![Patch Antenna[1]](https://static.fdokumen.com/doc/165x107/63158e4cc32ab5e46f0d5c89/patch-antenna1.jpg)
Landscape fragmentation, land-use legacy and propagule pressure promote plant invasion on coastal...
12
1 23 Landscape Ecology ISSN 0921-2973 Volume 29 Number 9 Landscape Ecol (2014) 29:1541-1550 DOI 10.1007/s10980-014-0074-3 Landscape fragmentation, land-use legacy and propagule pressure promote plant invasion on coastal dunes: a patch-based approach Marco Malavasi, Marta Carboni, Maurizio Cutini, Maria L. Carranza & Alicia T. R. Acosta
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Landscape fragmentation, land-use legacy and propagule pressure promote plant invasion on coastal...

1 23
Landscape Ecology ISSN 0921-2973Volume 29Number 9 Landscape Ecol (2014) 29:1541-1550DOI 10.1007/s10980-014-0074-3
Landscape fragmentation, land-use legacyand propagule pressure promote plantinvasion on coastal dunes: a patch-basedapproach
Marco Malavasi, Marta Carboni,Maurizio Cutini, Maria L. Carranza &Alicia T. R. Acosta

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media Dordrecht. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

RESEARCH ARTICLE
Landscape fragmentation, land-use legacy and propagulepressure promote plant invasion on coastal dunes:a patch-based approach
Marco Malavasi • Marta Carboni •
Maurizio Cutini • Maria L. Carranza •
Alicia T. R. Acosta
Received: 3 February 2014 / Accepted: 19 July 2014 / Published online: 6 August 2014
� Springer Science+Business Media Dordrecht 2014
Abstract Coastal dunes and sand areas are reported to
be among the habitats most invaded by alien species in
Europe. Landscape pattern could be a significant driver in
invasion processes in parallel with land-use legacy.
Fragmentation of natural habitats combined with the
availability of propagules from the surrounding matrix
may enhance the invisibility of ecological communities.
Based on multitemporal land cover maps (1954–2008)
and a floristic database, we analyzed how habitat
fragmentation, propagule pressure and land-use legacy
have affected alien plants’ presence and richness on
natural dune patches along the Lazio Coast (Central
Italy). Floristic data were derived from an existing geo-
database of random vegetation plots (64 m2). A set of
landscape patch-based metrics, considered to be adequate
proxies of the main processes affecting alien invasion and
richness, was calculated. First, we fit a generalized linear
model (GLM) with binomial errors to assess which
landscape metrics are influencing patch invasion. Sec-
ond, we extracted invaded patches and, with GLMs, we
investigated how landscape metrics affect average alien
species richness. Alien invasion and alien richness seem
to be affected by different processes: although alien
invasion of each patch is strongly associated with its land-
use legacy, the richness of aliens is more affected by
landscape fragmentation and by the propagule pressure to
which patch is exposed. By integrating spatial and
temporal landscape metrics with floristic data, we were
able to disentangle the relations of landscape fragmen-
tation, propagule pressure and land-use legacy with the
presence and richness of alien plants. The methodolog-
ical approach here adopted could be easily extended to
other alien species and ecosystems, offering scientifically
sound support to prevent the high economic costs derived
from both the control and the eradication of aliens.
Keywords Aliens � Landscape pattern � Land use
change � Mediterranean coastal dunes � Patch-based
metrics � Alien invasion � Alien richness
Introduction
Theoretical studies suggest that the spread of alien
species is closely connected to factors operating at the
Electronic supplementary material The online version ofthis article (doi:10.1007/s10980-014-0074-3) contains supple-mentary material, which is available to authorized users.
M. Malavasi � M. Carboni � M. Cutini � A. T. R. Acosta
Dipartimento di Scienze, Universita degli Studi di Roma
Tre, V.le Marconi 446, 00146 Rome, Italy
Present Address:
M. Carboni
Laboratoire d’Ecologie Alpine, Universite Joseph Fourier/
CNRS Grenoble, BP 53, 2233 Rue de la Piscine,
38041 Grenoble Cedex 9, France
M. L. Carranza (&)
EnviX-Lab. Dipartimento di Bioscienze e Territorio,
Universita degli Studi del Molise, C.da Fonte Lappone,
86090 Pesche, IS, Italy
e-mail: [email protected]
123
Landscape Ecol (2014) 29:1541–1550
DOI 10.1007/s10980-014-0074-3
Author's personal copy

landscape level (With 2002, 2004). There is no doubt
that landscape patterns could be significant drivers of
invasion processes (Higgins et al. 1999; Hobbs 2000;
Zechmeister et al. 2003; Essl and Dirnbock 2008;
Gonzalez-Moreno et al. 2013) in parallel with histor-
ical factors such as land use and the time of
introduction (Pysek and Hulme 2005). Surprisingly,
despite a large number of contributions focusing on
various aspects of the colonization, spread and control
of alien species in numerous habitats (Pysek and
Hulme 2005; Richardson and Pysek 2008), few studies
have addressed these issues through a landscape
approach (With 2002; Bruno et al. 2004; Vila and
Ibanez 2011; Thomas and Moloney 2013) and even
fewer have taken into account a historical component
or land-use legacy (Domenech et al. 2005; Kuhman
et al. 2010; Mattingly and Orrock 2013).
Invasive plants have the ability to thrive and spread
aggressively outside their native range. In particular,
the landscape fragmentation of natural habitats may
directly enhance the invasibility of ecological com-
munities (Hobbs 2000). Landscape ‘‘fragmentation’’
(Forman and Godron 1986) entails the reduction of the
extent of natural habitats and the isolation and decline
of the ecological quality of the remaining fragments.
The resulting fragmented pattern has the potential to
boost habitat colonization by alien species from the
surrounding matrix (Carranza et al. 2010; Vila and
Ibanez 2011). Among the agents of fragmentation,
urban sprawl may have consistent effects on alien
invasion by increasing the availability of alien prop-
agules, ready to move from artificial areas to adjacent
natural communities (e.g., O’Shea and Kirkpatrick
2000; McKinney 2006; Thuiller et al. 2006). Last but
not least, the vulnerability of a landscape to invasion
depends not only on its current spatial pattern but also
on the local land-use history and legacy (Vila et al.
2003; Vila and Ibanez 2011). In fact, it is well known
that land-use changes can alter ecosystems, providing
opportunities for exotic species to invade due to the
increased movement of seeds (Hobbs and Huenneke
1992; Trombulak and Frissell 2000). Historical pro-
cesses and unique events profoundly influence local
community structure (Ricklefs 1987), and the time and
circumstances of the introduction of alien species may
play a particularly central role in invasion success
(Lonsdale 1999).
Coastal dune ecosystems are increasingly threa-
tened by direct and indirect human pressures
(Schlacher et al. 2008) and have been reported to
harbor many alien species (Bruno et al. 2004; Dark
2004; Chytry et al. 2008). In Europe, coastal dune and
sand areas (EUNIS B1) (European Environment
Agency 2008) are among the most invaded habitats
by alien species (Chytry et al. 2008). In particular,
during the last decades, beach fruition by tourists, the
expansion of urban areas and the spread of agriculture
and afforestation activities have strongly shaped
Mediterranean coastal landscapes (Alados et al.
2004; Hesp and Martinez 2007; Malavasi et al.
2013), consistently modifying land use and their
spatial patterns (Carboni et al. 2009; Drius et al.
2013). Furthermore, humans have been responsible
not only for landscape transformation but also for
transporting species beyond their native ranges. The
arrival of an exotic species to a landscape generally
results from the intentional or accidental introduction
by humans, which involves overcoming geographical
barriers (With 2004; Carboni et al. 2010b). Although
different coastal dune habitats are known to undergo
different levels of invasion (Carboni et al. 2010a;
Carranza et al. 2011; Chytry et al. 2008), to our
knowledge no studies have considered the role of both
the spatial and the temporal pattern of the landscape in
influencing alien invasions.
In consideration of all the above, the aim of this
work is to investigate whether and how spatial
landscape characteristics and landscape legacy may
affect fine-scale alien plant invasion on natural
Mediterranean coastal dune habitats. In particular,
based on multitemporal land cover maps and on a
floristic database, we propose to investigate the effects
of habitat fragmentation, propagule pressure and land-
use legacy on alien plant invasion and alien richness in
natural dune patches.
Methods
Study area
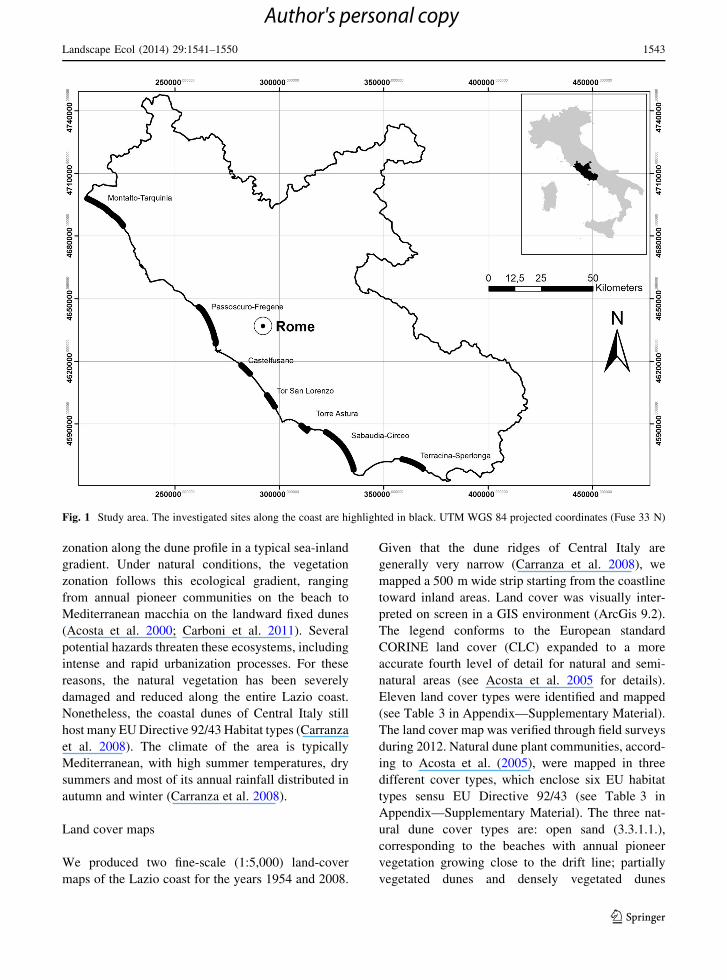
The study was carried out on the Tyrrhenian coast of
Central Italy and focused on recent coastal dunes
(Holocene). The area includes seven study sites along
the coast of the Lazio region (Fig. 1), with recent
dunes harboring remnant natural vegetation patches
along the seashore. Along the sea-inland ecotone,
abiotic conditions vary greatly, shaping habitat
1542 Landscape Ecol (2014) 29:1541–1550
123
Author's personal copy
zonation along the dune profile in a typical sea-inland
gradient. Under natural conditions, the vegetation
zonation follows this ecological gradient, ranging
from annual pioneer communities on the beach to
Mediterranean macchia on the landward fixed dunes
(Acosta et al. 2000; Carboni et al. 2011). Several
potential hazards threaten these ecosystems, including
intense and rapid urbanization processes. For these
reasons, the natural vegetation has been severely
damaged and reduced along the entire Lazio coast.
Nonetheless, the coastal dunes of Central Italy still
host many EU Directive 92/43 Habitat types (Carranza
et al. 2008). The climate of the area is typically
Mediterranean, with high summer temperatures, dry
summers and most of its annual rainfall distributed in
autumn and winter (Carranza et al. 2008).
Land cover maps
We produced two fine-scale (1:5,000) land-cover
maps of the Lazio coast for the years 1954 and 2008.
Given that the dune ridges of Central Italy are
generally very narrow (Carranza et al. 2008), we
mapped a 500 m wide strip starting from the coastline
toward inland areas. Land cover was visually inter-
preted on screen in a GIS environment (ArcGis 9.2).
The legend conforms to the European standard
CORINE land cover (CLC) expanded to a more
accurate fourth level of detail for natural and semi-
natural areas (see Acosta et al. 2005 for details).
Eleven land cover types were identified and mapped
(see Table 3 in Appendix—Supplementary Material).
The land cover map was verified through field surveys
during 2012. Natural dune plant communities, accord-
ing to Acosta et al. (2005), were mapped in three
different cover types, which enclose six EU habitat
types sensu EU Directive 92/43 (see Table 3 in
Appendix—Supplementary Material). The three nat-
ural dune cover types are: open sand (3.3.1.1.),
corresponding to the beaches with annual pioneer
vegetation growing close to the drift line; partially
vegetated dunes and densely vegetated dunes
Fig. 1 Study area. The investigated sites along the coast are highlighted in black. UTM WGS 84 projected coordinates (Fuse 33 N)
Landscape Ecol (2014) 29:1541–1550 1543
123
Author's personal copy

(3.3.1.2.), corresponding to the herbaceous dune
vegetation growing on fore dunes; and Mediterranean
macchia (3.2.3.1.), corresponding to the woody dune
vegetation growing on back dunes (Malavasi et al.
2013). To make the maps comparable, we interpreted
the year 1954 using the backward editing technique.
On the basis of the most recent map (2008), the other
time level was processed in reverse order by editing,
with information being adapted to the older time level
(Walz 2008).
Floristic data
Floristic data of the natural dune patches were derived
from an existing geo-database of random vegetation
plots performed on the study area during the spring in
the years 2005–2010 (Santoro et al. 2012). Plots were
geographically positioned with 3 m of accuracy using
a GPS. For each 8 m by 8 m (64 m2) plot, the list of all
vascular plant species (including aliens) was com-
piled. A total of 570 plots were available with 158
species recorded. Among these, eight are alien species,
identified according to Celesti-Grapow et al. (2009).
In this work, alien species are defined as plant taxa in a
given area whose presence is due to intentional or
unintentional human involvement (Celesti-Grapow
et al. 2009).
Landscape data
We overlaid the georeferenced plots and the landscape
maps, selecting only those coastal dune patches that
overlapped with at least two floristic plots. In this way,
we obtained a total of 186 patches containing random
plots used for further analyses. For each coastal dune
patch, we calculated a set of landscape metrics as
proxies for three main processes affecting alien
invasion and richness at the landscape level: fragmen-
tation, propagule pressure and land-use change.
Although, in some cases, the same patterns may result
from multiple processes, here we assigned each metric
only to the principal process to which it has been linked
in previous studies and ecological theory (Forman
1995; Carboni et al. 2009; Carranza et al. 2010).
Landscape fragmentation
For each natural dune patch, we calculated two
parameters that have been proven to be useful in
describing the spatial pattern of coastal landscapes
(Carboni et al. 2009; Carranza et al. 2010): patch size
(PS) and shape index (SI) (Table 1). PS is a good
parameter for describing the effects of human distur-
bance on coastal dune habitats: the smaller the size, the
higher the ongoing fragmentation process (Carboni
et al. 2009). We chose SI because of its power to
discriminate between long irregularly shaped patches,
characteristic of well-preserved coastal dune habitats,
and regular round patches, which often dominate in
disturbed and fragmented coastal dunes (Forman
1995; Carranza et al. 2010). Moreover, we calculated
the Euclidean Nearest Neighbor (ENN) based on the
shortest edge-to-edge distance. This parameter is
commonly used to infer the degree of patch isolation
and the level of fragmentation in many different
landscape contexts (Forman 1995).
Table 1 Landscape parameters calculated for each patch and
used as independent variables
Metric Description Process
PS (ha) Size of the focal patch Landscape
fragmentationSI
(adim.)
Shape Index of the focal patch
ENN
(m)
Euclidean Nearest Neighbor to
the nearest neighboring patch
of the same type based on
shortest edge-to-edge distance
SB_Art
(%)
Shared border with artificial
cover types (ART)
Propagule
pressure
SB_Sn
(%)
Shared border with semi-natural
cover types (WET, SHV,
SWV, REF, BFM)
SB_Du
(%)
Shared border with the other
dune cover types (BPV, HDV,
WDV)
DC_Art
(m)
Centroid-to-centroid distance
from the nearest artificial
polygon (ART)
DC_Sn
(m)
Centroid-to-centroid distance
from the nearest semi-natural
polygon (WET, SHV, SWV,
REF, BFM)
DC_Du
(m)
Centroid-to-centroid distance
from the nearest natural dune
polygon (BPV, HDV, WDV)
Durb (m) Difference between 1954 and
2006 distance to the artificial
nearest neighboring patch
based on shortest edge-to-edge
distance
Land use
legacy
1544 Landscape Ecol (2014) 29:1541–1550
123
Author's personal copy

Propagule pressure
As proxies of propagule pressure, we use the spatial
relationships of each patch with adjacent categories
applying boundary-based metrics (Rescia et al. 1994)
and the distance to other polygons (Table 1).
The analysis of shared borders, which allows us to
describe the contiguity among patches (SB_Art:
shared borders with artificial surfaces, SB_Sn: shared
borders with semi-natural cover types, SB_Du: shared
borders with other natural dune habitats), has been
previously used for assessing the conservation status
of coastal dune landscapes (Acosta et al. 2000;
Carboni et al. 2009; Drius et al. 2013). A positive
spatial association of dune cover types with artificial
surfaces is expected for anthropogenic landscapes
(Carranza et al. 2010), leading to higher levels of
propagule pressure in the adjacent natural communi-
ties (e.g., O’Shea and Kirkpatrick 2000; McKinney
2006; Thuiller et al. 2006). On the other hand,
although highly connected natural dune cover types
could guarantee dune integrity in landscapes with low
human pressure (Drius et al. 2013), in highly human
transformed sites, the series of adjacent natural
patches could facilitate the movement of aliens across
the landscape (Alofs and Fowler 2010).
The spatial position of natural dune cover types was
assessed by computing the centroid-to-centroid min-
imum Euclidean distance (DC—in meters) between
each natural dune patch and the patches of other cover
types (Table 1). This metric (DC_Art: distance from
artificial areas; DC_Sn: distance from semi-natural
cover types; DC_Du: distance from other natural dune
habitats) gives information about the proximity of
each patch to urban nuclei and/or to other natural dune
areas and is used here for the first time on coastal dune
mosaics. A shorter distance to urban patches could
enhance alien invasion by increasing the probability of
success of alien propagules, which move from artifi-
cial areas to surrounding natural communities (Thu-
iller et al. 2006). In addition, the proximity of invaded
semi-natural patches may facilitate alien establish-
ment elsewhere in the landscape (Pauchard and
Alaback 2004).
Land use legacy
Finally, we took the local history of each patch into
account by quantifying temporal urbanization trends
in the surrounding areas. Coastal areas have been
mostly shaped by urban expansion in most of the
Mediterranean countries (Hesp and Martinez 2007;
Malavasi et al. 2013). Thus, we selected the urbani-
zation process as a proxy of the overall historical land-
use legacy. First, we measured the shortest edge-to-
edge distance (in meters) between each focal natural
dune patch and artificial patches for the years 1954 and
2006. Then, for each patch, we computed the differ-
ence between these edge-to-edge distances (1954 -
2006) and obtained a Durb value (Table 1). Durb, which
expresses the urbanization trends in the surrounding
areas of each natural patch, is equal to 0 when the
distance between natural dunes and urban patches
across the last 60 years has remained stable, and it
increases as artificial areas expanded closer to the
natural patches (i.e., when natural and artificial areas
became closer).
Statistical analyses
For each natural dune patch, we calculated two
response variables: the presence of at least one
invaded plot within the patch (binary variable: invaded
or not) and the average number of aliens recorded in
the floristic plots within it (quantitative variable:
average alien species richness). In this way, alien
species richness is here considered at the plot level
(mean alien richness of all plots recorded in that
patch). Then, linear models were fit for each response
variable (alien invasion and alien richness) using
different landscape metrics of fragmentation (patch
size, shape index, and Euclidean Nearest Neighbor),
propagule pressure (shared borders and minimum
Euclidean distance to other cover types) and land-use
legacy (temporal variation of the distance to artificial
areas) as independent variables (Table 1). In addition,
by including the geographical coordinates of each
patch (x and y of the centroid) in the model, we also
accounted for spatial autocorrelation among patches.
In addition, the land cover type of each patch was
included in the models to accommodate the fact that
different coastal dune habitats are known to have
different levels of invasibility (Carboni et al. 2010a;
Chytry et al. 2008; Carranza et al. 2011).
First, we fit a generalized linear model (GLM) with
binomial errors to assess which landscape metric
influences patch invasion by alien plants. Second, we
extracted invaded patches and investigated how
Landscape Ecol (2014) 29:1541–1550 1545
123
Author's personal copy

landscape metrics affect the average alien species
richness by using GLMs.
All landscape metrics were log transformed
(excluding the shared borders values), and variance
inflation factor (VIF) analysis was used to reduce
collinearity among all the explanatory variables (R
package ‘‘car’’). Model simplification was performed
by implementing stepwise model selection (forward
and backward) based on the Akaike Information
Criterion (AIC). Finally, correlograms were used to
evaluate any remaining spatial dependencies among
the residuals from the various models (R package
‘‘spdep’’).
All modeling analyses were conducted in the
R-statistical environment (version 2.13.2; R Founda-
tion for Statistical Computing, Vienna, Austria. http://
www.R-project.org.).
Results
Alien invasion
The final model explains approximately 30 % of the
total variance (R2 = 0.28) (Table 2), suggesting that
the selected landscape characteristics can indeed
explain a large amount of variability in patch invasion
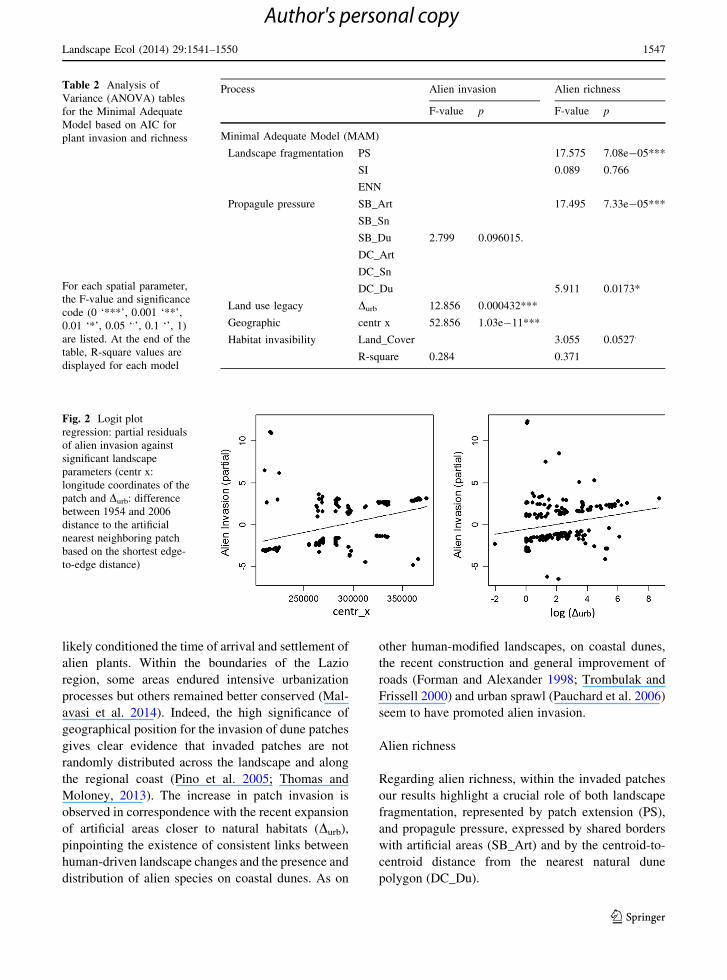
patterns. We found a significant relationship between
patch alien invasion and historic factors, expressed by
land-use legacy (Durb) and by the geographic position
of each patch (centr_x) (Table 2). The higher the
recent development of artificial areas in the surround-
ing areas of the patch (Durb; p \ 0.001), the higher the
probability of invasion (Fig. 2). The geographical
coordinates were also retained in the model
(p \ 0.001), suggesting that the geographical position
of the patches in the study area also play an important
role (Fig. 2). Moreover, the minimum adequate model
based on AIC retains shared borders with other dune
cover types (SB_Du) (Table 2). However, the F-test in
the ANOVA table (Table 2) highlights that this last
term was not significant and should be omitted from a
more conservative model (Zuur et al. 2007).
The correlogram showed that no spatial depen-
dency was left among the residuals of the model (see
Fig. 4 in Appendix—Supplementary Material). Given
that geographical coordinates were included in the
model, the absence of spatial dependency in the
residuals was expected.
Alien richness
The minimal adequate model for alien plant richness
within the invaded patches explains almost 40 % of
the total variance (R2 = 0.37) (Table 2). The model
suggests that patch extension (PS; p \ 0.001) and the
length of the shared borders with artificial cover types
(SB_Art; p \ 0.001) are negatively and positively
correlated with alien richness, respectively. The
smaller the patch size, the higher the average number
of alien species in the plots (Fig. 3); the higher the
amount of patch border shared with artificial areas, the
higher the average number of alien species in the plots
within the patch (Fig. 3). The minimal model for alien
richness based on AIC also included the centroid-to-
centroid distance from the nearest natural dune
polygon (DC_Du; p = 0.0173), the Shape Index (SI;
p = 0.766) and the land cover category (Land_Cover;
p = 0.0527) to which the patch belongs (Table 2).
However, F-tests showed that DC_Du, SI and Land_-
Cover were not significant terms and therefore should
be omitted from the final model (Zuur et al. 2007).
Correlograms highlighted limited spatial depen-
dency in both the original response variable (Alien
richness) and the residuals of the model but only when
considering first lag neighbors (see Fig. 5 in Appen-
dix—Supplementary Material). Indeed, the geograph-
ical coordinates were excluded from the minimal
adequate model.
Discussion
Based on our results, we can affirm that the three
analyzed processes affect alien plant invasion within
Mediterranean dune patches at different stages.
Although the presence of alien plants is strongly
related to land-use legacy, alien richness mainly
develops from fragmentation and propagule pressure
forces.
Alien invasion
We found that alien invasion is associated with the
land-use legacy of each patch and with its geograph-
ical position along the coast. The Italian coast has
experienced contrasting histories of human develop-
ment and landscape transformation over the last
60 years (Malavasi et al. 2013), which have most
1546 Landscape Ecol (2014) 29:1541–1550
123
Author's personal copy
likely conditioned the time of arrival and settlement of
alien plants. Within the boundaries of the Lazio
region, some areas endured intensive urbanization
processes but others remained better conserved (Mal-
avasi et al. 2014). Indeed, the high significance of
geographical position for the invasion of dune patches
gives clear evidence that invaded patches are not
randomly distributed across the landscape and along
the regional coast (Pino et al. 2005; Thomas and
Moloney, 2013). The increase in patch invasion is
observed in correspondence with the recent expansion
of artificial areas closer to natural habitats (Durb),
pinpointing the existence of consistent links between
human-driven landscape changes and the presence and
distribution of alien species on coastal dunes. As on
other human-modified landscapes, on coastal dunes,
the recent construction and general improvement of
roads (Forman and Alexander 1998; Trombulak and
Frissell 2000) and urban sprawl (Pauchard et al. 2006)
seem to have promoted alien invasion.
Alien richness
Regarding alien richness, within the invaded patches
our results highlight a crucial role of both landscape
fragmentation, represented by patch extension (PS),
and propagule pressure, expressed by shared borders
with artificial areas (SB_Art) and by the centroid-to-
centroid distance from the nearest natural dune
polygon (DC_Du).
Fig. 2 Logit plot
regression: partial residuals
of alien invasion against
significant landscape
parameters (centr x:
longitude coordinates of the
patch and Durb: difference
between 1954 and 2006
distance to the artificial
nearest neighboring patch
based on the shortest edge-
to-edge distance)
Table 2 Analysis of
Variance (ANOVA) tables
for the Minimal Adequate
Model based on AIC for
plant invasion and richness
For each spatial parameter,
the F-value and significance
code (0 ‘***’, 0.001 ‘**’,
0.01 ‘*’, 0.05 ‘.’, 0.1 ‘’, 1)
are listed. At the end of the
table, R-square values are
displayed for each model
Process Alien invasion Alien richness
F-value p F-value p
Minimal Adequate Model (MAM)
Landscape fragmentation PS 17.575 7.08e-05***
SI 0.089 0.766
ENN
Propagule pressure SB_Art 17.495 7.33e-05***
SB_Sn
SB_Du 2.799 0.096015.
DC_Art
DC_Sn
DC_Du 5.911 0.0173*
Land use legacy Durb 12.856 0.000432***
Geographic centr x 52.856 1.03e-11***
Habitat invasibility Land_Cover 3.055 0.0527.
R-square 0.284 0.371
Landscape Ecol (2014) 29:1541–1550 1547
123
Author's personal copy
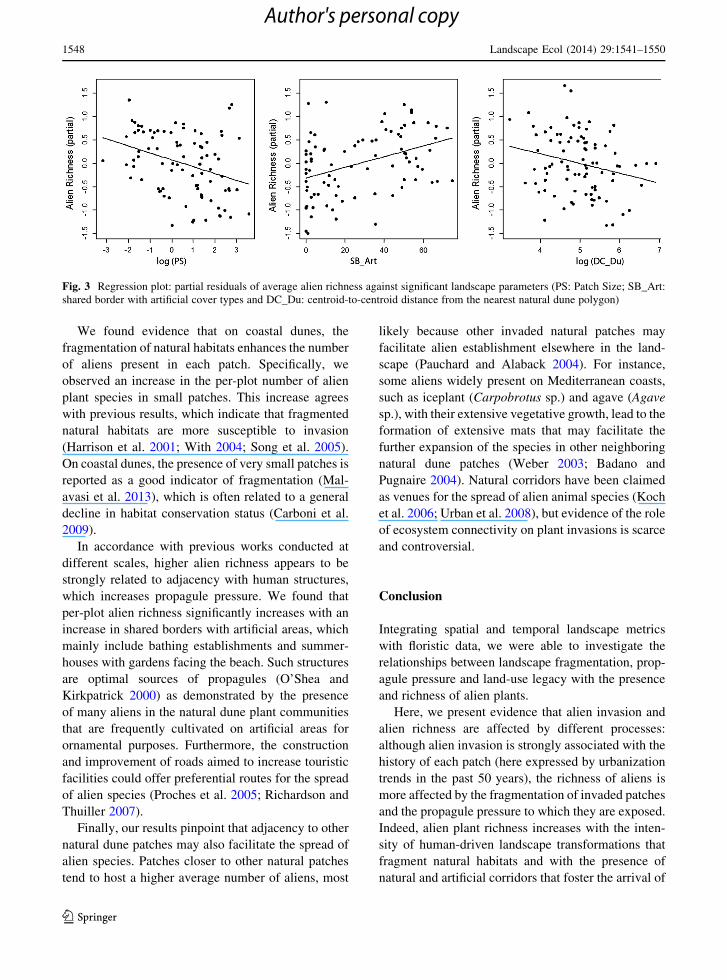
We found evidence that on coastal dunes, the
fragmentation of natural habitats enhances the number
of aliens present in each patch. Specifically, we
observed an increase in the per-plot number of alien
plant species in small patches. This increase agrees
with previous results, which indicate that fragmented
natural habitats are more susceptible to invasion
(Harrison et al. 2001; With 2004; Song et al. 2005).
On coastal dunes, the presence of very small patches is
reported as a good indicator of fragmentation (Mal-
avasi et al. 2013), which is often related to a general
decline in habitat conservation status (Carboni et al.
2009).
In accordance with previous works conducted at
different scales, higher alien richness appears to be
strongly related to adjacency with human structures,
which increases propagule pressure. We found that
per-plot alien richness significantly increases with an
increase in shared borders with artificial areas, which
mainly include bathing establishments and summer-
houses with gardens facing the beach. Such structures
are optimal sources of propagules (O’Shea and
Kirkpatrick 2000) as demonstrated by the presence
of many aliens in the natural dune plant communities
that are frequently cultivated on artificial areas for
ornamental purposes. Furthermore, the construction
and improvement of roads aimed to increase touristic
facilities could offer preferential routes for the spread
of alien species (Proches et al. 2005; Richardson and
Thuiller 2007).
Finally, our results pinpoint that adjacency to other
natural dune patches may also facilitate the spread of
alien species. Patches closer to other natural patches
tend to host a higher average number of aliens, most
likely because other invaded natural patches may
facilitate alien establishment elsewhere in the land-
scape (Pauchard and Alaback 2004). For instance,
some aliens widely present on Mediterranean coasts,
such as iceplant (Carpobrotus sp.) and agave (Agave
sp.), with their extensive vegetative growth, lead to the
formation of extensive mats that may facilitate the
further expansion of the species in other neighboring
natural dune patches (Weber 2003; Badano and
Pugnaire 2004). Natural corridors have been claimed
as venues for the spread of alien animal species (Koch
et al. 2006; Urban et al. 2008), but evidence of the role
of ecosystem connectivity on plant invasions is scarce
and controversial.
Conclusion
Integrating spatial and temporal landscape metrics
with floristic data, we were able to investigate the
relationships between landscape fragmentation, prop-
agule pressure and land-use legacy with the presence
and richness of alien plants.
Here, we present evidence that alien invasion and
alien richness are affected by different processes:
although alien invasion is strongly associated with the
history of each patch (here expressed by urbanization
trends in the past 50 years), the richness of aliens is
more affected by the fragmentation of invaded patches
and the propagule pressure to which they are exposed.
Indeed, alien plant richness increases with the inten-
sity of human-driven landscape transformations that
fragment natural habitats and with the presence of
natural and artificial corridors that foster the arrival of
Fig. 3 Regression plot: partial residuals of average alien richness against significant landscape parameters (PS: Patch Size; SB_Art:
shared border with artificial cover types and DC_Du: centroid-to-centroid distance from the nearest natural dune polygon)
1548 Landscape Ecol (2014) 29:1541–1550
123
Author's personal copy

new propagules. Thus, spatial and temporal landscape
patterns and the processes that drive alien invasions
seem to be inherently linked, and our understanding of
the spread and richness of alien species will be
substantially improved by considering them
simultaneously.
Finally, from a practical point of view, our results
offer a scientifically sound basis for orienting the
management and risk analysis of biological invasions.
They can help identify the most effective strategies for
sustainable management, prevent the high economic
costs derived from both the control and eradication of
aliens and, ultimately, prevent the loss of natural
mosaics and associated ecological services.
Acknowledgements This work was partially funded by
ENVEUROPE project (LIFE08 ENV/IT/000339).
References
Acosta ATR, Blasi C, Stanisci A (2000) Spatial connectivity and
boundary patterns in coastal dune vegetation in the Circeo
National Park, Central Italy. J Veg Sci 11:149–154
Acosta ATR, Carranza ML, Izzi CF (2005) Combining land
cover mapping of coastal dunes with vegetation analyses.
Appl Veg Sci 8:133–138
Alados CL, Pueyo Y, Barrantes O, Escos J, Giner L, Robles AB
(2004) Variations in landscape patterns and vegetation
cover between 1957 and 1994 in a semiarid Mediterranean
ecosystem. Landsc Ecol 19:543–559
Alofs KM, Fowler NL (2010) Habitat fragmentation caused by
woody plant encroachment inhibits the spread of an inva-
sive grass. J Appl Ecol 47:338–347
Badano IE, Pugnaire FI (2004) Invasion of Agave species
(Agavaceae) in south-east Spain: invader demographic
parameters and impacts on native species. Divers Distrib
10:493–500
Bruno JF, Kennedy CW, Rand TA, Grant MB (2004) Land-
scape-scale patterns of biological invasions in shoreline
plant communities. Oikos 107:531–540
Carboni M, Carranza ML, Acosta ATR (2009) Assessing con-
servation status on coastal dunes: a multiscale approach.
Landscape Urban Plan 91:17–25
Carboni M, Santoro R, Acosta ATR (2010a) Are some com-
munities of the coastal dune zonation more susceptible to
alien plant invasion? J Plant Ecol 3:139–147
Carboni M, Thuiller W, Izzi F, Acosta ATR (2010b) Disen-
tangling the relative effects of environmental versus human
factors on the abundance of native and alien plant species
in Mediterranean sandy shores. Divers Distrib 16:537–546
Carboni M, Santoro R, Acosta ATR (2011) Dealing with scarce
data to understand how environmental gradients and
propagule pressure shape fine-scale alien distribution pat-
terns on coastal dunes. J Veg Sci 22:751–765
Carranza ML, Acosta ATR, Stanisci A, Pirone G, Ciaschetti G
(2008) Ecosystem classification for EU habitat distribution
assessment in sandy coastal environments. Environ Monit
Assess 140:99–107
Carranza ML, Carboni M, Feola S, Acosta ATR (2010) Landscape-
scale patterns of alien plant species on coastal dunes. The case
of iceplant in central Italy. Appl Veg Sci 13:135–145
Carranza ML, Ricotta C, Carboni M, Acosta ATR (2011)
Habitat selection by invasive alien plants: a bootstrap
approach. Preslia 83:529–536
Celesti-Grapow L, Alessandrini A, Arrigoni PV, Banfi E, Ber-
nardo L, Bovio M, Brundu G, Cagiotti MR, Camarda I,
Carli E, Conti F, Fascetti S, Galasso G, Gubellini L, La
Valva V, Lucchese F, Marchiori S, Mazzola P, Peccenini S,
Poldini L, Pretto F, Prosser F, Siniscalco C, Villani MC,
Viegi L, Wilhalm T, Blasi C (2009) Inventory of the non-
native flora of Italy. Plant Biosyst 143:386–430
Chytry M, Maskell LC, Pino J, Pysek P, Vila M, Font X, Smart
SM (2008) Habitat invasions by alien plants: a quantitative
comparison among Mediterranean, subcontinental and
oceanic regions of Europe. J Appl Ecol 45:448–458
Dark SJ (2004) The biogeography of invasive alien plants in
California: an application of GIS and spatial regression
analysis. Divers Distrib 10:1–9
Domenech R, Vila M, Pino J (2005) Historical land-use legacy
and Cortaderia selloana invasion in the Mediterranean
region. Glob Change Biol 11:1054–1064
Drius M, Malavasi M, Santoro R, Acosta ATR, Ricotta C,
Carranza ML (2013) Boundary based analysis for the
assessment of coastal dune landscape integrity over time.
Appl Geog 45:41–48
Essl F, Dirnbock T (2008) Diversity of native and alien vascular
plants of dry grasslands in Central Europe. Appl Veg Sci
11:441–450
European Environment Agency (2008) European Nature
Information System (EUNIS) Habitats Classification.
Copenhagen, Denmark. Available from http://eunis.eea.
europa.eu/habitats.jsp. accessed Dec 2013
Forman RTT (1995) Land mosaics: the ecology of landscapes
and regions. Cambridge University Press, Cambridge
Forman RTT, Alexander LE (1998) Roads and their major
ecological effects. Annu Rev Ecol Syst 29:207–231
Forman RTT, Godron M (1986) Landscape Ecology. John
Wiley & Sons, New York
Gonzalez-Moreno P, Pino J, Carreras D, Basnou C, Fernandez-
Rebolalr I, Montserrat V (2013) Quantifying the landscape
influence on plant invasion in Mediterranean coastal hab-
itats. Landscape Ecol 28:891–903
Harrison S, Rice K, Maron J (2001) Habitat patchiness promotes
invasion by alien grasses on serpentine soil. Biol Cons
100:45–53
Hesp PA, Martinez ML (2007) Disturbance processes and
dynamics in coastal dunes. In: Johnson EA and Miyanishi
K (eds). Plant disturbance ecology. The process and the
response. Academic Press, London, pp 215-247
HigginsSI, Richardson DM,Cowling RM,Trinder-Smith TH (1999)
Predicting the landscape-scale distribution of alien plants and
their threat to plant diversity. Conserv Biol 12:303–313
Hobbs RJ (2000) Land use changes and invasion. In: Mooney
HA, Hobbs RJ (eds) Invasive species in a changing world.
Island Press, Washington, pp 55–64
Landscape Ecol (2014) 29:1541–1550 1549
123
Author's personal copy

Hobbs RJ, Huenneke LF (1992) Disturbance, diversity, and
invasion: implications for conservation. Conserv Biol
6:324–337
Koch FH, Cheshire HM, Devine HA (2006) Landscape-scale
prediction of hemlock woolly adelgid, Adelges tsugae
(Homoptera: adelgidae), infestation in the southern
Appalachian Mountains. Environ Entomol 35:1313–1323
Kuhman TR, Pearson SM, Turner MG (2010) Effects of land use
history and the contemporary landscape on non-native
plant invasion at local and regional scales in the forest-
dominated southern Appalachians. Landscape Ecol 25:
1433–1445
Lonsdale WM (1999) Global patterns of invasions and the
concept of invisibility. Ecology 80:223–228
Malavasi M, Santoro R, Cutini M, Acosta ATR, Carranza ML
(2013) What has happened to coastal dunes in the last half
century? A multitemporal coastal landscape analysis in
Central Italy. Landsc Urb Plan 119:54–63
Malavasi M, Santoro R, Cutini M, Acosta ATR (2014) Carranza
ML (2014) The impact of human pressure on landscape
patterns and plant species richness in Mediterranean coastal
dunes. Plant Biosyst. doi:10.1080/11263504.913730
Mattingly WB, Orrock JL (2013) Historic land use influences
contemporary establishment on invasive plant species.
Oecologia 172:1147–1157
McKinney ML (2006) Urbanization as a major cause of biotic
homogenization. Biol Conserv 127:247–260
O’Shea EM, Kirkpatrick JB (2000) The impact of suburbani-
zation on remnant coastal vegetation in Hobart, Tasmania.
Appl Veg Sci 3:243–252
Pauchard A, Alaback PB (2004) Influence of elevation, landuse,
and landscape context on patterns of alien plant invasions
along roadsides in protected areas of South- Central Chile.
Conserv Biol 18:238–248
Pauchard A, Aguayo M, Pena E, Urrutia R (2006) Multiple
effects of urbanization on the biodiversity of developing
countries: the case of a fast-growing metropolitan area
(Concepcion, Chile). Biol Cons 127:272–281
Pino J, Font X, Carbo J, Jove M, Pallares L (2005) Large-scale
correlates of alien plant invasion in Catalonia (NE of
Spain). Biol Cons 122:339–350
Proches S, Wilson JRU, Veldtman R, Kalwij JM, Richardson
DM, Chown SL (2005) Landscape corridors: possible
dangers? Science 310:779
Pysek P, Hulme PE (2005) Spatio-temporal dynamics of plant
invasions: linking pattern to process. Ecoscience. 12:302–315
Rescia AJ, Schmitz MF, Martind de Agar P, De Pablo CL, At-
auri JA, Pineda FD (1994) Influence of landscape com-
plexity and land management on woody plant diversity in
Northern Spain. J Veg Sci 5:505–516
Richardson DM, Pysek P (2008) Fifty years of invasion ecology—
the legacy of Charles Elton. Divers Distrib 14:161–168
Richardson DM, Thuiller W (2007) Home away from home—
objective mapping of high-risk source areas for plant
introductions. Divers Distrib 13:299–312
Ricklefs RE (1987) Community Diversity: relative roles of local
and regional processes. Science 235:167–171
Santoro R, Jucker T, Carboni M, Acosta ATR (2012) Patterns of
plant community assembly in invaded and non-invaded
communities along a natural environmental gradient. J Veg
Sci 23:483–494
Schlacher TA, Schoeman DS, Dugan J, Lastra M, Jones A,
Scapini F, McLachlan A (2008) Sandy beach ecosystems:
key features, sampling issues, management challenges and
climate change impacts. Marine Ecol 29:70–90
Song IJ, Hong SK, Kim HO, Byun B, Gin Y (2005) The pattern
of landscape patches and invasion of naturalized plants in
developed areas of urban Seoul. Landscape Urban Plan
70:205–219
Thomas SM, Moloney KA (2013) Hierarchical factors impact-
ing the distribution of an invasive species: landscape con-
text and propagule pressure. Landscape Ecol 28:81–93
Thuiller W, Richardson DM, Rouget M, Proches S, Wilson RU
(2006) Interactions between environment, species traits,
and human uses describe patterns of plant invasions.
Ecology 87:1755–1769
Trombulak SC, Frissell CA (2000) Review of ecological effects
of roads on terrestrial and aquatic communities. Conserv
Biol 14:18–30
Urban MC, Phillips BL, Skelly DK, Shine R (2008) A toad more
travelled: the heterogeneous invasion dynamics of cane
toads in Australia. Am Nat 171:134–148
Vila M, Ibanez I (2011) Plant invasions in the landscape.
Landscape Ecol 26:461–472
Vila M, Burriel JA, Pino J, Chamizo J, Llach E, Porterias M,
Vives M (2003) Association between Opuntia spp. Inva-
sion and changes in land-cover in the Mediterranean
region. Glob Change Biol 9:1234–1239
Walz U (2008) Monitoring of landscape change and functions in
Saxony (Eastern Germany)—Methods and indicators. Ecol
Indic 8:807–817
Weber E (2003) Invasive plant species of the world: a reference
guide to environmental weeds. CABI Publishing, Cambridge
With KA (2002) The landscape ecology of invasive spread.
Conserv Biol 16:1192–1203
With KA (2004) Assessing the risk of invasive spread in frag-
mented landscape. Risk Anal 24:803–815
Zechmeister HG, Schmitzberger I, Steurer B, Peterseil J, Wrbka
T (2003) The influence of land-use practices and eco-
nomics on plant species richness in meadows. Biol Cons
114:165–177
Zuur AF, Ieno EN, Smith GM (2007) Analysing ecological data.
Springer-Verlag, New York
1550 Landscape Ecol (2014) 29:1541–1550
123
Author's personal copy




















