
Jungia sellowii suppresses the carrageenan-induced inflammatory response in the mouse model of...
15
RESEARCH ARTICLE Jungia sellowii suppresses the carrageenan-induced inflammatory response in the mouse model of pleurisy Marina Nader • Geison Vicente • Julia Salvan da Rosa • Tamires Cardoso Lima • Alyne Machado Barbosa • Alan Diego Conceic ¸a ˜o Santos • Andersson Barison • Eduardo Monguilhott Dalmarco • Maique Weber Biavatti • Ta ˆnia Silvia Fro ¨de Received: 25 May 2014 / Accepted: 17 June 2014 Ó Springer Basel 2014 Abstract This study was conducted to explore the anti- inflammatory effect of Jungia sellowii (Asteraceae) using a murine model of pleurisy induced by carrageenan (Cg). This plant is used in southern Brazil to treat inflammatory diseases. J. sellowii leaves were extracted with ethanol/ water to obtain the crude extract (CE), which was frac- tionated with different solvents, yielding n-hexane (Hex), dichloromethane (DCM), ethyl acetate (EtOAc) and n- butanol (BuOH) fractions, and aqueous fraction (Aq). The major compounds succinic acid (SA) and lactic acid (LA) were isolated from Aq fraction, and their structures were determined by 1 H and 13 C NMR. Pleurisy was induced by Cg (Saleh et al. 1996). The leukocytes, exudation, myelo- peroxidase (MPO) and adenosine–deaminase (ADA) activities, metabolites of nitric oxide (NO x ) levels, protein levels and mRNA expression for interleukin 1 beta (IL-1b), tumour necrosis factor alpha (TNF-a), interleukin 17A (IL17A) and inducible of nitric oxide synthase (iNOs), and p65 protein phosphorylation (NF-jB) were analysed 4 h after pleurisy induction. Animals pre-treated with CE, BuOH, Aq, SA, or LA inhibited leukocytes, exudation, MPO and ADA activities, NO x , IL-1b, TNF-a, and IL-17A levels, and the mRNA expression for IL-1b, TNF-a, IL- 17A, iNOS, and p65 protein phosphorylation (NF-jB) (p \ 0.05). Our study demonstrated that J. sellowii can protect against inflammation induced by Cg by decreasing the leukocytes and exudation. Its effects are related to the decrease of either proinflammatory cytokines and/or NO x . The isolated compounds SA and LA may play an important role in this anti-inflammatory action by inhibiting all the studied parameters. The anti-inflammatory properties of these compounds are due to the downregulation of NF-jB. Keywords Jungia sellowii Anti-inflammatory properties Pleurisy induced by carrageenan NF-kappa B Lactic acid Succinic acid Introduction There is increasing interest in the biological activities of plant extracts, especially those related to their uses in tra- ditional medicine. Latin America, including Brazil holds a great number of underexploited and largely unknown plant varieties. These plants have nutritional and/or medicinal properties (NRC 1989; De-La-Cruz et al 2007). Traditional use of these plants by native population has guaranteed conservation of many species. Jungia is a genus from the Asteraceae family, which comprises shrubs, lianas and herbs and is widespread from M. Nader G. Vicente J. S. da Rosa E. M. Dalmarco T. S. Fro ¨de (&) Department of Clinical Analysis, Center of Health Sciences, Federal University of Santa Catarina (UFSC), Campus Universita ´rio-Trindade, Floriano ´polis, Santa Catarina 88040-970, Brazil e-mail: [email protected] T. C. Lima A. M. Barbosa M. W. Biavatti Department of Pharmaceutical Sciences, Center of Health Sciences, Federal University of Santa Catarina (UFSC), Campus Universita ´rio-Trindade, Floriano ´polis, Santa Catarina 88040-970, Brazil A. D. C. Santos A. Barison Department of Chemistry, Centre of Mathematical Sciences, Federal University of Parana ´ (UFPR), Curitiba, Parana ´ 81531990, Brazil Inflammopharmacol DOI 10.1007/s10787-014-0210-3 Inflammopharmacology 123
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Jungia sellowii suppresses the carrageenan-induced inflammatory response in the mouse model of...

RESEARCH ARTICLE
Jungia sellowii suppresses the carrageenan-induced inflammatoryresponse in the mouse model of pleurisy
Marina Nader • Geison Vicente • Julia Salvan da Rosa • Tamires Cardoso Lima •
Alyne Machado Barbosa • Alan Diego Conceicao Santos • Andersson Barison •
Eduardo Monguilhott Dalmarco • Maique Weber Biavatti • Tania Silvia Frode
Received: 25 May 2014 / Accepted: 17 June 2014
� Springer Basel 2014
Abstract This study was conducted to explore the anti-
inflammatory effect of Jungia sellowii (Asteraceae) using a
murine model of pleurisy induced by carrageenan (Cg).
This plant is used in southern Brazil to treat inflammatory
diseases. J. sellowii leaves were extracted with ethanol/
water to obtain the crude extract (CE), which was frac-
tionated with different solvents, yielding n-hexane (Hex),
dichloromethane (DCM), ethyl acetate (EtOAc) and n-
butanol (BuOH) fractions, and aqueous fraction (Aq). The
major compounds succinic acid (SA) and lactic acid (LA)
were isolated from Aq fraction, and their structures were
determined by 1H and 13C NMR. Pleurisy was induced by
Cg (Saleh et al. 1996). The leukocytes, exudation, myelo-
peroxidase (MPO) and adenosine–deaminase (ADA)
activities, metabolites of nitric oxide (NOx) levels, protein
levels and mRNA expression for interleukin 1 beta (IL-1b),
tumour necrosis factor alpha (TNF-a), interleukin 17A
(IL17A) and inducible of nitric oxide synthase (iNOs), and
p65 protein phosphorylation (NF-jB) were analysed 4 h
after pleurisy induction. Animals pre-treated with CE,
BuOH, Aq, SA, or LA inhibited leukocytes, exudation,
MPO and ADA activities, NOx, IL-1b, TNF-a, and IL-17A
levels, and the mRNA expression for IL-1b, TNF-a, IL-
17A, iNOS, and p65 protein phosphorylation (NF-jB)
(p \ 0.05). Our study demonstrated that J. sellowii can
protect against inflammation induced by Cg by decreasing
the leukocytes and exudation. Its effects are related to the
decrease of either proinflammatory cytokines and/or NOx.
The isolated compounds SA and LA may play an important
role in this anti-inflammatory action by inhibiting all the
studied parameters. The anti-inflammatory properties of
these compounds are due to the downregulation of NF-jB.
Keywords Jungia sellowii �Anti-inflammatory properties �Pleurisy induced by carrageenan � NF-kappa B �Lactic acid � Succinic acid
Introduction
There is increasing interest in the biological activities of
plant extracts, especially those related to their uses in tra-
ditional medicine. Latin America, including Brazil holds a
great number of underexploited and largely unknown plant
varieties. These plants have nutritional and/or medicinal
properties (NRC 1989; De-La-Cruz et al 2007). Traditional
use of these plants by native population has guaranteed
conservation of many species.
Jungia is a genus from the Asteraceae family, which
comprises shrubs, lianas and herbs and is widespread from
M. Nader � G. Vicente � J. S. da Rosa � E. M. Dalmarco �T. S. Frode (&)
Department of Clinical Analysis, Center of Health Sciences,
Federal University of Santa Catarina (UFSC), Campus
Universitario-Trindade, Florianopolis, Santa Catarina
88040-970, Brazil
e-mail: [email protected]
T. C. Lima � A. M. Barbosa � M. W. Biavatti
Department of Pharmaceutical Sciences, Center of Health
Sciences, Federal University of Santa Catarina (UFSC), Campus
Universitario-Trindade, Florianopolis, Santa Catarina
88040-970, Brazil
A. D. C. Santos � A. Barison
Department of Chemistry, Centre of Mathematical Sciences,
Federal University of Parana (UFPR), Curitiba, Parana
81531990, Brazil
Inflammopharmacol
DOI 10.1007/s10787-014-0210-3 Inflammopharmacology
123

North and Central America to South America, including
southern Brazil. The species Jungia sellowii less. (J. sel-
lowii) is also found in the South Brasil, state of Santa
Catarina (Katinas et al. 2008; Marchant et al. 2002).
The leaves, stems and flowers from Jungia paniculata
(J. paniculata) are used to disinfect external cuts and to
treat urinary tract inflammation, amigdalitis, tonsillitis,
stomach ulcers, nephritis and haemorrhoids, as well as an
antibacterial agent in traditional medicine in South Amer-
ica (De-La-Cruz et al. 2007; Hammond et al. 1998;
Rehecho et al. 2011). Additionally, the leaves and stems
from J. paniculata are used as a depurative of blood in
traditional medicine in South America (Goleniowski et al.
2006).
There are few pharmacological studies of anti-inflam-
matory effects of Jungia except for that described by
Casado et al. (2010), who demonstrated the anti-inflam-
matory and antioxidant effects of J. paniculata, a plant
used as topical anti-inflammatory herb in traditional med-
icine in South America. In that study the effect of the herb
was associated with the presence of polyphenols and
flavonoids.
Phytochemical analysis in our laboratory identified two
carboxylic acids, succinic acid (SA) and lactic acid (LA)
that were isolated from the aqueous fraction of the leaf
extract of J. sellowii.
In particular, LA can inhibit the inflammatory response
by decreasing interleukin-6 (IL-6) and tumour necrosis
factor-a (TNF-a) messenger ribonucleic acid (mRNA)
levels. Also LA prevented the activation of nuclear factor
NF-Kappa B (NF-jB) in these experiments (Jiang et al.
2013; Liu et al. 2011a, b; Xu et al. 2013).
However, thus far, the effect of J. sellowii as an anti-
inflammatory herb as indicated by its use in Brazilian tra-
ditional medicine has not been investigated. No scientific
study regarding this species was found in the literature to
support its use.
Taking into consideration: (1) the importance of the
regulation of cytokines and other mediators in inflamma-
tory diseases; (2) the increasing use of herbs to treat
inflammation; and (3) the relative lack of studies focusing
on the anti-inflammatory mechanism of action of plants
used as anti-inflammatory agents in traditional medicine. In
this study, the anti-inflammatory potential of J. sellowii
was explored in a murine model of pleurisy induced by
carrageenan (Cg).
This study tried to demonstrate the impact and the
possible mechanism of the anti-inflammatory action of the
crude extract (CE) of J. sellowii, its derived fractions and
isolated compounds on several proinflammatory parame-
ters. The following parameters were studied: leukocyte
content, degree of exudation, myeloperoxidase (MPO) and
adenosine-deaminase (ADA) activities, nitric oxide
metabolites (NOx), interleukin-1b (IL-1b), TNF-a and
interleukin-17A (IL-17A) levels and the mRNA expression
of all studied cytokines. Also the mRNA expression of
inducible nitric oxide synthase (iNOS) was analysed. Fur-
thermore, we investigated the effect of the isolated
compounds (SA and LA) on p65 protein phosphorylation
[p-p65 Nuclear factor NF-kappa B (NF-jB)].
Materials and methods
Plant material
The leaves of J. Sellowii were collected in Rio Negr-
inho, Santa Catarina, Brazil, in March 2012. Plant
identification was performed by the botanist Dr. Ademir
Reis from the botany department at the Federal Uni-
versity of Santa Catarina, and a voucher specimen (RB
number 537.991) is preserved in the Jardim Botanico at
Rio de Janeiro, Brazil.
Drugs and reagents
The following drugs and reagents used were purchased
from Sigma Chemical Co. (St Louis, MO, USA): carra-
geenan (degree IV), human neutrophil myeloperoxidase,
indomethacin, adenosine, Evans blue dye, ortho-dianisi-
dinedihydrochloride (3,30dimethoxybenzidine), phenol,
sulphanilamide, sodium azide, sodium nitroprussiate,
naphthylethylendiamidedihydrochloride, phosphoric acid,
vanadium chloride III and all primers. Dexamethasone
was purchased from Ache Pharmaceutical Laboratories
S.A. (Sao Paulo, SP, Brazil). Sodium hydrogen phos-
phate, zinc sulphate and hydrogen peroxide were
purchased from Vetec (Rio de Janeiro, RJ, Brazil).
Sodium hydroxide was purchased from Reagen (Rio de
Janeiro, RJ, Brazil). Turk–May–Grunwald dye was
obtained from Newprov (Pinhais, PR, Brazil), and
Giemsa dye was obtained from Laborclin (Pinhais, PR,
Brazil). ELISA kits for mouse IL-1b, TNF-a, IL-17A and
the Insta One Phospho-NF-jB p-65 (Ser536) were
obtained from IBL Immuno Biological Laboratories Co.,
Ltd. (Fujioka-city, Gunma, Japan), BD Biosciences
Pharmingen (San Diego, CA, USA) and eBioscience, Inc.
(San Diego, CA, USA), respectively. PCR-grade agarose
was purchased from Laboratorios Conda (Torrejon de
Ardoz, Madri, Spain), and Brazol� and ethidium bromide
were purchased from LGC Biotecnologia (Cotia, SP,
Brazil). DNase grade I� and the RevertAid H Minus First
Strand cDNA synthesis kit were purchased from Fer-
mentas (Burlington, Ontario, Canada). All other reagents
used were of analytical grade and obtained from several
commercial sources.
M. Nader et al.
123

Extraction and isolation
Fresh leaves of J. Sellowii (1.77 kg) were extracted
exhaustively at room temperature with ethanol and water
(3 L, 50:50, v/v) for 30 days. After evaporation of the
solvent under reduced pressure using a Rotavapor� R-210
(BuchiLabortechnik AG, Flawil, Switzerland), 65.9 g of
the hydro-alcoholic crude extract (CE) from J. sellowii was
obtained. This CE was then dissolved in water (600 mL)
and partitioned with solvents of increasing polarity,
yielding n-hexane (Hex: 0.414 g), dichloromethane (DCM:
0.973 g), ethyl acetate (EtOAc: 0. 590 g) and n-butanol
(BuOH: 0.574 g) fractions, as well as a residual aqueous
fraction (Aq) that was lyophilised to give a hygroscopic
solid matrix (31.9 g). An aliquot of this solid (9.5 g) was
solubilized in water and subjected to vacuum liquid chro-
matography (VLC) using RP-18 silica (230–400 mesh)
(Lichroprep Merck, Darmstadt, Germany). Elution was
carried out using different gradients of water and methanol
(100:0, 70:30, 50:50, 30:70, and 0:100), yielding five sub-
fractions (A, B, C, D and E, 100 mL each). Sub-fraction E
(0.185 g) presented a good chromatographic profile and
was partly soluble in methanol (20 mL), then filtered using
a C-18 cartridge (Varian ChemElut, CA, USA) to retain
non-polar compounds. Subsequently, the eluted sub-frac-
tion (0.154 g) was purified in a Sephadex LH-20 column
(GE Healthcare, UK) using acetone–methanol (1:1) as
eluent. 27 sub-fractions of 20 mL were obtained (Fig. 1).
Thin layer chromatography was performed, and sub-frac-
tions 10–14 and 24–27 exhibited the same chemical
pattern, so they were reunited, affording lactic acid (LA:
33.4 mg) and succinic acid (SA: 9.1 mg), respectively.
Characterisation of the isolated compounds
1H and 13C NMR spectra were obtained using a high-res-
olution Bruker AVANCE-400 MHz NMR spectrometer
(Bruker, Fremont, CA, USA), at a frequency of 400 MHz
for 1H and 100 MHz for 13C. Deuterium oxide (D2O) was
used as a solvent and tetramethylsilane (TMS) as a
chemical shift reference. The isolated compounds LA and
SA presented a spectral grade purity superior to 99 %.
Lactic acid
Colourless liquid. 1H NMR: d1.25 (d, J = 7 Hz, H-1), 4.22
(q, J = 7 Hz, H-2). 13C NMR: d 21.9 (C-1), 69.6 (C-2),
182.4 (C-3).
Succinic acid
White solid.1H NMR: d 2.47 (s, H-2 and H-3). 13C NMR: d34.4 (C-2 and C-3), 181.2 (C-1 and C-4).
Animals
One-month-old female Swiss mice weighing 18–21 g
were randomly divided into the following 13 groups of
five animals each: 1 = negative control group =
received only an intrapleural injection of sterile saline
(0.9 % NaCl, i.pl.), 2 = vehicle control group =
receive only an intrapleural injection of sterile saline
(0.9 % NaCl) plus dimethylsulfoxide (10 %),
3 = positive control group = received only Cg (1 %)
dissolved in 0.9 % NaCl administered intrapleurally
(i.pl.), 4 and 5 = reference anti-inflammatory drugs
groups = received an intraperitoneal (i.p.) injection of
dexamethasone (Dex: 0.5 mg/kg) or indomethacin
(Indo: 5 mg/kg, i.p.) 30 min before Cg, and groups
6–13 each received an intraperitoneal injection of dif-
ferent doses of CE, its derived fractions (Hex, DCM,
EtOAc, BuOH, and Aq) or isolated compounds (SA,
LA) 30 min before Cg. In order to dissolve the low
polarity of fractions (Hex, DCM and EtOAc) a maxi-
mum of 10 % of dimethylsulfoxide was added to the
fraction solution (0.9 % NaCl). The animals were
housed under standardised conditions (room tempera-
ture of 21 ± 2 �C, with 12 h alternating periods of
light and darkness and 50–60 % humidity) and had free
access to a standard mouse diet and water. This study
was approved by the Committee for Ethics in Animal
Research at Federal University of Santa Catarina
(license no. PP00757) and the experiments were per-
formed in accordance with the principles stated in the
Guide for Care and Use of Laboratory Animals
[National Council of Animal Experimentation Control
(COSEA)].
Induction of pleurisy
Pleurisy was induced by a single intrapleural injection of
0.1 mL 1 % Cg (diluted in 0.9 % NaCl), as previously
described by Saleh et al. (1996). After 4 h, animals were
euthanised with an overdose of pentobarbital (180 mg/kg,
i.p.). The thoracic cavity was exposed and washed with
1.0 mL sterile phosphate buffered saline [PBS, pH 7.6,
composition: NaCl (130 mmol); Na2HPO4 (5 mmol);
KH2PO4 (1 mmol), and distilled water (1,000 mL) con-
taining heparin (20 IU/mL)]. The fluid leakage was
collected for further quantification of total and differential
leukocyte content, degree of exudation, MPO and ADA
activities, as well as NOx and cytokine (IL-1b, TNF-a and
IL-17A) levels. Lung tissue samples were collected for
further the analysis of IL-1b, TNF-a, IL-17A, iNOS
mRNA expression and the measurement of p65 protein
phosphorylation (p-p65 NF-jB).
Jungia sellowii suppresses the carrageenan-induced inflammatory
123

Experimental protocol
To determine the doses of the plant material to be
used in subsequent experiments, dose–response curves
of the CE, its derived fractions and isolated com-
pounds were performed. Therefore, different groups of
animals were treated with different doses of CE
(10–50 mg/kg, i.p.), Hex (25–50 mg/kg, i.p.), DCM
(25–50 mg/kg, i.p.), EtOAc (25–50 mg/kg, i.p.), BuOH
(10–50 mg/kg, i.p.), Aq (1–25 mg/kg, i.p.), SA
(0.5–2.5 mg/kg, i.p.) or LA (0.5–2.5 mg/kg, i.p.), 0.5 h
prior to pleurisy induction. The total and differential
leukocyte content and the degree of exudation were
analysed after 4 h.
Based on the results of single doses of the plant material,
which were able to significantly inhibit the leukocyte
content and the degree of exudation, doses were selected
for each group (CE: 25 mg/kg, BuOH: 25 mg/kg, Aq:
5 mg/kg, SA: 1 mg/kg and LA: 1 mg/kg) to evaluate the
effect of the herb on MPO and ADA activities and NOx, IL-
1b, TNF-a, and IL-17A levels, IL-1b, TNF-a, IL-17A and
iNOS mRNA expression levels and p65 protein phos-
phorylation (p-p65 NF-jB). All parameters were analysed
4 h after pleurisy induction.
Fig. 1 Flow diagram of the
extraction process of Jungia
sellowii leaves. VLC vacuum
liquid chromatography
M. Nader et al.
123

Determination of leukocyte content and degree
of exudation
After euthanising the animals with an overdose of anaes-
thetic and exposing the pleural cavity, the fluid leakage was
collected for the analysis of total and differential leukocyte
content and the degree of exudation. The total leukocyte
count was performed in a veterinarian automatic counter
adjusted to mouse parameters (MINDRAY, BC-2800 Vet,
Nanshan, Shenzhen, China), and the cytospin preparations
were stained using May–Grunwald–Giemsa dye for the
differential count (Saleh et al. 1996).
The animals were injected with 0.2 mL Evans blue dye
(25 mg/kg) by the intravenous route 10 min before
receiving the herb treatment to evaluate the degree of
exudation. The concentration of the dye was determined by
interpolation from a standard Evans blue dye curve
(0.01–50 lg/mL) by colorimetric measurements at 620 nm
on an enzyme-linked immunosorbent assay (ELISA) plate
reader (OrganonTeknika, Roseland, NJ, USA).
Determination of MPO and ADA activities
The determination of MPO and ADA activities was con-
ducted in accordance with previously described
methodology (Giusti and Galanti 1984; Rao et al. 1993). In
both in-house assays, the concentration of the enzymes in
samples of the fluid leakage from the pleural cavity was
estimated by interpolation from a standard curve by col-
orimetric measurements at 450 and 630 nm, respectively,
on an ELISA plate reader (OrganonTecknica, Roseland,
NJ, USA).
Determination of NOx levels
The amount of nitrite (NO2-) and nitrate (NO3
-), as
indicators of NO production, was determined in samples
from the fluid leakage from the pleural cavity by the Griess
method (Green et al. 1982). The concentration of metab-
olites was determined by interpolation from a standard
curve of sodium nitrite (0–150 lM) by colorimetric mea-
surements at 540 nm on an ELISA plate reader
(OrganonTecknika, Roseland, NJ, USA).
Determinations of IL-1b, TNF-a and IL-17A levels
To quantify the proinflammatory cytokines (IL-1b, TNF-aand IL-17A) in the fluid leakage from the pleural cavity,
commercially available ELISA kits containing monoclonal
antibodies for each cytokine were used according to the
instructions of the manufacturer. The ranges of values
detected by these assays were as follows: IL-1 b,
11.72–750 pg/mL; TNF-a, 5–2.000 pg/mL; and IL-17A,
4–500 pg/mL. The intra-assay coefficients of variation
(CV) were as follows: IL-1b, 3.6 ± 0.5 %; TNF-a,
7.8 ± 0.9 %; and IL-17A, 7.5 ± 1.7 %. The inter-assay
CV were as follows: IL-1b, 9.0 ± 1.7 %; TNF-a,
9.6 ± 2.1 %; and IL-17A, 6.2 ± 0.9 %. Sensitivity values
were as follows: IL-1b, 1.67 pg/mL; TNF-a, 5.0 pg/mL;
and IL-17A, 4.0 pg/mL. The quantification of these cyto-
kines was obtained by interpolation from a standard curve
by colorimetric measurements at 450 nm on an ELISA
plate reader (OrganonTeknika, Roseland, NJ, USA).
Semi-quantification of the mRNA expression levels
of IL-1b, TNF-a, IL-17A and iNOS
Total ribonucleic acid (RNA) was extracted from lung
tissue samples (200 mg) using Brazol� (1,000 mL) plus
chloroform (200 mL) and 75 % isopropyl alcohol (500 lL,
V/V). After 15 min of centrifugation (12,0009g at 4 �C),
the precipitate was washed with 75 % ethanol (1,000 lL,
V/V). The pellets were dissolved in a 0.1 % diethylpyro-
carbonate (DEPC)/water solution (100 lL, V/V). The RNA
concentration was determined using a Picodrop Microliter
UV/Vis spectrophotometer (Picodrop Ltd., Hinxton, Cam-
bridge, UK). The total RNA (1 lg/lL) was mixed with
DNase Grade I (1 lL) to obtain the complementary
deoxyribonucleic acid (cDNA) according to the manufac-
turer’s instructions. The DNase-treated RNA (500 ng) was
added to a solution containing OligodT (12–18) (1 lL) and
reverse transcriptase (1 lL) in accordance with the manu-
facturer’s protocol. All reactions were performed in a
thermocycler (Life Technologies Corporation, Carlsbad,
CA, USA). The primer design for specific genes was based
on the sequences published in Gen-Bank and the poly-
merase chain reaction (PCR) conditions of our protocol
(Table 1). Primers for b-actin were used as the reverse
transcriptase control (internal control) for PCR analysis.
The PCR products were separated by electrophoresis using
an agarose gel (1 %) containing ethidium bromide (0.5 lg/
mL) and then visualised on an ultraviolet transilluminator
(Amersham Biosciences AB, Uppsala, Sweden). The rela-
tive amount of the reverse transcription-polymerase chain
reaction (RT-PCR) products was determined by digital
photography followed by densitometric analysis of the
ethidium bromide-stained agarose gels normalised against
the internal control using Photo-Capt image version 12.4
for Windows� to quantify the intensity.
Measurement of p65 protein phosphorylation
(p-p65-NF-jB)
In this assay, mouse lung tissue was collected (20 mg) to
evaluate the effect of the isolated compounds, SA and LA,
on p65 protein phosphorylation (NF-jB). A commercially
Jungia sellowii suppresses the carrageenan-induced inflammatory
123
available kit, Insta One Phospho-NF-jB p 65 (Ser536)
ELISA Kit (eBioscience, San Diego, CA, USA), with
monoclonal specific antibodies against phosphorylated
mouse p65 protein was used following the manufacturer’s
protocol. The absorbance of the samples was obtained at
450 nm on an ELISA plate reader (OrganonTeknika,
Roseland, NJ, USA). The total protein content was mea-
sured with a Lowry assay (Lowry et al. 1951) and adjusted
to 60 lg protein per sample in all studied groups to stan-
dardise the p65 protein phosphorylation analysis. The
results are expressed as relative fold change in comparison
with the negative control group, which represents the basal
level of p-65 phosphorylation.
Data analysis
To assess data normality and the homogeneity of variances,
the Shapiro–Wilks test was used. The data are presented as
the mean ± standard error of the mean (SEM) and percent
of five animals per group. Significant differences between
groups were determined by two-way analysis of variance
(ANOVA) followed by the Newman–Keuls post hoc test.
Values of p \ 0.05 were considered significant. All sta-
tistical analyses were performed with GraphPad Prism
software (version 3.0) (San Diego, CA, USA).
Results
Phytochemical analysis
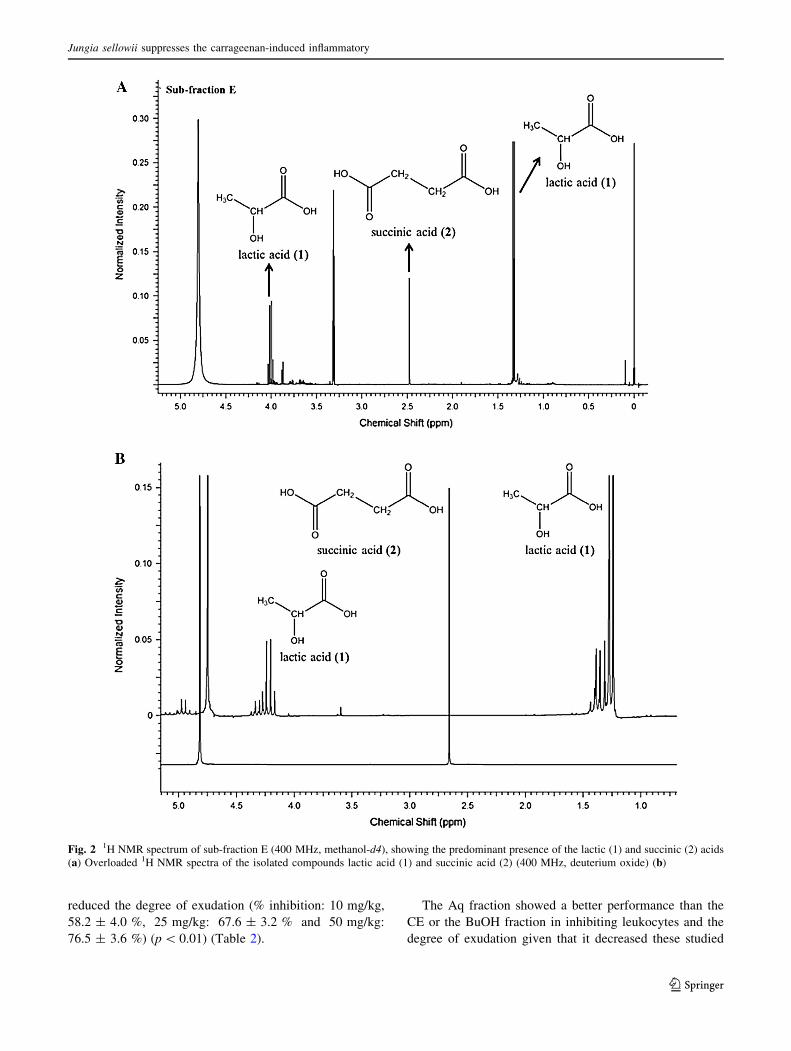
The phytochemical study of the aqueous residual sub-
fraction E from J. sellowii revealed the presence of two
major simple carboxylic acids, LA and SA, representing
57.2 and 14.3 % of this sub-fraction, respectively. These
compounds were identified using NMR spectral data, and
the proton and carbon chemical shift values were in
agreement with the published data (Williams et al. 2002;
Buzas et al. 2003; Nord et al. 2004) (Fig. 2a, b).
Inflammatory parameters analysis
It is important to note that there was no significant differ-
ences between animals treated with vehicle [0.9 % NaCl
plus dimethylsulfoxide (10 %)] in comparison with ani-
mals treated only with sterile saline (0.9 % NaCl) or
animals treated only with phosphate buffer saline (PBS, pH
7.6) upon the studied pro-inflammatory parameters
(p [ 0.05) (results not shown).
Effect of J. sellowii extract and fractions on leukocyte
content and degree of exudation
In comparison to the positive control group the CE sig-
nificantly inhibited, in a dose-dependent manner, the
leukocyte content at doses of 25 mg/kg (% inhibition:
42.8 ± 2.9 %) and 50 mg/kg (% inhibition: 66.8 ± 5.8 %)
(p \ 0.01). This inhibitory effect was associated with a
significant decrease in neutrophils (% inhibition: 25 mg/kg:
40.3 ± 3.4 % and 50 mg/kg: 65.8 ± 6.0 %) (p \ 0.01).
However, the same effect was not observed in relation to
mononuclear cells (p [ 0.05). Additionally, the CE at
doses of 10–50 mg/kg was able to reduce the degree of
exudation (% inhibition: 10 mg/kg: 31.2 ± 3.8 %, 25 mg/
kg: 51.4 ± 3.3 % and 50 mg/kg: 37.8 ± 2.8 %) (p \ 0.01)
(Table 2).
The BuOH fraction showed similar results to the CE,
inhibiting leukocytes (% inhibition: 25 mg/kg: 42.8 ±
5.3 % and 50 mg/kg: 29.8 ± 4.4 %) (p \ 0.01) and neu-
trophils (% inhibition: 25 mg/kg: 40.0 ± 5.9 % and 50 mg/
kg: 28.0 ± 4.4 %) (p \ 0.01) at the same studied doses. At
a dose of 25 mg/kg, this fraction markedly decreased
mononuclear cells (% inhibition: 25 mg/kg: 78.2 ± 4.6 %)
(p \ 0.05). All doses of this fraction (10–50 mg/kg)
Table 1 The primers and conditions used in RT-PCR reactions
Gene Primers Ampliconpb PCR
cycles
Temp. (oC)
of denaturing
Temp. (oC)
of annealing
Temp. (oC)
of extension
Genebank
register
b-actin CCAACCGTGAAAAGATGACC 616 30 95 52 72 BC138614.1
CAGTAATCTCCTTCTGCATCC
IL-1b CAGGCAGGCAGTATCACTCA 350 35 95 53 72 BC11437.1
GGCCACAGGTATTTTGTCG
TNF-a CTGGGACAGTGACCTGGACT 204 36 95 50 72 BC137720.1
CACCTCAGGGAAGAGTCTG
IL-17A TCTCTGATGCTGTTGCTGCT 195 38 95 48 72 BC119309.1
GTGGAACGGTTGAGGTAGT
iNOS GCATCCATGCAAAGAATGTG 282 40 95 51 72 NC000077.5
GCAGCCTCTTGTCTTTGACC
All PCR reactions were performed using a period of 5 min at 95 �C. The cycles were followed by a period of 7 min at 72 �C
M. Nader et al.
123
reduced the degree of exudation (% inhibition: 10 mg/kg,
58.2 ± 4.0 %, 25 mg/kg: 67.6 ± 3.2 % and 50 mg/kg:
76.5 ± 3.6 %) (p \ 0.01) (Table 2).
The Aq fraction showed a better performance than the
CE or the BuOH fraction in inhibiting leukocytes and the
degree of exudation given that it decreased these studied
Fig. 2 1H NMR spectrum of sub-fraction E (400 MHz, methanol-d4), showing the predominant presence of the lactic (1) and succinic (2) acids
(a) Overloaded 1H NMR spectra of the isolated compounds lactic acid (1) and succinic acid (2) (400 MHz, deuterium oxide) (b)
Jungia sellowii suppresses the carrageenan-induced inflammatory
123
parameters at lower doses (1–5 mg/kg). At doses of
5–25 mg/kg, this fraction reduced the leukocyte content (%
inhibition: 5 mg/kg: 49.8 ± 4.5 %, 10 mg/kg: 60.7 ±
5.2 % and 25 mg/kg: 47.4 ± 5.0 %) (p \ 0.01) and neu-
trophil content (% inhibition: 5 mg/kg: 47.8 ± 4.6 %,
10 mg/kg: 59.8 ± 5.2 % and 25 mg/kg: 45.9 ± 5.3 %)
(p \ 0.01). At doses of 1–15 mg/kg this fraction also
decreased both mononuclear cells (% inhibition: 1 mg/kg:
59.5 ± 6.9 %, 5 mg/kg: 77.2 ± 5.8 %, 10 mg/kg: 76.8 ±
4.4 % and 25 mg/kg: 70.7 ± 3.6 %) (p \ 0.05) and the
degree of exudation (% inhibition: 1 mg/kg: 58.4 ± 3.9 %,
5 mg/kg: 73.4 ± 3.2 %, 10 mg/kg: 41.4 ± 2.7 % and
25 mg/kg: 70.0 ± 3.1 %) (p \ 0.01) (Table 2).
The Hex, DCM and EtOAc fractions reduced the degree
of exudation (p \ 0.05), but did not decrease the leukocyte
content (p [ 0.05) (data not shown), and for this reason
these fractions were not analysed further.
Based on the results showed above, the Aq fraction was
chosen for further isolation of the active compounds SA
and LA.
The isolated compounds presented similar anti-inflam-
matory action patterns. SA and LA at doses of 1.0 and
2.5 mg/kg decreased the leukocyte content (% inhibition:
1 mg/kg SA: 24.9 ± 6.8 %, 2.5 mg/kg SA: 54.4 ± 2.9 %,
1 mg/kg LA: 31.2 ± 4.3 % and 2.5 mg/kg LA: 66.3 ±
5.3 %) (p \ 0.01) and neutrophils (% inhibition: 1 mg/kg
SA: 25.6 ± 6.3 %, 2.5 mg/kg SA: 53.2 ± 3.3 %, 1 mg/kg
LA: 31.8 ± 4.2 % and 2.5 mg/kg LA: 66.2 ± 5.2 %)
(p \ 0.01). These compounds also decreased mononuclear
cells at a dose of 2.5 mg/kg (% inhibition: SA:
73.9 ± 2.7 % and LA: 74.2 ± 5.6 %) (p \ 0.05). Further,
SA and LA reduced the degree of exudation at all tested
doses (% inhibition: 0.5 mg/kg SA: 15.0 ± 2.6 %, 1 mg/
kg SA: 41.7 ± 5.0 %, and 2.5 mg/kg SA: 42.9 ± 4.8 %,
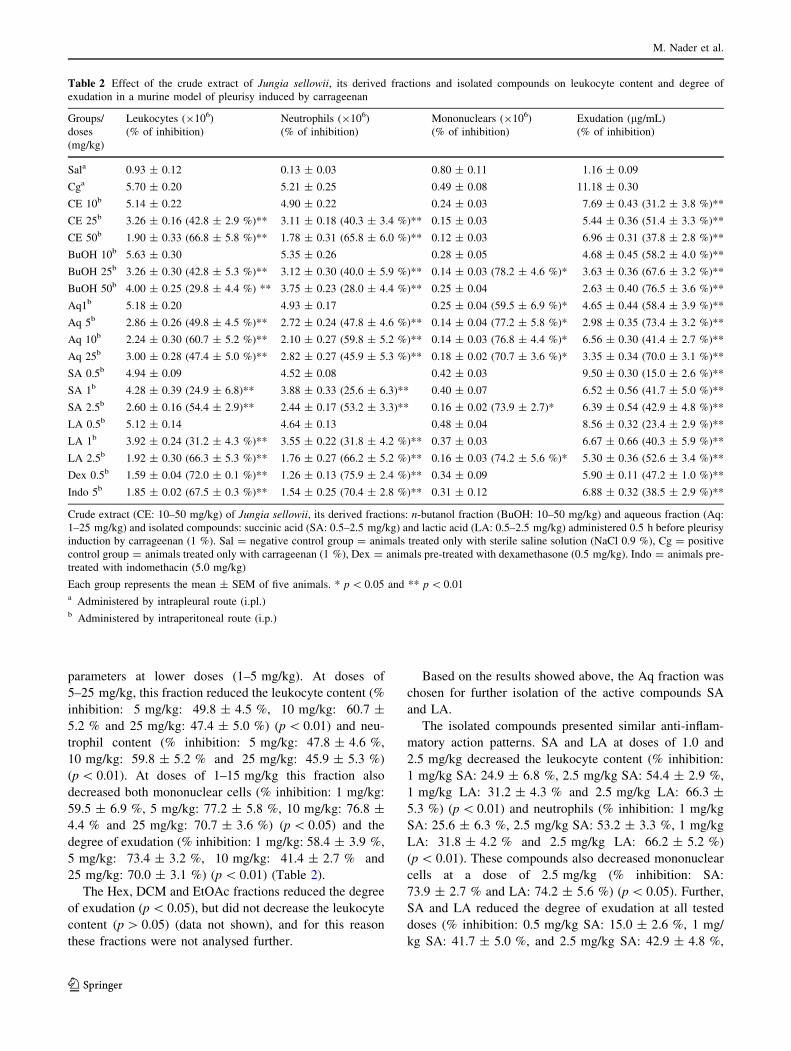
Table 2 Effect of the crude extract of Jungia sellowii, its derived fractions and isolated compounds on leukocyte content and degree of
exudation in a murine model of pleurisy induced by carrageenan
Groups/
doses
(mg/kg)
Leukocytes (9106)
(% of inhibition)
Neutrophils (9106)
(% of inhibition)
Mononuclears (9106)
(% of inhibition)
Exudation (lg/mL)
(% of inhibition)
Sala 0.93 ± 0.12 0.13 ± 0.03 0.80 ± 0.11 1.16 ± 0.09
Cga 5.70 ± 0.20 5.21 ± 0.25 0.49 ± 0.08 11.18 ± 0.30
CE 10b 5.14 ± 0.22 4.90 ± 0.22 0.24 ± 0.03 7.69 ± 0.43 (31.2 ± 3.8 %)**
CE 25b 3.26 ± 0.16 (42.8 ± 2.9 %)** 3.11 ± 0.18 (40.3 ± 3.4 %)** 0.15 ± 0.03 5.44 ± 0.36 (51.4 ± 3.3 %)**
CE 50b 1.90 ± 0.33 (66.8 ± 5.8 %)** 1.78 ± 0.31 (65.8 ± 6.0 %)** 0.12 ± 0.03 6.96 ± 0.31 (37.8 ± 2.8 %)**
BuOH 10b 5.63 ± 0.30 5.35 ± 0.26 0.28 ± 0.05 4.68 ± 0.45 (58.2 ± 4.0 %)**
BuOH 25b 3.26 ± 0.30 (42.8 ± 5.3 %)** 3.12 ± 0.30 (40.0 ± 5.9 %)** 0.14 ± 0.03 (78.2 ± 4.6 %)* 3.63 ± 0.36 (67.6 ± 3.2 %)**
BuOH 50b 4.00 ± 0.25 (29.8 ± 4.4 %) ** 3.75 ± 0.23 (28.0 ± 4.4 %)** 0.25 ± 0.04 2.63 ± 0.40 (76.5 ± 3.6 %)**
Aq1b 5.18 ± 0.20 4.93 ± 0.17 0.25 ± 0.04 (59.5 ± 6.9 %)* 4.65 ± 0.44 (58.4 ± 3.9 %)**
Aq 5b 2.86 ± 0.26 (49.8 ± 4.5 %)** 2.72 ± 0.24 (47.8 ± 4.6 %)** 0.14 ± 0.04 (77.2 ± 5.8 %)* 2.98 ± 0.35 (73.4 ± 3.2 %)**
Aq 10b 2.24 ± 0.30 (60.7 ± 5.2 %)** 2.10 ± 0.27 (59.8 ± 5.2 %)** 0.14 ± 0.03 (76.8 ± 4.4 %)* 6.56 ± 0.30 (41.4 ± 2.7 %)**
Aq 25b 3.00 ± 0.28 (47.4 ± 5.0 %)** 2.82 ± 0.27 (45.9 ± 5.3 %)** 0.18 ± 0.02 (70.7 ± 3.6 %)* 3.35 ± 0.34 (70.0 ± 3.1 %)**
SA 0.5b 4.94 ± 0.09 4.52 ± 0.08 0.42 ± 0.03 9.50 ± 0.30 (15.0 ± 2.6 %)**
SA 1b 4.28 ± 0.39 (24.9 ± 6.8)** 3.88 ± 0.33 (25.6 ± 6.3)** 0.40 ± 0.07 6.52 ± 0.56 (41.7 ± 5.0 %)**
SA 2.5b 2.60 ± 0.16 (54.4 ± 2.9)** 2.44 ± 0.17 (53.2 ± 3.3)** 0.16 ± 0.02 (73.9 ± 2.7)* 6.39 ± 0.54 (42.9 ± 4.8 %)**
LA 0.5b 5.12 ± 0.14 4.64 ± 0.13 0.48 ± 0.04 8.56 ± 0.32 (23.4 ± 2.9 %)**
LA 1b 3.92 ± 0.24 (31.2 ± 4.3 %)** 3.55 ± 0.22 (31.8 ± 4.2 %)** 0.37 ± 0.03 6.67 ± 0.66 (40.3 ± 5.9 %)**
LA 2.5b 1.92 ± 0.30 (66.3 ± 5.3 %)** 1.76 ± 0.27 (66.2 ± 5.2 %)** 0.16 ± 0.03 (74.2 ± 5.6 %)* 5.30 ± 0.36 (52.6 ± 3.4 %)**
Dex 0.5b 1.59 ± 0.04 (72.0 ± 0.1 %)** 1.26 ± 0.13 (75.9 ± 2.4 %)** 0.34 ± 0.09 5.90 ± 0.11 (47.2 ± 1.0 %)**
Indo 5b 1.85 ± 0.02 (67.5 ± 0.3 %)** 1.54 ± 0.25 (70.4 ± 2.8 %)** 0.31 ± 0.12 6.88 ± 0.32 (38.5 ± 2.9 %)**
Crude extract (CE: 10–50 mg/kg) of Jungia sellowii, its derived fractions: n-butanol fraction (BuOH: 10–50 mg/kg) and aqueous fraction (Aq:
1–25 mg/kg) and isolated compounds: succinic acid (SA: 0.5–2.5 mg/kg) and lactic acid (LA: 0.5–2.5 mg/kg) administered 0.5 h before pleurisy
induction by carrageenan (1 %). Sal = negative control group = animals treated only with sterile saline solution (NaCl 0.9 %), Cg = positive
control group = animals treated only with carrageenan (1 %), Dex = animals pre-treated with dexamethasone (0.5 mg/kg). Indo = animals pre-
treated with indomethacin (5.0 mg/kg)
Each group represents the mean ± SEM of five animals. * p \ 0.05 and ** p \ 0.01a Administered by intrapleural route (i.pl.)b Administered by intraperitoneal route (i.p.)
M. Nader et al.
123
and 0.5 mg/kg LA: 23.4 ± 2.9 %, 1 mg/kg LA:
40.3 ± 5.9 % and 2.5 mg/kg LA: 52.6 ± 3.4 %)
(p \ 0.01) (Table 2).
The reference anti-inflammatory drugs (Dex: 0.5 mg/kg
and Indo: 5 mg/kg) significantly inhibited these inflam-
matory parameters (p \ 0.01) with the exception of the
mononuclear cell migration (p [ 0.05) (Table 2).
Effect of J. sellowii on MPO and ADA activities
MPO and ADA are important markers of activated leuko-
cytes (Frode and Medeiros 2001). However, the cellular
mechanisms governing the response of these enzymes in the
mouse model of pleurisy induced by Cg have yet to be
elucidated. The effect of J. sellowii upon these enzymes can
assist in the elucidation of the cellular anti-inflammatory
action of this herb. When compared to the positive control
group, the CE (25 mg/kg), BuOH fraction (25 mg/kg), Aq
fraction (5 mg/kg) and the isolated compounds (SA: 1 mg/kg
and LA: 1 mg/kg), drastically reduced the MPO (% inhibi-
tion: CE: 60.1 ± 1.6 %, BuOH: 60.9 ± 1.3 %, Aq:
67.5 ± 1.1 %, SA: 58.8 ± 3.9 % and LA: 65.9 ± 2.8 %)
and ADA activities (% inhibition: CE: 45.2 ± 2.3 %, BuOH:
51.2 ± 8.0 %, Aq: 63.9 ± 5.8 %, SA: 37.5 ± 6.0 % and
LA: 64.4 ± 6.7 %) (p \ 0.01) (Table 3).
The anti-inflammatory drugs Dex (0.5 mg/kg) and Indo
(5 mg/kg) also inhibited MPO and ADA activities
(p \ 0.01) (Table 3).
Effect of J. sellowii on NOx levels and mRNA
expression of iNOS
Nitric oxide (NO) is known to participate in the inflammatory
process, increasing vascular permeability and inflammatory
cell infiltration (Redington 2006). As shown in Fig. 4a, NOx
levels were markedly inhibited by the CE, its derived frac-
tions and isolated compounds (p \ 0.01), with a percentage
inhibition by CE (25 mg/kg) of 40.7 ± 1.1 %,by BuOH
(25 mg/kg) of 62.0 ± 4.0 %, by Aq (5 mg/kg) of
70.4 ± 0.8 %, by SA (1 mg/kg) of 73.8 ± 2.6 % and by LA
(1 mg/kg) of 76.5 ± 1.4 % (Fig. 3a).
Furthermore, at the same conditions, this herb reduced
the mRNA expression levels of iNOS in lung tissues (%
inhibition: 25 mg/kg CE: 49.4 ± 1.6 %, 25 mg/kg BuOH:
68.5 ± 0.7 %, 5 mg/kg Aq: 56.7 ± 0.9 %, 1 mg/kg SA:
49.1 ± 1.1 % and 1 mg/kg LA: 59.1 ± 2.3 %) (p \ 0.01)
(Fig. 3b).
Dex (0.5 mg/kg) and Indo (5 mg/kg) also significantly
decreased both NOx levels and mRNA expression of iNOS
(p \ 0.01) (Fig. 3).
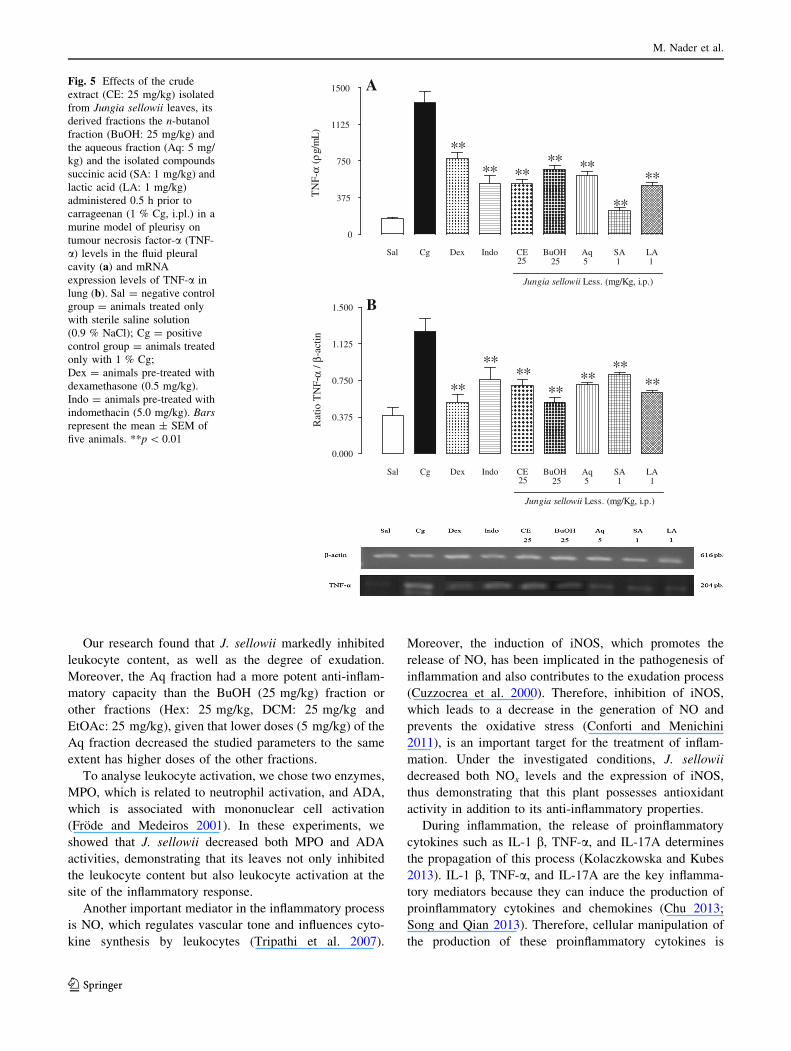
Effect of J. sellowii on of IL-1b, TNF-a and IL-17A
levels and mRNA expression levels of these cytokines
The proinflammatory cytokines IL-1b, TNF-a, and IL-17A
have an important role in leukocyte chemotaxis, and they
have been detected at high concentrations at sites of acute
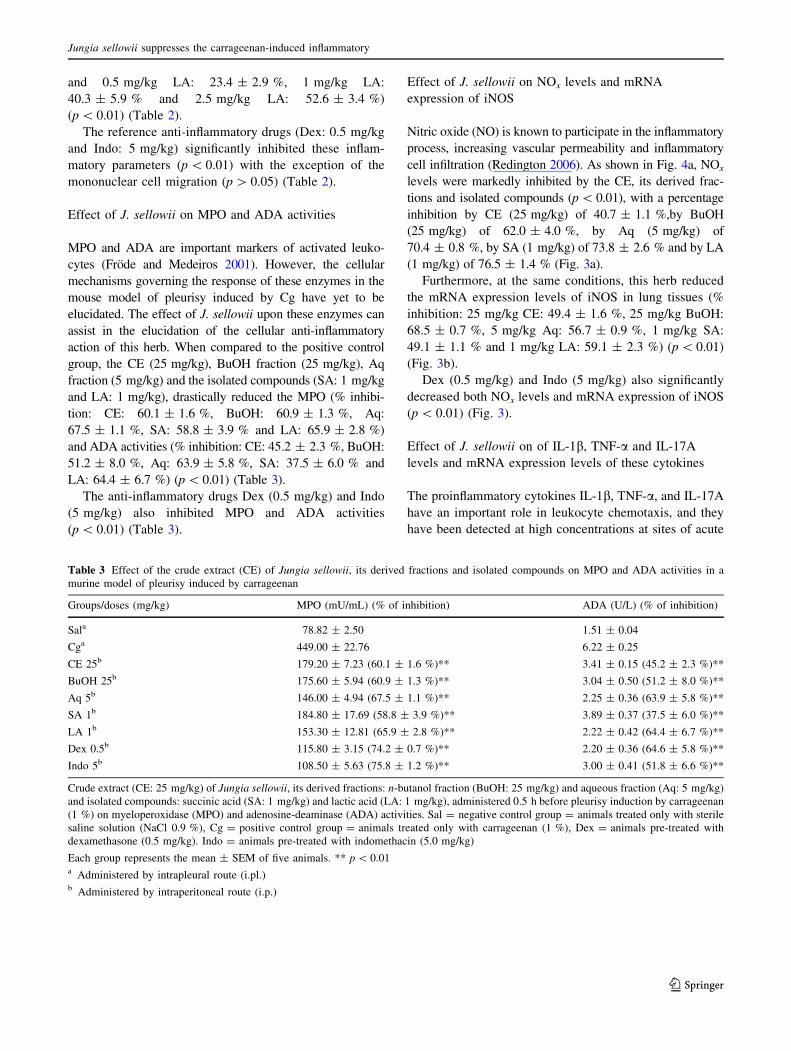
Table 3 Effect of the crude extract (CE) of Jungia sellowii, its derived fractions and isolated compounds on MPO and ADA activities in a
murine model of pleurisy induced by carrageenan
Groups/doses (mg/kg) MPO (mU/mL) (% of inhibition) ADA (U/L) (% of inhibition)
Sala 78.82 ± 2.50 1.51 ± 0.04
Cga 449.00 ± 22.76 6.22 ± 0.25
CE 25b 179.20 ± 7.23 (60.1 ± 1.6 %)** 3.41 ± 0.15 (45.2 ± 2.3 %)**
BuOH 25b 175.60 ± 5.94 (60.9 ± 1.3 %)** 3.04 ± 0.50 (51.2 ± 8.0 %)**
Aq 5b 146.00 ± 4.94 (67.5 ± 1.1 %)** 2.25 ± 0.36 (63.9 ± 5.8 %)**
SA 1b 184.80 ± 17.69 (58.8 ± 3.9 %)** 3.89 ± 0.37 (37.5 ± 6.0 %)**
LA 1b 153.30 ± 12.81 (65.9 ± 2.8 %)** 2.22 ± 0.42 (64.4 ± 6.7 %)**
Dex 0.5b 115.80 ± 3.15 (74.2 ± 0.7 %)** 2.20 ± 0.36 (64.6 ± 5.8 %)**
Indo 5b 108.50 ± 5.63 (75.8 ± 1.2 %)** 3.00 ± 0.41 (51.8 ± 6.6 %)**
Crude extract (CE: 25 mg/kg) of Jungia sellowii, its derived fractions: n-butanol fraction (BuOH: 25 mg/kg) and aqueous fraction (Aq: 5 mg/kg)
and isolated compounds: succinic acid (SA: 1 mg/kg) and lactic acid (LA: 1 mg/kg), administered 0.5 h before pleurisy induction by carrageenan
(1 %) on myeloperoxidase (MPO) and adenosine-deaminase (ADA) activities. Sal = negative control group = animals treated only with sterile
saline solution (NaCl 0.9 %), Cg = positive control group = animals treated only with carrageenan (1 %), Dex = animals pre-treated with
dexamethasone (0.5 mg/kg). Indo = animals pre-treated with indomethacin (5.0 mg/kg)
Each group represents the mean ± SEM of five animals. ** p \ 0.01a Administered by intrapleural route (i.pl.)b Administered by intraperitoneal route (i.p.)
Jungia sellowii suppresses the carrageenan-induced inflammatory
123
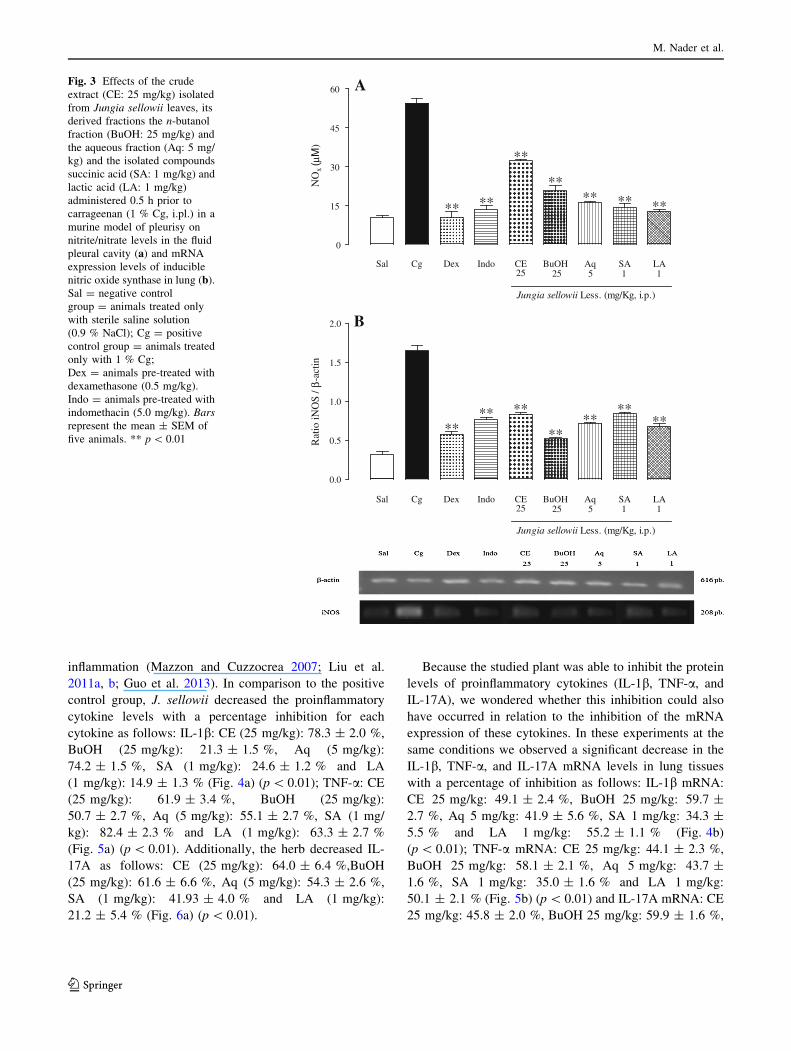
inflammation (Mazzon and Cuzzocrea 2007; Liu et al.
2011a, b; Guo et al. 2013). In comparison to the positive
control group, J. sellowii decreased the proinflammatory
cytokine levels with a percentage inhibition for each
cytokine as follows: IL-1b: CE (25 mg/kg): 78.3 ± 2.0 %,
BuOH (25 mg/kg): 21.3 ± 1.5 %, Aq (5 mg/kg):
74.2 ± 1.5 %, SA (1 mg/kg): 24.6 ± 1.2 % and LA
(1 mg/kg): 14.9 ± 1.3 % (Fig. 4a) (p \ 0.01); TNF-a: CE
(25 mg/kg): 61.9 ± 3.4 %, BuOH (25 mg/kg):
50.7 ± 2.7 %, Aq (5 mg/kg): 55.1 ± 2.7 %, SA (1 mg/
kg): 82.4 ± 2.3 % and LA (1 mg/kg): 63.3 ± 2.7 %
(Fig. 5a) (p \ 0.01). Additionally, the herb decreased IL-
17A as follows: CE (25 mg/kg): 64.0 ± 6.4 %,BuOH
(25 mg/kg): 61.6 ± 6.6 %, Aq (5 mg/kg): 54.3 ± 2.6 %,
SA (1 mg/kg): 41.93 ± 4.0 % and LA (1 mg/kg):
21.2 ± 5.4 % (Fig. 6a) (p \ 0.01).
Because the studied plant was able to inhibit the protein
levels of proinflammatory cytokines (IL-1b, TNF-a, and
IL-17A), we wondered whether this inhibition could also
have occurred in relation to the inhibition of the mRNA
expression of these cytokines. In these experiments at the
same conditions we observed a significant decrease in the
IL-1b, TNF-a, and IL-17A mRNA levels in lung tissues
with a percentage of inhibition as follows: IL-1b mRNA:
CE 25 mg/kg: 49.1 ± 2.4 %, BuOH 25 mg/kg: 59.7 ±
2.7 %, Aq 5 mg/kg: 41.9 ± 5.6 %, SA 1 mg/kg: 34.3 ±
5.5 % and LA 1 mg/kg: 55.2 ± 1.1 % (Fig. 4b)
(p \ 0.01); TNF-a mRNA: CE 25 mg/kg: 44.1 ± 2.3 %,
BuOH 25 mg/kg: 58.1 ± 2.1 %, Aq 5 mg/kg: 43.7 ±
1.6 %, SA 1 mg/kg: 35.0 ± 1.6 % and LA 1 mg/kg:
50.1 ± 2.1 % (Fig. 5b) (p \ 0.01) and IL-17A mRNA: CE
25 mg/kg: 45.8 ± 2.0 %, BuOH 25 mg/kg: 59.9 ± 1.6 %,
Sal Cg Dex Indo CE BuOH Aq SA LA
0
15
30
45
60
**** **
**
****
**
Jungia sellowii Less. (mg/Kg, i.p.)
25 525 1 1
Sal Cg Dex Indo CE BuOH Aq SA LA
Jungia sellowii Less. (mg/Kg, i.p.)
25 525 1 1
A
NO
x( μ
M)
0.0
0.5
1.0
1.5
2.0
**** **
****
****
BR
atio
iNO
S / β
-act
in
Fig. 3 Effects of the crude
extract (CE: 25 mg/kg) isolated
from Jungia sellowii leaves, its
derived fractions the n-butanol
fraction (BuOH: 25 mg/kg) and
the aqueous fraction (Aq: 5 mg/
kg) and the isolated compounds
succinic acid (SA: 1 mg/kg) and
lactic acid (LA: 1 mg/kg)
administered 0.5 h prior to
carrageenan (1 % Cg, i.pl.) in a
murine model of pleurisy on
nitrite/nitrate levels in the fluid
pleural cavity (a) and mRNA
expression levels of inducible
nitric oxide synthase in lung (b).
Sal = negative control
group = animals treated only
with sterile saline solution
(0.9 % NaCl); Cg = positive
control group = animals treated
only with 1 % Cg;
Dex = animals pre-treated with
dexamethasone (0.5 mg/kg).
Indo = animals pre-treated with
indomethacin (5.0 mg/kg). Bars
represent the mean ± SEM of
five animals. ** p \ 0.01
M. Nader et al.
123
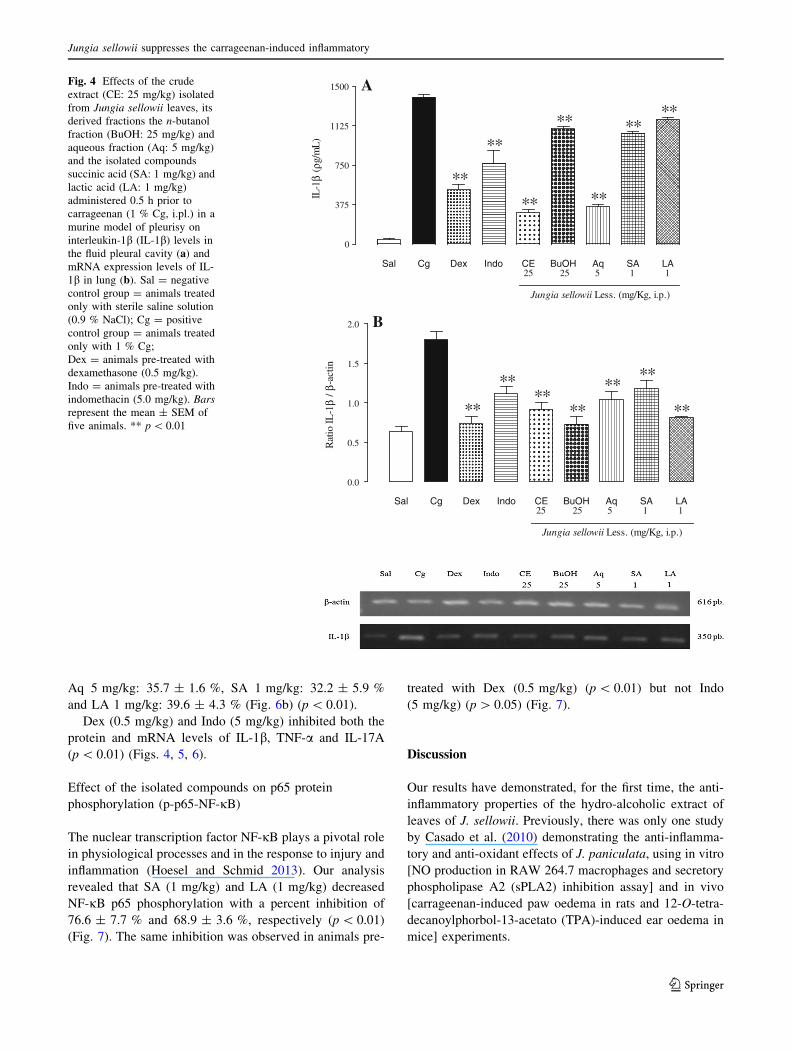
Aq 5 mg/kg: 35.7 ± 1.6 %, SA 1 mg/kg: 32.2 ± 5.9 %
and LA 1 mg/kg: 39.6 ± 4.3 % (Fig. 6b) (p \ 0.01).
Dex (0.5 mg/kg) and Indo (5 mg/kg) inhibited both the
protein and mRNA levels of IL-1b, TNF-a and IL-17A
(p \ 0.01) (Figs. 4, 5, 6).
Effect of the isolated compounds on p65 protein
phosphorylation (p-p65-NF-jB)
The nuclear transcription factor NF-jB plays a pivotal role
in physiological processes and in the response to injury and
inflammation (Hoesel and Schmid 2013). Our analysis
revealed that SA (1 mg/kg) and LA (1 mg/kg) decreased
NF-jB p65 phosphorylation with a percent inhibition of
76.6 ± 7.7 % and 68.9 ± 3.6 %, respectively (p \ 0.01)
(Fig. 7). The same inhibition was observed in animals pre-
treated with Dex (0.5 mg/kg) (p \ 0.01) but not Indo
(5 mg/kg) (p [ 0.05) (Fig. 7).
Discussion
Our results have demonstrated, for the first time, the anti-
inflammatory properties of the hydro-alcoholic extract of
leaves of J. sellowii. Previously, there was only one study
by Casado et al. (2010) demonstrating the anti-inflamma-
tory and anti-oxidant effects of J. paniculata, using in vitro
[NO production in RAW 264.7 macrophages and secretory
phospholipase A2 (sPLA2) inhibition assay] and in vivo
[carrageenan-induced paw oedema in rats and 12-O-tetra-
decanoylphorbol-13-acetato (TPA)-induced ear oedema in
mice] experiments.
Sal Cg Dex Indo CE BuOH Aq SA LA
0
375
750
1125
1500
** ****
**** **
**
Jungia sellowii Less. (mg/Kg, i.p.)
25 525 1 1
A
IL-1
β ( ρ
g/m
L)
Sal Cg Dex Indo CE BuOH Aq SA LA
0.0
0.5
1.0
1.5
2.0
Jungia sellowii Less. (mg/Kg, i.p.)
25
****
**
**
5
****
**
25 1 1
BR
atio
IL
-1β
/ β-a
ctin
Fig. 4 Effects of the crude
extract (CE: 25 mg/kg) isolated
from Jungia sellowii leaves, its
derived fractions the n-butanol
fraction (BuOH: 25 mg/kg) and
aqueous fraction (Aq: 5 mg/kg)
and the isolated compounds
succinic acid (SA: 1 mg/kg) and
lactic acid (LA: 1 mg/kg)
administered 0.5 h prior to
carrageenan (1 % Cg, i.pl.) in a
murine model of pleurisy on
interleukin-1b (IL-1b) levels in
the fluid pleural cavity (a) and
mRNA expression levels of IL-
1b in lung (b). Sal = negative
control group = animals treated
only with sterile saline solution
(0.9 % NaCl); Cg = positive
control group = animals treated
only with 1 % Cg;
Dex = animals pre-treated with
dexamethasone (0.5 mg/kg).
Indo = animals pre-treated with
indomethacin (5.0 mg/kg). Bars
represent the mean ± SEM of
five animals. ** p \ 0.01
Jungia sellowii suppresses the carrageenan-induced inflammatory
123
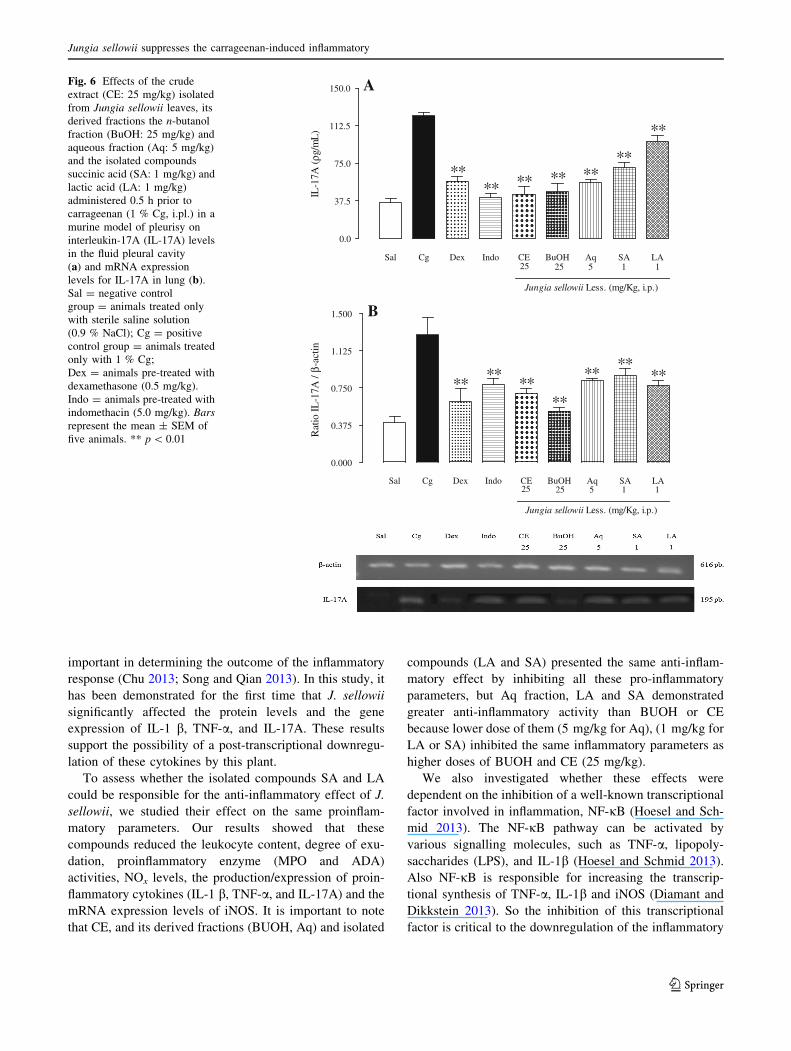
Our research found that J. sellowii markedly inhibited
leukocyte content, as well as the degree of exudation.
Moreover, the Aq fraction had a more potent anti-inflam-
matory capacity than the BuOH (25 mg/kg) fraction or
other fractions (Hex: 25 mg/kg, DCM: 25 mg/kg and
EtOAc: 25 mg/kg), given that lower doses (5 mg/kg) of the
Aq fraction decreased the studied parameters to the same
extent has higher doses of the other fractions.
To analyse leukocyte activation, we chose two enzymes,
MPO, which is related to neutrophil activation, and ADA,
which is associated with mononuclear cell activation
(Frode and Medeiros 2001). In these experiments, we
showed that J. sellowii decreased both MPO and ADA
activities, demonstrating that its leaves not only inhibited
the leukocyte content but also leukocyte activation at the
site of the inflammatory response.
Another important mediator in the inflammatory process
is NO, which regulates vascular tone and influences cyto-
kine synthesis by leukocytes (Tripathi et al. 2007).
Moreover, the induction of iNOS, which promotes the
release of NO, has been implicated in the pathogenesis of
inflammation and also contributes to the exudation process
(Cuzzocrea et al. 2000). Therefore, inhibition of iNOS,
which leads to a decrease in the generation of NO and
prevents the oxidative stress (Conforti and Menichini
2011), is an important target for the treatment of inflam-
mation. Under the investigated conditions, J. sellowii
decreased both NOx levels and the expression of iNOS,
thus demonstrating that this plant possesses antioxidant
activity in addition to its anti-inflammatory properties.
During inflammation, the release of proinflammatory
cytokines such as IL-1 b, TNF-a, and IL-17A determines
the propagation of this process (Kolaczkowska and Kubes
2013). IL-1 b, TNF-a, and IL-17A are the key inflamma-
tory mediators because they can induce the production of
proinflammatory cytokines and chemokines (Chu 2013;
Song and Qian 2013). Therefore, cellular manipulation of
the production of these proinflammatory cytokines is
Sal Cg Dex Indo CE BuOH Aq SA LA
0
375
750
1125
1500
**
******
****
**
Jungia sellowii Less. (mg/Kg, i.p.)
25 525 1 1
A
TN
F-α
( ρg/
mL
)
Sal Cg Dex Indo CE BuOH Aq SA LA
0.000
0.375
0.750
1.125
1.500
****
** ****
****
Jungia sellowii Less. (mg/Kg, i.p.)
25 525 1 1
BR
atio
TN
F -α
/ β-a
ctin
Fig. 5 Effects of the crude
extract (CE: 25 mg/kg) isolated
from Jungia sellowii leaves, its
derived fractions the n-butanol
fraction (BuOH: 25 mg/kg) and
the aqueous fraction (Aq: 5 mg/
kg) and the isolated compounds
succinic acid (SA: 1 mg/kg) and
lactic acid (LA: 1 mg/kg)
administered 0.5 h prior to
carrageenan (1 % Cg, i.pl.) in a
murine model of pleurisy on
tumour necrosis factor-a (TNF-
a) levels in the fluid pleural
cavity (a) and mRNA
expression levels of TNF-a in
lung (b). Sal = negative control
group = animals treated only
with sterile saline solution
(0.9 % NaCl); Cg = positive
control group = animals treated
only with 1 % Cg;
Dex = animals pre-treated with
dexamethasone (0.5 mg/kg).
Indo = animals pre-treated with
indomethacin (5.0 mg/kg). Bars
represent the mean ± SEM of
five animals. **p \ 0.01
M. Nader et al.
123
important in determining the outcome of the inflammatory
response (Chu 2013; Song and Qian 2013). In this study, it
has been demonstrated for the first time that J. sellowii
significantly affected the protein levels and the gene
expression of IL-1 b, TNF-a, and IL-17A. These results
support the possibility of a post-transcriptional downregu-
lation of these cytokines by this plant.
To assess whether the isolated compounds SA and LA
could be responsible for the anti-inflammatory effect of J.
sellowii, we studied their effect on the same proinflam-
matory parameters. Our results showed that these
compounds reduced the leukocyte content, degree of exu-
dation, proinflammatory enzyme (MPO and ADA)
activities, NOx levels, the production/expression of proin-
flammatory cytokines (IL-1 b, TNF-a, and IL-17A) and the
mRNA expression levels of iNOS. It is important to note
that CE, and its derived fractions (BUOH, Aq) and isolated
compounds (LA and SA) presented the same anti-inflam-
matory effect by inhibiting all these pro-inflammatory
parameters, but Aq fraction, LA and SA demonstrated
greater anti-inflammatory activity than BUOH or CE
because lower dose of them (5 mg/kg for Aq), (1 mg/kg for
LA or SA) inhibited the same inflammatory parameters as
higher doses of BUOH and CE (25 mg/kg).
We also investigated whether these effects were
dependent on the inhibition of a well-known transcriptional
factor involved in inflammation, NF-jB (Hoesel and Sch-
mid 2013). The NF-jB pathway can be activated by
various signalling molecules, such as TNF-a, lipopoly-
saccharides (LPS), and IL-1b (Hoesel and Schmid 2013).
Also NF-jB is responsible for increasing the transcrip-
tional synthesis of TNF-a, IL-1b and iNOS (Diamant and
Dikkstein 2013). So the inhibition of this transcriptional
factor is critical to the downregulation of the inflammatory
Sal Cg Dex Indo CE BuOH Aq SA LA
0.0
37.5
75.0
112.5
150.0
** ******
****
**
Jungia sellowii Less. (mg/Kg, i.p.)
25 525 1 1
A
IL-1
7A (
ρg/m
L)
Sal Cg Dex Indo CE BuOH Aq SA LA
0.000
0.375
0.750
1.125
1.500
****
****
Jungia sellowii Less. (mg/Kg, i.p.)
25 525 1 1
****
**
BR
atio
IL
-17A
/β-
actin
Fig. 6 Effects of the crude
extract (CE: 25 mg/kg) isolated
from Jungia sellowii leaves, its
derived fractions the n-butanol
fraction (BuOH: 25 mg/kg) and
aqueous fraction (Aq: 5 mg/kg)
and the isolated compounds
succinic acid (SA: 1 mg/kg) and
lactic acid (LA: 1 mg/kg)
administered 0.5 h prior to
carrageenan (1 % Cg, i.pl.) in a
murine model of pleurisy on
interleukin-17A (IL-17A) levels
in the fluid pleural cavity
(a) and mRNA expression
levels for IL-17A in lung (b).
Sal = negative control
group = animals treated only
with sterile saline solution
(0.9 % NaCl); Cg = positive
control group = animals treated
only with 1 % Cg;
Dex = animals pre-treated with
dexamethasone (0.5 mg/kg).
Indo = animals pre-treated with
indomethacin (5.0 mg/kg). Bars
represent the mean ± SEM of
five animals. ** p \ 0.01
Jungia sellowii suppresses the carrageenan-induced inflammatory
123
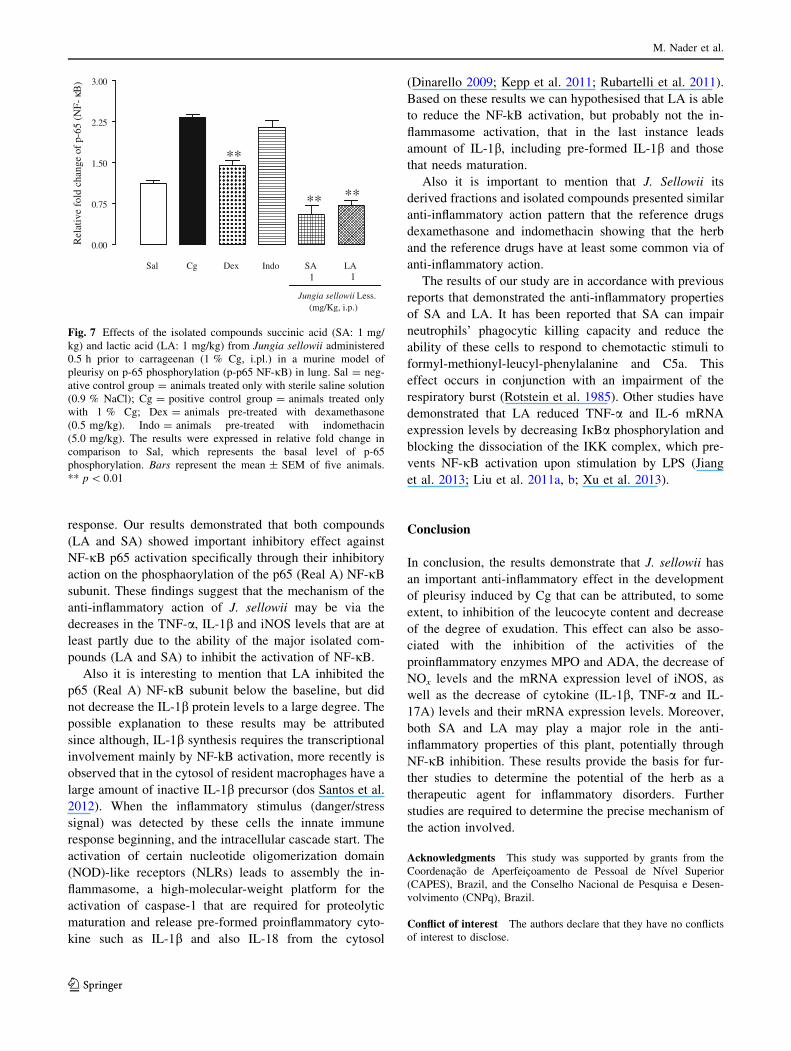
response. Our results demonstrated that both compounds
(LA and SA) showed important inhibitory effect against
NF-jB p65 activation specifically through their inhibitory
action on the phosphaorylation of the p65 (Real A) NF-jB
subunit. These findings suggest that the mechanism of the
anti-inflammatory action of J. sellowii may be via the
decreases in the TNF-a, IL-1b and iNOS levels that are at
least partly due to the ability of the major isolated com-
pounds (LA and SA) to inhibit the activation of NF-jB.
Also it is interesting to mention that LA inhibited the
p65 (Real A) NF-jB subunit below the baseline, but did
not decrease the IL-1b protein levels to a large degree. The
possible explanation to these results may be attributed
since although, IL-1b synthesis requires the transcriptional
involvement mainly by NF-kB activation, more recently is
observed that in the cytosol of resident macrophages have a
large amount of inactive IL-1b precursor (dos Santos et al.
2012). When the inflammatory stimulus (danger/stress
signal) was detected by these cells the innate immune
response beginning, and the intracellular cascade start. The
activation of certain nucleotide oligomerization domain
(NOD)-like receptors (NLRs) leads to assembly the in-
flammasome, a high-molecular-weight platform for the
activation of caspase-1 that are required for proteolytic
maturation and release pre-formed proinflammatory cyto-
kine such as IL-1b and also IL-18 from the cytosol
(Dinarello 2009; Kepp et al. 2011; Rubartelli et al. 2011).
Based on these results we can hypothesised that LA is able
to reduce the NF-kB activation, but probably not the in-
flammasome activation, that in the last instance leads
amount of IL-1b, including pre-formed IL-1b and those
that needs maturation.
Also it is important to mention that J. Sellowii its
derived fractions and isolated compounds presented similar
anti-inflammatory action pattern that the reference drugs
dexamethasone and indomethacin showing that the herb
and the reference drugs have at least some common via of
anti-inflammatory action.
The results of our study are in accordance with previous
reports that demonstrated the anti-inflammatory properties
of SA and LA. It has been reported that SA can impair
neutrophils’ phagocytic killing capacity and reduce the
ability of these cells to respond to chemotactic stimuli to
formyl-methionyl-leucyl-phenylalanine and C5a. This
effect occurs in conjunction with an impairment of the
respiratory burst (Rotstein et al. 1985). Other studies have
demonstrated that LA reduced TNF-a and IL-6 mRNA
expression levels by decreasing IjBa phosphorylation and
blocking the dissociation of the IKK complex, which pre-
vents NF-jB activation upon stimulation by LPS (Jiang
et al. 2013; Liu et al. 2011a, b; Xu et al. 2013).
Conclusion
In conclusion, the results demonstrate that J. sellowii has
an important anti-inflammatory effect in the development
of pleurisy induced by Cg that can be attributed, to some
extent, to inhibition of the leucocyte content and decrease
of the degree of exudation. This effect can also be asso-
ciated with the inhibition of the activities of the
proinflammatory enzymes MPO and ADA, the decrease of
NOx levels and the mRNA expression level of iNOS, as
well as the decrease of cytokine (IL-1b, TNF-a and IL-
17A) levels and their mRNA expression levels. Moreover,
both SA and LA may play a major role in the anti-
inflammatory properties of this plant, potentially through
NF-jB inhibition. These results provide the basis for fur-
ther studies to determine the potential of the herb as a
therapeutic agent for inflammatory disorders. Further
studies are required to determine the precise mechanism of
the action involved.
Acknowledgments This study was supported by grants from the
Coordenacao de Aperfeicoamento de Pessoal de Nıvel Superior
(CAPES), Brazil, and the Conselho Nacional de Pesquisa e Desen-
volvimento (CNPq), Brazil.
Conflict of interest The authors declare that they have no conflicts
of interest to disclose.
Sal Cg Dex Indo SA LA
0.00
0.75
1.50
2.25
3.00
** **
**
Jungia sellowii Less. (mg/Kg, i.p.)
1 1
Rel
ativ
e fo
ld c
hang
e of
p-6
5 (N
F-κB
)
Fig. 7 Effects of the isolated compounds succinic acid (SA: 1 mg/
kg) and lactic acid (LA: 1 mg/kg) from Jungia sellowii administered
0.5 h prior to carrageenan (1 % Cg, i.pl.) in a murine model of
pleurisy on p-65 phosphorylation (p-p65 NF-jB) in lung. Sal = neg-
ative control group = animals treated only with sterile saline solution
(0.9 % NaCl); Cg = positive control group = animals treated only
with 1 % Cg; Dex = animals pre-treated with dexamethasone
(0.5 mg/kg). Indo = animals pre-treated with indomethacin
(5.0 mg/kg). The results were expressed in relative fold change in
comparison to Sal, which represents the basal level of p-65
phosphorylation. Bars represent the mean ± SEM of five animals.
** p \ 0.01
M. Nader et al.
123

References
Buzas M, Chira N, Deleanu C, Rosca S (2003) Identification and
quantitative measurement by 1H-NMR spectroscopy of several
compounds present in Romanian wines. Rev Chim 54:831–833
Casado R, Landal A, Calvol JJ, Del Terencio MDC, Calvo MI (2010)
Anti-inflammatory and antioxidant activities of Jungia panicu-
lata. Pharm Biol 48:897–905
Chu W (2013) Tumor necrosis factor. Cancer Lett 328:222–225
Conforti F, Menichini F (2011) Phenolic compounds from plants as
nitric oxide production inhibitors. Curr Med Chem
18:1137–1145
Cuzzocrea S, Mazzon E, Calabro G, Dugo L, De Sarro A, Van De
Loo FAJ, Caputi AP (2000) Inducible nitric oxide synthase—
knockout mice exhibit resistance to pleurisy and lung injury
caused by carrageenan. Am J Respir Crit Care Med
162:1859–1866
De-La-Cruz H, Vilcapoma G, Zevallos PA (2007) Ethnobotanical
study of medicinal plants used by the Andean people of Canta,
Lima, Peru. J Ethnopharmacol 111:284–294
Diamant G, Dikkstein R (2013) Transcriptional control by NF-kappa
B: elongation in focus. Biochem Biophys Acta 1829:937–945
Dinarello CA (2009) Immunological and inflammatory functions of
the interleukin-1 family. Annu Rev Immunol 27:519–550
dos Santos G, Kutuzov MA, Ridge KM (2012) The inflammasome in
lung diseases. Am J Physiol Lung Cell Mol Physiol
303(8):L627–L633
Frode TS, Medeiros YS (2001) Myeloperoxidase and adenosine-
deaminase levels in the pleural fluid leakage induced by
carrageenan in the mouse model of pleurisy. Mediat Inflamm
10:223–227
Giusti G, Galanti B (1984) Adenosine deaminase: colorimetric
method. In: Bergmeyer HU (ed) Methods of enzymatic analysis.
Verlang Chemie Press, Weinheim, pp 315–323
Goleniowski ME, Bongiovanni GA, Palacio L, Nunez CO, Cantero JJ
(2006) Medicinal plants from the ‘‘Sierra de Comechigones’’,
Argentina. J Ethnopharmacol 107:324–341
Green IC, Wagner DA, Glowski J, Skipper PL, Wishnok JS,
Tannenbaum SB (1982) Analysis of nitrate, nitrite and [15N]
nitrate in biological fluids. Anal Biochem 126:131–138
Guo J, Gu N, Chen J, Shi T, Zhou Y, Rong Y, Zhou T, Yang W, Gui
X, Chen W (2013) Neutralization of interleukin-1 beta attenuates
silica-induced lung inflammation and fibrosis in C57BL/6 mice.
Arch Toxicol 87:1963–1973
Hammond GB, Fernandes ID, Villegas LF, Vaisberg AJ (1998) A
survey of traditional medicinal plants from the Callejon de
Huaylas, Department of Ancash, Peru. J Ethnopharmacol
61:17–30
Hoesel B, Schmid J (2013) The complexity of NF-jB signaling in
inflammation and cancer. Mol Cancer 12(86):1–15
Jiang J, Xu G, Shi Y, Qiao Y, Hu G, Ren X (2013) Lactic acid inhibits
lipopolysaccharide-induced translocation of NF-jB p65 from
cytoplasm to nucleus and transcription of nuclear factor-jB p65
and cyclooxygenase 2. AAS 44:204–209
Katinas L, Pruski J, Sancho G, Tellerıa MC (2008) The subfamily
Mutisioideae (Asteraceae). Bot Rev 74:469–716
Kepp O, Galluzzi L, Kroemer G (2011) Mitochondrial control of the
NLRP3 inflammasome. Nat Immunol 12:199–200
Kolaczkowska E, Kubes P (2013) Neutrophil recruitment and
function in health and inflammation. Nat Rev Immunol
13:159–175
Liu J, Xue J, Zhu Z, Hu G, Ren X (2011a) Lactic acid inhibits NF-jB
activation by lipopolysaccharide in rat intestinal mucosa micro-
vascular endothelial cells. ASC 10:954–959
Liu Y, Mei J, Gonzales L, Yang G, Dai N, Wang P, Zhang P, Favara
M, Malcom KC, Guttentag S, Worthen GS (2011b) IL-17A and
TNF-a exert synergistic effects on expression of CXCL5 by
alveolar type II cells in vivo and in vitro. J Immunol
186:3197–3205
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein
measurement with the folin phenol reagent. J Biol Chem
193:265–267
Marchant R, Almeida L, Behling H, Berrrio JC, Bush M, Cleef A,
Duivenvoorden J, Kappelle M, De Oliveira P, De Oliveira-Filho
AT, Lozano-Garcia S, Hooghiemstra H, Ledru M, Ludlow-
Wiechers B, Markgraf V, Mancini V, Paez M, Prieto A, Rangel
O, Salgado-Labouriau ML (2002) Distribution and ecology of
parent taxa of pollen lodged within the Latin American Pollen
Database. Rev Palaeobot Palynol 21:1–75
Mazzon E, Cuzzocrea S (2007) Role of TNF-a in lung tight junction
alteration in mouse model of acute lung inflammation. Respir
Res 8:1–19
National Research Council (1989) Roots and tubers in lost crops of
the Incas: Little known plants of the Andes with promise for
worldwide cultivation. National Academy Press, Washington,
pp 67–73
Nord LI, Vaag P, Duus J (2004) Quantification of organic and amino
acids in beer by 1H NMR spectroscopy. Anal Chem
76:4790–4798
Rao TS, Currie JL, Shaffer AF, Isakson PC (1993) Comparative
evaluation of arachidonic acid (AA)-and tetradecanoylphorbol
acetate (TPA)-induced dermal inflammation. Inflammation
17:723–741
Redington AE (2006) Modulation of nitric oxide pathways: thera-
peutic potential in asthma and chronic obstructive pulmonary
disease. Eur J Pharmacol 533:263–276
Rehecho S, Uriarte-Pueyo I, Calvo J, Vivas LA, Calvo MI (2011)
Ethnopharmacological survey of medicinal plants in Nor-
Yauyos, a part of the Landscape Reserve Nor-Yauyos-Cochas,
Peru. J Ethnopharmacol 133:75–85
Rotstein OD, Pruett TL, Firgel VD, Nelson RD, Simmons RL (1985)
Succinic acid, a metabolic by-product of Bacteroides species,
inhibits polymorfonuclear leukocyte function. Infect Immun
48:402–408
Rubartelli A, Gattorno M, Netea MG, Dinarello CA (2011) Interplay
between redox status and inflammasome activation. Trends
Immunol 32:559–566
Saleh TS, Calixto JB, Medeiros YS (1996) Anti-inflammatory effects
of theophylline, cromolyn and salbutamol in a murine model of
pleurisy. Br J Pharmacol 118:811–819
Song X, Qian Y (2013) IL-17 family cytokines mediated signaling in
the phatogenesis of inflammatory diseases. Cell Signal
25:2335–2347
Tripathi P, Tripathi LK, Singh V (2007) The role of nitric oxide in
inflammatory reactions. FEMS Immunol Med Microbiol
51:443–452
Williams LN, Petterson KA, Roberts JD (2002) The Conformations of
1,4-butanedioic acid as a function of solvent polarity in a series
of alcohols as determined by NMR spectroscopy. J Phys Chem A
106:7491–7493
Xu G, Jiang J, Wang M, Li J, Su J, Ren X (2013) Lactic acid reduced
LPS-induced TNF-a and IL-6 mRNA levels through decreasing
IjBa phosphorylation. JIA 12:1073–1078
Jungia sellowii suppresses the carrageenan-induced inflammatory
123




















