 10cm. e Trough cross-bedded sandstone (LFT VIII) with tangential toconcave shaped toe-sets and sharp base. Close to section F10. f Cross-section of a Thalassinoides-like, ovoid-shaped, vertical burrowpenetrated in sandy lithofacies, eastern part, close to section F6. gHummocky cross-stratified sandstone (LFT IX) with a wavelength of10–15 cm. Scale 10 cm
500 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

Palaeobio Palaeoenv (2014) 94:495–528 501
Author's personal copy

vigens, Rhaetipollis germanicus, Ricciisporites tuberculatusand other diagnostic forms and are therefore assigned to theGTr 18 zone (Heunisch 1999) and the Rhaetian part of theCorollina-Enzonalasporites zone (Lund 1977), respectively.However, the assemblages of other samples have wider rangesfrom the Late Norian to Early Rhaetian (Table 6).
A slightly different biostratigraphic age is suggested byconchostracans (Hauschke and Kozur 2011). The lower partsof both successions comprise a monospecific fauna of largeconchostracans with Shipingia gerbachmanni and Shipingiaetzoldi and can therefore be assigned to the Latest Sevatiangerbachmani zone (Barth and Kozur 2011). The uppermost partof the Fuchsberg succession comprises a monospecific faunadominated by small Gregoriusella polonica that constrains thisinterval to the polonica zone of the earliest Rhaetian age (Kozurand Weems 2010). The interval where large Shipingia andGregoriusella polonica co-occur represents the transitions fromthe gerbachmanni zone to polonica zone and is consideredNorian/Rhaetian boundary interval. It is located in the upperpart of the exposed strata at Fuchsberg about 4 m below the so-called ‘Grüne Grenzhorizont’ (Fig. 2, section F2).
Lithofacies analysis
Depositional cycles and events
The quarry faces at Fuchsberg and Langenberg are orientedW–E (Fig. 2). As the axes of measured trough cross-bedded units(n=13) are directed towards the south with a meanpalaeocurrent of 183°, the quarries provide a cross-sectionalview perpendicular to the mean palaeoflow direction.Internally, the exposures are composed of vertically stackedmetre-scale sedimentary bodies (Fig. 4). Despite slight lateralfacies shifts, three characteristic vertical successions oflithofacies are to be observed. These centimetre- to metre-scalesuccessions are related to shifts in grain sizes and beddingstructure and thus reflect diagnostic changes of depositionalmode. Because of their repeated occurrence, they are consid-ered depositional cycles (Shukla et al. 2006; Zimmermann et al.2014) of lower delta plain environments. In contrast, character-istic centimetre-scale successions of coarse-grained lithofaciesoccur only irregularly and are scattered in both exposures. Theyare considered episodic depositional events of higher energy.
Small-scale coarsening upward cycles
Up to 2-m-thick depositional cycles show gradual shifts fromhorizontal-laminated siltstone-sandstone to ripple cross-beddedsandstone lithofacies. The silty to sandy successions are em-bedded in heterolithic lithofacies and their internal gradationalshift from silty to sandy lithologies is recognised as overallcoarsening upward trend in grain sizes. At the base, the
transition zone from heterolithic lithologies to horizontal-laminated siltstone-sandstone varies from some centimetres tosome decimetres in thicknesses. In some cases, the horizontal-laminated lithofacies is missing and ripple cross-beddedsiltstone-sandstone evolves from heterolithic lithologies due toupward increase in sand and decrease in shales. At the top, thisis always sharp and pronounced ripple cross-bedded lithofaciesappears diffused and often superimposed by hummocky cross-stratification (Table 1; Figs. 3g and 4d).
The gradual vertical change from heterolithic backgroundfacies to sand-dominated lithofacies and vice versa suggest thesubsequent progradation or shift of more proximal sedimen-tary environments during a depositional event. Heterolithesreflect variable input of suspended load into subaquatic envi-ronments and subsequent modification by wave action andunidirectional flows (Dumas and Arnott 2006). The change tohorizontal-laminated siltstone-sandstone lithofacies at the ba-se of individual metre-scale coarsening upward cycles pointsto an increase of sand content within the suspended load. Asmodification by wave action is strongly reduced, an increasein sedimentation rate may be suggested. The subsequentcoarsening in grain sizes triggered the change from asuspended load- to a bed load-dominated mode of transport.Low energy currents of lower flow regime formed currentripples and their successive migration produced up to a fewdecametres thick cosets of non-climbing ripple cross-beddedlithofacies (Reineck and Singh 1980; Allen 1982; Southardand Boguchwal 1990). The diffused top and its superimposi-tion by hummocky cross-stratification indicate subsequentreduced sediment input and reworking by wave action(Harms 1975). Accordingly, the heterolithic background fa-cies terminated the meter-scale coarsening upward cycles.
Small-scale fining upward cycles
Depositional cycles of up to 2 m thicknesses that show internalfining upward in grain sizes are laterally associated to coarseningupward cycles (Fig. 4a). These cycles represent the fill of broadand shallow channel-like structures that cut a few decametresdeep into ripple cross-bedded lithofacies. Following the sharpand erosional base large-scale trough cross-bedded lithofaciespredominates and individual troughs reach up to 4 m in widthand 0.6 m in depth (Figs. 4b and 10). Towards the top, the scaleof troughs decreases successively and ripple cross-beddedlithofacies forms the upper part of the channel fill. At the top,ripple cross-bedded lithofacies becomes diffused and issuperimposed by hummocky cross-stratification (Fig. 3g). Assubordinated inclined bounding surfaces that would represent
�Fig. 4 Depositional cycles and depositional events: a Mouth barprogradation cycle. b Distributary channel fill cycle. c Interdistributarybay fill cycles. d Tempestite. fsi fine silt, msi medium silt, csi coarse silt,vfs very fine sand, fs fine sand, ms medium sand
502 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

Palaeobio Palaeoenv (2014) 94:495–528 503
Author's personal copy

accretion surfaces are absent, the channel fill appears simple andis therefore considered single-storied.
The succession of lithofacies clearly shows that the formationof fining upward cycles was associated to erosion and highertransport velocities in the early stage, successively lowered trans-port velocities in the late stage and reworking by wave action inthe final stage. Higher transport velocities in the early stage areevidenced by slight incision of broad and shallow channelsaccompanied by non-deposition and sediment bypass. The sim-ple fill of channels with large-scale trough cross-beddedlithofacies points to lowered but still high transport velocities ofthe lower flow regime as indicated by concave- to tangential-shaped toe-sets (Allen and Collinson 1974; Allen 1982; Ashley1990). The subsequent decrease in the scale of trough cross-bedded sets and the change to ripple cross-bedded lithofaciestowards the top reflects the decrease to slow transport velocitiesof the lower flow regime (Southard and Boguchwal 1990).Finally, transport ceased and the deposited strata became subjectto modification by wave action.
Micro-scale coarsening upward cycles
At Fuchsberg and Langenberg, an up to 1 m thick successionof laminated claystone-siltstone lithofacies (LFT II) is formedof numerous up to 3 cm thick individual couplets of dark greylaminated claystone and yellowish to greyish laminated silt-stone. Each couplet is bounded by a more or less sharp baseand top and characterised by internal coarsening up in grainsizes (Fig. 4c). At bedding planes, plant debris and smallremains of vertebrates and invertebrates occur abundantly.Modification of physically bedding structure by faunal activi-ties has not been observed. Both lithologies forming a couplethave been deposited from suspended load of a low energysubaquatic environment (Elliott 1974). The shift in grain size iscommonly attributed to increased input of suspended loadfrom a neighbouring feeder system (Coleman et al. 1964;Elliott 1974).
The overall stacking pattern of couplets reveal their cyclicnature and resembles a ‘varve-like’ structure. Within this stackingpattern of small-scale cycles, the repeated variations of grain sizesand thicknesses over a series of couplets suggest four bundles ofcouplets. These bundles comprise 105, 30, 20 and 7 individualcouplets and evidence variations of input superordinated to dis-charge variations that produced individual couplets (Barth 2011).Such superordinated input variations may originate from lateralshifts of feeder systems or their progradation (Tye and Coleman1989). Both, couplets and their bundles, contribute to the fill of alow energy subaquatic environment.
Tempestite beds
Within the successions at Fuchsberg and Langenberg, a prom-inent up to 10 cm thick bed is formed of coarser grained
lithofacies types. According to conchostracan biostratigraphyand the corresponding order of strata, the beds occur in com-parable levels and are therefore considered exposures of asingle marker bed (Figs. 2 and 10).
With a sharp and slightly erosive base, sandy and clayeyheterolithics are terminated by coarse grained and low-anglecross-bedded sandstone lithofacies (Fig. 4d). Sets of thislithofacies are up to 7 cm high and comprise partly abundantrip-up clasts. At the top, they show fining up in grain sizes andare followed by ripple cross-bedded sandstone lithofacies.Isolated trains of ripples are up to 3 cm thick. Their internalasymmetric structure shows parallel foreset lamina that dip inthe same direction and suggest the formation at the lee side ofmigrating current ripples (Jopling and Walker 1968; Kumarand Sanders 1976; Aigner 1985; Myrow and Southard 1996).In contrast, the external shape of the ripples is symmetric andindicates modification by wave action subsequent to theirformation as current ripples (Aigner and Reineck 1982,1983; Hunter and Clifton 1982; Aigner 1985). According tothis, the top of the marker bed comprises a prominent rippledsurface with wave ripples of up to 3 cm amplitude and amaximum wave length of 15 cm (Fig. 4d). In addition, thetop of the coarse-grained marker bed is highlighted by a sharpcontrast in lithology to the overlying dark laminated claystonelithofacies (Fig. 4d). The claystone bed is up to 7 cm thick andgrades towards the top into sandy and clayey heterolithics.
The heterolithic lithofacies below and above the marker bedis commonly attributed to variable input of suspended load andsubsequent modification by waves and slow currents (Reineckand Singh 1980). Here, it represents the background facies of arather low energy environment. According to its grain size andphysical bedding, the marker bed clearly evidences higher ener-getic conditions for at least a short time. The erosive reworking atthe base responsible for the formation of rip-up clasts suggesterosion and transport by high-energy currents. The mud clastsare intraformational in origin and contradict a longer transport(Smith 1972). Subsequent lowering of transport velocitiesenabled the deposition of low-angle cross-bedded sand-stone lithofacies under conditions of critical flow (Miall1977; Rust 1978). Following this, the further decrease oftransport velocities produced slow migrating trains of cur-rent ripples of the lower flow regime (Jopling and Walker1968). Cessation of directed transport is indicated by theformation of symmetric ripples under oscillating conditionsthat are superimposed on the top of the marker bed (Aignerand Reineck 1982, 1983; Harms 1975). The dark laminatedclaystone lithofacies at the top represents depositedsuspended load and segues into the heterolithic backgroundfacies.
The isolated occurrence of this thin marker bed and itsinternal succession of coarse-grained lithofacies that dif-fers remarkably from the strata below and above, and areevidence of its formation as an event layer. Compared to
504 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

the background facies, the short-term establishment ofhigh-energy currents in a lower energetic environmentare the main features concluded. These currents intro-duced coarse-grained sands from more proximal to distaldepositional environments and incorporated bone frag-ments and teeth of aquatic animals as well as erodedmud clasts. As the deposition has been triggered by a decreasein velocity, we conclude that this load has been transported insuspension. The short-term occurrence of suspension currents thatintroduce coarser sediment fromproximal tomore distal facies is atypical feature of tempestites of shallow storm-influenced deposi-tional environments (Kumar and Sanders 1976; Reineck andSingh 1980; Aigner 1985; Morton et al. 2007).
Granulometry
Sandstones from coarsening upward and fining upward cyclescan be readily distinguished according to their grain size
distribution. Sandstones from the former are mainly com-posed of very fine sand and comprise only subordinatedproportions of finer (up to 12.3 % silt) and coarser grain sizeclasses (up to 20.9% fine sand). Sandstones from the latter aremainly composed of fine sand with subordinated finer (up to34% very fine sand and silt) and coarser grain size classes (4.3%medium sand). Two samples are composed of medium sandwith up to 47.2 % finer and 0.7 % coarser grains (Fig. 5a). Asripple cross-bedded lithofacies is mainly present in coarseningupward cycles and trough cross-bedded lithofacies is onlypresent in fining upward cycles the differences are clearlyrelated to the mode of transport. Accordingly sandstones fromfining upward cycles have mean grain sizes between 2.09 and2.87 Φ, are very well to moderate sorted (0.27–0.73 Φ) andnear symmetrical to strongly fine-skewed (0.0–0.43 Φ).Sandstones from coarsening upward cycles have mean grainsizes between 2.09 and 2.87 Φ, are generally well sorted(0.35–0.43 Φ) and fine to coarse skewed (−0.23–0.24 Φ).
well
moderate well
0.7
0.8
0.6
0.5
0.4
0.3
Sort
ing
[Φ]
mouth bars
very well
Skew
ness
[Φ]
–0.2
0
0.2
0.4
0.6
distributaries
mouth bars distributaries
0
mean grain size [Φ]
skew
ness
[Φ]
mouth bars
0
20
40
60
80
100
0 1 2 3 4 5 6mean grain size [Φ]
perc
enta
ge [%
]
moderate
coarse-skewed
nearsymmetrical
fine-skewed
stronglyfine-skewed
coarsesand
mediumsand
finesand
very finesand
coarsesilt
mediumsilt
mouth barsdistributaries
distributaries
1.0 1.5 2.0 2.5 3.0 3.5 4.0–1.0
–0.5
0.5
1.0
ba
dc
Fig. 5 Grain size distributions and statistical values. a Cumulative grain-size distribution and grain-size classes according to Wentworth (1922). bBox-Whisker-Plot of skewness shows on average near symmetricalskewedmouth bar sands and on average fine-skewed distributary channel
sands. c Box-Whisker-Plot of sorting demonstrates well-sorted mouthbars sands and very well to moderate sorted distributary channel sands. dSkewness versus mean grain-size discriminates mouth bars sands fromdistributary channel sands
Palaeobio Palaeoenv (2014) 94:495–528 505
Author's personal copy

The impact of transport mode on grain size distribution isfurther demonstrated by plotting skewness valuesstandardised to mean grain-size values (Fig. 5).
Biofacies analyses
Most of the invertebrate fossils (insects, limulids,conchostracans, malacostracans) described from Fuchsbergand Langenberg have been found in the up to 50 cm thickfossiliferous layer referred to as ‘Main Fossil Layer’ (MFL) or‘Fossilführende Wechselfolge’ according to Hauschke andWilde (1996). In addition, some cm-thin layers of laminatedclaystone-siltstone lithofacies yielded accumulations of insectand plant remains and conchostracans. In particular, plantremains are not bounded to distinct layers or lithologies asthey practically occur throughout the successions (Fig. 2). Offurther importance are two bonebeds as they comprise count-less ‘microvertebrate remains’. The term applies to small,microscopic fossil remains of vertebrates, in particular tosmall teeth of cartilaginous and bony fishes, dermal denticlesof selachians and scales of bony fishes. Amphibian or reptilianmicrovertebrate remains could not be identified with certainty.The first bonebed is associated to a tempestite layer that occursin the successions exposed at Langenberg and Fuchsberg
(bonebed I, Figs. 2, 4d and 10a). The second bonebed occursin the small exposure north of the main quarry at Fuchsbergand therefore may be slightly younger (bonebed II, Fig. 2b).Bulk samples were taken from both bonebeds. The samplesfrom the second bonebed were processed according to themethod described by Thies et al. (in prep) and yielded a richmaterial of microvertebrate remains comprising teeth of elas-mobranchs and actinopterygians, dermal denticals of elasmo-branchs and scales of bony fishes. So far, only a small part ofthe material from the first bonebed was desintegrated withhydrogene peroxide and superficially monitored formicrovertebrate remains.
Only the remains of insects, micro- and macrovertebratesand macroflora will be discussed in detail. Other fossil groups(articulated fishes, coelacanthid scales, malacostracans) arecurrently under revision. Most of the collected invertebratematerial from Fuchsberg and Langenberg which is describedin Hauschke andWilde (1987, 1996), Barth (2011), Barth andKozur (2011) and Hauschke and Kozur (2011) is stored in theGeological and Palaeontological Collections, Martin-Luther-University Halle-Wittenberg (Germany) and in the privatecollection of J. Ansorge (Horst, Germany). The holotype ofPaleolimulus fuchsbergensis and a part of the macroflora isstored in Forschungsinstitut Senckenberg (Frankfurt,Germany). The micro- and macrovertebrate remains (includ-ing the Plateosaurus femur) are deposited in the Roemer- and
Table 3 Taxonomy and occurrence of insect taxa at Fuchsberg andLangenberg. For position of sections and MFL see Figs. 1 and 2. D=dominant, R=rare. Classification partly after C. Brauckmann (Clausthal),
E. Gröning (Clausthal), J. Ansorge (Greifswald), J. Schneider (Freiberg),G. Bechly (Stuttgart), A.G. Ponomarenko (Moscow), P. Vršanský (Bra-tislava), and W. Zessin (Schwerin)
Super family/family Specimen Figure Occurrencein section
Order Frequencyof the order
Ephemeroptera Hyatt & Arms, 1891 (mayflies) 1 not attributed larvae R
Odonata Fabricius, 1792 (dragonflies) Triassolestidae Italophlebia baueri Barth et al. 2013
Langenberg, MFL1 not attributed wing fragment
R
Blattodea Brunner, 1882 (cockroaches) Voltziablattidae indet.
Caloblattinidae
24 wings in at least 3 different taxaincluding 1 articulated pair of hindwings
Langenberg +Fuchsberg, MFL
D
Orthoptera Olivier, 1789 (“grasshoppers“) Ensifera: Elcanidae Archelcana Sharov, 1968with color patterns
Langenberg, MFL
?Hagloidea indet. Langenberg, MFL
R
Fuchsberg, MFL R
Scytinopteroidea: Ipsviciidae Ipsvicia langenbergensis Barth et al. 2011
Prosboloidea: Dysmorphoptilidae Dysmorphoptilidae indet.2 imagines with wings, 1 isolated wing
Langenberg, MFL
Langenberg, MFL
Coleoptera Linnaeus, 1758 (beetles) Adephaga: ?Triaplidae ?Triaplidae n. sp.
Hydradephaga Hydradephaga indet.
Polyphaga(complete body preservation)
Family indet.
7f Fuchsberg, MFL
7a, 12a
7c
7e
7d
7bFuchsberg, MFL+ section F2
D
Diptera Linnaeus, 1758 (flies) Nematocera: Eoptychopteridae Eoptychopteridae indet. RFuchsberg
Mecoptera Hyatt & Arms, 1891 (scorpion flies) Pseudopolycentropus Handlirsch, 1906
Mesotanyderus Riek, 1955
RFuchsberg
Hymenoptera Linnaeus, 1758 (wesps, bees, ants) Sepulcidae ?Sepulenia Rasnitsyn, 19681 fore wing
RFuchsberg
Langenberg +Fuchsberg, MFL
Protomyrmeleontidae 2 wings, some fragments
Hemiptera Linnaeus, 1758 (true bugs) Membracoidea: Auchenorrhyncha
Polyphaga indet. ?n. sp.
3 not attributed coleopteran elytra
Membracoidea indet.
Not attributed insect remains 21 wings/fragments1 wing ?Palaeoptera indet.several aquatic larvae
506 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

Tab
le4
Taxonomyandoccurrence
ofchondrichthyan
andosteichthyan
taxa
from
Fuchsberg
Class
Subclass/series
Order
Fam
ilySp
ecies
Figures
Occurrence
inbonebed
Frequency
Chondrichthyes
Huxley,1880
Elasm
obranchii
Bonaparte,1838
Hybodontiformes
Miasey,1975
Polyacrodontidae
Glik
man,
1964
Polyacrodus
sp.
6aI
IIR
Lonchidiid
aeHerman,1977
Lissodus
nodosus(Seilacher,1943)
III
D
PseudodalatiidaeReif,1978
Pseudodalatiasbarnstonensis(Sykes,
1971)
IR
Synechodontiformes
Duffin&
Ward,1993
PalaeospinacidaeRegan,1906
Synechodus
n.sp.1
6b,12h
III
D
Synechodus
n.sp.2
IIR
Rhomphaiodon
minor
(Agassiz,1837)
III
D
Rhomphaiodon
nicolensisDuffin,1993
IIR
Family
inc.sed.
Synechodontiformes
n.gen.n.sp.
6dII
D
Order
andfamily
inc.sed.
PseudocetorhinuspickfordiD
uffin,1998
IIR
Osteichthyes
Huxley,1880
Actinopterygii
Klein,1885
Chondrostei
Müller,1844
[sensu
Patterson,
1982]
Palaeonisciformes
Hay,1929
Acrolepididae
Aldinger,1937
GyrolepisalbertiiAgassiz,1835
IIR
Fam
ilyinc.sed.
Severnichthysacum
inatus
(Agassiz,
1835)
III
D
Neopterygii
Regan,1923
[sensu
Rosen
etal.
1981]
Semionotiformes
Arambourg&
Bertin
,1958
(Under
revision)
7remains
8aMFL
atFuchs-berg(F1)
Perleidiform
esBerg,1937
[sensu
Tintori&
Lom
bardo,
1996]
Colobodontid
aeAndersson,
1916
[sensu
Mutter,2004]
Colobodus
?sp.
IIR
Fam
ilyinc.sed.
Serrolepissuevicus
Dam
es,1888
IIR
Sarcopterygii
Rom
er,1955
Coelacanthiform
esBerg,1937
Coelacanthiform
esinc.sed.,4
scales
8e,12f
MFL
atFuchs-berg(F1)
Ddominant,Rrare.F
orpositio
nof
MFL
,see
Figs.1and2
Palaeobio Palaeoenv (2014) 94:495–528 507
Author's personal copy

Tab
le5
Listo
fplantfossilsin
Seinstedt
(literature
andtoday)
Brauns(1862)
Brauns(1866)
Schenk(1867)
Schim
per(1869,1870–72,1874)
Jüngst(1928)
Thispaper
Anomozam
itessp.
Cyclopteriscrenata
Cyclopteriscrenata
Cyclopteriscrenata
Cyclopteriscrenata
Ginkgocrenata
Aphlebia?
Cam
ptopterisfagifolia
;Cam
ptopterisplanifo
lia;
Clathropterismeniscoides
Cam
ptopterisplanifo
liaCam
ptopterisplanifo
liaClathropterisplatyphylla
Clathropterisplatyphylla
Clathropterismeniscoides
Odontopteriscycadea;Odontopterislaevis
Odontopteriscycadea;
Odontopterislaevis
Pterophyllumsp.;Pterophyllum
crassinerve
Ctenozamitescf.w
olfia
na
Cam
ptopterisexilis
Cam
ptopterisexilis
Dictyophyllu
macutilo
bum
Dictyophyllu
macutilo
bum
Dictyophyllu
mexile
Dictyophyllu
mexile
Thuitesschloenbachi
Thuitesschloenbachi
Hirmerella
muensteri
Laecopterisalternifo
lia;
Pterophyllumsp.dub.
Calam
itesgümbeli;
Pecopteris
grum
brechti
?Laccopterisgoepperti??;
Asplenitesottonis
Pecopterisgumbrechtii;
Asplenitesottonis
Lepidopterisottonis
Lepidopterisottonis
?Taeniopterisvitta
taTaeniopterisvitta
taTaeniopterismuensteri
Angiopteridiummuensteri
Marattia
interm
edia
Graminieen
remains
Calam
iteshoerensis
Calam
iteshoerensis
Equisetiteshoerensis
Neocalamiteshoerensis
Nilssonialin
earis;Za
mitessp.dub.
Calam
itesguem
beli
Calam
itesguem
beli
Equisetiteslehm
anni
Neocalamiteslehm
annianus
Arundinitesdubius;A
rundinitespriscus
Calam
iteshoerensis
Neocalamitessp.
Cycaditesrectangularis
Cycaditesasterocarpoides
Cycaditesrectangularis
Cycaditesrectangularis
Phlebopterisangustilo
ba
Pterophyllummaximum
Pterophyllummaximum
Pterophyllumbraunsii
Anomozam
itesbraunsii
Pterophyllumbraunsii
?Nilssoniasp.
Nilssoniablasii
Nilssoniablasii
Pterophyllumblasii
Pterozamitesblasii
Pterophyllumblasii
Pterozamitesblasii
Nilssoniablasii
Nilssoniablasii
Pterophyllumblasii
Pterozamitesblasii
Ptilozam
itesnilssoni
Ptilozam
itesheeri
Taeniopteristenuinervis
Taeniopteristenuinervis
Taeniopteristenuinervis
Oleandridiumtenuinerve
Taeniopteristenuinervis
Taeniopteristyporientalis
Equisetitesmuensteri
nonvide
Nilssoniaelongata
Nilssoniaelongata
gen.indet.sp.indet.
nonvide
Nilssoniasp.dub.
Nilssoniasp.
nonvide
Pterophyllummuensteri
nonvide
508 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

Pelizaeus-Museum Hildesheim (RPMH). The larger part ofthe described macroflora is stored at the Museum fürNaturkunde/Leibnitz Institute for Research on Evolution andBiodiversity at the Humboldt University Berlin (Mb.Pb.suffix).
Molluscs
Schlönbach (1862) described several marine bivalves andsmall gastropods from shaly and sandy lithofacies of the sandyquarries near Seinstedt: Cardium rhaeticum Merian,Taeniodon praecursor nov. sp., Isocyprina (Taeniodon)ewaldi Bornemann, 1854, Leda deffneri Oppel and Suess,1856?, Mytilus minutus Goldfuss, Gervillia praecursorQuenstedt, 1856, Gervillia inflata Schafhäutl, 1851, andRhaetavicula (Avicula) contorta Portlock, 1843. Most of thesebivalves are typical for Late Triassic environments andthought to indicate marine conditions (e.g. Gervillia) of aprobably slightly younger age (Schlönbach 1862, Benda1963).
Bivales living in most cases microphagous feeding ondetritus-feeding organism (foraminifers, ostracods, diatomsor small molluscs) or plankton (bacteria, green algae,diatoms, flagellates, protozoans or metazoan larvae;Lehmann and Hillmer 1997; Ziegler 1998a). Some bivalvesare scavengers hunting small worms or crustaceans (Lehmannand Hillmer 1997). The occurrence of bivalves is closelyrelated to water temperature, water depth, salinity, substrateand water energy (Ziegler 1998a).
Arthropods
‘Conchostraca’
Hauschke and Wilde (1996), Barth (2011), Barth and Kozur(2011) and Hauschke and Kozur (2011) described 264conchostracans from Fuchsberg and Langenberg, includingShipingia gerbachmanni Hauschke and Kozur, 2011(Fig. 8d), Shipingia n. sp. aff. Shipingia etzoldi Hauschkeand Kozur, 2011, Euestheria sp., and Gregoriusella polonicaKozur and Weems (Fig. 8d).
Conchostracans occur throughout the successions exposedat Langenberg and Fuchsberg and are not bounded to a distinctstratum or lithofacies. They are accumulated at bedding planesaround the MFL and often associated with insect and plantremains (Fig. 2). The majority of Branchiopods (includingconchostracans) are adapted to freshwater habitats (lakes,ponds, fluvial) but also accept brackish habitats or habitatssubject to repeated salinity changes (Retallack and Clifford1980; Storch and Welsch 2004; Lecointre and Le Guyader2006). In contrast to recent forms, Triassic conchostracansprobably did not tolerate higher salinities (Reible 1962). Inaddition, Kobayashi (1954) proposed a correlation between
the temperature of the habitat and the size of theconchostracans, with an optimum under subtropical climate.Conchostracans are feeding on disintegrated plant material(Stern and Stern 1971; Strenth and Sissom 1975; Royan1976; Martin 1989).
Malacostraca
From theMain Fossil Layer, at least two taxa (probably of twosuperorders Syncarida and Eucarida) and nine unassignedfragments of crustacean remains have been reported byHauschke and Wilde (1996) (Fig. 8b). All known remainsare currently under revision (N. Hauschke, MLU Halle-Wittenberg, personal communication). The majority of mala-costracan crustaceans are living in euryhaline marine habitatsbecause of their ability to adjust their osmotic system (Ziegler1998b). But a number of these marine forms are adapted tobrackish habitats, especially tidal environments (Ziegler1998b). Crustaceans are feeding on carrion of epibenthic biota(Lecointre and Le Guyader 2006). Fossil representatives ofthe superorder Syncarida are considered mainly freshwaterforms as only a few have been described from clear marinehabitats, whereas fossil representatives of the superorderEucarida include both marine and freshwater forms (Storchand Welsch 2004).
Limulids
Four specimens of probably one taxon, including their tracks,are described in Hauschke and Wilde (1987, 1996) and Barth(2011). Three of them, including the holotype Paleolimulusfuchsbergensis Hauschke and Wilde 1987 (Figs. 8c and 12g),were discovered at Fuchsberg and one at Langenberg. Asalready mentioned in Hauschke and Wilde (1987, 1996),limulids are very rarely preserved as fossils in Late Triassicstrata. Triassic limulids possessed an aquatic lifestyle, andrecent representatives tolerate large fluctuations of salinity,temperature and suspended load but prefered clean sandyshallow water habitats (Malz and Poschmann 1993; Storchand Welsch 2004; Lecointre and Le Guyader 2006).Xiphosurids are omnivores feeding on bivalves but alsoworms, small arthropods and carrion (Malz and Poschmann1993; Storch and Welsch 2004; Lecointre and Le Guyader2006). During their mating season, limulids are able to leavetheir marine aquatic habitats and temporarily occupy brackishand even terrestrial habitats (Fisher 1979, 1984; Storch andWelsch 2004). Hauschke and Wilde (1991) discussed therelationship between the size of mesozoic limulids and theirpossible habitats and proposed that, contemporaneous tolarge marine specimens, smaller specimens may haveoccupied freshwater environments. However, it cannotbe excluded that the small specimens from Fuchsbergrepresent juvenile forms.
Palaeobio Palaeoenv (2014) 94:495–528 509
Author's personal copy

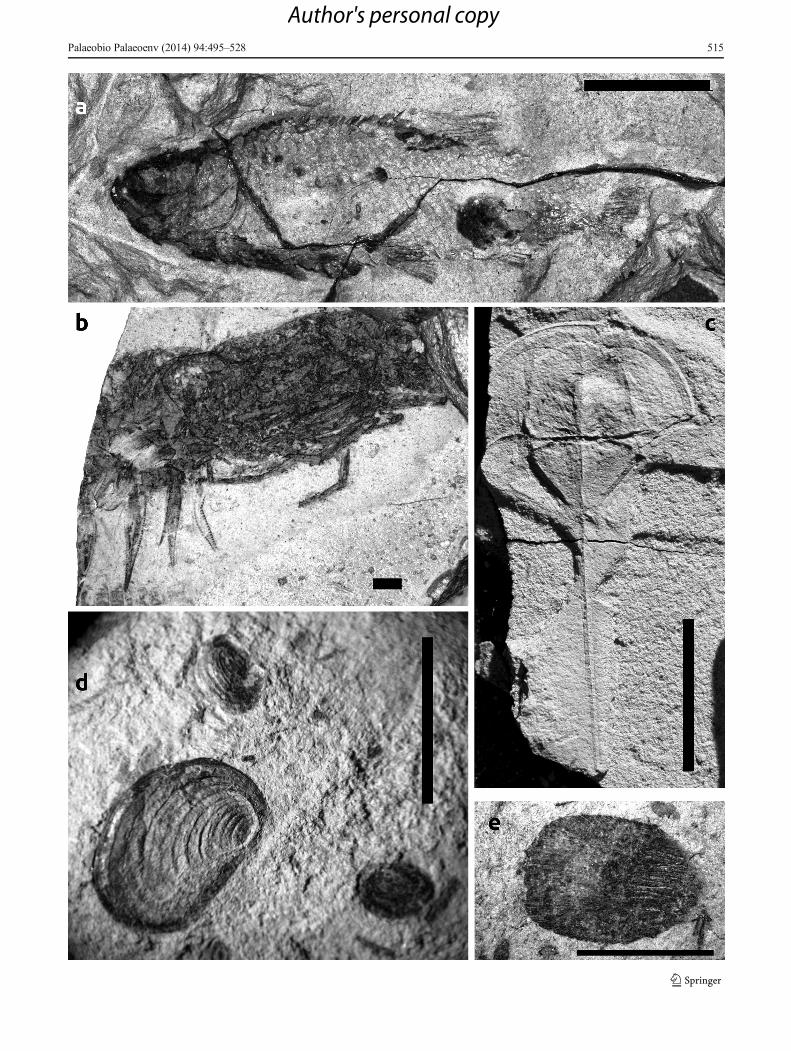
Insects
More than 60 moderately to well-preserved wing/body remains(predominantly isolated wings), often associated with plant debrisand conchostracans, are recorded (Fig. 2). Fine bristles and wingvenation are preserved in most cases. The insect fauna of theSeinstedt area contains typical Late Triassic representatives of atleast nine orders (Table 3). Because some of them are known onlyfrom the Early Jurassic (e.g. Elcanidae, Dysmorphoptilidae; seeTable 3), the coleopteran fauna from Fuchsberg is considered ‘tooevolved for Triassic habitats’ (Ponomarenko, Moscow, personalcommunication). Most known insect remains were found withinthe MFL at Langenberg and Fuchsberg and the strata above, aswell as the lowermost part of section F2 (Fig. 2), and describedand illustrated in Hauschke and Wilde (1996), Barth (2011) andBarth et al. (2011, 2013). In general, the characterisation of insecthabitats remains difficult because of their typical allochthonoustaphocoenosis. However, some details could indicate an at leastpara-autochthonuous taphocoenosis: for example, a pair ofcockroach-hindwings (Fig. 7c) was documented by Barth(2011) that would not have been resistant against transportation.In addition, entirely articulated dysmorphoptilids indicate the veryclose relation between habitat and fossil-lagerstätte (Fig. 7d).
Concerning the order Ephemeroptera (mayflies), a larva(Fig. 7f) reliably indicates good water quality (Hubbard andPeters 1978). Immature mayflies are entirely aquatic and arevery sensitive to water pollution and fluctuations in oxygena-tion (Merrit and Cummins 1978; Ward 1992; Grimaldi andEngel 2005). After Dettner and Peters (2010), recent represen-tatives of mayflies are adapted to habitats in moderate climatesbut also occur in mountainous regions. Mayflies attach theirlarva at or underneath stones or at the ground of shallow waterhabitats. Mayflies are seldom scavengers, mostly they feed onorganic detritus or algae (Dettner and Peters 2010).
True bugs (order Hemiptera) and representatives of theorder Orthopterans (grasshoppers and others) are typical phy-tophagous feeder. Shcherbakov (1992) supposed that the he-mipteran representative Karajassus, which is very similar toone iso la t ed hemip te ran wing (Membraco idea ,MLU.Sei2010.009) from Fuchsberg, was sucking plants.Also, the Dysmorphoptilids are phytophagous feeders(Shcherbakov 2000). Cockroaches (order Blattodea) are om-nivores, and dragonflies (order Ordonata; Figs. 7a and 12a)and partly beetles (order Coleoptera) are scavengers. Recentdragonflies prefer vegetated habitats at lakes and ponds.Recent cockroaches live mostly in subtropical to tropicalclimates, whereas orthopterans are in most cases related tograss-covered plain-like habitats.
Fishes
The occurrences and distribution of chondrichthyes andosteichthyes (micro- and macrovertebrate remains) in the
two bonebeds and the Main Fossil Layer (MFL) atFuchsberg are shown in Table 4. Altogether, ninechondrichthyan and at least six osteichthyan taxa were iden-tified. From the upper part of the MFL, well-preservedsemionotid-like fishes (Fig. 8a) and scales assigned tocoalcanthid fishes are described and illustrated in Hauschkeand Wilde (1996) and Barth (2011). Due to high energytransport and abrasion, most of the microvertebrate remainsfrom the two bonebeds are poorly preserved. The new elas-mobranch taxa Synechodus n. sp. 1 and Synechodontiformesn. gen. n. sp. will be described elsewhere (Thies et al. in prep).
Synechodontiforms are supposed to include only taxa ofmarine environments (e.g. Underwood and Ward 2004).Lissodus nodosus was, however, reported from questionablebrackish and freshwater deposits (Fischer 2008). The occur-rences of both marine and brackish to freshwater sharks in thetwo bonebeds either confirm a brackish habitat of marine andfreshwater influences or can be attributed to processes ofconcentration that formed the bonebeds as ‘concentrateLagerstätte’. According to recent forms, all sharks from theFuchsberg/Langenberg biocoenosis are aquatic carnivors atthe top of the food chain feeding on other fishes and/orcrustaceans (Fig. 12h)
Semionotid fishes are frequently found in Middle to UpperKeuper lacustrine environments and are therefore assigned tofreshwater habitats (e.g. Etzold and Schweizer 2005; Havliket al. 2013). However, elsewhere this fish group is also de-scribed from marine sediments (e.g. McCune 1986; López-Arbarello 2008). According to their jaw structure, semionotidsare aquatic carnivores ‘able to engulf prey by sucking theminto their mouths’ (Murry 1989) or they ‘may have also beenbrowsers or nibblers and their diet may have included a widevariety of nectonic, planktonic and benthonic animals’(Schaeffer and Rosen 1961; Schaeffer 1967; Murry 1989).The articulated duripartic preservation of completesemionotid-like fishes in the MFL suggest that they have beenpart of the autochthonous population and following their deathrapidly embedded without decomposition. However, this issupported by the lack of any bioturbation feature of the MFLbut in contrast to epibenthic crustaceans (malacostracans)which are considered part of a necrophage decomposer fauna.
The habitat of Late Triassic coelacanthiformes is still underdiscussion. According to Müller (in Hauschke and Wilde1996) coalcanthid fishes were adapted to marine
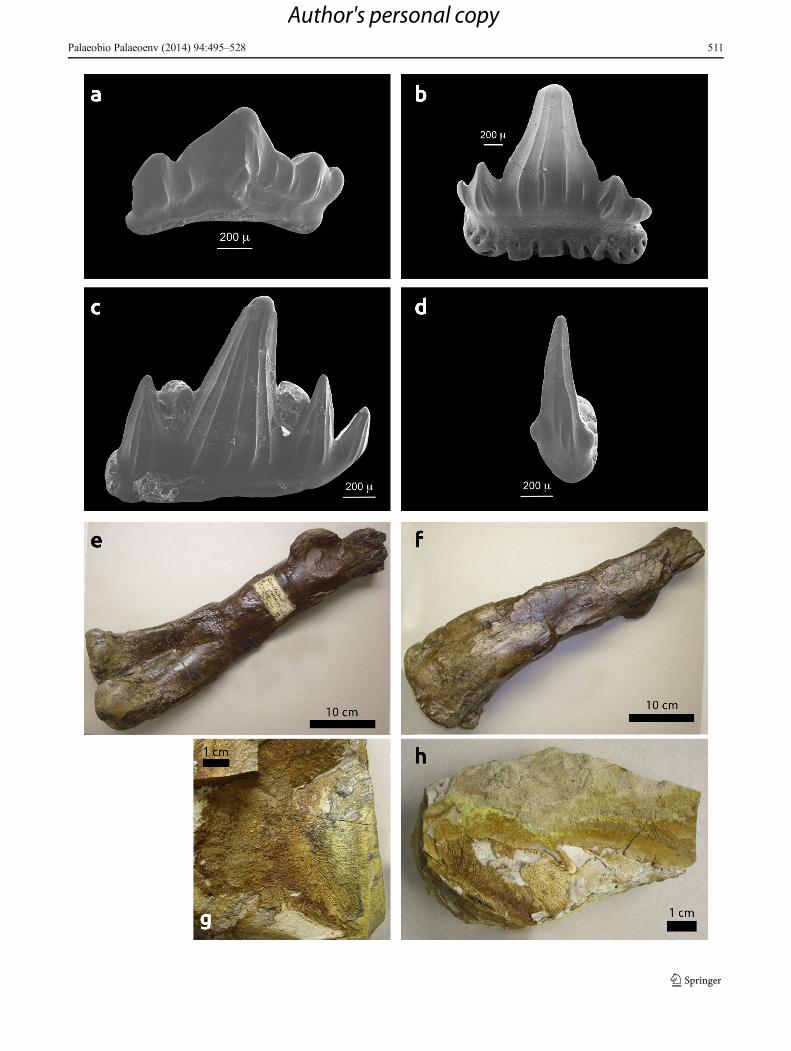
�Fig. 6 Micro- and macrovertebrate remains. a Polyacrodus sp., lateraltooth, from bonebed in section F13, RPMH, cat. no. NKP 3985. bSynechodus n. sp. 1, postero-lateral tooth, from bonebed in section F13,RPMH, NKP 3969. c Rhomphaiodon nicolensis, antero-lateral tooth,f r om bonebed i n s e c t i o n F13 , RPMH, NKP 3986 . dSynechodontiformes n. gen. n. sp., antero-lateral tooth, from bonebed insection F13, RPMH, NKP 3978. e Plateosaurus laevis, sinistral femur,length 0.51 m, RPMH, NKP 13466. f Same specimen from behind. gUndetermined amphibian, imprint of a bony plate, RPMH, NKP 13477. hAnother specimen, RPMH, NKP 13473
510 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy
Palaeobio Palaeoenv (2014) 94:495–528 511
Author's personal copy

environments. In contrast, Caroll (1997) mentioned a fewfreshwater forms among the marine Actinistia of theMesozoic. Also the Late Triassic genus Chinlea Schaeffer,1967 from the Chinle-Formation in North America occurs interrestrial sediments (Yabumoto 2008). However, remnants ofthis fish group are very rare in the Germanic Triassic (Schultzeand Kriwet 1999) and all described remnants have been foundinmarine sediments (Winkler 1880; Seilacher 1943; Gall et al.1974; Schweizer 1966).
The semionotid-like fishes and coelacanthid scales fromFuchsberg are currently under revision (Fig. 12f).
Phytoplankton
Schlönbach (1862) did not mention further specified remnantsof diatoms from Fuchsberg. Accordingly, recent palynologicalinvestigations of Heunisch in Seeling (1999) and Barth (2011)confirm a marine influence (Table 6; Heunisch 1998, 2011a,b). Following the Sporomorph Eco Group (SEG) model ofAbbink (1998) and Abbink et al. (2004a, b), modified for theTriassic by Ruckwied (2009) and Kustatscher et al. (2012), theprasinophycacean genera Tasmanites and Botryococcus areattributed to lagoonal-brackish environments (ALB SEG). Inaddition, Tasmanites may suggest slightly hypersaline condi-tions (Heunisch 1998). The prasinophycacean generaLeiosphaeridia is assigned to marine environments (AMSEG). The taxonomic position of Schizosporis found at baseof section F1 is still not clear; either it belongs to pollen or toZygnematacean algae.
Ichnofossils
Several unbranched, vertical, ovoid-shaped and unlined bur-rows (some cm length, 0.5 cm in diameter) are documented insandy and heterolithic layers (Fig. 3f). Concerning shape, size,orientation and substrate, this ichnofossil probably belongs tothe ichnogenus Thalassinoides. Decapod crustaceans arecommonly considered producers of such dwelling structuresin sandy to muddy soft ground of intertidal, lagoonal andshoreface environments (Pemberton and Ichnology ResearchGroup IRG 1998; Buatois and Mángano 2011). However,only vertical tubes have been observed that may haveprovided excess to branched horizontal mazes. In addition,Hauschke and Wilde (1987) mentioned small, paralleltrended, V-shaped imprints as limulid tracks.
Tetrapods
Plateosauridae
One sinistral femur of the prosauropod dinosaur Plateosauruslaevis (Quenstedt) was published by Fraas (1897) as‘Zanclodon laevis’ (Figs. 6e, f). The femur was found 1896
in the locality ‘Langenberge bei Hedeper’ (Fraas 1897).Jüngst (1928: p. 109) relocated it to the eastern part of theFuchsberg quarry. Due to damage during World War II, onlythe distal part is now preserved in the collections of theRoemer- and Pelizaeus-Museum Hildesheim Nr. NKP13466. The overall length of the femur is 51 cm (Fig. 6e, f).According to Sander (1992, 1999), Plateosaurids were thefirst herbivores feeding on plants (conifer branches andtwigs) at an elevation of more than 1 m above ground. InLate Triassic times, Plateosaurids have been widely dis-tributed and populated various terrestrial environments(Jaekel 1911; von Huene 1926; Galton 1984) (Fig. 12d).A few localities in Central and southern Germany provid-ed excellent complete and articulated individuals andgroups that have been embedded by catastrophicsheetfloods (Haubold 1989, Sander 1992, 1999).Disarticulated bones of Plateosaurus were transported byfluvial processes over longer distances (Weishampel 1984;Weishampel and Westphal 1986).
Amphibians
Besides the Plateosaurus femur, several fragments of unde-termined amphibian cephalic remains from the phosphoriticlayer at section F1 are preserved as imprints because theoriginal bone matter was mostly resolved during diagenesis(Fig. 6g, h). The amphibian remains were collected from theso-called phosphoritic horizon of the Fuchsberg quarry, seeHauschke and Wilde (1996: Fig. 4) and Fig. 2. This materialwas originally discovered by Dr. J. Solcher in June 2008.Because of the poor preservation, the taxonomic determina-tion and therefore the further characterisation of amphibianswithin the trophic structure of the Fuchsberg ecosystem is notpossible (Fig. 12e).
Plants
Plant remains (conifer shoots, cones, scales, stem and leaffragments) were found in the laminated silty to clayey strataas well as sandy layers. The occurrence of the plant debris isallochthonous and not bound on distinct layers. Most of theplant remains are disarticulated and badly preserved; conifer
�Fig. 7 Fossils from Fuchsberg and Langenberg, a Italophlebia baueri,fore wing, Late Norian, MFL, Langenberg, MLU.Sei2010.253a, scale 1cm, see also Fig. 12a b Polyphaga indet., dorsal side, Late Norian, MFL,Fuchsberg-west, MLU.Sei2010.028, scale 1 mm. c Voltziablattinidaeindet, pair of hind wings, Late Norian, MFL, Langenberg,MLU.Sei2010.242, scale 0.5 cm. d Dysmorphoptilidae indet., entirebody imprint, Late Norian, MFL, Langenberg, MLU.Sei.2010.271a,scale 0.5 cm. e Ipsvicia langenbergensis, holotype, Late Norian, MFL,Langenberg, MLU.Sei2010.182, scale 1 mm. f Ephemeroptera indet.,larva, Late Norian, MFL, Fuchsberg-west, MLU.Sei2010.012, scale 1mm
512 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

Palaeobio Palaeoenv (2014) 94:495–528 513
Author's personal copy

reproductive organs, leaves and other plant remains are rarelycollected.
Seinstedt has been known for its plant remains since thenineteenth century (e.g. Brauns 1862, 1866; Schenk 1867; seealso Table 5). Brauns (1862, 1866) described plant remainsfrom a 9-m (30-foot)-deep quarry northeast of Seinstedt, whathe considered the most important outcrop of this area andwhat is probably to equate with the Fuchsberg quarry.According to the author, the flora is composed ofsphenophytes (Calamites), ‘ferns’ (Taeniopteris (Fig. 9d and12b), Odontopteris, Laecopteris, Cyclopteris, Clathropteris,Camptopteris, Pecopteris, Cyatheites) as well as cycado-phytes (Nilssonia, Pterophyllum) and ‘Graminaceae’(Arundinites; Brauns 1862). Schenk (1867) and Schimper(1869, 1870–72, 1874) re-attributed some of the species ofBrauns (see Table 5). Jüngst (1928) enlisted from the north-east slope of Fallstein (near Fuchsberg) for the first timeginkgophyte (Ginkgo), conifer (Thuites) and seed fern(Ptilozamites, Lepidopteris) remains. Finally, Hauschke andWilde (1996) ment ioned for the f i rs t t ime thecheirolepidiaceous Hirmeriella muensteri.
New material was collected at Fuchsberg by one of theauthors (G.B.) and the figures in the historical publicationswere studied for a better understanding of the flora ofSeinstedt. The historical collection of Fuchsberg in Berlin(25 specimens) is dominated by Hirmeriella muensteri(Schenk) Jung 1968 shoots and male cones (Fig. 9a), whilethe newly collected material also shows badly preserved frag-ments of Clathropteris meniscoides Brongniart, 1828,sphenophytes stem fragments and Ctenozamites cf. wolfiana(Gothan) van Konijenburg–van Cittert et al. 1998 leaf remains(Fig. 9b, c). The material is mostly badly preserved, whencoalified material is preserved it seldom yielded cuticles. Thehistorical collection of Seinstedt stored at Berlin (about 120specimens) is preserved as impressions on coarse sandstone;in most cases venation and finer details are not preserved. Thecollection is dominated by narrow elongated leaves with anentire margin labelled Oleandrium tenuinerve Brauns, 1862(about 60 specimens; Fig. 9d) which probably belong toTaeniopteris typ orientalis Heer, 1878 orMarattia intermedia(Münster) Kilpper, 1964 known from the Rhaeto–Liassic ofIran, Afghanistan (Schweitzer et al. 1997, 2000) and Bayreuth(Weber 1968), and from the Early Jurassic of Nürnberg(Gothan 1914). Unfortunately, the venation pattern is not wellenough preserved for a specific determination. At least onefragment probably belongs to Anomozamites Schimper, 1870(Fig. 9e). Pterophyllum maximum Germar, 1879 is known tothe authors only from literature. Since the preservation is notgood enough for cuticle analyses, an attribution of this mate-rial to the bennettitalean genus Anomozamites as proposed bySchimper, (1870–72) is not possible. Actually, generalmacromorphology suggests rather an attribution to thecycadalean genus Nilssonia Bronn, 1835 (personal
communication and Van Konijnenburg-van Cittert, 2013).Nilssonia elongata Brongniart, 1828 and Pterophyllummuensteri Göppert, 1844 are mentioned in the literature, buthave not been found so far. Sphenophytes (about 20 speci-mens labelled partly as Arundinites priscus Brauns, 1862 andCalamites guembeli Schenk, 1867) are preserved as impres-sions of the stems with vascular bundles. Similar fragmentshave been described as Neocalamites lehmannianus(Göppert) Weber, 1968 from the Rhaeto-Liassic of Bayreuth(Weber 1968). Other material figured in the literature belongsto Neocalamites hoerensis (Schimper) Halle, 1907, while noEquisetites muensteri Sternberg, 1828, mentioned by Jüngst(1928), has been found in the collections. The ferns are rare(about 10 specimens) with frond remains of Dictyophyllumexile (Brauns) Nathorst, 1878 (Fig. 9f) and Phlebopterisangustiloba (Presl in Sternberg) Hirmer and Hörhammer,1934. The frond fragments labelled and figured asCyclopteris crenata Brauns, 1862 do not show any venation;the shape and the leathery texture suggest that they mayrepresent aphlebia of ferns. The seed ferns are poorly and veryfragmentarily preserved and rare (8 specimens). Nonetheless,four different taxa have been distinguished (Ctenozamites cf.wolfiana (Gothan) van Konijenburg–van Cittert et al. 1998,Lepidopteris ottonis (Göppert) Schimper, 1869, Ptilozamitesblasii (Brauns) Nathorst, 1879, Ptilozamites heeri Nathorst,1878; Fig. 9g), well known from the Rhaetian of Europe (e.g.Harris 1932, 1937; Kelber and van Konijnenburg–van Cittert1997; van Konijenburg–van Cittert et al. 1998; Kustatscherand van Konijenburg–van Cittert 2007; Zavialova and vanKonijnenburg–van Cittert 2011). The bonebed of Seinstedtalso yielded conifer remains labelled as Thuites schloenbachiSchenk, 1867 which belong to Hirmeriella muensteri(Fig. 9a), a typical species of the Rhaetian–Liassic ofGermany and Poland (e.g. Jung 1968; Weber 1968;Barbacka et al. 2007 and references therein). The plant re-mains from Seinstedt have no biostratigraphic value as mostof them have so far been recorded from Rhaetian–Liassicsediments or even have a more extensive stratigraphic record.The general composition with a high abundance of cycado-phytes, common ferns and horsetail remains suggest upperdelta plain or fluvial floodplain environments as habitats. Asthe successions at Fuchsberg and Langenberg are
�Fig. 8 Fossils from Fuchsberg, a Semionotus-like actinopterygian, LateNorian, MFL, Fuchsberg-west, MLU.Sei2010.076. b Malacostracaindet., Late Norian, MFL, Fuchsberg-west, MLU.Sei2010.092. cPaleolimulus fuchsbergensis, holotype, Late Norian, MFL, Fuchsberg-west, SMF VIII 311, picture from Hauschke and Wilde (1987). dConchostracans: Shipingia gerbachmanni (bottom left) and twoGregoriusella polonica (top centre and right) on one bedding plane,latest Sevatian to earliest Rhaetian, between MFL and ‘GrüneGrenzhorizont’, MLU.Sei2010.118. e Coelacanthid scale, Late Norian,MFL, Fuchsberg-west, MLU.Sei2010.078. All scales 1 cm, except b 0.5cm
514 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy
Palaeobio Palaeoenv (2014) 94:495–528 515
Author's personal copy

generally rich in plant remains (mostly debris) repeatedsheet flooding or crevassing is concluded for upstreamlowland environments. Shoots, leaves and reproductiveorgans of the land plants were introduced via distributarychannels and reworked during transport. The conifers areso far represented only by the cheirolepidaceaeHirmeriella. Since a near-sea environment has been pos-tulated (e.g. Weber 1968), Hirmeriella probably was partof lower delta plain plant communities that vegetatedmargins of interdistributary bays (Fig. 12c).
Indications for plant-feeding by insects were not recorded.
Spores and pollen
More than 30 samples for palynological analyses were takenat Fuchsberg and Langenberg (Table 6). The samples aredominated by Coroll ina (Classopoll is) spp. andGranuloperculatipollis rudis; spores are represented by fewe l emen t s ( e . g . De l t o i d o s po ra , Ca l amospo ra ,Punctatisporites). Typical elements of the Keuper such asEnzonalasporites, Vallasporites and Ovalipollis are well rep-resented. Ricciisporites tuberculatus is rarer then Ovalipollisspp. Notable is the presence of Spiritisporites spirabilis pre-viously found together with Granuloperculatipolles rudis inthe upper Gipskeuper (Mainhardt Formation), and lowerSteinmergelkeuper (Heunisch and Nitsch 2011). This couldindicate a slightly younger age for the lower samples. Thesections can generally be attributed to an Early Rhaetian age(also due to the presence of Ricciisporites tuberculatus), cor-responding to the Corollina-Enzonalasporites zone of Lund(1977) and Granuloperculatipollis subzone followingHeunisch (1999). The palynofacies is composed of sporesand pollen from land plants and comprises abundant Inertite.Both originates from vegetated environments of the upperdelta plain or fluvial floodplains.
Depositional environment
Facies associations and depositional architecture
According to lithofacies types and their stacking pattern,sedimentary architecture and fossil content, a fluvial-dominated delta plain that was dissected by distributary chan-nels is reconstructed. Laterally distributary channels wereassociated to mouth bars and interdistributary bays.
Lower delta plain wetlands
The exposures at Fuchsberg and Langenberg comprise a sig-nificant portion of heterolithic and horizontal laminatedlithofacies forming tabular and laterally persistent sediment
bodies. In an overall aggradational architecture these tabularbodies are intercalated with those of mouth bars, distributarychannels and interdistributary bays but lack internal trendsthat can be attributed to sedimentary cyclicity (Fig. 10). Theheterolithic and horizontal laminated lithofacies suggest de-position of suspended load and modification by wave actionand unidirectional flows (Reineck 1963; Reineck and Singh1980). Further evidence of frequent modification under oscil-latory conditions is given by wave ripples and hummockycross-stratification (de Raaf et al. 1977; Arnott and Southard1990; Dumas and Arnott 2006). The occasional occurrence ofconvolute bedding results from loading of soft and unconsol-idated fine clastics due to at least temporally higher sedimen-tation rates. As indications of at least temporal subaerialexposure like mud cracks, incipient soils and rooted horizonsare lacking, the permanent exposure to subaquatic conditionsis concluded. The described autochthonous aquatic biota(mostly conchostracans) tolerate either freshwater or brackishto freshwater conditions (Retallack and Clifford 1980; Storchand Welsch 2004; Lecointre and Le Guyader 2006). In par-ticular, the recorded phytoplankton forms Botryococcus,Tasmanites and Leiosphaeridia provide more reliable argu-ments for a brackish influence. Accordingly, the distal part ofa lower delta plain with wetlands that where permanentlyexposed to subaquatic conditions is reconstructed (Fig. 12).
The delta plain forms the flat-lying topset strata of a delta(Gilbert 1885) and is defined by the presence of distributarychannels (Bhattacharya 2006). Its landward limit is commonlydrawn at the point where a trunk streams become unconfinedand distributive; seawards, it is limited by the shoreline of thedelta front (Elliott 1986). The landward limit of marine incur-sions enable the subdivision into upper and lower delta plain(Coleman and Prior 1982). Both the upper and larger parts of thelower delta plain are subaerial exposed and vegetated and onlythe distal part of the lower delta plain is permanently exposed tosubaquatic conditions (Coleman and Prior 1982; Elliott 1986).
The abundant plant remains as well as spores and pollenrecorded at Fuchsberg and Langenberg give considerableinsights into upstream environments. Cycadophytes, commonferns and horsetail remains are considered part of a persistentflora that vegetated floodplains and lowlands. These plants (aswell as the Plateosaurus femur) were washed away duringsheet flows and deposited in the lower delta plain throughdistributary channels. The cheirolepidaceae Hirmeriella pre-ferred moist habitats of the upper and proximal lower delta
�Fig. 9 Macroplant remains from Seinstedt area, aHirmeriella muensteri,MB.PB.2013_1055. b Clathropteris meniscoides, MB.PB.2013_1637. cCtenozamites, MB.PB.2013_1078. d Taeniopteris orientalis,MB.PB.2013_1656. e Anomozamites, MB.PB.2013_1571. Note thefeeding traces. f Dictyophyllum exile, MB.PB.2013_1544. gPtilozamites heeri, MB.PB.2013_1579. All scales 1 cm
516 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

Palaeobio Palaeoenv (2014) 94:495–528 517
Author's personal copy

plain (Weber 1968) and, probably, vegetated levees ofdistributaries.
Distributary channels
At Fuchsberg, two more or less symmetric channels of up to100 m width and up to 3 m thickness are exposed (Fig. 12).The channels have convex downward bases and flat tops, andare incised into tabular sedimentary bodies mainly formed ofripple cross-bedded lithofacies. The fill is composed of finingupward cycles that comprise stacked sets of trough cross-bedded lithofacies (Fig. 10b). In terms of grain sizes, thesesets are noticeable coarser compared to strata below and above(Fig. 5). At the top, the channel fill is composed of ripplecross-bedded lithofacies and often superimposed by hum-mocky cross-stratification (Fig. 10). The simple fill of chan-nels appears single-storied as subordinated inclined boundingsurfaces that would indicate lateral shifting has not beenobserved. The channels were lateral only confined by incision;strata that can be attributed to natural levees were not ob-served. At Langenberg, a comparable channel fill is partlyexposed (Fig. 10a).
The simple symmetric and single-storied architecture ofchannels and their fining upward fill suggest straight channelsthat were incised during stages of higher transport velocitiesand later subsequently filled during stages of lowered trans-port velocities. Reactivation or erosion within the channel fillshave not been observed. Accordingly, only the lower parts ofthe channel fills comprises coarse grained trough cross-bedded lithofacies, whereas the upper parts are either modifiedby wave action (HCS) or formed of finer grained ripple cross-
bedded lithofacies (Fig. 10b). This indicates that the respec-tive channel was only active for a shorter interval of time andlater became inactive due to lateral shift or avulsion.
Channels of comparable architecture have been describedfrom the straight distal ends of distributaries of high-constructive fluvial-dominated delta systems (Fisher et al.1972; Tye and Coleman 1989; Reading 1996). Such sys-tems are, by definition, progradational and characterised bybifurcation of distributary channels at their distal ends aswell as avulsion of delta lobes (Fisher et al. 1972).Bifurcation of a main distributary into two or more minordistributaries is a common feature at the lower delta plainwhere the main channel is not longer confined by subaque-ous levees (Russel 1967, Fisher et al. 1972). Such a bifur-cation is possibly exposed in the lower parts at Fuchsberg/Langenberg, as the lower channel at Fuchsberg is time-constrained to the channel at Langenberg by means ofconchostracan biostratigraphy and the corresponding orderof strata. Lateral distributary shift or avulsion of a delta lobeis indicated by laminated interdistributary bay lithologiesthat follows distributary channel fills at Fuchsberg andLangenberg (Figs. 2, 10a).
Mouth bars
Tabular sedimentary bodies of up to more than 100 m lateralextent and up to 4 m thickness form prominent sandstones atFuchsberg/Langenberg (Fig. 3). On a larger scale, these sand-stones show aggradational architecture and interfinger withdelta plain and interdistributary bay lithologies. On a smallerscale, they are composed of up to 2-m-thick coarsening
Table 6 Listing of all known palynomorph data in section Langenberg and Fuchsberg
Column ‘outcrop’: L Langenberg, F Fuchsberg
518 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

upward depositional cycles (Fig. 10). The vertical shift fromhorizontal-laminated siltstone-sandstone to ripple cross-bedded sandstone lithofacies results from progradational pat-tern and increasing importance of bed load transport (Fig. 4).At their top, hummocky cross-stratification indicates modifi-cation by wave action (Figs. 3, 10).
Similar upward-coarsening successions have been com-monly described from progradational mouth bars of moderndeltas (Scruton 1960; Coleman and Wright 1975; Elliott1986). At the mouth of a distributary channel, transport ve-locity decreases and transported load forms a larger mouth bar(Fisk et al. 1954; Fisher et al. 1972). Due to wave action,mouth bars are smoothed and elongated parallel to the shore-line (Wright 1977; Fielding et al. 2005) and form sand bodiesof up to several kilometres long and wide (Reynolds 1999).Vertical successions of lateral associated distributary channelsand mouth bars form if the delta progrades basinwards and the
channel overrides and incises into precursor mouth bar de-posits (Bhattacharya and Walker 1992). At Fuchsberg, suchvertical successions of precursor mouth bars and incised dis-tributary channels form ‘complexes’. In the lower part, twolarger and proximal mouth bar/channel complexes are attrib-uted to successive progradation of the delta, whereas thesmaller and distal mouth bar/channel complex in the upperpart is rather attributed to the interdistributary bay fill(Fig. 10).
Interdistributary bay
Both the exposures at Fuchsberg and Langenberg comprise anup to 1-m-thick succession of laminated claystone-siltstonelithofacies that can be correlated due to conchostracan bio-stratigraphy and the corresponding order of strata (Fig. 2). Bymeans of this, an at least several hundred meters wide and 1-
distal mouth bars
distributaries mouth bars lower deltaic plaininterdistributary bay fill
20 0 m
0
4 m
proximal channel/mouth bar complex
60 cm
400 cm
Mouth Bar
Distributary channel
Heterolithics, HCSerosive base
clay
coar
se s
and
med
ium
san
dfin
e sa
ndsi
lt
grain size
F8
F9 F10 F11F12
NorianRhaetian
storm eventlayers (tempestite)
b
b proximalmouth bars
distal channel/mouth bar complex
a
Mouth BarHeterolithics, HCS
Distributary channelerosive base
20 cm150 cm
0 20 m0
2 m
F3F2
F4
F5F7
F6
n = 13mean = 183°
N
90270
c Through cross-bedded setsd
c
Palaeobio Palaeoenv (2014) 94:495–528 519
Fig. 10 Facies architecture at Fuchsberg and Langenberg. For key seeFig. 2. a Overview, note bifurcation of two time-constrained distributarychannels at Langenberg and Fuchsberg. The storm event layer partlycontains the bonebed I. b c Detailed sketches of proximal and distal
mouth bar/channel complexes exposed at Fuchsberg with sections F8–F12 and F2–F7. d Transport directions of distributary channel fills atFuchsberg
Author's personal copy

m-thick tabular sediment body can be reconstructed thataggrades onto a mouth bars/distributary channel complex(Figs. 10, 12). The horizontal-laminated fine grainedlithofacies point to suspended load as dominant mode ofdeposition and 1- to 2-cm-thick micro-scale coarsening up-wards cycles suggest slight variations in discharge(Bhattacharya 2006). Besides the ‘varve-like’ lithofacies, theabundant aquatic biota recorded in the MFL point to a largeraquatic ecosystem at the lower delta plain that received onlylimited detrital input. In particular, the phytoplankton forms(Schizosporis, Botryococcus, Leiosphaeridia) suggest at leasta temporal brackish influence and the overall trend to-wards silty to sandy lithologies reflects a successiveshallowing (Walker and Harms 1971; Bhattacharya andWalker 1991). All these characteristics are described fromrecent analogues of interdistributary bays (Coleman et al.1964; Donaldson et al. 1970; Elliott 1974; Tye andColeman 1989).
Interdistributary bays are sub-environments of deltaplains and form shallow water bodies or swamps betweenthe downstream end of distributary channels and theirassociated mouth bars. These bays are commonly open tothe sea but may also be (partially) enclosed and conditionsrange from saline to nearly fresh (Coleman et al. 1964;Gould 1970; Elliott 1974; Tye and Kosters 1986). In gen-eral, the fills of interdistributary bays are dominated bylaminated light to dark grey fine clastic lithologies (clays,silts, fine sands) introduced due to overbank spilling orcrevassing (Elliott 1974; Tye and Coleman 1989).
At Fuchsberg/Langenberg, the interdistributary bayevolved on top of a mouth bar/distributary channel complexand was terminated by a mouth bar/distributary channel com-plex its development was probably linked to avulsion ofdistributary channels (Mackey and Bridge 1995; Holbrook1996).
Geochemistry
At Fuchsberg, gamma ray readings are increased toabout 150 cps within a distinct horizon 120–150 cmabove the MFL (Fig. 11b, black curve). This horizoncan be traced around Seinstedt and was prospected foruranium and thorium in the 1960s. According to Benda(1963), these elements are concentrated in phosphoriteconcretions as phosphatic complexes or minerals(apatite) with a maximum uranium content of 775ppm. This can be confirmed by the covalent trend ofP2O5 and the U/Th ratio (Fig. 11b); however, new datasuggest only strong increased thorium values whereasuranium values are only slightly increased. Accordingly,nickel and cobalt values are increased in samples withhigher thorium and uranium values (Table 7). FollowingPaul and Siggelkow (2004), oxidized uranium of erodedrocks was mobilized as uranyl-ion and adsorbed on clayminerals . Entering subaquatic environments atFuchsberg, this complex was reduced by sulfide-richwater and fixed in sediment as hardly dissolveableUO2. In this way, the lower delta plain was considered
Langenberg – L1
Fuchsberg – F10
1
2
3
4
5
6
7
8
9
1 0
0 1 2 3 4 5 6 7
Redox: Ni/Co vs. U/Th
Ni/C
o
U/Th
Langenberg
Fuchsberg
dyso
xic
anoxicoxic d
anox
icox
icb
0 6 8 100
1
2
4
3
5
6
7
0
1
2
4
3
5
6
7
0
1
2
4
3
5
6
7
0
1
2
4
3
5
6
7
19
a
Ni/Co2 4 0 1 2 3
U/Th
MFL
oxic dys anoxic oxic d anoxic
0Ni/Co
0 2 4 6 8 1 0 1 2 3U/Th
oxic d anoxic
oxic dys anoxic
height[m]
height[m]
MFL
5.318.4
P2O5
GR
50 100 150 cps
1 2 3 4 %
18
P2O5
520 Palaeobio Palaeoenv (2014) 94:495–528
Fig. 11 a Cross-plot of palaeoredox indices U/Th versus Ni/Co (Jonesand Manning 1994). b Palaeoredox indices U/Th and Ni/Co measured atLangenberg (left) and Fuchsberg (right) point to spatial and temporal
restricted dysoxic or anoxic conditions only; note: base of section F1 doesnot correspond to base of section L1. For data, see Table 7
Author's personal copy
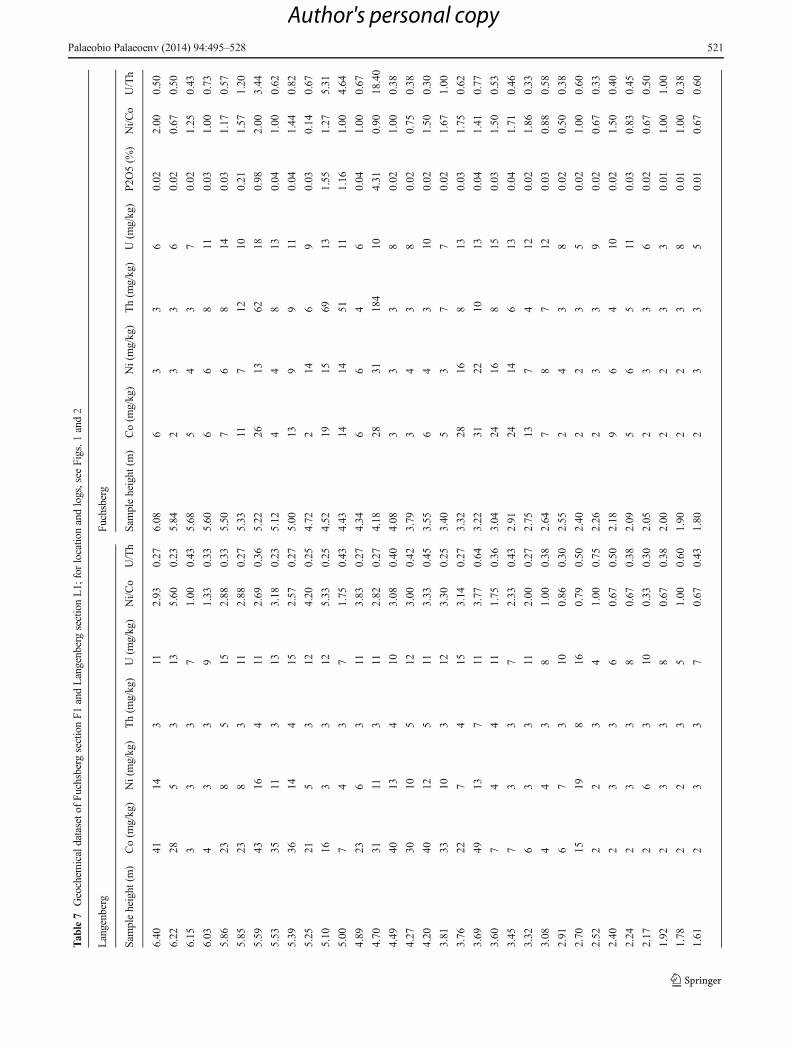
Tab
le7
Geochem
icaldataseto
fFuchsberg
sectionF1andLangenbergsectionL1;
forlocatio
nandlogs,see
Figs.1
and2
Langenberg
Fuchsberg
Sampleheight
(m)
Co(m
g/kg)
Ni(mg/kg)
Th(m
g/kg)
U(m
g/kg)
Ni/C
oU/Th
Sam
pleheight
(m)
Co(m
g/kg)
Ni(mg/kg)
Th(m
g/kg)
U(m
g/kg)
P2O5(%
)Ni/C
oU/Th
6.40
4114
311
2.93
0.27
6.08
63
36
0.02
2.00
0.50
6.22
285
313
5.60
0.23
5.84
23
36
0.02
0.67
0.50
6.15
33
37
1.00
0.43
5.68
54
37
0.02
1.25
0.43
6.03
43
39
1.33
0.33
5.60
66
811
0.03
1.00
0.73
5.86
238
515
2.88
0.33
5.50
76
814
0.03
1.17
0.57
5.85
238
311
2.88
0.27
5.33
117
1210
0.21
1.57
1.20
5.59
4316
411
2.69
0.36
5.22
2613
6218
0.98
2.00
3.44
5.53
3511
313
3.18
0.23
5.12
44
813
0.04
1.00
0.62
5.39
3614
415
2.57
0.27
5.00
139
911
0.04
1.44
0.82
5.25
215
312
4.20
0.25
4.72
214
69
0.03
0.14
0.67
5.10
163
312
5.33
0.25
4.52
1915
6913
1.55
1.27
5.31
5.00
74
37
1.75
0.43
4.43
1414
5111
1.16
1.00
4.64
4.89
236
311
3.83
0.27
4.34
66
46
0.04
1.00
0.67
4.70
3111
311
2.82
0.27
4.18
2831
184
104.31
0.90
18.40
4.49
4013
410
3.08
0.40
4.08
33
38
0.02
1.00
0.38
4.27
3010
512
3.00
0.42
3.79
34
38
0.02
0.75
0.38
4.20
4012
511
3.33
0.45
3.55
64
310
0.02
1.50
0.30
3.81
3310
312
3.30
0.25
3.40
53
77
0.02
1.67
1.00
3.76
227
415
3.14
0.27
3.32
2816
813
0.03
1.75
0.62
3.69
4913
711
3.77
0.64
3.22
3122
1013
0.04
1.41
0.77
3.60
74
411
1.75
0.36
3.04
2416
815
0.03
1.50
0.53
3.45
73
37
2.33
0.43
2.91
2414
613
0.04
1.71
0.46
3.32
63
311
2.00
0.27
2.75
137
412
0.02
1.86
0.33
3.08
44
38
1.00
0.38
2.64
78
712
0.03
0.88
0.58
2.91
67
310
0.86
0.30
2.55
24
38
0.02
0.50
0.38
2.70
1519
816
0.79
0.50
2.40
22
35
0.02
1.00
0.60
2.52
22
34
1.00
0.75
2.26
23
39
0.02
0.67
0.33
2.40
23
36
0.67
0.50
2.18
96
410
0.02
1.50
0.40
2.24
23
38
0.67
0.38
2.09
56
511
0.03
0.83
0.45
2.17
26
310
0.33
0.30
2.05
23
36
0.02
0.67
0.50
1.92
23
38
0.67
0.38
2.00
22
33
0.01
1.00
1.00
1.78
22
35
1.00
0.60
1.90
22
38
0.01
1.00
0.38
1.61
23
37
0.67
0.43
1.80
23
35
0.01
0.67
0.60
Palaeobio Palaeoenv (2014) 94:495–528 521
Author's personal copy

Tab
le7
(contin
ued)
Langenberg
Fuchsberg
Sampleheight
(m)
Co(m
g/kg)
Ni(mg/kg)
Th(m
g/kg)
U(m
g/kg)
Ni/C
oU/Th
Sampleheight
(m)
Co(m
g/kg)
Ni(mg/kg)
Th(m
g/kg)
U(m
g/kg)
P2O5(%
)Ni/C
oU/Th
1.40
23
38
0.67
0.38
1.70
113
514
0.03
3.67
0.36
1.32
23
34
0.67
0.75
1.66
23
36
0.02
0.67
0.50
1.22
23
34
0.67
0.75
1.58
22
33
0.02
1.00
1.00
1.10
23
35
0.67
0.60
1.47
23
35
0.01
0.67
0.60
1.02
23
35
0.67
0.60
1.41
63
39
0.04
2.00
0.33
0.91
23
37
0.67
0.43
1.30
324
811
0.05
8.00
0.73
0.82
23
34
0.67
0.75
1.22
285
2018
0.09
5.60
1.11
0.75
23
36
0.67
0.50
1.12
42
34
0.02
2.00
0.75
0.61
22
34
1.00
0.75
1.00
23
36
0.02
0.67
0.50
0.46
22
37
1.00
0.43
0.89
23
34
0.02
0.67
0.75
0.33
23
36
0.67
0.50
0.78
33
35
0.02
1.00
0.60
0.19
23
38
0.67
0.38
0.72
33
35
0.02
1.00
0.60
0.03
23
39
0.67
0.33
0.61
23
34
0.02
0.67
0.75
0.49
74
39
0.03
1.75
0.33
0.39
23
39
0.02
0.67
0.33
0.30
23
35
0.02
0.67
0.60
0.22
23
35
0.01
0.67
0.60
522 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

a trap for uranium and other elements like copper, zincor lead precipitated as sulfides (Paul and Siggelkow2004). Lateral variations in redox conditions are indi-cated as the corresponding horizon at Langenberg isonly slightly enriched in nickel and cobalt but notthorium and uranium (Fig. 11b). Palaeoredox indicesU/Th and Ni /Co measured a t Fuchsberg andLangenberg successions suggest deposition under
general oxic conditions (Jones and Manning 1994).Temporal dysoxic or anoxic conditions were only local-ly restricted (Fig. 11a, b) and may be responsible forthe occurrence of well-preserved fishes in an individuallayer of the MFL (Hauschke and Wilde 1996).However, persistent dysoxic or anoxic conditions ofinterdistributary bay environments (MFL) are not indi-cated, based on these indices.
Fig. 12 Reconstruction of deltaic environments and habitats of theSeinstedt area, all drawings by Frederik Spindler (Freiberg, Germany).a Italophlebia baueri, body length around 7 cm. b Taeniopteris sp., heightaround 30 cm. c conifer and seedfern forest, height of trees around 20 m.d Plateosaurus sp. cadaver, length of femur 0.5 m, see Fig. 6e, f;
Plateosaurus body height not to scale at position ‘d’ in the environmentalreconstruction. e Mastodonsaurus sp., length around 1 m. fcoelacanthiform fish, length around 1.5 m. g Paleolimulusfuchsbergensis, length around 3 cm. h Synechodus sp. (length around0.5 m) and single tooth (height=around 1 mm, see Fig. 6b)
Palaeobio Palaeoenv (2014) 94:495–528 523
Author's personal copy

Delta plain and floodplains: a brief synopsis of habitats
Based on litho- and biofacies analyses, the delta plain of afluvial-dominated delta type is reconstructed. The recordedfauna and flora enable the recognition of an autochthonouslower delta plain exposed at Fuchsberg and Langenberg andallochthonous upper delta plain and floodplain environmentsexposed upstream (Fig. 12).
The lower delta plain includes subenvironments of (1) sub-aqueous wetlands, (2) mouth bar/distributary channel com-plexes and (3) interdistributary bays. In particular, theautochthonous ecosystem of the interdistributary bay canbe characterised by successive trophic levels that formedfood chains. Phytoplankton and plant remnants are con-sidered primary producers that fed primary consumerslike conchostracans, malacostracans, insect larvae andjuvenile semionotid and coelacanthid fishes. Adult fishes,limulids and small sharks (teeth remains found inbonebed I and II) are considered secondary consumersthat fed on primary consumers. Possible tertiary con-sumers and/or top predators are represented by amphib-ians and larger sharks. In addition, malacostracans andlimulids acted as decomposers.
Most of the groups that inhabited the lower delta plain andthe interdistributary bay are thought to tolerate larger fluctua-tions of salinity (bivalves, coelacanthids, sharks, limulids).However, in particular, the prasinophycacean generaLeiosphaeridia and the bivalve genera Gervillia andRhaetavicula provide a robust argument for a marine influ-ence and therefore a brackish salinity is reconstructed.Oxygenation of water is suggested by epibentic fauna likecrustaceans, insect larvae and geochemistry (palaeoredox in-dices: U/Th, Ni/Co).
Due to excellent taphonomical conditions within theinterdistributary bay, at least some trophic levels ofparautochthonous riparian ecosystems of the lower deltaplain can be reconstructed. Plants that populated the banksof the interdistributary bay like the cheirolepidacean coni-fer Hirmeriella represent primary producers that fed prima-ry consumers like herbivore dysmorphoptilids and grass-hoppers as well as omnivore cockroaches. Carnivore drag-onflies (Figs. 7a, 12a) and beetles as well as omnivorecockroaches are considered secondary consumers.However, further trophic levels of the riparian ecosystemwere not recorded.
Floral and faunal remnants of upper delta plain andfloodplain environments were introduced via fluvial and dis-tributary channels. These upstream environments were vege-tated with a flora composed of abundant cycadophytes, fernsand horsetails. The Plateosaurus femur recorded at Fuchsbergindicate that large herbivores fed on this rich vegetation(Fig. 12d). The vegetated upper delta plain and floodplainenvironments are considered basic trophic level of a number
of terrestrial ecosystems that are, due to taphonomical bias,nor preserved at Fuchsberg and Langenberg.
Conclusions
1. The comprehensive revision of both exposures revealeda well-documented ecosystem of a fluvial-dominateddelta plain. Delta formation was controlled byprogradation, bifurcation of distributary channels andavulsion of delta lobes.
2. The exposures at Fuchsberg and Langenberg areconstrained to the Norian/Rhaetian boundary interval bymeans of conchostracan and palynomorph biostratigra-phy. The transition from the polonica/gerbachmanni zoneto the polonica zone and from the GTr 17 zone to the GTr18 zone is well documented at Fuchsberg (Fig. 2). Bothexposures can be reliably correlated by means ofconchostracan biostratigraphy and the corresponding or-der of strata.
3. At Fuchsberg and Langenberg, the distal lower deltaplain is exposed with brackish subaqueous deltaplain wetlands, mouthbar/distributary channel com-plexes and interdistributary bay subenvironments.
4. A diverse fauna is documented from the specificsubenvironments: limulids (1 taxon), malacostracansand conchostracans (several taxa) from the lowerdelatic plain, amphibians (at least 1 taxon), sharks(several taxa) and osteichthyan fishes (at least 6taxa) from the interdistributary bay and at least 20taxa of 9 insect orders and plateosaurids from ripar-ian or upstream environments. The trophic systemsof autochthonous subaqueous and parautochthonousriparian habitats are well preserved within fossilif-erous interdistributary bay lithologies.
5. The reconstruction of vegetated upstream environ-ments at the upper delta plain and floodplain isenabled by abundant remnants of cycadophytes,ferns, horsetails and large vertebrates.
6. The marine-influenced lower delta plain atFuchsberg/Langenberg, documented by, e.g. phyto-plankton and sharks, confirms an early transgressionof the ‘Rhaetian Sea’ as far east as SE LowerSaxony.
Acknowledgements We acknowledge field assistance of K.Rauppach (Neubrandenburg), J. Solcher (Egestorf) and J.Zimmermann (Freiberg), and K. Bauer (Bozen) for photographingthe macroflora from Seinstedt. G.B. thanks N. Hauschke fromGeological and Palaeontological Collections, Martin-Luther-University Halle-Wittenberg for making the Fuchsberg andLangenberg invertebrate material available, and E.K. thanks C.Hiller, M. Ruhl and B. Mohr from Museum für Naturkunde–
524 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

Leibnitz Institute for Research on Evolution and Biodiversity atHumboldt University Berlin for making the Seinstedt material avail-able. This study was funded by ‘Bundesministerium für Umwelt,Naturschutz und Reaktorsicherheit’ (FKZ 0325285), the project‘The biodiversity of the Late Permian floras in the GermanZechstein Basin’ (DE-TAF-3195) and supported by SYNTHESYS(EC Research Infrastructure Action, Program FP7 ‘Structuring theEuropean Research Area’). E.K. thanks J.H.A. Van Konijnenburg-Van Cittert (University of Leiden and Utrecht) for sharing her expe-rience on the Late Triassic floras of Germany and acknowledgesfinancial support from the Alexander von Humboldt-Foundation(3.3-ITA/1141759STP). D.T. and J.V. thank J. Mutterlose (Bochum)for preparation of SEM pictures of fish teeth. We acknowledge J.Ansorge (Horst) for discussions, F. Spindler (Freiberg, Germany) fordrawing the reconstructions and C. Brauckmann (Clausthal-Zellerfeld) and N. Hauschke (Halle/Saale) for careful reviews.
References
Abbink OA (1998) Palynological identification in the Jurassic of theNorth Sea region. LPP Contrib Ser 8:1
Abbink OA, van Konijnenburg-van Cittert JHA, Visscher H (2004a) Asporomorph ecogroup model for the northwest European Jurassic-Lower Cretaceous I: Concepts and framework. Netherl J Geosc 83:17–31
Abbink OA, van Konijnenburg-van Cittert JHA, van der Zwan CJ,Visscher H (2004b) A sporomorph ecogroup model for the north-west European Jurassic-Lower Cretaceous II. Application to anexploration well from the Dutch North Sea. Netherl J Geosc 83:81–91
Aigner T (1985) Storm depositional systems: dynamic stratigraphy inmodern and ancient shallow-marine sequences. Lecture notes inearth sciences, vol 3. Springer, Berlin
Aigner T, Reineck H-E (1982) Proximality trends in modern storm sandsfrom the Helgoland Bight (North Sea) and their implications forbasin analysis. Senck marit 14:183–215
Aigner T, Reineck H-E (1983) Seasonal variation of wave base on theshoreface of the barrier island Norderney, North Sea. Senck marit15:87–92
Allen JRL (1982) Sedimentary structures: their character and physicalbasis. Elsevier, Amsterdam
Allen JRL, Collinson J (1974) The superposition and classification ofdunes formed by unidirectional flow. Sed Geol 12:169–178
Appel D (1981) Petrographie und Genese der Sandsteine des Unter- undMittelräts im nördlichen Harzvorland (Ostniedersachsen). Mitt GeolInst TU Hannover 20:1–133
Arnott RW, Southard JB (1990) Exploratory flow-duct experiments oncombined-flow bed configurations, and some implications forinterpreting storm-event stratification. J Sed Petrol 60:211–219
Ashley GM (1990) Classification of large-scale subaqueous bedforms: anew look at an old problem. J Sed Petrol 60:160–172
Bachmann GH, Grosse S (1989) Struktur und Entstehung des Nord-deutschen Beckens – geologische und geophysikalische Inter-pretation einer verbesserten Bouguer-Schwerekarte. Nieders AkadGeow Veröffentl 2:23–47
Bachmann GH, Geluk M, Warrington J, Becker A, Beutler G,Hagdorn H, Hounslow M, Nitsch E, Röhling H-G, Simon T,Szulc A, Dusar M, Nielsen LH, Barnasch J, Franz M (2010)Triassic. In: Doornenbal H, Stevenson A (eds) PetroleumGeological Atlas of the Southern Permian Basin Area. EAGE,Houton, pp 149–173
Barbacka M, Ziaja J, Wcislo-Luraniec E, Reymanowna M (2007)Hirmerella muensteri (Schenk) Jung from Odrowąż (Poland), with
female and male cones, and in situ Classopollis pollen grains. ActaPalaeobot 47:339–357
Barth G (2011) Das Ober-Norium von Seinstedt (Niedersachsen) –Sedimentologie, Geochemie und Arthropodenfunde (Insekten,Conchostraken). Diploma thesis, Martin-Luther-Universität Halle-Wittenberg
Barth G, Kozur HW (2011) A Latest Norian age for insect-bearing bedsof the Fuchsberg and Langenberg near Seinstedt, northern forelandof the Harz Mountains (Lower Saxony, Germany). In: Sullivan R,Lucas S, Spielmann J (eds) Fossil Record 3. New Mexico Museumof Natural History and Science, Albuquerque, pp 157–165
Barth G, Ansorge J, Brauckmann C (2011) First record of the genusIpsvicia (Hemiptera: Ipsviciidae) outside Gondwana – AnAustralian congener from the Upper Triassic of Germany. Pol JEntom 80:645–657
Barth G, Nel A, FranzM (2013) Two new odonate-like insect wings fromthe latest Norian of northern Germany. Pol J Entom 82:127–142
Behrend F (1927) Erläuterungen zur Geologischen Karte von Preußen,Blatt Hornburg und Blatt Hessen. Lfg. 245, Berlin
Benda L (1960) Bericht über die Anreicherung von Uran in PhosphoritenNiedersachsens, unter besonderer Berücksichtigung des Phos-phorites von Seinstedt. MTB Hornburg, vol 3929
Benda L (1963) Über die Anreicherung von Uran und Thorium inPhosphoriten und Bonebeds des nördlichen Harzvorlandes. GeolJb 80:313–348
Benek R, Kramer W, McCann T, Scheck M, Negendank JFW, Korich D,Huebscher H-D, Bayer U (1996) Permo-Carboniferous magmatismof the Northeast German Basin. Tectonophysics 266:379–404
Bhattacharya JP (2006) Deltas. In: Posamentier HW, Walker RG (eds)Facies models revisited. Special publication 84. Society forSedimentary Geology, Tulsa, pp 237–292
Bhattacharya JP, Walker RG (1991) Facies and facies successions inriver- and wave-dominated depositional systems of the UpperCretaceous Dunvegan Formation, northwestern Alberta. B CanPetrol Geol 39:165–191
Bhattacharya JP, Walker RG (1992) Deltas. In: Walker RS, James NP(eds) Facies models: response to sea-level change. GeologicalAssociation of Canada, pp 157–177
Brauns D (1862) Der Sandstein bei Seinstedt unweit des Fallsteins unddie in ihm vorkommenden Pflanzenreste. Palaeontographica 9:47–62
Brauns D (1866) Der Sandstein bei Seinstedt unweit des Fallsteins unddie in ihm vorkommenden Pflanzenreste, nebst Bemerkungen überdie Sandsteine gleichen Niveaus anderer Örtlichkeiten Nord-deutschlands. Palaeontographica 13:237–246
Buatois LA, Mángano MG (2011) Ichnology: Organism–substrate inter-actions in space and time. Cambridge University Press, Cambridge
Caroll RL (1997) Paläontologie und Evolution der Wirbeltiere. Thieme,Stuttgart
Coleman JM, Prior DB (1982) Deltaic Environments. In: Scholle PA,Spearing D (eds) Sandstone depositional environments. TheAmerican Association of Petroleum Geologists, Tulsa, pp 139–178
Coleman JM, Wright LD (1975) Modern river deltas: variability ofprocesses and sand bodies. In: Broussard ML (ed) Deltas, Modelsfor Exploration. Houston Geological Society, Houston, pp 99–149
Coleman JM, Gagliano SM, Webb JE (1964) Minor sedimentary struc-tures in a prograding distributary. Mar Geol 1:240–258
De Raaf JFM, Boersma JR, van Gelder A (1977) Wave generatedstructures and sequences from a shallow marine succession. LowerCarboniferous, County Cork, Ireland. Sedimentology 4:1–52
Dettner K, Peters W (eds) (2010) Lehrbuch der Entomologie. Spektrum,Heidelberg
Donaldson AC, Martin RH, Kanes WH (1970) Holocene Guadelupedelta of Texas Guld Coast. In: Morgan JP (ed) Deltaic sedimenta-tion, modern and ancient, Soc. Econ. Paleontologists andMineralogists Special Publications, vol 15, pp 107–137
Palaeobio Palaeoenv (2014) 94:495–528 525
Author's personal copy

Dumas S, Arnott RWC (2006) Origin of hummocky and swaley cross-stratification—The controlling influence of unidirectional currentstrength and aggradation rate. Geology 34:1073–1076
Elliott T (1974) Interdistributary bay sequences and their genesis.Sedimentology 21:611–622
Elliott T (1986) Deltas. In: Reading HG (ed) Sedimentary environmentsand facies. Blackwell Scientific Publications, Oxford, pp 113–154
Etzold A, Schweizer V (2005) Der Keuper in Baden-Württemberg. In:Deutsche Stratigraphische Kommission (eds) Stratigraphie vonDeutschland IV – Keuper. Cour Forsch-Inst Senckenberg 253:214–264
Fielding CR, Trueman J, Alexander J (2005) Sharp-based mouth barsands from the Burdekin River Delta of northeastern Australia:extending the spectrum of mouth bar facies, geometry, and stackingpatterns. Int J Sed Res 75:55–66
Fischer J (2008) Brief synopsis of the hybodontiform form taxonLissodus Brough, 1935, with remarks on the environment andassociated fauna. FFH C528:1–23
Fisher DC (1979) Evidence for subaerial activity of Euproops danae(Merostomata, Xiphosurida). In: Nitecki MH (ed) Mazon Creekfossils. Academic Press, New York, pp 379–447
Fisher DC (1984) TheXiphosurida: archetypes or bradytely? In: EldredgeN, Stanley SM (eds) Living fossils. Springer, New York, pp196–213
Fisher WL, Brown LF, Scott AJ, McGowen JH (1972) Delta systems inthe exploration for oil and gas. Bureau of Economic Geology, TheUniversity of Texas, Austin
Fisk HN, McFarlan E, Kolb CR, Wilbert LJ (1954) Sedimentary frame-work of the modern Mississippi delta. J Sed Petrol 24:76–99
Folk RL, Ward WC (1957) Brazos River bar: a study in the significanceof grain size parameters. J Sed Petrol 27:3–26
Fraas E (1897) Reste von Zanclodon aus dem oberen Keuper vomLangenberge bei Wolfenbüttel. ZDGG 49:482–485
Franz M (2008) Litho- und Leitflächenstratigraphie, Chronostratigraphie,Zyklo- und Sequenzstratigraphie des Keupers im östlichenZentraleuropäischen Becken (Deutschland, Polen) und DänischenBecken (Dänemark, Schweden). Dissertation, Martin-Luther-Universität Halle-Wittenberg
Gall J-C, Grauvogel L, Lehman JP (1974) Faune du Buntsandstein. V. –Les poissons fossiles de la collection Grauvogel-Gall. Annales depaléontologie, Vertébrés 60:129–147
Galton PM (1984) Cranial anatomy of the prosauropod dinosaurPlateosaurus from the Knollenmergel (Middle Keuper, UpperTriassic) of Germany. I. Two complete skulls from Trossingen/Württ. With comments on the diet. Geol Pal 18:139–171
Gilbert GK (1885) The topographic features of lake shores. Ann Rep USGeol Survey 5:69–123
Gothan W (1914) Die unter-liassische (rhätische) Flora der Umgegendvon Nürnberg. Abh Nat Ges 19:91–186
Gould HR (1970) The Mississippi delta complex. In: Morgan JP, ShaverRH (eds) Deltaic sedimentation modern and ancient. Spec Publ SocEcon Paleont Miner, vol 15, Tulsa, pp 3–30
Grimaldi D, Engel MS (2005) Evolution of the Insects. CambridgeUniversity Press, Cambridge
Harms JC (1975) Stratification and sequence in prograding shorelinedeposits. SEPM, Short Course 2:81–102
Harris TM (1932) The fossil flora of Scoresby Sound East Greenland –Part 2: Description of seed plants incertae sedis together with adiscussion of certain Cycadophyte cuticles. Medd om Grønl 85:1–112
Harris TM (1937) The fossil flora of Scoresby Sound. Part 5.Stratigraphic relations of the plant beds. Medd Grønl 112:1–114
Haubold H (1989) Die Dinosaurier. A. Ziemsen, WittenbergHauschke N, Kozur HW (2011) Two new conchostracan species from the
Late Triassic of the Fuchsberg, northern foreland of the HarzMountains northeast of Seinstedt (Lower Saxony, Germany). In:
Sullivan R, Lucas S, Spielmann J (eds) Fossil Record 3, vol 53.New Mexico Museum of Natural History and Science,Albuquerque, pp 187–194
Hauschke N, Wilde V (1987) Paleolimulus fuchsbergensis n. sp.(Xiphosura, Merostomata) aus der oberen Trias vonNordwestdeutschland, mit einer Übersicht zur Systematik undVerbreitung rezenter Limuliden. Paläontol Z 61:87–108
Hauschke N, Wilde V (1991) Zur Verbreitung und Ökologiemesozoischer Limuliden. N Jb Geol Paläont, Abh 183:391–411
Hauschke N, Wilde V (1996) Fazies und Fossilführung im Unterrhät desFuchsberges bei Seinstedt (nördliches Harzvorland, Niedersachsen).Hall Jb Geowiss B18:141–152
Havlik P, Aiglstorfer M, El Atfy H, Uhl D (2013) A peculiar bonebedfrom the Norian Stubensandstein (Löwenstein Formation, LateTriassic) of southern Germany and its palaeoenvironmental inter-pretation. N Jb Geol Paläont, Abh 269:321–337
Heunisch C (1998) Palynologischer Untersuchungsbericht, Fuchsberg.Technical report, 1743_2864. State Authority for Mining, Energyand Geology, Hannover
Heunisch C (1999) Die Bedeutung der Palynologie für Biostratigraphieund Fazies in der Germanischen Trias. In: Hauschke N, WildeV (eds) Trias – Eine ganz andere Welt. Verlag Dr. FriedrichPfeil, München, pp 207–220
Heunisch C (2011a) Mikropaläontologischer Untersuchungsbericht,Fuchsberg/Langenberg. Technical report, 2010_3467, StateAuthority for Mining, Energy and Geology, Hannover
Heunisch C (2011b) Mikropaläontologischer Untersuchungsbericht,Fuchsberg/Langenberg. Technical report, 2010_3500, StateAuthority for Mining, Energy and Geology, Hannover
Heunisch C, Nitsch E (2011) Eine seltene Mikroflora aus der Mainhardt-Formation (Keuper, Trias) von Baden-Württemberg (Süd-deutschland). Jb Mitt Oberrheinischen Geol Vereins, NF 93:1–22
Holbrook JM (1996) Complex fluvial response to low gradients at max-imum regression: a genetic link between smooth sequence-boundarymorphology and architecture of overlying sheet sandstone. Int J SedRes 66:713–722
Hubbard MD, Peters WL (1978) Environmental requirements and pollu-tion tolerance of Ephemeroptera. Environmental Protection Agency,Springfield
Hunter RE, Clifton HE (1982) Cyclic deposition and hummocky cross-stratification of probable storm origin in Upper Cretaceous rocks ofthe Cape Sebastian Area, southwestern Oregon. J Sed Petrol 52:127–143
Jaekel O (1911) Die Wirbeltiere. Eine Übersicht über die fossilen undlebenden Formen. Bornträger, Berlin
Jones B, Manning DAC (1994) Comparison of geochemical indices usedfor the interpretation of palaeoredox conditions in ancient mud-stones. Chem Geol 111:111–129
Jopling AV, Walker RG (1968) Morphology and origin of ripple-driftcross-lamination with examples from the Pleistocene ofMassachusetts. J Sediment Petrol 38:971–984
Jung W (1968) Hirmerella muensteri (Schenk) Jung nov. comb., einebedeutsame Konifere des Mesozoikums. Palaeontographica B122:53–93
Jüngst H (1928) Rhät, Psilonoten- und Schlotheimienschichten imnördlichen Harzvorlande. Geol Pal Abh 20 (NF 16):1–194
Jüngst H (1929) Zur vergleichenden Stratigraphie des Rät zwischenHarz und Elsaß. Dissertation, Technische Universität,Darmstadt
Kelber K-P, van Konijnenburg–van Cittert JHA (1997) A new Rhaetianflora from the neighbourhood of Coburg (Germany) – preliminaryresults. Mededelingen Nederlands Instituut voor ToegepasteGeowetenschappen 58:105–113
Kobayashi T (1954) Fossil Estherians and allied fossils. J Fac Sci,University of Tokyo, Section 2, 9:1–192
526 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy

Koch G, Schneider W (1985) Der Obere Keuper Südost-Niedersachsensin Übertage-Aufschlüssen – Fazies und Mineralbestand. Ber NatGes Hannover 125:19–48
Kozur HW, Weems R (2010) The biostratigraphic importance ofconchostracans in the continental Triassic of the northern hemisphere.In: Lucas S (ed) The Triassic Timescale, vol 334, Geological Societyof London. Special Publications, London, pp 315–417
Kumar N, Sanders JE (1976) Characteristics of shoreface storm deposits:Modern and ancient examples. J Sed Petrol 46:145–162
Kustatscher E, van Konijenburg–van Cittert JHA (2007) Taxonomicaland palaeogeographic considerations on the seedfern genusPtilozamites. N Jb Geol Paläont, Abh 243:71–100
Kustatscher E, Heunisch C, van Konijenburg–van Cittert JHA (2012)Taphonomical implications of the Ladinian megaflora andpalynoflora of Thale (Germany). Palaios 27:753–764
Lecointre G, Le Guyader H (2006) Biosystematik. Springer, BerlinLehmann U, Hillmer G (1997) Wirbellose Tiere der Vorzeit. Enke, StuttgartLewandowski J (1988) Sedimentpetrographische Untersuchungen im
Keuper Nordwestdeutschlands. Dissertation, Ruhr-Universität,Bochum
Littke R, Scheck-Wenderoth M, Brix MR, Nelskamp S (2008)Subsidence, inversion and evolution of the therman field. In:Littke R, Bayer U, Gajewski D, Nelskamp S (eds) Dynamics ofcomplex sedimentary basins. The example of the Central EuropeanBasin system. Springer, Berlin, pp 125–152
Look E-R (1984) Geologie und Bergbau im Braunschweiger Land(Nördliches Harzvorland, Asse, Elm – Lappwals, Peine –Salzgitter, Allertal). Dokumentation zur Geologischen Wander-karte 1:100.000. Ber Nat Ges Hannover 127:1–467
López-Arbarello A (2008) Revision of Semionotus bergeriAgassiz, 1833(Upper Triassic, Germany), with comments on the taxonomic statusof Semionotus (Actinopterygii, Semionotiformes). Paläontol Z 82:40–54
Lund JJ (1977) Rhaetic to Lower Liassic palynology of the onshoresouth-eastern North Sea Basin. Geological Survey of Denmark, II.Series 109:1–129
Mackey SD, Bridge JS (1995) Three dimensional model of alluvialstratigraphy: theory and application. Int J Sed Res B65:7–31
Malz H, Poschmann M (1993) Erste Süßwasser-Limuliden (Arthropoda,Chelicerata) aus dem Rotliegenden der Saar-Nahe-Senke.Osnabrücker naturwiss Mitt 19:21–34
Martin JW (1989) Morphology of feeding structures in the Conchostracawith special reference to Lynceus. In: Felgenhauer BE, Watling L,Thistle AB (eds) Functional morphology of feeding and groomingin Crustacea, vol 5, Crustacean Issues, Balkema
McCune AR (1986) A revision of Semionotus (Pisces: Semionotidae)from the Triassic and Jurassic of Europe. Palaeontology 29:213–233
Merrit RW, Cummins KW (1978) An introduction to the aquatic insectsof North America. Kendall-Hunt, Dubuque
Miall AD (1977) A review of the braidedriver depositional environment.Earth Sci Rev 13:1–62
Morton RA, Gelfenbaum G, Jaffe BE (2007) Physical criteria fordistinguishing sandy tsunami and storm deposits using modernexamples. Sed Geol 200:184–207
Murry, PA (1989) Paleoecology and vertebrate faunal relationships of theUpper Triassic Dockum and Chinle Formations, southwesternUnited States. In: Lucas SG, Hunt AP (eds) Dawn of the Age ofdinosaurs in the American Southwest. New Mexico Museum ofNatural History, pp 375–400
Myrow PM, Southard JB (1996) Tempestite deposition. Int J Sed Res 66:875–887
Nitsch E (1996) Fazies, Diagenese und Stratigraphie der Grabfeld-Gruppe Süddeutschlands (Keuper, Trias). Dissertation, UniversitätKöln
Paul J, Siggelkow D (2004) Lakustrische Sedimente der Solling-Folge(Mittlerer Buntsandstein, Norddeutschland). Geol Jb Hessen 131:61–79
Pemberton SG, Ichnology Research Group (IRG) (1998) IchnologyAtlas. University of Alberta at Edmonton and University ofColorado at Boulder, electronic source
Pflücker y Rico L (1868) Das Rhät (die Rhätische Gruppe) in derUmgebung von Göttingen. Dissertation, Universität Göttingen
Pieńkowski G, Niedźwiedzki G, Waksmundzka M (2012)Sedimentological, palynological and geochemical studies of theterrestrial Triassic—Jurassic boundary in northwestern Poland.Geol Mag 149:308–332
Reading HG (1996) Sedimentary environments: processes, facies andstratigraphy. Blackwell , Oxford
Reible P (1962) Die Conchostraken (Branchiopoda, Crustacea) derGermanischen Trias. N Jb Geol Paläont, Abh 114:169–244
Reineck H-E (1963) Sedimentgefüge im Bereich der südlichen Nordsee.Abh Senckenberg naturforsch Ges 505:1–138
Reineck HE, Singh IB (1980) Depositional sedimentary environments –with reference to terrigenous clastics. Springer, Berlin
Reinhardt L, RickenW (2000) The stratigraphic and geochemical record ofplaya cycles: monitoring a Pangean monsoon-like system (Triassic,Middle Keuper, S. Germany). Palaeogeogr Palaeoclimatol Palaeoecol195:99–124
Retallack JT, Clifford HF (1980) Periodicity of crustaceans in a salineprairie stream of Alberta, Canada. Am Midl Nat 103:123–132
Reynolds AD (1999) Dimensions of paralic sandstone bodies. AAPGBull 83:211–229
Roemer H (1874) Ueber ein neues Vorkommen des Räth bei Hildesheim.ZDGG 26:349–354
Röhling H-G (2002) Rhätsandsteine am Fuchsberg östlich von Seinstedt.NLfB Geotop Info Flyer
Rosen DE, Forey PL, Gardiner BG, Patterson C (1981) Lungfishes,tetrapods, paleontology and plesiomorphy. Bull Am Mus Nat Hist167:159–276
Royan JP (1976) Studies on the gut contents of Leptestheriellamaduraiensis (Conchostraca: Branchiopoda) Nayar and Nair.Hydrobiologia 51:209–212
Ruckwied K (2009) Palynology of Triassic/Jurassic boundary key sec-tions of the NW Tethyan Realm (Hungary and Slovakia).Dissertation, Technische Universität Darmstadt
Russel RJ (1967) River and delta morphology: Louisiana StateUniversity. Coastal Studies Institute, Tech Rept No 52:55
Rust BR (1978) Depositional models for braided alluvium. In: Miall AD(ed) Fluvial sedimentology, Canadian Society of PetroleumGeologists Memoir, vol 5, pp 605–625
Sander PM (1992) The Norian Plateosaurus bonebeds of central Europeand their taphonomy. Palaeogeogr Palaeoclimatol Palaeoecol 93:255–299
Sander PM (1999) Die Plateosaurus-Friedhöfe im Keuper: Belege füreinen Massentod? In: Hauschke N, Wilde V (eds) Trias – Eine ganzandere Welt. Verlag Dr. Friedrich Pfeil, München, pp 419–425
Schaeffer B (1967) Late Triassic fishes from the western United States.Bull Am Mus Nat Hist 135:287–342
Schaeffer B, Rosen DE (1961) Major adaptive levels in the evolution ofthe actinopterygian feeding mechanism. Am Zool 1:187–204
Scheck M, Bayer U (1999) Evolution of the Northeast German Basin–inferences from a 3D structural model and subsidence analysis.Tectonophysics 313:145–169
Scheck M, Bayer U, Lewerenz B (1996) Lithosphere structure under-neath the Northeast German Basin: constraints by gravity modeling.Phys Chem Earth 21:313–318
Schenk A (1867) Die fossile Flora der Grenzschichten des Keupers undLias Frankens. Kreidel, Wiesbaden
Schimper WP (1869) Traitè de Paléontologie végétale ou la flore dumonde primitif dans ses rapports avec les formations géologiqueset la flore du monde actuel. Tome premier. Bailliére et Fils, Paris
Schimper WP (1870–1872) Traitè de Paléontologie végétale ou laflore du monde primitif dans ses rapports avec les formations
Palaeobio Palaeoenv (2014) 94:495–528 527
Author's personal copy

géologiques et la flore du monde actuel. Tome second.Bailliére et Fils, Paris
Schimper WP (1874) Traitè de Paléontologie végétale ou la flore dumonde primitif dans ses rapports avec les formations géologiqueset la flore du monde actuel. Tome troisième. Bailliére et Fils, Paris
Schlönbach A (1861) Mittheilung über das Bonebed bei Seinstedt unweitHornburg. ZDGG XIII:17 and 353
Schlönbach A (1862) Beitrag zur genauen Niveau-Bestimmung des aufder Grenze zwischen Keuper und Lias im Hannoverischen undBraunschweigischen auftretenden Sandsteins. N Jb Miner GeognGeol Petrefaktenk 1862:146–177
Schultze H-P, Kriwet J (1999) Die Fische der Germanischen Trias. In:Hauschke N, Wilde V (eds) Trias – Eine ganz andere Welt. VerlagDr. Friedrich Pfeil, München, pp 239–250
Schweitzer HJ, van Konijnenburg-van Cittert JHA, van der Burgh J(1997) The Rhaeto–Jurassic flora of Iran & Afghanistan. 10.Bryophyta, Lycophyta, Sphenophyta, Pterophyta-Eusporangiatae& -Protoleptosporangiatae. Palaeontographica B243:103–192
Schweitzer HJ, Kirchner M, van Konijnenburg-van Cittert JHA (2000)The Rhaeto-Jurassic flora of Iran & Afghanistan. 12. CycadophytaII. Nilssoniales. Palaeontographica B254
Schweizer R (1966) Ein Coelacanthide aus dem Oberen MuschelkalkGöttingens. N Jb Geol Paläont, Abh 125:216–226
Scruton PC (1960) Delta building and the deltaic sequence. In: ShepardFP, Phleger FB, van Andel TH (eds) Recent sediments, northwestGulf of Mexico. American Association of Petroleum Geologists,Tulsa, pp 82—102
Seeling M (1999) Fazielle und sequenzstratigraphische Untersuchungenim Rhätkeuper (Exter-Formation) Thüringens und des nördlichenHarzvorlandes. Diploma thesis, Martin-Luther-Universität Halle-Wittenberg
Seilacher A (1943) Elasmobranchier-Reste aus dem oberen Muschelkalkund dem Keuper Württembergs. N Jb Miner Geol Paläont,Mh 10:256–292
Shcherbakov DE (1992) The earliest leafhopper (Hemiptera,Karajassidae n. fam) from the Jurassic of Karatau. N Jb GeolPaläont, Mh 1992:39–51
ShcherbakovDE (2000) Permian faunas of Homoptera (Hemiptera) in relationto phytogeography and the Permo-Triassic Crisis. Pal J 34:251–267
Shukla UK, Bachmann GH, Beutler G, Barnasch J, Franz M (2006)Extremely distal fluvial sandstones within the playa system ofArnstadt Formation (Norian, Late Triassic), Central Germany.Facies 52:541–555
Smith ND (1972) Flume experiments on the durability of mudclasts. JSed Petrol 42:378–384
Southard JB, Boguchwal LA (1990) Bed configurations in steady unidi-rectional water flows. Part 2. Synthesis of flume data. J Sed Petrol60:658–679
Stampfli GM, Kozur HW (2006) Europe from the Variscan to the Alpinecycles. In: Gee DG, Stephenson RA (eds) European lithospheredynamics. Geological Society Memoir, vol 32, London, pp 333–343
Stern DH, Stern MS (1971) Morphology and culture of Eulimnadiadiversa (Crustacea, Conchostraca) found in Louisiana. Trans AmMicrosc Soc 90:483–486
Stollhofen H, Barnasch J, Bayer U, Beutler G, FranzM, Kästner M, Legler B,Mutterlose J, Radies D (2008) Upper Rotliegend to Lower Cretaceousbasin development. In: LittkeR, Bayer U,Gajewski D,NelskampS (eds)Dynamics of complex sedimentary basins. The example of the CentralEuropean basin system. Springer, Berlin, pp 181–210
Storch V, Welsch U (2004) Systematische Zoologie. Spektrum, BerlinStrenth NE, Sissom SL (1975) A morphological study on the post-
embryonic stages of Eulimnadia texana Packard (Conchostraca,Crustacea). Texas J Sci 26:137–154
Tye RS, Coleman JM (1989) Depositional processes and stratigraphy offluvially dominated lacustrine deltas: Mississippi Delta plain. J SedPetrol 59:973–996
Tye RS, Kosters EC (1986) Styles of interdistributary basins sedimenta-tion: Mississippi delta plain, Louisiana. Trans Gulf Coast Ass GeolSoc 36:575–588
Underwood CJ, Ward DJ (2004) Environmental distribution of Bathonian(Middle Jurassic) neoselachians in southern England. In: Arratia G,Tintori A (eds) Mesozoic Fishes 3 – Systematics, Paleoenvironmentsand Biodiversity. Verlag Dr. Friedrich Pfeil, München, pp 111–122
van Konijenburg–van Cittert JHA, Schmeißner S, Hauptmann S,Hauptmann T (1998) Neue Ergebnisse zu Ctenozamiteswolfiana (Pteridosperma) und Pseudoctenis prossii nov. spec.(Cycadophyta) aus dem Unteren Lias (Jura, Bayern). Doc Nat117:13–33
von Engelhardt W (1942) Untersuchungen an den Schwermineralen desnordwestdeutschen Rät. Öl Kohle 38:259–265
von Huene F (1926) Vollständige Osteologie eines Plateosauriden ausdem schwäbischen Keuper. Geol Pal Abh, NF 15:139–179
von Strombeck A (1852) Ueber den oberen Keuper bei Braunschweig.ZDGG 4:54–82
Walker RG, Harms JC (1971) The ‘Catskill Delta’: A prograding muddyshoreline in central Pennsylvania. J Geol 79:381–399
Ward JV (1992) Aquatic insect ecology, Part 1: Biology and habitat.Wiley, New York
WarrenG (1974) Simplified form of the Folk-Ward skewness parameter. JSed Petrol 44:259
Weber R (1968) Die fossile Flora der Rhät-Lias-Übergangsschichten vonBayreuth (Oberfranken) unter besonderer Berücksichtigung derCoenologie. Erl Geol Abh 72:3–73
Weishampel DB (1984) Trossingen: E. Fraas, F. von Huene, R. Seemann,and the ‘Schwäbische Lindwurm’ Plateosaurus. In: Reif W-E,Westphal F (eds) Third Symposium on Mesozoic TerrestrialEcosystems. Attempto, Tübingen, pp 249–253
Weishampel DB, Westphal F (1986) Die Plateosaurier vonTrossingen. Attempto, Tübingen
Wentworth C (1922) A scale of grade and class terms for clastic sedi-ments. J Geol 30:377–392
Winkler T (1880) Description de quelques restes du poissonsfossiles des terrains triasiques des environs de Wurzburg.Arch Musée Teyler 5:1–41
Wright DL (1977) Sediment transport and deposition at river mouths: asynthesis. Geol Soc Am Bull 88:857–868
Yabumoto Y (2008) A new Mesozoic coelacanth from Brazil(Sarcopterygii, Actinistia). Pal Res 12:329–343
Zavialova N, van Konijnenburg–van Cittert JHA (2011) Exineultrastructure of in situ peltasperm pollen from the Rhaetianof Germany and its implications. Rev Palaeobot Palyno 168:720
Ziegler PA (1990) Geological Atlas ofWestern and Central Europe. ShellInternational Petroleum, Den Haag
Ziegler B (1998a) Einführung in die Paläobiologie, Teil 2: SpeziellePaläontologie: Protisten, Spongien und Coelenteraten. Mollusken.E. Schweizerbart, Stuttgart
Ziegler B (1998b) Einführung in die Paläobiologie, Teil 3: SpeziellePaläontologie: Würmer, Arthropoden, Lophophoraten.
Echinodermen. E. Schweizerbart, StuttgartZimmermann J, Franz M, Wolfgramm M (2014) The Late Aalenian
Polyplocussandstein Formation in SE Lower Saxony,Germany: Meandering distributary and crevasse splay sedi-mentation on a lower deltaic plain. N Jb Geol Paläont, Abh271:69–94
528 Palaeobio Palaeoenv (2014) 94:495–528
Author's personal copy




















