
Large variations in absolute wall shear stress levels within one species and between species
11
Atherosclerosis 195 (2007) 225–235 Review Large variations in absolute wall shear stress levels within one species and between species Caroline Cheng a,∗,1 , Frank Helderman a,1 , Dennie Tempel a , Dolf Segers a , Beerend Hierck b , Rob Poelmann b , Arie van Tol e , Dirk J. Duncker a , Danielle Robbers-Visser c , Nicolette T.C. Ursem d , Rien van Haperen e , Jolanda J. Wentzel a , Frank Gijsen a , Anton F.W. van der Steen a , Rini de Crom e,f , Rob Krams a,g a Department of Cardiology, Thoraxcenter, Erasmus MC, Rotterdam, The Netherlands b Department of Embryology and Anatomy, LUMC, Leiden, The Netherlands c Department of Pediatric Cardiology, Erasmus MC, Rotterdam, The Netherlands d Department of Obstetrics, Erasmus MC, Rotterdam, The Netherlands e Department of Cell Biology, Erasmus MC, Rotterdam, The Netherlands f Department of Vascular Surgery, Erasmus MC, Rotterdam, The Netherlands g Department of Physiology, Medical Physics and Vascular Surgery, VUMC, Amsterdam, The Netherlands Received 7 August 2006; received in revised form 25 October 2006; accepted 15 November 2006 Available online 12 December 2006 Abstract Wall shear stress (WSS), the frictional force between blood and endothelium, is an important determinant of vascular function. It is generally assumed that WSS remains constant at a reference value of 15 dyn/cm 2 . In a study of small rodents, we realized that this assumption could not be valid. This review presents an overview of recent studies in large and small animals where shear stress was measured, derived from velocity measurements or otherwise, in large vessels. The data show that large variations exist within a single species (human: variation of 2–16 N/m 2 ). Moreover, when we compared different species at the same location within the arterial tree, an inverse relationship between animal size and wall shear stress was noted. When we related WSS to diameter, a unique relationship was derived for all species studied. This relationship could not be described by the well-known r 3 law of Murray, but by the r 2 law introduced by Zamir et al. in 1972. In summary, by comparing data from the literature, we have shown that: (i) the assumption of a physiological WSS level of ∼15 dyn/cm 2 for all straight vessels in the arterial tree is incorrect; (ii) WSS is not constant throughout the vascular tree; (iii) WSS varies between species; (iv) WSS is inversely related to the vessel diameter. These data support an “r 2 law” rather than Murray’s r 3 law for the larger vessels in the arterial tree. © 2006 Elsevier Ireland Ltd. All rights reserved. Keywords: Wall shear stress; Animal; Clinical; Patients; Mice; Mouse; Rat; Rabbit; Dog; In vivo; Human; Doppler Contents 1. Introduction ........................................................................................................ 226 2. Variations in mean WSS in the arterial system ......................................................................... 226 2.1. Human individuals ........................................................................................... 226 2.2. Cross-species differences ...................................................................................... 231 ∗ Corresponding author at: Erasmus MC, Room Ee1073b, Dr. Molewaterplein 50, 3015 GE Rotterdam, The Netherlands. Tel.: +31 10 4089393; fax: +31 10 4089494. E-mail address: [email protected] (C. Cheng). 1 Authors have contributed equally to this manuscript. 0021-9150/$ – see front matter © 2006 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.atherosclerosis.2006.11.019
Transcript of Large variations in absolute wall shear stress levels within one species and between species

A
anv
sr
aW©
K
C
f
0d
Atherosclerosis 195 (2007) 225–235
Review
Large variations in absolute wall shear stress levelswithin one species and between species
Caroline Cheng a,∗,1, Frank Helderman a,1, Dennie Tempel a, Dolf Segers a, Beerend Hierck b,Rob Poelmann b, Arie van Tol e, Dirk J. Duncker a, Danielle Robbers-Visser c,
Nicolette T.C. Ursem d, Rien van Haperen e, Jolanda J. Wentzel a, Frank Gijsen a,Anton F.W. van der Steen a, Rini de Crom e,f, Rob Krams a,g
a Department of Cardiology, Thoraxcenter, Erasmus MC, Rotterdam, The Netherlandsb Department of Embryology and Anatomy, LUMC, Leiden, The Netherlands
c Department of Pediatric Cardiology, Erasmus MC, Rotterdam, The Netherlandsd Department of Obstetrics, Erasmus MC, Rotterdam, The Netherlands
e Department of Cell Biology, Erasmus MC, Rotterdam, The Netherlandsf Department of Vascular Surgery, Erasmus MC, Rotterdam, The Netherlands
g Department of Physiology, Medical Physics and Vascular Surgery, VUMC, Amsterdam, The Netherlands
Received 7 August 2006; received in revised form 25 October 2006; accepted 15 November 2006Available online 12 December 2006
bstract
Wall shear stress (WSS), the frictional force between blood and endothelium, is an important determinant of vascular function. It is generallyssumed that WSS remains constant at a reference value of 15 dyn/cm2. In a study of small rodents, we realized that this assumption couldot be valid. This review presents an overview of recent studies in large and small animals where shear stress was measured, derived fromelocity measurements or otherwise, in large vessels.
The data show that large variations exist within a single species (human: variation of 2–16 N/m2). Moreover, when we compared differentpecies at the same location within the arterial tree, an inverse relationship between animal size and wall shear stress was noted. When weelated WSS to diameter, a unique relationship was derived for all species studied.
This relationship could not be described by the well-known r3 law of Murray, but by the r2 law introduced by Zamir et al. in 1972.
In summary, by comparing data from the literature, we have shown that: (i) the assumption of a physiological WSS level of ∼15 dyn/cm2 forll straight vessels in the arterial tree is incorrect; (ii) WSS is not constant throughout the vascular tree; (iii) WSS varies between species; (iv)SS is inversely related to the vessel diameter. These data support an “r2 law” rather than Murray’s r3 law for the larger vessels in the arterial tree.2006 Elsevier Ireland Ltd. All rights reserved.
eywords: Wall shear stress; Animal; Clinical; Patients; Mice; Mouse; Rat; Rabbit; Dog; In vivo; Human; Doppler
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2. Variations in mean WSS in the arterial system . . . . . . . . . . . . . . . . . . .
2.1. Human individuals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.2. Cross-species differences . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
∗ Corresponding author at: Erasmus MC, Room Ee1073b, Dr. Molewaterplein 50ax: +31 10 4089494.
E-mail address: [email protected] (C. Cheng).1 Authors have contributed equally to this manuscript.
021-9150/$ – see front matter © 2006 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.atherosclerosis.2006.11.019
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 226. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 226. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 226
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 231
, 3015 GE Rotterdam, The Netherlands. Tel.: +31 10 4089393;

2
1
beIhmwtamfwetTtv
toa0WiiWca1mc
∼dvtuptrp[tdmb
26 C. Cheng et al. / Atherosclerosis 195 (2007) 225–235
3. Modification of Murray’s law may explain a non-uniform WSS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2324. Limitations of the assessed studies. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2335. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 233
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 233References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 233
. Introduction
Wall shear stress (WSS), the frictional force betweenlood and endothelium, is an important determinant ofndothelial cell function, gene expression, and structure.ndeed, a variety of studies provided evidence that WSSas to be maintained between certain limits in order toaintain vascular haemostasis. WSS is actively maintainedithin limits during intrauterine growth, during the neona-
al period and early childhood, and during exercise in thedult. Inappropriate values of WSS have been associated withaladaptive growth, patent ductus arteriosus, congenital mal-
ormations of the heart and atherosclerosis [1–3]. Indeed,hen WSS is reduced by 30% in vivo in ApoE mice, the
xpression of several atherogenic genes is induced, whichriggers the development of large atherosclerotic lesions [4].o avoid these conditions, the endothelium in the arterial sys-
em should be responsive to WSS within a narrow range ofalues that are considered “normal”.
At present, it remains unclear whether the endotheliumhroughout the arterial system is primed with the same rangef WSS values. Currently, researchers in the field oftenssume mean WSS levels of ∼15 dyn/cm2 (1 dyne/cm2 =.1 N/m2) as acceptable, because it represents the averageSS values over the cardiac cycle of the large straight arter-
es experiencing steady laminar flow. This is based on studiesn patients and animal models, which provide evidence that
SS actively influences vessel wall remodeling [5–8]. Thisompensatory response mediated by the endothelium aimst the maintenance of a WSS magnitude of approximately5–20 dyn/cm2. Partially based on this notion, it is also com-only assumed that this acceptable range of WSS is rather
onstant throughout the vascular system [9–14].Another argument for a constant WSS value of
15 dyn/cm2 at different locations in the arterial system iserived from the principle of minimal work for the cardio-ascular system as proposed by Murray [15]. He stated thathe total energy to drive the blood and to maintain blood vol-me is minimized in the arterial system. Deducted from thisrinciple is Murray’s law [16], which states that the cube ofhe radius of the mother vessel equals the sum of cubes of theadii of the daughter vessels. While this optimization princi-le predicts a constant WSS throughout the vascular system17], a number of recent publications show a broad range in
WSS varies with the location across the cardiovascular sys-tem within one species, and that there are cross-species differ-ences. In spite of this, a paucity of data exists which compareWSS at different anatomical locations [21,26] or betweenspecies in one type of vessel. Until now, no reviews are avail-able which summarize the separate WSS values found inliterature to provide an adequate overview about this subject.
In this review, we present evidence from literature that sup-ports the concept that WSS levels are not identical throughoutthe vascular tree. We also provide evidence that WSS levelsdiffer between species. The interpretation of these data willbe discussed in relation to a modification of Murray’s law.Acceptance of this concept would have significant impli-cations for further WSS research, as the importance of theeffects of anatomical localization of the studied endothelialcells and the species from which they are derived is oftenoverlooked in current studies.
2. Variations in mean WSS in the arterial system
2.1. Human individuals
Vascular disease affects the vascular remodeling capaci-ties of arteries, which could result in an alteration of the meanWSS level. Accordingly, only studies performed in healthyhuman subjects are included in this review.
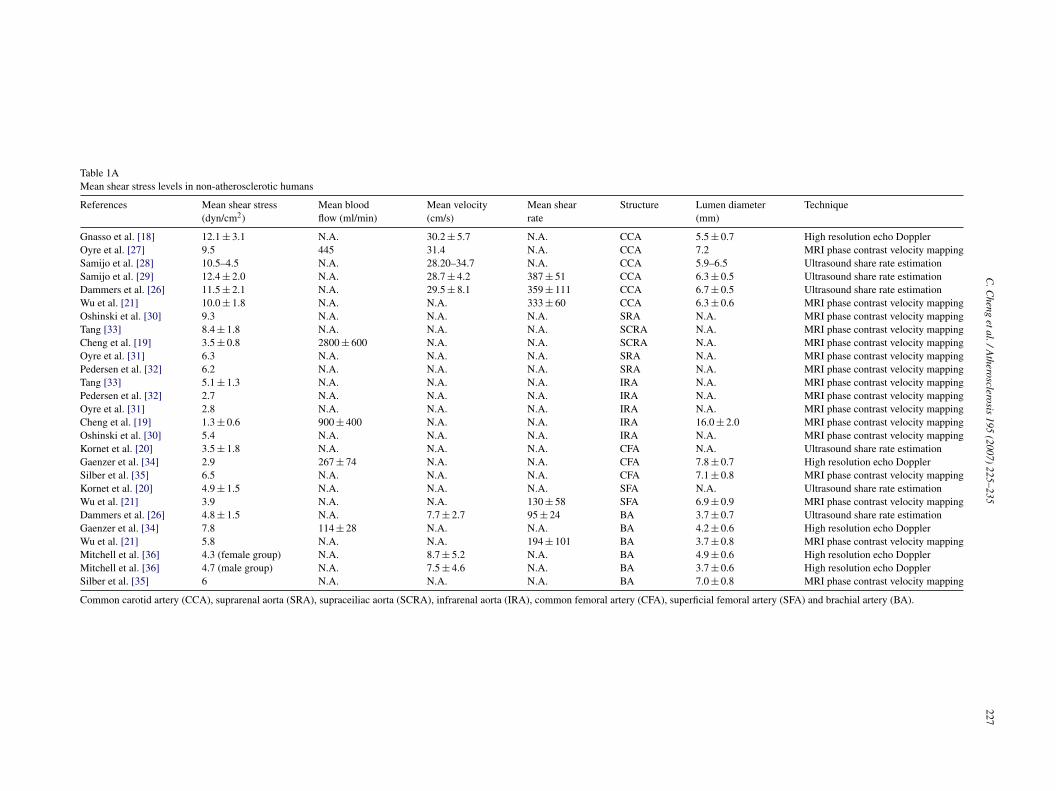
In Table 1A, the WSS values in different types of arter-ies derived from experimental data of several investigatorsare summarized [18–21,26–39]. All WSS values are derivedfrom in vivo measurements in conscious human subjects, byapplying either ultrasound or MRI techniques. All valuesare stationary WSS values or recalculated to become sta-tionary WSS values, and therefore the time dependence ofthe WSS measurements are not taken into account. Excel-lent studies and reviews are present on this topic [40–42].For more information on the studies that we have includedin Table 1A concerning background, and details in meth-ods, we refer to Table 1B. It should be mentioned that theWSS values in Table 1A are obtained from studies testingdifferent hypothesizes (e.g. the effect of smoking or exerciseon blood blow), with different study designs testing specificparameters. However, only WSS values obtained from thecontrol (non-treatment) groups were included in this review.
he actual mean WSS levels that could be measured in theifferent types of arteries in humans [18–21]. Flow measure-ents in animal models also show differences in WSS levels
etween species [22–25]. These data therefore indicate that
Ssyl
election criteria for the incorporated studies are as followed:ubjects of the selected control group are younger than 40ears old, do not show clinical manifestation of cardiovascu-ar disease, are non-smokers and do not receive any prescribed
C.C
hengetal./A
therosclerosis195
(2007)225–235
227
Table 1AMean shear stress levels in non-atherosclerotic humans
References Mean shear stress(dyn/cm2)
Mean bloodflow (ml/min)
Mean velocity(cm/s)
Mean shearrate
Structure Lumen diameter(mm)
Technique
Gnasso et al. [18] 12.1 ± 3.1 N.A. 30.2 ± 5.7 N.A. CCA 5.5 ± 0.7 High resolution echo DopplerOyre et al. [27] 9.5 445 31.4 N.A. CCA 7.2 MRI phase contrast velocity mappingSamijo et al. [28] 10.5–4.5 N.A. 28.20–34.7 N.A. CCA 5.9–6.5 Ultrasound share rate estimationSamijo et al. [29] 12.4 ± 2.0 N.A. 28.7 ± 4.2 387 ± 51 CCA 6.3 ± 0.5 Ultrasound share rate estimationDammers et al. [26] 11.5 ± 2.1 N.A. 29.5 ± 8.1 359 ± 111 CCA 6.7 ± 0.5 Ultrasound share rate estimationWu et al. [21] 10.0 ± 1.8 N.A. N.A. 333 ± 60 CCA 6.3 ± 0.6 MRI phase contrast velocity mappingOshinski et al. [30] 9.3 N.A. N.A. N.A. SRA N.A. MRI phase contrast velocity mappingTang [33] 8.4 ± 1.8 N.A. N.A. N.A. SCRA N.A. MRI phase contrast velocity mappingCheng et al. [19] 3.5 ± 0.8 2800 ± 600 N.A. N.A. SCRA N.A. MRI phase contrast velocity mappingOyre et al. [31] 6.3 N.A. N.A. N.A. SRA N.A. MRI phase contrast velocity mappingPedersen et al. [32] 6.2 N.A. N.A. N.A. SRA N.A. MRI phase contrast velocity mappingTang [33] 5.1 ± 1.3 N.A. N.A. N.A. IRA N.A. MRI phase contrast velocity mappingPedersen et al. [32] 2.7 N.A. N.A. N.A. IRA N.A. MRI phase contrast velocity mappingOyre et al. [31] 2.8 N.A. N.A. N.A. IRA N.A. MRI phase contrast velocity mappingCheng et al. [19] 1.3 ± 0.6 900 ± 400 N.A. N.A. IRA 16.0 ± 2.0 MRI phase contrast velocity mappingOshinski et al. [30] 5.4 N.A. N.A. N.A. IRA N.A. MRI phase contrast velocity mappingKornet et al. [20] 3.5 ± 1.8 N.A. N.A. N.A. CFA N.A. Ultrasound share rate estimationGaenzer et al. [34] 2.9 267 ± 74 N.A. N.A. CFA 7.8 ± 0.7 High resolution echo DopplerSilber et al. [35] 6.5 N.A. N.A. N.A. CFA 7.1 ± 0.8 MRI phase contrast velocity mappingKornet et al. [20] 4.9 ± 1.5 N.A. N.A. N.A. SFA N.A. Ultrasound share rate estimationWu et al. [21] 3.9 N.A. N.A. 130 ± 58 SFA 6.9 ± 0.9 MRI phase contrast velocity mappingDammers et al. [26] 4.8 ± 1.5 N.A. 7.7 ± 2.7 95 ± 24 BA 3.7 ± 0.7 Ultrasound share rate estimationGaenzer et al. [34] 7.8 114 ± 28 N.A. N.A. BA 4.2 ± 0.6 High resolution echo DopplerWu et al. [21] 5.8 N.A. N.A. 194 ± 101 BA 3.7 ± 0.8 MRI phase contrast velocity mappingMitchell et al. [36] 4.3 (female group) N.A. 8.7 ± 5.2 N.A. BA 4.9 ± 0.6 High resolution echo DopplerMitchell et al. [36] 4.7 (male group) N.A. 7.5 ± 4.6 N.A. BA 3.7 ± 0.6 High resolution echo DopplerSilber et al. [35] 6 N.A. N.A. N.A. BA 7.0 ± 0.8 MRI phase contrast velocity mapping
Common carotid artery (CCA), suprarenal aorta (SRA), supraceiliac aorta (SCRA), infrarenal aorta (IRA), common femoral artery (CFA), superficial femoral artery (SFA) and brachial artery (BA).
228C
.Cheng
etal./Atherosclerosis
195(2007)
225–235
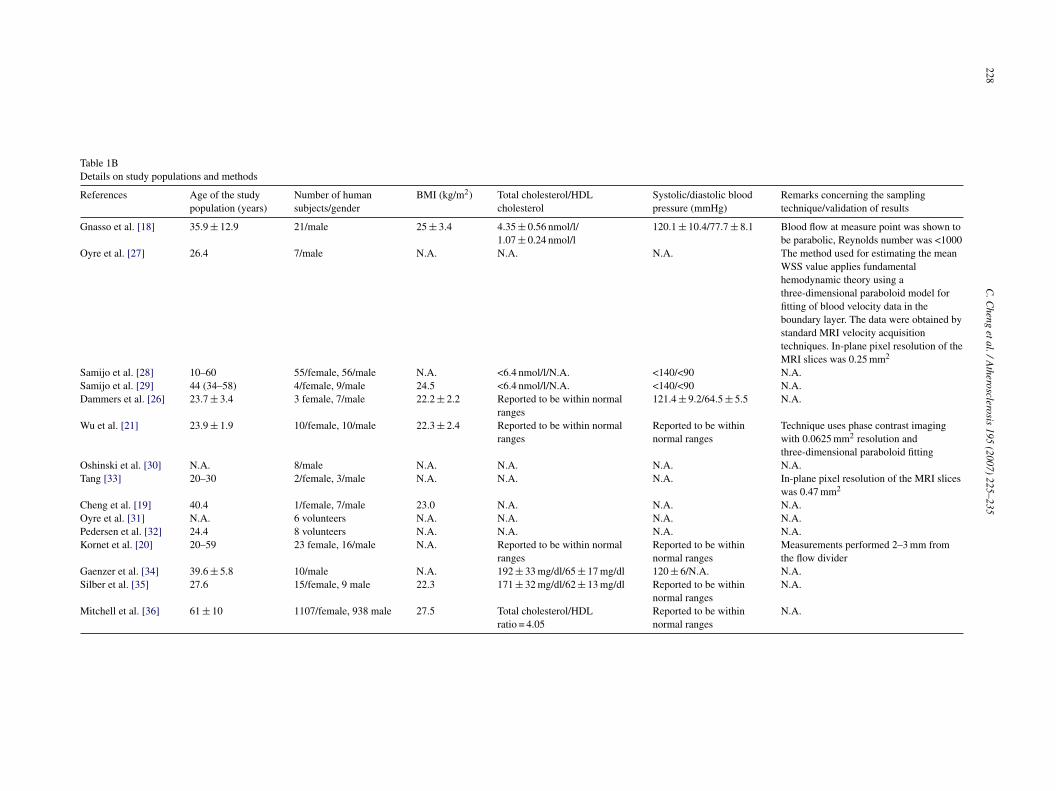
Table 1BDetails on study populations and methods
References Age of the studypopulation (years)
Number of humansubjects/gender
BMI (kg/m2) Total cholesterol/HDLcholesterol
Systolic/diastolic bloodpressure (mmHg)
Remarks concerning the samplingtechnique/validation of results
Gnasso et al. [18] 35.9 ± 12.9 21/male 25 ± 3.4 4.35 ± 0.56 nmol/l/1.07 ± 0.24 nmol/l
120.1 ± 10.4/77.7 ± 8.1 Blood flow at measure point was shown tobe parabolic, Reynolds number was <1000
Oyre et al. [27] 26.4 7/male N.A. N.A. N.A. The method used for estimating the meanWSS value applies fundamentalhemodynamic theory using athree-dimensional paraboloid model forfitting of blood velocity data in theboundary layer. The data were obtained bystandard MRI velocity acquisitiontechniques. In-plane pixel resolution of theMRI slices was 0.25 mm2
Samijo et al. [28] 10–60 55/female, 56/male N.A. <6.4 nmol/l/N.A. <140/<90 N.A.Samijo et al. [29] 44 (34–58) 4/female, 9/male 24.5 <6.4 nmol/l/N.A. <140/<90 N.A.Dammers et al. [26] 23.7 ± 3.4 3 female, 7/male 22.2 ± 2.2 Reported to be within normal
ranges121.4 ± 9.2/64.5 ± 5.5 N.A.
Wu et al. [21] 23.9 ± 1.9 10/female, 10/male 22.3 ± 2.4 Reported to be within normalranges
Reported to be withinnormal ranges
Technique uses phase contrast imagingwith 0.0625 mm2 resolution andthree-dimensional paraboloid fitting
Oshinski et al. [30] N.A. 8/male N.A. N.A. N.A. N.A.Tang [33] 20–30 2/female, 3/male N.A. N.A. N.A. In-plane pixel resolution of the MRI slices
was 0.47 mm2
Cheng et al. [19] 40.4 1/female, 7/male 23.0 N.A. N.A. N.A.Oyre et al. [31] N.A. 6 volunteers N.A. N.A. N.A. N.A.Pedersen et al. [32] 24.4 8 volunteers N.A. N.A. N.A. N.A.Kornet et al. [20] 20–59 23 female, 16/male N.A. Reported to be within normal
rangesReported to be withinnormal ranges
Measurements performed 2–3 mm fromthe flow divider
Gaenzer et al. [34] 39.6 ± 5.8 10/male N.A. 192 ± 33 mg/dl/65 ± 17 mg/dl 120 ± 6/N.A. N.A.Silber et al. [35] 27.6 15/female, 9 male 22.3 171 ± 32 mg/dl/62 ± 13 mg/dl Reported to be within
normal rangesN.A.
Mitchell et al. [36] 61 ± 10 1107/female, 938 male 27.5 Total cholesterol/HDLratio = 4.05
Reported to be withinnormal ranges
N.A.

scleros
mbtp6t13lw
cf5ittlootothi(lpaadv
ptvcfp(
idaeamWdbtctDtbTodadclearly differs between certain vascular locations, as the
F
C. Cheng et al. / Athero
edication. An exception to this rule is the study performedy the group of Mitchell et al. that provides values of WSS forhe brachial artery (see Table 1B). This study includes a largeopulation of subjects (N = 2045), with an average age of0 ± 10 years. One has to take into account that from this par-icular population, 13% had prevalent cardiovascular disease,2.5% had diabetes, 45% was diagnosed with hypertension,2.5% took antihypertensive medication, 20.6% took lipid-owering medications, 29.5% took aspirin daily, and 13.5%ere smokers.WSS levels range from 9.5 to 15.0 dyn/cm2 in the common
arotid artery, from 6.2 to 9.3 dyn/cm2 in the suprarenal aorta,rom 3.5 to 8.4 dyn/cm2 in the supraceiliac aorta, from 1.3 to.4 dyn/cm2 in the infrarenal aorta, from 2.9 to 6.5 dyn/cm2
n the common femoral artery, from 3.9 to 4.9 dyn/cm2 inhe superficial femoral artery, from 4.3 to 7.8 dyn/cm2 inhe brachial artery, and from 10.9 to 15.7 dyn/cm2 in theeft anterior descending artery. When calculating the averagef these WSS values per type of artery, it becomes obvi-us that there are clear differences in WSS values betweenhe anatomical locations (Fig. 1), which seem dependentn the distance from the aortic root. The more downstreamhat the vessel is located, the lower is its WSS value. Theighest average values are found in the left anterior descend-ng artery (12.7 dyn/cm2), and the common carotid artery11.6 dyn/cm2). The values in the aorta are dependent on theocation, and seem to decrease downstream from the branch-oints of the renal arteries (7.3 dyn/cm2 in the suprarenalorta to 4.2 dyn/cm2 in the supraceiliac/infrarenal aorta). In
ddition, estimation of the WSS in the fetal descending aortauring the second half of pregnancy demonstrated a meanalue of 22 dyn/cm2 [43]. This exceeded level of WSS mayrrr
ig. 1. Stationary WSS values in the large arterial vessels of conscious non-atheros
is 195 (2007) 225–235 229
rovide a stimulus for outward remodeling of the fetus. Fur-her downstream in the arterial tree, the average of the WSSalues remains around the level of the suprarenal aorta in theommon femoral artery (4.3 dyn/cm2) and in the superficialemoral artery (4.4 dyn/cm2). In the brachial artery, a com-arable value for the WSS can be calculated (5.6 dyn/cm2)Fig. 1).
These data clearly indicate that there is non-uniformityn mean WSS levels in the human vascular system. Theifferences in WSS values could be the result of thepplication of different techniques to measure flow and diam-ter in the separate studies. Recently, two groups becameware of this phenomenon and conducted excellent experi-ents in which the mean WSS and mean shear rate (meanSS = mean shear rate × viscosity) were compared between
ifferent arteries. Wu et al. showed significant differencesetween the superficial femoral artery (3.9 ± 1.8 dyn/cm2),he brachial artery (5.8 ± 3.0 dyn/cm2), and the commonarotid artery (10.0 ± 1.8 dyn/cm2) in one study withinhe same human subject and experimental protocol [21].ammers et al. also showed significant differences between
he common carotid artery (11.5 ± 2.1 dyn/cm2) and therachial artery (4.8 ± 1.5 dyn/cm2) within one study [26].hese observations confirm the concept of non-uniformityf mean WSS in the human arterial tree. The remain-er of the data is summarized in Table 1A. Furthermore,lthough the comparison between several arteries is con-ucted with data from separate studies, the mean WSS
ange values show no overlap (e.g. common carotid artery,ange 9.5–15.0 dyn/cm2 versus common femoral artery,ange 2.9–6.5 dyn/cm2).
clerotic humans. Note that the location determines the actual WSS values.

230C
.Cheng
etal./Atherosclerosis
195(2007)
225–235Table 2Mean shear stress levels in laboratory animalsReference Animals Mean shear stress (dyn/cm2) Mean blood
flow (ml/min)Mean velocity(cm/s)
Structure Vessel diameter (mm) Technique Anesthesia
Pohl et al. [45] Dogs 4.9 (OD), 9.5 (LD) 99.0 N.A. CFA 4.7 (OD), 3.8 (LD) Electromagnetic flow probe(perivascular)
Nembutal
Serhatlioglu et al. [46] Dogs 15.6 (OD), 19.3 (LD) N.A. 59.3 (systolic) CFA 4.6 (OD), 3.7 (LD) Doppler sonography Awake animalsLie et al. [47] Dogs 5.1 (OD), 10.0 (LD) 79.6 N.A. CFA 4.3 (OD), 3.4 (LD) Electromagnetic flow probe
(perivascular)Nembutal
Kamiya and Togawa [22] Dogs 15.8 (LD) N.A. 19.0 CCA 3.0 (LD) Electromagnetic flow probe(perivascular)
Nembutal
Lee et al. [48] Dogs 46.0 (LD) N.A. 115.0 (systolic) CCA 3.0 (LD) Doppler sonography Awake animalsLee et al. [48] Dogs 44.0 (LD) N.A. 110.0 (systolic) CFA 3.0 (LD) Doppler sonography Awake animalsZderic et al. [64] Rabbits (New Zealand White) 156.8 (LD) N.A. 61.4 (systolic?) CFA 0.94 (LD) Doppler sonography AcepromazineTronc et al. [65] Rabbits (New Zealand White) 19.4 (LD) 22.2 N.A. CCA 1.8 (LD) Electromagnetic flow probe
(perivascular)Nembutal and ketaminehydrochloride
Walpola et al. [66] Rabbits (New Zealand White) 12.2 (LD) 13.9 N.A. CCA 2.0 (LD) Electromagnetic flow probe(perivascular)
Xylazine and Ketamine
Marano et al. [23] Rabbits (New Zealand White) 60.9 (LD) 42.0 N.A. CCA 1.6 (LD) Doppler sonography IsofluraneLangille et al. [67] Rabbits (New Zealand White) 13.8 (OD), 17.3 (LD) N.A. 15 CCA 2.6 (OD), 2.1 (LD) Electromagnetic flow probe
(perivascular)Xylazine and Ketamine
Sho et al. [68] Rabbits (Japanese White) 10.2 (LD, using histomorphometry) ? N.A. CCA ? (LD, using histomorphometry) Electromagnetic flow probe(perivascular)
Xylazine and ketamine,sevoflurane
Masuda et al. [69] Rabbits (Japanese White) 12.0 (OD), 20.9 (LD) 19.4 N.A. CCA 2.1(OD), 1.7 (LD) Electromagnetic flow probe(perivascular)
Xylazine and ketamine,sevoflurane
Lu et al. [70] Rats (Wistar) 53.7 (OD), 102.9 (LD) 5 N.A. CFA 0.78 (OD), 0.62 (LD) Electromagnetic flow probe(perivascular)
Hyponorm and Dormicum
Ross et al. [24] Rats (Sprague–Dawley) 15.0 (assumed OD diameter), 28.8(assumed LD diameter)
1.4 N.A. CFA 0.78 (assumed OD diameter), 0.62(assumed LD diameter)
Electromagnetic flow probe(perivascular)
Pentobarbione sodium
Ibrahim et al. [71]* Rats (GH) 23.6 (OD), 40.8 (LD) 2.8 N.A. CCA 0.88 (OD), 0.70 (LD) Electromagnetic flow probe(perivascular)
Xylazine, Ketamine, andAcepromazine
Ibrahim et al. [71]* Rats (Fisher) 25.7 (OD), 43.3 (LD) 2.4 N.A. CCA 0.82 (OD), 0.66 (LD) Electromagnetic flow probe(perivascular)
Xylazine, Ketamine, andAcepromazine
Ibrahim et al. [71]* Rats (BN, hypertensive) 18.8 (OD), 48.4 (LD) 2.4 N.A. CCA 0.79 (OD), 0.63 (LD) Electromagnetic flow probe(perivascular)
Xylazine, Ketamine, andAcepromazine
Ibrahim et al. [71]* Rats (SHR-SP, hypertensive) 22.4 (OD), 36.8 (LD) 2.7 N.A. CCA 0.90 (OD), 0.72 (LD) Electromagnetic flow probe(perivascular)
Xylazine, Ketamine, andAcepromazine
Ross et al. [24] Rats (Sprague–Dawley) 42.6 (assumed OD diameter), 84.0(assumed LD diameter)
5.7 N.A. CCA 0.88 (assumed OD diameter), 0.70(assumed LD diameter)
Electromagnetic flow probe(perivascular)
Pentobarbione sodium
Miyashiro et al. [72]* Rats (Fisher) 22.1 (OD), 43.0 (LD) 4.2 N.A. CCA 0.99 (OD), 0.79 (LD) Electromagnetic flow probe(perivascular)
Xylazine, Ketamine, andAcepromazine
Miyashiro et al. [72]* Rats (Fisher, juvenile) 28.9 (OD), 56.3 (LD) 2.9 N.A. CCA 0.80 (OD), 0.64 (LD) Electromagnetic flow probe(perivascular)
Xylazine, Ketamine, andAcepromazine
Tohda et al. [73] Rats (Sprague–Dawley) 20.0 (LD, using histomorphometry withcorrections)
2.6 N.A. CCA ? (LD, using histomorphometry withcorrections)
Electromagnetic flow probe(perivascular)
Sodium pentobarbital
Hartley et al. [74] Mice (C57Bl/6J) 36.0 (assumed OD diameter), 45.0(assumed LD diameter) (calculated from15 khz)
N.A. N.A. CCA 0.50 (assumed OD diameter), 0.4(assumed LD diameter)
Electromagnetic flow probe Isolurane
Li et al. [25] Mice (C57Bl/6J) 81.0 (assumed OD diameter), 102.0(assumed LD diameter)
N.A. 34.0 (peak) CCA 0.50 (assumed OD diameter), 0.4(assumed LD diameter)
Electromagnetic flow probe Pentobarbital
Korshunov and Berk [75] Mice (C57Bl/6J) 40.0 (LD, using histomorphometry) 0.5 N.A. CCA ? (LD, using histomorphometry) Electromagnetic flow probe(perivascular)
Xylazine, Ketamine
Castier et al. [76] Mice (C57Bl/6J) 37.0 (LD, using histomorphometry) 0.64 N.A. CCA ? (LD, using histomorphometry) Electromagnetic flow probe Xylazine, KetamineRudic et al. [77] Mice (C57Bl/6J) 31.0 (assumed OD diameter), 60.0
(assumed LD diameter)0.75 N.A. CCA 0.50 (assumed OD diameter), 0.4
(assumed LD diameter)Electromagnetic flow probe Xylazine, Ketamine
Sullivan and Hoying [78] Mice (50% Black Swiss, 50%129 SV)
142.0 (LD, using histomorphometry) 0.75 N.A. CCA 0.30 (LD, using histomorphometry) Electromagnetic flow probe Xylazine, Ketamine
Schiffers et al. [79] Mice (C57Bl/6J, 129 Sv) 14.0 (OD), 28.0 (LD) 0.35 N.A. CCA 0.50 (OD), 0.4 (LD) Ultrasonic probe Xylazine, Ketamine
Common femoral artery (CFA), common carotid artery (CCA), outer diameter vessel (OD), lumen diameter = 80% of outer diameter vessel (LD), systolic velocity (systolic), mean velocity value was used tocalculate mean shear stress value is 25% of systolic velocity, mean of right and left common carotid artery (*).

scleros
2
aaWmItttotrtuWcfaflct
c1atofiCcrta6hbwHth
Fd
itbmFmd9dawaioashcro1mkospafauuvtpli
se
C. Cheng et al. / Athero
.2. Cross-species differences
In Table 2, the mean WSS values in the common carotidrtery and the common femoral artery of different laboratorynimals and of several investigators are compared. All the
SS values are calculated using in vivo data from measure-ents of both lumen and outer diameter of the blood vessel.
f no lumen diameter is presented, it can be calculated fromhe outer diameter as: lumen diameter = 80% of outer diame-er [44], thereby assuming that wall thickness is proportionalo lumen diameter. In rodents, and in rabbits in particular, theuter diameter is often used for WSS calculations, whereashe lumen diameter provides a more correct value for theadius of a vessel. The radius of a vessel is inversely relatedo the WSS. Consequently, recalculations of the WSS valuessing lumen diameter instead of outer diameter yields higherSS values for rabbits, rats and mice. Therefore, further dis-
ussion in this review will be concerning WSS calculatedrom the lumen diameter values only. All values are station-ry WSS values omitting the pulsatile nature of the bloodow. The blood flow and velocity were measured in bothonscious and anesthetized animals, by applying ultrasoundechniques.
In anesthetized animals, WSS levels in the dog commonarotid artery are 15.8 dyn/cm2, and ranged from 10.2 to56.8 dyn/cm2 in rabbits, from 20.0 to 84 dyn/cm2 in rats,nd from 28.0 to 142.0 dyn/cm2 in mice. When calculatinghe average of these WSS values per species in the same typef artery, cross-species differences in mean WSS values areound (Fig. 2), which are 15.8 dyn/cm2 in dogs, 23.3 dyn/cm2
n rabbits, 46.6 dyn/cm2 in rats, and 64.8 dyn/cm2 in mice.omparing these data to the average mean WSS in theommon carotid artery in conscious humans (11.6 dyn/cm2)eveals a higher WSS in smaller species (Fig. 2). In anes-hetized animals, the average WSS in the common femoralrtery is 9.8 dyn/cm2 in dogs, 156.8 dyn/cm2 in rabbits, and5.9 dyn/cm2 in rats, compared to 4.3 dyn/cm2 in consciousumans. Together these data indicate a variation in WSSetween species when the data are ordered per type of artery,
ith a tendency for WSS to increase in smaller animals.owever, when interpreting the collected data we shouldake into account that in contrast to the data provided forumans, the majority of the experiments are conducted in an
ig. 2. Average mean WSS values in the common carotid artery of fiveifferent species.
bfiacmuciaatlooWso
is 195 (2007) 225–235 231
nvasive manner using different anesthetics. Consequently,he data for one species for one type of vessel may varyetween different studies (e.g. WSS ranges for the com-on carotid artery of the rabbit from 10.2 to 156.8 dyn/cm2).or example, comparing the data acquired from the com-on femoral artery and the common carotid artery in
ogs anesthetized with Nembutal (common femoral artery:.5 dyn/cm2, common carotid artery: 15.8 dyn/cm2) with theata from conscious animals (common femoral artery = 19.3nd 44.0 dyn/cm2, common carotid artery = 46.0 dyn/cm2),e found clear indications that the WSS levels are presum-
bly higher in non-anesthetized dogs [22,45–48]. However,t remains unclear if this also holds true for different typesf anesthetics in different laboratory animals, as the effect ofnesthetics on the cardiovascular system can vary betweenpecies. Most of the commonly used anesthetics do have aeart rate lowering effect, which could lead to a reduction inardiac output, thereby lowering the blood flow in the arte-ial tree. In rats, for example, the often used combinationf ketamine–xylazine reduces the heart rate by ∼20% up toh after administration [49]. A recent study conducted inice compared the effect of different anesthetics (isoflurane,
etamine–xylazine and pentobarbital sodium) on the cardiacutput with the hemodynamic condition measured in con-cious animals. The results indicated that the mean arterialressure and the cardiac output were decreased during allnesthetic interventions, with the smallest effects observedor isoflurane and the largest for ketamine–xylazine [50]. Asll mouse studies summarized in this review were carried outsing one of these types of anesthetics, the mean WSS val-es in the conscious mice are likely to be higher than thealues presented in Table 2. One should take into accounthat in Fig. 2, the WSS values in conscious humans are com-ared to anesthetized animals, and that the mean WSS in ateast conscious dogs, rats and mice is presumably higher thanndicated.
Despite these restrictions, a trend was noticed betweenpecies, indicating an effect of body size, in addition to theffect of location on WSS values. As both observations haveeen ignored to a great extent, it is of importance to test thesendings in one study. In our laboratory, mice experimentsre routinely carried out under isoflurane anesthetics. Weonducted a small experiment in which we non-invasivelyeasured the blood velocity in wildtype C57BL/6J mice
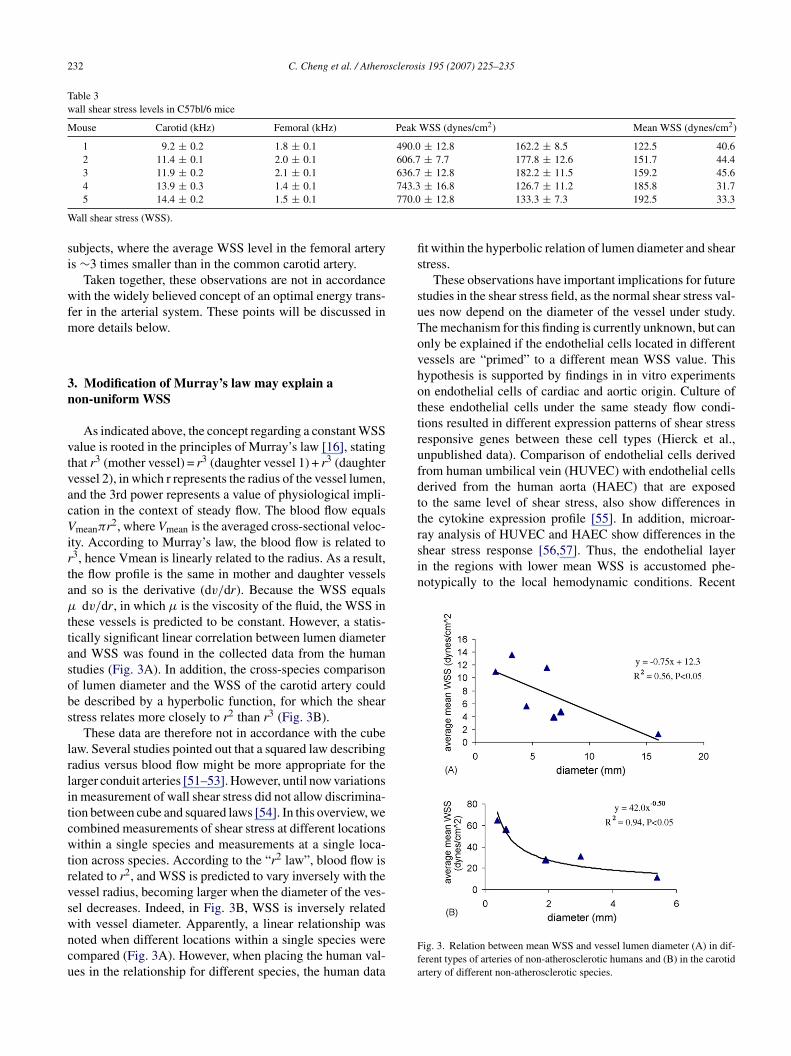
sing a 30-MHz Doppler probe. The femoral artery andommon carotid artery were compared. The measured veloc-ty was subsequently used for WSS calculations, using anssumed luminal diameter for the common carotid arterynd the femoral artery. We observed that under the condi-ions of current experimental protocol: (i) the average WSSevel in mice are indeed much higher than the WSS valuesbserved in larger animals and humans, (ii) the average WSS
f the femoral artery is ∼4 times smaller than the averageSS in the common carotid artery (39.6 ± 2.7 dyn/cm2 ver-us 170.6 ± 14.8 dyn/cm2, respectively) (Table 3). The lastbservation is comparable to the findings in healthy human
232 C. Cheng et al. / Atherosclerosis 195 (2007) 225–235
Table 3wall shear stress levels in C57bl/6 mice
Mouse Carotid (kHz) Femoral (kHz) Peak WSS (dynes/cm2) Mean WSS (dynes/cm2)
1 9.2 ± 0.2 1.8 ± 0.1 490.0 ± 12.8 162.2 ± 8.5 122.5 40.62 11.4 ± 0.1 2.0 ± 0.1 606.7 ± 7.7 177.8 ± 12.6 151.7 44.43 11.9 ± 0.2 2.1 ± 0.1 636.7 ± 12.8 182.2 ± 11.5 159.2 45.6
743.3770.0
W
si
wfm
3n
vtvacVirtaμ
ttasobs
lrlitcwtrvswncu
fis
suTovhottrufdttray analysis of HUVEC and HAEC show differences in theshear stress response [56,57]. Thus, the endothelial layerin the regions with lower mean WSS is accustomed phe-notypically to the local hemodynamic conditions. Recent
4 13.9 ± 0.3 1.4 ± 0.15 14.4 ± 0.2 1.5 ± 0.1
all shear stress (WSS).
ubjects, where the average WSS level in the femoral arterys ∼3 times smaller than in the common carotid artery.
Taken together, these observations are not in accordanceith the widely believed concept of an optimal energy trans-
er in the arterial system. These points will be discussed inore details below.
. Modification of Murray’s law may explain aon-uniform WSS
As indicated above, the concept regarding a constant WSSalue is rooted in the principles of Murray’s law [16], statinghat r3 (mother vessel) = r3 (daughter vessel 1) + r3 (daughteressel 2), in which r represents the radius of the vessel lumen,nd the 3rd power represents a value of physiological impli-ation in the context of steady flow. The blood flow equalsmeanπr2, where Vmean is the averaged cross-sectional veloc-
ty. According to Murray’s law, the blood flow is related to3, hence Vmean is linearly related to the radius. As a result,he flow profile is the same in mother and daughter vesselsnd so is the derivative (dv/dr). Because the WSS equals
dv/dr, in which μ is the viscosity of the fluid, the WSS inhese vessels is predicted to be constant. However, a statis-ically significant linear correlation between lumen diameternd WSS was found in the collected data from the humantudies (Fig. 3A). In addition, the cross-species comparisonf lumen diameter and the WSS of the carotid artery coulde described by a hyperbolic function, for which the sheartress relates more closely to r2 than r3 (Fig. 3B).
These data are therefore not in accordance with the cubeaw. Several studies pointed out that a squared law describingadius versus blood flow might be more appropriate for thearger conduit arteries [51–53]. However, until now variationsn measurement of wall shear stress did not allow discrimina-ion between cube and squared laws [54]. In this overview, weombined measurements of shear stress at different locationsithin a single species and measurements at a single loca-
ion across species. According to the “r2 law”, blood flow iselated to r2, and WSS is predicted to vary inversely with theessel radius, becoming larger when the diameter of the ves-el decreases. Indeed, in Fig. 3B, WSS is inversely related
ith vessel diameter. Apparently, a linear relationship wasoted when different locations within a single species wereompared (Fig. 3A). However, when placing the human val-es in the relationship for different species, the human dataFfa
± 16.8 126.7 ± 11.2 185.8 31.7± 12.8 133.3 ± 7.3 192.5 33.3
t within the hyperbolic relation of lumen diameter and sheartress.
These observations have important implications for futuretudies in the shear stress field, as the normal shear stress val-es now depend on the diameter of the vessel under study.he mechanism for this finding is currently unknown, but cannly be explained if the endothelial cells located in differentessels are “primed” to a different mean WSS value. Thisypothesis is supported by findings in in vitro experimentsn endothelial cells of cardiac and aortic origin. Culture ofhese endothelial cells under the same steady flow condi-ions resulted in different expression patterns of shear stressesponsive genes between these cell types (Hierck et al.,npublished data). Comparison of endothelial cells derivedrom human umbilical vein (HUVEC) with endothelial cellserived from the human aorta (HAEC) that are exposedo the same level of shear stress, also show differences inhe cytokine expression profile [55]. In addition, microar-
ig. 3. Relation between mean WSS and vessel lumen diameter (A) in dif-erent types of arteries of non-atherosclerotic humans and (B) in the carotidrtery of different non-atherosclerotic species.

scleros
pmsaPlsldeatticsedive
4
DPtsoibsitaavaTmItst
vvstsmiti
5
tWaoWel
A
dt
R
[
[
[
[
C. Cheng et al. / Athero
ublications have identified endothelial shear stress sensingechanisms that can alter their capacity by adjusting their
hear stress sensors in response to WSS (e.g. hyaluroniccid glycosaminoglycans in the glycocalyx [58,59], andECAM-1 [60,61]). The endothelium may be attuned to
ocal WSS conditions by specific modification of these sheartress sensors. The priming of the endothelial cells to theocal WSS conditions may take place during endothelial cellevelopment. In adult endothelial cells, WSS regulates genexpression by inducing epigenetic modification of histonesnd activation of transcription complexes bearing acetyl-ransferase activity [62]. In a recent study, it was shownhat WSS could also epigenetically modify histones andnfluence cell differentiation in mouse embryonic stem (ES)ells, committing them to become cardiovascular precur-ors [63]. It is interesting to hypothesize that perhaps duringmbryonic development, the geometry in the vasculatureetermines the fate of these vascular precursors by prim-ng them to respond athero-protectively to the local WSSalue, resulting in the adaptation of the subsequently formedndothelium.
. Limitations of the assessed studies
Estimation of the magnitude of shear stress based onoppler measurements indeed relies on the validity ofoiseuille’s law. This law is valid under a number of assump-
ions, including rigid vessel walls, sufficient inlet length,tationary Newtonian flow, and excluding bends/bifurcationsf the vessel. These parameters have an effect on the veloc-ty, and therefore the shear stress, distribution in arteries. Allut one of these parameters were investigated in an excellenteries of studies by the group of Hoeks. They measured veloc-ty profiles in the common carotid artery, and they showed thathe velocity profiles follow the Womersley profiles reason-bly well. This implies that, although momentary velocitynd shear stress patterns may deviate from the steady flowalues based on Poiseuille’s law, the time averaged valuesre approximated fairly well by the steady flow assumptions.he one parameter that was not studied in the straight com-on carotid arteries, is the effect of geometrical variation.
ndeed, the presence of bends and branching points may leado local differences in shear stress that can be large. As theseites are not the territory we are interested in, we still believehat the observations in this review are valid.
Anesthetics and exercise may have an effect on the WSSalues in Tables 1A and 2. Therefore, we only included WSSalues of the human studies that were measured in a con-cious, resting state in a lying down position in order to keephe WSS values comparable between the different humantudies. The type of anesthetics used in the animal experi-
ents was also reported. However, the WSS values presentedn this paper depend on the mobile state and anatomical posi-ion of the subjects and could deviate from the WSS valuesn different conditions (e.g. during exercise).
[
[
is 195 (2007) 225–235 233
. Conclusion
In summary, by comparing the data provided from litera-ure we have shown that: (i) the assumption of a physiological
SS level of ∼15 dyn/cm2 for all the straight vessels in therterial tree is incorrect; (ii) WSS is not constant through-ut the vascular tree; (iii) WSS varies between species; (iv)SS is approximately inversely related to the vessel diam-
ter. These data support a “r2 law” rather than Murray’s r3
aw for larger vessels in the arterial tree.
cknowledgements
This work was supported by The Netherlands Heart Foun-ation (NHS), grant 2002T045, and by the grant supplied byhe translational research program of the Erasmus MC.
eferences
[1] Hogers B, DeRuiter MC, Gittenberger-de Groot AC, Poelmann RE.Unilateral vitelline vein ligation alters intracardiac blood flow patternsand morphogenesis in the chick embryo. Circ Res 1997;80:473–81.
[2] Hove JR, Koster RW, Forouhar AS, Acevedo-Bolton G, Fraser SE,Gharib M. Intracardiac fluid forces are an essential epigenetic factorfor embryonic cardiogenesis. Nature 2003;421:172–7.
[3] Gimbrone Jr MA, Topper JN, Nagel T, Anderson KR, Garcia-CardenaG. Endothelial dysfunction, hemodynamic forces, and atherogenesis.Ann N Y Acad Sci 2000;902:230–9 [discussion 239–240].
[4] Cheng C, Tempel D, van Haperen R, et al. Atherosclerotic lesion sizeand vulnerability are determined by patterns of fluid shear stress. Cir-culation 2006;113:2744–53.
[5] Malek AM, Alper SL, Izumo S. Hemodynamic shear stress and its rolein atherosclerosis. JAMA 1999;282:2035–42.
[6] LaBarbera M. Principles of design of fluid transport systems in zoology.Science 1990;249:992–1000.
[7] Girerd X, London G, Boutouyrie P, Mourad JJ, Safar M, Laurent S.Remodeling of the radial artery in response to a chronic increase inshear stress. Hypertension 1996;27:799–803.
[8] Kamiya A, Bukhari R, Togawa T. Adaptive regulation of wallshear stress optimizing vascular tree function. Bull Math Biol1984;46:127–37.
[9] Rossitti S, Lofgren J. Vascular dimensions of the cerebral arteries followthe principle of minimum work. Stroke 1993;24:371–7.
10] Gnasso A, Carallo C, Irace C, et al. Association between wall shearstress and flow-mediated vasodilation in healthy men. Atherosclerosis2001;156:171–6.
11] Jiang Y, Kohara K, Hiwada K. Low wall shear stress contributes toatherosclerosis of the carotid artery in hypertensive patients. HypertensRes 1999;22:203–7.
12] Joannides R, Bizet-Nafeh C, Costentin A, et al. Chronic ACEinhibition enhances the endothelial control of arterial mechanicsand flow-dependent vasodilatation in heart failure. Hypertension2001;38:1446–50.
13] Taber LA, Ng S, Quesnel AM, Whatman J, Carmen CJ. InvestigatingMurray’s law in the chick embryo. J Biomech 2001;34:121–4.
14] Taber LA. An optimization principle for vascular radius including theeffects of smooth muscle tone. Biophys J 1998;74:109–14.
15] Murray C. The physiological principle of minimum work. I. The vas-cular system and the cost of blood volume. Proc Natl Acad Sci USA1926;12:207–14.

2 scleros
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
34 C. Cheng et al. / Athero
16] Murray C. The physiological principle of minimum work applied tothe angle of the branching of arteries. J Gen Physiol 1926;9:835–41.
17] Zamir M. Shear forces and blood vessel radii in the cardiovascularsystem. J Gen Physiol 1977;69:449–61.
18] Gnasso A, Carallo C, Irace C, et al. Association between intima-mediathickness and wall shear stress in common carotid arteries in healthymale subjects. Circulation 1996;94:3257–62.
19] Cheng CP, Herfkens RJ, Taylor CA. Abdominal aortic hemody-namic conditions in healthy subjects aged 50–70 at rest and duringlower limb exercise: in vivo quantification using MRI. Atherosclerosis2003;168:323–31.
20] Kornet L, Hoeks AP, Lambregts J, Reneman RS. Mean wall shear stressin the femoral arterial bifurcation is low and independent of age at rest.J Vasc Res 2000;37:112–22.
21] Wu SP, Ringgaard S, Oyre S, et al. Wall shear rates differ between thenormal carotid, femoral, and brachial arteries: an in vivo MRI study. JMagn Reson Imaging 2004;19:188–93.
22] Kamiya A, Togawa T. Adaptive regulation of wall shear stress to flowchange in the canine carotid artery. Am J Physiol 1980;239:H14–21.
23] Marano G, Palazzesi S, Vergari A, Ferrari AU. Protection by shear stressfrom collar-induced intimal thickening: role of nitric oxide. ArteriosclerThromb Vasc Biol 1999;19:2609–14.
24] Ross G, White FN, Brown AW, Kolin A. Regional blood flow in therat. J Appl Physiol 1966;21:1273–5.
25] Li YH, Reddy AK, Taffet GE, et al. Doppler evaluation of peripheralvascular adaptations to transverse aortic banding in mice. UltrasoundMed Biol 2003;29:1281–9.
26] Dammers R, Stifft F, Tordoir JH, et al. Shear stress depends on vascularterritory: comparison between common carotid and brachial artery. JAppl Physiol 2003;94:485–9.
27] Oyre S, Ringgaard S, Kozerke S, et al. Accurate noninvasive quanti-tation of blood flow, cross-sectional lumen vessel area and wall shearstress by three-dimensional paraboloid modeling of magnetic resonanceimaging velocity data. J Am Coll Cardiol 1998;32:128–34.
28] Samijo SK, Willigers JM, Barkhuysen R, et al. Wall shear stress in thehuman common carotid artery as function of age and gender. CardiovascRes 1998;39:515–22.
29] Samijo SK, Barkhuysen R, Willigers JM, et al. Wall shear stress assess-ment in the common carotid artery of end-stage renal failure patients.Nephron 2002;92:557–63.
30] Oshinski JN, Ku DN, Mukundan Jr S, Loth F, Pettigrew RI. Determina-tion of wall shear stress in the aorta with the use of MR phase velocitymapping. J Magn Reson Imaging 1995;5:640–7.
31] Oyre S, Pedersen EM, Ringgaard S, Boesiger P, Paaske WP. In vivowall shear stress measured by magnetic resonance velocity mappingin the normal human abdominal aorta. Eur J Vasc Endovasc Surg1997;13:263–71.
32] Pedersen EM, Oyre S, Agerbaek M, et al. Distribution of earlyatherosclerotic lesions in the human abdominal aorta correlates withwall shear stresses measured in vivo. Eur J Vasc Endovasc Surg1999;18:328–33.
33] Tang BT. Abdominal aortic hemodynamics in young healthy adults atrest and during lower limp exercise: quantification using image-basedcomputer modelling, in review.
34] Gaenzer H, Neumayr G, Marschang P, et al. Flow-mediated vasodi-lation of the femoral and brachial artery induced by exercise inhealthy nonsmoking and smoking men. J Am Coll Cardiol 2001;38:1313–9.
35] Silber HA, Ouyang P, Bluemke DA, et al. Why is flow-mediated dilationdependent on arterial size? Assessment of the shear stimulus usingphase-contrast magnetic resonance imaging. Am J Physiol Heart CircPhysiol 2005;288:H822–8.
36] Mitchell GF, Parise H, Vita JA, et al. Local shear stress and brachialartery flow-mediated dilation: the Framingham Heart Study. Hyperten-sion 2004;44:134–9.
37] Matsuda K, Teragawa H, Fukuda Y, et al. Response of the left ante-rior descending coronary artery to acetylcholine in patients with chest
[
is 195 (2007) 225–235
pain and angiographically normal coronary arteries. Am J Cardiol2003;92:1394–8.
38] Fukuda Y, Teragawa H, Matsuda K, Yamagata T, Matsuura H, ChayamaK. Tetrahydrobiopterin restores endothelial function of coronary arter-ies in patients with hypercholesterolaemia. Heart 2002;87:264–9.
39] Eleuteri E, Scapellato F, Temporelli PL, Giannuzzi P. Evaluation ofthe left anterior descending coronary artery flow velocity by transtho-racic echo-Doppler without contrast enhancement. Ital Heart J 2002;3:520–4.
40] Liepsch D. An introduction to biofluid mechanics–basic models andapplications. J Biomech 2002;35:415–35.
41] Dammers R, Tordoir JH, Hameleers JM, Kitslaar PJ, Hoeks AP.Brachial artery shear stress is independent of gender or age and doesnot modify vessel wall mechanical properties. Ultrasound Med Biol2002;28:1015–22.
42] Long Q, Xu XY, Ariff B, Thom SA, Hughes AD, Stanton AV.Reconstruction of blood flow patterns in a human carotid bifur-cation: a combined CFD and MRI study. J Magn Reson Imaging2000;11:299–311.
43] Struijk PC, Stewart PA, Fernando KL, et al. Wall shear stressand related hemodynamic parameters in the fetal descending aortaderived from color Doppler velocity profiles. Ultrasound Med Biol2005;31:1441–50.
44] Bots ML, Grobbee DE, Hofman A, Witteman JC. Common carotidintima-media thickness and risk of acute myocardial infarction: therole of lumen diameter. Stroke 2005;36:762–7.
45] Pohl U, Holtz J, Busse R, Bassenge E. Crucial role of endotheliumin the vasodilator response to increased flow in vivo. Hypertension1986;8:37–44.
46] Serhatlioglu S, Kiris A, Kocakoc E, Canpolat I, Bozgeyik Z, Han MC.Evaluation of the effects of sildenafil citrate (Viagra) on canine renalartery, carotid and aortic blood flow with the aid of color Dopplersonography. Urol Int 2003;71:103–7.
47] Lie M, Sejersted OM, Kiil F. Local regulation of vascular cross sec-tion during changes in femoral arterial blood flow in dogs. Circ Res1970;27:727–37.
48] Lee K, Choi M, Yoon J, Jung J. Spectral waveform analysis of majorarteries in conscious dogs by Doppler ultrasonography. Vet RadiolUltrasound 2004;45:166–71.
49] Sumitra M, Manikandan P, Rao KV, Nayeem M, Manohar BM,Puvanakrishnan R. Cardiorespiratory effects of diazepam-ketamine,xylazine-ketamine and thiopentone anesthesia in male Wistar rats—acomparative analysis. Life Sci 2004;75:1887–96.
50] Janssen BJ, De Celle T, Debets JJ, Brouns AE, Callahan MF, SmithTL. Effects of anesthetics on systemic hemodynamics in mice. Am JPhysiol Heart Circ Physiol 2004;287:H1618–24.
51] Uylings HB. Optimization of diameters and bifurcation angles in lungand vascular tree structures. Bull Math Biol 1977;39:509–20.
52] Gafiychuk VV, Lubashevsky IA. On the principles of the vascularnetwork branching. J Theor Biol 2001;212:1–9.
53] Zamir M, Sinclair P, Wonnacott TH. Relation between diameterand flow in major branches of the arch of the aorta. J Biomech1992;25:1303–10.
54] Karau KL, Krenz GS, Dawson CA. Branching exponent heterogeneityand wall shear stress distribution in vascular trees. Am J Physiol HeartCirc Physiol 2001;280:H1256–63.
55] Morris TE, Mattox PA, Shipley GD, Wagner CR, Hosenpud JD.The pattern of cytokine messenger RNA expression in human aor-tic endothelial cells is different from that of human umbilical veinendothelial cells. Transpl Immunol 1993;1:137–42.
56] Chen BP, Li YS, Zhao Y, et al. DNA microarray analysis of geneexpression in endothelial cells in response to 24-h shear stress. Physiol
Genomics 2001;7:55–63.57] Garcia-Cardena G, Comander J, Anderson KR, Blackman BR, Gim-brone Jr MA. Biomechanical activation of vascular endothelium asa determinant of its functional phenotype. Proc Natl Acad Sci USA2001;98:4478–85.

scleros
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
C. Cheng et al. / Athero
58] Mochizuki S, Vink H, Hiramatsu O, et al. Role of hyaluronic acidglycosaminoglycans in shear-induced endothelium-derived nitric oxiderelease. Am J Physiol Heart Circ Physiol 2003;285:H722–6.
59] Gouverneur M, Spaan JA, Pannekoek H, Fontijn RD, Vink H. Fluidshear stress stimulates incorporation of hyaluronan into endothelial cellglycocalyx. Am J Physiol Heart Circ Physiol 2006;290:H458–62.
60] Tzima E, Irani-Tehrani M, Kiosses WB, et al. A mechanosensory com-plex that mediates the endothelial cell response to fluid shear stress.Nature 2005;437:426–31.
61] Cheng C, van Haperen R, de Waard M, et al. Shear stress affects theintracellular distribution of eNOS: direct demonstration by a novel invivo technique. Blood 2005;106:3691–8.
62] Illi B, Nanni S, Scopece A, et al. Shear stress-mediated chromatinremodeling provides molecular basis for flow-dependent regulation ofgene expression. Circ Res 2003;93:155–61.
63] Illi B, Scopece A, Nanni S, et al. Epigenetic histone modification andcardiovascular lineage programming in mouse embryonic stem cellsexposed to laminar shear stress. Circ Res 2005;96:501–8.
64] Zderic V, Keshavarzi A, Noble ML, et al. Hemorrhage control in arteriesusing high-intensity focused ultrasound: a survival study. Ultrasonics2006;44:46–53.
65] Tronc F, Wassef M, Esposito B, et al. Role of NO in flow-inducedremodeling of the rabbit common carotid artery. Arterioscler ThrombVasc Biol 1996;16:1256–62.
66] Walpola PL, Gotlieb AI, Langille BL. Monocyte adhesion and changesin endothelial cell number, morphology, and F-actin distribution elicitedby low shear stress in vivo. Am J Pathol 1993;142:1392–400.
67] Langille BL, Bendeck MP, Keeley FW. Adaptations of carotid arteries
of young and mature rabbits to reduced carotid blood flow. Am J Physiol1989;256:H931–9.68] Sho E, Nanjo H, Sho M, et al. Arterial enlargement, tortuosity, andintimal thickening in response to sequential exposure to high and lowwall shear stress. J Vasc Surg 2004;39:601–12.
[
is 195 (2007) 225–235 235
69] Masuda H, Zhuang YJ, Singh TM, et al. Adaptive remodeling of inter-nal elastic lamina and endothelial lining during flow-induced arterialenlargement. Arterioscler Thromb Vasc Biol 1999;19:2298–307.
70] Lu X, Zhao JB, Wang GR, Gregersen H, Kassab GS. Remodeling ofthe zero-stress state of femoral arteries in response to flow overload.Am J Physiol Heart Circ Physiol 2001;280:H1547–59.
71] Ibrahim J, Miyashiro JK, Berk BC. Shear stress is differentially regu-lated among inbred rat strains. Circ Res 2003;92:1001–9.
72] Miyashiro JK, Poppa V, Berk BC. Flow-induced vascular remodelingin the rat carotid artery diminishes with age. Circ Res 1997;81:311–9.
73] Tohda K, Masuda H, Kawamura K, Shozawa T. Difference in dilata-tion between endothelium-preserved and -desquamated segments inthe flow-loaded rat common carotid artery. Arterioscler Thromb1992;12:519–28.
74] Hartley CJ, Reddy AK, Madala S, et al. Hemodynamic changes inapolipoprotein E-knockout mice. Am J Physiol Heart Circ Physiol2000;279:H2326–34.
75] Korshunov VA, Berk BC. Flow-induced vascular remodeling in themouse: a model for carotid intima-media thickening. ArteriosclerThromb Vasc Biol 2003;23:2185–91.
76] Castier Y, Brandes RP, Leseche G, Tedgui A, Lehoux S.p47phox-dependent NADPH oxidase regulates flow-induced vascularremodeling. Circ Res 2005;97:533–40.
77] Rudic RD, Bucci M, Fulton D, Segal SS, Sessa WC. Temporal eventsunderlying arterial remodeling after chronic flow reduction in mice:correlation of structural changes with a deficit in basal nitric oxidesynthesis. Circ Res 2000;86:1160–6.
78] Sullivan CJ, Hoying JB. Flow-dependent remodeling in the carotid
artery of fibroblast growth factor-2 knockout mice. Arterioscler ThrombVasc Biol 2002;22:1100–5.79] Schiffers PM, Henrion D, Boulanger CM, et al. Altered flow-inducedarterial remodeling in vimentin-deficient mice. Arterioscler ThrombVasc Biol 2000;20:611–6.




















