
juliana frança dos reis costa
157
JULIANA FRANÇA DOS REIS COSTA Influência dos leucócitos do colostro no desenvolvimento da microbiota intestinal, resposta imune inata e incidência de diarreias em bezerras recém-nascidas São Paulo 2016
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of juliana frança dos reis costa

JULIANA FRANÇA DOS REIS COSTA
Influência dos leucócitos do colostro no desenvolvi mento da
microbiota intestinal, resposta imune inata e incid ência de diarreias
em bezerras recém-nascidas
São Paulo
2016

JULIANA FRANÇA DOS REIS COSTA
Influência dos leucócitos do colostro no desenvolvi mento da
microbiota intestinal, resposta imune inata e incid ência de
diarreias em bezerras recém-nascidas
Dissertação apresentada ao Programa de Pós-Graduação em Clínica Veterinária da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para a obtenção do título de Mestre em Ciências
Departamento:
Clínica Médica
Área de concentração:
Clínica Veterinária
Orientadora:
Profa. Dra. Viviani Gomes
São Paulo
2016

Autorizo a reprodução parcial ou total desta obra, para fins acadêmicos, desde que citada a fonte.
DADOS INTERNACIONAIS DE CATALOGAÇÃO NA PUBLICAÇÃO
(Biblioteca Virginie Buff D’Ápice da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo)
T.3332 Costa, Juliana França dos Reis FMVZ Influência dos leucócitos do colostro no desenvolvimento da microbiota intestinal,
resposta imune inata e incidência de diarreias em bezerras recém-nascidas / Juliana França dos Reis Costa. -- 2016.
156 f. : il. Dissertação (Mestrado) - Universidade de São Paulo. Faculdade de Medicina
Veterinária e Zootecnia. Departamento de Clínica Médica, São Paulo, 2016.
Programa de Pós-Graduação: Clínica Veterinária. Área de concentração: Clínica Veterinária.
Orientador: Profa. Dra. Viviani Gomes.
1. Colostro congelado. 2. Colostro fresco. 3. Contagem bacteriana. 4. Neutrófilos.5. Fagocitose. I. Título.


FOLHA DE AVALIAÇÃO
Autor: COSTA, Juliana França dos Reis
Título: Influência dos leucócitos do colostro no desenvolvi mento da microbiota
intestinal, resposta imune inata e incidência de di arreias em bezerras
recém-nascidas
Dissertação apresentada ao Programa de Pós-Graduação em Clínica Veterinária da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção do titulo de Mestre em Ciências
Data: _____/_____/_____
Banca Examinadora
Prof. Dr._____________________________________________________________
Instituição:__________________________ Julgamento:_______________________
Prof. Dr._____________________________________________________________
Instituição:__________________________ Julgamento:_______________________
Prof. Dr._____________________________________________________________
Instituição:__________________________ Julgamento:_______________________

DEDICATÓRIA
Dedico este trabalho a Deus que sempre guiou
meus passos, permitindo-me ter saúde e força para
enfrentar todos os obstáculos impostos pela vida e
também por me cobrir de bênçãos e alegrias.
À minha mãe Sylvia que nunca mediu esforços para
me ajudar em todos os momentos da minha vida; ao
meu pai Esio, que de onde estiver sei que sente
imenso orgulho de mim; ao meu irmão Bruno, que
desempenhou papel importante ao me guiar nos
primeiros estudos, me ensinando muito.
Ao meu marido Thiago, pelo amor, carinho e
companheirismo que me inspira e motiva
diariamente. Juntos, iremos sempre mais longe.
À minha orientadora Profa. Dra. Viviani Gomes, por
toda determinação e excelência com que
desempenha seu trabalho diariamente.

AGRADECIMENTOS
Agradeço a Deus pela minha saúde... pela incrível força, paz e serenidade que me
fez seguir adiante nos momentos de dificuldade e por ser tão bom e generoso ao
proporcionar inúmeras realizações e alegrias em minha vida.
À minha mãe, Sylvia de França Guimarães, companheira e melhor amiga de uma
vida inteira. Obrigada por todas as ajudas, pelos nossos momentos “mãe e filha”, por
me orientar, aconselhar, me salvar sempre que precisei de última hora, jamais
medindo esforços para isso. Muito obrigada por sacrificar a sua vida, pela minha.
Serei eternamente grata e espero conseguir ser uma mãe tão boa para meus filhos!
Ao meu pai, Esio dos Reis Filho, que me deixou em meio à correria do experimento,
em que tivemos a oportunidade de nos vermos pela última vez num domingo, dentro
do laboratório da FMVZ, na sala do fluxo já que infelizmente estava muito difícil
conciliar tempo para ir visitá-lo. Me desculpe pelos momentos de ausência! Sei do
tanto que valorizava o estudo e principalmente a USP. Sei que você foi embora
orgulhoso do que viu por mais que, apesar de médico, tenha achado “complexo
demais” acompanhar todo este avanço. Obrigada por ter exigido tanto de mim
durante a vida inteira. Inúmeras vezes eu achei essa cobrança excessiva; mas, sem
isso, talvez eu não tivesse chegado até aqui. Obrigada! Saudades, pai!
Ao meu irmão, Bruno França dos Reis, a pessoa mais inteligente que já conheci,
meu grande orgulho. Obrigada por ter dividido comigo sempre seu imenso
conhecimento sobre tudo e ter me ensinado sobre tantas coisas nesta vida.
Obrigada pela amizade, parceria e irmandade que nos une e unirá eternamente.
Ao meu querido marido Thiago João Costa, meu grande companheiro, amor da
minha vida. Obrigada por tudo. Por sempre me incentivar a crescer, estar ao meu
lado e por valorizar cada conquista. Você é um exemplo de profissional,
comprometido, batalhador e merecedor de todas as suas conquistas. Obrigada por
entender e aceitar sempre que precisei estar longe de casa, por ser um marido tão

dedicado à nossa família e por estarmos juntos construindo, dia após dia, cada um
dos nossos sonhos. Te amo muito!
À minha orientadora, Profa. Dra. Viviani Gomes, primeiramente pela oportunidade
que me proporcionou ao abrir as portas de sua equipe desde a Iniciação Científica
em 2013, sem ainda me conhecer muito bem, proporcionando-me não apenas esta
conquista, mas também inúmeras outras oportunidades nesta trajetória. Obrigada
pela confiança depositada em mim e no meu trabalho. Você é um exemplo de
persistência e a prova de que com muito empenho, força de vontade e dedicação
chegaremos lá! Muito obrigada!
Ao ilustre diretor da FMVZ/USP Prof. Dr. José Antonio Visintin e aos admirados
professores do Departamento de Clínica Médica: Profa. Dra. Alice Maria Melville
Paiva Della Libera, Prof. Dr. Archivaldo Reche Junior, Profa. Dra. Carla Bargi Belli,
Prof. Dr. Carlos Eduardo Larsson, Profa. Dra. Denise Saretta Schwartz, Prof. Dr.
Enrico Lippi Ortolani, Prof. Dr. Fábio Celidonio Pogliani, Prof. Dr. Fernando José
Benesi, Profa. Dra. Lílian Gregory, Profa. Dra. Márcia de Oliveira Sampaio Gomes,
Profa. Dra. Marcia Mery Kogika, Profa. Dra. Maria Cláudia Araripe Sucupira, Profa.
Dra. Maria Helena Matiko Akao Larsson, Profa. Dra. Mitika Kuribayashi Hagiwara,
Profa. Dra. Raquel Yvona Arantes Baccarin, Profa. Dra. Silvia Regina Ricci, Profa.
Dra. Viviani Gomes e Prof. Dr. Wilson Roberto Fernandes.
À Profa. Dra. Karina Medici Madureira pela parceria, colaboração nas análises
laboratoriais e artigos, bem como pelos momentos de descontração em suas vindas
à FMVZ/USP. Ao Jean Silva Ramos pela prontidão em ajudar a todos à sua volta.
Obrigada!
À Rejane dos Santos Sousa pela ajuda nas análises de cortisol e por ser sempre
atenciosa com a nossa equipe.
Ao Prof. Dr. David John Hurley, da "University of Georgia" por auxiliar nas ideias,
técnicas e análises da proposta inicial deste trabalho, além de transmitir seus
conhecimentos e sugestões sempre que solicitamos sua ajuda. Obrigada por me

receber em sua Universidade durante o pós doc da professora Viviani e me mostrar
um pouco da fascinante realidade da pesquisa nos Estados Unidos.
À Profa. Dra. Alice Maria Melville Paiva Della Libera pelo carinho comigo nesses
anos, e por sempre nos mostrar que é possível buscar um equilíbrio saudável entre
a vida pessoal e uma carreira de sucesso. À Profa. Dra. Carla Maria Bittar por já ter
contribuído bastante com seu conhecimento à nossa equipe, e agora agradeço por
terem aceito o convite para compor a banca desta dissertação.
Aos colegas de equipe: Sylvia Marquart Fontes Novo e Camila Costa Baccili além de
excelentes profissionais, por terem sido maravilhosos seres humanos ao agarrarem
como de vocês toda a minha parte a campo do experimento, no momento em que
precisei me ausentar pelo súbito falecimento do meu pai. Agradeço do fundo do meu
coração, pois vocês foram além do limite para me ajudar nesse momento tão difícil e
só por isso foi possível finalizar este trabalho com a máxima qualidade. Agradeço
também à nossa orientadora que mesmo estando em seu pós doc nos EUA, soube
nos auxiliar e conduzir esta situação, ajudando da melhor forma a todos. Muito
obrigada por, acima de tudo, terem demonstrado apoio, carinho e amizade!
Aos membros da equipe que, mesmo já tendo saído, tiveram participação
importante em minha trajetória: Cynthia Pereira da Costa e Silva, Vinicius Alvim
Passos Baldacim pelos saudosos momentos do IC e Bruno Toledo Silva, muito
obrigada! Aos mais novos: Camila Cecília Martin obrigada pelo apoio, convívio
alegre e diário e Natalia Meirelles Sobreira pela ajuda com os animais na fazenda.
Boa sorte nesta fase que se inicia! A todos, muito sucesso!
Stephanie Blima e Milena Maia, obrigada pela ajuda durante o experimento. Pamella
Lorenci, amiga de tantos anos, obrigada pela ajuda na fazenda e por me mostrar
que tudo sempre tem o lado bom. À Ana Pérola por toda a atenção, pelas inúmeras
vezes que me ajudou e por ser muito querida comigo. À Karen Nascimento da Silva,
muito obrigada pela ajuda na reta final da dissertação. Ao nosso padrinho Luciano
Donizeti Varanda pela amizade, companheirismo e ensinamentos em todos os
momentos.

Aos colegas do Programa de Pós-Graduação, boa sorte a todos!
À minha família, amigos do Colégio Santa Cruz (G8) e amigas da Universidade
Anhembi Morumbi, muito obrigada pelo carinho, por me apoiarem, me ajudarem, me
darem força e coragem mesmo sem entender inúmeras vezes o que eu realmente
fazia por aqui. Obrigada por aceitarem e respeitarem os momentos em que eu
precisei me ausentar e por nunca me cobrarem por isso. À Franciane Corrêa
Cardoso que desde a minha graduação me ensina muito na área da veterinária e,
hoje em dia, principalmente, sobre a vida. Amo vocês! Obrigada a todos!
À biomédica Cláudia Regina Stricagnolo pela fundamental e inigualável ajuda
durante o período em que estive aqui, desde a Iniciação Científica até o mestrado.
Muito obrigada por todas as bactérias identificadas, placas na estufa, empréstimos
de material e companhia nas tardes de laboratório.
À Samantha Ive Miyashiro por toda a ajuda com o citômetro e pelo carinho de
sempre! Muito obrigada! À Dinha por tornar o ambiente mais alegre e engraçado,
além de manter sempre os materiais impecáveis para a nossa equipe! Obrigada! À
toda equipe dos Laboratórios da FMVZ/USP, Clara Mori, Maria Helena da Silva
Pelissari e Marly Elizabeth Ferreira. À Nicolle Gilda Teixeira de Queiroz
Hazarbassanov do Laboratório do Departamento de Patologia pelos conhecimentos
transmitidos, sempre pronta a ajudar desde a época da Iniciação.
Gostaria de agradecer a toda a equipe da Fazenda Colorado que nos abriu as portas
para que o experimento pudesse ser realizado lá. Aos veterinários Natália Meirelles
Sobreira e Sergio Soriano, além de todos os funcionários que colaboraram direta ou
indiretamente, muito obrigada!
Aos residentes da Clínica de Bovinos e Pequenos Ruminantes (CBPR) de 2013 a
2016, muito obrigada! Todos vocês sempre nos ajudaram prontamente em algum
momento! Em especial aos residentes Rodrigo, Camila, Marcela, Joel, Mailson e
Giuliana que nos ajudaram nas coletas do mestrado. Muito obrigada e sucesso para
vocês!

À Fundação de Amparo à Pesquisa do Estado de São Pa ulo - FAPESP pela
concessão do auxílio de pesquisa (2013/06152-7).

“O saber a gente aprende com os mestres e os livros. A sabedoria, se
aprende é com a vida e com os humildes.”
Cora Coralina
“Seja quem você for, seja qualquer posição que você tenha na vida,
do nível altíssimo ao mais baixo, tenha sempre como meta muita força,
muita determinação e sempre faça tudo com muito amor
e com muita fé em Deus que um dia você chega lá,
de alguma maneira você chega lá...”
Ayrton Senna

RESUMO
COSTA, J. F. R. Influência dos leucócitos do colostro no desenvolv imento da microbiota intestinal, resposta imune inata e incid ência de diarreias em bezerras recém-nascidas. [The influence of colostrum leukocytes in the development of intestinal microbiota, innate immune response, and incidence of diarrhea in newborn calves]. 2016. 156 f. Dissertação (Mestrado em Ciências) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2016.
O objetivo geral deste trabalho foi avaliar a influência dos leucócitos do colostro
bovino na imunidade inata, desenvolvimento da microbiota intestinal e ocorrência de
diarreias em bezerras Holandesas recém-nascidas. Para isso, 20 bezerras
Holandesas foram acompanhadas nos seguintes momentos: antes da mamada do
colostro (D0); 1-2 (D2); 7 (D7); 14 (D14); 21 (D21) e 28 (D28) dias após nascimento
e foram distribuídas em dois grupos experimentais: grupo COL+ recebeu colostro
fresco (4L) proveniente de suas respectivas mães; e grupo COL- recebeu colostro
congelado e acelular (4L) de vacas doadoras. Capítulo 1 – O objetivo deste capítulo
foi avaliar a presença e influência dos leucócitos do colostro na colonização do trato
gastrintestinal. Para isso, amostras de colostro foram semeadas em TSA e
MacConkey para CBT e CCT; e amostras de fezes em agar sangue, MacConkey e
Salmonella-Shiguella. Bezerras apresentaram maior frequência de diarreia no D14
(COL+=78%; COL-=60%). O grupo COL- apresentou maior CBT e CCT/mL
(3,93x106 ufc/mL e 3,01x105 ufc/mL) em relação ao COL+ (0,94x106 ufc/mL e
0,78x105ufc/mL). Sobre as espécies bacterianas isoladas, as que foram mais
frequentes no COL+ foram Proteus mirabilis (29,91%), Escherichia coli (28,04%),
Citrobacter freundii (5,61%) e Staphylococcus spp (5,61%), no COL- foram
Escherichia coli (28,70%), Proteus mirabilis (27,78%), Klebsiella pneumoniae
(11,32%) e Morganella morganii (5,66%). O momento com maior isolamento em
ambos os grupos foi o D2 (COL+ = 26 cepas; COL- = 27 cepas), período em que
iniciou a diarreia nas bezerras. Não foi possível detectar diferenças entre as
frequências de microrganismos entre os grupos COL+ e COL-. A administração de
colostro COL+ e COL- não influenciou na proporção de bactérias aeróbias presentes
nas fezes e na ocorrência de diarreia das bezerras durante o período neonatal.
Capítulo 2 – O objetivo deste capítulo foi avaliar a influência dos leucócitos do
colostro na resposta imune inata em bezerras Holandesas. Os animais foram

submetidos ao exame clínico, seguido da colheita das amostras sanguíneas para
realização das provas laboratoriais. As concentrações de cortisol sérico e
haptoglobina não apresentaram diferenças estatísticas, enquanto a concentração de
ferro foi diferente entre os grupos no D7. No leucograma não foi detectada diferença
para os leucócitos totais e neutrófilos absolutos, mas houve tendência entre os
grupos no D7 para os valores de neutrófilos relativos. O marcador CH138+CD62L+
não apresentou diferença entre os grupos, apesar da maior expressão ter sido
detectada no COL-. A expressão de CH138+CD62L- foi maior no COL+ no D14. A
fagocitose e produção de H2O2 pelos neutrófilos sanguíneos apresentou perfil
semelhante em relação ao S. aureus e E. coli entre os grupos, apesar do COL-
apresentar maior intensidade de fagocitose do D7 ao D14. Houve tendência para o
aumento da proporção de granulócitos liberando H2O2 basal no D14 para o COL-,
entretanto o COL + apresentou tendência para maior intensidade da fagocitose no
D21. A proporção de granulócitos (%), estimulados com S. aureus, liberando H2O2
foi maior no COL+ em D7, enquanto o estímulo com a E. coli não resultou em
diferenças entre os grupos. A intensidade de fluorescência foi maior e gradual no
COL+ quando as células foram estimuladas, porém não foram encontradas
diferenças estatísticas. As células do colostro influenciaram no perfil sanitário
apresentado pelas bezerras, observando-se maior intensidade de diarreias, menores
teores de ferro sérico e anemias no grupo que recebeu colostro congelado. A
migração dos granulócitos sanguíneos foi mais rápida e intensa no COL+ em relação
ao COL-, após exposição natural aos patógenos causadores de diarreia. O índice de
fagocitose (%) dos granulócitos apresentou semelhança entre os grupos, entretanto
a intensidade da fluorescência foi mais intensa no COL-. Em contrapartida, o COL+
demonstrou maior habilidade em produzir H2O2.
Palavras-chave: Colostro congelado. Colostro fresco. Contagem bacteriana.
Neutrófilos. Fagocitose.

ABSTRACT
COSTA, J. F. R. The influence of colostrum leukocytes in the devel opment of intestinal microbiota, innate immune response, and incidence of diarrhea in newborn calves. [Influência dos leucócitos do colostro no desenvolvimento da microbiota intestinal, resposta imune inata e incidência de diarreias em bezerras recém-nascidas]. 2016. 156 f. Dissertação (Mestrado em Ciências) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2016.
The aim of this study was to evaluate the influence of bovine colostrum leukocytes in
innate immunity, in the development of the intestinal microbiota and the occurrence
of diarrhea in newborn Holstein calves. For this, 20 Holstein calves were followed in
the following moments: before colostrum intake (D0); 1-2 (D2); 7 (D7); 14 (D14); 21
(D21) and 28 (D28) days after birth. They were divided into two experimental groups:
COL+ group, which received fresh colostrum (4L) from their mothers; and COL-
group which received frozen and acellular colostrum (4L) of donor cows. Chapter 1 -
The aim of this chapter was to evaluate the presence and influence of colostrum
leukocytes in the colonization of the gastrointestinal tract. For this, colostrum samples
were seeded in TSA and MacConkey for TPC and TCC; and stool samples on blood
agar, MacConkey and Salmonella-Shiguella. Calves presented higher frequency of
diarrhea in D14 (COL+ = 78%; COL- = 60%). The COL- group showed higher TPC
and TCC/mL (3,93x106cfu/mL and 3,01x105 cfu/mL) compared to COL+ (0,94x106
cfu/mL and 0,78x105 cfu/mL). Regarding the isolated bacterial species, those that
were more frequent in the COL+ were Proteus mirabilis (29.91%), Escherichia coli
(28.04%), Citrobacter freundii (5.61%) and Staphylococcus spp (5.61%); in the COL-
were Escherichia coli (28.70%), Proteus mirabilis (27.78%), Klebsiella pneumoniae
(11.32%) and Morganella morganii (5.66%). The moment with more insolation in both
groups was the D2 (COL+ = 26 strains; COL- = 27 strains), period in which started
the diarrhea in calves. It was not possible to detect differences between the
frequencies of microorganisms between the COL+ and COL- groups. The COL+ and
COL- colostrum management did not influence the proportion of aerobic bacteria
present in feces and the occurrence of diarrhea in calves during the neonatal period.
Chapter 2 – The aim of this chapter was to evaluate the influence of colostrum
leukocytes in the innate immune response in Holstein calves. The animals were

submitted to clinical examination, followed by blood samples collection to perform
laboratory tests. The serum cortisol and haptoglobin concentrations did not show
statistical differences while the iron concentration was different between groups at
D7. The leukogram did not show difference to total leukocytes and absolute
neutrophils, but there was a trend between groups at D7 for relative values of
neutrophils. The CH138+CD62L+ marker showed no difference between groups,
despite an increased expression was detected in COL-. The expression of
CH138+CD62L- was greater in COL+ at D14. The phagocytosis and production of
H2O2 by blood neutrophils showed similar profile in relation to S. aureus and E. coli
between groups, although COL- presented greater phagocytosis intensity from D7 to
D14. There was a trend for the increase of granulocyte proportion, releasing basal
H2O2 at D14 for COL-, however COL+ presented trend to a higher intensity of
phagocytosis at D21. The proportion of granulocytes (%) stimulated with S. aureus
releasing H2O2 was greater in COL+ at D7, while stimulation with E. coli resulted in
no differences between the groups. The fluorescence intensity was higher and
gradual in COL+ when cells were stimulated; however, it was not detected statistical
differences. The colostrum cells influenced in the health profile presented by the
calves, with a higher intensity of diarrhea, lower levels of serum iron and anemia in
the group that received frozen colostrum. The migration of blood granulocytes was
faster and more intense in COL+ in relation to COL- after natural exposure to
pathogens that cause diarrhea. The phagocytosis rate (%) of granulocytes showed
similarity between groups, although the fluorescence intensity of COL- was more
intense. In contrast, the COL+ demonstrated greater ability in producing H2O2.
Keywords: Frozen colostrum. Fresh colostrum. Bacterial count. Neutrophils.
Phagocytosis

LISTA DE FIGURAS
Figura 1 - Esquema da transferência de leucócitos do colostro materno até a
absorção pelo epitélio intestinal dos bezerros recém-nascidos –
São Paulo – 2016 ................................................................................. 35
Figura 2 - Resposta imune intestinal para bactérias entéricas e seus antígenos –
São Paulo – 2016 ................................................................................. 41
Figura 3 - Nascimento em baias de parição (a); colostragem em baias de
transição (b), localizadas na maternidade – São Paulo – 2016 ........... 46
Figura 4 - Esfregaço da secreção mamária do colostro fresco fornecido para as
bezerras do grupo com células viáveis (COL+) – São Paulo – 2016 ... 48
Figura 5 - Alojamento das bezerras durante o primeiro mês de vida – São Paulo
– 2016 .................................................................................................. 50
Figura 6 - Escores de fezes adotados para detecção da diarreia em bezerras no
primeiro mês de vida – São Paulo – 2016 ............................................ 52
Figura 7 - Etapas para a diluição seriada e plaqueamento das amostras de
colostro fornecido às bezerras (COL+) e (COL-) – São Paulo –
2016 ..................................................................................................... 53
Figura 8 - Crescimentos bacterianos na placa de TSA – CBT (a); crescimento de
bactérias Gram-negativas na placa com meio de cultura
MacConkey (b) – São Paulo – 2016..................................................... 54
Figura 9 - Demonstração da coleta de fezes com luva estéril direto da ampola
retal – São Paulo – 2016 ...................................................................... 55
Figura 10 - Fezes foram semeadas no fluxo, na presença do fogo em placas
agar sangue, agar MacConkey e SS – São Paulo – 2016 ................... 56
Figura 11 - Esquema do procedimento realizado no laboratório para identificação
das bactérias aeróbias das fezes – São Paulo – 2016......................... 57
Figura 12 - Exemplos dos crescimentos obtidos após 24 a 48 horas de
incubação das placas em estufa a 37°C – São Paulo – 2016 .............. 57
Figura 13 - Frequências (%) do escore fecal em bezerras Holandesas
amamentadas com colostro fresco (COL+) e colostro sem células
viáveis (COL- ) durante o primeiro mês de vida – São Paulo – 2016 ... 61

Figura 14 - Contagem bacteriana total (CBT/mL) e contagem de coliformes totais
(CCT/mL) nas amostras de colostro com (COL+) e sem células
(COL-) – São Paulo – 2016 .................................................................. 63
Figura 15 - Correlações entre IgG, contagem bacteriana total (CBT), contagem
de coliformes totais (CCT) em amostras de colostro com (COL+) e
sem células viáveis (COL-) - São Paulo – 2016 ................................... 65
Figura 16 - Correlação entre sólidos totais, contagem bacteriana total (CBT),
contagem de coliformes totais (CCT) em amostras de colostro com
(COL+) e sem células viáveis (COL-) – São Paulo – 2016 .................. 66
Figura 17 - Comparação entre as diferentes espécies bacterianas isoladas a
partir das fezes das bezerras Holandesas (COL+) e (COL-) durante
o primeiro mês de vida – São Paulo – 2016 ......................................... 71
Figura 18 - Número de isolamentos dos principais gêneros bacterianos isolados
nas amostras de fezes das bezerras Holandesas que ingeriram
colostro com (COL+) e sem células viáveis (COL-) – São Paulo –
2016 ..................................................................................................... 73
Figura 19 - Nascimento em baias de parição (a); colostragem em baias de
transição (b), localizadas na maternidade – São Paulo – 2016 ........... 95
Figura 20 - Esfregaço da secreção mamária do colostro fresco fornecido para as
bezerras do grupo com células viáveis (COL+) – São Paulo – 2016 ... 97
Figura 21 - Alojamento das bezerras durante o primeiro mês de vida – São Paulo
– 2016 .................................................................................................. 98
Figura 22 - Escores de fezes adotado para detecção da diarreia em bezerras no
primeiro mês de vida – São Paulo – 2016 .......................................... 100
Figura 23 - Avaliação da proporção de granulócitos no sangue de bezerras
Holandesas no primeiro mês de vida – São Paulo – 2016 ................. 104
Figura 24 - Ensaio de avaliação da L-selectina – São Paulo – 2016 ...................... 105
Figura 25 - Ensaio de avaliação do CH138+ – São Paulo – 2016 .......................... 105
Figura 26 - Ensaio de avaliação do duplo positivo para CD62L+ e CH138+ – São
Paulo – 2016 ...................................................................................... 106
Figura 27 - Avaliação da proporção de PMN no sangue de bezerras Holandesas
– São Paulo – 2016 ............................................................................ 108
Figura 28 - Avaliação da produção de H2O2 basal, sem estímulos bacterianos,
pelas células PMN com DCFH-DA. Seleção do eixo FL1+ para

avaliação da produção de H2O2 pelos PMN sem estímulo
bacteriano – São Paulo – 2016 .......................................................... 108
Figura 29 - Avaliação da fagocitose das células PMN do sangue de bezerras
Holandesas que fagocitaram Staphylococcus aureus e Escherichia
coli – São Paulo – 2016 ..................................................................... 109
Figura 30 - Avaliação da produção de H2O2 e fagocitose pelas células PMN do
sangue de bezerras Holandesas com DCFH-DA e Staphylococcus
aureus – São Paulo – 2016 ................................................................ 110
Figura 31 - Avaliação da produção de H2O2 e fagocitose pelas células PMN do
sangue de bezerras Holandesas com DCFH-DA e Escherichia coli
– São Paulo – 2016 ............................................................................ 111
Figura 32 - Frequência do escore fecal e incidência de diarreia em bezerras
Holandesas (COL+) e (COL-) durante o primeiro mês de vida – São
Paulo – 2016 ...................................................................................... 114
Figura 33 - Frequência de anemias em bezerras Holandesas (COL+) e (COL-)
durante o primeiro mês de vida - São Paulo - 2016 ........................... 115
Figura 34 - Concentração de cortisol sérico (µg/dL) em bezerras Holandesas que
receberam colostro com (COL+) e sem células viáveis (COL-),
durante o primeiro mês de vida - São Paulo - 2016 ........................... 116
Figura 35 - Haptoglobina sérica em bezerras Holandesas recém-nascidas que
receberam colostro com (COL+) e sem células viáveis (COL-) –
São Paulo – 2016 ............................................................................... 118
Figura 36 - Valores de concentração sérica de ferro (µM/L) em bezerras
Holandesas recém-nascidas que receberam colostro com (COL+) e
sem células (COL-) - São Paulo – 2016 ............................................. 119
Figura 37 - Leucócitos totais (WBC) e neutrófilos (NEUTR) do sangue de
bezerras Holandesas recém-nascidas que receberam colostro com
(COL+) e sem células (COL-) - São Paulo – 2016 ............................ 121
Figura 38 - Proporção de granulócitos CH138+ expressando o receptor CD62L
em bezerras Holandesas que receberam colostro com (COL+) e
sem células viáveis (COL-), no primeiro mês de vida - São Paulo –
2016 ................................................................................................... 123
Figura 39 - Proporção (%) e intensidade da fagocitose (Média Geométrica– Geo
Mean) da fagocitose pelos granulócitos mediante liberação

espontânea ou estimulados, em bezerras Holandesas recém-
nascidas que receberam colostro com (COL+) e sem células
viáveis (COL-) – São Paulo - 2016 ..................................................... 128
Figura 40 - Produção de peróxido de hidrogênio (% e intensidade – Média
Geométrica – Geo Mean) pelos granulócitos, mediante liberação
espontânea ou estimulados, em bezerras Holandesas recém-
nascidas que receberam colostro com (COL+) e sem células
viáveis (COL-) - São Paulo - 2016...................................................... 129

LISTA DE TABELAS
Tabela 1 - Valores médios, mínimos e máximos de imunoglobulinas avaliados
pelo colostrômetro e Brix para (COL+) e (COL-) – São Paulo –
2016 ..................................................................................................... 59
Tabela 2 - Mediana, mínimo, máximo, média e desvio padrão da CBT/mL (x106 e
log10) e CCT/mL (x105 e log10) nas amostras de colostro com
(COL+) e sem células (COL-) – São Paulo – 2016 .............................. 62
Tabela 3 - Distribuição das amostras de colostro com (COL+) e sem células
viáveis (COL-) de acordo com a CBT e CCT/mL – São Paulo –
2016 ..................................................................................................... 64
Tabela 4 - Valores de P para a correlação de Pearson em relação às
concentrações de IgG e sólidos totais correlacionados com CBT e
CCT obtidas a partir das amostras de colostro com (COL+) e sem
células viáveis (COL-) – São Paulo – 2016 .......................................... 64
Tabela 5 - Frequências (%) e número de isolamentos por gêneros bacterianos
nas amostras de fezes das bezerras que ingeriram colostro com
(COL+) e sem células viáveis (COL-) do nascimento aos 28 dias de
vida – São Paulo – 2016 ...................................................................... 68
Tabela 6 - Número de espécies bacterianas e frequência dos isolamentos das
amostras de fezes das bezerras que ingeriram colostro com
(COL+) e sem células viáveis (COL-) do nascimento aos 28 dias de
vida – São Paulo – 2016 ...................................................................... 70
Tabela 7 - Concentração de cortisol sérico (µg/dL) em bezerras Holandesas que
receberam colostro com (COL+) e sem células viáveis (COL-),
durante o primeiro mês de vida - São Paulo - 2016 ........................... 116
Tabela 8 - Valores da concentração (mg/dL) e densidade óptica (DO) para a
haptoglobina sérica em bezerras Holandesas que receberam
colostro com (COL+) e sem células viáveis (COL-), durante o
primeiro mês de vida - São Paulo - 2016 ........................................... 117
Tabela 9 - Valores da concentração de ferro sérico (µM/L) em bezerras
Holandesas que receberam colostro com (COL+) e sem células
(COL-), durante o primeiro mês de vida - São Paulo – 2016 ............. 118

Tabela 10 - Médias e desvios-padrão para os leucócitos totais (WBC) e
neutrófilos (NEUTR) do sangue de bezerras Holandesas que
receberam colostro com (COL+) e sem células viáveis (COL-) - São
Paulo - 2016 ....................................................................................... 120
Tabela 11 - Médias e desvios-padrão para as proporções de granulócitos
CH138+ expressando o receptor CD62-L em bezerras Holandesas
que receberam colostro com (COL+) e sem células viáveis (COL-),
no primeiro mês de vida - São Paulo – 2016 .................................... 122
Tabela 12 - Proporção (%) e intensidade da fagocitose (Média Geométrica – Geo
Mean) e produção de peróxido de hidrogênio pelos granulócitos em
bezerras Holandesas que receberam colostro com (COL+) e sem
células viáveis (COL-) – São Paulo – 2016 ........................................ 127

LISTA DE QUADROS
Quadro 1 - Intervalos de referência para as funções vitais em bezerras no
primeiro mês de vida ............................................................................ 47
Quadro 2 - Escores de fezes adotado para detecção da diarreia em bezerras
no primeiro mês de vida ....................................................................... 51
Quadro 3 - Escores de broncopneumonia adotados para detecção da doença
em bezerras no primeiro mês de vida .................................................. 52
Quadro 4 - Fatores para conversão de números de colônias na placa TSA e
MacConkey para CBT/mL ou CCT/mL de colostro .............................. 54
Quadro 5 - Intervalos de referência para as funções vitais em bezerras no
primeiro mês de vida ............................................................................ 96
Quadro 6 - Escores de fezes adotado para detecção da diarreia em bezerras
no primeiro mês de vida ..................................................................... 100
Quadro 7 - Escores de broncopneumonia adotado para detecção da doença
em bezerras no primeiro mês de vida ................................................ 101
Quadro 8 - Ensaio de fagocitose e produção de H2O2 do sangue de bezerras
Holandesas ........................................................................................ 107

LISTA DE ABREVIATURAS E SIGLAS
FC Frequência Cardíaca
FR Frequência Respiratória
bpm Batimentos por minuto
mpm Movimentos por minuto
T Temperatura
L Litros
Igs Imunoglobulinas
g gramas
ML mililitros
m metro
D dia
FITC Fluorescent Isothiocyanate
CBT Contagem Bacteriana Total
CCT Contagem de Coliformes Totais
TSA Tryptic Soy Agar
MC MacConkey
SOL Solução
SS Solmonella Shigella
PBS Phosphate-Buffered Saline
h Horas
mg miligramas
n número
DP Desvio Padrão
mM milimolar
q.s.p. quantidade suficiente para
VCM Volume Corpuscular Médio
HCM Hemoglobina Corpuscular Média
nm Nanômetro
EDTA Ethylene Diamine Tetraacetic Acid
PE Phycoerythrin

FSC Forward Scatter
SSC Side Scatter
E. coli Escherichia coli
PMN Polimorfonucleares
WBC White Blood Cells
CD Cluster of Differentiation
BHI Brain Heart Infusion
PAMPs Pathogen Associated Molecular Pattern
PRRs Pattern Recognition Receptors
RBC Red Blood Cells
APC Allophycocyanin
SFB Soro Fetal Bovino
NEUTR Neutrófilos
H2O2 Peróxido de Hidrogênio
PI Iodeto de Propídio
µl Microlitro
µg Micrograma
ufc Unidades formadoras de colônia

LISTA DE SÍMBOLOS
°C Graus Celsius
- Negativo
± Mais ou menos
% Porcentagem
® Marca Registrada
> Maior que
≤ Menor ou igual que
< Menor que
= Igual
G Força G
β Beta
* Diferença estatística
† Tendênca
≥ Maior ou igual que
n° Número

SUMÁRIO
1 INTRODUÇÃO GERAL ............................... ............................................... 29
2 REVISÃO DE LITERATURA .......................... ............................................ 30
2.1 TRANSFERÊNCIA DE IMUNIDADE PASSIVA .......................................... 30
2.1.1 Aspectos gerais ........................................................................................ 30
2.1.2 Células do colostro e seus produtos ...................................................... 32
2.1.3 Papel das células na resposta imune dos recé m-nascidos .................. 35
2.1.4 Papel das células do colostro como carreador as de bactérias ........... 38
2.1.5 Desenvolvimento da Resposta Imune de mucosa intestinal ................ 39
CAPÍTULO 1 - INFLUÊNCIA DAS CÉLULAS DO COLOSTRO NA COLONIZAÇÃO
DO INTESTINO POR BACTÉRIAS AERÓBIAS ............... ....................................... 42
3 INTRODUÇÃO ........................................................................................... 43
4 MATERIAL E MÉTODOS ............................. .............................................. 46
4.1 BEZERRAS ................................................................................................ 46
4.2 MOMENTOS DE AVALIAÇÃO ................................................................... 50
4.3 AVALIAÇÃO CLÍNICA PERIÓDICA ........................................................... 51
4.4 ANÁLISES .................................................................................................. 52
4.4.1 CBT e CCT ................................................................................................. 53
4.4.2 Isolamento das bactérias das fezes ........................................................ 54
4.4.2.1 Amostra de fezes ........................................................................................ 55
4.4.2.2 Processamento das amostras de fezes ...................................................... 55
4.5 ANÁLISE ESTATÍSTICA ............................................................................ 58
5 RESULTADOS ..................................... ...................................................... 59
5.1 QUALIDADE DO COLOSTRO ................................................................... 59
5.2 STATUS SANITÁRIO ................................................................................. 59
5.2.1 CBT e CCT no colostro ............................................................................ 62
5.2.2 Relação entre a CBT / CCT com a concentração de Igs no colostro ... 64
5.2.3 Isolamento enteropatógenos aeróbios ................................................... 66
6 DISCUSSÃO .............................................................................................. 75
6.1 QUALIDADE DO COLOSTRO ................................................................... 75
6.2 STATUS SANITÁRIO ................................................................................. 76

6.2.1 CBT e CCT no colostro ............................................................................ 77
6.2.2 Isolamentos bacterianos das fezes ........................................................ 80
7 CONCLUSÃO ...................................... ...................................................... 85
REFERÊNCIAS .......................................................................................... 86
CAPÍTULO 2 - INFLUÊNCIA DOS LEUCÓCITOS DO COLOSTRO NA RESPOSTA
IMUNE INATA DE BEZERRAS HOLANDESAS RECÉM-NASCIDAS . ................... 91
8 INTRODUÇÃO ........................................................................................... 92
9 MATERIAL E MÉTODOS ............................. .............................................. 95
9.1 BEZERRAS ................................................................................................ 95
9.2 MOMENTOS DE AVALIAÇÃO ................................................................... 99
9.3 AVALIAÇÃO CLÍNICA PERIÓDICA ........................................................... 99
9.4 AMOSTRAS ............................................................................................. 101
9.5 ANÁLISES ................................................................................................ 101
9.5.1 Hemograma ............................................................................................. 101
9.5.2 Cortisol .................................................................................................... 102
9.5.3 Haptoglobina ........................................................................................... 102
9.5.4 Ferro ........................................................................................................ 103
9.5.5 Imunofenotipagem dos Neutrófilos ...................................................... 103
9.5.6 Fagocitose e Produção Peróxido de Hidrogênio ................................. 106
9.6 ANÁLISE ESTATÍSTICA .......................................................................... 112
10 RESULTADOS .................................... ..................................................... 113
10.1 STATUS SANITÁRIO ............................................................................... 113
10.2 CORTISOL ............................................................................................... 115
10.3 HAPTOGLOBINA ..................................................................................... 117
10.4 FERRO ..................................................................................................... 118
10.5 LEUCÓCITOS TOTAIS E NEUTRÓFILOS SANGUÍNEOS ...................... 119
10.6 EXPRESSÃO DE MARCADORES NOS GRANULÓCITOS ..................... 121
10.7 FUNÇÃO DOS NEUTRÓFILOS ............................................................... 123
11 DISCUSSÃO ............................................................................................ 130
11.1 QUALIDADE DO COLOSTRO ................................................................. 130
11.2 STATUS SANITÁRIO DAS BEZERRAS................................................... 131
11.3 CORTISOL ............................................................................................... 134
11.4 HAPTOGLOBINA ..................................................................................... 135

11.5 FERRO ..................................................................................................... 136
11.6 PROPORÇÃO E FUNÇÃO DOS NEUTRÓFILOS .................................... 137
12 CONCLUSÕES ........................................................................................ 142
REFERÊNCIAS ........................................................................................ 143
13 CONCLUSÃO GERAL ............................... .............................................. 148
REFERÊNCIAS ........................................................................................ 149
ANEXOS .................................................................................................. 153

29
1 INTRODUÇÃO GERAL
Os bezerros recém-nascidos são agamaglobulinêmicos, imunossuprimidos e
possuem sistema imune específico imaturo ao nascimento, sendo dependentes do
colostro materno no primeiro mês de vida (CORTESE, 2009).
O colostro bovino é rico em componentes imunológicos como leucócitos,
citocinas e imunoglobulinas (NOCEK; BRAUND; WARNER, 1984; WILLIAMS, 1993;
CROSS; GILL, 2000; KORHONEN; MARNILA; GILL, 2000). O papel dos anticorpos
do colostro nos mecanismos imunes para a defesa das bezerras recém-nascidas
está bem consolidado, entretanto pouco se sabe sobre os mecanismos que
envolvem as células e seus produtos.
As células do colostro são absorvidas pelos enterócitos e transferidas para o
sangue e órgãos linfoides das bezerras após colostragem (TUBOLY et al., 1988
LIEBLER-TENORIO; RIEDEL-CASPARI; POHLENZ, 2002; REBER et al., 2006). A
participação dessas células na resposta imune é amplamente aceita, porém pouco
se sabe sobre a intersecção e os mecanismos que envolvem as células maternas e
a imunidade dos recém-nascidos.
As poucas pesquisas realizadas nesta linha focaram seus estudos na
influência dos leucócitos do colostro, através da marcação de linfócitos sanguíneos,
na resposta imune de bezerras que ingeriram colostro celular e acelular. A resposta
imune inata das bezerras que receberam colostro fresco e congelado foi avaliada
apenas por Riedel-Caspari e Schimidt (1991a,b) e Riedel-Caspari (1993).
Diante do contexto apresentado, esta pesquisa avaliou a influência das
células do colostro na resposta imune inata e ocorrência de doenças em bezerras
Holandesas do nascimento aos 28 dias de vida, motivada pela busca de novas
estratégias de manejo de colostro para a otimização da imunidade e sanidade em
bezerras, durante a fase de desenvolvimento.
O objetivo geral desta pesquisa foi avaliar se os leucócitos do colostro
influenciam na colonização da microbiota intestinal, resposta imune inata e na
ocorrência de diarreias no período neonatal.

30
2 REVISÃO DE LITERATURA
Este tópico apresentará os principais aspectos relacionados à transferência
de imunidade passiva com ênfase no papel das células do colostro em relação à
colonização intestinal e desenvolvimento da resposta imune de bezerros recém-
nascidos.
2.1 TRANSFERÊNCIA DE IMUNIDADE PASSIVA
Os bezerros recém-nascidos são agamaglobulinêmicos, imunossuprimidos e
possuem sistema imune específico imaturo ao nascimento, sendo dependentes do
colostro materno no primeiro mês de vida (CORTESE, 2009).
2.1.1 Aspectos gerais
O colostro dos bovinos é composto por fatores imunes como imunoglobulinas
(Igs), células e seus subprodutos (GONZÁLEZ et al., 2013). Além disso, a secreção
mamária também possui elevadas concentrações de gordura, vitaminas solúveis em
gordura (retinol, tocoferol e β-caroteno), vitaminas solúveis em água (niacina,
tiamina, riboflavina, vitamina B12, piridoxal, piridoxamina e piridoxina), minerais (Ca,
P, Mg, Na, K, Zn, Fe, Cu, S e Mn), fatores antimicrobianos (lactoferrina, lisozima e
lactoperoxidase) e fator de crescimento semelhante à insulina que estimula o
crescimento dos recém-nascidos (REITER, 1978; TODHUNTER; SMITH;
SCHOENBERGER, 1985; KEHOE; JAYARAO; HEINRICHS, 2007).
A qualidade imunológica do colostro tem sido avaliada pelas concentrações
de imunoglobulinas (Igs). O colostro bovino é composto por aproximadamente 85-
90% de IgG, 5% de IgA e 7% de IgM que atravessam a barreira do epitélio da
glândula mamária migrando do fluido extracelular para a glândula mamária através
do mecanismo de endocitose (LARSON; HEARY; DEVERY, 1980).

31
Esta avaliação pode ser feita através do imunoensaio para a determinação
direta dos teores de IgG (IgG > 50g/L), entretanto esse teste é caro e sua leitura
deve ser realizada entre 18 a 24 horas de incubação. Este fato torna a escolha de
testes rápidos a campo mais adequada, tais como colostrômetro e refratômetro
(CHIGERWE et al., 2008; BIELMANN et al., 2010; QUIGLEY et al., 2013).
Recentemente foram estabelecidos novos pontos de corte para o teor de IgG (IgG ≥
80g/L) e sólidos totais (≥ 23%), respectivamente, determinados pelo colostrômetro e
refratômetro Brix (BARTIER; WINDEYER; DOEPEL, 2015).
A absorção das Igs pelas células epiteliais intestinais ocorre por pinocitose.
Durante as primeiras horas de vida, o abomaso das bezerras apresenta pequena
produção de ácido clorídrico, atividade mínima da pepsina gástrica, presença de
fator inibidor de tripsina e baixa atividade proteolítica intestinal. Estes fatores
protegem as Igs do colostro do processo de digestão (KRUSE, 1983). No intestino,
as Igs se ligam aos receptores Fc dos enterócitos (FcRn) responsáveis pelo
transporte ativo dessa substância do lúmen intestinal para a circulação sanguínea
dos neonatos. A permeabilidade do epitélio intestinal às macromoléculas ocorre
apenas entre as primeiras 18 a 24 horas de vida, possivelmente devido à
substituição das células intestinais que expressavam os receptores FcRn (GODDEN,
2008).
A quantidade de colostro fornecida para os bezerros é outro fator importante.
Recentemente vem sendo indicado o fornecimento de volume equivalente a 15% do
peso vivo ao nascimento, dentro das primeiras 12 horas de vida (PAULA, 2016),
entretanto McGuirk e Collins (2004) recomendaram o volume equivalente a 8-10%
do peso vivo ao nascimento, considerando que neste volume há a quantidade
suficiente de imunoglobulinas necessária para a transferência de imunidade passiva
e garantia da saúde e desenvolvimento das bezerras.
Após a ingestão do colostro, os níveis séricos de imunoglobulinas atingem
concentrações significativas em poucas horas. Bezerras apresentam falha na
transferência de imunidade passiva quando ocorrem concentrações de IgG menores
que 10mg/dL, quando avaliadas entre 24 e 48 horas de vida (NATIONAL ANIMAL
HEALTH MONITORING SYSTEM, 1996; WEAVER et al., 2000).

32
2.1.2 Células do colostro e seus produtos
Os leucócitos foram observados no colostro humano pela primeira vez por
Alexander Donné (1844) 1 apud Mandyla et al. (1982, p.995). Entretanto, a
transferência de imunidade celular da mãe ao recém-nascido foi sugerida no artigo
de revisão de Watson (1980).
Estima-se que o colostro bovino possui entre 1x106 e 2,5x106 células
somáticas/mL, dentre as quais cerca de 32% são viáveis (MCDONALD;
ANDERSON, 1981; LIEBLER-TENORIO; RIEDEL-CASPARI; POHLENZ, 2002;
GOMES et al., 2011; SILVA, 2014).
Os leucócitos colostrais apresentam perfil diferenciado em relação ao sangue
periférico. A população de leucócitos no colostro de primeira ordenha, colhido até
seis horas pós-parto, consiste em 13,3% de neutrófilos, 16,4% de linfócitos, 69,5%
de monócitos e células epiteliais e 0,27% de eosinófilos (GOMES et al., 2011).
Megank et al. (2014) avaliaram os leucócitos colostrais através da separação das
células por centrifugação e densidade de gradiente e obtiveram a proporção de
25,4% de linfócitos T, 2,9% de linfócitos B, e 32,7% de macrófagos.
A avaliação da atividade funcional das células do colostro é limitada pela
dificuldade do seu isolamento, devido à quantidade abundante de debris celulares,
gordura, proteínas e partículas auto-fluorescentes, que dificultam a separação das
células do colostro para sua avaliação funcional (GOMES et al., 2014).
Tuboly et al. (1988) foram os primeiros autores a comprovarem a
transferência de células do colostro para 23 leitões filhos de quatro porcas (A a D).
Os leucócitos do colostro foram isolados por gradiente de densidade com uso de
Ficoll-Paque, sendo posteriormente coradas com Tecnécio (Na99mTcO4) em solução
de NaCl. Após sete a dez horas de nascimento, 5mL da solução contendo as células
marcadas foram injetados por laparotomia diretamente no estômago dos leitões da
porca A e no jejuno dos leitões provenientes da porca B. Os leitões oriundos das
porcas C e D receberam a suspensão celular através de sonda naso-esofágica. Os
animais dos referidos grupos receberam células oriundas do colostro da própria mãe
ou doadoras. Três leitões das porcas B e C receberam células inativadas a 56ºC.
1 DONNÉ, A. Cours de microscopie: Complementaire des études medicales, v.1, n.1, Bailliere, Paris, 1844.

33
Após oito horas da aplicação do colostro os animais foram eutanasiados e os
fragmentos de duodeno, jejuno e linfonodos foram examinados por autorradiografia.
Tuboly et al. (1988) observaram que os leitões que mamaram colostro oriundo
de suas próprias mães apresentaram células colostrais na região cortical dos
linfonodos mesentéricos, enquanto os leitões que receberam colostro de doadoras
não apresentaram células nos ductos linfáticos, nem nos linfonodos, mas sim no
tecido epitelial do intestino. Em leitões que mamaram colostro de suas próprias
mães foram encontrados linfócitos no lúmen e frequentemente nas micro-
vilosidades, sozinhos ou agrupados. Em animais que mamaram colostro de
doadoras, muitas células colostrais foram encontradas no lúmen intestinal, porém,
no epitélio intestinal, foram encontradas apenas ocasionalmente. Assim, os autores
concluem que há migração intracelular dos leucócitos do colostro. Não houve
absorção de linfócitos que receberam colostro com células inativadas pela alta
temperatura.
Tuboly et al. (1995) também confirmaram a transferência de células maternas
para ovinos recém-nascidos.
Williams (1993) estudou a absorção intestinal de leucócitos colostrais
marcados com FITC em 49 leitões. Este autor observou a presença dos leucócitos
marcados na circulação sanguínea dos leitões em duas horas após a alimentação,
observando-se pico em relação à presença dessas células entre 5-7 horas após a
ingestão. Após 24 horas de ingestão, as células foram encontradas também no
fígado, pulmão, linfonodos, baço e tecidos gastrintestinais. Os autores encontraram
células marcadas no duodeno e jejuno, porém não foram encontradas células na
região do íleo, fora da placa de Peyer.
Liebler-Tenorio, Riedel-Caspari e Pohlenz (2002) comprovaram a absorção
dos leucócitos do colostro proveniente de vacas através da marcação dessas
células. O colostro foi ordenhado em até 15 minutos após o parto e as células
separadas por mecanismo de centrifugação. Ao pellet celular foi adicionado
fluorescent isothiocyanate (FITC). Os animais foram anestesiados e através de
laparotomia foi obtido acesso a duas regiões do intestino: no jejuno médio onde não
existe placa de Peyer e na região distal do íleo onde as placas de Peyer estão
presentes. Nestas regiões foram injetados 10 a 15 mL da suspensão celular
contendo as células maternas marcadas. Após 1h30 e 2 horas de exposição, uma
porção da alça intestinal de cada região previamente estabelecida foi removida e

34
fixada com solução de formalina. Em seguida, animais foram eutanasiados com
pentobarbital. Os fragmentos teciduais foram submetidos à imuno-histoquímica para
a detecção das células maternas.
Liebler-Tenorio, Riedel-Caspari e Pohlenz (2002) detectaram células
maternas marcadas no lúmen intestinal, na superfície, entre vilosidades e nas placas
de Peyer em todos os animais (n=3). Apenas macrófagos e linfócitos puderam ser
discernidos pela morfologia. Foi possível notar diferença na quantidade de leucócitos
observados no corte intestinal, provavelmente por diferenças na proporção das
células efetivamente marcadas ou perda de células durante o procedimento de
coloração, considerando que foi injetada a mesma quantidade da suspensão celular
(10-15mL). Foram encontradas mais células coradas nos seios ao redor dos
folículos linfóides das placas de Peyer em comparação ao epitélio e espaço
subepitelial, indicando rápido e eficiente transporte através do epitélio. Os autores
acreditam que através dos seios linfáticos ocorre a drenagem das células para os
linfonodos mesentéricos; além disso, os linfócitos absorvidos do colostro podem
recircular e serem encontrados em outros órgãos e tecidos que não o intestino.
Reber et al. (2006) compararam o efeito do colostro acelular e sem gordura
(congelados a -20ºC), acrescidos de células mononucleares do sangue periférico
materno, na resposta imune de bezerros recém-nascidos. As células foram isoladas
do sangue e marcadas com substância fluorescente (PKH26-GL, lido em FL2),
sendo administradas aos bezerros com seis horas de nascimento. Foram coletadas
amostras sanguíneas dos bezerros com 4, 8, 12, 24, 36 e 48 horas pós ingestão do
colostro. As células maternas no colostro foram encontradas na corrente sanguínea
de bezerros, avaliando assim, a capacidade de absorção e sua resposta imune. A
circulação destas células nos recém-nascidos foi monitorada por citometria de fluxo.
Os autores observaram que células maternas marcadas ganharam a circulação dos
bezerros após absorção pelo epitélio intestinal. Os autores consideraram que o
fenótipo das células maternas da glândula mamária pode facilitar a transferência dos
leucócitos maternos para a circulação do neonato. O pico de leucócitos na
circulação sanguínea dos bezerros ocorreu entre 12 a 24 horas após a ingestão do
colostro e grande parte das células que foram marcadas, foram encontradas no
sangue e, por fim, as células marcadas desapareceram da circulação apos 36 horas
de ingestão do colostro (Figura 1).

35
Reber et al. (2006) compararam bezerros que mamaram colostro fresco,
colostro com células mononucleares periféricas oriundas do sangue materno
marcadas com substância fluorescente, com bezerros que mamaram colostro
acelular. Os autores encontraram em seu experimento maior quantidade de
expressão de receptores indicativos de inflamação no grupo que mamou colostro
sem células (CD11a, CD11c, CD43 e CD62L).
Figura 1 - Esquema da transferência de leucócitos do colostro materno até a absorção pelo
epitélio intestinal dos bezerros recém-nascidos – São Paulo – 2016
Fonte: adaptado de: Reber (2000) por Costa, J. F. R. (2016)
2.1.3 Papel das células na resposta imune dos recém -nascidos
Experimentos recentes têm demonstrado que a presença das células no
colostro é um diferencial na resposta imune das bezerras recém-nascidas.
Entretanto, o papel destas células nos diferentes mecanismos da resposta imune
que conduzem ao amadurecimento imunológico das bezerras ainda são pobremente
compreendidos.

36
Riedel-Caspari (1993) administrou 109 unidades formadoras de colônia da
Escherichia coli enteropatogênica de forma oral para 20 bezerros em até três horas
pós-nascimento. Após a infecção e durante os próximos dois dias, 10 bezerros
receberam pool de colostro suplementado com células colostrais (COL+) e 10
bezerros receberam pool de colostro acelular (COL-). As bezerras pertencentes ao
grupo COL+ excretaram significativamente menor quantidade de bactérias E. coli
enteropatogênica nas fezes durante a primeira semana após a infecção e atingiram
o limite mínimo de detecção antes que o grupo COL-. A concentração de IgA e IgM
específicos contra E. coli no soro das bezerras COL+ foi significativamente superior
no período pós-natal do que no grupo COL-, e permaneceu um pouco acima durante
todo o período estudado.
Reber et al. (2008a) buscaram entender o impacto que as células do colostro
materno, quando absorvidas pelo epitélio intestinal, causam no desenvolvimento do
sistema imune dos recém-nascidos, avaliando a resposta adaptativa por meio da
fenotipagem de linfócitos. Para isso, 10 bezerros foram alimentados com colostro
com (COL+) e sem (COL-) células. Amostras de sangue foram colhidas
semanalmente durante o primeiro mês de vida. Todos os bezerros apresentaram
uma baixa quantidade de anticorpos circulantes ao nascimento, e após a ingestão
do colostro, receberam quantidade semelhante de IgG, o que aumentou a
concentração sérica de IgG no sangue para aproximadamente 4.000 mg/dL.
Reber et al. (2008a) também observaram que os bezerros que receberam
colostro acelular (COL-) apresentaram maior número e intensidade de fluorescência
para os linfócitos expressando CD11a, quando comparados ao grupo que recebeu
colostro com células (COL+). O CD11a é relacionado pelos autores como um
marcador que indica inflamação sistêmica, sugerindo aumento na susceptibilidade
dos bezerros que não receberam leucócitos colostrais aos desafios ambientais e
agentes infecciosos. Bezerros COL- apresentaram demora no desenvolvimento do
sistema imune em 1 a 2 semanas quando comparados ao COL+, demonstrada pela
baixa expressão de receptores CD25+ e CD26+ pelos linfócitos e produção de
citocinas abaixo do valor considerado ótimo para as células responsivas. A
expressão de MHC de classe I rapidamente aumentou durante a primeira semana
de vida em ambos os grupos. Os autores concluem que cada vez mais tem sido
comprovado que as células do colostro representam uma importante arma para o
desenvolvimento do sistema imune do recém-nascido e que o grupo que mamou

37
colostro com células apresentou uma menor expressão dos marcadores que estão
relacionados com ativação linfocitária e estresse fisiológico.
Reber et al. (2008b) também examinaram o efeito dos leucócitos do colostro
materno no desenvolvimento e maturação das células apresentadoras de antígeno
dos neonatos. Ao nascimento, bezerros receberam colostro com (COL+) e sem
células (COL-). Semanalmente, amostras de sangue foram coletadas e marcadores
de células associadas com estresse fisiológico da linhagem dos monócitos foram
adicionados aos leucócitos sanguíneos. A maior diferença encontrada entre os
marcadores ocorreu nos primeiros sete dias de vida. Os bezerros que receberam
colostro sem células apresentaram maior expressão dos marcadores indicativos de
processos inflamatórios (CD11a, CD11c e CD14), quando comparados com o grupo
que mamou colostro com células. Os bezerros do COL- apresentaram um aumento
no número de monócitos no sangue periférico durante as primeiras duas semanas,
entretanto, essas células apresentaram menores níveis de expressão do CD25 e
MHC de classe I comparado com o grupo COL+, que apresentou maior ativação
celular e apresentação de antígenos. Os autores demonstram assim que a
transferência de leucócitos colostrais afeta a maturação da linhagem celular de
monócitos. Isso inclui um aumento na capacidade de apresentação de antígeno,
como indicado pelo aumento do MHC de classe I nos monócitos.
Langel et al. (2015) examinaram o efeito das células do colostro materno na
saúde e perfil imune das células de bezerras leiteiras, tendo como hipótese que
bezerras alimentadas com colostro fresco, contendo células viáveis (COL+), teriam
níveis de parâmetros sanguíneos aumentados no primeiro mês de vida, quando
comparados aos bezerros que fossem alimentados com colostro sem células viáveis
(COL-). Para isso, 37 bezerras das raças Holandesas e Jersey receberam colostro
fresco com (COL+) e sem células viáveis (COL-). O colostro fornecido foi de boa
qualidade (índice Brix > 32%). Os momentos avaliados foram: antes da mamada do
colostro (0h), 6h, D1, D3, D7, D14, D21 e D28. As células sanguíneas foram
avaliadas através da imunofenotipagem, além do acompanhamento da sanidade por
meio dos escores de fezes e broncopneumonia.
Langel et al. (2015) não observaram diferenças para os tratamentos nas
concentrações de IgG, IgM e IgA antes e depois da ingestão do colostro. Não foram
encontradas diferenças para o perfil sanitário das bezerras durante o estudo. O
momento de maior ocorrência de diarreias ocorreu no D14 para ambos os grupos.

38
Escores respiratórios aumentaram para os dois grupos no D12. O grupo COL-
apresentou valores inferiores de T auxiliar CD4+ e TCD4+CD62L+CD45RO- no D1, e
menos também de TCD4+CD62L+CD45RO+ no D1 e D3 do COL-, quando
comparado ao COL+. Em contrapartida, as bezerras COL- apresentaram maior
expressão de TCD4+CD62L-CD45RO+ com 6h de vida, 1, 3 e 7 dias e maior
proporção de monócitos no D7. A diferença entre a proporção de células
CD4+CD62L+CD45RO- no D1 para o grupo COL- pode ser explicado pela migração
das células que expressam CD62L através do intestino para o sangue. Não foram
detectadas diferenças entre os tratamentos para as células T gamma-delta. Desta
forma, os autores concluem que a transferência de células imunes intactas através
do colostro fresco pode desempenhar um papel importante no status imune do
neonato. Aumentando a capacidade imune da bezerra recém-nascida, aumentará a
resistência a doenças, resultando em animais saudáveis para o rebanho.
2.1.4 Papel das células do colostro como carreadora s de bactérias
O papel das células do colostro como carreadoras de bactérias maternas para
o intestino dos recém-nascidos tem sido suscitado. O isolamento de microrganismos
do colostro humano empregando-se técnicas tradicionais de cultivo bacteriano tem
sido mal sucedida. Entretanto, material genético de bactérias intestinais tem sido
detectado no interior das células presentes na secreção mamária em humanos
(PÉREZ et al., 2007).
Em bovinos, bactérias do grupo Staphylococcus foram detectadas a partir das
células do colostro cultivadas em meio enriquecido para Staphylococcus (24/72),
ágar sangue (26/72) e manitol (32/72). As principais células identificadas no colostro
foram células apresentadoras de antígenos - macrófagos (ADKINS et al., 2015).
A similaridade entre a microbiota das fezes e leite materno com as fezes dos
recém-nascidos tem suscitado a hipótese de que existe uma via entero-mamária, na
qual as bactérias do intestino materno são capturadas pelas células dendríticas e
transferidas para a secreção mamária. A maior biodiversidade bacteriana presente
nas células dendríticas de mulheres gestantes e lactentes, quando comparada com

39
as não gestantes e não lactentes, reforçam a retro-referida hipótese (PÉREZ et al.,
2007).
Os microrganismos apresentam padrões repetidos em sua superfície
conhecidos como PAMPs (Pathogen-Associated Molecular Pattern) reconhecidos
pelos PRRs (Pattern Recognition Receptors) presentes nas células apresentadoras
de antígenos. Acredita-se que as células maternas migram para os linfonodos
mesentéricos e Placa de Peyer, induzindo a ativação e diferenciação dos linfócitos
naïve.
2.1.5 Desenvolvimento da Resposta Imune de mucosa i ntestinal
A resposta imune inata das mucosas possui mecanismos para impedir o
contato dos microrganismos residentes com o epitélio intestinal e circulação
sistêmica, pois não são inócuos e podem induzir resposta inflamatória intensa e
sepse (BRANDTZAEG, 2010; VERHASSELT, 2010; RESCIGNO, 2011; KOBOZIEV
et al., 2014).
Mecanismos da imunidade inata impedem o contato e destroem
microrganismos da microbiota residente para controle populacional. A barreira
intestinal é composta por muco e fatores antimicrobianos, secretados por diferentes
tipos celulares que compõem a mucosa intestinal, como as células Globet que
secretam grande quantidade de muco e proteína glicosilada altamente resistente à
digestão enzimática. A perda da camada de muco, penetração e aderência de
bactérias ao epitélio intestinal pode desencadear processo inflamatório. Outras
células epiteliais intestinais secretam potentes peptídeos microbianos como a
lecitina, capazes de destruir diretamente bactérias Gram-positivas. Células Paneth
localizadas nas criptas intestinais secretam defensinas e catelicidinas que
permeabilizam a membrana das bactérias. Também são produzidas lipocalina-2 e
catepsina-K (BRANDTZAEG, 2010; VERHASSELT, 2010; RESCIGNO, 2011;
KOBOZIEV et al., 2014).
A secreção de IgA é outro mecanismo usado para impedir o contato da
bactéria com o epitélio intestinal. Evidências sugerem que as células dendríticas da
Placa de Peyer do intestino delgado podem penetrar o epitélio intestinal, capturando

40
continuamente a microbiota residente do lúmen intestinal via endocitose. Estes
microrganismos são carregados pelas células dendríticas aos linfonodos
mesentéricos ou permanecem na Placa de Peyer, induzindo a ativação e
diferenciação das células B naïve para produção de IgA específica para bactérias
comensais. Estas células migram através da circulação sistêmica para a lâmina
própria do epitélio intestinal e secretam IgA no interstício intestinal. As IgA são
transportadas pelas células epiteliais até seu ápice e secretadas no lúmen intestinal
para ligar-se à microbiota residente, limitando sua habilidade em penetrar a mucosa
intestinal (BRANDTZAEG, 2010; KOBOZIEV et al., 2014).
Em função do grande número de bactérias no lúmen intestinal, é inevitável
que algumas consigam violar a barreira epitelial e alcançar a lâmina própria. Neste
caso, os invasores encontram a segunda linha de defesa do sistema imune
composta por fagócitos. Geralmente estes microrganimos são fagocitados e
destruídos por macrófagos intestinais através de ação enzimática ou produção de
radicais livres. É importante ressaltar que macrófagos intestinais produzem menores
concentrações de citocinas pró-inflamatórias que aqueles da circulação sistêmica
para limitação do processo inflamatório. Além disso, o macrófago intestinal é
extremamente hábil para reparação tecidual (BRANDTZAEG, 2010; VERHASSELT,
2010; RESCIGNO, 2011; KOBOZIEV et al., 2014).
Os microrganismos que colonizam a microbiota intestinal podem ajudar a
suprimir respostas imunes inadvertidas que poderiam ser desencadeadas por outros
patógenos (SARTOR, 2008).
Microrganimos invasores dispararam resposta imune envolvendo células T,
associadas a macrófagos e células mielóides. Antígenos associados às células
dendríticas nos linfonodos mesentéricos podem ativar, diferenciar e proliferar células
T para produzir resposta Th1 (celular) ou Th17. Estas células também migram dos
linfonodos para a lâmina própria intestinal onde estão as bactérias residentes, via
circulação sistêmica. Células Th1 e Th17 liberam citocinas pró-inflamatórias, como
IFN-gama, IL-17, IL-22, IL-21 e TNF-alfa, que ativam células apresentadoras de
antígeno presentes no tecido (macrófagos), resultando na geração adicional das
citocinas pró-inflamatórias IL-1 beta, IL-6, IL-8 e IL-12 e espécies reativas do
oxigênio (superóxido, peróxido de hidrogênio) e óxido nítrico. Estes mediadores
inflamatórios amplificam a atividade microbicida de macrófagos e aumentam a
expressão de moléculas de adesão (ICAM-1, VICAM-1 e E-selectina) na superfície

41
das células endoteliais, facilitando o recrutamento adicional de neutrófilos,
macrófagos e linfócitos para o tecido epitelial, colaborando com a defesa do
hospedeiro. A supressão da resposta inflamatória é realizada pelas células T CD4+
reguladoras (Treg). As bactérias também podem ser fagocitadas por endocitose
diretamente pelas células dendríticas e seguirem diretamente para os linfonodos
mesentéricos realizando o mesmo processo de apresentação aos linfócitos T e
expansão clonal (AUJLA; DUBIN; KOLLS, 2007; BRANDTZAEG, 2010;
VERHASSELT, 2010; IVANOV; LITTMAN, 2011; RESCIGNO, 2011; KOBOZIEV et
al., 2014) (Figura 2).
Figura 2 - Resposta imune intestinal para bactérias entéricas e seus antígenos – São Paulo – 2016
.
Fonte: adaptado de: Mowat (2003); Koboziev et al. (2014) por Costa, J. F. R. (2016). Legenda: Gut: intestino; Peyer`s Patch: placa de Peyer

42
CAPÍTULO 1 - INFLUÊNCIA DAS CÉLULAS DO COLOSTRO NA COLONIZAÇÃO
DO INTESTINO POR BACTÉRIAS AERÓBIAS

43
3 INTRODUÇÃO
O colostro apresenta uma diversidade de bactérias que possuem duas
origens distintas: microbiota materna e aquelas oriundas do processo de
contaminação entre a colheita e o fornecimento de colostro para as bezerras. O
papel das bactérias oriundas destes distintos grupos ainda é pobremente discutido
no que se refere à espécie bovina.
McGuire e McGuire (2015) ressaltam que a secreção mamária pode ser
considerada um probiótico devido à presença de microrganismos oriundos da
microbiota materna.
Staphylococcus, Streptococcus e Enterobacteriaceae representam os
principais grupos de bactérias aeróbias isoladas no leite materno humano (MARTIN
et al., 2012). O perfil de bactérias aeróbias também foi determinado no colostro e
leite de vacas em transição, observando-se a presença de Staphylococcus
coagulase negativa, Staphylococcus aureus, Streptococcus, Corynebacterium e
Serratia (SILVA, 2014).
Bifidobacterium e Lactobacilllus são os principais anaeróbios isolados no leite
materno humano (MARTIN et al., 2009), porém estes dados ainda não foram
relatados para os bovinos.
A importância das células do colostro como carreadoras de bactérias
maternas participantes do processo de colonização do intestino dos recém-nascidos
tem sido suscitada. Em bovinos, bactérias do grupo Staphylococcus foram
detectadas a partir das células do colostro cultivadas em meio enriquecido para
Staphylococcus (24/72), agar sangue (26/72) e manitol (33/72). As principais células
identificadas no colostro foram células apresentadoras de antígenos - macrófagos
(ADKINS et al., 2015).
Apesar da conotação positiva das bactérias do colostro provenientes da
microbiota materna, as pesquisas em bovinos têm sido focadas na presença do
grupo de bactérias oriundas da contaminação desse alimento da ordenha à
mamadeira das bezerras recém-nascidas. Nesta linha, os pesquisadores da área
buscam alternativas para reduzir a contagem bacteriana total (CBT) e a contagem
de coliformes totais (CCT) nas amostras de colostro fornecidas às bezerras.

44
Godden et al. (2012) comprovaram que a pasteurização do colostro a 60oC
por 30 minutos reduziu a CBT e CCT/mL (Log10 = 3,5 e 2,1; respectivamente) em
relação ao colostro fresco (Log10 = 5,6 e 4,7; respectivamente). A CBT e CCT/mL
apresentou correlação negativa com a concentração sérica de IgG, fato que explica
a maior incidência de diarreias e doenças totais nas bezerras que receberam o
colostro fresco.
Elizondo-Salazar e Heinrichs (2009) também determinaram o efeito do uso de
três diferentes tratamentos de colostro: sem aquecimento e congelado a -20oC
imediatamente após ordenha – baixa concentração de bactérias; pasteurização:
60oC por 30 minutos e em seguida congelados a -20oC; e sem aquecimento – alta
concentração de bactérias devido à exposição ambiental do colostro por 24 horas a
20oC. Os valores encontrados para CBT/mL e CCT/mL (log10) foi igual a 3,97 e 2,02;
5,61 e 3,16; 2,81 e 0,00 para os grupos sem aquecimento – baixa bactéria, sem
aquecimento – alta bactéria e pasteurização, respectivamente. As amostras de
colostro foram descongeladas e examinadas para Staphylococcus coagulase-
negativa, Streptococcus ambiental, não-coliforme Gram-negativos, Streptococcus
agalactiae e Staphylococcus aureus, de acordo com Jayarao et al. (2004). Não
foram observadas diferenças entre as concentrações de IgG em relação aos grupos
experimentais.
Muitas propriedades vêm utilizando colostro pasteurizado com o intuito de
diminuir a contaminação por microrganismos. Neste processo, o colostro é
submetido a alta temperatura (60ºC) e os leucócitos presentes nessa secreção são
destruídos (GODDEN et al., 2012). Apesar deste fato, Johnson et al. (2007) não
encontraram alterações no número total de leucócitos periféricos, neutrófilos e
linfócitos em animais que receberam colostro fresco ou pasteurizado.
A colonização bacteriana do intestino é um processo complexo, que se inicia
em pequena escala durante o período fetal, que é comprovada pela presença de
bactérias no líquido amniótico, mecônio e cordão umbilical de recém-nascidos
humanos. O contato com microrganismos pertencentes à microbiota vaginal,
intestinal e mamária da mãe, além daqueles pertencentes ao meio ambiente ao qual
o neonato está inserido, intensifica o processo de colonização intestinal após seu
nascimento. A composição da microbiota materna, local, forma de nascimento e
padrão de dieta desempenham importante papel na colonização intestinal (MARTIN
et al., 2012).

45
Staphylococcus, Bifidobacterium, Lactobacilllus, Streptococcus e
Enterobacteriaceae são os principais grupos bacterianos isolados no leite materno e
fezes de recém-nascidos humanos (MARTIN et al., 2012; FERNÁNDEZ et al., 2013).
Testes de biologia molecular confirmaram que mãe e recém-nascido compartilham
as mesmas cepas bacterianas, indicando que o leite materno pode contribuir para a
transferência de bactérias e colaborar com a colonização do intestino infantil
(MARTIN et al., 2012).
O perfil da microbiota das fezes das bezerras em aleitamento parece ser
semelhante aos humanos, observando-se predomínio de coliformes, Lactobacillus e
Bifidobacterium nos primeiros 21 dias de vida (RADA et al., 2006; VLKOVÁ;
TROJANOVÁ; RADA, 2006).
As pesquisas relatadas buscaram bactérias específicas e não foram
encontrados trabalhos que avaliassem o perfil dos microrganismos nas fezes das
bezerras por métodos tradicionais de cultivo. A desvantagem desta metodologia está
associada à ineficiência na detecção de anaeróbios como os Lactobacillus e
Bifidobacterium. Por outro lado, a identificação destes microrganismos requer o
emprego de técnicas de biologia molecular que exige treinamento e maior
investimento financeiro.
Apesar da grande relevância dos bovinos para a economia mundial, muitos
aspectos da sua biologia são ainda desconhecidos, incluindo a composição da sua
microflora normal e alterações relacionadas às doenças. A diversidade da população
bacteriana em bovinos ainda é muito pouco conhecida. Saber a complexidade e
ecologia de microrganismos presentes no intestino dos bovinos, principalmente no
período neonatal e suas alterações decorrentes das mudanças na forma de
aleitamento e alimentação é de extrema importância para o desenvolvimento de
estratégias eficazes para reduzir ou bloquear os distúrbios gastrintestinais.
A hipótese desta pesquisa é de que as células do colostro bovino influenciam
no perfil de enterobactérias aeróbias do trato gastrintestinal de bezerras Holandesas
recém-nascidas. Assim, o objetivo desta pesquisa foi avaliar se o fornecimento de
colostro com (fresco) e sem células viáveis (congelado) influencia na proporção das
espécies de bactérias aeróbias do trato gastrintestinal e ocorrência de diarreias no
período neonatal.

46
4 MATERIAL E MÉTODOS
Esta pesquisa foi aprovada pelo Comitê de Ética no Uso de Animais da
Faculdade de Medicina Veterinária e Zootecnia da USP, protocolo nº 2934/2013.
4.1 BEZERRAS
As bezerras desta pesquisa eram oriundas de fazenda comercial de alta
produção, localizada no município de Araras – São Paulo. Este experimento foi
conduzido de julho a outubro de 2014.
Os partos foram assistidos pelos funcionários e veterinários da propriedade
para auxílio obstétrico, separação das recém-nascidas e controle do manejo de
colostro (Figura 3).
A equipe de pesquisa era acionada assim que os funcionários detectavam o
início do trabalho de parto para o deslocamento de pessoal e preparo do material
para a colheita das amostras.
Figura 3 - Nascimento em baias de parição (a); colostragem em baias de
transição (b), localizadas na maternidade – São Paulo – 2016
(a) (b) Fonte: (COSTA, J. F. R., 2016).

47
Bezerras hígidas advindas de partos eutócicos foram selecionadas por meio
do exame clínico geral, segundo os procedimentos descritos por Dirksen et al.
(2008). Foram aferidas as frequências cardíaca (FC), respiratória (FR) e temperatura
corpórea (Quadro 1). Outros critérios foram usados para determinar a vitalidade das
bezerras, tais como: tônus muscular, irritabilidade reflexa e coloração das mucosas
oculares (RODRIGUES, 2008).
Quadro 1 - Intervalos de referência para as funções vitais em bezerras no primeiro mês de vida
Parâmetros Intervalos de Referência
1hora após nascimento 1ª semana 2ª semana 3ª semana 4ª semana
FC (bpm) 100,0-150,0 124,0±2,0 105,0±3,0 101,0±3,0 86,0±2,0
FR (mpm) 50,0-75,0 59,0±8,8 46,0±7,0 40,0±8,4 33,0±9,0
TEMP (°C) 39,0-39,5 38,6±0,3 39,0±1,0 39,0±0,4 39,0±0,3,0 Fonte: (SILVA et al., 2016, no prelo)2
Vinte bezerras hígidas foram triadas e distribuídas em dois grupos
denominados COL+ (n=10) e COL- (n=10). As bezerras do grupo COL+ receberam
colostro fresco com células proveniente de suas respectivas mães, enquanto os
animais COL- receberam colostro congelado sem células viáveis, oriundo de vacas
doadoras.
Vacas e doadoras foram ordenhadas imediatamente após a parição em
tronco de contenção localizado no interior da maternidade. Inicialmente foi realizada
a lavagem com solução clorada e secagem dos tetos com papel toalha individual.
O colostro das doadoras foi armazenado pelos funcionários da fazenda em
garrafas plásticas de 2L, mantidas sob refrigeração (4 - 8°C), por 24 horas. Por fim,
as garrafas foram estocadas a -20°C durante o período mínimo de 24 horas e
máximo de 3 meses.
Inicialmente, para justificar a ausência de leucócitos viáveis nas amostras
congeladas, foi realizado um projeto piloto pela equipe. Alíquotas de colostro fresco, 2 SILVA, B. T.; HENKLEN, A.; MARQUES, R. S.; OLIVEIRA, P.; L.; LEITE, S. B. P.; NOVO, S. M. F.; BACCILI, C. C.; REIS, J. F.; GOMES, V. Vital parameters of Holstein calves from birth to weaning. Revista Brasileira de Medicina Veterinária. [200-]. (no prelo).

48
refrigerado e congelado foram avaliadas quanto à sua viabilidade celular, de hora
em hora, durante as primeiras seis horas e, após, entre 18-24 horas. Com esse
estudo foi possível comprovar que as amostras de colostro congeladas não
apresentaram viabilidade celular após o congelamento (0%) desde os momentos
iniciais (NOVO et al., 2014).
A avaliação indireta da concentração de Igs do colostro foi realizada por meio
de colostrômetro e refratômetro Brix (23-32%). Foi possível determinar amplitudes
de variação entre 80 e 120g/L de Igs no colostro das vacas COL+; e 70 a 120g/L
para as vacas COL-.
A quantidade de células somáticas (1,9 milhões/mL; log10: 6,28) encontrada
no colostro foi estimada a partir das amostras de colostro fresco utilizando-se como
metodologia a contagem microscópica direta adaptada por Gomes et al. (2011)
(Figura 4).
Figura 4 - Esfregaço da secreção mamária do colostro fresco fornecido para as bezerras do grupo com células viáveis (COL+) – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016). Legenda: (a) macrófagos presentes em predominância no colostro; (b) neutrófilo; (c) glóbulos de gordura.
A primeira garrafa de colostro foi descongelada lentamente a 37°C para
alimentar as bezerras COL– dentro das primeiras seis horas de vida. A segunda
alimentação foi realizada no intervalo máximo de doze horas de vida, utilizando-se
outra garrafa de colostro descongelado.
(a)
(b)
(c)

49
Bezerras COL+ foram alimentadas com dois litros de colostro fresco nas
primeiras seis horas de vida. O colostro para a segunda mamada foi mantido sob
refrigeração a 4°C, levemente aquecido a 37°C pouco antes da sua administração
entre seis a 12 horas pós-nascimento.
A cura do umbigo foi realizada pela imersão completa do cordão umbilical em
recipiente contendo tintura de iodo a 10% por pelo menos 1 minuto e meio,
repetindo-se o processo após 12 horas. A partir do segundo dia, foi utilizada tintura
de iodo a 5% duas vezes ao dia até mumificação do cordão umbilical.
As bezerras COL+ e COL- foram mantidas inicialmente nas baias dentro da
maternidade, local onde foram realizados os exames clínicos iniciais, colostragem e
cura do umbigo, sendo posteriormente transferidas para o bezerreiro às 6 horas de
vida, onde permaneceram em baias de transição feitas de madeira não suspensas
até os primeiros três primeiros dias pós-nascimento. Por fim, os animais foram
transferidos e permaneceram em gaiolas de metal individuais suspensas, medindo
1,2m x 1m, em local com capacidade para 100 gaiolas (Figura 5).

50
Figura 5 - Alojamento das bezerras durante o primeiro mês de vida – São Paulo – 2016
(a)
(c) (b)
Fonte: (COSTA, J. F. R., 2016). Legenda: (a) alojamento das bezerras recém-nascidas durante as primeiras horas de vida para exame clínico geral, colostragem e cura do umbigo; (b) baia de transição não suspensa onde ficaram até o terceiro dia de vida; (c) bezerreiro para onde as bezerras foram transferidas e permaneceram até o fim do experimento.
A partir deste momento, os animais foram alimentados em baldes, duas vezes
ao dia, sendo fornecido um total de seis litros de leite sem antibiótico proveniente do
rebanho, além do fornecimento de ração comercial3 e água ad libitum.
4.2 MOMENTOS DE AVALIAÇÃO
As bezerras foram avaliadas periodicamente logo após o nascimento (D0),
entre 24 a 48 horas de vida (D2) e nos dias 7, 14, 21 e 28 pós-nascimento (D7 a
D28). 3 Rumileite 20®, Guabi

51
4.3 AVALIAÇÃO CLÍNICA PERIÓDICA
Os animais foram avaliados clinicamente utilizando-se os escores de diarreia
e broncopneumonia de acordo com o Calf Heath Scoring Criteria previamente
publicado na University of Wiscosin - Madison (McGUIRK, 2008).
A consistência das fezes foi categorizada em escores, sendo 0 – consistência
normal, 1 – consistência pastosa e semi-formada, 2 – consistência aquosa com
maior quantidade de água, com conteúdo fecal aderido no períneo e cauda, 3 –
líquida, conteúdo fecal aderido no períneo e cauda. Diarreia foi considerada quando
o escore obtido foi 2 ou 3 (Quadro 2 e Figura 6).
Broncopneumonia foi estabelecida considerando-se os seguintes parâmetros:
temperatura retal, tosse, secreção nasal e ocular e posição de orelhas. Para cada
item foi atribuído o escore de zero a 3 de acordo com a severidade dos sintomas. A
soma dos sintomas acima de 5 foi utilizada para a detecção da Doença Respiratória
Bovina (Quadro 3).
A região umbilical externa foi avaliada por inspeção e palpação periódica para
a detecção de sinais inflamatórios.
Quadro 2 - Escores de fezes adotado para detecção da diarreia em bezerras no primeiro mês de vida
Parâmetros Escores
Consistência normal: firme, coloração amarronzada, períneo e cauda limpos e secos 0 Pastosa, semi-formada 1 Pastosa com maior quantidade de água: permanece sobre a “cama”; conteúdo fecal aderido ao períneo e cauda 2
Líquida: aquosa, conteúdo fecal aderido ao períneo e cauda 3 Fonte: (McGUIRK, 2008).

52
Figura 6 - Escores de fezes adotados para detecção da diarreia em bezerras no primeiro mês de vida – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
Quadro 3 - Escores de broncopneumonia adotados para detecção da doença em bezerras no primeiro mês de vida
Escores Temperatura retal ( OC) Tosse Secreção nasal Secreção ocular Posicionamento
das orelhas 0 Ausente Serosa Serosa Normal
1 38,3 - 38,8 Presente e única,
quando estimulada
Pouca quantidade,
unilateral Pouca quantidade
Balançar das orelhas ou
cabeça
2 38,9 - 39,3
Presente e repetida quando estimulada, ou
ocasional quando espontânea
Excessiva, mucosa e bilateral
Moderada quantidade
bilateral
Ligeiramente pendente, unilateral
3 ≥ 39,4 Presente, repetida
e espontânea
Abundante, mucopurulenta,
bilateral
Intensa quantidade,
bilateral
Pendentes intensamente,
bilateral ou torção da cabeça
Fonte: (POULSEN; McGUIRK, 2009).
4.4 ANÁLISES
A contagem bacteriana total (CBT) e de coliformes totais (CCT) do colostro foi
analisada através da cultura das amostras em meio Tryptic Soy agar (TSA) (Anexo
A) e agar MacConkey (MC) (Anexo B).

53
4.4.1 CBT e CCT
Amostras de colostro fornecidas às bezerras dos grupos COL+ e COL- foram
coletadas de forma asséptica de dentro da mamadeira e armazenadas em tubos
plásticos estéreis com capacidade para 15 mL. Os tubos foram armazenados em
freezer a -80°C até o momento do processamento das amostras.
Amostras de colostro foram descongeladas até 4°C, homogeneizadas com o
uso de vortex e submetidas a 5 diluições seriadas 1:10. Cada diluição foi pipetada
(100 µL) em placa de Petri contendo meio agar MacConkey para a CCT e meio TSA
para a CBT, conforme descrito por Godden (2012) (Figuras 7 e 8). As placas foram
incubadas por 48 horas, a 37°C, e o número de colônias foi convertido para ufc/mL
de acordo com quadro 4.
Figura 7 - Etapas para a diluição seriada e plaqueamento das amostras de colostro
fornecido às bezerras (COL+) e (COL-) – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016). Legenda: SOL: Solução

54
Figura 8 - Crescimentos bacterianos na placa de TSA – CBT (a); crescimento de bactérias Gram-negativas na placa com meio de cultura MacConkey (b) – São Paulo – 2016
(a) (b)
Fonte: (COSTA, J. F. R., 2016).
Quadro 4 - Fatores para conversão de números de colônias na placa TSA e MacConkey
para CBT/mL ou CCT/mL de colostro
Diluição 10-1 Multiplicar o no de colônias que cresceram na placa por 100
Diluição 10-2 Multiplicar o no de colônias que cresceram na placa por 1.000
Diluição 10-3 Multiplicar o no de colônias que cresceram na placa por 10.000
Diluição 10-4 Multiplicar o no de colônias que cresceram na placa por 100.000
Diluição 10-5 Multiplicar o no de colônias que cresceram na placa por 1.000.000
Fonte: (COSTA, J. F. R., 2016).
4.4.2 Isolamento das bactérias das fezes
A colonização bacteriana do trato gastrintestinal das bezerras foi analisada
através da cultura das fezes em meio agar sangue de carneiro 5% (Anexo C), agar
MacConkey e Salmonella-Shigella (SS) (Anexo D).

55
4.4.2.1 Amostra de fezes
As amostras de fezes foram obtidas usando luvas estéreis, diretamente da
ampola retal e foram armazenadas em tubos coletores universais estéreis (Figura 9).
Em seguida, uma alça flexível foi utilizada para coletar amostra de dentro do tubo
coletor que foi transferida para tubos de vidro contendo meio de transporte
Tetrationato (10mL) (Anexo E) e Iodo-Iodeto (200µL) (Anexo F). Ambas as amostras
foram mantidas refrigeradas a 4°C e processadas no prazo máximo de seis horas.
Figura 9 - Demonstração da coleta de fezes com luva estéril direto da ampola retal – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016). 4.4.2.2 Processamento das amostras de fezes
Ao chegarem no laboratório, as amostras no frasco com tetrationato foram
incubadas em estufa bacteriológica durante 24 horas a 37°C. Após 24 horas de
incubação o frasco foi homogeneizado e a amostra foi semeada na placa contendo
agar Salmonella-Shigella (SS) nas condições descritas por Edwards; Ewing (1972).

56
As amostras mantidas no tubo coletor universal foram semeadas em placas
contendo agar sangue, agar MacConkey e SS. As placas foram incubadas a 37°C,
durante 24 horas. Após 24 horas de incubação, as placas que apresentaram
crescimento foram retiradas e protegidas em papel filme para melhor
armazenamento na geladeira até o momento da realização da série bioquímica. As
outras que não apresentaram crescimento nas primeiras 24 horas foram mantidas
na estufa até completarem 48 horas (Figuras 10 a 12).
Os microrganismos foram identificados segundo suas características de cultivo
morfo-tintoriais e séries bioquímicas com os açúcares: fenilalanina, citrato, indol,
vermelho de metil, avaliação da motilidade, produção de gás, lisina e ornitina
(KRIEG; HOLT, 1984) (Figura 11).
Figura 10 - Fezes foram semeadas no fluxo, na presença do fogo em placas agar sangue, agar MacConkey e SS – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).

57
Figura 11 - Esquema do procedimento realizado no laboratório para identificação das bactérias aeróbias das fezes – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016). Figura 12 - Exemplos dos crescimentos obtidos após 24 a 48 horas de incubação das placas em
estufa a 37°C – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).

58
4.5 ANÁLISE ESTATÍSTICA
A análise estatística foi realizada utilizando-se o programa estatístico SPSS
19.0 (IBM Corp. Released 2011. IBM SPSS Statistics for Windows, Version 19.0.
Armonk, NY: IBM Corp.). As análises foram consideradas significativas quando
P≤0,05 e tendência quando P>0,05 (*) e <0,1 (†).
Os dados da CBT e CCT inicialmente foram submetidos a uma transformação
logarítmica de base 10 para que fosse possível a realização de análise paramétrica.
Sendo assim, o teste T de Student para amostras independentes foi adotado para
comparação dos valores obtidos no colostro com (COL+) e sem células (COL-), e a
análise de variância (ANOVA) realizada para comparação entre os tempos (D0 a
D28) dentro do mesmo grupo. No caso onde ocorreram as transformações
logarítmicas, o p apresentado nas tabelas diz respeito aos testes relacionados com
os valores em log10, enquanto os valores descritos são reais.
As espécies e gêneros bacterianos obtidos a partir das fezes estão
apresentas em frequências. As espécies que obtiveram frequência superior a 5%
foram submetidas ao teste Qui-quadrado e Exato de Fisher para verificar se houve
associação com o tipo de colostro oferecido (COL+/COL-) e com a presença de
diarreia.
Nem todas as bezerras apresentaram sinais clínicos de doenças, entretanto,
as frequências positivas e negativas de escore para cada parâmetro foram avaliadas
pelo teste Qui-quadrado.
A correlação das IgG com as bactérias do colostro nos dois grupos COL+ e
COL- foram realizadas a partir da correlação de Pearson.

59
5 RESULTADOS
Este trabalho avaliou a influência das células do colostro na colonização do
trato gastrointestinal de bezerras Holandesas no período neonatal.
5.1 QUALIDADE DO COLOSTRO
A avaliação indireta da concentração de Igs e sólidos totais do colostro
administrado para as bezerras COL+ e COL- foi avaliada por meio de colostrômetro
e índice Brix, respectivamente. Foi possível determinar amplitudes de variação entre
80 e 120g/L de Igs no colostro das vacas COL+; e 70 a 120g/L para as vacas COL-
(Tabela 1).
Tabela 1 - Valores médios, mínimos e máximos de imunoglobulinas avaliados pelo colostrômetro e Brix para (COL+) e (COL-) – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
5.2 STATUS SANITÁRIO
A diarreia foi a doença mais frequente no período neonatal, seguida por
inflamações umbilicais e Doença Respiratória Bovina.
A incidência de diarreias foi avaliada a partir da tabulação de dados do
Parâmetros Colostrômetro (mg/dL)
p COL+ COL- Média 9700±1,17 10200±1,20 0,575 Mínimo 8000 7000 Máximo 12000 12000 Brix (%)
COL+ COL-
Média 27,6±3,69 28,5±3,78 0,596 Mínimo 23,0 23,0
Máximo 32,0 32,0

60
escore de fezes estabelecida segundo McGuirk (2008). A diarreia teve início no D2 e
a frequência aumentou do D7 (COL+ = 56%; COL- = 70%) ao D14 (COL+ = 78%;
COL- = 60%). Após estes momentos as bezerras apresentaram uma ligeira
diminuição na frequência de diarreias no D21 COL+ = 56%; COL- = 50%) e D28
(COL+ = 44%; COL- = 50%) (Figura 13).
A região umbilical externa foi avaliada através da inspeção e palpação
periódica para a detecção de sinais inflamatórios. Desta forma, foi possível detectar
que três bezerras do grupo COL- apresentaram inflamação umbilical no D14 (n=2) e
D28 (n=1). Além disso, também foram detectadas duas bezerras com
broncopneumonia em ambos os grupos: COL+ no D14 (n=1) e COL- no D21 (n=1).
Apesar da maior frequência de doenças encontradas no grupo COL-, não foi
possível comprovar diferenças estatísticas entre a frequência de diarreias ou
doenças totais usando o teste Qui-Quadrado.

61
Figura 13 - Frequências (%) do escore fecal em bezerras Holandesas amamentadas com colostro fresco (COL+) e colostro sem células viáveis (COL- ) durante o primeiro mês de vida – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
0
20
40
60
80
100
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D0
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D2
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D7
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D14
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D21
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D28
COL+
COL-

62
5.2.1 CBT e CCT no colostro
Os valores obtidos para CBT e CCT no colostro fornecido às bezerras COL+
e COL- estão expressos na tabela 2 e figura 14.
Foram obtidos valores médios de 0,94 x 106 ufc/mL e 3,93 x 106 ufc/mL para a
CBT do colostro fresco e congelado, respectivamente.
Para a CCT observou-se valores de 0,78 x 105 ufc/mL e 3,01 x 105 ufc/mL no
colostro fresco e congelado, respectivamente.
Não foi possível identificar diferenças entre as contagens bacterianas em
relação aos grupos COL+ e COL-.
Tabela 2 - Mediana, mínimo, máximo, média e desvio padrão da CBT/mL (x106 e log10) e CCT/mL (x105 e log10) nas amostras de colostro com (COL+) e sem células (COL-) – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
CBT/mL Log10 CCT/mL Log10
TOTAL DE BEZERRAS
(n = 20)
Mediana 1,0x106 6,02 0,83x105 4,92
Mínimo 0,046 x106 4,66 0,02x105 3,30
Máximo 15,0x106 7,19 22,1x105 6,34
COL +
Mediana 0,5 x 106 5,76 0,63x105 4,80
Mínimo 0,076 x106 4,88 0,06x105 3,79
Máximo 3,1x106 6,49 1,81x105 5,26
Média e DP 0,94x106 ± 0,92x106 5,98 0,78x105± 0,58 x105 4,90
COL -
Mediana 1,4 x106 6,17 0,99x105 5,00
Mínimo 0,046x106 4,66 0,02x105 3,30
Máximo 15,6x106 7,19 22,1x105 6,34
Média e DP 3,93x106 ± 5,14 x106 6,60 3,01x105± 6,7x105 5,48 Significância p 0,215 0,730

63
Figura 14 - Contagem bacteriana total (CBT/mL) e contagem de coliformes totais (CCT/mL) nas amostras de colostro com (COL+) e sem células (COL-) – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
As amostras de colostro com (COL+) e sem células viáveis (COL-) foram
distribuídas de acordo com a CBT/mL e CCT/mL (Tabela 3).
Em relação à CBT/mL, foram obtidas frequências de 40 e 30; 20 e 10; 20 e
10; 20 e 50% para os intervalos de 0 a 500.000; 500.001 a 1.000.000; 1.000.001 a
1.500.000; e > 1.500.001 nas amostras com (COL+) e sem células viáveis (COL-),
respectivamente.
Em relação à CCT/mL, foram obtidas frequências de 50 e 30; 10 e 20; 30 e
30; 10 e 0 e 0 e 20% para os intervalos de 0 a 500.000; 500.001 a 1.000.000;
1.000.001 a 1.500.000; e > 1.500.001 nas amostras com (COL+) e sem células
viáveis (COL-), respectivamente.
0
1
2
3
4
5
6
7
CBT/mL CCT/mL
Lo
g 10
COL+
COL-

64
Tabela 3 - Distribuição das amostras de colostro com (COL+) e sem células viáveis (COL-) de acordo com a CBT e CCT/mL – São Paulo – 2016
CBT/mL COL+ COL - Total
% Nº % Nº % Nº 0 a 500.000 40 4 30 3 35 7
500.001 a 1.000.000 20 2 10 1 15 3
1.000.001 a 1.500.000 20 2 10 1 15 3
> 1.500.000 20 2 50 5 35 7
Total 100 10 100 10 100 20
CCT/mL COL+ COL - Total
% Nº % Nº % Nº 0 a 50.000 50 5 30 3 40 8
50.001 a 100.000 10 1 20 2 15 3
100.001 a 150.000 30 3 30 3 30 6
150.001 a 200.000 10 1 0 0 5 1
> 200.000 0 0 20 2 10 2
Total 100 10 100 10 100 20
Fonte: (COSTA, J. F. R., 2016).
5.2.2 Relação entre a CBT / CCT com a concentração de Igs no colostro
Os dados obtidos a partir do colostrômetro e refratômetro Brix para a
concentração de IgG foram correlacionados com a contagem bacteriana no colostro
com (COL+) e sem células viáveis (COL-) (Figuras 15 e 16).
Não foi possível observar correlação entre a CBT/mL ou CCT/mL e a
qualidade do colostro avaliada pelo colostrômetro (IgG) ou Brix (sólidos totais). Os
valores de P obtidos a partir da correlação podem ser encontrados na tabela 4.
Tabela 4 - Valores de P para a correlação de Pearson em relação às concentrações de IgG e sólidos totais correlacionados com CBT e CCT obtidas a partir das amostras de colostro com (COL+) e sem células viáveis (COL-) – São Paulo – 2016
Parâmetros CBT CCT
COL+ COL- TOTAL COL+ COL- TOTAL
IgG Colostrômetro (mg/dL) 0,959 0,419 0,651 0,524 0,329 0,369
IgG Brix (%) 0,399 0,319 0,948 0,880 0,159 0,361
Fonte: (COSTA, J. F. R., 2016).

65
Figura 15 - Correlações entre IgG, contagem bacteriana total (CBT), contagem de coliformes totais (CCT) em amostras de colostro com (COL+) e sem células viáveis (COL-) - São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
y = -0,0224x + 4,125R² = 0,0394
3,80
3,85
3,90
3,95
4,00
4,05
4,10
0,00 2,00 4,00 6,00 8,00
IgG
(m
g/d
L)
CBT Log10
Geraly = -0,0278x + 4,1248
R² = 0,0606
3,80
3,85
3,90
3,95
4,00
4,05
4,10
-1,00 1,00 3,00 5,00 7,00
IgG
(m
g/d
L)
CCT Log10
Geral
y = -0,0079x + 4,0271R² = 0,0032
3,85
3,90
3,95
4,00
4,05
4,10
0,00 2,00 4,00 6,00 8,00
IgG
(m
g/d
L)
CBT Log10
COL+y = -0,0281x + 4,1146
R² = 0,0354
3,85
3,90
3,95
4,00
4,05
4,10
0,00 2,00 4,00 6,00
IgG
(m
g/d
L)
CCT Log10
COL+
y = -0,0378x + 4,2334R² = 0,1248
3,80
3,85
3,90
3,95
4,00
4,05
4,10
0,00 2,00 4,00 6,00 8,00
IgG
(m
g/d
L)
CBT Log10
COL-y = -0,0296x + 4,1449
R² = 0,0897
3,80
3,85
3,90
3,95
4,00
4,05
4,10
0,00 2,00 4,00 6,00 8,00
IgG
(m
g/d
L)
CCT Log10
COL-

66
Figura 16 - Correlação entre sólidos totais, contagem bacteriana total (CBT), contagem de coliformes totais (CCT) em amostras de colostro com (COL+) e sem células viáveis (COL-) – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
5.2.3 Isolamento enteropatógenos aeróbios
O cultivo bacteriano das amostras fecais possibilitou a observação de uma
vasta variedade de espécies que colonizam o trato gastrintestinal das bezerras
Holandesas desde o nascimento até completarem o primeiro mês de vida.
y = -1,1064x + 34,632R² = 0,0413
0
5
10
15
20
25
30
35
0,00 2,00 4,00 6,00 8,00
Índ
ice
Bri
x (%
)
CBT Log10
Geraly = -1,5452x + 35,441
R² = 0,0802
0
5
10
15
20
25
30
35
0,00 2,00 4,00 6,00 8,00
IgG
Bri
x (%
)
CCT Log10
Geral
y = 0,6664x + 23,763R² = 0,0082
0
5
10
15
20
25
30
35
0,00 2,00 4,00 6,00 8,00
Índ
ice
Bri
x (%
)
CBT Log10
COL+y = -0,8788x + 31,756
R² = 0,0122
0
5
10
15
20
25
30
35
0,00 2,00 4,00 6,00
Índ
ice
Bri
x (%
)
CCT Log10
COL+
y = -2,263x + 42,391R² = 0,2241
0
5
10
15
20
25
30
35
0,00 2,00 4,00 6,00 8,00
Índ
ice
Bri
x (%
)
CBT Log10
COL-y = -1,8295x + 37,35
R² = 0,1711
0
5
10
15
20
25
30
35
0,00 2,00 4,00 6,00 8,00
Índ
ice
Bri
x (%
)
CCT Log10
COL-

67
A frequência e o número de gêneros bacterianos isolados a partir das
amostras de fezes das bezerras que receberam colostro fresco (COL+) e congelado
(COL-), do nascimento aos 28 dias de vida, estão expressos na tabela 5.
Observou-se similaridade entre o número de isolados nas fezes das bezerras
COL+ e COL-. O menor número de cepas foi obtido no D0 (COL+=8; COL-=8), e o
pico máximo de isolamentos foi observado no D2 para ambos os grupos (COL+=26;
COL-=27). Ao final do experimento, o número de isolados diminuiu (COL+=12; COL-
=16).
Em ambos os grupos, o momento com menor número de isolamentos foi o D0
com 8 isolamentos. Os gêneros presentes no grupo COL+ foram: Bactéria não
fermentativa (25%), Enterobacter spp (25%), Klebsiella spp (25%), Citrobacter spp
(12,50%) e Staphylococcus spp (12,50%). Os gêneros presentes no mecônio do
grupo COL- foram: Enterobacter spp (25%), Klebsiella spp (25%), Citrobacter spp
(12,50%), Bactéria não fermentativa (12,50%), Morganella spp (12,50%) e Proteus
spp (12,50%).
No grupo COL+ a bactéria mais isolada foi Escherichia coli com 9 isolamentos
que correspondem a 45,00% dos isolamentos no D14 e 9 isolamentos de Proteus
spp que também correspondem a 45,00% no D7.
No grupo COL- a bactéria mais isolada também foi Escherichia coli, observou-
se 9 isolamentos (52,94%) no D21. Isolou-se 9 cepas de Proteus spp (33,33%) no
D2 e 9 cepas (47,37%) no D7.

68
Tabela 5 - Frequências (%) e número de isolamentos por gêneros bacterianos nas amostras de fezes das bezerras que ingeriram colostro com (COL+) e sem células viáveis (COL-) do nascimento aos 28 dias de vida – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016)
As espécies de bactérias aeróbias mais frequentes isoladas das amostras de
fezes das 20 bezerras do nascimento aos 28 dias de vida foram: Proteus mirabilis
(28,84%), Escherichia coli (28,37%), e Klebsiella pneumoniae (6,05%) (Tabela 6).
Foi possível observar que as espécies que estiveram em maior frequência
nas fezes durante o estudo para o grupo COL+ foram: Proteus mirabilis (29,91%),
Escherichia coli (28,04%), Citrobacter freundii (5,61%) e Staphylococcus spp
(5,61%) (Tabela 6).
Gêneros
COL+
D0 D2 D7 D14 D21 D28 TOTAL
% No % No % No % No % No % No % No
Bactéria não fermentativa 25,00 2 0,00 0 0,00 0 0,00 0 0,00 0 7,69 1 2,83 3
Citrobacter spp 12,50 1 19,23 5 5,00 1 5,00 1 5,26 1 0,00 0 8,49 9
Enterobacter spp 25,00 2 11,54 3 0,00 0 5,00 1 0,00 0 7,69 1 6,60 7
Escherichia spp 0,00 0 19,23 5 25,00 5 45,00 9 36,84 7 30,77 4 28,30 30
Klebsiella spp 25,00 2 15,38 4 0,00 0 10,00 2 15,79 3 7,69 1 11,32 12
Morganella spp 0,00 0 3,85 1 10,00 2 0,00 0 0,00 0 0,00 0 2,83 3
Proteus spp 0,00 0 26,92 7 45,00 9 35,00 7 31,58 6 30,77 4 31,13 33 Pseudomonas spp 0,00 0 0,00 0 0,00 0 0,00 0 0,00 0 0,00 0 0,00 0 Salmonella spp 0,00 0 3,85 1 0,00 0 0,00 0 0,00 0 0,00 0 0,94 1 Staphylococcus spp 12,50 1 0,00 0 15,00 3 0,00 0 5,26 1 7,69 1 5,66 6 Streptococcus spp 0,00 0 0,00 0 0,00 0 0,00 0 5,26 1 0,00 0 0,94 1 Total cepas 100 8 100 26 100 20 100 20 100 19 100 12 100 105
Gêneros
COL -
D0 D2 D7 D14 D21 D28 TOTAL
% No % No % No % No % No % No % No
Bactéria não fermentativa 12,50 1 0,00 0 0,00 0 5,26 1 5,88 1 18,75 3 5,66 6
Citrobacter spp 12,50 1 11,11 3 0,00 0 0,00 0 0,00 0 0,00 0 3,77 4
Enterobacter spp 25,00 2 14,82 4 5,26 1 5,26 1 5,88 1 0,00 0 8,49 9
Escherichia spp 0,00 0 18,52 5 26,32 5 31,59 6 52,94 9 37,50 6 29,25 31
Klebsiella spp 25,00 2 11,11 3 15,79 3 5,26 1 5,88 1 12,50 2 11,32 12
Morganella spp 12,50 1 7,41 2 5,26 1 5,26 1 0,00 0 6,25 1 5,66 6
Proteus spp 12,50 1 33,33 9 47,37 9 42,11 8 17,66 3 18,75 3 31,13 33 Pseudomonas spp 0,00 0 0,00 0 0,00 0 0,00 0 5,88 1 6,25 1 1,89 2 Salmonella spp 0,00 0 3,70 1 0,00 0 0,00 0 0,00 0 0,00 0 0,94 1 Staphylococcus spp 0,00 0 0,00 0 0,00 0 5,26 1 5,88 1 0,00 0 1,89 2 Streptococcus spp 0,00 0 0,00 0 0,00 0 0,00 0 0,00 0 0,00 0 0,00 0 Total cepas 100 8 100 27 100 19 100 19 100 17 100 16 100 106

69
As espécies que estiveram em maior frequência nas fezes durante o estudo
para o grupo COL- foram: Escherichia coli (28,70%), Proteus mirabilis (27,78%),
Klebsiella pneumoniae (11,32%) e Morganella morganii (5,66%) (tabela 6).
As diferentes espécies bacterianas isoladas nas fezes das bezerras podem
ser observadas na figura 17. Foi possível observar comportamento semelhante entre
as espécies para ambos os grupos.
Inicialmente, no D0, foram observadas poucas bactérias quando comparado
aos outros momentos (COL+ = 7 e COL- = 8 espécies diferentes). O pico de
isolamento ocorreu no D2 (COL+ = 12 e COL- = 10 espécies diferentes). Neste
momento foram observadas diversas espécies bacterianas como: Citrobacter
farmeri, Citrobacter freundii, Citrobacter koseri, Enterobacter agglomerans,
Enterobacter gergoviae, Escherichia coli, Klebsiella ornithinolytica, Klebsiella
ozaenae, Klebsiella pneumoniae, Morganella morganii, Proteus mirabilis, Salmonella
enteritidis no COL+ e Citrobacter koseri, Enterobacter agglomerans, Enterobacter
cloacae, Enterobacter sakazakii, Escherichia coli, Klebsiella pneumoniae, Morganella
morganii, Proteus mirabilis, Proteus vulgaris e Salmonella enteridis paratyphia A no
COL-. Nos momentos seguintes houve um decréscimo gradual no número de
isolamentos para o grupo COL+ que atingiu o D28 com uma variedade de 6
espécies diferentes isoladas (Bactéria não fermentativa, Enterobacter agglomerans,
Escherichia coli, Klebsiella pneumoniae, Proteus mirabilis e Staphylococcus sp). O
COL- apresentou um decréscimo porém menos acentuado no número de
isolamentos, finalizando o período também com com 6 espécies diferentes (Bactéria
não fermentativa, Escherichia coli, Klebsiella pneumoniae, Morganella morganii,
Proteus mirabilis e Pseudomonas aeruginosa).

70
Tabela 6 - Número de espécies bacterianas e frequência dos isolamentos das amostras de fezes das bezerras que ingeriram colostro com (COL+) e sem células viáveis (COL-) do nascimento aos 28 dias de vida – São Paulo – 2016
Espécies bacterianas
Total isolados COL+ COL- No % No % No %
Bactéria não fermentativa 9 4,19 3 2,80 6 5,56 Citrobacter farmeri 1 0,47 1 0,93 0 0,00 Citrobacter freundii 6 2,79 6 5,61 0 0,00 Citrobacter koseri 6 2,79 2 1,87 4 3,70 Enterobacter aerogenes 1 0,47 0 0,00 1 0,93 Enterobacter agglomerans 6 2,79 4 3,74 2 1,85 Enterobacter cloacae 6 2,79 2 1,87 4 3,70 Enterobacter gergoviae 1 0,47 1 0,93 0 0,00 Enterobacter sakazakii 2 0,93 0 0,00 2 1,85 Escherichia coli 61 28,37 30 28,04 31 28,70 Klebsiella ornithinolytica 2 0,93 1 0,93 1 0,93 Klebsiella oxytoca 2 0,93 1 0,93 1 0,93 Klebsiella ozaenae 6 2,79 5 4,67 1 0,93 Klebsiella pneumoniae 13 6,05 4 3,74 9 8,33 Klebsiella rhinoscleromatis 1 0,47 1 0,93 0 0,00 Morganella morganii 1 4,19 3 2,80 6 5,56 Proteus mirabilis 9 28,84 32 29,91 30 27,78 Proteus vulgaris 62 3,26 2 1,87 5 4,63 Pseudomonas aeruginosa 7 0,93 0 0,00 2 1,85 Salmonella enteritidis 2 0,47 1 0,93 0 0,00 Salmonella enteridis paratyphia A 1 0,47 0 0,00 1 0,93 Staphylococcus sp 8 3,72 6 5,61 2 1,85 Streptococcus sp 1 0,47 1 0,93 0 0,00 TOTAL 214 100 106 100 108 100
Fonte: (COSTA, J. F. R., 2016)

71
Figura 17 - Comparação entre as diferentes espécies bacterianas isoladas a partir das fezes das
bezerras Holandesas (COL+) e (COL-) durante o primeiro mês de vida – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
0
5
10
15
20
25
30
D0 D2 D7 D14 D21 D28 D0 D2 D7 D14 D21 D28
Col+ Col-
Bactéria não fermentativa Citrobacter farmeri
Citrobacter freundii Citrobacter koseri
Enterobacter aerogenes Enterobacter agglomerans
Enterobacter cloacae Enterobacter gergoviae
Enterobacter sakazakii Escherichia coli
Klebsiella ornithinolytica Klebsiella oxytoca
Klebsiella ozaenae klebsiella pneumoniae
Klebsiella rhinoscleromatis Morganella morganii
Proteus mirabilis Proteus vulgaris
Pseudomonas aeruginosa Salmonella enteritidis
Salmonella enteridis paratyphia A Staphylococcus sp
Streptococcus sp

72
O comportamento dos principais gêneros bacterianos encontrados para os
dois grupos pode ser observado na figura 18.
Klebsiella spp esteve presente em todos os momentos analisados com
exceção do D7 para o grupo COL+. Por outro lado, o momento com maior
isolamento no COL+ se deu na semana anterior, no D2, com 4 isolamentos. O
número máximo de cepas isoladas no grupo COL- ocorreu nos momentos D2 e D7,
com 3 isolamentos em cada tempo.
O momento onde mais observa-se Citrobacter spp dentre o período analisado
foi o D2, pico de COL+ com 5 isolamentos, e pico de COL- com 3 isolamentos. O
COL- apenas apresentou isolamento de Citrobacter spp nos momentos iniciais D0 e
D2, após isso, só foi possível observar para o grupo COL+.
As bactérias não fermentativas estiveram presentes, em maior escala, nos
extremos dos momentos analisados. No D0 o grupo COL+ apresentou 2
isolamentos, enquanto o COL- apresentou apenas 1. No D28, o COL+ apresentou 3
isolamentos e o COL- apenas 1 isolado. No D2 e D7 não foi possível isolar esse
gênero em ambos os grupos.
Enterobacter spp apresentou-se crescente do D0 ao D2, onde se deu o pico
(COL+ = 3 e COL- = 2), em seguida observou-se decréscimo no número de cepas.
No D7 e D21 não foram observados isolamentos para o grupo COL+.
O pico de isolamentos da Morganella spp no grupo COL+ ocorreu no D7,
enquanto o grupo COL- ocorreu no D2, com 2 isolamentos em ambos os grupos. No
D21 não foi possível observar nenhum isolamento em ambos os grupos.
O pico de Staphylococcus spp ocorreu no D7 para o grupo COL+ com 3
isolamentos. O grupo COL- apresentou apenas dois isolamentos sendo 1 no D14 e
outro no D21.
Os isolamentos de Proteus spp em ambos os grupos foram crescentes, tendo
o pico no dia 7 para COL+ e COL-, obtendo-se 9 isolamentos, seguido de um
decréscimo nos momentos subsequentes até alcançar 4 e 3 isolamentos no D28
para os grupos COL+ e COL-, respectivamente (Figura 18).
O perfil de isolamentos para a Escherichia coli foi diferente entre os grupos.
Observou-se pico de isolamentos para o COL+ no D14, entretanto, o pico no COL-
ocorreu no D21.

73
Figura 18 - Número de isolamentos dos principais gêneros bacterianos isolados nas amostras de fezes das bezerras Holandesas que ingeriram colostro com (COL+) e sem células viáveis (COL-) – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
0
2
4
6
8
10
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
D0 D2 D7 D14 D21 D28
No
iso
lam
ento
s
Escherichia coli
0
2
4
6
8
10
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
D0 D2 D7 D14 D21 D28
No
iso
lam
ento
s
Proteus spp
0
1
2
3
4
5
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
D0 D2 D7 D14 D21 D28
No
iso
lam
ento
s
Klebsiella spp
0
1
2
3
4
5
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
D0 D2 D7 D14 D21 D28
No
iso
lam
ento
s
Enterobacter spp
0123456
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
D0 D2 D7 D14 D21 D28
No
iso
lam
ento
sCitrobacter spp
0
1
2
3
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
D0 D2 D7 D14 D21 D28
No
iso
lam
ento
s
Morganella spp
0
1
2
3
4
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
D0 D2 D7 D14 D21 D28
No
iso
lam
ento
s
Staphylococcus spp
0
1
2
3
4
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
CO
L+
CO
L-
D0 D2 D7 D14 D21 D28
No
iso
lam
ento
s
Bactéria não fermentativa

74
Além disso, outros gêneros foram isolados, porém com menor frequência
durante o experimento, sendo eles: Pseudomonas spp, Salmonella spp e
Streptococcus spp.
As Pseudomonas spp não estiveram presentes durante o período estudado
para o grupo COL+ e foram isoladas apenas nos momentos D21 e D28 do grupo
COL-, obtendo-se 1 isolamento em cada tempo.
A Salmonella spp foi isolada duas vezes durante o experimento, uma em
cada grupo. Ambos os grupos apresentaram um isolamento de Salmonella spp no
D2.
O Streptococcus spp foi isolado apenas uma vez no grupo COL+, no D21. O
grupo COL- não apresentou esse isolamento durante o período estudado.
Para avaliarmos a possível influência das bactérias mais frequentemente
isoladas (Escherichia coli, Proteus mirabilis e Klebsiella pneumoniae) em relação ao
tipo de colostro fornecido e frequência de diarreias, foram realizados os testes Qui-
Quadrado e Exato de Fischer, porém, não foi possível estabelecer nenhum tipo de
relação (P>0,05).
A primeira associação realizada foi entre bactéria x grupo (COL+/ COL-) onde
tentou-se estabelecer associação para verificar se a frequência bacteriana havia
sido influenciada pelo tipo de colostro fornecido (P>0,05).
A segunda associação estabelecida foi entre a diarreia e a bactéria para
avaliar se a diarreia (ausente/presente) influenciou na frequência das bactérias mais
frequentemente encontradas (P>0,05).

75
6 DISCUSSÃO
O colostro bovino possui múltiplas funções, além daquelas relacionadas à
transferência de fatores imunes aos recém-nascidos. Esta pesquisa comprovou que
o colostro tem potencial para carrear inúmeras bactérias aos recém-nascidos, que
podem ter interferido diretamente na colonização do trato gastrintestinal, bem como
influenciado no status de saúde das bezerras que desenvolveram episódios
importantes de diarreia, os quais serão discutidos a seguir. Entretanto, a presença
de células no colostro não influenciou na colonização do intestino por bactérias
aeróbias isoladas a partir das fezes.
6.1 QUALIDADE DO COLOSTRO
A celularidade do colostro fornecido às bezerras deste experimento foi
avaliada apenas no grupo COL+ para leucócitos totais, pois o congelamento do
colostro fornecido ao grupo COL- promoveu a lise das células. Esta análise foi
realizada para verificar a quantidade de células recebidas pelas bezerras.
A CCS (1.895.844 células/mL) do colostro obtida neste estudo foi superior ao
valor de 878.000 células/mL encontrado por Gomes et al. (2011). Considerando-se a
concentração celular obtida, pode-se estimar que as bezerras receberam um total de
células de 7,58x109 em quatro litros de colostro. Silva (2014) relatou que o colostro
de primeira ordenha apresenta 32% de células viáveis. Desta forma, pode-se
estimar a ingestão de aproximadamente 2,42x109 células viáveis no volume mínimo
equivalente a quatro litros de colostro administrado às bezerras recém-nascidas.
A concentração de Igs do colostro dos dois grupos experimentais
apresentaram valores adequados, segundo o limiar (50g/L) estabelecido por
Chigerwe et al. (2008); Godden (2008); Morril et al. (2012) e Quigley et al. (2013).
Assim, foi possível afirmar que todas as bezerras deste estudo receberam colostro
de qualidade, com teores de Igs acima do ponto de corte recomendado pela
literatura.

76
A ausência de diferenças estatísticas entre os valores obtidos para a
concentração de imunoglobulinas do colostro para o COL+ e COL- permitiu a
confirmação da padronização da qualidade do colostro fornecido para os dois grupos
experimentais.
6.2 STATUS SANITÁRIO
A frequência da diarreia observada neste estudo foi maior no COL+ (44-78%)
e COL- (50-70%) do que a prevalência (19,1%) e índice de risco (21,2%) reportados
por outros pesquisadores (BARTELS et al., 2010; WINDEYER et al., 2014). Na
fazenda onde este experimento foi desenvolvido, as mães foram vacinadas no
período pré parto para Escherichia coli, Rotavirus e Coronavirus. Assim, a diarreia
encontrada neste experimento deveria ser menor do que a real incidência da
propriedade. Megank et al. (2015) provaram que a vacinação das mães reduziu a
frequência de diarreia de 39,9% para 14,3% em bezerros recém-nascidos.
A frequência das diarreias foi similar entre os grupos, entretanto apenas o
grupo COL- apresentou inflamações umbilicais (n=3). Foram detectadas duas
bezerras com broncopneumonia, uma pertencente a cada grupo. Langel et al. (2015)
avaliaram a saúde das bezerras que receberam colostro fresco e congelado usando
escores de fezes e escores respiratórios. Estes autores também não encontraram
diferenças estatísticas para a frequência de diarreia entre os grupos, entretanto o
escore respiratório foi maior no grupo COL- no D38. Nossos achados de
broncopneumonia não eram esperados pela faixa etária em que as bezerras se
encontravam, pois, inicialmente a bezerra está protegida pelos anticorpos maternos
adquiridos pelo colostro (RIDIPATH; BOLIN, 1995); entretanto, a metabolização
destes componentes e a diminuição da sua concentração no plasma são fatores
predisponentes para a instauração de quadros clínicos, como os respiratórios, a
partir do primeiro mês de vida (BABIUK et al., 2004).

77
6.2.1 CBT e CCT no colostro
Foram obtidos valores médios de 0,94 x 106 ufc/mL (Log10: 5,98) e 3,93 x 106
ufc/mL (Log10: 6,60) para a CBT do colostro fresco e congelado, respectivamente.
Para a CCT observou-se valores de 0,78 x 105 ufc/mL (Log10: 4,90) e 3,01 x 105
ufc/mL (Log10: 5,48) no colostro fresco e congelado, respectivamente. Considerando
a CBT e CCT do colostro congelado pode-se afirmar que a maioria das bactérias
eram coliformes fecais; sendo assim, pressupõe-se que a maior contagem
bacteriana total destes grupos pode estar associada à contaminação entre a colheita
e o fornecimento do colostro às bezerras.
Godden et al. (2012), obtiveram valores inferiores de CBT (3,5 log10 CBT/mL)
e valor superior para a CCT (5,6 log10 CCT/mL) em amostras de colostro fresco,
quando comparadas aos valores obtidos neste experimento. As diferenças nos
valores, apesar de não terem sido muito discrepantes, podem ter se dado por
importantes fatores de risco para a contaminação do colostro, desde a ordenha até a
coleta das amostras frescas, levando-se em consideração os seguintes itens:
identificação e separação de vacas infectadas; impedimento da bezerra de mamar
diretamente da mãe; preparação da higiene da glândula mamária antes e após a
ordenha; higienização do equipamento de ordenha; armazenamento e limpeza dos
equipamentos de alimentação, além da influência do local, pois um experimento foi
realizado no Brasil, com suas condições climáticas e o trabalho em questão foi
realizado em Minnesota, EUA, o que pode ter influenciado na diferente carga
bacteriana das amostras do colostro.
Apesar de não terem sido detectadas diferenças estatísticas entre os grupos
de bezerras que ingeriram colostro fresco (COL+) e congelado (COL-), tanto a
mediana quanto a média da CBT e CCT foram superiores no grupo que ingeriu
colostro congelado. Este resultado não seria inicialmente esperado, caso
considerássemos que a amostra de colostro fornecida às bezerras COL- tivesse sido
congelada imediatamente após ter sido ordenhada e envazada. Infelizmente isto não
ocorreu conforme o planejado, pois este estudo foi realizado em fazenda comercial e
não havia a possibilidade de interferir diretamente no manejo e protocolo de
colostragem da propriedade.

78
O colostro é mantido em geladeira 24 horas antes do seu congelamento,
aguardando caso alguma bezerra necessite tomá-lo. Caso não seja necessário o
fornecimento deste colostro, a mesma garrafa é retirada da geladeira e armazenada
no freezer, onde fica congelado até o momento em que precisar descongelar para
fornecer a alguma bezerra ou, no caso do experimento, para fornecer ao grupo
COL-. É interessante ressaltar que este manejo, que tem ótima intenção por parte da
fazenda, que tenta prezar pela viabilidade dos leucócitos colostrais ao mantê-los
refrigerados e não congelados, não é o ideal, pois estudos prévios da nossa equipe
comprovaram que a viabilidade celular dos colostros armazenados em geladeira se
mantém estável até 6 horas após ordenha; depois disso a viabilidade diminui
consideravelmente (NOVO et al., 2014).
O tempo de permanência do colostro na geladeira por 24 horas,
provavelmente viabilizou este aumento na proliferação das bactérias psicrotróficas
das amostras, aquelas que apresentam ótima taxa de crescimento em temperaturas
abaixo de 7ºC (FRANK et al., 1992).
As maiores frequências obtidas para a CBT no colostro fresco foram de 40%
para o menor intervalo (0 e 500.000 ufc/mL) e 50% para o maior intervalo (>
1.500.000) no colostro congelado.
Apenas duas amostras estiveram dentro dos valores recomendados como
meta por McGuirk e Collins (2004) para a contagem bacteriana total (<100.000
ufc/mL). Isso quer dizer que 10% de todo o colostro fornecido estava dentro do limiar
de bactérias totais preconizado pelos autores e aceito por grande parte de outros
pesquisadores da literatura.
Para a CCT também observou-se maior frequência das amostras no menor
intervalo (0 a 50.000 ufc/mL) para o colostro fresco (50%). As maiores frequências
para o colostro congelado foram encontradas nos intervalos de 0 a 50.000 (30%) e
100.001 a 150.000 ufc/mL (30%).
Apenas três amostras estiveram dentro dos valores recomendados para CCT
por McGuirk e Collins (2004) (<10.000 ufc/mL). Destas, duas amostras eram
pertencentes ao COL- e uma ao COL+. O fato de 17/20 amostras estarem acima dos
índices mundialmente aceitos por outros pesquisadores da área, nos faz pensar em
duas questões: a primeira delas é a qualidade higiênica do colostro que se é
fornecido às bezerras e no manejo que se é realizado com o colostro desde a
ordenha até o fornecimento ao animal, uma vez que neste intervalo ocorre a

79
proliferação de bactérias; a outra questão a ser considerada é se estes valores
preconizados por McGuirk e Collins (2004) e aceitos por outros pesquisadores, não
seriam um tanto quanto baixos e utópicos ao considerar a realidade ambiental e da
rotina da fazenda.
Outro ponto importante a ser considerado é a possibilidade de alguns grupos
bacterianos presentes no colostro serem em parte positivos para o desenvolvimento
do hospedeiro ao invés de patogênicos, como por exemplo os Lactobacillus e
Bifidobacteria, que são grupos bacterianos isolados do colostro e leite materno
humano e comumente utilizados como probióticos. Estas bactérias são importantes
para o crescimento saudável do hospedeiro, pois em estudos humanos,
comprovaram que elas atuam inibindo o crescimento e atividades de bactérias
patogênicas no lúmen do intestino e em bovinos neonatos são responsáveis por
maior ganho de peso e menor índice de diarreia (ABE; ISHIBASHI; SHIMAMURA,
1995; ISOLAURI; SALMINEN; OUWEHAND, 2004).
Os valores de imunoglobulinas obtidos a partir do colostrômetro e
refratômetro Brix para a concentração de IgG foram correlacionados com as
bactérias do colostro (CBT e CCT), porém não foram encontradas correlações.
Todas as bezerras deste experimento foram colostradas com colostro de alta
qualidade e a transferência de imunidade passiva destes animais foi confirmada
através da mensuração da proteína total do soro das bezerras por Blima (2015).
Pesquisadores têm avaliado a contagem bacteriana em comparação não só
ao colostro fresco e congelado, mas também ao colostro pasteurizado, como
importante minimizador da carga bacteriana (GODDEN et al., 2012). Dentro do
contexto apresentado e da alta presença de bactérias no colostro congelado, apesar
de não podermos afirmar se elas eram patogênicas ou benéficas para a microbiota,
pode-se recomendar o uso de pasteurizador de colostro para melhorar o manejo
desta fazenda e minimizar a CCT, pois, mesmo considerando todo o cuidado e
higiene no atual protocolo de manejo, ainda assim não é suficiente para manter os
níveis de bactérias na amostra próximos aos valores recomendados. Ao pasteurizar
o colostro a 60ºC por 60 minutos, Godden et al. (2012) notaram que não houve
alteração na concentração de IgG do colostro e diminuiu riscos de doenças até o
desmame, minimizando índices de morbidade e mortalidade, porém, mencionaram
um ponto negativo que seria a diminuição da viabilidade de leucócitos colostrais.

80
Além disso, ressalta-se a perda das bactérias benéficas Bifidobacterium e
Lactobacillus.
Outros autores também demonstraram que a pasteurização foi bastante
eficaz ao atingir o objetivo de redução de CBT e CCT do colostro (JOHNSON et al.,
2007; ELIZONODO-SALAZAR; HEINRICHS, 2009).
O ideal seria o fornecimento de colostro fresco, pelo menos na primeira
mamada, com baixas CBT e CCT para garantir a viabilidade das células do colostro
e bactérias benéficas. A pasteurização seria uma boa alternativa considerando
rebanhos com altas taxas de mastite no pós-parto imediato, que resulta na falta de
volume adequado de colostro para amamentar as bezerras no primeiro dia de vida.
Recomenda-se que o colostro pasteurizado seja fornecido apenas na segunda
mamada de colostro. Outra questão a ser considerada é o custo do pasteurizador,
por volta de U$33.000 com capacidade para 100 litros.
6.2.2 Isolamentos bacterianos das fezes
O cultivo bacteriano das amostras fecais nos possibilitou acompanhar a
dinâmica bacteriana das bezerras durante o primeiro mês de vida.
A maioria das bezerras ao nascimento apresentaram pouco crescimento
bacteriano no mecônio. Há quem acredite que o útero é um ambiente
completamente estéril (BARRINGTON; PARISH, 2001), mas sabe-se que nem
sempre é assim que ocorre (NOAKES; WALLACE; SMITH, 1991). Hoje em dia
existem técnicas de biologia molecular capazes de determinar a microbiota residente
dos tecidos, como do útero por exemplo. Desta forma, a observação de algumas
bactérias no mecônio foi um resultado esperado.
Após a colostragem, exame clínico e cura de umbigo, os animais foram
transferidos da baia de dentro da maternidade para as baias de madeira no chão, no
ambiente externo da maternidade, no bezerreiro. O pico de isolamento observado
nesta pesquisa ocorreu no D2 (COL+= 12 e COL-= 10 espécies diferentes); próxima
coleta após os animais terem sido transferidos de lugar. Neste momento foram
observadas diversas espécies bacterianas dos gêneros: Citrobacter spp,
Enterobacter spp, Klebsiella spp, Morganella spp, Proteus spp Salmonella spp e

81
Escherichia coli, no COL+. No COL- os gêneros bacterianos isolados foram:
Citrobacter spp, Enterobacter spp, Klebsiella spp, Morganella spp, Proteus spp e
Salmonella spp e Escherichia coli no COL-.
Ao analisarmos as bactérias isoladas neste momento, pode-se observar uma
grande quantidade de bactérias próprias da microbiota, aquelas chamadas de
residentes, como as pertencentes aos gêneros Enterobacter spp e Klebsiella spp.
Estas bactérias são consideradas próprias da microbiota pois são encontradas em
fezes de animais com e sem diarreia (RIBEIRO et al., 2000).
A Salmonella spp e Escherichia coli encontradas em ambos os grupos são
conhecidas por serem grandes responsáveis pela diarreia neonatal bovina. O
mecanismo de virulência principal da Salmonella spp consiste na capacidade de
invadir a mucosa intestinal, se multiplicar em tecidos linfoides e evadir do sistema de
defesa do organismo. A diarreia por Salmonella spp pode se apresentar de forma
aquosa, mucoide e com presença de estrias de sangue. A Escherichia coli
enterotoxigênica (ETEC) é a principal bacteria relacionada à diarreia neonatal
bovina, que produz K99, antígeno de adesão. A ETEC afeta o epitélio intestinal e se
multiplica nos enterócitos das vilosidades do intestino. A região de predileção da
ETEC é a porção distal do intestino delgado devido ao baixo pH da região (ACRES
et al., 1985; FRANCIS et al., 1989; NATARO; KAPER, 1998; CHO; YOON, 2014;
SEEDY et al., 2016).
Este alto número de isolamentos está relacionado a diversos fatores
epidemiológicos como: alteração de ambiente; mudanças climáticas; aumento da
densidade populacional; maior contato com funcionários da fazenda; manejo ao qual
os animais foram submetidos, que se tornam desafios para o animal jovem
(FERREIRA et al., 2009). As bezerras do experimento foram alocadas nesta baia de
madeira no chão (D2), onde há pouca ventilação, pouca entrada de luz, e
principalmente é de difícil higienização, sendo que o ideal seria a realização de
vassoura de fogo com frequência; porém, a fazenda acabava higienizando com cal
virgem e água. Este fato pode ter influenciado no aumento da presença da diarreia
encontrado neste experimento.
No grupo COL+ (D14 = 45%) e COL- (D21= 52,9%) a bactéria mais isolada foi
Escherichia coli. Os momentos deste experimento em que foram isoladas E. coli em
alta quantidade diferem do achado de Rada et al. (2006) que relataram maior
frequência da população bacteriana em questão no terceiro dia, observando-se

82
diminuição acentuada até o sétimo dia. Os valores obtidos a partir de então
oscilaram pouco, apresentando leve aumento do D7 ao D21.
Rada et al. (2006) relacionaram o isolamento de Escherichia coli com os
isolamentos de bifidobacteria. No dia 3, quando a população de E. coli esteve em
alta, a de bifidobacteria estava baixa. Logo em seguida, a população de
bifidobacteria aumentou, enquanto a de E. coli diminuiu, apresentando inversão nos
valores. Essa inversão acentuada de população bacteriana foi justificada pelos
autores por possível alteração na alimentação, uma vez que a bifidobacteria é
considerada um excelente probiótico que possivelmente competiu por espaço com a
Escherichia coli, utilizando-se de mecanismos antibacterianos como, por exemplo, a
diminuição do pH gastrintestinal pela produção de compostos ácidos que inibem o
crescimento de patógenos e inibe a aderência de Escherichia coli aos receptores por
mecanismo de competição, mecanismos estes que minimizam o potencial
inflamatório dos enteropatógenos (BRANDT; CARNEIRO SAMPAIO; MIUKI, 2006;
MARTIN et al., 2012; FERNÁNDEZ et al., 2013).
O gênero Proteus spp que foi o segundo gênero mais isolado neste
experimento, após a Escherichia coli, não esteve associado à diarreia pelo teste de
Qui-Quadrado. Oliveira Filho (2006) também isolou Proteus spp nas amostras de
fezes de bezerros sadios e diarreicos, concluindo que ela não tem relação direta
com a diarreia e faz parte da microbiota residente.
Citrobacter spp foi outro gênero comumente encontrado durante todo o
experimento para o grupo COL+ com exceção do D28. O momento em que essa
bactéria foi mais isolada foi no D2, com 19,23% dos isolamentos. Por outro lado,
esse gênero no grupo COL- foi encontrado apenas no início do experimento no D0 e
D2 (12,50 e 11,11%). O gênero Citrobacter spp não está diretamente associado com
a diarreia pois autores relacionam o gênero com a flora normal do trato
gastrintestinal, porém, em grande quantidade pode ter um possível papel patogênico
(RIBEIRO et al., 2000).
Os principais patógenos causadores de diarreias em bezerros neonatos são
Escherichia coli, Cryptosporidium spp e viroses representadas por Rotavírus e
Coronavírus, respectivamente, encontrados antes e após a primeira semana de vida
(FEITOSA et al., 2008; GULLIKSEN et al., 2009; BARTELS et al., 2010). A natureza
multifatorial da diarreia em bezerros (bactérias, vírus e protozoários) torna esta

83
doença difícil de controlar com eficácia. Além disso, em muitos casos, as diarreias
vêm acompanhadas de outras doenças concomitantes (CHO; YOON, 2014).
Apesar de considerarmos as bactérias como possíveis principais agentes
causadores da diarreia, o que observamos foi que provavelmente outros agentes
que não apenas os bacterianos, estiveram à ela relacionados. Estes dados fazem
com que se possa concluir que a diarreia apresentada pelas bezerras deste
experimento não tenha sido causada necessariamente pelas bactérias que foram
isoladas. Estas bactérias podem, em sua maioria, ser apenas microrganismos
pertencentes à microbiota residente.
As bactérias tradicionalmente identificadas e classificadas utilizando técnicas
de cultura e provas bioquímicas como as desenvolvidas neste trabalho, requerem
um processo bastante demorado e trabalhoso. Muitas bactérias não são fáceis de
cultivar e isolar, principalmente as anaeróbias. O período de cultura pode variar,
dependendo da bactéria e do material utilizado para o isolamento, fazendo com que
o tempo de leitura e resultado sejam variados (ELKJAER et al., 2013).
Embora as técnicas de cultura bacteriana tenham auxiliado na identificação
dos microrganismos, estudos utilizando essas técnicas podem subestimar a
quantidade dos microrganismos presentes no trato gastrintestinal de bovinos
(SANTOS et al., 2011; PENGE et al., 2013). Desta forma, em 1996 foi desenvolvido
o pirosequenciamento por Ronaghi e colaboradores que permitiu o sequenciamento
genético via extração de DNA bacteriano e rDNA ribosomal auxiliando na
compreensão e caracterização mais completa do microbioma (WANG et al., 2012;
AZIZ et al., 2013; KNUDSEN et al., 2014; VAZIRI et al., 2015).
Como consequência do desenvolvimento e difusão das técnicas de
sequenciamento e estudos em metagenômica, aumentou consideravelmenete o
número de trabalhos relacionados ao microbioma humano e animal. Entre os vários
ambientes animais explorados através da metagenômica, o trato gastrointestinal
bovino e o reprodutivo são os mais estudados, no entanto as pesquisas ainda estão
apenas começando (KNUDSEN et al., 2014).
Estes métodos apresentam vantagens em comparação à técnica de cultura
bacteriana tradicional, uma vez que são capazes de detectar o DNA bacteriano e
indicar todas as bactérias presentes na amostra. Esta técnica não depende do
potencial de crescimento bacteriano e detecta todas as bactérias, não apenas as
que podem ser cultivadas. No entanto, apesar de todas essas vantagens, os custos

84
por amostra ainda são muito elevados, limitando o uso desse método (ELKJAER et
al., 2013). As amostras deste experimento estão armazenadas em freezer -80ºC
para futuro sequenciamento metagenômico pois, após o início deste experimento, foi
aprovada a verba para fazermos a análise do DNA bacteriano. Em breve, estes
resultados trarão mais respostas aos nossos questionamentos.

85
7 CONCLUSÃO
Com base nos dados obtidos, pode-se concluir:
(a) Os colostros celular e acelular apresentavam a mesma concentração de
imunoglobulinas, contagem bacteriana total e contagem de coliformes;
(b) A administração de colostro com (COL+) e sem células viáveis (COL-) não
influenciou na proporção de bactérias aeróbias presentes nas fezes das
bezerras durante o período neonatal;
(c) A administração de colostro com (COL+) e sem células viáveis (COL-) não
influenciou na ocorrência de diarreias em bezerras durante o período neonatal.

86
REFERÊNCIAS
ABE, F.; ISHIBASHI, N.; SHIMAMURA, S. Effect of administration of bifidobacteria and lactic acid bacteria to newborn calves and piglets, Journal of Dairy Science, v. 78, p. 2838-2846, 1995. ACRES, S. D. Enterotoxigenic Escherichia coli infections in newborn calves: a review, Journal of Dairy Science ,v. 68, p. 229—256, 1985. ADKINS, M. L. Ecology of transferred maternal Staphylococci and neonatal Staphylococcal immunity. In: SCIENCE OF VETERINARY MEDICINE SYMPOSIUM, 2014, Georgia: University of Georgia, 2014.
AZIZ, Q.; DORE, J.; EMMANUEL, A.; GUARNE, F.; QUIGLEY, E. M. M. Gut microbiota and gastrointestinal health: current concepts and future directions, Neurogastroenterology Motility , v.25, p.4-15, 2013. BABIUK, S.; BACA-ESTRADA, M. E.; FOLDVARI, M.; MIDDLETON, D. M.; RABUSSAY, D.; WIDERA, G., BABIUK, L. A. Increased gene expression and inflammatory cell infiltration caused by electroporation are both important for improving the efficacy of DNA vaccines. Journal of Biotechnology , v. 110, p. 1-10, 2004. BARRINGTON, G. M.; PARISH, S. M. Bovine neonatal immunology. Veterinary Clinics of North America: food animal practice, v. 17, n. 3. p. 463-476, 2001. BARTELS, C. J. M.; HOLZHAUER, M.; JORRITSMA, R.; SWART, W. A. J. M.; LAM, T. J. G. M. Prevalence, prediction and risk factors of enteropathogens innormal and non-normal faeces of young Dutch dairy calves. Preventive Veterinary Medicine , v. 93, p. 162–169, 2010. BLIMA, S. Avaliação de métodos para estimar a qualidade do co lostro bovino e sua relação com os teores de imunoglobulinas transf eridas aos recém-nascidos . São Paulo: [s. n.], 2015. Programa de Iniciação Científica da Universidade de São Paulo. BRANDT, K. G.; SAMPAIO, M. M. S. C.; MIUKI, C. J. Importância da microflora intestinal. Revisões e Ensaios: Pediatria , São Paulo, v. 28, n. 2, p. 117-127, 2006. CHIGERWE, M.; TYLER, J. W.; MIDDLETON, J. R.; SPAIN, J. N.; DILL, J. S.; STEEVENS, B. J. Comparison of four methods to assess colostral IgG concentration in dairy cows. Journal of the American Veterinary Medical Association , v. 233, p. 761-766, 2008. CHO, YONG-IL.; YOON, KYOUNG-JIN. An overview of calf diarrhea - infectious etiology, diagnosis, and intervention. Journal Veterinary Science, v. 15 n.1, p. 1-17, 2013.

87
DIRKSEN, G.; GRÜNDER, H. D.; STÖBER, M. R. Exame clínico dos bovinos . Rio de Janeiro: Guanabara Koogan, 2008. EDWARDS, P. R.; EWING, W. H. Identification of enterobacteriaceae. Minneapolis: Burgess Publication . 3th edition, p. 85-90, 1972. ELIZONDO-SALAZAR, J. A.; HEINRICHS, A. J. Feeding heat-treated colostrum or unheated colostrum with two different bacterial concentrations to neonatal dairy calves. Journal of Dairy Science , v.92, n. 9, p. 4565-4571, 2009. ELKJAER, K.; ANCKER, M. L.; GUSTAFSSON, H.; FRIGGENS, N. C.; WALDMANN, A.; MOLBAK, L.; CALLESEN, H. Uterine bacterial flora in postpartum Danish Holstein dairy cows determined using DNA-based fingerprinting: correlation to uterine condition and calving management. Animal reproduction science , v.138, n. 1, p. 39-48, 2013. FEITOSA, F. L. F.; SHIMAMURA, G. M.; ROBERTO, T.; MENDES, L. C. N.; PEIRÓ,J. R.; FÉRES, F. C.; BOVINO, F.; PERRI, S. H. V.; MEIRELES, M. V. Importância de Cryptosporidium spp. como causa de diarréia em bezerros. Pesquisa Veterinária Brasileira, v. 28 n. 10, p. 452-456, 2008. FERNANDÉZ, L.; LANGA, S.; MARTÍN, V.; MALDONADO, A.; JIMÉNEZ, E.; MARTÍN, R.; RODRÍGUEZ, J. M. The human milk microbiota: origin and potential roles in health and disease. Pharmacological Research , v. 69, p. 1-10, 2013. FERREIRA, M. G.; FACURY FILHO, E. J.; HEINEMANN, M. B.; CARVALHO, A. Ú.; LAGE, A. Ú.; FERREIRA, P. M.; FREITAS, M. D. Prevalência de Eimeria, Helmintos, Escherichia coli, Salmonella, Rotavirus, Coronavirus e Cryptosporidium parvum em propriedades leiteiras de Minas Gerais, Brasil. Ciência animal Brasileira, v. 1, p.524-529, 2009. FRANCIS, D. H.; ALLEN, S. D.; WHITE, R.D. Influence of bovine intestinal fluid on the expression of K99 pili by Escherichia coli. American Journal of Veterinary Research , v. 50, p. 822-826, 1989. FRANK, J. F. Tests for groups of microrganisms. In: MARSHALL, R.T. (Ed.). Standard methods for the examination of dairy products. 16.ed. Washington: American Public Health Association , p.271-286, 1992. GODDEN, S. Colostrum management for dairy calves. Veterinary Clinics North America: food animal practice, v. 24, p. 19- 39, 2008. GODDEN, S. M.; SMOLENSKI, D. J.; DONAHUE, M.; OAKES, J. M.; BEY, R.; WELLS, S.; SREEVATSAN, S.; STABEL, J.; FETROW, J. Heat-treated colostrum and reduced morbidity in preweaned dairy calves: Results of a randomized trial and examination of mechanisms of effectiveness. Journal of Dairy Science, v.95, p. 4029-4040, 2012. GOMES, V.; MADUREIRA, K. M.; DELLA LIBERA, A. M. M. P.; ALVES, M.; BAPTISTELLA, F.; BENESI, F. J. Dinâmica da celularidade do colostro de vacas

88
holandesas no pós-parto imediato. Arquivo Brasileiro de Medicina Veterinária e Zootecnia , v. 63, n. 5, p.1047-1053, 2011. GULLIKSEN, S. M.; JOR, E.; LIE, K. I.; HAMNES, I. S.; LOKEN, T.; ÅKERSTEDT, J.; OSTERÅS, O. Enteropathogens and risk factors for diarrhea in Norwegiandairy calves. Journal of Dairy Science , v. 92, p. 5057–5066, 2009. ISOLAURI, E.; SALMINEN, S.; OUWEHAND, A. C. Microbial-gut interactions in health and disease. Probiotics. Best Practice and Research Clinical Gastroenterology , v. 2, p.299-313, 2004. JAYARAO, B. M.; PILLAI, S. R.; SAWANT, A. A.; WOLFGANG, D. R.; HEGDE, N. V. Guidelines for monitoring bulk tank milk somatic cell and bacterial counts. Journal of Dairy Science , v. 87, p. 3561–3573, 2004. JOHNSON, J. L.; GODDEN, S. M.; MOLITOR, T.; AMES, T. HAGMAN, D. Effects of feeding heat-treated colostrum on passive transfer of immune and nutritional parameters in neonatal dairy calves. Journal of Dairy Science , v. 90, p. 5189-5198, 2007. KNUDSEN, L. R. V.; SCHOU, K. K.; JENSEN, T. K.; ANGEN, O. Molecular characterisation of the uterine microbiome of dairy cows suffering from endometritis, metritis and pyometra. Kgs. Lyngby: Technical University of Denmark , 2014 KRIEG, N. R., HOLT, J.C. Bergey´s manual of systematic bacteriology. 9. Ed. Baltimore: Willians & Wilkings , p. 1599, 1984. LANGEL, S. N.; WARK, W. A.; GARST, S. N.; JAMES, R. E.; McGILLIARD, M. L.; PETERSSON-WOLFE, C. S.; KANEVSKY-MULLARKY, I. Effect of feeding whole compared with cell-free colostrum on calf immune status: The neonatal period. Journal of Dairy Science , v. 98, n. 6, p. 3729-3740, 2015. MARTIN, R.; JIMÉNEZ, E.; HEILIG, H.; FERNÁNDEZ, L.; MARÍN, M. L.; ZOETENDAL, E. G.; RODRIGUEZ, J. M. Isolation of bifidobacteria from breast milk and assessment of the bifidobacterial population by PCR-denaturing gradient gel electrophoresis and quantitative real-time PCR. Applied and Environmental Microbiology , v. 75, n. 4, p. 965- 969, 2009. MARTIN, R.; LANGA, S.; REVIRIEGO, C.; JIMÉNEZ, E.; MARÍN, ML.; XAUS, J.; RODRIGUEZ, J. M. Human milk is a source of lactic acid bacteria for the infant gut. Journal of Pediatrics , V. 28, p. 36-44, 2012. MCGUIRE M. K.; MCGUIRE M. A. Human milk: mother nature’s prototypical probiotic food? American Society for Nutrition, v.6, p. 112–123, 2015. MCGUIRK, S. M. Disease management of dairy calves and heifers. Veterinary Clinics of North America, food animal practice, v. 24, n.1, p. 139-156, 2008. MCGUIRK, S. M., AND M. COLLINS. Managing the production, storage and delivery of colostrum, Veterinary Clinics of North America: Food Animal Pr actice, v.20, p. 593-603, 2004.

89
MEGANCK, V.; OPSOMER, G.; PIEPERS, S.; COX, E.; GODDEERIS, B. M. Maternal colostral leukocytes appear to enhance cell-mediated recall response, but inhibit humoral recall response in prime–boost vaccinated calves. Journal of Reproductive Immunology, 2015. DOI: /dx.doi.org/doi:10.1016/j.jri.2015.11.004 MORRILL, K. M.; QUIGLEY, J. D.; LAGO, A.; TYLER, H. D. Estimate of colostral IgG concentration using refractometry without or with caprylic acid fractionation. Journal of Dairy Science , v. 95, p. 3987–3996, 2012. NATARO, J. P.; KAPER, J. B. Diarrheagenic Escherichia coli. Clinical Microbiology Reviews , v.11, p. 142-201, 1998. NOAKES D. E.; WALLACE, L.; SMITH, G.R. Bacterial flora of the uterus of cows after calving on two hygienically contrasting farms, Veterinary Record, v. 128, n. 19, p. 440-442, 1991. NOVO, S. M. F.; REIS, J. F.; SOBREIRA, N. M.; GOMES, V. Effect of temperature and conservation time on the feasibility of bovine colostrum cells. In: WORLD BUIATRICS CONGRESS, 28., 2014, Cairns, Austrália. Anais... Cairns, Austrália, 2014. OLIVEIRA FILHO, J. P. Diarreia em bezerros da raça Nelore criados extensivamente: estudo clinic e etiológico. 2006. 109 f. Dissertação (mestrado). Universidade Estadual Paulista, Faculdade de Medicina Veterinária e Zootecnia, Botucatu, 2006. PENGE, Y.; WANG, Y.; HANG, S.; ZHU, W. Microbial diversity in uterus of healthy and metritic postpartum Holstein dairy cows. Folia Microbiologica , v. 58, n. 6, p. 593-600, 2013. POULSEN, K. P.; MCGUIRK, S. M. Respirator y diseaseof the bovine neonate. Veterinary Clinics of North America : food animal practice, v. 25, p. 121–137, 2009. QUIGLEY, J. D.; LAGO, A.; CHAPMAN, C.; ERICKSON, P.; POLO, J. Evaluation of the Brix refractometer to estimate immunoglobulin G concentration in bovine colostrum. Journal of Dairy Science , v. 96, p. 1148–1155, 2013. RADA, V.; VLOKOVÁ, E.; NEVORAL, J.; TROJANOVÁ, I. Comparison of bacterial flora and enzymatic activity in faeces of infants and calves. Microbiology Letters , v. 258, p. 25-28, 2006. RIBEIRO, M. G.; LANGONI, H.; JEREZ, J. A.; LEITE, D. S.; FERREIRA, F.; GENNARI, S. M. Identification of enteropathogens from buffalo calves with and without diarrhoea in the Ribeira Valley, State of São Paulo, Brazil. Brazilian Journal of Veterinary Research and Animal Science, v. 37, n.2, 2000. DOI: http://dx.doi.org/10.1590/S1413-95962000000200004

90
RIDPATH, J. F.; BOLIN, S. R. The genomic sequence of a virulent bovine viral diarrhea virus (BVDV) from the type 2 genotypes: detection of a large genomic insertion in a nonpathogenic BVDV. Virology , v. 212, n. 1, p. 39-46, 1995. RODRIGUES, J. A. Assistência clínica materna e neonatal à diferentes condições obstétricas em bovinos da raça Holandesa. 2008. 94 f. Dissertação (Mestrado em Medicina Veterinária) - Faculdade de Medicina Veterinária, Universidade de São Paulo, São Paulo, 2008. SANTOS, T. M. A.; GILBERT, R. O.; BICALHO, R. C. Metagenomic analysis of the uterine bacterial microbiota in healthy and metritic postpartum dairy cows. Journal of dairy science, v. 94, n. 1, p. 291-302, 2011. SEEDY, F. R.; ABED, A. H.; YANNI, H. A.; RAHMAN, S. S. A. Prevalence of Salmonella and E. coli in neonatal diarrheic calves. Beni-Suef University Journal of Basic and Applied Sciences, v. 5, n. 1, p. 45-51, 2016. SILVA, C. P. C. Imunidade inata no sangue e secreção mamária de vac as holandesas no período periparto . São Paulo, 2014. 142 f. Dissertação (Mestrado em Clínica Veterinária) – Faculdade de Medicina Veterinária, Universidade de São Paulo, São Paulo, 2014. VAZIRI, N. D.; ZHAO, Y. Y.; PAHL, M. V. Altered intestinal microbial flora and impaired epithelial barrier structure and function in CKD: the nature, mechanisms, consequences and potential treatment. Nephrology Dialysis transplantation , p. 1-10, 2015. VLKOVÁ, E.; TROJANOVÁ, I.; RADA, V. Distribution of Bifidobacteria in the gastrointestinal tract of cows. Folia Microbiologica , v. 51, n. 4, p. 325-328, 2006. WANG, S.; ZHU, H.; LU, C.; KANG Z.; LUO, Y.; FENG, L.; LU, X. Fermented milk supplemented with probiotics and prebiotics can effectively alter the intestinal microbiota and immunity of host animals. Journal of Dairy Science , v. 95, n. 9, p. 4813-4822, 2012. WINDEYER, M. C.; LESLIE, K. E.; GODDEN, S. M.; HODGINS, D. C.; LISSEMORE, K. D.; LEBLANC, S. J. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age. Preventive Veterinary Medicine , v.113, n.2, p. 231-240, 2013.

91
CAPÍTULO 2 - INFLUÊNCIA DOS LEUCÓCITOS DO COLOSTRO NA RESPOSTA
IMUNE INATA DE BEZERRAS HOLANDESAS RECÉM-
NASCIDAS

92
8 INTRODUÇÃO
Os bezerros recém-nascidos nascem imaturos e agamaglobulinêmicos,
devido à impermeabilidade da placenta sineptéliocorial dos bovinos, cuja principal
função é a proteção do ambiente fetal contra microrganismos patogênicos. Desta
forma, os animais emergem do útero e iniciam a sua vida mediante à exposição aos
inúmeros microrganismos do meio ambiente ao qual está inserido (CHASE;
HURLEY; REBER, 2008).
A imunossupressão é outro evento associado ao estresse materno-fetal que
possui impacto no status imune dos recém-nascidos, devido ao aumento no cortisol
principalmente quando o parto é distócico (VANNUCCHI et al., 2015). O efeito do
cortisol sob o perfil leucocitário é classicamente relatado nos primeiros três dias de
vida, caracterizado por leucocitose, neutrofilia, monocitose, linfopenia e eosinopenia.
Assim, pode-se afirmar que bezerros recém-nascidos apresentam predomínio de
neutrófilos e baixos valores de linfócitos ao nascimento, observando-se inversão
deste perfil a partir do terceiro dia de vida (BENESI et al., 2012; NOVO et al., 2015).
O cortisol estimula a liberação de neutrófilos segmentados e bastonetes pela medula
óssea do compartimento estoque para a circulação (TORNQUIST; RIGAS, 2010).
O status imune do recém-nascido ao nascimento o torna dependente do
colostro materno. Falhas no gerenciamento do manejo do colostro na criação de
bezerras estão associadas à elevada prevalência (19,1%) e incidência (21,2%) de
diarreia neonatal causada por Escherichia coli, Rotavírus, Coronavirus e
Cryptosporidium parvum (WINDEYER et al., 2013; MEGANCK; HOFLACK;
OPSOMER, 2014). Além disso, animais que receberam colostro com baixos teores
de imunoglobulinas apresentaram altos índices de escore de diarreia que
perduraram por mais tempo, alta mortalidade e menor ganho de peso, quando
comparados à animais que receberam colostro com alto teor de imunoglobulinas
(NOCEK; BRAUND; WARNER,1984).
O colostro contém altas concentrações de nutrientes e hormônios que dão
suporte ao crescimento e maturação no desenvolvimento fisiológico do bezerro. O
colostro também contém fatores imunológicos importantes como citocinas,
anticorpos e uma grande quantidade de leucócitos maternos para a proteção do
neonato. O papel dos anticorpos colostrais na proteção do neonato tem sido bem

93
estabelecido, entretanto, o papel dos leucócitos maternos e citocinas do colostro
ainda não foi completamente estabelecido.
O colostro bovino contém entre 1x106 e 2,5x106 células somáticas/mL, dentre
as quais cerca de 32% são viáveis (MCDONALD; ANDERSON, 1981; LIEBLER-
TENORIO; RIEDEL-CASPARI; POHLENZ, 2002; GOMES et al., 2011; SILVA, 2014).
A população de leucócitos no colostro de primeira ordenha consiste em 13,3% de
neutrófilos, 16,4% de linfócitos, 69,5% de monócitos e células epiteliais e 0,27% de
eosinófilos (GOMES et al., 2011). Meganck et al. (2014) avaliaram a proporção dos
leucócitos colostrais e obtiveram 25,4% de linfócitos T, 2,9% de linfócitos B, e 32,7%
de macrófagos.
As células do colostro bovino são absorvidas pelo epitélio intestinal dos
recém-nascidos e migram para a placa de Peyer e linfonodos mesentéricos
(LIEBLER-TENORIO; RIDEL-CASPARI; POHLENZ, 2002). As células maternas
também foram encontradas na circulação sanguínea dos recém-nascidos,
detectando-se pico máximo de observação 24 horas após a ingestão do colostro.
Em seguida, as células maternas desapareceram da circulação sanguínea com 36
horas devido à da migração das mesmas para outros tecidos e órgãos linfoides
secundários com intuito de promover proteção ao recém-nascido (REBER et al.,
2006).
O perfil imune dos bezerros após a ingestão de colostro celular e acelular
parece ser diferenciado, de acordo com os resultados obtidos nas poucas pesquisas
realizadas nesta área. Além disso, a maioria dos dados refere-se à avaliação do
perfil imune específico e pouco se sabe sobre o padrão da resposta imune inata em
bezerros que ingeriram colostro fresco ou congelado.
Riedel-Caspari (1993) infectou 20 bezerras com 109 colônias de Escherichia
coli enteropatogênica, através de inoculação oral, três horas após o nascimento e
antes do fornecimento de colostro. Após a inoculação, as bezerras foram
alimentadas com colostro com e sem células. A autora observou que as células do
colostro podem desempenhar papel importante na manutenção de saúde do
bezerro, uma vez que as bezerras que receberam colostro com células
apresentaram maior vitalidade durante o período estudado. Os animais que
receberam colostro sem células apresentaram maior temperatura retal,
principalmente entre os dias 9 e 16, do que o COL+. Nitidamente estes animais
apresentaram maior carga de E. coli. Além disso, alguns animais do COL- se

94
recusaram a ingerir o substituto de leite que foi oferecido igualmente para ambos os
grupos, enquanto todos os animais do COL+ o ingeriram normalmente. O grupo
COL+ excretou menos E. coli durante a primeira semana de infecção e apresentou
pico de excreção depois do COL-. O COL+ atingiu o limite inferior de detecção de
bactérias mais cedo em relação ao COL-, que por sua vez apresentou outro pico de
detecção após o COL+ já ter cessado a liberação nas fezes. Os animais do COL-
atingiram o limite mínimo de detecção quatro dias após o COL+. Em relação às
concentrações específicas de anticorpos (IgA e IgM), os autores observaram que a
concentração foi maior no soro das bezerras COL+ durante todo o período
analisado. A autora conclui que leucócitos colostrais aumentaram a resistência dos
animais contra a infecção artificial desde o início, sinalizando para uma menor
resposta inflamatória do grupo, e que essas células contribuem para a imunidade
passiva e a resistência do recém-nascido, ressaltando que a qualidade e quantidade
de leucócitos são cruciais para a sua eficiência. Por outro lado, Langel et al. (2015)
não encontraram diferenças para os escores de diarreia em bezerras que receberam
colostro fresco e congelado.
Bezerros que mamaram colostro acelular apresentaram maior expressão dos
receptores CD11a, CD11c, CD43 e CD62L nas células mononucleares, em
comparação aos bezerros que receberam colostro celular, indicando maior
ocorrência de processos inflamatórios. O CD11a e CD43 são importantes receptores
ligados à migração celular da corrente sanguínea para o epitélio intestinal e o CD62L
está relacionado à sinalização dos leucócitos para que eles diminuam a velocidade
de circulação e fiquem aderidos à parede dos vasos sanguíneos facilitando a adesão
e migração (REBER et al., 2006). Em contrapartida, Stieler et al. (2012) não
observaram diferenças na proporção ou índice de fagocitose de neutrófilos e
macrófagos entre os bezerros que receberam colostro fresco e congelado.
A hipótese desta pesquisa é de que as células do colostro bovino influenciam
na resposta imune inata e consequentemente no perfil sanitário de bezerras
Holandesas durante o primeiro mês de vida. Assim, o objetivo desta pesquisa foi
avaliar se o fornecimento de colostro com (fresco) e sem células viáveis (congelado)
influencia na proporção e função dos neutrófilos sanguíneos e ocorrência de doença
durante o período neonatal.

95
9 MATERIAL E MÉTODOS
Esta pesquisa foi aprovada pelo Comitê de Ética no Uso de Animais da
Faculdade de Medicina Veterinária e Zootecnia da USP, protocolo nº2934/2013.
9.1 BEZERRAS
As bezerras desta pesquisa eram oriundas de fazenda comercial de alta
produção, localizada no município de Araras – São Paulo. Este experimento foi
conduzido de julho a outubro de 2014.
Os partos foram assistidos pelos funcionários e veterinários da propriedade
para auxílio obstétrico, separação das recém-nascidas e controle do manejo de
colostro (Figura 19).
A equipe de pesquisa era acionada assim que os funcionários detectavam o
início do trabalho de parto para o deslocamento de pessoal e preparo do material
para a colheita das amostras.
Figura 19 - Nascimento em baias de parição (a); colostragem em baias de transição
(b), localizadas na maternidade – São Paulo – 2016
(a) (b) Fonte: (COSTA, J. F. R., 2016).
Bezerras hígidas advindas de partos eutócicos foram selecionadas por meio
do exame clínico geral, segundo os procedimentos descritos por Dirksen et al.

96
(2008). Foram aferidas as frequências cardíaca (FC), respiratória (FR) e temperatura
corpórea (Quadro 5). Outros critérios foram usados para determinar a vitalidade das
bezerras, tais como: tônus muscular, irritabilidade reflexa e coloração das mucosas
oculares (RODRIGUES, 2008).
Quadro 5 - Intervalos de referência para as funções vitais em bezerras no primeiro mês de vida
Parâmetros Intervalos de Referência
1hora após nascimento 1ª semana 2ª semana 3ª semana 4ª semana
FC (bpm) 100,0-150,0 124,0±2,0 105,0±3,0 101,0±3,0 86,0±2,0
FR (mpm) 50,0-75,0 59,0±8,8 46,0±7,0 40,0±8,4 33,0±9,0
TEMP (°C) 39,0-39,5 38,6±0,3 39,0±1,0 39,0±0,4 39,0±0,3,0 Fonte: (SILVA et al., 2016, no prelo)4
Vinte bezerras hígidas foram triadas e distribuídas em dois grupos
denominados COL+ (n=10) e COL- (n=10). As bezerras do grupo COL+ receberam
colostro fresco com células proveniente de suas respectivas mães, enquanto os
animais COL- receberam colostro congelado sem células viáveis oriundo de vacas
doadoras.
Vacas e doadoras foram ordenhadas imediatamente após a parição em
tronco de contenção localizado no interior da maternidade. Inicialmente foi realizada
a lavagem com solução clorada e secagem dos tetos com papel toalha individual.
O colostro das doadoras foi armazenado pelos funcionários da fazenda em
garrafas plásticas de 2L, mantidas sob refrigeração (4 - 8°C), por 24 horas. Por fim,
as garrafas foram estocadas a -20°C durante o período mínimo de 24 horas e
máximo de 3 meses.
Inicialmente, para justificar a ausência de leucócitos viáveis nas amostras
congeladas, foi realizado um projeto piloto pela equipe onde alíquotas de colostro
fresco, refrigerado e congelado foram avaliadas quanto à sua viabilidade celular, de
hora em hora, durante as primeiras seis horas e, após, entre 18-24 horas. Com esse
estudo foi possível comprovar que as amostras de colostro que foram congeladas
4 SILVA, B. T.; HENKLEN, A.; MARQUES, R. S.; OLIVEIRA, P. ;L.; LEITE, S. B. P.; NOVO, S. M. F.; BACCILI, C. C.; REIS, J. F.; GOMES, V. Vital parameters of Holstein calves from birth to weaning. Revista Brasileira de Medicina Veterinária. [200-]. (no prelo).

97
não apresentaram viabilidade celular após o congelamento (0%) desde os
momentos iniciais (NOVO et al., 2014).
A avaliação indireta da concentração de Igs do colostro administrado para as
bezerras COL+ e COL- foi avaliada por meio de colostrômetro e refratômetro Brix
(23-32%). Foi possível determinar amplitudes de variação entre 80 e 120g/L de Igs
no colostro das vacas COL+; e 70 a 120g/L para as vacas COL-.
A quantidade de células somáticas (1,9 milhões/mL; log10: 6,28) encontrada
no colostro foi estimada a partir das amostras de colostro fresco utilizando-se como
metodologia a contagem microscópica direta adaptada por Gomes et al. (2011)
(Figura 20).
Figura 20 - Esfregaço da secreção mamária do colostro fresco fornecido para as bezerras do grupo com células viáveis (COL+) – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016). Legenda: (a) macrófagos presentes em predominância no colostro; (b) neutrófilo; (c) glóbulos de gordura.
A primeira garrafa de colostro foi descongelada lentamente a 37°C para
alimentar as bezerras COL- dentro das primeiras seis horas de vida. A segunda
alimentação foi realizada no intervalo máximo de doze horas de vida, utilizando-se
outra garrafa de colostro descongelado.
Bezerras COL+ foram alimentadas com dois litros de colostro fresco nas
primeiras seis horas de vida. O colostro para a segunda mamada foi mantido sob
refrigeração a 4°C, levemente aquecido a 37°C pouco antes da sua administração
entre seis a 12 horas pós-nascimento.
A cura do umbigo foi realizada pela imersão completa do cordão umbilical em
recipiente contendo tintura de iodo a 10% por pelo menos 1 minuto e meio,
(a)
(b)
(c)

98
repetindo-se o processo após 12 horas. A partir do segundo dia, foi utilizada tintura
de iodo a 5% duas vezes ao dia até mumificação do cordão umbilical.
As bezerras COL+ e COL- foram mantidas inicialmente nas baias dentro da
maternidade, local onde foram realizados os exames clínicos iniciais, colostragem e
cura do umbigo, sendo posteriormente transferidas para o bezerreiro às 6 horas de
vida, onde permaneceram em baias de madeira não suspensas até os primeiros três
primeiros dias pós-nascimento. Por fim, os animais foram transferidos e
permaneceram em gaiolas de metal individuais suspensas, medindo 1,2m x 1m, em
local com capacidade para 100 gaiolas, até o final desta pesquisa (Figura 21).
Figura 21 - Alojamento das bezerras durante o primeiro mês de vida – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
Legenda: (a) alojamento das bezerras recém-nascidas durante as primeiras horas de vida para exame clínico geral, colostragem e cura do umbigo; (b) baia de transição não suspensa onde ficaram até o terceiro dia de vida; (c) bezerreiro para onde as bezerras foram transferidas e permaneceram até o fim do experimento.
a (b) (a)
(c)

99
A partir deste momento, os animais foram alimentados em baldes, duas vezes
ao dia, sendo fornecido um total de seis litros de leite sem antibiótico proveniente do
rebanho, além do fornecimento de ração comercial5 e água ad libitum.
9.2 MOMENTOS DE AVALIAÇÃO
As bezerras foram avaliadas periodicamente logo após o nascimento (D0),
entre 24 a 48 horas de vida (D2) e nos dias 7, 14, 21 e 28 pós-nascimento (D7-D28).
9.3 AVALIAÇÃO CLÍNICA PERIÓDICA
Os animais foram avaliados clinicamente utilizando-se os escores de diarreia
e broncopneumonia realizado de acordo com o Calf Heath Scoring Criteria
previamente publicado na University of Wiscosin - Madison (McGUIRK, 2008).
O escore fecal foi categorizado em escores sendo 0- consistência normal, 1-
consistência pastosa e semi-formada, 2- consistência aquosa com maior quantidade
de água, com conteúdo fecal aderido no períneo e cauda, 3- líquida, conteúdo fecal
aderido no períneo e cauda. A diarreia foi considerada presente quando o escore
obtido foi 2 ou 3 (Quadro 6 e Figura 22).
Broncopneumonia foi estabelecida considerando-se os seguintes parâmetros:
temperatura retal, tosse, secreção nasal e ocular e posição das orelhas. Para cada
item foi atribuído o escore de zero a 3 de acordo com a severidade dos sintomas. A
soma dos sintomas acima de 5 foi utilizada para a detecção da Doença Respiratória
Bovina (Quadro 7).
A região umbilical foi avaliada por inspeção e palpação periódica da região
umbilical para a detecção de sinais inflamatórios.
As anemias foram avaliadas a partir do eritrograma das bezerras. O
hematócrito, hemoglobina ou hemácias abaixo dos valores de referência
5 Rumileite 20®, Guabi

100
estabelecidos por Brun-Hansen, Kampen e Lund (2006) foram utilizados para a
interpretação dos eritrogramas e determinação das anemias. A classificação das
anemias foi baseada nos índices hematimétricos (BRUN-HANSEN; KAMPEN; LUND,
2006).
Quadro 6 - Escores de fezes adotado para detecção da diarreia em bezerras no primeiro mês de vida
Parâmetros Escores
Consistência normal: firme, coloração amarronzada, períneo e cauda limpos e secos 0
Pastosa, semi-formada 1
Pastosa com maior quantidade de água: permanece sobre a “cama”; conteúdo fecal aderido no períneo e cauda
2
Líquida: aquosa, conteúdo fecal aderido no períneo e cauda 3 Fonte: (MCGUIRK, 2008).
Figura 22 - Escores de fezes adotado para detecção da diarreia em bezerras no primeiro mês de vida – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).

101
Quadro 7 - Escores de broncopneumonia adotado para detecção da doença em bezerras no primeiro mês de vida
Escores Temperatura retal ( OC) Tosse Secreção nasal Secreção
ocular Posicionamento das
orelhas 0 37,7 - 38,2 Ausente Serosa Serosa Normal
1 38,3 - 38,8 Presente e única,
quando estimulada
Pouca quantidade,
unilateral
Pouca quantidade
Balançar das orelhas ou cabeça
2 38,9 - 39,3
Presente e repetida quando estimulada, ou
ocasional quando espontânea
Excessiva, mucosa e bilateral
Moderada quantidade
bilateral
Ligeiramente pendente, unilateral
3 ≥ 39,4 Presente, repetida
e espontânea
Abundante, mucopurulenta,
bilateral
Intensa quantidade,
bilateral
Pendentes intensamente, bilateral ou torção da cabeça
Fonte: (POULSEN; MCGUIRK, 2009).
9.4 AMOSTRAS
Foram obtidas amostras de sangue por meio de punção da veia jugular,
utilizando sistema a vácuo, em tubos siliconados sem e com os anticoagulantes
EDTA (Ethylene Diamine Tetraacetic Acid) (Anexo G) e heparina de sódio.
9.5 ANÁLISES
A partir das amostras sanguíneas foram realizadas dosagens para avaliar o
perfil inflamatório das bezerras.
9.5.1 Hemograma
Os componentes hematológicos (hemácias, Red Blood Cells-RBC;
hemoglobina, hematócrito, Volume Corpuscular Médio - VCM, Hemoglobina
Corpuscular Média - HCM, Concentração de Hemoglobina Corpuscular Média - HCM
e leucócitos totais) foram obtidos por sistema automático6. A contagem diferencial de
6 BC 2800Vet®

102
leucócitos foi realizada observando-se as características morfológicas das células
por microscopia óptica, aumento de 1.000x.
9.5.2 Cortisol
As determinações hormonais de cortisol foram realizadas por ensaio
quimiluminescente, utilizando-se kit comercial 7 em analisador de imunoensaios
Immulite 1000® (Siemens). Para a medição quantitativa do cortisol foi utilizado um
imunoensaio competitivo de fase sólida, de enzima químico - luminosas, com ciclo
de incubação de 1x30 minutos. O procedimento de ensaio segue o princípio básico
de enzima imunoensaio onde existe uma competição entre um antígeno não
marcado e um antígeno marcado com enzima, por um número determinado de sítios
de ligação no anticorpo. A quantidade de antígeno marcado com enzima é
inversamente proporcional à concentração do analítico presente não marcado. O
material não ligado é removido por decantação e lavagem das cavidades. Com os
níveis de refração da luz, o equipamento mensura a quantidade de cortisol presente
na amostra.
9.5.3 Haptoglobina
A concentração de haptoglobina sérica foi avaliada com base na sua
habilidade de ligação com a hemoglobina. Uma solução-padrão purificado de
haptoglobina de 100 a 1,5 mg/dL, foi utilizado para a preparação da curva padrão.
As amostras de soro controle (10µL) foram diluídas em solução salina (90µL) e foi
adicionado 50µL de solução de meta-hemoglobina bovina (30mg/dL). A reação foi
incubada durante dez minutos à temperatura ambiente, no escuro e em seguida
foram adicionados 150µL de substrato de guaiacol e 50 µL de peróxido de
hidrogênio. A placa ficou incubada por mais dez minutos no escuro e a leitura da
7 Cortisol Siemens® Ref: LKCO1, Lot: 0403

103
absorbância através da densidade óptica foi realizada no leitor de microplacas com
comprimento de onda de 450-500 nm.
9.5.4 Ferro
Alíquotas de soro foram descongeladas em geladeira (overnight),
homogeneizadas em homogeneizador automático para posterior realização dos
testes bioquímicos em analisador bioquímico automático5. Para a dosagem da
concentração de ferro foi utilizado kit comercial6 (Ferro UIBC, Randox®),
empregando-se a metodologia descrita pelo fabricante.
9.5.5 Imunofenotipagem dos Neutrófilos
As subpopulações e ativação dos leucócitos sanguíneos foram avaliadas
utilizando-se a técnica de citometria de fluxo. Inicialmente, alíquotas do sangue total
obtidas em tubos com EDTA (100µL) foram distribuídas em tubos de citometria de
fluxo. Os eritrócitos foram lisados pela adição de 900µL de solução comercial8
(Facslysing), e os tubos foram mantidos em temperatura ambiente por 15 minutos.
Inicialmente, dois anticorpos monoclonais primários: mouse anti-bovine (Washington
State University) foram adicionados para a marcação celular na mesma diluição
baseados nas titulações. Desta forma, tanto CH138 (IgM - CH138A), quanto CD62-L
(IgG1 - CC32) foram diluídos em 1:400.
As amostras foram incubadas por 40 minutos a 4°C para que houvesse
marcação celular e em seguida, foram submetidas a três lavagens com solução de
Phosphate Buffered Saline (PBS) (Anexo H). Posteriormente, foram adicionados
anticorpos monoclonais secundários para emissão das fluorescências identificadas
pelo citômetro de fluxo 9 . O fluorocromo APC (Allophycocyanin) (INVITROGEN,
U.S.A.) foi utilizado como anticorpo secundário para o CH138 (APC – M31505) e o
fluorocromo PE (Phycoerythrin) (BENCTON DICKINSON, U.S.A.) foi utilizado como
8 BD nº catálogo: 349202 9 FACSCalibur (Becton Dickinson Immunocytometry System®, San Diego, CA)

104
anticorpo secundário para o CD62-L (PE – P-21129). Os anticorpos secundários
foram previamente diluídos em 1:200, adicionados às células, incubados por 30
minutos refrigeradas e mantidos em ambiente escuro.
As amostras foram então submetidas a três lavagens com PBS e
ressuspendidas em 300µL e reservadas para avaliação das fluorescências emitidas
nos comprimentos de onda 650-660nm (APC) e 480nm (PE) em citômetro de fluxo.
Os eventos foram adquiridos no citômetro utilizando o software CellQuest,
selecionando os granulócitos a partir do seu tamanho - forward scatter (FSC) e
granulosidade - side scatter (SSC). Para isso, foram adquiridas dez mil células para
os neutrófilos no gate correspondente aos granulócitos.
As células foram avaliadas quanto à fluorescência emitida pelos anticorpos
secundários utilizando-se o software FlowJo® (Tree Star, EUA), versão 7.2.5. Os
resultados foram expressos em porcentagem do gate de células positivas para cada
marcador e os histogramas foram estabelecidos para avaliar a proporção de células
em cada fluorescência. Os dot plot (gráficos emitidos a partir do citômetro) foram
interpretados a partir das cores para estabelecer as populações celulares (Figuras
23 a 26).
Figura 23 - Avaliação da proporção de granulócitos no sangue de bezerras Holandesas no primeiro mês de vida – São Paulo – 2016
(a) (b)
Fonte: (COSTA, J. F. R., 2016) Legenda: (a) Distribuição dos leucócitos sanguíneos; (b) seleção da população de granulócitos em relação ao total de eventos adquiridos
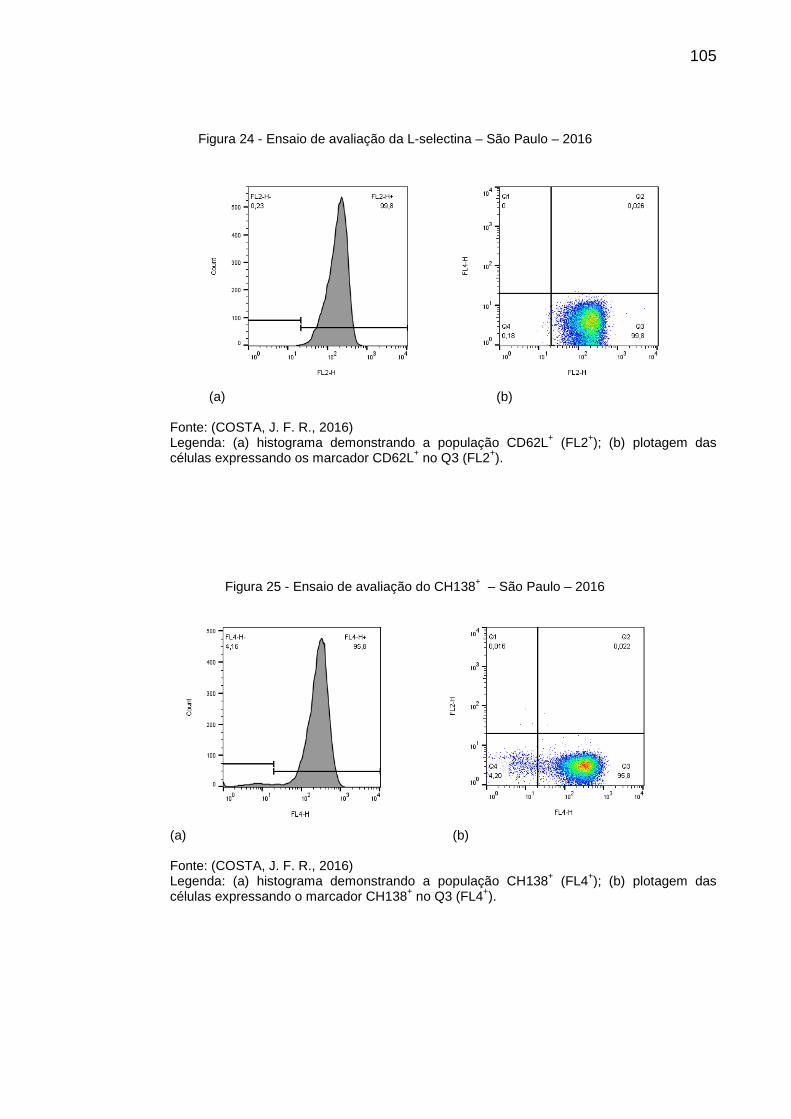
105
Figura 24 - Ensaio de avaliação da L-selectina – São Paulo – 2016
(a) (b)
Fonte: (COSTA, J. F. R., 2016) Legenda: (a) histograma demonstrando a população CD62L+ (FL2+); (b) plotagem das células expressando os marcador CD62L+ no Q3 (FL2+).
Figura 25 - Ensaio de avaliação do CH138+ – São Paulo – 2016
(a) (b)
Fonte: (COSTA, J. F. R., 2016) Legenda: (a) histograma demonstrando a população CH138+ (FL4+); (b) plotagem das células expressando o marcador CH138+ no Q3 (FL4+).

106
Figura 26 - Ensaio de avaliação do duplo positivo para CD62L+ e CH138+ – São Paulo – 2016
(a) (b) (c)
Fonte: (COSTA, J. F. R., 2016) Legenda: (a) histograma demonstrando a população CD62L+ (FL2+); (b) histograma demonstrando a população CH138+ (FL4+); (c) plotagem das células expressando os marcadores CD62L+CH138+ no quadrante duplo positivo (circulado).
9.5.6 Fagocitose e Produção Peróxido de Hidrogênio
Para realização desta prova, foram utilizadas Staphylococcus aureus e
Escherichia coli isoladas do leite de vaca com mastite, gentilmente cedidas pelo
Prof. Dr. Nilson Roberti Benites. As bactérias foram marcadas com Iodeto de
Propídeo segundo procedimento descrito por Silva (2014).
Para a realização dos ensaios (Quadro 8), foram adicionados 100µL de
sangue heparinizado contendo aproximadamente 2x105 células em tubos de
citometria de fluxo. Em seguida, foram adicionados 200µL da solução de uso do
DCFH-DA10 (Anexo I), 50µL da bactéria marcada, ajustando-se o volume final da
mistura para 1100µL, utilizando PBS suplementado com Soro Fetal Bovino (SFB) a
10%. A mistura foi incubada em estufa a 37°C por 30 minutos, mantida em
suspensão com o uso de homogeneizador. Em seguida, foram adicionados 2mL de
EDTA 3mM (Anexo I) gelado, nos ensaios onde as bactérias foram acrescidas. Logo
após, foi realizada a lise das hemácias para lavagem das células e retirada do
excesso de Iodeto de Propídeo.
10 Sigma® Aldrich, St. Louis, EUA, nº cat. D6883.

107
Para a lise das hemácias foi adicionado 1mL de água destilada ao pellet,
homogeneizado no agitador de tubos por 15 segundos, e imediatamente foi
acrescido 1mL de PBS 2x concentrado (Anexo J). Imediatamente em seguida, foi
adicionado 3mL de PBS gelado. Os tubos foram centrifugados por 5 minutos,
783xG, 4ºC. O sobrenadante foi desprezado e o pellet e ressuspendido em 1mL de
PBS. O ensaio foi centrifugado novamente nas mesmas condições (5 minutos,
783xG, 4ºC) e o pellet foi ressuspendido em 300µl de PBS para realização da leitura
no citômetro de fluxo 11 , onde foram adquiridos 20.000 eventos utilizando-se o
software CellQuest®.
A análise das fluorescências emitidas foi realizada utilizando-se software
FlowJo®. Inicialmente, a população de granulócitos foi selecionada no ensaio
contendo células sem anticorpos (branco) e copiada para os demais ensaios. Em
seguida, a produção de H2O2 basal e determinação do limite para a fluorescência
FL1 foi avaliada nos tubos onde foram adicionados células e o substrato DCFH. Os
próximos tubos da análise eram os que continham apenas as células e bactérias
Staphylococcus aureus e Escherichia coli para a determinação do limite da
fluorescência FL2. Por fim, após terem sido estabelecidos os cortes a partir dos
controles, contendo células não marcadas e marcações simples, realizou-se às
análises dos ensaios que continham DCFH-DA e bactérias. Neste tubo, foram
analisados tanto a fluorescência FL1 e FL2 para a determinação da fagocitose e
produção de peróxido de hidrogênio na presença dos estímulos bacterianos (Figuras
27 a 31).
Quadro 8 - Ensaio de fagocitose e produção de H2O2 do sangue de bezerras Holandesas
Tubo Fluorescência Sangue DCFH BACT PBS SFB 10% Evento
A Branco S - - S Branco
B FL1 S S - S Burst Basal
C FL2 S - Sapi S Fagocitose
D FL2 S - E.coli S Fagocitose
E Ensaio 1 S S Sapi S Fagocitose/Burst
F Ensaio 2 S S E.coli S Fagocitose/Burst
Fonte: (COSTA, J. F. R., 2016). Legenda: Bact – Bactéria; S – Acrescido no tubo.
11
FACSCalibur Flow Cytometer (Becton Dickinson Immunocytometry System®), San Diego, CA

108
Figura 27 - Avaliação da proporção de PMN no sangue de bezerras Holandesas – São Paulo – 2016
(a)
(a) (b)
Fonte: (COSTA, J. F. R., 2016) Legenda: (a) Distribuição dos leucócitos sanguíneos; (b) Seleção dos PMN em relação ao total de eventos adquiridos.
Figura 28 - Avaliação da produção de H2O2 basal, sem estímulos bacterianos, pelas células PMN com DCFH-DA. Seleção do eixo FL1+ para avaliação da produção de H2O2 pelos PMN sem estímulo bacteriano – São Paulo – 2016
(a) (b)
Fonte: (COSTA, J. F. R., 2016) Legenda: (a) histograma demonstrando a produção de H2O2 basal (FL1+); (b) plotagem das células produzindo H2O2 basal (FL1+).

109
Figura 29 - Avaliação da fagocitose das células PMN do sangue de bezerras Holandesas que fagocitaram Staphylococcus aureus e Escherichia coli – São Paulo – 2016
(a) (b)
(c) (d)
Fonte: (COSTA, J. F. R., 2016) Legenda: (a) Histograma demonstrando a fagocitose dos PMN estimulados com Staphylococcus aureus (FL2+); (b) plotagem das células fagocitando Staphylococcus aureus (FL2+); (c) Histograma demonstrando a fagocitose dos PMN estimulados com Escherichia coli (FL2+); (d) plotagem das células fagocitando Escherichia coli (FL2+).

110
Figura 30 - Avaliação da produção de H2O2 e fagocitose pelas células PMN do sangue de bezerras Holandesas com DCFH-DA e Staphylococcus aureus – São Paulo – 2016
(a) (b)
(c) (d)
Fonte: (COSTA, J. F. R., 2016) Legenda: (a) Histograma para avaliação da produção de H2O2 pelas células PMN (FL1+); (b) plotagem das células para avaliação da produção de H2O2 pelas células PMN (FL1+); (c) histograma para avaliação da fagocitose pelas células PMN estimuladas com S. aureus (FL2+); (d) plotagem para avaliação da fagocitose pelas células PMN estimuladas com S. aureus (FL2+).

111
Figura 31 - Avaliação da produção de H2O2 e fagocitose pelas células PMN do sangue de bezerras Holandesas com DCFH-DA e Escherichia coli – São Paulo – 2016
(a) (b)
(c) (d)
Fonte: (COSTA, J. F. R., 2016)
Legenda: (a) Histograma para avaliação da produção de H2O2 pelas células PMN (FL1+); (b) plotagem das células para avaliação da produção de H2O2 pelas células PMN (FL1+); (c) histograma para avaliação da fagocitose pelas células PMN estimuladas com E. coli (FL2+); (d) plotagem para avaliação da fagocitose pelas células PMN estimuladas com E. coli (FL2+).

112
9.6 ANÁLISE ESTATÍSTICA
Os dados foram armazenados em planilhas (Microsoft Excel® 2010), e
posteriormente analisados com o software estatístico SPSS 19.0 (IBM Corp.
Released 2011. IBM SPSS Statistics for Windows, Version 19.0. Armonk, NY: IBM
Corp.).
Para que fosse possível uma análise paramétrica, a transformação
logarítmica foi utilizada. Sendo assim, o Teste t de Student para amostras
independentes foi realizado para comparação entre os grupos (COL+ e COL-) e a
análise de variância (ANOVA) realizada para comparação entre os tempos (D0 a
D28) dentro do mesmo grupo. No caso onde ocorreram as transformações
logarítmicas, o p apresentado nas tabelas diz respeito aos testes relacionados com
os valores em log10, enquanto os valores descritos são reais.
Todas as análises foram consideradas estatisticamente significativas quando
p<0,05 e tendência quando p>0,05 e p<0,1. Análises descritivas foram apresentadas
pela média e desvio padrão.

113
10 RESULTADOS
Este trabalho avaliou a influência das células do colostro na resposta imune
inata e saúde de bezerras Holandesas durante o primeiro mês de vida.
10.1 STATUS SANITÁRIO
A diarreia teve início no D2 e a frequência aumentou do D7 (COL+ = 56%;
COL-= 70%) ao D14 (COL+ = 78%; COL-= 60%). Após estes momentos, as
bezerras apresentaram uma ligeira diminuição da frequência no D21 COL+ = 56%;
COL-= 50%) e D28 (COL+ = 44%; COL-= 50%) (Figura 32).
A região umbilical externa foi avaliada através da inspeção e palpação
periódica para a detecção de sinais inflamatórios. Desta forma, foi possível detectar
que três bezerras do grupo COL- apresentaram inflamação umbilical no D14 (n=2) e
D28 (n=1). Além disso, também foram detectadas duas bezerras com
broncopneumonia em ambos os grupos: COL+ no D14 (n=1) e COL- no D21 (n=1).
Apesar da maior frequência de doenças encontradas no grupo COL-, não foi
possível comprovar diferenças estatísticas entre a frequência de diarreia ou doenças
totais usando o teste Qui-Quadrado.
A interpretação individual do eritrograma de cada bezerra mostrou anemia do
D2 até o D28 de acordo com os valores de referência estabelecidos por Brun-
Hansen; Kampen; Lund (2006) (Figura 33).
As frequências de anemias observadas foram de 20 e 30% no D2, 10 e 10%
no D7, 20 e 50% no D14, 0 e 40% no D21 e 0 e 50% no D28 para bezerras do COL+
e COL-, respectivamente. As bezerras do COL- apresentaram maior frequência de
anemia no D21 (COL+= 0%; COL-= 40%; P=0,03) e D28 (COL+= 0%; COL-= 50%;
P=0,02).
Durante todo o experimento foram observados 23 episódios de anemia. A
anemia predominantemente de acordo com a morfologia dos eritrócitos foi a
normocítica hipocrômica. As frequências de anemias microcítica, macrocítica e
normocítica foram 21,7% (5/23), 56,52% (13/23) e (21,7%) 5/23, respectivamente.

114
Anemia normocítica foi observada em 11/23 bezerras (47,8%) e 12/23 (52,17%),
respectivamente. Anemia normocítica foi observada em 11/23 (47,8%) e 12/23
(52,17%) dos casos.
Figura 32 - Frequência do escore fecal e incidência de diarreia em bezerras Holandesas (COL+) e (COL-) durante o primeiro mês de vida – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016).
0
20
40
60
80
100
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D0
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D14
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D2
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D21
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D7
COL+
COL-
0
20
40
60
0 1 2 3
Fre
quên
cia
(%)
Escore Fecal
D28
COL+
COL-

115
Figura 33 - Frequência de anemias em bezerras Holandesas (COL+) e (COL-) durante o primeiro mês de vida - São Paulo - 2016
Fonte: (COSTA, J. F. R., 2016). Legenda: S.A.: sem alteração; An No Hipo: anemia normocítica hipocrômica; An Mac Hipo: anemia macrocítica hipocrômica; An No No: anemia normocítica normocrômica; An Mi No: anemia microcítica normocrômica; An Mi Hipo: anemia microcítica hipocrômica.
10.2 CORTISOL
O estresse ao qual as bezerras foram submetidas nos primeiros dias de vida
foi mensurado através da dosagem do cortisol, observando-se perfil semelhante os
0
20
40
60
80
100
S. A. An NoHipo
AnMacHipo
An NoNo
An MiNo
An MiHipo
Fre
qu
ênci
a (%
)
D0
COL+
COL-
0
20
40
60
80
100
S. A. An NoHipo
AnMacHipo
An NoNo
An MiNo
An MiHipo
Fre
qu
ênci
a (%
)
D14
COL+
COL-
0
20
40
60
80
100
S. A. An NoHipo
AnMacHipo
An NoNo
An MiNo
An MiHipo
Fre
qu
ênci
a (%
)
D2
COL+
COL-
0
20
40
60
80
100
S. A. An NoHipo
AnMacHipo
An NoNo
An MiNo
An MiHipo
Fre
qu
ênci
a (%
)
D21
COL+
COL-
0
20
40
60
80
100
S. A. An NoHipo
AnMacHipo
An NoNo
An MiNo
An MiHipo
Fre
qu
ênci
a (%
)
D7
COL+
COL-
0
20
40
60
80
100
S. A. An NoHipo
AnMacHipo
An NoNo
An MiNo
An MiHipo
Fre
qu
ênci
a (%
)
D28
COL+
COL-

116
grupos COL + e COL-. O pico máximo de cortisol foi observado no D0 (COL+: 12,27
µg/dL; COL-: 11,53 µg/dL), em seguida, os valores apresentaram um decréscimo
gradual até o D28 (COL+: 0,81µg/dL; COL-: 0,35 µg/dL). Apesar de não terem sido
detectadas diferenças estatísticas entre os grupos, foi possível observar diferença
entre os momentos D0 e D21 no grupo COL+, onde p = 0,048 (Tabela 7 e Figura
34).
Tabela 7 - Concentração de cortisol sérico (µg/dL) em bezerras Holandesas que receberam
colostro com (COL+) e sem células viáveis (COL-), durante o primeiro mês de vida - São Paulo - 2016
Fonte: (COSTA, J. F. R., 2016) Legenda: letras minúsculas na mesma coluna demonstram diferença entre os tempos, sendo (a) valores diferentes do D0; (b) valores diferentes do D2; (c) valores diferentes do D7; (d) valores diferentes do D14; (e) valores diferentes do D21; (f) valores diferentes do D28, quando p<0,05; tendência quando P≤0,10
Figura 34 - Concentração de cortisol sérico (µg/dL) em bezerras Holandesas que receberam colostro com (COL+) e sem células viáveis (COL-), durante o primeiro mês de vida - São Paulo - 2016
Fonte: (COSTA, J. F. R., 2016).
Momentos (dias) D0 D2 D7 D14 D21 D28
Cortisol (µg/dL)
COL+ 12,27
±2,23 e 6,05 ±3,43
3,16 ±1,68
1,70 ±1,49
0,95 ±0,73 a
0,81 ±0,80
COL- 11,53 ±2,38
4,81 ±2,59
3,45 ±1,12
1,09 ±0,95
0,71 ±1,01
0,35 ±0,75
p 0,479 0,150 0,886 0,163 0,483 0,359
0
2
4
6
8
10
12
14
0 7 14 21 28
Co
rtis
ol(
µg/
dL
)
Dias
Col+
Col-

117
10.3 HAPTOGLOBINA
As concentrações e densidades ópticas obtidas para a haptoglobina sérica
durante o primeiro mês de vida estão dispostas na tabela 8 e figura 35. Os valores
obtidos para a concentração de haptoglobina foram crescentes do D0 (COL+: 1,24;
COL-: 1,34 mg/dL), atingindo pico máximo no D7 (COL+: 5,38; COL-: 5,27 mg/dL) e
D14 (COL-: 5,70 mg/dL), observando-se decréscimo dos valores nos momentos
subsequentes (COL+: 1,67-2,24; COL-: 1,31-1,71 mg/dL). Apesar destas variações
observadas, não foram detectadas diferenças estatísticas entre os grupos e
momentos para a concentração e densidade óptica da haptoglobina.
Tabela 8 - Valores da concentração (mg/dL) e densidade óptica (DO) para a haptoglobina sérica em bezerras Holandesas que receberam colostro com (COL+) e sem células viáveis (COL-), durante o primeiro mês de vida - São Paulo - 2016
Fonte: (COSTA, J. F. R., 2016). Legenda: † indica tendência entre os grupos naquele momento (p>0,05 e < 0,1).
Momentos (dias) D0 D2 D7 D14 D21 D28
Hp (mg/dL)
COL+ 1,24 ±0,41
1,28 ±0,50
5,38 ±9,17
1,67 ±0,99
2,24 ±3,24
1,80 ±1,93
COL- 1,34 ±0,59
1,56 ±0,49
5,27 ±6,18
5,70 ±12,28
1,31 ±0,45
1,71 ±0,91
P 0,802 0,873 0,682 0,454 0,686 0,365
Hp (DO)
COL+ 0,37 ±0,03
0,37 ±0,03
0,48 ±0,22
0,41 ±0,08
0,39 ±0,13
0,38 ±0,09
COL- 0,35 ±0,02
0,35 ±0,01
0,44 ±0,18
0,45 ±0,18
0,38 ±0,05
0,34 ±0,03
P 0,178 0,083 † 0,620 0,589 0,828 0,170

118
Figura 35 - Haptoglobina sérica em bezerras Holandesas recém-nascidas que receberam colostro com (COL+) e sem células viáveis (COL-) – São Paulo – 2016
(a) (b)
Fonte: (COSTA, J. F. R., 2016). Legenda: (a) concentração da haptoglobina em mg/dL; (b) valores da densidade óptica da haptoglobina; † indica tendência entre os grupos naquele momento (p>0,05 e < 0,1).
10.4 FERRO
A concentração de ferro sérico obtido para bezerras recém-nascidas estão
dispostos na tabela 9 e figura 36.
O pico máximo de ferro observado para ambos os grupos foi no D0 (COL+ =
21,76µM/L e COL- =15,41µM/L). A partir do D7, o grupo COL- (5,61 µM/L)
apresentou menor concentração de ferro sérico quando comparado ao COL+
(10,74 µM/L). Este perfil permaneceu até o D28. Foi possível observar diferença
estatística entre os grupos no D7 (p = 0,031) e diferença entre os momentos D0 e
D7 no grupo COL-.
Tabela 9 - Valores da concentração de ferro sérico (µM/L) em bezerras Holandesas que receberam colostro com (COL+) e sem células (COL-), durante o primeiro mês de vida - São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016) Legenda: letras minúsculas na mesma coluna demonstram diferença entre os tempos, sendo (a) valores diferentes do D0; (b) valores diferentes do D2; (c) valores diferentes do D7; (d) valores diferentes do D14; (e) valores diferentes do D21; (f) valores diferentes do D28, quando p<0,05; tendência quando P≤0,10.
Momentos (dias) D0 D2 D7 D14 D21 D28
Ferro (µM/L)
COL+ 21,67 ±9,14
8,98 ±2,68
10,74 ±6,21
13,26 ±10,68
9,73 ±6,32
11,15 ±10,48
COL- 15,41 ±9,81c
10,08 ±4,20
5,61 ±2,75 a
9,60 ±6,94
7,30 ±3,15
10,67 ±6,55
P 0,144 0,698 0,031 * 0,404 0,525 0,689
0,30
0,35
0,40
0,45
0,50
0 7 14 21 28
Hap
togl
ob
ina
(Den
sid
ade
Óp
tica
)
Dias
COL+
COL-
†
0
1
2
3
4
5
6
7
0 7 14 21 28
Hap
togl
ob
ina
(mg/
dL
)
Dias
Col+
Col-

119
Figura 36 - Valores de concentração sérica de ferro (µM/L) em bezerras Holandesas recém-nascidas que receberam colostro com (COL+) e sem células (COL-) - São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016). Legenda: * indica diferença entre os grupos naquele momento (p>0,05).
10.5 LEUCÓCITOS TOTAIS E NEUTRÓFILOS SANGUÍNEOS
Os valores médios e desvio padrão absolutos e relativos para leucócitos
totais (White Blood Cells-WBC) e neutrófilos (NEUTR) estão apresentados na
tabela 10 e figura 37.
As bezerras de ambos os grupos apresentaram pico máximo do WBC no D7
(COL+= 12,37x103/µL e COL- = 11,75 x103/µL), observando-se diminuição dos
valores nos momentos subsequentes até D21 (COL+= 7,55x103/µL e COL- =
9,61x103/µL). No D28 detectou-se o menor valor médio de WBC para o COL-
(6,91x103/µL), em contrapartida, bezerras COL+ apresentaram aumento no número
dessas células (9,42x103/µL).
Os valores de neutrófilos decresceram do pico que ocorreu no D7
(8,47x103/µL) até o D21 (3,07x103/µL) no grupo COL+. O grupo COL- apresentou
perfil diferente, observando decréscimo gradual dos valores desde o D0
(7,81x103/µL) até o D28 (2,54 x103/µL).
A comparação entre os grupos COL+ e COL- para WBC e NEUTR não
possibilitou a detecção de diferenças entre os grupos, porém foi possível observar
tendência na proporção de neutrófilos no momento D7 (P= 0,073).
0
5
10
15
20
25
0 7 14 21 28
Fer
ro (
uM
/L)
Dia
Col+
Col-
*

120
As diferenças obtidas para as comparações múltiplas entre os momentos
pode ser visualizada na tabela 10.
Tabela 10 - Médias e desvios-padrão para os leucócitos totais (WBC) e neutrófilos (NEUTR) do sangue de bezerras Holandesas que receberam colostro com (COL+) e sem células viáveis (COL-) - São Paulo - 2016
Fonte: (COSTA, J. F. R., 2016) Legenda: letras minúsculas na mesma coluna demonstram diferença entre os tempos, sendo (a) valores diferentes do D0; (b) valores diferentes do D2; (c) valores diferentes do D7; (d) valores diferentes do D14; (e) valores diferentes do D21; (f) valores diferentes do D28, quando p<0,05; tendência quando P≤0,10.
Momentos (dias) D0 D2 D7 D14 D21 D28
WBC (103/µL)
COL+ 10,89 ±2,72
10,19 ±5,88
12,37 ±5,23
8,03 ±2,75
7,55 ±3,39
9,42 ±3,92
COL- 11,41 ±4,23 f
9,61 ±4,23
11,75 ±5,81
9,52 ±4,20 9,61±3,18
6,91 ±1,65 a
P 0,772 0,827 0,802 0,336 0,275 0,134
NEUTR (103/µL)
COL+ 7,96
±2,34 def 7,60 ±5,73
8,47 ±5,06
3,38 ±1,92 a
3,07 ±2,87 a
3,93 ±2,30 a
COL- 7,81
±3,65 f 6,61 ±3,63
6,28 ±4,13
5,03 ±3,30
4,61 ±2,72
2,54 ±1,10 a
P 0,930 0,690 0,349 0,196 0,325 0,167
NEUTR (%)
COL+ 72,70
±9,83 def 66,30
±21,07 e 63,50
±14,28 def 39,90
±13,11 ac 36,00
±15,29 abc 39,60
±11,54 ac
COL- 67,20 ±9,31 ef
64,00 ±13,94 ef
49,60 ±12,64
49,30 ±11,92
44,30 ±14,92 ab
35,30 ±12,18 ab
P 0,239 0,783 0,073 † 0,143 0,296 0,455

121
Figura 37 - Leucócitos totais (WBC) e neutrófilos (NEUTR) do sangue de bezerras Holandesas recém-nascidas que receberam colostro com (COL+) e sem células (COL-) - São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016) Legenda: † indica tendência entre os grupos naquele momento (p>0,05 e < 0,1)
10.6 EXPRESSÃO DE MARCADORES NOS GRANULÓCITOS
As proporções de células e marcadores dos granulócitos estão dispostos na
tabela 11 e figura 38.
A proporção de PMN no COL+ aumentou levemente do D0 (40,25%) ao D7
(45,83%), em seguida, os valores diminuíram (27,46-34,44%). O COL- apresentou
média mínima no D0 (34,06%), pico máximo no D2 (53,25%), leve redução entre
D7 ao D21 (41,79-45,14%) seguida por redução marcante no D28 (36,76%). Foi
possível detectar diferença estatística entre os grupos no D21 (p=0,036) e
tendência no D14 (p=0,056). Não foi possível observar diferença estatística entre os
0
2
4
6
8
10
12
14
0 7 14 21 28
WB
C (
10
3/µ
L)
Dias
Col+
Col-
0
2
4
6
8
10
0 7 14 21 28
NE
UT
R (
10
3/µ
L)
Dias
Col+
Col-
0
10
20
30
40
50
60
70
80
0 7 14 21 28
NE
UT
R (
%)
Dias
Col+
Col-
†

122
momentos.
O perfil apresentado pelos grupos experimentais em relação à proporção de
granulócitos CH138+ expressando ou não o receptor CD62L+ foi diferente apesar da
ausência de diferenças estatísticas (P=0,095).
Foi observada elevada proporção de granulócitos CH138+ expressando L-
selectina em ambos os grupos no D0 (COL+: 87,73%; COL-: 88,51%). A partir
deste momento, os animais apresentaram perfil diferenciado, observando-se
aumento gradual das proporções destas células no COL- (45,14%) e queda no
COL+ (80,65%) até D14. Após, o COL- apresentou queda brusca (78,70%) e
aumento subsequente no D28 (98,88%), enquanto o COL+ apresentou aumento
(89,96%) no D21 e ligeira diminuição no D28 (85,78%).
Tabela 11 - Médias e desvios-padrão para as proporções de granulócitos CH138+ expressando o
receptor CD62-L em bezerras Holandesas que receberam colostro com (COL+) e sem células viáveis (COL-), no primeiro mês de vida - São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016) Legenda: * indica diferença entre os grupos naquele momento (p>0,05); † indica tendência entre os grupos naquele momento (p>0,05 e < 0,1)
Momentos (dias) D0 D2 D7 D14 D21 D28
PMN
(%)
COL+ 40,25 ± 20,57
41,39 ±17,02
45,83 ±19,47
34,44 ±13,27
27,46 ±11,28
33,92 ±12,94
COL- 34,06 ±16,4
53,25 ±21,55
42,79 ±16,23
45,14 ±10,04
41,79 ±13,64
36,76 ±15,27
P 0,530 0,308 0,819 0,056 † 0,036 * 0,851
CH138+CD62L+
(%)
COL+ 87,73
± 30,70 87,71 ±16,67
85,94 ±24,65
80,65 ±29,67
89,96 ±14,78
85,78 ±32,31
COL- 88,51 ±30,58
91,34 ±11,21
95,19 ±5,41
97,03 ±1,91
78,70 ±43,99
98,88 ±1,72
P 0,839 0,539 0,290 0,303 0,401 0,332
CH138+CD62L-
(%)
COL+ 0,68 ±0,84
2,61 ±5,94
2,03 ±6,28
2,40 ±5,00
0,00 ±0,01
0,02 ±0,06
COL- 9,93 ±9,51
3,03 ±6,99
0,44 ±0,61
0,09 ±0,16
0,47 ±0,97
0,02 ±0,06
P 0,295 0,819 0,859 0,095 † 0,512 0,854

123
Figura 38 - Proporção de granulócitos CH138+ expressando o receptor CD62L em bezerras Holandesas que receberam colostro com (COL+) e sem células viáveis (COL-), no primeiro mês de vida - São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016) Legenda: * indica diferença entre os grupos e † indica tendência entre os grupos naquele momento (p>0,05 e < 0,1)
10.7 FUNÇÃO DOS NEUTRÓFILOS
A proporção e intensidade (Média Geométrica) da fagocitose e produção de
peróxido de hidrogênio (H2O2) pelos granulócitos, estimulados ou não, estão
dispostos na tabela 12 e figuras 39 e 40.
A proporção de granulócitos que fagocitaram S. aureus foi semelhante entre
as bezerras que ingeriram colostro com (COL+) e sem células viáveis (COL-). A
proporção mínima de células fagocitando foi observada no D0 para ambos os
grupos (COL+ = 45,78%; COL- = 33,76%), em seguida, observou-se rápido
aumento no índice de fagocitose no D2 (COL+ = 89,29%; COL- = 89,04%). Os
valores decresceram gradualmente até o D21 (COL+ = 62,82%; COL- = 65,01%),
25
30
35
40
45
50
55
0 7 14 21 28
PM
N (
%)
Dias
Col+
Col-
† *
0
2
4
6
8
10
12
0 7 14 21 28
CH
13
8+
CD
62
L-(%
)
Dias
Col+
Col-
†
70
75
80
85
90
95
100
0 7 14 21 28
CH
13
8+
CD
62
L+
(%)
Dias
Col+
Col-

124
observando-se decréscimo para o COL- (61,8%) e acréscimo para o COL+
(76,35%) no D28. Não foi possível detectar diferenças estatísticas entre os grupos
durante o primeiro mês de vida (P≥0,110).
A intensidade da fagocitose da bactéria Staphylococcus aureus pelos
granulócitos foi mínima no D0 para ambos os grupos (COL+: 15,15; COL-: 5,1),
observando-se rápido aumento em D2 (COL+: 204,54; COL-: 158,07). A partir do
D2, o perfil apresentado entre os grupos foi diferenciado, detectando-se queda
gradativa no COL + até D28 (52,27) e aumento gradativo no COL- até D14 (272,08)
com queda nos momentos subsequentes (29,45-36,27). Apesar destas variações,
foi possível detectar tendência estatística apenas no 28 (P=0,081).
A proporção de granulócitos que fagocitaram Escherichia coli também foi
semelhante entre as bezerras que ingeriram colostro com (COL+) e sem células
viáveis (COL-) (P≥0,102). Ambos os grupos apresentaram valores mínimos no D0
(COL+ = 35,68%; COL- = 32,36%), observando-se aumento acentuado no D2 para
o COL+ (87,44%) e COL- (77,92%). Após, os valores de ambos os grupos
decresceram no D21 (COL+ = 55,04%; COL- = 55,71%) e aumentaram no
momento subsequente (COL+ = 73,67%; COL- = 65,47%).
A intensidade da fagocitose da bactéria Escherichia coli pelos granulócitos
foi mínima no D0 para ambos os grupos (COL+: 4,38; COL-: 4,59), observando-se
rápido aumento em D2 (COL+: 179,17; COL-: 118,28). A partir do D2, o perfil
apresentado entre os grupos foi diferenciado, detectando-se queda gradativa até
D21 (23,70) e aumento no D28 (79,97) no COL +, em contrapartida, o COL- teve
aumento gradativo até D14 (252,98) com queda nos momentos subsequentes
(18,31-43,50). Foi possível detectar diferenças estatísticas entre os grupos no D7
(P=0,034).
As diferenças obtidas para a fagocitose das bactérias S.aureus e E.coli,
considerando-se as comparações múltiplas entre os momentos, pode ser
visualizada na tabela 12.
A produção espontânea de peróxido de hidrogênio (H2O2) pelos granulócitos
(%) apresentou perfil diferenciado entre os grupos COL+ e COL-. O grupo COL+
apresentou leves variações (89,64-83,51%) durante o estudo considerando-se a
proporção de células produzindo H2O2, enquanto o COL- apresentou aumento do
D0 (COL- = 89,38%) ao D14-D28 (91,25-94,41%). Foi possível detectar tendência
estatística entre os grupos no D14 (P=0,061), momento em que o COL- (94,41%)

125
apresentava maior proporção de células produzindo H2O2 que o COL+ (90,43%).
Apesar das variações relatadas entre os momentos, não foi possível detectar
diferenças estatísticas.
A intensidade da produção espontânea de peróxido de hidrogênio (H2O2)
pelos granulócitos foi mínima no D0 (COL+: 234,55; COL-: 280,14), em seguida o
perfil foi diferenciado entre os grupos. O grupo COL+ apresentou aumento gradual
na intensidade da fluorescência até D21 (727,50) e redução em D28 (395,20), em
contrapartida, o COL- apresentou leves oscilações no período com pico máximo no
D7 (406,80). Foi possível observar tendência estatística entre os grupos no D21,
observando-se maior intensidade de fluorescência no COL+ (727,50) em relação ao
COL- (243,40) (P=0,094). Apesar das variações relatadas entre os momentos, não
foi possível detectar diferenças estatísticas.
O perfil apresentado para a proporção e intensidade da produção de H2O2
pelos granulócitos foram semelhantes para as duas bactérias usadas como
estímulo, entretanto ficou nítida a diferença entre os grupos que receberam colostro
com (COL+) e sem células viáveis (COL-).
A produção espontânea de peróxido de hidrogênio (H2O2) pelos granulócitos
(%), quando estimulados pelo S.aureus, sofreram poucas variações. O COL+
(95,73%) maiores valores que o COL- (92,37%) em D7, observando-se diferença
estatística (P=0,008). Foi observada tendência estatística no D0 entre os grupos
(P=0,092). Não foi possível detectar diferenças entre os momentos.
A intensidade da produção espontânea de peróxido de hidrogênio (H2O2)
pelos granulócitos quando estimulados com S.aureus foi crescente no COL + do D0
(584) ao D21-D28 (929,5-1011,8), em contrapartida, o COL- apresentou aumento
do D0 (359,2) ao D2 (739,1) e valores constantes nos momentos subsequentes
(739,1-864,9). Foi observada tendência estatística no D0 entre os grupos
(P=0,089). Não foi possível detectar diferenças entre os momentos.
A produção espontânea de peróxido de hidrogênio (H2O2) pelos granulócitos
(%), quando estimulados pelo E.coli sofreram poucas variações. O COL+
apresentou variações nos valores, detectando-se quedas do D0 (94,94%) ao D2
(89,66%), D14-D21 (86,51-85,81%) e aumento em D28 (93,57%). O COL-
apresentou valores constantes do D0 (91,21%) ao D14 (90,43%), detectando-se
leve aumento no D21 (92,14%) e queda em D28 (85,98%). Foi possível detectar
tendências no D1 (0,084) e diferenças estatísticas entre os grupos no D28

126
(P=0,040). Não foi possível detectar diferenças entre os momentos, apesara das
variações relatadas.
A intensidade da produção espontânea de peróxido de hidrogênio (H2O2)
pelos granulócitos quando estimulados com E.coli foi constante no COL- do D0 ao
D28 (406,6-528,10), enquanto o COL+ apresentou aumento crescente no decorrer
do estudo (505,9-746,80). Apesar destas variações não foi possível detectar
diferenças entre os momentos e grupos experimentais.

127
Tabela 12 - Proporção (%) e intensidade da fagocitose (Média Geométrica – Geo Mean) e produção
de peróxido de hidrogênio pelos granulócitos em bezerras Holandesas que receberam colostro com (COL+) e sem células viáveis (COL-) – São Paulo – 2016
Fonte: (COSTA, J. F. R., 2016) Legenda: letras minúsculas na mesma coluna demonstram diferença entre os tempos, sendo (a) valores diferentes do D0; (b) valores diferentes do D2; (c) valores diferentes do D7; (d) valores diferentes do D14; (e) valores diferentes do D21; (f) valores diferentes do D28, quando p<0,05 e tendência quando P≤0,10. * indica diferença entre os grupos e † indica tendência entre os grupos naquele momento (p>0,05 e < 0,1); * indica diferença entre os grupos e † indica tendência entre os grupos naquele momento (p>0,05 e < 0,1).
Momentos (dias) D0 D2 D7 D14 D21 D28
S.aureus
Índice Fagocitose
(%)
COL+ 45,78 ±27,6
89,29 ±11,15
79,98 ±26,26
71,13 ±22,79
62,82 ±22,02
76,35 ±14,27
COL- 33,76 ±18,72bce
89,04 ±10,77 a
84,71 ±15,43 a
77,82 ±27,04
65,1 ±18,26 a
61,8 ±26,67
S.aureus
Fagocitose (Geo Mean)
COL+ 15,15 ±28,63bcf
204,54 ±187,45 ae
117,99 ±101,49 a
103,98 ±152,02
25,72 ±20,43 b
52,27 ±41,14 a
COL- 5,1 ±3,38 bcdef
158,07 ±142,41 a
211,6 ±155,27 a
272,08 ±198,71 a
29,45 ±28,15 a
36,27 ±55,49 a
E.coli
Índice Fagocitose
(%)
COL+ 35,68 ±22,47
87,44 ±11,68
72,49 ±23,96
65,19 ±28,37
55,04 ±16,80
73,67 ±16,54
COL- 32,36 ±20,41c
77,92 ±17,06 e
81,63 ±13,06 a
82,52 ±21,66
55,71 ±18,75 b
65,47 ±13,08
E.coli
Fagocitose (Geo Mean)
COL+ 4,38 ±1,32bcef
179,17 ±164,14 a
89,47 ±82,91 a
63,65 ±103,77
23,70 ±32,70 a
79,97 ±95,05 a
COL- 4,59 ±1,50bcdf
118,28 ±128,14 a
167,08 ±110,44 a
252,98 ±244,98 a
18,32 ±18,64f
43,50 ±54,98 ae
Produção espontânea
H2O2 (%)
COL+ 91,44 ±6,19
89,64 ±7,09
91,82 ±7,59
90,43 ±5,77
91,03 ±8,74
93,51 ±3,93
COL- 84,20 ±14,31
91,03 ±5,45
89,21 ±7,74
94,41 ±1,85
93,20 ±5,51
91,25 ±11,72
Produção espontânea
H2O2 (Geo Mean)
COL+ 234,55 ±236,24
281,47 ±292,64
300,86 ±290,58
437,30 ±248,76
727,50 ±857,22
395,20 ±212,87
COL- 280,14 ±261,76
299,77 ±181,45
406,80 ±373,55
343,40 ±110,91
243,40 ±85,67
372,30 ±149,54
S.aureus Produção H2O2
(%)
COL+ 93,88 ±4,58
87,65 ±12,31
95,73 ±1,77
88,88 ±6,77
87,51 ±12,33
88,1 ±16,07
COL- 89,38 ±6,38
91,45 ±9,02
92,37 ±3,01
91,35 ±3,8
92,45 ±4,9
90,9 ±6,09
S.aureus Produção H2O2
(Geo Mean)
COL+ 584 ±392,64
603,1 ±526,54
722,7 ±609,84
954,7 ±762,59
1011,8 ±1023,82
929,5 ±511,97
COL- 359,2 ±220,75 d f
739,1 ±505,25
801 ±767,01
864,9 ±306,87 a
692,8 ±253,67
812,4 ±219,56 a
E.coli Produção H2O2
(%)
COL+ 94,94 ±2,51
89,66 ±6,59
94,08 ±4,05
86,51 ±8,30
85,81 ±12,01
93,57 ±4,31
COL- 91,21 ±5,79
91,14 ±5,86
91,47 ±4,13
90,43 ±5,42
92,14 ±5,71
85,98 ±10,98
E.coli Produção H2O2
(Geo Mean)
COL+ 505,90 ±234,58
427,44 ±367,05
557,50 ±669,89
661,60 ±584,71
771,70 ±692,03
746,80 ±310,67
COL- 406,60 ±252,43
457,56 ±236,20
508,10 ±410,26
534,30 ±207,22
490,80 ±219,55
528,10 ±187,24

128
Figura 39 - Proporção (%) e intensidade da fagocitose (Média Geométrica– Geo Mean) da fagocitose pelos granulócitos mediante liberação espontânea ou estimulados, em bezerras Holandesas recém-nascidas que receberam colostro com (COL+) e sem células viáveis (COL-) – São Paulo - 2016
Fonte: (COSTA, J. F. R., 2016). Legenda: * indica diferença entre os grupos e † indica tendência entre os grupos naquele momento (p>0,05 e < 0,1)
30
40
50
60
70
80
90
100
0 7 14 21 28
Fag
oci
tose
(%
)
Dias
E. coli
Col+
Col-
0
50
100
150
200
250
300
0 7 14 21 28
Fag
oci
tose
(G
M)
Dias
S. aureus
Col+
Col-
†
0
50
100
150
200
250
300
0 7 14 21 28
Fag
oci
tose
(G
M)
Dias
E. coli
Col+
Col-
30
40
50
60
70
80
90
100
0 7 14 21 28
Fag
oci
tose
(%
)
Dias
S. aureus
Col+
Col-

129
Figura 40 - Produção de peróxido de hidrogênio (% e intensidade – Média Geométrica – Geo Mean)
pelos granulócitos, mediante liberação espontânea ou estimulados, em bezerras Holandesas recém-nascidas que receberam colostro com (COL+) e sem células viáveis (COL-) - São Paulo - 2016
Fonte: (COSTA, J. F. R., 2016). Legenda: * indica diferença entre os grupos e † indica tendência entre os grupos naquele momento (p>0,05 e < 0,1)
8486
8890
9294
9698
0 7 14 21 28
Bu
rst
(%)
Dias
E. coli
Col+
Col-
†
8486889092949698
0 7 14 21 28
Bu
rst
(%)
Dias
S. aureus
Col+
Col-
† *
350450550650750850950
1050
0 7 14 21 28
Bu
rst
(GM
)
Dias
S. aureus
Col+
Col-
350450550650750850950
1050
0 7 14 21 28
Bu
rst
(GM
)
Dias
E. coli
Col+
Col-
82
84
86
88
90
92
94
96
0 7 14 21 28
Bu
rst
bas
al (
%)
Dia
Liberação espontânea
Col+
Col-
†
200
300
400
500
600
700
800
0 7 14 21 28
Bu
rst
bas
al (
GM
)
Dia
Liberação espontânea
Col+
Col-
†

130
11 DISCUSSÃO
Esta pesquisa avaliou a influência das células do colostro na resposta imune
inata e ocorrência de diarreias em bezerras recém-nascidas.
11.1 QUALIDADE DO COLOSTRO
A CCS (1.895.844 células/mL) do colostro obtida neste estudo foi superior ao
valor de 878.000 células/mL encontrado por Gomes et al. (2011). Considerando-se a
concentração celular obtida, pode-se estimar que as bezerras receberam um total de
células de 7,58x109 em quatro litros de colostro. Silva (2014) relatou que o colostro
de primeira ordenha apresenta 32% de viabilidade celular, assim pode-se estimar a
ingestão de aproximadamente 2,42x109 células viáveis no volume mínimo
equivalente a quatro litros de colostro administrado às recém-nascidas.
Novo et al. (2014) demonstraram que não há células viáveis após
congelamento das amostras em freezer -20ºC. No estudo da equipe foram feitos
testes a partir de uma hora de congelamento, com intervalos de hora em hora até 6
horas no freezer e após 24 horas. Em todas as análises foi possível observar que o
congelamento lisou as células, tornando-as inviáveis. Langel et al. (2015) utilizaram
os mesmos grupos experimentais (COL+ e COL-) para desenvolver seu trabalho,
porém com a diferença que o colostro congelado que as bezerras mamaram foi
congelado em nitrogênio líquido para que o processo de lise das células ocorresse
de forma rápida e, assim, fosse possível que a bezerra recebesse o colostro
congelado de sua própria mãe. Desta forma, não precisaram mamar colostro de
doadoras, como ocorreu neste experimento.
A concentração de Igs do colostro e sólidos totais dos dois grupos
experimentais apresentaram valores adequados, segundo o limiar inicial e bem
consolidado de 50g/L estabelecidos por Chigerwe et al. (2008); Godden (2008);
Morril et al. (2012) e Quigley et al. (2013). Se considerarmos o ponto de corte
determinado mais recentemente para o teor de IgG (IgG ≥ 80g/L) e sólidos totais
(≥23%), respectivamente, determinados pelo colostrômetro e refratômetro Brix, os

131
valores continuam acima deste limiar, garantindo assim que foi fornecido colostro de
alta qualidade para as bezerras deste experimento, acima do recomendado pela
literatura atual (BARTIER; WINDEYER; DOEPEL, 2015).
A ausência de diferenças estatísticas entre os valores obtidos para a
concentração de imunoglobulinas e sólidos totais do colostro para o COL+ e COL-
permitiu a confirmação da padronização da qualidade do colostro fornecido para os
dois grupos experimentais.
11.2 STATUS SANITÁRIO DAS BEZERRAS
A principal doença que acometeu as bezerras deste experimento no período
neonatal foi a diarreia. Através da avaliação dos escores pode-se observar que a
doença teve início no D2 e perdurou pelas próximas três semanas. A maior
ocorrência de diarreia foi observada em ambos os grupos no D7 e D14.
Nesta pesquisa não foi observada diferença para as frequências de diarreias
entre os grupos e momentos avaliados, porém o grupo COL- apresentou maior
número de bezerras apresentando diarreia severa, com escore de fezes igual a 3.
Riedel-Caspari (1993) após infectar as bezerras com E. coli observou a
manifestação de diarreias em ambos os grupos de bezerras que haviam recebido
colostro com (COL+) e sem células viáveis (COL-). As bezerras que mamaram
colostro acelular apresentaram quadro clínico de maior intensidade, considerando o
aumento de temperatura retal durante todo o período, além da maior eliminação por
tempo prolongado de E. coli nas fezes. Estes dados coincidem com os achados da
presente pesquisa, considerando-se maior frequência de escore de fezes 3 nas
bezerras COL-. A autora ressalta que os leucócitos colostrais, bem como outras
substâncias do colostro, melhoraram a resistência do animal contra a infecção
artificial à qual elas foram submetidas desde o estágio inicial e após a infecção. A
autora levantou duas hipóteses sobre os mecanismos de defesa envolvendo as
células do colostro: a primeira é de que os leucócitos do colostro podem ter atuado
através de efeito antimicrobiano ou então, as células podem ter estimulado a
resistência endógena da bezerra à infecção.

132
Langel et al. (2015) analisaram o escore de fezes de bezerras Holandesas e
Jersey recém-nascidas, durante o primeiro mês, que receberam colostro fresco ou
congelado. Estes autores também não encontraram diferenças para o escore de
fezes em relação aos grupos experimentais. A maior taxa de diarreia observada
pelos retro-referidos autores foi aos 14 dias de vida, período coincidente com os
achados deste experimento.
A elevada ocorrência de diarreia observada nas bezerras COL+ (50-70%) e
COL- (44-78%) no primeiro mês de vida foi superior à prevalência (19,1%) e
incidência (21,2%) reportados por outros pesquisadores (BARTELS et al., 2010;
WINDEYER et al., 2014).
A elevada frequência de diarreia a partir do D2 nas bezerras que ingeriram
colostro com (COL+) e sem células viáveis (COL-) pode estar relacionada a diversos
fatores de risco.
O alojamento e manejo ao qual as bezerras foram submetidas durante o
período experimental pode ter influenciado na elevada ocorrência de diarreia. As
bezerras eram transferidas das baias individuais e suspensas localizadas na
maternidade para o bezerreiro aproximadamente seis horas após o parto. A
estrutura do bezerreiro contempla uma área coberta de aproximadamente 1040 m2,
com a possibilidade de fechamento lateral com cortinas de tecido sintético,
composto por 100% de polipropileno. No bezerreiro, as bezerras inicialmente foram
alojadas em baias de madeira não suspensas onde era fornecido o leite de
transição. Após três dias de vida, as bezerras foram novamente movidas para
gaiolas suspensas de metal (1,2 m2), onde foram mantidas até 70 dias de vida. No
período de aleitamento, as bezerras recebiam seis litros de leite sem antibióticos, em
baldes, proveniente do rebanho.
As baias da maternidade eram higienizadas a cada cinco dias, sendo que
uma vez por semana as paredes eram pintadas com uma mistura de cal com água.
As baias de transição não suspensas e localizadas no bezerreiro também eram
lavadas e desinfetadas com água e cal. O chão desta baia era um tapete de
borracha, onde os funcionários colocavam cal e capim por cima. O chão e o teto do
barracão eram higienizados antes da entrada das bezerras com água pressurizada
e, em seguida, com cal. O cloro era usado como desinfetante para a limpeza dos
baldes e mamadeiras. Nos tanques onde o leite era armazenado eram usados
produtos de limpeza de ordenha: alcalino clorado e ácido.

133
O maior risco para a ocorrência da diarreia no manejo supra-descrito está
relacionado à higienização das baias individuais, onde, apesar dos responsáveis
pela fazenda terem consciência que o ideal seria a troca da cama e higienização a
cada nova bezerra, o fato de não trocarem o material constantemente permite que
haja contaminação entre os animais que são inseridos em ambiente já utilizado por
outras bezerras.
Outros fatores de risco citados por McGuirk (2008) também podem ser
mencionados considerando-se a realidade da fazenda onde este experimento foi
realizado, tais como: contaminação oro-fecal pelo colostro ou ambiente; presença de
outros animais no recinto como gatos; possível higienização não adequada de
utensílios como mamadeiras, sondas esofágicas, bem como da limpeza do uniforme
dos funcionários, como botas, macacões além da higiene das mãos dos cuidadores
das bezerras.
Os principais patógenos causadores de diarreia em bezerros neonatos são
Escherichia coli, Rotavírus, Coronavírus e Cryptosporidium parvum, identificados
antes e após a primeira semana de vida (GULLIKSEN et al., 2009; BARTELS et al.,
2010; CHO; YOON, 2013; MEGANCK et al., 2015). A identificação do patógeno
causador da diarreia não foi contemplada no objetivo deste estudo, porém a maior
ocorrência de diarreia entre 7 e 14 dias de vida aponta para maior possibilidade para
agentes como Rotavirus, Coronavirus, Cryptosporidium (MCGUIRK, 2008; CHO;
YOON, 2013).
As vacas desta fazenda são vacinadas no período pré-parto para Escherichia
coli, Rotavírus e Coronavírus, assim, a ocorrência de diarreia deveria ter sido menor
do que os valores obtidos neste experimento. Meganck et al. (2015) mostraram que
a vacinação materna reduz a frequência da diarreia neonatal de 39,9% para 14,3%.
Broncopneumonia foi observada em apenas uma bezerra do COL+ aos 14
dias de vida e COL- aos 28 dias de vida. Langel et al. (2015) também avaliaram o
escore de broncopneumonia em bezerras que receberam colostro fresco e
congelado. Estes autores detectaram maior ocorrência de escore respiratório aos 12
dias de vida no COL-, porém sem diferença estatística. O período onde foi detectada
diferença estatística foi aos 38 dias, fase que não avaliamos neste experimento.
Durante todo o experimento foram observados 23 episódios de anemia. A
anemia, predominantemente de acordo com a morfologia dos eritrócitos, foi a
normocítica hipocrômica e o grupo mais anêmico foi o COL-. No caso da anemia da

134
inflamação, a absorção e distribuição das reservas de ferro não são suficientes para
satisfazer a síntese de hemoglobina na medula óssea, mesmo tendo adequada
ingestão de ferro na alimentação (GANZ, 2005). Acredita-se que a anemia
encontrada no grupo COL- foi decorrente da resposta inflamatória causada a partir
dos agentes responsáveis pela diarreia. Ganz (2005) afirmaram que este tipo de
anemia pode ser um efeito colateral da resposta de defesa do hospedeiro para o
controle da multiplicação bacteriana, considerando-se que o ferro é fundamental ao
seu metabolismo.
O diferente status sanitário apresentado pelos grupos experimentais pode
estar relacionado à ingestão e absorção de células maternas do colostro.
11.3 CORTISOL
O perfil da concentração do cortisol foi semelhante entre os grupos que
receberam colostro com (COL+) e sem células viáveis (COL-). Ambos os grupos
apresentaram pico de cortisol no D0, o que coincide com o momento pós-
nascimento. O momento do parto é um período crítico e estressante, tanto para a
vaca quanto para o bezerro, mesmo no parto eutócico, onde são observadas
alterações endócrinas com a liberação de cortisol, noradrenalina e adrenalina. O
momento do parto é bastante delicado, fazendo-se necessário um acompanhamento
rigoroso durante as primeiras horas de vida (VANNUCCHI et al., 2015).
Uma das principais funções do cortisol é manter o equilíbrio da glicose do
sangue. O aumento do cortisol serve, inicialmente, para prevenir a hipoglicemia
durante estresse agudo ou prolongado como, por exemplo, do parto (BENFIELD et
al., 2014).
Os altos níveis de cortisol que os bezerros produzem durante o parto
permanecem elevados durante a primeira semana de vida, dependendo do
oferecimento de condições ambientais e de manejo favoráveis à adaptação orgânica
do recém-nascido. Ao nascer, bezerras necessitam passar por um processo de
adaptação orgânica ao ambiente extrauterino. Algumas destas adaptações foram

135
avaliadas do nascimento ao desmame por Silva et al. (2016, no prelo) 12 ,
observando-se que a primeira semana pós-natal foi marcada por importantes
alterações fisiológicas inerentes à adaptação das funções cardiorrespiratórias, com
algumas oscilações nos momentos seguintes, que foram atribuídas a oscilações
climáticas do ambiente onde foram criadas. Os autores relataram impactos diretos
do período de adaptação extrauterina nos parâmetros fisiológicos dos neonatos. A
inadequada adaptação pode prolongar o estresse e os elevados níveis de cortisol.
Nesta pesquisa observou-se rápida diminuição dos níveis de cortisol já na
primeira semana de vida. Esta adaptação em relação aos níveis de cortisol pode
estar relacionada às baias de transição, pois provavelmente elas devem ter
fornecido condições que otimizassem esta estabilidade orgânica pós-nascimento.
O acúmulo desses hormônios tem um efeito supressivo nas respostas imunes
dos bezerros, afetando diretamente a resposta imune inata, causando diminuição na
fagocitose e na atividade bactericida dos neutrófilos e macrófagos (BARRINGTON;
PARISH, 2001; MOREIN et al., 2002; CHASE; HURLEY; REBER, 2008).
11.4 HAPTOGLOBINA
As proteínas de fase aguda são um excelente marcador inflamatório utilizado
para mensurar a inflamação. Dentre as proteínas de fase aguda, a haptoglobina é a
proteína de fase aguda (PFA) mais indicada para a mensuração de inflamação em
bovinos (MURRATA et al., 2004; PETERSEN et al., 2004).
As PFA são produzidas em resposta ao estímulo bacteriano e podem ser
usadas como um indicador quantitativo da infecção, inflamação, lesão tecidual e
estresse. A hp reduz o estresse pró-oxidativo e pró-inflamatório através da formação
de complexos pela ligação da hp com a hemoglobina e retirando-a da circulação
(MURRATA et al., 2004; PETERSEN et al., 2004).
Bezerros nascem com baixa concentração de hp, que podem estar ligadas ao
sistema imunológico. Kruger et al. (2015) acreditam que o aumento da hp deve ser
12
SILVA, B. T.; HENKLEN, A.; MARQUES, R. S.; OLIVEIRA, P. ;L.; LEITE, S. B. P.; NOVO, S. M. F.; BACCILI, C. C.; REIS, J. F.; GOMES, V. Vital parameters of Holstein calves from birth to weaning. Revista Brasileira de Medicina Veterinária. [200-]. (no prelo).

136
visto como um evento positivo que indica inflamação nos primeiros dias de vida e
consideram este fenômeno como algo normal e necessário para que haja a
adaptação do animal ao ambiente extrauterino. Por outro lado, o que foi observado
nesta pesquisa é que a hp aumentou apenas nos momentos em que as bezerras
apresentaram diarreia, desta forma, contradizendo o autor supracitado e reafirmando
a possibilidade de ela ser considerada um marcador inflamatório.
A haptoglobina das bezerras deste experimento apresentou perfil diferente
entre os dois grupos, apesar da ausência de diferenças estatísticas. Foi possível
observar que ambos começaram com baixos valores, tanto no nascimento quanto no
D2. Já no D7 houve um acentuado aumento, atingindo o pico para o COL+ que logo
em seguida decresceu de forma acentuada, observando-se valores próximos ao
basal no D14. Já o COL- apresentou valores elevados em D7 e D14. Analisando os
resultados, pode-se observar que os bezerros que ingeriram colostro sem células
viáveis (COL-) apresentaram uma resposta inflamatória mais prolongada,
principalmente nos momento em que houve diarreia. Pourjafar et al. (2011) também
avaliaram a hp de bezerras durante o primeiro mês de vida e encontraram picos de
hp coincidentes com o período em que elas apresentaram diarreia e provaram que a
hp é um indicador confiável da severidade da diarreia em bezerras recém-nascidas.
Possivelmente as citocinas pró-inflamatórias, especialmente a interleucina 6,
estimularam o fígado para a produção de proteínas de fase aguda, dentre elas a
haptoglobina (HEINRICH; CASTELL; ANDUS, 1990). A haptoglobina é induzida pela
citocina IL-6 e é caracterizada por ter um aumento na concentração sérica mais
tardio, permanecendo elevado até a segunda semana (PETERSEN et al., 2004).
Os valores apresentados neste trabalho confirmam que foi possível mensurar
a inflamação através da formação dos complexos da haptoglobina, pois o pico de
concentração de hp coincide com os momentos em que as bezerras apresentaram
diarreia.
11.5 FERRO
Os animais do grupo COL- apresentaram menores valores de hemácias,
hematócrito e hemoglobina, especialmente no D2. Este momento coincide com

137
acentuada diminuição nos valores de ferro sérico das bezerras que ingeriram
colostro sem células viáveis.
Citocinas pró-inflamatórias, especialmente IL-1 e IL-6 são responsáveis pela
indução da produção de proteínas de fase aguda, como por exemplo a hepcidina.
Essa substância se liga com a porção férrica dos enterócitos, hepatócitos e
macrófagos para bloquear a liberação de ferro armazenado nestas células. O efeito
desta interação resulta na inibição da absorção de ferro da dieta e limita a
exportação do mineral a partir do macrófago e hepatócito. O ferro é fundamental
para as funções vitais das bactérias, tais como síntese de DNA e transporte de
elétrons (NEMETH; GANZ, 2006).
11.6 PROPORÇÃO E FUNÇÃO DOS NEUTRÓFILOS
O leucograma apresentou perfil ligeiramente diferenciado entre os grupos
COL- e COL+. A oscilação observada no gráfico dos leucócitos totais e dos
neutrófilos foi muito semelhante para o grupo COL+ e COL-, porém a queda do
número de neutrófilos a partir do D7 foi mais intensa no COL+. Este fato pode ser
associado com a maior eficiência dos neutrófilos das bezerras COL+ em migrar a
partir dos episódios de diarreia (ZWAHLEN; ROTH, 1990).
Novo (2015) utilizou as mesmas bezerras deste experimento para o seu
estudo. O alto número de células T gamma-delta ativadas presentes no grupo COL+
pode ser explicado pela resposta inflamatória mais rápida em comparação ao tempo
que COL- levou para responder à inflamação. Acredita-se que os neutrófilos das
bezerras que mamaram o colostro fresco tiveram maior agilidade em perceber o
estímulo e se mover no sentido do tecido para combater a inflamação. As células T
gamma-delta representam a principal fonte de IL-17, que é responsável pelo
recrutamento para o tecido inflamado, e essa migração dos neutrófilos observada no
D7 coincidiu com os episódios detectados de diarreia. (ROARK et al., 2008)
Foi possível observar diferença entre os momentos para os leucócitos totais
do grupo COL- onde o D0 foi diferente do D28. Em relação ao número de neutrófilos
no grupo COL+, o D0 foi diferente em relação ao D14, D21, D28.

138
Nagahata et al. (2000) sugeriram que o aumento do cortisol sérico poderia
resultar em uma diminuição da expressão da L-selectina na superfície dos
neutrófilos. Este fato justificaria a diminuição da ligação dessas células com as
endoteliais, aumentando o número dos neutrófilos na circulação com redução no
compartimento marginal; portanto, o aumento, dos neutrófilos ocorre pela somatória
entre os neutrófilos circulantes e aqueles liberados do estoque da medula óssea. O
que observamos neste experimento foi que no momento em que o cortisol esteve
alto (D0), a expressão de L-selectina na superfície dos neutrófilos estava ao redor de
85-90%. Talvez o efeito do cortisol sobre a expressão da L-selectina tenha sido
mínimo considerando as condições de manejo oferecidas pelas bezerras.
A avaliação da influência das células do colostro sobre a proporção das
subpopulações de leucócitos mononucleares do sangue das bezerras tem sido
avaliada pela técnica de imunofenotipagem (REBER et al., 2006; REBER et al.,
2008a,b; LANGEL et al., 2015; NOVO, 2015), porém, pouco ainda se tem na
literatura a respeito da proporção e função das células polimorfonucleares CH138+
expressando ou não da L-selectina.
O grupo COL+ apresentou queda brusca na proporção de PMN do D7 ao D21
que coincidiu com a redução na proporção de células expressando CD62L,
provavelmente porque já haviam migrado para o tecido inflamado. Em contrapartida,
o COL- apresentou queda lenta e gradual do PMN que coincidiu com maior
expressão CD62L na circulação sanguínea devido ao lento processo de migração
destas células aos tecidos.
A população CH138+CD62L+ é representada por granulócitos ativados. A
ativação acentuada dos neutrófilos através da L-selectina ocorreu para que
houvesse o mecanismo de adesão dos neutrófilos às células endoteliais dos vasos
para a migração das células para o tecido através da diapedese. Este processo
parece ter sido mais rápido nos bezerros que ingeriram colostro materno com células
(COL+) observado no D14 em relação ao COL- (D21). Esta hipótese é reforçada
pelo perfil dos valores absolutos de neutrófilos determinados pelo leucograma, no
qual o COL+ apresentou queda brusca do D7 ao D14, enquanto o COL- apresentou
queda lenta e gradual no decorrer do estudo.
A população CH138+CD62L- circulante apresentou tendência para diferentes
perfis, observando-se maiores proporções no COL+ em relação ao COL-. Este fato
pode indicar diferenças na intensidade da diarreia apresentada pelos dois grupos,

139
considerando que o COL- apresentou maior frequência de escore 3. Os granulócitos
CH138+ das bezerras COL- estavam sendo recrutados em quase totalidade para a
defesa contra a infecção. O COL+ apresentou um pouco mais de neutrófilos
circulantes sem que estivessem sendo recrutados ao tecido, mais um sinal que
comprova que estes animais estiveram mais sadios e menos debilitados,
necessitando um pouco menos da atividade.
Reber et al. (2006) também não encontraram diferenças entre os grupos
COL+ e COL- para a expressão de L-selectina das células mononucleares
sanguíneas. Os autores acreditam que a L-selectina desempenha um papel menos
importante na migração de células através da parede do intestino devido ao menor
fluxo dos vasos na região intestinal.
Langel et al. (2015) observaram que a expressão de L-selectina foi maior nos
linfócitos no grupo acelular. Os autores acreditam que possa ter havido expressão
do CD62-L na superfície celular em resposta a antígenos ou estímulo de citocinas.
Gomes et al. (2014) avaliaram a influência do colostro no desenvolvimento da
resposta imune inata de bezerros recém-nascidos pela imunofenotipagem do
CH138+ das células sanguíneas. Os autores observaram que as proporções de
neutrófilos CH138+ estiveram maiores no nascimento, e permaneceram altas até o
15º dia de vida. Este perfil se assemelhou ao encontrado nesta pesquisa para
CH138+CD62L+ os quais também permaneceram altos praticamente nos mesmos
períodos, tendo baixa nos valores no 14º dia. Estas alterações na população de
CH138+ podem estar relacionadas ao estresse do nascimento e neste caso não
foram relacionadas a processos inflamatórios, pois os autores detectaram baixos ou
indetectáveis níveis de citocinas pró-inflamatórias.
A partir dos dados apresentados para o perfil de COL+ e COL- foi possível
notar uma maior e mais rápida resposta nas bezerras COL+, observando maior
dificuldade do COL- em reagir aos desafios. A hipótese que levantamos para
justificar esse achado é de que as bezerras COL+ apresentaram uma maior resposta
Th17 através do estímulo das células gamma-delta. Possivelmente, o que possa ter
ocorrido é que ao ingerir o colostro com células viáveis, as mesmas foram
absorvidas pelo epitélio intestinal. Ao nascimento, ambos os grupos foram
desafiados pelos microrganismos do meio ambiente. Desta forma, os patógenos que
adentraram o organismo e estiveram presentes na parede do intestino foram
fagocitados por células dendríticas e macrófagos, seguiram aos linfonodos

140
mesentéricos e placas de Peyer onde estimularam a expansão clonal de linfócitos T.
Esta expansão clonal estimulou a produção de células T gamma-delta que, por sua
vez, ao liberarem a citocina IL-17, atraíram mais neutrófilos para combater a
infecção no foco da inflamação observados no COL-.
Esta hipótese pode ser sustentada pelos achados de NOVO (2015) que, ao
utilizar as mesmas bezerras para o experimento e mensurar a IL-17, obteve que as
células T gamma-delta das bezerras COL+ apresentaram produção precoce da
citocina, recrutando neutrófilos de forma mais ágil. Shibata et al. (2007) analisaram a
produção de IL-17 por células T gamma-delta em ratos infectados com E. coli e
demonstraram que estas células são importantes para a resposta a estímulos
bacterianos.
Para avaliar a função dos neutrófilos cabe explicar o motivo pelo qual não foi
utilizado o marcador CH138+ para definir o local exato onde os neutrófilos estavam
dentro do gate de polimorfonucleares. Em experimento anterior da equipe, SILVA
(2014) avaliou a fagocitose e produção de burst oxidativo das células
polimorfonucleares do sangue de vacas no periparto pela mesma técnica de
fagocitose de S. aureus e E. coli desenvolvida neste experimento. Neste trabalho,
SILVA (2014) observou que a maioria das células dentro do gate PMN eram células
CH138+ (96%).
A metodologia empregada para a mensuração da fagocitose e produção de
radicais livres foi testada e comprovada por Kampen et al. (2004) que, dentre cinco
metodologias testadas, concluiu que a citometria de fluxo é ideal por ser considerada
relativamente fácil, por usar uma pequena quantidade de sangue, e por permitir
isolamento da população celular de interesse.
Os PMN apresentaram perfis diferentes em ambos os grupos, o que permitiu
a detecção de algumas diferenças ou tendências estatísticas. Os neutrófilos do
COL- foram mais reativos durante praticamente todo o período estudado,
detectando-se tendência no D14 e diferença no D21 entre grupos.
O índice de fagocitose realizada pelas células polimorfonucleares,
estimulados com S. aureus e Escherichia coli, foram semelhantes entre os grupos
de bezerras que ingeriram colostro com (COL+) e sem células viáveis (COL-),
observando-se pico máximo no D2. Em contrapartida, a intensidade da fagocitose
foi maior no COL- que apresentou valores crescentes do D7 ao D14 e queda brusca
em D21. Diferença estatística foi comprovada no D7 para a E.coli, onde o COL-

141
apresentou maior intensidade de fluorescência. Este fato aponta que os PMN do
grupo COL- apresentavam maior quantidade de bactérias em seu interior e maior
atividade celular.
O perfil apresentado para a produção de peróxido de hidrogênio foi diferente
entre os grupos, apesar das poucas diferenças estatísticas obtidas. A proporção de
células produzindo o H2O2 basal foi maior no COL- no D14, entretanto as bezerras
que ingeriram colostro com células (COL+) apresentaram maior intensidade de
fluorescência no D21. Estes achados indicam que as células PMN das bezerras
COL+ possuem maior capacidade para produzir peróxido de hidrogênio.
A proporção de células PMN produzindo H2O2 após a estimulação com o
S.aureus e E.coli foi constante no COL- e sofreram variações no COL+ observando-
se pico máximo em D7 e redução entre D14 e D28. Este fato pode apontar para um
constante desafio no COL- ao longo do estudo e redução da função dos PMN no
COL+ devido ao melhor status sanitário dessas bezerras.
A intensidade da fluorescência obtida para a produção de H2O2 quando
estimuladas com S.aureus e E.coli também foi mais intensa no COL+ em relação ao
COL- apesar da ausência de diferenças estatísticas entre os grupos ao longo do
estudo.
Batista et al. (2015) avaliaram o sistema imune inato das bezerras através da
fagocitose e produção de radicais livres por fagócitos CH138+ e CD14+ estimulados
com S. aureus e E. coli durante os primeiros 90 dias de vida. Os autores
encontraram maior intensidade de fagocitose de S. aureus na primeira semana. Na
presente pesquisa, a maior intensidade foi observada na primeira semana de vida
para o COL+ e na segunda semana para o COL-. Possivelmente o perfil se
assemelhou ao encontrado no COL+ devido ao melhor status sanitário desse grupo.
O índice de fagocitose (%) dos granulócitos apresentou semelhança entre os
grupos, entretanto a intensidade da fluorescência foi mais expressiva no COL-. Em
contrapartida, o COL+ demonstrou maior habilidade em produzir H2O2. Desta forma,
a grande quantidade de bactérias no interior dos granulócitos das bezerras que
ingeriram colostro sem células viáveis pode ser explicada pela menor eficiência na
produção de peróxido de hidrogênio.

142
12 CONCLUSÕES
Com base nos dados obtidos, pode-se concluir:
(a) As células do colostro influenciaram no perfil sanitário apresentado pelas
bezerras, observando-se maior intensidade de diarreias, menores teores de ferro
sérico e anemias no grupo que recebeu colostro congelado;
(b) A migração dos granulócitos sanguíneos foi mais rápida e intensa no COL+ em
relação às bezerras que receberam colostro congelado (COL-), após exposição
natural aos patógenos causadores de diarreia;
(c) O índice de fagocitose (%) dos granulócitos apresentou semelhança entre os
grupos, entretanto a intensidade da fluorescência foi mais intensa no COL-. Em
contrapartida, o COL+ demonstrou maior habilidade em produzir H2O2.

143
REFERÊNCIAS
BARRINGTON, G. M.; PARISH, S. M. Bovine neonatal immunology. Veterinary Clinics of North America : food animal practice, v. 17, n. 3. p. 463-476, 2001. BARTELS, C. J. M.; HOLZHAUER, M.; JORRITSMA, R.; SWART, W. A. J. M.; LAM, T. J. G. M. Prevalence, prediction and risk factors of enteropathogens innormal and non-normal faeces of young Dutch dairy calves. Preventive Veterinary Medicine , v. 93, p. 162–169, 2010. BARTIER, A. L.; WINDEYER, M. C.; DOEPEL, L. Evaluation of on-farm tools for colostrum quality measurement. Journal of Dairy Science, v. 98, p. 1878-1884, 2015. BATISTA, C. F.; BLAGITZ, M.G.; BERTAGNON,H. G.; GOMES, R. C.; SANTOS, K. R.; DELLA LIBERA, A. M. M. P. Evolution of phagocytic function in monocytes and neutrophils blood cellls of healthy calves. Journal of Dairy Science , v. 98, p. 8882-8888, 2015. BENESI, F. J.; TEIXEIRA, C. M. C.; LEAL, M. L. R.; LISBOA, J. A. N.; MIRANDOLA, R. M. S.; SHECAIRA, C. L.; GOMES, V. Leukograms of healthy Holstein calves within the first month of life. Pesquisa. Veterinária Brasileira , v. 32, n. 4, p. 352-356, 2012. BENFIELD, R. D.; NEWTON, E. R.; TANNER, C. J.; HEITKEMPER, M. M. Cortisol as a biomarker of stress in term human labor: physiological and methodological issues. Biological Research for Nursing, v.16, p. 64–71, 2014. BRUN-HANSEN, H. C.; KAMPEN, A. H.; LUND, A. Hematologic values in calves during the first 6 months of life. Veterinary Clinical Pathology, v. 35, n. 2, p. 182-187, 2006. BURTON, J. L.; KEHRLI JR.; M. E.; KAPIL S.; HORST R. L. Regulation of L-selectin and CD18 on bovine neutrophils by glucocorticoids: effects of cortisol and dexamethasone. Journal of Leukocyte Biology , v. 57, p. 317–325, 1995. CHASE, C. C. L.; HURLEY, D. J.; REBER, A. J. Neonatal immune development in the calf and its impact on vaccine response. Veterinary Clinics of North America: food animal practice, v. 24, p. 87–104, 2008. CHIGERWE, M.; TYLER, J. W.; MIDDLETON, J. R.; SPAIN, J. N.; DILL, J. S.; STEEVENS, B. J. Comparison of four methods to assess colostral IgG concentration in dairy cows. Journal of the American Veterinary Medical Association , v. 233, p. 761-766, 2008. CHO, YONG-IL.; YOON, KYOUNG-JIN. An overview of calf diarrhea - infectious etiology, diagnosis, and intervention. Journal Veterinary Science, v. 15 n.1, p. 1-17, 2013.

144
CORTESE, V. S. Neonatal immunology. Veterinary Clinics North America: food animal practice , v. 25, n.1, p. 221-227, 2009. CROSS, M. L.; GILL, H. S. Immunomodulatory properties of milk. British Journal of Nutrition , v. 84, p. S81-S89, 2000. Supplement, 1. DIRKSEN, G; GRÜNDER, H D; STÖBER, M R. Exame clínico dos bovinos . Rio de Janeiro: Guanabara Koogan, 2008. DONOVAN, D. C.; REBER, A. J.; GABBARD, J. D.; ACEVES-AVILA, M.; GALLAND, K. L.; HOLBERT, K. A.; ELY, L. O.; HURLEY, D. J. Effect of maternal cells transferred with colostrum on cellular responses to pathogen antigens in neonatal calves. American Journal of Veterinary Research , v. 68, n. 7, p. 778-82, 2007. GANZ, T. Hepcidin: A regulator of intestinal iron absorption and iron recycling by macrophages. Best Practice & Research Clinical Haematology , v. 18, n. 2, p. 171–182, 2005. GODDEN, S. Colostrum management for dairy calves. Veterinary Clinics North America: food animal practice , v. 24, p. 19- 39, 2008. GOMES, V.; BACCILI, C. C.; BALDACIM, V. A. P.; MADUREIRA, K. M.; GUILLOUX, A. G. A.; POZZI, C. R.; GOMES, C. O. M. S. Development of the innate immune response and influence of colostrum suckling in calves. American Journal of Animal and Veterinary Sciences, v. 9, n. 2, p. 77-83, 2014. GOMES, V.; MADUREIRA, K. M.; DELLA LIBERA, A. M. M. P.; ALVES, M.; BAPTISTELLA, F.; BENESI, F. J. Dinâmica da celularidade do colostro de vacas holandesas no pós-parto imediato. Arquivo Brasileiro de Medicina Veterinária e Zootecnia , v. 63, n. 5, p.1047-1053, 2011. GULLIKSEN, S. M.; JOR, E.; LIE, K. I.; HAMNES, I. S.; LOKEN, T.; ÅKERSTEDT, J.; OSTERÅS, O. Enteropathogens and risk factors for diarrhea in Norwegiandairy calves. Journal of Dairy Science , v. 92, p. 5057–5066, 2009. HEINRICH, P. C.; CASTELL, J. C. ; ANDUS, T. Interleukin-6 and the acute phase response. Biochemical Journal, v. 265, p. 621–636, 1990. KAMPEN, A. H.; TOLLERSRUD, T.; LARSEN, S.; ROTH, J. A.; FRANK, D. E; LUND, A. Repeatability of flow cytometric and classical measurement of phagocytosis and respiratory burst in bovine polymorphonuclear leukocytes. Veterinary Immunology and Immunopathology, v. 97, p.105–114, 2004. KRUGER, L.; BEITZ, D. C.; STUART, R. L.; STABEL, J. R. Early Inflammation Disorder in Neonatal Calves. Animal Industry Report, v. 661, n. 1, p. 32-42, 2015.

145
LANGEL, S. N.; WARK, W. A.; GARST, S. N.; JAMES, R. E.; McGILLIARD, M. L.; PETERSSON-WOLFE, C. S.; KANEVSKY-MULLARKY, I. Effect of feeding whole compared with cell-free colostrum on calf immune status: The neonatal period. Journal of Dairy Science , v. 98, n. 6, p. 3729-3740, 2015. LIEBLER-TENORIO, E. M.; RIEDEL-GASPARI, G.; POHLENZ, J. F. Uptake of colostral leukocytes in the intestinal tract of newborn calves. Veterinary Immunology and Immunopathology, v. 85, n.1-2, p. 33-40, 2002. McDONALD, J.S.; ANDERSON, A.J. Total and diferential somatic cell counts in secretions from no ninfected bovine mammary glands: the peripartum period. American Journal of Veterinary Research , v.45, p.1.366-1.368, 1981. McGUIRK, S. M. Disease management of dairy calves and heifers. Veterinary Clinics of North America, food animal practice , v. 24, n.1, p. 139-156, 2008. MEGANCK, V.; GODDEERIS, B. M.; STUYVEN, E.; PIEPERS, S.; COX, E.; OPSOMER, G. Development of a method for isolating bovine colostrum mononuclear leukocytes for phenotyping and functional studies.The Veterinary Journal , v. 200, n. 2, p. 294-298, 2014. MEGANCK, V.; HOFLACK, G.; OPSOMER, G. Advances in prevention and therapy of neonatal dairy calf diarrhoea: a systematical review with emphasis on colostrum management and fluid therapy. Acta Veterinary Scandinavica , p. 56-75, 2014. MEGANCK, V.; OPSOMER, G.; PIEPERS, S.; COX, E.; GODDEERIS, B. M. Maternal colostral leukocytes appear to enhance cell-mediated recall response, but inhibit humoral recall response in prime–boost vaccinated calves. Journal of Reproductive Immunology, 2015. DOI: /dx.doi.org/doi:10.1016/j.jri.2015.11.004 MOREIN, B.; ABUSUGRA, I.; BLOMQVIST, G. Immunity in neonates. Veterinary Immunology and Immunopathology, v. 87, p. 207-213, 2002. MORRILL, K. M.; QUIGLEY, J. D.; LAGO, A.; TYLER, H. D. Estimate of colostral IgG concentration using refractometry without or with caprylic acid fractionation. Journal of Dairy Science , v. 95, p. 3987–3996, 2012. MURATA, H.; SHIMADA, N.; YOSHIOKA, M. Current research on acute phase proteins in veterinary diagnosis: An overview. The Veterinary Journal . v.168, p. 28–40, 2004. NAGAHATA, H.; HIGUCHI, H.; YAMASHIKI, N.; YAMAGUCHI, M. Analysis of the functional characteristics of L-selectin and its expression on normal and CD18-deficient bovine neutrophils. Immunology and Cell Biology , v. 78, n. 3, p. 264-271, 2000. NEMETH, E.; GANZ, T. Regulation of iron metabolism by hepcidin. The Annual Review of Nutrition, v. 26, p. 323-342, 2006.

146
NOCEK, J. E.; BRAUND D. G.; WARNER, R. G. Influence of Neonatal Colostrum Administration, Immunoglobulin, and Continued Feeding of Colostrum on Calf Gain, Health, and Serum Protein. Journal of Dairy Science , v. 67, n. 2, 1984. NOVO, S. Efeito dos leucócitos do colostro materno na respos ta imune de bezerros recém-nascidos. São Paulo, 2015. 128 f. Dissertação (Mestrado em Clínica Veterinária) – Faculdade de Medicina Veterinária, Universidade de São Paulo, São Paulo, 2015. NOVO, S. M. F.; FREITAS, R. L.; SILVA, C. P. C..; BALDACIM, V. A. P.; BACCILI, C. C.; REIS, J. F.; HAGIWARA, M. K.; GOMES, V. Hematological adaptation in Holstein calves during the neonatal period. Brazilian Journal of Veterinary Research and Animal Science, v. 52, n. 3, p. 212-216, 2015. NOVO, S. M. F.; REIS, J. F.; SOBREIRA, N. M.; GOMES, V. Effect of temperature and conservation time on the feasibility of bovine colostrum cells. In: WORLD BUIATRICS CONGRESS , 28., 2014, Cairns, Austrália. Anais... Cairns, Austrália, 2014. PETERSEN, H.; NIELSEN, J.; MIKAEL, P.; HEEGAARD, H. Application of acute phase protein measurements in veterinary clinical chemistry. Veterinary Research, BioMed Central , v. 35, n. 2, p.163-187, 2004. POULSEN, K. P.; MCGUIRK, S. M. Respirator y diseaseof the bovine neonate. Veterinary Clinics of North America: food animal pr actice , v. 25, p. 121–137, 2009. POURJAFAR, M., BADIEI, K., NAZIFI, S. NAGHIB, S. M. Acute phase response in Holstein dairy calves affected with diarrhoea. Bulgarian.Journal of Veterinary Medicine , v.14, n. 3, p. 142−149, 2011. QUIGLEY, J. D.; LAGO, A.; CHAPMAN, C.; ERICKSON, P.; POLO, J. Evaluation of the Brix refractometer to estimate immunoglobulin G concentration in bovine colostrum. Journal of Dairy Science , v. 96, p. 1148–1155, 2013. REBER, A. J.; DONOVAN, D. C.; GABBARD, J.; GALLAND, K.; ACEVES-AVILA, M.; HOLBERT, K. A.; MARSHALL, L.; HURLEY, D. J. Transfer of maternal colostral leukocytes promotes development of the neonatal immune system Part II. Effects on neonatal lymphocytes. Veterinary Immunology and Immunopathology , v. 123, n. 3-4, p. 305-313, 2008a. REBER, A. J.; DONOVAN, D. C.; GABBARD, J.; GALLAND, K.; ACEVES-AVILA, M.; HOLBERT, K. A.; MARSHALL, L.; HURLEY, D. J. Transfer of maternal colostral leukocytes promotes development of the neonatal immune system Part I. Effects on monocyte lineage cells. Veterinary Immunology and Immunopathology , v. 123, n. 3-4, p. 186–196, 2008b.

147
REBER, A. J.; LOCKWOOD, A.; HIPPEN, A. R.; HURLEY, D. J. Colostrum induced phenotypic and trafficking changes in maternal mononuclear cells in a peripheral blood leukocyte model for study of leukocyte transfer to the neonatal calf. Veterinary Immunology and Immunopathology , v. 109, n. 1-2, p. 139-50, 2006. RIEDEL-CASPARI, G. The influence of colostral leukocytes on the course of an experimental Escherichia coli infection and serum antibodies in neonatal calves. Veterinary Immunology and Immunopatology , v. 35, p. 275-288, 1993. ROARK, C. L.; SIMONIAN, P. L.; FONTENOT, A. P.; BORN, W. K.; O'BRIEN, R. L. gammadelta T cells: an important source of IL-17. Current Opinion in Immunology , v. 20, n. 3, p. 353-357, 2008. RODRIGUES, J. A. Assistência clínica materna e neonatal à diferentes condições obstétricas em bovinos da raça Holandesa , 2008. 94 f. Dissertação (Mestrado em Medicina Veterinária) - Faculdade de Medicina Veterinária, Universidade de São Paulo, São Paulo, 2008. SHIBATA, K.; YAMADA, H.; HARA, H.; KISHIHARA, K.; YOSHIKAI, Y. Resident Vdelta1+ gamma-delta T cells control early infiltration of neutrophils after Escherichia coli infection via IL-17 production. The Journal of Immunology , v. 178, p. 4466-4472, 2007. SILVA, C. P. C. Imunidade inata no sangue e secreção mamária de vac as Holandesas no período periparto. São Paulo, 2014. 142 f. Dissertação (Mestrado em Clínica Veterinária) – Faculdade de Medicina Veterinária, Universidade de São Paulo, São Paulo, 2014. STIELER, A.; BERNARDO, B. S.; DONOVAN, G. A. Neutrophil and Monocyte Function in Neonatal Dairy Calves Fed Fresh or Frozen Colostrum. International Journal of Applied Research in Veterinary Medicine , v. 10, n. 4, 2012. TORNQUIST, S. J.; RIGAS, J. Interpretation of ruminant leukocyte. In: SCHALM, O. W. Veterinary hematology . 2nd ed. Philadelphia, PA: Lea and Febiger, 2010. p. 307-312. VANNUCHCHI, C. I.; RODRIGUES. J. A.; SILVA, L. C. G.; LÚCIO, C. F.; VEIGA, G. A. L.; FURTADO, P. V.; OLIVEIRA, C. A.; NICHI, M.; Association between birth conditions and glucose and cortisol profiles of periparturient dairy cows and neonatal calves. Veterinary Record , 2015, doi: 10.1136/vr.102862. WINDEYER, M. C.; LESLIE, K. E.; GODDEN, S. M.; HODGINS, D. C.; LISSEMORE, K. D.; LEBLANC, S. J. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age. Preventive Veterinary Medicine , v.113, n. 2, p. 231-240, 2013. ZWAHLEN, R. D.; ROTH, D. R . Chemotactic Competence of Neutrophils from Neonatal calves: Functional Comparison with Neutrophils from Adult Cattle. Inflammation , v. 14, n. 1, p. 109-123, 1990.

148
13 CONCLUSÃO GERAL
A administração de colostro com (COL+) e sem células viáveis (COL-) não
influenciou na colonização da microbiota do intestino, porém a maior habilidade de
migração e produção de peróxido de hidrogênio pelos granulócitos sanguíneos das
bezerras COL+ pode justificar o melhor status sanitário desses animais,
considerando-se que as bezerras que ingeriram colostro congelado apresentaram
maior intensidade da diarreia e anemia do processos inflamatório.

149
REFERÊNCIAS
ADKINS, M. L. Ecology of transferred maternal Staphylococci and neonatal Staphylococcal immunity. In: SCIENCE OF VETERINARY MEDICINE SYMPOSIUM, 2014, Georgia: University of Georgia, 2014. AUJLA, S. J.; DUBIN, P. J.; KOLLS, J. K. Th17 cells and mucosal host defense. Seminars in Immunology , v. 19, p. 377-382, 2007. BARTIER, A. L.; WINDEYER, M. C.; DOEPEL, L. Evaluation of on-farm tools for colostrum quality measurement. Journal of dairy science , v. 98, n. 3, p. 1878-1884, 2015. BIELMANN, V.; GILLAN, J.; PERKINS, N. R.; SKIDMORE, A. L.; GODDEN, S.; LESLI, K. E. An evaluation of Brix refractometry instruments for measurement of colostrum quality in dairy cattle. Journal of dairy science , v. 93, p. 3713-3721, 2010. BRANDTZAEG, P. The mucosal immune system and its integration with the mammary glands. The Journal of Pediatrics , v. 156, n. 2, p. 8-15, 2010. CHIGERWE, M.; TYLER, J. W.; MIDDLETON, J.R, SAPAIN, J. N.; DILL, J. S.; STEEVENS, B. J. Comparison of four methods to assess colostral IgG concentration in dairy cows. Journal of the American Veterinary Medical Associat ion , v. 233, p. 761-766, 2008.
CORTESE, V. S. Neonatal immunology. Veterinary Clinics North America: food animal practice, v. 25, n.1, p. 221-227, 2009.
CROSS, M. L.; GILL, H. S. Immunomodulatory properties of milk. British Journal of Nutrition , v. 84, p. S81-S89, 2000. Supplement, 1.
GODDEN, S. Colostrum management for dairy calves. Veterinary Clinics North America: food animal practice, v. 24, p. 19- 39, 2008.
GOMES, V.; BACCILI, C. C.; SILVA, C. P. C.; GOMES, C. O. M. S. Development of specific immune system and detection of risk factors for diseases in Holstein calves. In: WORLD BUIATRICS CONGRESS, 28., 2014, Cairns, Austrália. Anais… Cairns, Austrália, 2014.
GOMES, V.; MADUREIRA, K. M.; DELLA LIBERA, A. M. M. P.; ALVES, M.; BAPTISTELLA, F.; BENESI, F. J. Dinâmica da celularidade do colostro de vacas holandesas no pós-parto imediato. Arquivo Brasileiro de Medicina Veterinária e Zootecnia , v. 63, n. 5, p.1047-1053, 2011.
GONZALEZ, D. D.; RIMONDI, A.; AGUIRREBURUALDE, M. S. P.; MOZGOVO J. M.; BELLIDO, D.; WIGDOROVITZ, A.; DUS SANTOS, M. J. Quantitation of cytokine gene expression by real time PCR in bovine milk and colostrum cells from cows immunized with a bovine rotavirus VP6 experimental vaccine. Research in Veterinary Science , v. 95, p. 703-708, 2013.

150
IVANOV, I. I.; LITTIMAN, D. R. Modulation of immune homeostasis by commensal bacteria. Current Opinion in Microbiology , v. 14, p.106–114, 2011. KEHOE, S. I.; JAYARAO, B. M.; HEINRICHS, A. J. A Survey of Bovine Colostrum Composition and Colostrum Management Practices on Pennsylvania Dairy Farms. Journal of dairy science , v.90, n. 9, p. 4108-4116, 2007. KOBOZIEV, I.; WEBB, C. R.; FURR, W. B. K.; GRISHAM, M. B. Role of the enteric microbiota in intestinal homeostasis and inflammation. Free Radical Biology and Medicine G8 , p. 122-133, 2014. KORHONEN, H.; MARNILA, P.; GILL, H. S. Milk immunoglobulins and complement factors. British Journal of Nutrition , v. 84, n. 1, p. 75-80, 2000. KRUSE, P. E. The importance of colostral immunoglobulins and their absorption from the intestine of the newborn animals. Annales de Recherches vétérinaires , v. 14, n. 4,p. 349-353, 1983. LANGEL, S. N.; WARK, W. A.; GARST, S. N.; JAMES, R. E.; McGILLIARD, M. L.; PETERSSON-WOLFE, C. S.; KANEVSKY-MULLARKY, I. Effect of feeding whole compared with cell-free colostrum on calf immune status: The neonatal period. Journal of Dairy Science , v. 98, n. 6, p. 3729-3740, 2015. LARSON, B. L.; HEARY, H. L.; DEVERY, J. E. Immunoglobulin Production and Transport by the Mammary Gland. Journal of Dairy Science , v. 63, n. 4, p. 665-667, 1980. LIEBLER-TENORIO, E. M.; RIEDEL-GASPARI, G.; POHLENZ, J. F. Uptake of colostral leukocytes in the intestinal tract of newborn calves. Veterinary Immunology and Immunopathology , v. 85, n.1-2, p. 33-40, 2002. MANDYLA, H.; XANTHOU, M.; MARVELIAS, C.; BAUM, D.; MATSANIOTIS, N. Antibody dependent cytotoxicity of humam colostrum phagocytes. Pediatrics Research , v.16, p. 995-999, 1982. McDONALD, J. S.; ANDERSON, A. J. Total and differential somatic cell counts in secretions from no infected bovine mammary glands: the peripartum period. American Journal of Veterinary Research , v.45, p.1.366-1.368, 1981. MCGUIRK, S. M.; COLLINS, M. Managing the producton, storage, and delivery of colostro. Veterinary Clinics North America , v. 20, p. 593-603, 2004. MEGANCK, V.; GODDEERIS, B. M.; STUYVEN, E.; PIEPERS, S.; COX, E.; OPSOMER, G. Development of a method for isolating bovine colostrum mononuclear leukocytes for phenotyping and functional studies. The Veterinary Journal , v. 200, n. 2, p. 294-298, 2014.

151
MEGANCK, V.; OPSOMER, G.; PIEPERS, S.; COX, E.; GODDEERIS, B. M. Maternal colostral leukocytes appear to enhance cell-mediated recall response, but inhibit humoral recall response in prime–boost vaccinated calves. Journal of Reproductive Immunology, 2015. DOI: /dx.doi.org/doi:10.1016/j.jri.2015.11.004 MOWAT, A. M. Anatomical basis of tolerance and immunity to intestinal antigens. Nature Reviews Immunology, v, 3, p. 331–341, 2003. NATIONAL ANIMAL HEALTH MONITORING SYSTEM. National dairy health evaluation project. Dairy heifer morbidity, mortali ty, and health management focusing on preweaned heifers. Ft. Collins (CO): USDA-APHIS Veterinary Services, 1996.
PAULA, M. R. Avaliação da saúde e desempenho de bezerros leiteir os recebendo suplemento de colostro associado ao colos tro materno . 2016. 111 f. Tese (Doutorado em Ciêncas) – Escolar Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo, São Paulo. 2016.
PEREZ, P. F.; DORÉ, J.; LECLERC, M.; LEVENEZ, F.; BENYACOUB, J.; SERRANT, P.; ROGGERO, I. S.; SCHIFFRIN, E. J.; HUGHES, A. D. Bacterial imprinting of the neonatal immune system: lessons from maternal cells? Pediatrics , v.119, n. 3, p. 724-732, 2007. QUIGLEY, J. D.; LAGO, A.; CHAPMAN, C.; ERICKSON, P.; POLO, J. Evaluation of the Brix refractometer to estimate immunoglobulin G concentration in bovine colostrum. Journal of Dairy Science , v. 96, p. 1148–1155, 2013. REBER, A. J. The influence of the colostral environment on the c apacity of maternal leucocytes to cross intestinal epithelium and modulate the neonatal immune system. (2000) 103 f. (PhD Dissertation) Paper 267. South Dakota State University, 2000. REBER, A. J.; DONOVAN, D. C.; GABBARD, J.; GALLAND, K.; ACEVES-AVILA, M.; HOLBERT, K. A.; MARSHALL, L.; HURLEY, D. J. Transfer of maternal colostral leukocytes promotes development of the neonatal immune system Part II. Effects on neonatal lymphocytes. Veterinary Immunology and Immunopathology , v. 123, n. 3-4, p. 305-313, 2008a. REBER, A. J.; DONOVAN, D. C.; GABBARD, J.; GALLAND, K.; ACEVES-AVILA, M.; HOLBERT, K. A.; MARSHALL, L.; HURLEY, D. J. Transfer of maternal colostral leukocytes promotes development of the neonatal immune system Part I. Effects on monocyte lineage cells. Veterinary Immunology and Immunopathology , v. 123, n. 3-4, p. 186–196, 2008b. REBER, A. J.; LOCKWOOD, A.; HIPPEN, A. R.; HURLEY, D. J. Colostrum induced phenotypic and trafficking changes in maternal mononuclear cells in a peripheral blood leukocyte model for study of leukocyte transfer to the neonatal calf. Veterinary Immunology and Immunopathology , v. 109, n. 1-2, p. 139-50, 2006. REITER, B. Rewiew of the progress of dairy Science: antimicrobial system in milk. Journal of Dairy Research , v. 45, p. 131-147, 1978.

152
RESCIGNO, M. The intestinal ephitelial barrier in the control of homeostasis and immunity. Trends in Immunology , v.32, n. 6, p. 256-264, 2011. RIEDEL-CASPARI, G. The influence of colostral leukocytes on the course of an experimental Escherichia coli infection and serum antibodies in neonatal calves. Veterinary Immunology and Immunopathology , v. 35, p. 275-288, 1993. RIEDEL-CASPARI, G.; SCHMIDT, F. W. The influence of colostral leukocytes on the immune system of the neonatal calf. I Effects on lymphocyte responses. Deutsche Tierärztliche Wochenschrift , v. 98, p. 77-116, 1991a. RIEDEL-CASPARI, G.; SCHMIDT, F. W. The influence of colostral leukocytes on the immune system of the neonatal calf. II Effects on passive and active immunization. Deutsche Tierärztliche Wochenschrift , v. 98, p. 165-204, 1991b. SARTOR, R. B. Microbial influences in inflammatory bowel diseases. Gastroenterology , v.134, p. 577–594, 2008. SILVA, C. P. C. Imunidade inata no sangue e secreção mamária de vac as Holandesas no período periparto. São Paulo, 2014. 142 f. Dissertação (Mestrado em Clínica Veterinária) – Faculdade de Medicina Veterinária, Universidade de São Paulo, São Paulo, 2014. TODHUNTER, D. A.; SMITH, l. K.; SCHOENBERGER, P. S. In Vitro Growth of Mastitis Associated Streptococci in Bovine Mammary Secretions. Journal of Dairy Science , v. 68, n. 9, p. 2337-2346, 1985. TUBOLY, S.; BERNATH, S.; GLAVITS, R.; KOVACS, A.; MEGYERI, Z. Intestinal absorption of colostral lymphocytes in newborn lambs and their role in the development of the immune status. Acta Veterinaria Hungarica , v. 43, p. 105-115, 1995. TUBOLY, S.; BERNATH, S.; GLAVITS, R.; MEDVECZKY, I.;. Intestinal absorption of colostral lymphoid cells in newborn piglets. Veterinary Immunology and Immunopathology , v. 20, p. 75-85, 1988. VERHASSELT, V. Neonatal tolerance under breastfeeding influence. Current Opinion in Immunology , v. 22, p. 623-630, 2010. WATSON, D. L. Immunological functions of the mammary gland and its secretion-comparative review. Australian Journal of Biological Sciences , v. 33, n. 4, p. 403-422, 1980. WEAVER, D. M.; TYLER, J. W.; VANMETRE, D. C.; HOSTETLER, D. E.; BARRINGTON, G. M. Passive Transfer of Colostral Immunoglobulins in Calves. Jornal of Veterinary Internal Medicine , v.14, p. 569–577, 2000. WILLIAMS, P. P. Immunomodulating effects of intestinal absorbed maternal colostral leukocytes by neonatal pigs. Canadian Journal of Veterinary Research , v. 57, p.1-8, 1993.

153
ANEXOS
ANEXO A –TSA (Tryptic Soy Agar)
Meio de agar TSA .................................................................................................. 16 g
Água destilada q.s.p.......................................................................................... 400 mL
Colocar em um Erlenmeyer o pó e água destilada no fogo e homogeneizar até
dissolver completamente. Autoclavar por 15 minutos a 121°C, esperar esfriar e
distribuir nas placas.
ANEXO B – Agar MacConkey
Meio de agar MacConkey ..................................................................................... 48 g
Água destilada q.s.p.......................................................................................... 800 mL
Colocar em um Erlenmeyer o pó e água destilada no fogo e homogeneizar até
dissolver completamente. Autoclavar por 15 minutos a 121°C, esperar esfriar e
distribuir nas placas.
ANEXO C – Agar Sangue de carneiro 5%
Meio de agar sangue ............................................................................................. 32 g
Água destilada q.s.p.......................................................................................... 800 mL
Colocar em um Erlenmeyer o pó e água destilada no fogo e homogeneizar até
dissolver completamente. Autoclavar por 15 minutos a 121°C, esperar esfriar até
aproximadamente 50ºC e adicionar o sangue ovino 5%-8% desfibrinado e estéril. Em
seguida, distribuir nas placas.

154
ANEXO D – Salmonella-Shigella (SS)
Meio Salmonella-Shigella......................................................................................... 6 g
Água destilada q.s.p.......................................................................................... 100 mL
Colocar em um Erlenmeyer o pó e água destilada no fogo e homogeneizar até
dissolver completamente. Autoclavar por 15 minutos a 121°C, esperar esfriar e
distribuir nas placas.
ANEXO E – Tetrationato
Meio Tetrationato …............................................................................................ 4,60 g
Água destilada q.s.p.......................................................................................... 100 mL
Colocar em um Erlenmeyer o pó e água destilada no fogo e homogeneizar até
dissolver completamente. Aquecer até ferver e esperar esfriar até aproximadamente
60°C. Em seguida, distribuir 10mL da solução em tubos estéreis de vidro compridos
com tampa de rosca.
ANEXO F – Iodo-Iodeto
Cristais de Iodo ….................................................................................................... 6 g
Iodeto de Potássio .................................................................................................. 5 g
Água destilada q.s.p............................................................................................ 20 mL
Misturar e armazenar em frasco de vidro âmbar.

155
ANEXO G – Preparo da solução 100mM EDTA
EDTA sal dissódico (C10H14N2O8Na2.2H2O 372,24mM)................................. 3,722g
Água destilada.....................................................................................................100mL
Homogeneizar até dissolver completamente o sal e colocar autoclave
ANEXO H – PBS 10x (Concentrado)
Na2HPO47H2O.....................................................................................................26,79g
NaH2PO4H2O.........................................................................................................4,14g
NaCl.........................................................................................................................82g
Água destilada...................................................................................................1000mL
PBS uso: 1 PBS 10x: 9 de água destilada
ANEXO I – Soluções utilizadas no protocolo de Fagoc itose e Produção de H 2O2
DCFH – Solução de estoque – 25 mM Pó comercial 24,35 mg Etanol 2 mL Proteger da luz – Manter em freezer -20ºC DCFH – Solução de uso Solução de estoque 100 µL PBS 8,2 mL Adaptar a quantidade da solução a ser preparada, para a quantidade necessária apenas para o uso imediato, respeitando as mesmas proporções. EDTA 3 mM EDTA (C10H14N2O8Na22H2O-PM 372,24 g/mol) 1,12g PBS (1x) 1000 mL Manter refrigerado.

156
ANEXO J – PBS 2x (Concentrado)
PBS 10x Concentrado (Anexo H) ...................................................................... 200mL
Água Milli Q.........................................................................................................800mL




















