
Keys for and morphological character variation in some Egyptian cultivars of Cucurbitaceae
14
1 23 Genetic Resources and Crop Evolution An International Journal ISSN 0925-9864 Genet Resour Crop Evol DOI 10.1007/s10722-012-9924-5 Keys for and morphological character variation in some Egyptian cultivars of Cucurbitaceae Sami Rabei, Reda M. Rizk & Abdel- Hamid A. Khedr
Transcript of Keys for and morphological character variation in some Egyptian cultivars of Cucurbitaceae

1 23
Genetic Resources and CropEvolutionAn International Journal ISSN 0925-9864 Genet Resour Crop EvolDOI 10.1007/s10722-012-9924-5
Keys for and morphological charactervariation in some Egyptian cultivars ofCucurbitaceae
Sami Rabei, Reda M. Rizk & Abdel-Hamid A. Khedr

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media Dordrecht. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

RESEARCH ARTICLE
Keys for and morphological character variation in someEgyptian cultivars of Cucurbitaceae
Sami Rabei • Reda M. Rizk •
Abdel-Hamid A. Khedr
Received: 7 May 2012 / Accepted: 10 October 2012
� Springer Science+Business Media Dordrecht 2012
Abstract The cultivated varieties and landraces of
Cucurbitaceae growing in Egypt are presented, 27
taxa, belonging to three genera, six species, and five
subspecies. These characters are arranged according to
their usefulness for identifications as follow: fruit
characters, seed characters and trichome type. Fruit
characters are a good taxonomic tool at varietal level
when combined with the other vegetative characters.
Two keys to 27 cultivated varieties of Cucurbitaceae
(Cucumis, Cucurbita and Luffa) were constructed
using the DELTA software system. One key is used for
identification, and then the other is used as a confir-
matory key. Those keys were built using 36 morpho-
logical characters include vegetative, floral, fruit and
seed characters. There is a significant correlation
between seed size and growth parameters. The growth
parameters of the studied taxa included leaf length/
size, petiole length and corolla length. There is a
highly positive, significant correlation between seed
volume and leaf size (r = 0.81831, P B 0.001), seed
volume and petiole length (r = 0.79112, P B 0.001),
seed volume and leaf length (r = 0.83566, P B 0.001),
and seed volume and corolla length (r = 0.59108,
P B 0.001).
Keywords Computer generating key � Cucumis �Cucurbita �DELTA �Growth parameters � Landraces �Luffa
Introduction
Cucurbitaceae is a family of 118–122 genera and 900
species (Simpson 2010) of monoecious or dioecious
herbs and erect shrubs. The Cucurbitaceae has world
wide distribution, but occur mostly in tropical regions
and are poorly represented in temperate regions.
The family includes economically important food
crops such as Citrullus lanatus (Thunb.) Matsum. &
Nakai (watermelon), Cucumis melo L. (melons),
Cucumis sativus L. (cucumber), Cucurbita pepo
L. and other species (squash, pumpkins). The dried
fruit of several species are used as gourds, those of
Luffa L. (louf) are used as sponges, and some taxa have
medicinal uses. According to Jeffrey (1990), Cucur-
bitaceae is subdivided into two well-defined subfam-
ilies viz.: Zanonioideae and Cucurbitoideae, and eight
tribes representing varying degrees of circumscriptive
cohesiveness. The subfamily Cucurbitoideae has the
most important cucurbit crops, such as Cucumis,
Cucurbita, and Luffa.
S. Rabei (&) � A.-H. A. Khedr
Department of Botany, Faculty of Science,
Damietta University, P.O. Box 34517, Damietta, Egypt
e-mail: [email protected]
R. M. Rizk
Herbarium, National Gene Bank (NGB),
Agricultural Research Center, Giza, Egypt
123
Genet Resour Crop Evol
DOI 10.1007/s10722-012-9924-5
Author's personal copy

The Cucurbitaceae are distinctive in being mostly
monoecious vines with simple, palmately veined and/
or lobed leaves, usually with tendrils; female flowers
are epigynous with parietal placentation and three
carpels; the fruit is a berry, pepo, capsule or samara.
The genus Cucumis is divided into subgenus
Cucumis, containing C. sativus, and subgenus Melo
(Mill.) C. Jeffrey, containing C. melo. The subgenera
are widely separated, to the point that they have been
proposed as two distinct genera (Jeffrey 1980).
Willdenow (1805) recognized 5 cultivated species
of Cucumis. While Naudin (1859) grouped Willde-
now’s species as synonyms under C. melo and divided
the species into 10 entities. Kurz (1877) grouped
C. melo, C. dudaim, C. chate and C. flexuosus into one
group.
Cucumis pubescens Willd. received the name
C. melo L. var. pubescens (Willd.) Kurz, while
Cogniaux and Harms (1924) grouped C. melo,
C. dudaim, C. chate, C. pubescens and C. flexuosus
into C. melo and divided it into two varieties viz.: var.
melo and var. agrestis Naud. A recent monographic
treatment (Kirkbride 1993) recognized 32 species,
including two major groups, cucumbers (C. sativus)
and melons (C. melo).
Linnaeus (1753) used various references, including
herbals, to describe C. pepo. However, the diversity of
cultivars caused him to define some C. pepo varieties
as distinct species. Early classifications described the
cultivars in greater detail (Naudin 1856; Goff 1888),
and Alefeld (1866) considered these varieties to
belong to C. pepo. Primarily on the basis of fruit
morphology, he defined eight varietal groups includ-
ing 66 cultivars.
Castetter (1925) divided edible C. pepo cultivars
into six horticultural groups according to differences
in fruit shape, color, and size. Decker (1988) proposed
a new classification of C. pepo for North and South
America and divided the species into two subspecies
viz.: subsp. pepo and subsp. ovifera (L.) Decker. Paris
(1986) described eight groups of C. pepo edible
cultivars.
Pitrat et al. (2000) gave some notes on infraspecific
classification of cultivars of melon, they adopted the
system of Jeffrey (1980) and classified C. melo into
two subgenera including 16 varieties, five of which
assigned to subsp. agrestis and 11 to subsp. melo.
Nesom (2011) gave an account on certain species and
species complex of Citrullus, Cucumis and Cucurbita
in attempt to apply consistent taxonomic ranking to
wild and related domesticated forms.
Hassib (1938) reported fourteen genera of Cucur-
bitaceae in Egypt, which included wild and cultivated
as well as ornamental species. Citrullus and Cucumis
included both cultivated and wild species, while Luffa
and Cucurbita included only cultivated species.
Montasir and Hassib (1956) recorded seven genera
including both cultivated and wild taxa, and Rizk
(2001) studied cucurbit cultivars and landraces grow-
ing in Egypt.
Tackholm (1974) reported the presence of seven
genera and 11 wild species in Egypt. According to
Boulos (2000) Cucurbitaceae is represented in Egypt
by 8 genera and 13 species, while Heneidaq (2008)
recognized 13 wild species belonging to 9 genera of
Cucurbitaceae in Egypt. In his survey of Cucurbita-
ceae in Egypt, Hassib (1938) reported three varieties
of C. pepo (var. ovifera (L.) Sickenb., var. alba (Harz)
Sickenb. and var. typica Sickenb.)
Identification is the process of determining the
taxon to which a specimen belongs; several methods
are available as aids for this process (Pankhurst 1991).
The most important are conventional identification
keys (Dallwitz et al. 2002). DELTA is a modern
software system for managing taxonomic research
(Dallwitz 2009). DELTA is a collection of sophisti-
cated and powerful data-base programs which stores
morphological data for export in a number of different
formats. In this way it acts as a system for management
of taxonomic data which can be used on a local,
regional or world wide basis to make identifications of
the biota available for whomever needs to use them
(Coleman et al. 2010).
The aim of the present work is to update the
scientific names of the present day cultivated
varieties and landraces of Cucurbitaceae in Egypt.
It also aims to record, identify and collect the
available characters to be used for computer gener-
ation of keys. The keys will help the expert and
non-expert to correctly identify varieties and land-
races of Cucurbitaceae.
Materials and methods
Viable seeds were collected from ripened fruits
and market dealers in Damietta, Egypt. Table 1
Genet Resour Crop Evol
123
Author's personal copy

gives the reference numbers of the investigated
seeds.
The collected seeds were cultivated in fields at the
El-Karawan area of the Kafer El-Bateikh district,
Damietta Governorate, Egypt. The revised taxa are
arranged after Jeffrey (1990). Herbarium voucher
specimens and clean mature seeds are deposited at the
Department of Botany, Faculty of Science, Damietta
University, Egypt.
Materials for trichome and leaf epidermal investi-
gations were obtained from the cultivated plants, and
were cleared in 1 % warm lactic acid solution before
direct examination and photographing with a light
microscope (LM).
For each taxon, a photograph of seed and LM
photomicrographs of trichomes are provided.
For the seed descriptions, the measurements given
are the mean of twenty-five well-developed seeds with
the standard error (±) of the means (SE). The
terminology of the seed characters follows Stearn
(1973).
Data for the 37 qualitative and quantitative char-
acters listed in the ‘‘Appendix’’ were collected. The
DELTA programs (Dallwitz et al. 2000) were used to
construct the keys for these taxa, and the character data
were coded in DELTA format. The data were
converted using the TOKEY directives file, and the
KEY program was used to generate two different keys
(Dallwitz et al. 2000; Rabei and El-Gazzar 2007;
Coleman et al. 2010; Rabei 2011).
Results and discussion
Certain morphological characters were investigated
for the distinction of the cultivated taxa of Cucur-
bitaceae in Egypt. These characters are arranged
according to their usefulness for identifications as
follow:
Fruit characters
Fruit characters were used by Hassib (1938) to
distinguish Cucurbitaceae taxa at the varietal level.
Recently, Munger and Robinson (1991) used fruit
characters to distinguish between seven horticulturally
important melon groups.
Fruits of C. melo are oblong or slender and curved
or coiled, pubescent or becoming glabrous, and often
melon scented. The fruit of C. sativus is globular to
oblong to short cylindrical, 14–20 9 5–7 cm, prickly
when immature, flesh firm, green, not sweet and with a
cucumber odour.
Table 1 List of investigated taxa
Ref. Taxa
1 C. melo L. subsp. melo ‘Melo’
2 C. melo L. subsp. melo ‘Shahd’
3 C. melo L. subsp. melo ‘Santawi’
4 C. melo L. subsp. melo ‘Santawi Taweel’
5 C. melo L. subsp. melo ‘Senani’
6 C. melo L. subsp. melo ‘Ananas Kheshen’
7 C. melo L. subsp. melo ‘Ananas Naiem’
8 C. melo L. subsp. melo ‘Mangawi’
9 C. melo L. subsp. melo var. aegyptiaca Sickenb.
‘Mansouri’
10 C. melo L. subsp. melo. var. aegyptiaca Sickenb.
11 C. melo L. subsp. melo var. dudaim (L.) Naudin
‘Waraqi’
12 C. melo L. subsp. agrestis (Naudin) Pangalo var. dudaim(L.) Naudin ‘Bassusi’
13 C. melo L. subsp. agrestis (Naudin) Pangalo var. dudaim(L.) Naudin ‘Kisaan El-Assal’
14 C. melo L. subsp. agrestis (Naudin) Pangalo var. dudaim(L.) Naudin ‘Qatta Saidi’
15 C. melo L. subsp. melo var. flexuosus (L.) Naudin
‘Firany’
16 C. melo L. subsp. melo var. flexuosus (L.) Naudin
‘Khadra’
17 C. melo L. subsp. melo var. flexuosus (L.) Naudin
‘Beida’
18 C. melo L. subsp. agrestis (Naudin) Pangalo var. chate(Hasselq.) Sageret ‘Aggour’
19 C. sativus L. var. sativus
20 Cucurbita maxima Duchesne ex Lam. ‘Etampes
Pumpkin’ (Abied)
21 C. maxima Duchesne ex Lam. ‘Spanish Pumpkin’(Ahmar)
22 Cucurbita moschata Duchesne ex Lam. ‘Napple
Squash’
23 C. moschata Duchesne ex Lam.‘Long Neapolitan
Squash’
24 C. pepo L. var. ovifera (L.) Alef. ‘Eskandarani’
25 C. pepo L. var. ovifera (L.) Alef. ‘Baladi Khadra’
26 C. pepo L. var. ovifera (L.) Alef. ‘Baladi Beida’
27 Luffa aegyptiaca Mill.
Nomenclature following Hassib (1938), Pitrat et al. (2000),
Jeffrey (2001) and Nesom (2011)
Genet Resour Crop Evol
123
Author's personal copy
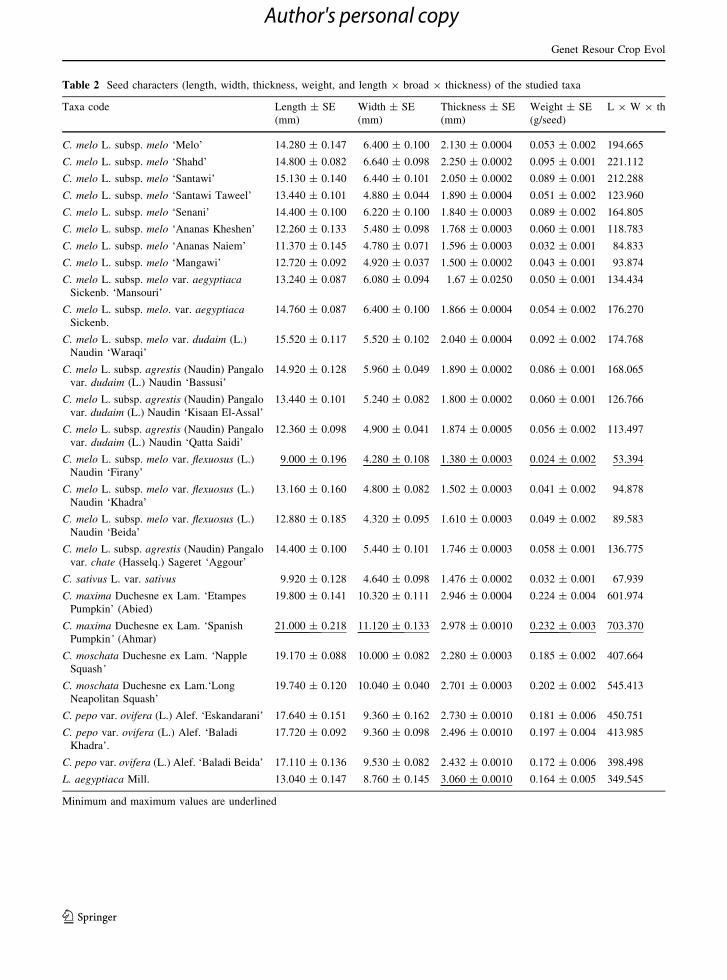
Table 2 Seed characters (length, width, thickness, weight, and length 9 broad 9 thickness) of the studied taxa
Taxa code Length ± SE
(mm)
Width ± SE
(mm)
Thickness ± SE
(mm)
Weight ± SE
(g/seed)
L 9 W 9 th
C. melo L. subsp. melo ‘Melo’ 14.280 ± 0.147 6.400 ± 0.100 2.130 ± 0.0004 0.053 ± 0.002 194.665
C. melo L. subsp. melo ‘Shahd’ 14.800 ± 0.082 6.640 ± 0.098 2.250 ± 0.0002 0.095 ± 0.001 221.112
C. melo L. subsp. melo ‘Santawi’ 15.130 ± 0.140 6.440 ± 0.101 2.050 ± 0.0002 0.089 ± 0.001 212.288
C. melo L. subsp. melo ‘Santawi Taweel’ 13.440 ± 0.101 4.880 ± 0.044 1.890 ± 0.0004 0.051 ± 0.002 123.960
C. melo L. subsp. melo ‘Senani’ 14.400 ± 0.100 6.220 ± 0.100 1.840 ± 0.0003 0.089 ± 0.002 164.805
C. melo L. subsp. melo ‘Ananas Kheshen’ 12.260 ± 0.133 5.480 ± 0.098 1.768 ± 0.0003 0.060 ± 0.001 118.783
C. melo L. subsp. melo ‘Ananas Naiem’ 11.370 ± 0.145 4.780 ± 0.071 1.596 ± 0.0003 0.032 ± 0.001 84.833
C. melo L. subsp. melo ‘Mangawi’ 12.720 ± 0.092 4.920 ± 0.037 1.500 ± 0.0002 0.043 ± 0.001 93.874
C. melo L. subsp. melo var. aegyptiacaSickenb. ‘Mansouri’
13.240 ± 0.087 6.080 ± 0.094 1.67 ± 0.0250 0.050 ± 0.001 134.434
C. melo L. subsp. melo. var. aegyptiacaSickenb.
14.760 ± 0.087 6.400 ± 0.100 1.866 ± 0.0004 0.054 ± 0.002 176.270
C. melo L. subsp. melo var. dudaim (L.)
Naudin ‘Waraqi’
15.520 ± 0.117 5.520 ± 0.102 2.040 ± 0.0004 0.092 ± 0.002 174.768
C. melo L. subsp. agrestis (Naudin) Pangalo
var. dudaim (L.) Naudin ‘Bassusi’
14.920 ± 0.128 5.960 ± 0.049 1.890 ± 0.0002 0.086 ± 0.001 168.065
C. melo L. subsp. agrestis (Naudin) Pangalo
var. dudaim (L.) Naudin ‘Kisaan El-Assal’
13.440 ± 0.101 5.240 ± 0.082 1.800 ± 0.0002 0.060 ± 0.001 126.766
C. melo L. subsp. agrestis (Naudin) Pangalo
var. dudaim (L.) Naudin ‘Qatta Saidi’
12.360 ± 0.098 4.900 ± 0.041 1.874 ± 0.0005 0.056 ± 0.002 113.497
C. melo L. subsp. melo var. flexuosus (L.)
Naudin ‘Firany’
9.000 ± 0.196 4.280 ± 0.108 1.380 ± 0.0003 0.024 ± 0.002 53.394
C. melo L. subsp. melo var. flexuosus (L.)
Naudin ‘Khadra’
13.160 ± 0.160 4.800 ± 0.082 1.502 ± 0.0003 0.041 ± 0.002 94.878
C. melo L. subsp. melo var. flexuosus (L.)
Naudin ‘Beida’
12.880 ± 0.185 4.320 ± 0.095 1.610 ± 0.0003 0.049 ± 0.002 89.583
C. melo L. subsp. agrestis (Naudin) Pangalo
var. chate (Hasselq.) Sageret ‘Aggour’
14.400 ± 0.100 5.440 ± 0.101 1.746 ± 0.0003 0.058 ± 0.001 136.775
C. sativus L. var. sativus 9.920 ± 0.128 4.640 ± 0.098 1.476 ± 0.0002 0.032 ± 0.001 67.939
C. maxima Duchesne ex Lam. ‘Etampes
Pumpkin’ (Abied)
19.800 ± 0.141 10.320 ± 0.111 2.946 ± 0.0004 0.224 ± 0.004 601.974
C. maxima Duchesne ex Lam. ‘Spanish
Pumpkin’ (Ahmar)
21.000 ± 0.218 11.120 ± 0.133 2.978 ± 0.0010 0.232 ± 0.003 703.370
C. moschata Duchesne ex Lam. ‘Napple
Squash’19.170 ± 0.088 10.000 ± 0.082 2.280 ± 0.0003 0.185 ± 0.002 407.664
C. moschata Duchesne ex Lam.‘Long
Neapolitan Squash’
19.740 ± 0.120 10.040 ± 0.040 2.701 ± 0.0003 0.202 ± 0.002 545.413
C. pepo var. ovifera (L.) Alef. ‘Eskandarani’ 17.640 ± 0.151 9.360 ± 0.162 2.730 ± 0.0010 0.181 ± 0.006 450.751
C. pepo var. ovifera (L.) Alef. ‘Baladi
Khadra’.
17.720 ± 0.092 9.360 ± 0.098 2.496 ± 0.0010 0.197 ± 0.004 413.985
C. pepo var. ovifera (L.) Alef. ‘Baladi Beida’ 17.110 ± 0.136 9.530 ± 0.082 2.432 ± 0.0010 0.172 ± 0.006 398.498
L. aegyptiaca Mill. 13.040 ± 0.147 8.760 ± 0.145 3.060 ± 0.0010 0.164 ± 0.005 349.545
Minimum and maximum values are underlined
Genet Resour Crop Evol
123
Author's personal copy
Fruit of C. melo subsp. melo var. flexuosus is
distinguished from the other studied taxa by the
absence of a central cavity, depressed lines with
hairy surface and skin color, which is white, light
green or dark and light green mottled. Fruits of
Luffa aegyptiaca are cylindrical or elongate-subcla-
vate, glabrous, and dry with spongy mesocarp when
ripe.
Seed characters
Mature-seed characters of the studied taxa are sum-
marized in Table 2, and the seeds are shown in Fig. 5.
Seeds of all examined taxa are ovate to ovate oblong in
outline, flattened to compressed, and the funicular
attachment is terminal.
In C. pepo and C. moschata, seeds have a double
rim, while those of C. maxima have a single rim. In L.
aegyptiaca the seed margin is winged, and those of
Cucumis taxa are smooth, without a rim.
Seeds vary in color from yellow to creamy white or
white in taxa of Cucumis and Cucurbita, and the seeds
of L. aegyptiaca are black.
The seed surface is smooth in Cucumis taxa and
dusty in taxa of Cucurbita and Luffa.
The seed apex tapers in C. melo cultivars; is
truncate in C. sativus var. sativus and cultivars of
Cucurbita; and obtuse in seeds of L. aegyptiaca.
The present investigation showed a wide range of
variation and combinations of seed characters. Seeds
of L. aegyptiaca are easily distinguished by their black
Y =-56.63692+1.22927* x , r = 0.81831
0 200 400 600 800
0
200
400
600
800
1000
1 234
567
89
1011
12
131415
1617
18
19
20
21
22
23
24
25
26
27
Lea
f si
ze (
cm)
Seed volume (mm )3
Fig. 1 Regression of seed volume (mm3) and leaf size of the
studied taxa of Cucurbitaceae. For the taxa names (see Table 1)
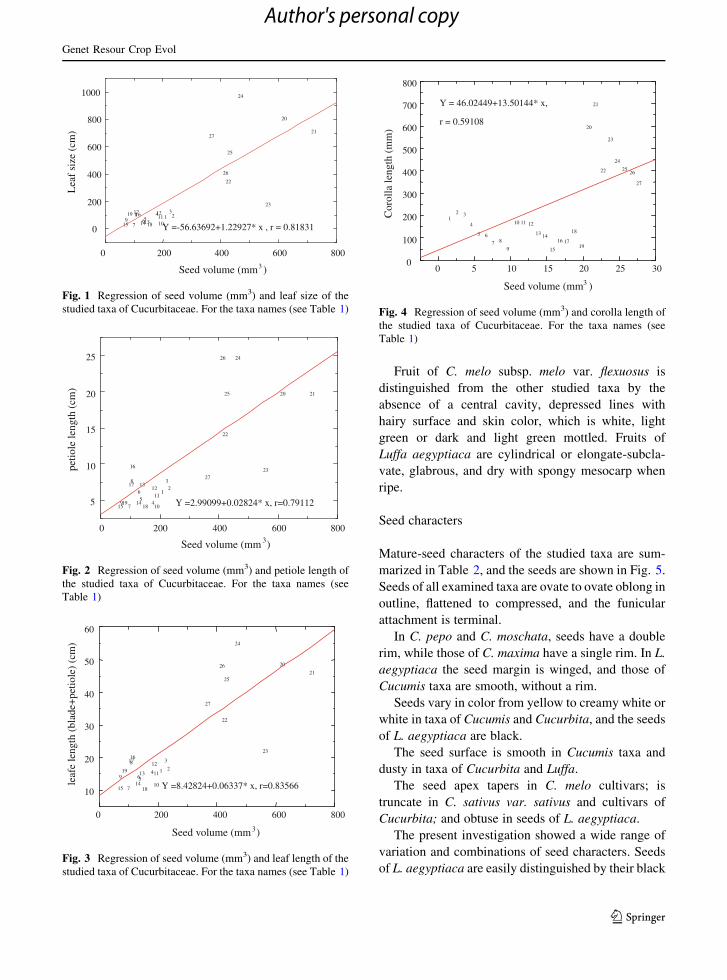
Y =2.99099+0.02824* x, r=0.79112
0 200 400 600 800
5
10
15
20
25
12
3
45
6
7
8
910
1112
13
1415
16
17
1819
20 21
22
23
24
25
26
27
petio
le le
ngth
(cm
)
Seed volume (mm )3
Fig. 2 Regression of seed volume (mm3) and petiole length of
the studied taxa of Cucurbitaceae. For the taxa names (see
Table 1)
Y =8.42824+0.06337* x, r=0.83566
0 200 400 600 800
10
20
30
40
50
60
1 2
3
456
7
8
9
10
11
12
13
1415
1617
18
19
20
21
22
23
24
25
26
27
leaf
e le
ngth
(bl
ade+
petio
le)
(cm
)
Seed volume (mm )3
Fig. 3 Regression of seed volume (mm3) and leaf length of the
studied taxa of Cucurbitaceae. For the taxa names (see Table 1)
0 5 10 15 20 25 300
100
200
300
400
500
600
700
800
12 3
4
5 67 8
9
10 11 12
13 14
15
16 17
18
19
20
21
22
23
24
2526
27
Cor
olla
leng
th (
mm
)
Seed volume (mm )3
Y = 46.02449+13.50144* x,
r = 0.59108
Fig. 4 Regression of seed volume (mm3) and corolla length of
the studied taxa of Cucurbitaceae. For the taxa names (see
Table 1)
Genet Resour Crop Evol
123
Author's personal copy

Fig. 5 Seed photographs of
the studied taxa. The scaleline 10 cm. For the taxa
names (see Table 1)
Fig. 6 Microphotographs showing different trichome types. For descriptions of the trichome types see the text
Genet Resour Crop Evol
123
Author's personal copy

color, winged margin and obtuse apex. The seed
characters in Cucurbita are useful at species level
only. C. melo subsp. melo var. flexuosus ‘Firany’ is
easily distinguished by its small-sized seeds
(9.000 ± 0.196 9 4.28 ± 0.108 mm.).
According to Leishman and Westoby (1994), seed
size is positively correlated with growth-form size.
From the data of the studied taxa, there is a significant
correlation between seed size and growth parameters.
The growth parameters of the studied taxa included
leaf size, petiole length, leaf length and corolla length.
There are highly positive significant correlations
between seed volume and leaf size (r = 0.81831,
P B 0.001, Fig. 1), seed volume and petiole length
(r = 0.79112, P B 0.001, Fig. 2), seed volume and
leaf length (r = 0.83566, P B 0.001, Fig. 3), and seed
volume and corolla length (r = 0.59108, P B 0.001,
Fig. 4).
Trichome type
The differences in trichome types are more useful in
taxonomic work than their density (Cutler 1979). This
is because the length, size and density of the trichomes
are more liable to vary with environmental factors than
type (Metcalfe and Chalk 1972).
Among the studied taxa, eight trichome types are
distinguished (Fig. 6).
Type ‘‘A’’: eglandular, multicellular uniseriate,
soft, long with long cells and punctatus hair surface.
Type ‘‘B’’: eglandular, multicellular uniseriate,
with two raised basal cells, long, hard, thick walled
and granular hair surface.
Type ‘‘C’’: eglandular uniseriate, with multicellular
base, hard, thick with rugulose surface.
Type ‘‘D’’: eglandular, multicellular uniseriate,
short with 3–4 cells, hard, thick walled with
granular hair surface.
Type ‘‘E’’: glandular, multicellular, uniseriate base
and long multicellular neck, and multiseriate glo-
boid head, often with 6 cells, soft, thin-walled with
smooth surface.
Type ‘‘F’’: glandular, multicellular, uniseriate with
four cells base and 6–8 trunk cells, thin-walled and
smooth surface.
Type ‘‘G’’: glandular, multicellular uniseriate con-
sists of at least 20 cells, soft, thin-walled and smooth
surface.
Type ‘‘H’’: glandular, multicellular, uniseriate
neck and multiseriate globoid head, thin-walled
and smooth surface.
The distribution of the different types of tric-
homes on mature stems, leaves, petioles, flower and
fruits among the studied taxa is summarized in
Table 3.
Trichome type ‘‘A’’ is present in all taxa studied
except those of Cucurbita, where Type ‘‘E’’ is
recorded. Trichome type ‘‘F’’ is only present on the
flower of C. sativus L. var. sativus.
Stomata and epidermal cells
All the studied taxa have anomocytic stomata on the
upper and lower leaf surfaces. Four types of epidermal
cells have been identified:
1. Straight in the varieties of C. melo subsp. melo.
2. Straight or slightly curved in the varieties of
C. melo subsp. melo var. aegyptiaca, C. melo
subsp. melo var. dudaim, C. melo subsp. melo var.
flexuosus and C. melo subsp. agrostis var. chate
‘Aggour’.
3. Folded in the varieties of Cucurbita and C. sativus
var. sativus.
4. Curved in the upper and folded in the lower
surface of L. aegyptiaca.
Leaves
Leaves mostly simple, palmately veined and lobed.
C. sativus has triangular, ovate often 3-lobed
with a pointed apex, and rounded ovate- reniform
with obtuse apex in C. melo and angled in
L. aegyptiaca.
Keys
Key 1
In the following key, vegetative, fruit and seed
characters are used. The quantitative characters are
omitted except for character 6 (the depth of the leaf
basal sinus). This character is very important for
identification at the level of cultivars (see the key;
led 18).
Genet Resour Crop Evol
123
Author's personal copy

Genet Resour Crop Evol
123
Author's personal copy

Key 2 (confirmatory)
The following key uses fruit and seed characters, and
vegetative and flower characters are omitted except
character 6 (depth of leaf basal sinus). The fruit and
seed characters can be used to identify most of the
Egyptian cultivars.
Genet Resour Crop Evol
123
Author's personal copy

Table 3 Trichomes types and epidermal cells characters among the studied taxa
Taxon Trichomes Epidermal cell
Stem Petiole Leaves Flower Fruit
Stalk Fruit
C. melo L. subsp. melo ‘Melo’ B, D C, D C, D A, D, G, H D – Straight
C. melo L. subsp. melo ‘Shahd’ B, D C, D B, C, D A, D, G, H D – Straight
C. melo L. subsp. melo ‘Santawi’ A, B, D B, D, H B, D A, D, G, H D – Straight
C. melo L. subsp. melo ‘Santawi
Taweel’
A, B, D B, D, H B, D A, D, G, H D – Straight
C. melo L. subsp. melo ‘Senani’ A, B, D B, C, D B, D A, D, G, H D – Straight
C. melo L. subsp. melo ‘Ananas
Kheshen’
B, C, D B, C, D B, C, D A, D, G, H D – Straight
C. melo L. subsp. melo ‘Ananas
Naiem’
C, D, B B, C, D B, C, D A, D, G, H D – Straight
C. melo L. subsp. melo ‘Mangawi’ B, D B, C, D B, D A, D, G, H D – Straight
C. melo L. subsp. melo var.
aegyptiaca Sickenb. ‘Mansouri’
B, C, D B, C, D B, C, D A, D, G, H – – Straight or slightly
curved
C. melo L. subsp. melo. var.
aegyptiaca Sickenb.
B, C, D C, D B, C, D A, D, G, H – – Straight or slightly
curved
C. melo L. subsp. melo var.
dudaim (L.) Naudin ‘Waraqi’
B, C, D B, C, D B, C, D A, D, G, H – – Straight or slightly
curved
C. melo L. subsp. agrestis(Naudin) Pangalo var. dudaim(L.) Naudin ‘Bassusi’
C, D C, D B, D A, D, G, H – – Straight or slightly
curved
C. melo L. subsp. agrestis(Naudin) Pangalo var. dudaim(L.) Naudin ‘Kisaan El-Assal’
C, D C, D B, D A, D, G, H – – Straight or slightly
curved
C. melo L. subsp. agrestis(Naudin) Pangalo var. dudaim(L.) Naudin ‘Qatta Saidi’
B, C, D B, C, D B, D A, D, G, H D – Straight or slightly
curved
C. melo L. subsp. melo var.
flexuosus (L.) Naudin ‘Firany’
B, C, D B, C, D B, D A, B, D, G, H A, D A, D Straight or slightly
curved
C. melo L. subsp. melo var.
flexuosus (L.) Naudin ‘Khadra’
B, C, D B, C, D B, D A, B, D, G, H D D Straight or slightly
curved
C. melo L. subsp. melo var.
flexuosus (L.) Naudin ‘Beida’
B, C, D B, C, D B, D A, B, D, G, H D D Straight or slightly
curved
C. melo L. subsp. agrestis(Naudin) Pangalo var. chate(Hasselq.) Sageret ‘Aggour’
A, B, D A, B, D B, D A, D, G, H D – Straight or slightly
curved
C. sativus L. var. sativus C, D, H C, D, H B, C, D A, D, F, H D D Folded
C. maxima Duchesne ex Lam.
‘Etampes Pumpkin’ (Abied)
C, D C, D B, C, D B, D, E, H – – Folded
C. maxima Duchesne ex Lam.
‘Spanish Pumpkin’ (Ahmar)
C, D C, D B, C, D B, D, E, H – – Folded
C. moschata Duchesne ex Lam.
‘Napple Squash’
B, D B, D D B, D, E, H – – Folded
C. moschata Duchesne ex
Lam.‘Long Neapolitan Squash’
B, D B, D D B, D, E, H – – Folded
C. pepo var. ovifera (L.) Alef.
‘Eskandarani’
C, D C, D B, C, D B, D, E, H, G C, D D Folded
Genet Resour Crop Evol
123
Author's personal copy

From the above keys, it can be concluded that
vegetative, flower and fruit charters are not enough for
separating all the studied taxa. However, depth of leaf
basal sinus and seed characters are very important in
the identification of the varieties and cultivars.
Based on a limited sampling of the morphological
characters and cultivars of Cucurbitaceae some sig-
nificant patterns of variation are clear. They can be
arranged according to their usefulness for identifica-
tions as follows: fruit characters, seed characters,
trichome type and epidermal cell type.
Acknowledgments The authors dedicate this work to the late
Prof. Dr. M. N. El-Hadidi, Botany Department, Cairo
University, who initiated this work. We thank Prof. Dr.
Hasnaa Hosni, Professor of Plant taxonomy in Botany
Department, Faculty of Science, Cairo University, Egypt for
reviewing through the manuscript and making valuable
suggestions. Special thanks to Prof. Dr. J. H. Kirkbride, Jr.,
USDA-ARS, US National Arboretum, Floral and Nursery Plants
Research Unit, 3501 New York Avenue NE, Washington, DC
20002-1958 USA for his valuable suggestions and comments on
the manuscript. Valuable references are provided by Prof. Dr.
De Wilde at National Herbarium Nederland, Universiteit Leiden
Branch, P.O. Box 9514, NL-2300 RA Leiden, The Netherlands.
Appendix
See Tables 2 and 3, Figs. 5 and 6.
*Character list
#1. Stem surface/ 1. Hispid/ 2. Hirsute/ 3.
Roughish/ 4. Scabrous/
#2. Tendril/ 1. Simple/ 2. Multified/
#3. Leaf size/ 1. \15 9 15 cm / 2. More than
15*15 cm/
#4. Leaf apex/ 1. Acute/ 2. Obtuse/ 3. Acuminate/
#5. Leaf margin/ 1. Dentate/ 2. Microdentate/
#6. Depth of leaf basal sinus/ 1. \8 mm/ 2. More
than 8 mm/
#7. Petiole length/ 1.\12 cm/ 2. More than 12 cm/
#8. Leaf epidermal cells/ 1. Straight/ 2. Straight or
slightly curved/ 3. Curved upper folded lower/
4. Folded/
#9. Flower colour/ 1. Yellow/ 2. Orange/
#10. Flower pedicel/ 1.\15 mm long/ 2. More than
15 mm long/
#11. Receptacle length/ 1.\7 mm/ 2. More than 7 mm
#12. Calyx length/ 1. \10 mm/ 2. 10–25 mm/ 3.
More than 25 mm/
#13. Corolla lobe apex/ 1. Obtuse/ 2. Acute/
#14. Corolla length/ 1. \15 mm/ 2. 15–30 mm/ 3.
40–100 mm/ 4. More than 100 mm/
#15. Corolla tube/ 1. Less than half the length of
corolla/ 2. More than half the length of corolla/
#16. Fruit shape/ 1. Globoid/ 2. Elongate/
#17. Fruit skin/ 1. Smooth/ 2. Smooth–rough/ 3.
Rough-netted/
#18. Fruit surface/ 1. Glabrous/ 2. Hairy/
#19. Fruit ends/ 1. Flat at both ends/ 2. Tapering at
both ends/ 3. Tapering one end/
#20. Fruit colour/ 1. Orange-reddish/ 2. Yellow/ 3.
Greenish/
#21. Fruit furrows/ 1. Present/ 2. Absent/
#22. Fruit odour/ 1. Strong melon/ 2. Melon odour/
3. Musk odour/ 4. Chat-cucumber odour/
#23. Fruit flesh-color/ 1. Orange–yellow/ 2. White–
greenish/
#24. Fruit central cavity/ 1. Present/ 2. Absent/
#25. Fruit width/ 1.\12 cm/ 2. More than 12 cm/
#26. Seed shape/ 1. Ovate/ 2. Oblong–ovate/
Table 3 continued
Taxon Trichomes Epidermal cell
Stem Petiole Leaves Flower Fruit
Stalk Fruit
C. pepo var. ovifera (L.) Alef.
‘Baladi Khadra’.
C, D C, D B, C, D B, D, E, H, G C, D D Folded
C. pepo var. ovifera (L.) Alef.
‘Baladi Beida’
C, D C, D B, C, D B, D, E, H, G C, D D Folded
L. aegyptiaca Mill. B B, D D, H A, B, D, G B – Curved upper/
folded lower
– Absent
Genet Resour Crop Evol
123
Author's personal copy

#27. Seed colour/ 1. Whitish/ 2. Yellowish or
darker/ 3. Black/
#28. Seed base/ 1. Obtuse/ 2. Tapering/
#29. Seed apex/ 1. Tapering/ 2. Truncate/ 3.
Truncate–oblique/ 4. Obtuse/
#30. Seed surface/ 1. Smooth/ 2. Dusty {What does
this mean?}/
#31. Seed rim/ 1. Rimless/ 2. Single rim/ 3. Double
rim/
#32. Seed margin/ 1. Smooth/ 2. Rough/ 3. Wavy/ 4.
Winged/
#33. Seed length/ 1.\15 mm/ 2. More than 15 mm/
#34. Seed width/ 1. \5 mm/ 2. 5–8 mm/ 3. More
than 8 mm/
#35. Seed thickness/ 1.\2 mm/ 2. 2–3 mm/ 3. More
than 3 mm/
#36. Seed weight (/seed)/ 1. \100 mg/ 2.
100–200 mg/ 3. More than 200 mg/
#37. Seed (length 9 width 9 thickness)/ 1. \100/
2. 100–300/ 3. 300–500/ 4. More than 500/
References
Alefeld F (1866) Landwirtschaftliche Flora. Wiegandt und
Hempel, Berlin
Boulos L (2000) Flora of Egypt, vol 2. Al Hadara, Cairo,
pp 134–143
Castetter EF (1925) Horticultural groups of cucurbits. Proc Am
Soc Hort Sci 22:338–340
Cogniaux A, Harms H (1924) Cucurbitaceae. Cucurbiteae–Cu-
cumerimae. In: Engler, Das Pflanzenreich 88(IV.275.II):
1–246
Coleman CO, Lowry JK, Macfarlane T (2010) DELTA for
beginners: an introduction into the taxonomy software
package DELTA. ZooKeys 45:1–75
Cutler DF (1979) The scanning electron microscope in sys-
tematic plant anatomy. In: Metcalfe CR, Chalk L (eds)
Anatomy of the dicotyledons, vol 1, 2nd edn. Clarendon
Press, Oxford, p 91
Dallwitz MJ (2009) Overview of the DELTA system.
http://delta-intkey.com/www/overview.htm
Dallwitz MJ, Paine TA, Zurcher EJ (2000) User’s guide to the
DELTA system: a general system for processing taxo-
nomic descriptions, 4.12th edn. CSIRO Division of Ento-
mology, Canberra
Dallwitz MJ, Paine TA, Zurcher EJ (2002) Interactive identifi-
cation using the internet. In: Saarenmaa H, Nielsen ES
(eds) Towards a global biological information infrastruc-
ture—challenges, opportunities, synergies, and the role of
entomology. European Environment Agency technical
report 70. EEA, Copenhagen, pp 23–33
Decker DS (1988) Origin(s), evolution, and systematics of
Cucurbita pepo (Cucurbitaceae). Econ Bot 42:4–15
Goff ES (1888) Report of the horticulturist. N Y St Agric Exp
Stat Ann Rep 6:76–306
Hassib M (1938) Cucurbitaceae in Egypt. Faculty of Science,
Fouad 1 University, Cairo, Egypt. Publ. 3. Imp. F. E. Noury
& Fils, p 173
Heneidaq S (2008) Taxonomic revision of wild species of Cu-
curbitaceae in Egypt. Taeckholmia 28:21–55
Jeffrey C (1980) A review of the Cucurbitaceae. Bot J Linn Soc
81:233–247
Jeffrey C (1990) An outline classification of the Cucurbitaceae.
In: Bates DM, Robinson RW, Jeffrey C (eds) Biology and
utilization of the Cucurbitaceae. Cornell University Press,
Ithaca, pp 449–463
Jeffrey C (2001) Cucurbitaceae. In: Hanelt P, Institute of Plant
Genetics and Crop Plant Research (eds) Mansfeld’s ency-
clopedia of agricultural and horticultural crops, vol 3.
Springer, Berlin, pp 1510–1557
Kirkbride JH (1993) Biosystematic monograph of the genus
Cucumis (Cucurbitaceae). Parkay, Boone, p 159
Kurz S (1877) Contributions towards a knowledge of the Bur-
mese flora, Cucurbitaceae. J Asia Soc Bengal 46(2):47–58
Leishman MR, Westoby M (1994) Hypotheses on seed size:
tests using the semiarid flora of western New South Wales,
Australia. Am Nat 143:890–906
Linnaeus C (1753) Species Plantarum. Haunae
Metcalfe CR, Chalk L (1972) Anatomy of the dicotyledons, vol
1. Clarendon Press, Oxford
Montasir AM, Hassib M (1956) Manual flora of Egypt. I. Im-
primerie Misr, SAE
Munger HM, Robinson RW (1991) Nomenclature of Cucumismelo L. Cucurbit Genet Co-op Rep 14:43
Naudin C (1856) Nouvelles recherches sur les caracteres spe-
cifiques et les varietes des plantes du genre Cucurbita. Ann
Sci Nat Bot IV 6:5–73
Naudin C (1859) Essaia d’une monographie des especes et des
variet es du genre Cucumis. Ann Sci Natl Bot ser 4 11:5–87
Nesom GL (2011) Toward consistency of taxonomic rank in
wild/domesticated Cucurbitaceae. Phytoneuron 13:1–33
Pankhurst RJ (1991) Practical taxonomic computing. Cam-
bridge University Press, Cambridge
Paris HS (1986) A proposed subspecific classification for
Cucurbita pepo. Phytologia 61:133–138
Pitrat M, Hanelt P, Hammer K (2000) Some comments on
infraspecific classification of cultivars of Cucumis melo.
Acta Hort 510:29–36
Rabei S (2011) A morphological, anatomical and taxonomic
study of Umbelliferae in Egypt. LAP Lambert, Koln
Rabei S, El-Gazzar A (2007) Computer-generated keys to spe-
cies of Hyptis s.l. (Labiatae). Taeckholmia 27:11–39
Rizk RMH (2001) Cytotaxonomic studies on the Egyptian taxa
of Cucurbitaceae. Unpublished Ph.D. thesis, Department of
Botany, Faculty of Science at Damietta, Mansoura Uni-
versity, Egypt
Simpson M (2010) Plant systematics, 2nd edn. Academic Press,
London, pp 341–344
Stearn WT (1973) Botanical Latin. David & Charles, Newton
Abbott
Tackholm V (1974) Students’ flora of Egypt, 2nd edn. Cairo
University, Cairo, pp 372–376
Willdenow CL (1805) Species Plantarum, vol 4, 4th edn. Imp-
ensis G. C. Nauk, Berlin
Genet Resour Crop Evol
123
Author's personal copy




















