
J. Physiol. (I949) I09, 380-39I - NCBI
13
380 J. Physiol. (I949) I09, 380-39I 6I2.OI4.46I:547.96 THE MODE OF REACTION OF INTERSTITIAL CONNECTIVE TISSUE WITH WATER BY T. D. DAY FProm the Department of Pathology, St Thomas's Hospital Medical School, London (Received 15 November 1948) When water was injected into the subcutaneous tissues of a freshly killed animal, it was observed to accumulate around the point of injection (Ranvier's 'Boule d'cedeme'). Other fluids, such as alcohol, did not localize in this way, but spread evenly and rapidly into the surrounding tissues. These facts suggested a peculiar avidity of the tissue for- water. Examination of the swollen tissue with the dark ground microscope showed separation, without visible alteration, of the collagen fibrils. For this reason, it was considered probable that the water had combined with a substance which lay between the fibrils rather than with the fibrils themselves (Day., 1948). A substance of this kind had been previously described (Day, 1947 a), and Flemming's observation (1876), that, in acid solutions of strength sufficient to cause the collagen fibres to swell, this substance became shrunken, was confirmed. The present investigation is concerned with those changes in the morphology of interstitial connective tissue which took place, in aqueous solutions, as the result of experimental variation of pH and of neutral salt concentration. The study was undertaken with the object of gaining further knowledge of the mode of reaction of this tissue with water with particular regard to the part played by the interfibrillary substance. METHODS The material studied was the deep fascia immediately covering the ventral aspect of the thigh muscles of the rat. Pieces up to 1 cm. wide were removed from a freshly killed animal. Each was placed at once in at least 20 ml. of the fluid in which it was to be studied. The pH of the fluids was measured electrometrically, with a glass electrode apparatus, at the beginning and at the end of the period of immersion of the tissue. The pH of the fluids was varied by the addition of dilute HCI or NaOH. Where solutions of salts were concerned, the quantity of Na+ or Cl- so added, was so very small in comparison with the concentrations of the salt ions already present that they could be ignored. Usually, the tissue was left in the fluids overnight and at a room temperature between 18 and 200C. The changes which took place were observed with the naked eye and also microscopically using dark ground illuimination. A Leitz 'cardioid' type of condenser was employed in conjunction with a Pointolite illuminant.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of J. Physiol. (I949) I09, 380-39I - NCBI

380
J. Physiol. (I949) I09, 380-39I 6I2.OI4.46I:547.96
THE MODE OF REACTION OF INTERSTITIALCONNECTIVE TISSUE WITH WATER
BY T. D. DAY
FProm the Department of Pathology, St Thomas's Hospital Medical School, London
(Received 15 November 1948)
When water was injected into the subcutaneous tissues of a freshly killedanimal, it was observed to accumulate around the point of injection (Ranvier's'Boule d'cedeme'). Other fluids, such as alcohol, did not localize in this way,but spread evenly and rapidly into the surrounding tissues. These facts suggesteda peculiar avidity of the tissue for- water. Examination of the swollen tissuewith the dark ground microscope showed separation, without visible alteration,of the collagen fibrils. For this reason, it was considered probable that thewater had combined with a substance which lay between the fibrils rather thanwith the fibrils themselves (Day., 1948). A substance of this kind had beenpreviously described (Day, 1947 a), and Flemming's observation (1876), that,in acid solutions of strength sufficient to cause the collagen fibres to swell, thissubstance became shrunken, was confirmed.The present investigation is concerned with those changes in the morphology
of interstitial connective tissue which took place, in aqueous solutions, as theresult of experimental variation of pH and of neutral salt concentration. Thestudy was undertaken with the object of gaining further knowledge of the modeof reaction of this tissue with water with particular regard to the part playedby the interfibrillary substance.
METHODS
The material studied was the deep fascia immediately covering the ventral aspect of the thighmuscles of the rat. Pieces up to 1 cm. wide were removed from a freshly killed animal. Each wasplaced at once in at least 20 ml. of the fluid in which it was to be studied. The pH of the fluidswas measured electrometrically, with a glass electrode apparatus, at the beginning and at the endof the period of immersion of the tissue. The pH of the fluids was varied by the addition of diluteHCI or NaOH. Where solutions of salts were concerned, the quantity of Na+ or Cl- so added, wasso very small in comparison with the concentrations of the salt ions already present that theycould be ignored. Usually, the tissue was left in the fluids overnight and at a room temperaturebetween 18 and 200C.The changes which took place were observed with the naked eye and also microscopically using
dark ground illuimination. A Leitz 'cardioid' type of condenser was employed in conjunction witha Pointolite illuminant.

HYDRATION OF CONNECTIVE TISSUE
RESULTS
Changes re8ult$ng from variation in pH alone(a) Pieces of fascia were put into weak alkali at pH 11i1. They became at
first curled up, white and opaque. Within a few minute the edges became grey.This process spread inwards so that within an hour each piece had becomeentirely grey, transparent and greatly swollen.Dark ground examination showed broadening of the bundles of collagen
fibrils, the majority measuring from 10 to 20,m. across (P1. lb). The bundlesthemselves were separated by spaces up to 100 . wide. Their constituent fibrilswere also separated and, for the most part, quite distinct from each other.They were not swollen.
Pieces were then transferred to weak acid at pH 4-3. Within a few minutes,white opaque areas appeared in them.'' After half an hour, each was entirelyopaque and also markedly shrunken. All parts of it clung together andresisted disintegration by teasing. Microscopically, it was seen that the collagenbundles had become knotted together and superimposed to such an extentthat no spaces could be observed between them. The bundles were greatlynarrowed (mostly less than 5 IL. wide) and remarkably crimped and coiledup. It was impossible to distinguish individual fibrils within the condensedbundles.By placing pieces of tissue alternately in weak acid and alkali, within the
limits of pH 4*5-11*0, the processes of shrinkage and swelling were repeatedseveral times. But after 24 hr. the tissue responded more slowly and the effectswere less pronounced.
(b) Pieces of fascia were treated overnight at a series of pH values rangingfrom 3 0 to 12-0. The initial and final pH value of the fluids is recorded in theorder given. The changes which took place were as follows:pH 3-8-3-8. Tissue transparent and swollen. Microscopically, considerable
swelling ofthe collagen bundl6s up to 50 ,. in widthwith completedisappearanceof outline of their constituent fibrils.pH 4-54*7. Tissue white, opaque, very shrunken and hard. Microscopically,
collagen bundles narrowed to less than 5,. wide, very crimped and knottedtogether. No spaces between them other than those produced artificially byteasing.-pH 5*05-7. Tissue opaque, grey-white, and slightly shrunken. Micro-
scopically, collagen bundles narrow and condensed to less than 5 U. wide, butwell separated from each other (P1. la).pH 7-67-3. Tissue grey-white and semi-opaque. Microscopically, collagen
bundles mostly from 10 to 5 ,u. wide. Their individual fibrils somewhat com-pressed but distinguishable from each other. Bundles well separated byapparently empty spaces.
381

pH 112-10 9. Tissue grey and transparent. Microscopically, collagen bundlesmostly from 20 to 10 ,u. wide, their constituent fibres being definitely distin-guishable and in some bundles exhibiting a continuous shimmering vibration.Bundles themselves also well separated (P1. 1 b).pH 11-7-11-5. Tissue transparent and swollen. Microscopically, considerable
sweling of collagen bundles, with swelling and disappearance of outline oftheir constituent fibrils.
It is to- be concluded that variation in external pH between 4-5 and llO wasaccompanied by changes in the volume of the tissue to which neither the fibresnor the cells appeared to contribute. Increase in acidity caused shrinkage, anddecrease caused swelling of the tissue. Swelling was accompanied by separationof the collagen bundles and fibrils. With shrinkage these structures becamedrawn together. These changes were reversible. Swelling which occurred atpH values immediately outside this range was of a different character, inasmuchas it manifestly involved the collagen fibrils themselves.
Changes resulting from variation in neutral salt concentration alonePieces of fascia were treated overnight in approximately neutral solutions of
NaCl at the following concentrations: 4-0, 2-0, 1-0, 0-5, 0-25, 0-125, 0'1, 0 05,0-025, 0-012M. One further piece was left in neutral distilled water. The pH ofall the fluids used, measured before and after these periods of immersion of thefascia, lay between 8-3 and 6*2. The majority of the values were closer toneutrality. The pieces were examined and then transferred to neutral distilledwater for a further period of 18 hr. They were then examined again.
It was found that the change which took place in concentrations of NaClbetween 2-0 and 0-125m was identical with that which occurred in weak alkali.The tissue became swollen and this swelling was accortpanied by separation,without increase in size, of collagen fibrils. The change was reversed by treat-ment with neutral distilled water.The concentration of NaCl in which the change began to take place was
critical. It was observed in a concentration of 0-125M but not in one of 01 M.The pieces which had been treated in solutions of 0 1M strength and under didnot differ in either naked eye or microscopic appearance from that which hadbeen treated with neutral distilled water alone.The change which took place in a 40M solution was not completely reversible
and differed in other respects from that which was seen in the lower con-centrations.
Changes resultingfrom variation in both neutral salt concentration and pHThe following concentrations of NaCl were prepared: 1'0, 0'5, 0.25, 0-125,
0*062M. Six test-tubes were filled with solution at each of these concentrations.Six further tubes were filled with distilled water. The fluid in one out of each
.382 T. D. DAY

HYDRATION OF CONNECTIVE TISSUEgroup of six tubes was adjusted as nearly as possible to one of the following pHvalues: 3-0, 4-0, 5-0, 7-0, 11-0 and 12-0. The precise pH value of the fluid in eachof the forty-two tubes was then measured and a piece of fascia inserted. Thetubes were left overnight at a temperature of between 18 and 200 C. The pHvalue of the fluids was then measured again. The naked eye and microscopicappearance of each piece was then examined.The results of this experiment are recorded in Table 1. The appearances of
the tissue are represented by the letters 0, T, SO and C. 0, T and SO indicatenaked eye changes which contrasted with each other to an extent which quiteexcluded subjective error. Microscopically these changes were seen to beaccompanied by alteration in the spatial relationships of the collagen fibrils toeach other, but by no visible change in the fibrils themselves. The letter C, onthe other hand, indicates swelling of the tissue accompanied by visible swellingof collagen fibrils.
TABLE 1. Changes in rat fascia after immersion in aqueous solutions ofsodium chloride of varied strength and at different pH values
Approx. pHNaCl con-centration 3 4 5 7 11 12
M 0 0 T T T CM/2 0 0 T T T CM/4 C 0 0 T T CM/8 C 0 0 T T CM/16 C 0 O SO T CWater ' C 0 0 so T C
0=Opacityandshrinkage. T=Transparencyandswelling. SO=Semi-opacitywithoutnotablechange in size. C =Swelling of oollagen fibres. Time of immersion= 18 hr. Temperature, 20-18o C.
From the evidence summarized in Table 1 two main conclusions were drawn:(1) That at pH values in the region of 4-0, salt, even in molar concentration,had no swelling action on the tissue. (2) That with variation of pH from 4-0 to11-0, the concentration ofsalt necessary to cau'se swellingbecame proportionatelyless.
Comparison of the effects of different neutral saltsIt has been mentioned both here and previously (Day, 1947 a) that, in certain
circumstances, collagen fibrils may exhibit a continuous shimmering vibration.The vibration has been observed to continue for several days in the samepreparation and is in the nature of Brownian movement. It was only seen intissue which had become swollen and appeared to be related to the degree ofseparation of the fibrils.
It was found that, under standard conditions of temperature, illumination,and pH, the lowest concentration in which Brownian movement of collagenfibrils could just be detected, was always the same for each particular salt. Ifit is assumed that the occurrence of this phenomenon is a measure of a certain
383

degree of inteifibrillary hydration of the tissue, amessment of this minimalconcentration, under standard conditions, may be expected to provide a meansof comparing the hydrating capacity of each of a series of salts.
It is to be emphasized, however, that the swelling effect of salts on the tissuecan be seen with the naked eye. Where, as in the case of NaCl, and Na3C.H507,the salts differ greatly in their hydrating capacity, this difference may easily beobserved by mere naked eye inspection. The value of Brownian movement, asa criterion of a precise. degree of hydration, was mainly in the comparison ofsaltts between which the difference in hydrating capacity was slight.
TABLz 2. The lowest concentrations of neutral salts in which Brownian movement of thecollagen fibres was observed compared with the nature of their ions
pH value of solution at Lowest dilution in whichbeg_nnin and end of period Brownian movement of Valency of
Salt of immersion of tissue collagen fibres was observed anionNaCl 9'86-9*30 M/7 1Na2SO4 9*46-8870 M/16 2Na3C6HO7 8-32-8*12 M/25 3
NaFNaClNaBrNal
LiC1NaClKlCRbCl
642-6*-546*20-6-206-306-306-08-607
M/3M/6m/7M/8
6-40"6-90 x/46*20A-20 M/65-84-5-84 M/7560-5*54 M/8
Time of immersion, - 3k hr. Temperature 200 C.
Length ofcrystal radiusof anion (A)
1-361*811*952-16
Length ofcrystal radius
of kation0-600951*331*48
The salts were compared in groups, each group being investigated on thesame day. The pH values of the dilutions of salts in each group were adjusted,as nearly as possible, to the same figure by the addition of weak HCI or NaOHdiluted in a solution ofthe appropriate salt. The experiments were carried out atroom temperature adjusted to 200 C. Pieces were left for at least 31 hr. It wasfound that Browman movement could be seen best in bundles which had beendissociated from the main mass of the tissue. After removal from the tube,each piece was, therefore, gently teased before being mounted under a cover-slip in a drop of the solution in which it had been lying. The precise pH valuesof the solutions were measured at the beginning and at the end of the period ofimmersion of the tissue. Each preparation was examined as soon as it had beenmade, using aA objective and a x 10 eyepiece. The same source of light wasused throughout and was adjusted to maximum intensity for each observation.The results of this experiment are recorded in Table 2.
384 T. D. DAY

HYDRATION OF CONNECTIVE TISSUE
The effect of neutral salts compared wirth that of urea and of glycinePieces were placed in 1.0, 0 5, 0*25, 0-125, 0-062m solutions of urea. They
were left for 6 hr. at a temperature of 200 C. The pH values of the solutionsranged between pH 5*4 and 6-8. No appearance was observed in any of thepieces other than would be expected, between these pH values, in water alone.
Pieces of fascia were treated over the same period with the same range ofconcentration of glycine. The pH values of the solutions remained at pH 6-0throughout the period ofthe experiment. No change in the tissue was observedwhich could be attributed to the presence of glycine.
DISCUSSION
The swelling and shrinkage which takes place in fascial connective tissue as theresult of variation in external salt concentration and of pH between the values4-5 and 11.0, has been shown to be related to the degree of separation of thecollagen fibrils. It is probable, therefore, that these changes occur in theinterfibriary substance. Other possibilities must, however, be considered.
In the first place the changes may be due, at least in part, to alteration of thefibrils themselves, a possibility suggested by the extreme cnrmping which theywere seen to undergo around pH 4-5-5-0. Against this is the observation (Day,1947a) that isolated fibrils from tendon in a rat's tail, which are not associatedwith interfibrillary substance, do not crimp in weak acid. It might be arguedthat the fibrils of tendon and fascia have a different composition. H6wever,both types of fibril become swollen in approximately the same minimal strengthsof acid or alkali and their staining reactions and appearance in stained sectionsare similar. It would seem unlikely, therefore, that the fibrils of fascia andtendon differ sufficiently in composition as to explain this marked contrast inbehaviour.A further possibility is that the fibrils are not joined together into bundles
by an interfibrillary substance but owe their cohesion entirely to physico-chemical forces. On this supposition, the separation of the fibrils which hasbeen observed at certain ionic concentrations is due to weakening or abolitionof these cohesive forces. Such a process would, however, be expected to resultfinally in the complete separation ofthe fibrils and in permanent disorganizationof their arrangement in bundles. It has been shown that this result is notcharacteristic of the action of either neutral salts or weak alkali. Even in tissuewhich has become very greatly swollen, the fibrils still remained arranged inbundles and maintained a fixed relationship to each other: furthermore, thechange was reversible so that with removal of the external conditions whichdetermined swelling, the fibrils in each bundle came together again.The action of trypsin, which digests the interfibrillary substance, leads to
quite different effects.- Individual fibrils then become entirely dissociated from
385

each other and the spaces between fibrils can no longer be made to vary in sizewith change in ionic environment. Moreover, the change produced by trypsinis irreversible and leads to permanent disorganization of the tissue.A point of some interest is that tryptic digestion of the interfibrillary sub-
stance does not lead to the appearance of Brownian movement of the fibrils,even when transferred to conditions which would have brought about thisphenomenon in fresh tissue. Fibrils which become completely isolated andwhich lie suspended free in the fluid, occasionally show a sluggish movementbut the vigorous shimmering vibration described above was not seen. Itwould appear that this vibration is not a property of the fibrils themselves butdepends upon their association with the interfibrillary substance.On the whole, there would seem to be good grounds to suppose that the
changes in rat fascia described above, are mainly due to the presence in it of aninterfibrillary substance. It is proposed, therefore, to consider the nature ofthis substance more fully.The nature ofthe interfibrillary substance. In spite of its hydrophilic character,
it is impossible to leach the substance from the tissue by prolonged perfusionwith water (Day, 1948). Above, it has also been shown that the extremedegrees of swelling and shrinkage, which have been produced experimentally,are reversible and could be repeated several times without any disorganizationof the tissue detectable with the dark ground microscope. Extreme hydrationofthe substance does not, therefore, lead to its solution. In view ofthe obviouslycolloidal nature of the substance, this fact can best be explained by assumingthat the non-aqueous component of the colloid must exist as a continulimrather than as separate particles.The structured nature of the substance may also be inferred from histological
evidence. In certain circumstances it may be altered in such a way as to beeasily visible with the dark ground microscope. It then appears in the form ofmeImbranes of extreme thinness containing very delicate, branching fibrils(Day, 1947 b).The assumption, that the substance consists of molecules held together in
a continunm, also implies the existence in it of chemical linkages of a kidcompatible with polymerization. It is of considerable interest, therefore, thatit can be digested by trypsin (Day, 1947 a), and that the contraction of thesubstance which followed treatment either with 60% ethyl alcohol-or byadjustment of eexternal pH to values around 4-0, greatly accelerated thedigestion (Day, 1948). Since trypsin specifically attacks peptide linkages,these observations strongly suggest that it is by these that the molecules of thecontinuum are joined together.
It was observed that, over the entire range studied, the interfibrillary sub-stance responded by change in volume to slight variations in external pH. Forthis reason the microscopic appearance of the tissue was quite typical for any
386 T. D. DAY

HYDRATION OF CONNECTIVE TISSUEparticular pH value. This would imply that its structure may be modified byvery small forces. In contrast with fibres, therefore, its internal scaffoldingmust be considered to be weak. The fact that the substance may readily betransformed into visible membranes suggests that the continuum of molecules,which has been postulated, may be arranged two-dimensionally: an arrangementto which an extensible trellis would seem to provide a suitable analogy. It isclear that a two-dimensional structure of this kind would not afford such goodopportunity for the formation of intermolecular linkages as does the lineararrangement of molecules in a fibre. It is to be concluded that the inter-fibrillary substance belongs essentially to the class of structured proteins. Butthis conclusion does not exclude the possibility that molecules of a non-proteinnature may be articulated in some fashion to the main framework.
The non-participation of collagen fibrils in the changes described. It has beenshown that pieces of fascia underwent considerable changes in volume whenimmersed in aqueous solutions of varied pH and salt content. In all con-centrations of salt, and at pH values between 4-5 and 11.0, these changes couldnot be accounted for by variation in the size of the collagen fibrils. With slightvariation of pH beyond this range, the swelling of the fibrils was, however,obvious and practically maximal.The forces which lead to the swelling of a collagen fibril arise as the result
of ionization of its protein molecules (Lloyd & Shore, 1938, ch. xi) andare therefore proportional to variation in pH, relative to the isoelectric pointof collagen. The fact that swelling can only take place at pH values outsidea certain range suggests that, within this range, the forces which cause swellingare counterbalanced by those of cohesion.Astbury (1933, pp. 35-6) has vividly described the way in which cohesion
between molecules increases in proportion to their length. Collagen fibresare built up of long molecules arrainged in a more or less parallel fashion(Lloyd, 1941). This arrangement not only facilitates the formation of laterallinkages between adjacent molecules but also enormously increases theircohesive effect. Even as weak a link as the hydrogen bond is considered tocontribute very greatly to the structural stability of a collagen fibre on accountof the parallel orientation of the long molecules between which it developsLloyd, 1941, p. 282). On purely theoretical grounds, therefore, the internalscaffolding of a collagen fibril is likely to be strong and it is to its strengththat the observed non-participation of collagen fibrils in swelling changesmust largely be due.The essentially colloidal nature of interstitial connective tissue has been
emphasized by Schade (1913, 1927) and by Schade & Menschel (1922). Theseworkers recognized that the cells in this tissue were too sparse to contributesignificantly to the great degree of swelling of which it is capable. They con-sidered that swelling was due to physico-chemical changes in an extracellular
PH. CIX. 25
387

' colloid mass' (Schade, 1927) consisting of fibres and an undifferentiated groundsubstance. Swelling of the fibres and of the ground substance was deter-mined by precisely opposite conditions. This finding led to the conclusionthat the water content of connective tissue was controlled by a swellingantagonism-' Quellungsantagonismus '-between these two main colloidalcomponents.
This conclusion is not supported by the observations which have beenrecorded here. The ranges ofpH and of salt concentration in which no swellingof collagen fibrils was observed both included and exceeded such concentrationsas are likely to be present in living tissue. It is clear, therefore, that as far asthe living animal is concerned, Schade's thesis of a swelling antagonism betweenfibres and interfibrillary substance cannot be sustained.
Changes in the substance due to variation in pH. It was observed that freshpieces of fascia, when put into different aqueous solutions, immediatelycontracted. This contraction occurred irrespectively of the composition of thesolutions and must be presumed to be due to the retraction of elastic fibres.In alkaline solutions this contraction was gradually overcome so that in abouthalf an hour the pieces were greatly swollen. In acid, on the other hand, thecontraction was intensified to such an extent that dissociation of the tissuewith a needle was almost impossible. These observations suggest that theswelling and contraction of the tissue which results from change in external pHis accompanied by the development of considerable mechanical force. It isproposed to consider how such forces may be generated.
It has been concluded that the interfibrillary substance is essentially a con-tinuum of protein molecules. It has been shown (Day, 1947 a) that it becamemaximally constricted at external pH values between 4x2 and 3x8. It is there-fore probable that the isoelectric point of these molecules lies between thesevalues. The influence of variation in pH upon the hydration of a system ofthis kind has been treated theoretically by Lloyd & Phillips (1933). Thus(p. 137) 'The molecules of an organized protein can approach as closely as thelength of their side chains permits. At the isoelectric point, when each moleculeof the structure contains anionic and kationic centres, they will tend to bedrawn together by electrostatic attraction which will favour the closest degreeof packing and hence the lowest degree of hydration. On either side of theisoelectric point each protein molecule contains predominantly either anionicor kationic centres and the molecules mutually repel each other. By this means,the spaces between the molecules in an organized protein are increased, leadingto an increase in the amount of water which can enter freely between theindividual molecules.'
Swelling, however, is assisted by other factors. Repulsion of like chargesmust lead to isolated charged centres around which, on account of their dipolarnature, water molecules will become oriented. Furthermore, the outer surface
388 Ts. D. DAY

HYDRATION OF CONNECTIVE TISSUEof the piece of tissue, as its swells, must be considered as a semipermeablemembrane, since the protein molecules, being joined together, cannot passbeyond it. This the diffusible ions, which increase in number proportionatelyas the external pH is removed from the isoelectric point, are certainly ableto do. In accordance with the theory first elaborated in relation to theswelling of gelatin by Procter (Lloyd & Shore, 1938, ch. xI) the resultingelectrostatic inequality must lead to the further indrawing of water into thesystem.
Fundamentally, therefore, the forces which lead to swelling and shrinkageof the tissue are electrostatic: they are generated as the result of ionization orsuppression of ionization of the multipolar molecules of which the interfibrillarysubstance must be considered to be composed.
Swelling caused by neutral salts. It has been shown that the non-electrolyteurea and the dipolar ion glycine did not cause the tissue to swell. This fact alonesuggests that the swelling effect of neutral salts is due to their existence as freeions. In 1905 J. Mellanby wrote: 'Solution of globulin by a neutral salt is dueto forces exerted by its free ions. Ions with equal valencies, whether positive ornegative, are equally efficient, and the efficiencies of ions of different valenciesare directly proportional to the squares of their valencies.' It is probable thatit is to precisely the same interionic forces which Mellanby has shown to causesolution of globulin, that the hydration of the interfibrillary substance is due.The minimal concentration of NaCl necessary to cause visible swelling was
found to be critical. It is probable that here, as in the case of the swelling ofcollagen fibrils, visible change can only occur when the forces which promoteswelling have overcome those of internal cohesion; that is to say when thestrength of the forces exerted by the free ions in the solution is sufficient. Thedegree of cohesion within the interfibrillary substance has been shown to varysensitively with external pH. The observed relationship between the concentra-tion of NaCl necessary to cause swelling and the prevailing pH (Table 1) istherefore to be expected.Comparison of the capacities of NaCl, N%2S04and Na3C6H507, to bring about
the same degree of interfibrillary hydration (Table 2), revealed a relationshipbetween the effective concentration of each salt and the valency of its anion.Although the minimal effective concentrations of these salts differed con-siderably (0.14, 0-06, 0-04M), in the order given, the ionic strengths of theseconcentrations, as calculated from u= E c1v2 (where ,t is the ionic strength,cl the actual concentration of each ion, and v, its valency), lay much closertogether (0-14, 0-18, 0 24).The observed relationship between the effective concentration of a salt and
the valency of its anion would suggest, therefore, that the capacity of a neutralsalt solution to cause interfibrillary hydration is determined fundamentally byits ionic strength.
25-2
389'

Comparison of the effects of the chlorides of alkali metals and of the halidesof sodium, showed that differences in capacity to promote swelling existedbetween univalent ions of the same sign. These differences appeared to berelated to the position of the ions in the lyotropic series (Lloyd & Shore, 1938,p. 405), and may therefore indicate a competition for water between thesalt ions and the interfibrillary substance: a competition which, in higherconcentration, would result in 'salting out'. But, as has been shown in Table 2,there would also seem to be a relationship between the capacity of ions tohydrate the substance and the length of their radii. Since this is a factor whichto some extent determines the strength of interionic forces (Lloyd & Shore,1938, p. 412), the observed differences in the hydrating capacity of these saltsmay not be due entirely to differences in lyotropism.
Study of the effect of pH upon the interfibrillary substance has been shownto be fundamental to the elucidation of its structure and mode of hydration.In the living body, however, it is to be expected that variation in pH must playan insignificant role in determining the condition of connective tissue-sinceliving cells could not survive at pH values such as have been shown to causechanges in the tissue in vitro. This is not so of salt. Acting at pH valuesapproximating to that of the blood, the lowest concentration of NaCl whichoccasioned visible change in the tissue was 0125M (0.73%). With increase ofconcentration to 0-25M (1.46%) the hydration of the tissue was very greatlyincreased.' It is of considerable significance that these values lie on either sideof that concentration of NaCl which is isotonic with the cells of the body.
SUMMARY
1. Changes which take place in rat interstitial connective tissue, as theresult of variation in external pH between 4-5 and 110 and in up to molarconcentrations of neutral salts, occur exclusively in the interfibrillary substance.The fibres and cells of the tissue take no significant part.
2. The interfibrillary substance is a hydrophilic colloid having a continuousnon-aqueous component held together by linkages which are disintegrated bytrypsin. It therefore belongs to the category of organized protein. The isoelectricpoint of this protein lies between pH 4-2 and 3-8.
3. The internal scaffolding of the substance is weak and yields to smallforces. It is for this reason that its hydration is modified by very slight changesin external pH and that changes occur in it in circumstances in which collagenfibres remain unaltered.
4. Variation in the hydration of the substance under the influence of changein external pH is considered to be due to attraction or repulsion of chargedcentres which develop as the result of ionization or suppression of ionizationof its multipolar protein molecules.
390 T.-D. DAY
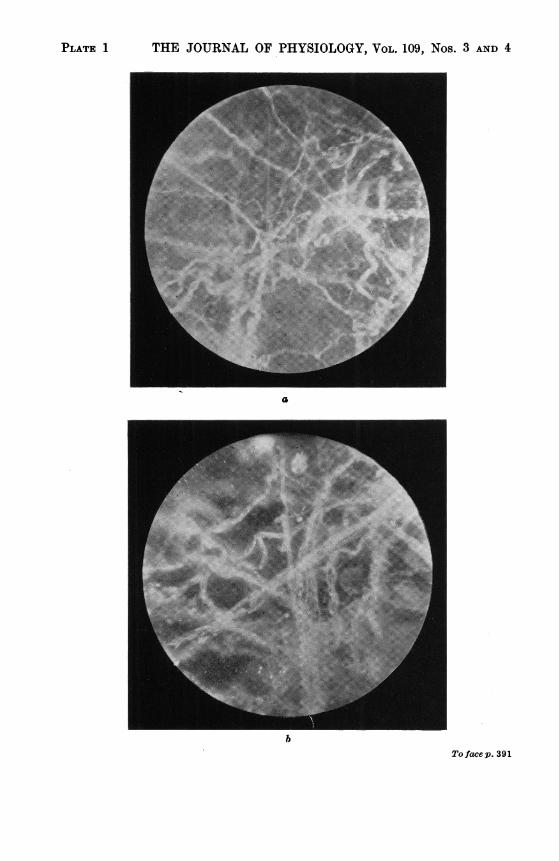
PLATE 1 THE JOURNAL OF PHYSIOLOGY, VOL. 109, Nos. 3 AND 4
a
bTo facep. 391

HYDRATION OF CONNECTIVE TISSUE 391
5. The hydrating action of neutral salts on the substance is due to forces setup by their free ions. In the living animal this action of neutral salts is probablyof greater importance than that of pH.
6. Schade's conception of a 'swelling antagonism' between fibres and inter-fibrillary substance is criticized in the light of these findings.
I wish to thank Mr W. W. Holland, student ofthis Medical School, for his enthusiastic assistance.
REFERENCES
Astbury, W. T. (1933). Fundamental8 offibre 8tructure. Oxford University Press, London.Day, T. D. (1947a), J. Path. Bact. 59, 567.Day, T. D. (1947b). Lancet, ii, 945.Day, T. D. (1948). J. Path. Bact. 60, 150.Flemming, W. (1876). Arch. mikr. Anat. 12, 391.Lloyd, D. J. (1941). J. Soc. Dy. Col., Bradford, 57,,281.Lloyd, D. J. & Phillips, H. (1933). Trans. Faraday Soc. 29, 132.Lloyd, D. J. & Shore, A. (1938). Chemistry of the Proteins& 2nd ed. London: Churchill.Mellanby, J. (1905). J. Physiol. 33, 338.Schade, H. (1913). Z. exp. Path. Ther. 14, 1.Schade, H. (1927). Ergebn. inn. Med. Kinderheilk. 32, 425.Schade, H. & Menschel, H. (1922). Kolloidzwchr. 31, 171.
EXPLANATION OF PLATE
a. Dark ground microscopic appearance of fascia which has come to equilibrium with an externalpH of 5-7 The collagen bundles are compact and their constituent fibres cannot be identified.With further acidification to pH 4 5, the bundles would themselves be drawn together, to suchanextent that the entire microscopic field would becomeopaqueandunsuitableforphotography.x 360.
b. Dark ground microscopic appearance of fascia at equilibrium with an external pH of I10.The bundles are widened and the individual fibrils are separated from each other and can beidentified individually. x 360.




















