
ISÓTOPOS ESTABLES EN ORNITOLOGÍA: UNA INTRODUCCIÓN BREVE
18
95 ORNITOLOGIA NEOTROPICAL 19 (Suppl.): 95–112, 2008 © The Neotropical Ornithological Society ISÓTOPOS ESTABLES EN ORNITOLOGÍA: UNA INTRODUCCIÓN BREVE Guillermo E. Pérez 1 , Jorge E. Schondube 2 , & Carlos Martínez del Rio 3,4 1 Sitka Biological Services, 4621 Koksilah Road, Duncan, British Columbia, Canada V9L 6N6. E-mail: [email protected] 2 Laboratorio de Ecología Funcional, Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, Campus Morelia. E-mail: [email protected] 3 University of Wyoming, Department of Zoology, Laramie, WY 82071-3166, USA. E-mail: [email protected] Abstract. – Stable isotopes in ornithology: A brief introduction. – Over the last ten years, stable iso- topes have become a widely used tool in ornithology and animal ecology. Here we review the nomencla- ture and methods used to analyze the isotopic composition of bird tissues and describe the uses of this technology in the study of birds. We argue that stable isotope analyses allow researching the niche dimen- sions characterized by George Evelyn Hutchinson as scenopoetic and bionomic. The use of stable iso- topes to populate the scenopetic axes of the niche depends on the existence of “isoscapes” generated by the hydrogen and oxygen isotopic composition of environmental water, as well as by the strontium com- position of soils. These isoscapes allow pinpointing the geographic site of origin of birds, the climatic envi- ronment that they inhabit, and to tack their local and migratory movements. The metabolic processes of primary producers, such as photosynthesis and nitrogen fixation, create a large number of isotopic signals that permit exploring the bionomic axes of the ecological niche. The isotopic composition of carbon and nitrogen can be used to find out the type of resources used by birds and/or their position in food webs. Stable isotopes are a especially useful tool to investigate the trophic diversity of individuals, populations, and communities. Resumen. – En los últimos 10 años el análisis de isótopos estables se ha convertido en una herramienta ampliamente usada en ecología animal y ornitología. En esta revisión presentamos la nomenclatura y métodos usados para analizar la composición isotópica de tejidos de aves, y describimos las aplicaciones de esta tecnología como herramientas para el estudio de las aves. Argumentamos que el análisis de isótopos estables permite investigar las dimensiones del nicho ecológico caracterizadas como scenopoéticas y bio- nómicas por George Evelyn Hutchinson. El uso de isótopos para poblar los ejes scenopoéticos del nicho depende de la existencia de “isopaisajes” generados por la composición isotópica del agua ambiental (espe- cialmente de los isótopos de hidrógeno y oxígeno) y de los suelos (especialmente el estroncio). Estos iso- paisajes permiten determinar el sitio de origen geográfico de las aves, el ambiente climático en donde viven, y rastrear sus movimientos locales y migratorios. Los procesos metabólicos de los productores pri- marios, tales como la fotosíntesis y la fijación de nitrógeno, producen una gran variedad de señales isotópi- cas que permiten explorar las dimensiones bionómicas del nicho ecológico. La composición isotópica de carbono y nitrógeno puede ser usada para determinar el tipo de recursos utilizados por las aves, y/o su posición dentro de las cadenas tróficas. Los isótopos estables son una herramienta espacialmente útil para ______________ 4 Autor corresponsal.
Transcript of ISÓTOPOS ESTABLES EN ORNITOLOGÍA: UNA INTRODUCCIÓN BREVE

95
ORNITOLOGIA NEOTROPICAL 19 (Suppl.): 95–112, 2008© The Neotropical Ornithological Society
ISÓTOPOS ESTABLES EN ORNITOLOGÍA: UNA INTRODUCCIÓN BREVE
Guillermo E. Pérez1, Jorge E. Schondube2, & Carlos Martínez del Rio3,4
1Sitka Biological Services, 4621 Koksilah Road, Duncan, British Columbia, Canada V9L 6N6. E-mail: [email protected]
2Laboratorio de Ecología Funcional, Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, Campus Morelia. E-mail: [email protected]
3University of Wyoming, Department of Zoology, Laramie, WY 82071-3166, USA. E-mail: [email protected]
Abstract. – Stable isotopes in ornithology: A brief introduction. – Over the last ten years, stable iso-topes have become a widely used tool in ornithology and animal ecology. Here we review the nomencla-ture and methods used to analyze the isotopic composition of bird tissues and describe the uses of thistechnology in the study of birds. We argue that stable isotope analyses allow researching the niche dimen-sions characterized by George Evelyn Hutchinson as scenopoetic and bionomic. The use of stable iso-topes to populate the scenopetic axes of the niche depends on the existence of “isoscapes” generated bythe hydrogen and oxygen isotopic composition of environmental water, as well as by the strontium com-position of soils. These isoscapes allow pinpointing the geographic site of origin of birds, the climatic envi-ronment that they inhabit, and to tack their local and migratory movements. The metabolic processes ofprimary producers, such as photosynthesis and nitrogen fixation, create a large number of isotopic signalsthat permit exploring the bionomic axes of the ecological niche. The isotopic composition of carbon andnitrogen can be used to find out the type of resources used by birds and/or their position in food webs.Stable isotopes are a especially useful tool to investigate the trophic diversity of individuals, populations,and communities.
Resumen. – En los últimos 10 años el análisis de isótopos estables se ha convertido en una herramientaampliamente usada en ecología animal y ornitología. En esta revisión presentamos la nomenclatura ymétodos usados para analizar la composición isotópica de tejidos de aves, y describimos las aplicaciones deesta tecnología como herramientas para el estudio de las aves. Argumentamos que el análisis de isótoposestables permite investigar las dimensiones del nicho ecológico caracterizadas como scenopoéticas y bio-nómicas por George Evelyn Hutchinson. El uso de isótopos para poblar los ejes scenopoéticos del nichodepende de la existencia de “isopaisajes” generados por la composición isotópica del agua ambiental (espe-cialmente de los isótopos de hidrógeno y oxígeno) y de los suelos (especialmente el estroncio). Estos iso-paisajes permiten determinar el sitio de origen geográfico de las aves, el ambiente climático en dondeviven, y rastrear sus movimientos locales y migratorios. Los procesos metabólicos de los productores pri-marios, tales como la fotosíntesis y la fijación de nitrógeno, producen una gran variedad de señales isotópi-cas que permiten explorar las dimensiones bionómicas del nicho ecológico. La composición isotópica decarbono y nitrógeno puede ser usada para determinar el tipo de recursos utilizados por las aves, y/o suposición dentro de las cadenas tróficas. Los isótopos estables son una herramienta espacialmente útil para______________4Autor corresponsal.
96
PÉREZ ET AL.
la investigación de la diversidad trófica de individuos, poblaciones y comunidades. Aceptado el 15 de Octubrede 2007.
Key words: Diets, stable isotopes, migration, ecological niches
INTRODUCCIÓN
Los elementos químicos con frecuencia tie-nen varias formas atómicas llamadas isótopos.Los isótopos de un elemento son átomos quetienen el mismo número de protones (i.e., tie-nen el mismo número atómico), pero quedifieren en el número de neutrones, y por lotanto difieren en su masa. Los isótopos pue-den ser estable o inestables (radiogénicos,Rundel et al. 1989). En este articulo, nos enfo-camos en el uso de isótopos estables comomarcadores de procesos ecológicos. Losisótopos estables son marcadores ecológicosde gran utilidad debido a que muchos proce-sos fisicoquímicos dependen de diferenciasen la energía de disociación de las moléculasque participan en ellos, y esta energía varíacon la masa atómica de los elementos en estasmoléculas (Lajtha & Marshall 1994). Porejemplo, los procesos enzimáticos que losorganismos usan para construir y transformarbiomoléculas pueden favorecer a un isótoposobre otro. Debido a que es mas fácil formary romper enlaces químicos que contengan elisótopo mas ligero, las moléculas que contie-nen el isótopo ligero son mas abundantes enlos productos de reacciones químicas (Hoeffs2004). En consecuencia, los residuos que noreaccionan tienden a quedar enriquecidos enel isótopo pesado. Efectos similares ocurrenen procesos físicos, como la evaporación y lacondensación (Lajtha & Marshall 1994).
Estos efectos fisicoquímicos que generandiferencias en la composición isotópica de lasmoléculas son muy útiles para los ecólogos.La composición isotópica de muchos materia-les, incluyendo los recursos que los animalesusan y los mismos tejidos de los animales, confrecuencia contienen la señal de los procesos
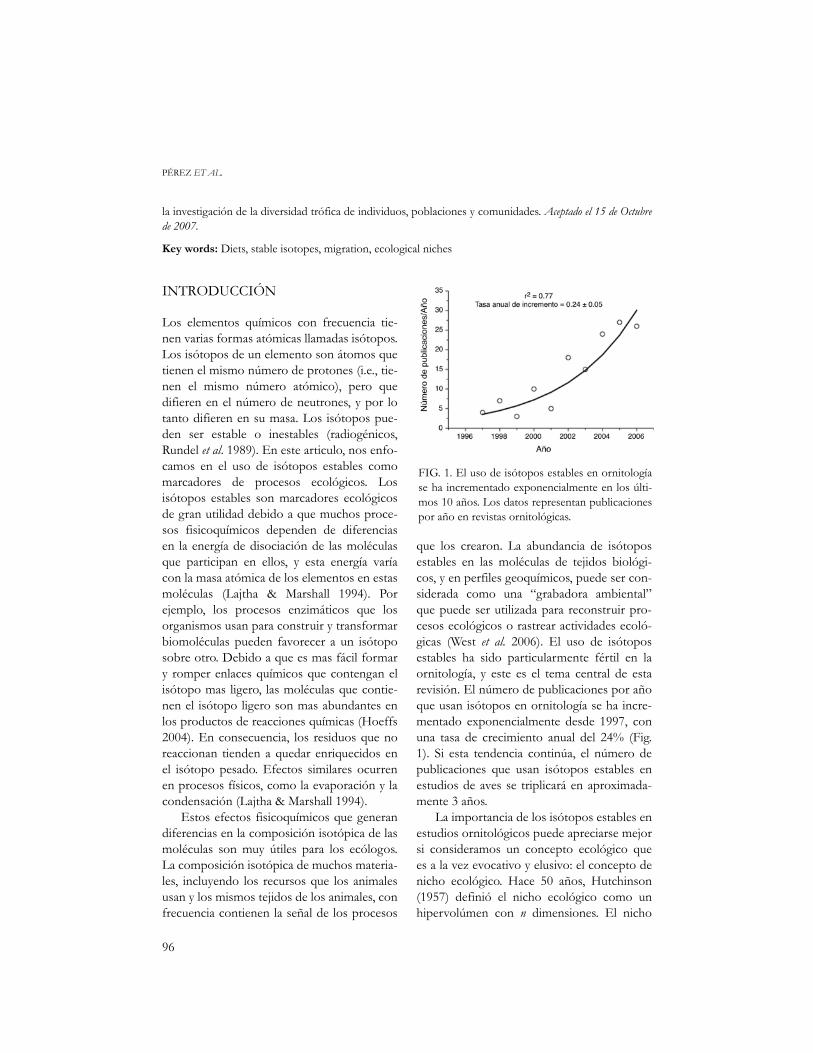
que los crearon. La abundancia de isótoposestables en las moléculas de tejidos biológi-cos, y en perfiles geoquímicos, puede ser con-siderada como una “grabadora ambiental”que puede ser utilizada para reconstruir pro-cesos ecológicos o rastrear actividades ecoló-gicas (West et al. 2006). El uso de isótoposestables ha sido particularmente fértil en laornitología, y este es el tema central de estarevisión. El número de publicaciones por añoque usan isótopos en ornitología se ha incre-mentado exponencialmente desde 1997, conuna tasa de crecimiento anual del 24% (Fig.1). Si esta tendencia continúa, el número depublicaciones que usan isótopos estables enestudios de aves se triplicará en aproximada-mente 3 años.
La importancia de los isótopos estables enestudios ornitológicos puede apreciarse mejorsi consideramos un concepto ecológico quees a la vez evocativo y elusivo: el concepto denicho ecológico. Hace 50 años, Hutchinson(1957) definió el nicho ecológico como unhipervolúmen con n dimensiones. El nicho
FIG. 1. El uso de isótopos estables en ornitologíase ha incrementado exponencialmente en los últi-mos 10 años. Los datos representan publicacionespor año en revistas ornitológicas.

97
ISÓTOPOS ESTABLES Y AVES
recibió considerable atención después de serdefinido, pero el interés en este concepto lan-guideció hacia el fin del siglo XX para renacera principios del siglo XXI. Definiciones delnicho abandonadas como inoperantes hansido rejuvenecidas. El concepto de Grinnell(1917) del nicho como “hábitat” reencarnó enel concepto de nicho bioclimático desarro-llado por los modeladores de áreas de distri-bución (Kearney 2006), mientras que el nichoEltoniano parece haber renacido como elnicho funcional de Chase & Leibold (2003).El resurgimiento del concepto de nicho enecología parece deberse a avances tecnológi-cos y conceptuales. El nicho bioclimático es elresultado de avances notables en la tecnologíade sistemas de información geográfica, y de lahabilidad de computadores de manejar enor-mes volúmenes de datos espacialmenteestructurados (Elith et al. 2006). El nicho fun-cional depende fundamentalmente del con-cepto de isoclinas de crecimiento cero(Tilman 1988).
Recientemente, Bearhop et al. (2004) yNewsome et al. (2007) propusieron que el aná-lisis de isótopos estables, que es una tecnolo-gía relativamente nueva, puede ser utilizadopara investigar muchas de la preguntas quetradicionalmente son consideradas como ele-mentos en el estudio del nicho ecológico.Estos autores han reconocido la similitud for-mal entre el nicho Hutchinsoniano y el modocomo los ecólogos isotópicos presentan susdatos. Típicamente, en un estudio isotópicolos datos son presentados en un espacio carte-siano en el que los ejes representan la abun-dancia relativa de cada isótopo (Newsome etal. 2007). El volumen ocupado en estos espa-cios puede ser considerado como la versiónisotópica del nicho ecológico. Existe una simi-litud notable entre el nicho isotópico yla formulación Huthinsoniana del nicho.Hutchinson (1978) estableció una valiosa dis-tinción entre los ejes “scenopoéticos” y losejes “bionómicos” del nicho. Los ejes sceno-
poéticos son aquellos que determinan el esce-nario bioclimático en el que las especiesactúan sus obras evolutivas (Hutchinson1978). Los ejes bionómicos, son aquellos queidentifican los recursos que los organismosutilizan. Los ecólogos han reconocido enrepetidas ocasiones las ideas capturadas porlos términos bionómico y scenopoético. Aus-tin (2002) se refiere a variables “directas” y de“recursos” y Begon et al. (2006) usan los tér-minos “condiciones” y “recursos”. Debido aque estos dos tipos de variables interactúan,no pueden ser considerados como estricta-mente independientes, sin embargo represen-tan una primera aproximación útil (Soberón2007).
El crecimiento explosivo que ha experi-mentado el uso de isótopos estables en ecolo-gía animal (incluyendo la ornitología) en lasúltimas dos décadas tiene dos corrientes quecorresponden de un modo sorprendente conlos ejes scenopoeticos y bionómicos de Hut-chinson. Una de estas corrientes utiliza losisótopos estables para seguir los movimientosde los animales (ver Rubenstein & Hobson2004, y referencias ahí citadas). La otra, losutiliza para investigar como los animales usanrecursos (Bearhop et al. 2004). La primeraaplicación se basa en que diversos factoresfísicos crean patrones geográficos predecibles,o “isopaisajes”, en la composición isotopicadel hidrógeno, oxígeno y estroncio del agua ylos minerales (West et al. 2006). Los factoresfísicos que moldean la topografía de estos iso-paisajes incluyen el origen geológico de lossuelos (Koch et al. 1995), la altitud, la latitud,la distancia a la costa y la temperatura (Bowenet al. 2005). Consecuentemente, los isopaisajeshan sido usados con frecuencia para rastrearlos movimientos locales y las migraciones delos animales (Hobson 2005). Cuando un ani-mal puede ser asociado a una localidad geo-gráfica, puede a la vez ser asociado con unconjunto de variables bioclimáticas (Pearson& Dawson 2003). Por tanto, teóricamente es

98
PÉREZ ET AL.
posible establecer vínculos conceptuales entrela ecología isotópica y los esfuerzos de losmodeladores de áreas geográficas de distribu-ción. La primera aplicación que revisaremoses el uso de isótopos estables en el estudio delos movimientos de las aves.
La segunda corriente en ecología isotópicadepende de otra observación significativa: losproductores, en la base de las cadenas tróficas,con mucha frecuencia confieren, a las molé-culas orgánicas que fabrican, firmas o señalescaracterísticas en la composición isotópica decarbono y nitrógeno (Farquhar et al. 1989,Robinson 2001). Debido a que los consumi-dores incorporan estas señales isotópicas ensus tejidos, podemos usarlas para identificarsu dependencia por plantas con distintas víasfotosintéticas, tales como C3, C4 y CAM(Sponheimer et al. 2003, Wolf & Martínez delRio 2004). Como demostraremos en lasegunda aplicación, podemos usar una combi-nación de análisis de carbono y nitrógenopara determinar la contribución de tramasmarinas y terrestres a la dieta de un ave, o paraestimar su posición trófica (Post 2002, Stapp& Polis 2003, Sabat et al. 2006a). Estos ejem-
plos son la razón que llevó a Bearhop et al.(2004) a proponer que los análisis de isótoposestables son la herramienta ideal para investi-gar la estructura de nichos tróficos. Reciente-mente, Layman et al. (2006) han desarrolladoesta idea y han propuesto una serie de medi-das/métricas que permiten investigar laestructura trófica de comunidades. En seccio-nes subsiguientes revisamos críticamenteestas medidas y sugerimos como es que losornitólogos pueden usarlas.
La aplicación de isótopos estable en estosdos amplios temas requiere que primero defi-namos la jerga y los métodos de la disciplina.
EL LENGUAJE Y LOS MÉTODOSDE LOS ISOTOPOS ESTABLES
Los isotópos mas usados. Los elementos mas uti-lizados en estudios ecológicos son los ele-mentos “livianos”: nitrógeno (N), carbono(C), azufre (S), hidrógeno (H) y oxígeno (O).El elemento “pesado” mas usado en ecologíaisotópica es el estroncio (Sr). Sin embargo losanálisis de Sr son caros y este elemento esusado con menor frecuencia que el C, N, H,
TABLA 1. Abundancia promedio de los isotopos estables importantes para entender sistemas ecológicos yestándares. El azufre y el estroncio tienen mas de dos isótopos estables. Los isótopos estables adicionalesdel azufre y estroncio son 33S, 36S, 84Sr y 88Sr, respectivament (Tabla tomada de Karasov & Martínez del Rio2007).
Elemento Ligero Abundancia (%)
Pesado Abundancia (%)
Estándar
Hidrógeno
Carbono
Nitrógeno
Oxígeno
Azufre
Estronio
1H
12C
14N
17O
2S
86Sr
99,984
98,89
99,63
99,759
95,00
9,86
2H (D)
13C
15N
18O
34S
87Sr
0,016
1,11
0,37
0,204
8,22
7,02
SMOW: Media del agua estándar del océano
PDB: Belemnita PeeDee
N2 (atm.): Nitrógeno Atmosférico
SMOW
CD: Triolita del meteorito del Diablo Canyon

99
ISÓTOPOS ESTABLES Y AVES
O y S. La Tabla 1 enlista los elemento usadosen ecología isotópica, sus abundancias relati-vas en el planeta, y los estándares que se usanpara medirlos.
La notación delta y sus alternativas – ¿Cómo semiden los isótopos estables? La abundancia rela-tiva de un isótopo en relación al otro es expre-sada como el cociente (Rx) del isótopo pesadoy el ligero, como la proporción del isótopopesado (fx), o mas comunmente usando lanotación delta (δX). Como lo muestra la Tabla1, en general los isótopos pesados son consi-derablemente menos abundantes que los lige-ros y por tanto, en general, Rx y fx tienenvalores muy pequeños (Werner & Brand2001). Las diferencias proporcionales expre-sadas en la notación delta son intuitivamentemas claras. El valor de delta (en unidades departes por mil ‰) se estima usando lasiguiente formula:
Rmuestra – Restandar δX = [ —————– ] x 1000 ecuación 1 Restandar
en donde Rmuestra y Restándar son los cocientesde las abundancias relativas del isó-topo pesado sobre el ligero en la muestra y elestándar, respectivamente. Las medidasnecesarias para aplicar la ecuación 1 sonhechas en un espectrómetro de masas.Speakman (2005) y Karasov & Martínez delRio (2007) describen en detalle el fun-cionamiento y uso de este instrumento. En elrango natural de abundancias relativas, elporcentaje del isótopo pesado (i.e., %X =(fx) x 100) depende linealmente del valor de δ.Fry (2006) proporciona las ecuaciones nece-sarias para transformar valores de delta aporcentajes. Muestras “enriquecidas” en elisótopo pesado tiene valores mas positivos dedelta, mientras que muestras “empobrecidas”tienen valores de δ mas negativos, y por endeuna abundancia relativa mayor del isótopoligero.
Fraccionamiento y discriminación isotópica. La varia-ción isotópica natural en la abundancia de unisótopo en un material es la consecuencia delas pequeñas variaciones en masa que hacenque los diferentes isótopos se comporten demodos diferentes en los procesos físicos oquímicos que generaron el material. La dife-rencia isotópica entre dos especies molecula-res (A y B) que participan de un proceso sellama fraccionamiento (αA-B):
ecuación 2
Fry (2006) y Martínez del Rio & Wolf (2004)describen los procesos que generan fracciona-miento isotópico de interés para biólogos.Con frecuencia los tejidos de un animal difie-ren en su composición isotópica de los mate-riales que este ingiere como parte de su dieta.Esta diferencia es con frecuencia (e incorrec-tamente) llamada “fraccionamiento”. Cerling& Harris (1999) y Martínez del Rio & Wolf(2004) proponen restringir el uso de la palabrafraccionamiento al efecto de procesos quími-cos y físicos, y sugieren el uso del término“factor de discriminación” para la diferenciaisotópica entre tejidos y dieta:
∆tejido–dieta = δXtejido – δXdieta ecuación 3
Restringir el uso del término fraccionamientoa procesos fisicoquímicos bien entendidos esnecesario debido a que la diferencia entre losvalores isotópicos de la dieta y los tejidos deun animal es resultado de una gran cantidadde factores. Hobson & Clark (1992b),Rubenstein & Hobson (2004) y Martínez delRio & Wolf (2004) describen los factores quegeneran diferencias isotópicas entre tejidos ydieta.
El factor de discriminación es fundamen-talmente importante y poco entendido. Esimportante por dos razones: 1) los valores de∆ son necesarios para reconstruir proporcio-
α A− B =RA
RB
=(1000 + δ A )
(1000 + δ B )

100
PÉREZ ET AL.
nes de cada recurso en la dieta de un animalusando modelos de combinación o mezcla-do (Phillips & Gregg 2001, Phillips & Koch2002, Phillips & Gregg 2003), y 2) son re-queridos para estimar el nivel trófico de unanimal. En las secciones siguientes, considera-remos la segunda razón, aquí enfatizamos laprimera. Supongamos que un animal con-sume una proporción p de un alimento 1 concomposición isotópica δX1, y una proporción1-p de un alimento 2 con composición δX2. Silos dos alimentos tienen mas o menos lamisma composición nutricional y son asimila-dos con igual eficiencia, entonces:
δXtejidos = p(δX1 + ∆1) + (1-p)(δX2 + ∆ 2)
ecuación 4
en donde ∆1 y ∆2 son los valores de discrimi-nación para las dietas 1 y 2. La ecuación 4 esel modelo de mezclado mas simple (Phillips &Koch 2002). Este modelo puede ser fácil-mente extendido para dilucidar 3 tipos de ali-mento con dos isótopos (o n + 1 fuentes conn isótopos, Phillips & Gregg 2003), y lassuposiciones de igualdad en composiciónnutricional e igual asimilación pueden ser mo-dificadas (ver Martínez del Rio & Wolf 2004).Debe notarse que para aplicar la ecuación 4, osus extensiones multivariadas, deben cono-cerse los factores de discriminación. Desafor-tunadamente estos valores varían en respuestaa diferentes factores como: 1) el tipo de tejido(Hobson & Clark 1992b), 2) la dieta (Hobson& Clark 1992a), y 3) el estado fisiológico delorganismo (McKechnie et al. 2004). Frecuen-temente, los valores promedio de ∆13C(≈ 1‰, Arneson & MacAvoy 2005) y ∆15N(≈ 3,4‰, Post 2002, Mirón et al. 2006) sonusados incorrectamente en estudios ecológi-cos. Consideramos que su uso es incorrectodebido a que estos valores son promedios dedistribuciones con varianzas altas y son deri-vados de estudios en los que los investigado-
res midieron la composición isotópica enmúsculo o en organismos completos (DeNiro& Epstein 1978, Fry et al. 1984, Fry 1988),dos tipos de tejidos poco usados en ornitolo-gía. Uno de los retos de la ecología isotópicaes la elaboración de un catalogo de valores de∆ como función de los factores que influyenen su magnitud para ser usados en las situa-ciones que cada investigación demanda.
ESTUDIOS ISOTÓPICOS DE MIGRA-CIÓN
Establecer patrones de movimientos tempo-rales de animales migratorios y su rol en elecosistema es crucial para nuestro entendi-miento de sus ciclos de vida, comportamientoy ecología. En teoría, al saber la ubicación delos sitios de verano, invierno y de paradadurante la migración, podemos conservar conmayor eficiencia estos animales y los hábitatsque utilizan. Muchos animales se mueven dia-riamente entre hábitats y transfieren nutrien-tes entre ellos. Otros, como las avesmigratorias de largas distancias, desempeñansus funciones ecológicas en lugares geográfi-camente y ecológicamente distintos, conec-tando comunidades a través de continentes endiferentes temporadas del año. Aunque losmovimientos de los animales son centrales enecología, nuestra capacidad para rastrear losmovimientos de individuos es rudimentaria.Especialmente al rastrear aves paserinaspequeñas, los programas de rastreo conven-cionales como anillamiento y radiotelemetríano han dado resultados contundentes (Hob-son 2003). Afortunadamente, nuestra habili-dad para rastrear movimientos migratorios seha incrementado con el uso de métodosmoleculares, como isótopos estables y gené-tica (Webster et al. 2002) y el uso de radiotransmisores cada vez mas pequeños.
Nuestro enfoque es isótopos estables. Los isótoposestables de carbono, deuterio y estroncio han

101
ISÓTOPOS ESTABLES Y AVES
sido utilizados para rastrear movimientos deanimales migratorios (Hobson 1999). El deu-terio (D) es el isótopo mas útil y mas frecuen-temente usado en estudios migratorioslatitudinales, porque la composición de deute-rio en lluvia presenta un patrón continentalamplio en Norte América. En Norte América,el contenido de deuterio en agua de lluviadecrece desde el sudeste hacia el noroeste for-mando un isopaisaje latitudinal muy claro(Hobson & Wassenaar 1997). En Norte Amé-rica, existen también gradientes altitudinalessignificativos (Meehan et al. 2004). En general,δD decrece con la elevación (Hobson et al.2003). Debido a que el δD y el δ18O covaríanen el agua de lluvia (llamada agua “meteórica”por los geoquímicos), los dos isótopos mues-tran patrones similares (Bowen et al. 2005). Larazón de este patrón geográfico es que,durante la evaporación de un cuerpo de agua,las moléculas de agua “livianas” escapan masfácilmente hacia la atmósfera que las molécu-las de agua mas “pesadas”. El vapor de aguaatmosférica esta siempre mas empobrecidoque las aguas oceánicas, y mas empobrecidocerca de los polos (ver Bowen et al. 2005). Lasituación opuesta ocurre cuando el agua secondensa en la atmósfera. La condensaciónfavorece a las moléculas de agua pesadas (ricasen D y 18O) en la lluvia y en la nieve. Las pri-meras lluvias que caen de nubes formadassobre el océano tienen valores de δD y 18Ocercanos a las aguas oceánicas donde se for-maron las nubes. Así, la precipitación mas alinterior del continente es derivada de unareserva de vapor que tiene valores delta aúnmas negativos que las aguas oceánicas masrecientemente evaporadas (Karasov & Martí-nez del Rio 2007).
Muchos estudios han establecido correla-ciones entre la composición de D/H en aguameteórica durante la temporada de creci-miento y la de la biomasa en plantas (Yapp &Epstein 1982). Los animales comen estas bio-masas e incorporan esta señal isotópica en sus
tejidos. La esencia de esta metodología fun-ciona bajo la noción de que la abundancia deδD en los tejidos de un consumidor refleja laúltima fuente de δD en precipitación a travésde la cadena. Por eso, al medir los valores deδD en los tejidos de animales es posible infe-rir la latitud aproximada (o posición en el iso-paisaje) en donde los tejidos fueronsintetizados (Pérez & Hobson 2007). Las plu-mas de vuelo de las aves han sido el tejido masutilizado para inferir conexiones migratorias(ver Hobson 2005 y referencias ahí citadas)porque, una vez, sintetizadas, pasan a sermateria inerte, conservando la firma isotópicadel sitio donde fueron sintetizadas hasta lapróxima muda (Pérez & Hobson 2006). Hob-son & Wassenaar (1997) y Chamberlain et al.(1997) encontraron una muy buena relaciónentre la composición isotópica de deuterio enplumas de aves insectívoras y el valor prome-dio de δD en precipitación durante el periodode crecimiento. El factor de discriminación deδD entre precipitación y plumas es mas omenos constante y cercano a –25‰ (Wasse-naar & Hobson 2000, Pérez & Hobson 2007),aunque Hobson (2005) y Norris et al. (2006)advierten que este factor no siempre puedeser aplicado a todas las áreas del continente,en todas las aves, edades (Meehan et al. 2003,Langin et al. 2007) y estados fisiológicos(McKechnie et al. 2004). Conocer los factoresque determinan la discriminación isotópicaentre plumas, dieta y agua preformada debeser una de las prioridades en los estudios iso-tópicos de migración. Wassenaar & Hobson(2000), Hobson & Wassenaar (2001) yRubenstein et al. (2002) usaron la composiciónisotópica de δD y 13C en plumas para delinearlos orígenes geográficos de Agelaius phoenicus,Lanius ludovicianus, y Dendroica caerulescens, res-pectivamente. Ellos encontraron que la com-posición de deuterio se correlacionó con lalatitud, mientras que 13C se correlaciono isotó-picamente con distintas cadenas alimenticias,como cultivos C4 (zahína y maíz) y trigo e

102
PÉREZ ET AL.
insectos con firmas 13C. Estos análisis, ilus-tran la idea de que algunas medidas isotópicasesclarecen variables scenopoéticas del nicho(δD), mientras que otras dilucidan variablesbionómicas (δC, Newsome et al. 2007).
Poco a poco el método se ha comenzadoa aplicar en otros continentes como Europa(Hobson et al. 2004), Africa (Yohannes et al.2007) y América del Sur (Farmer et al. 2003,
Hobson et al. 2003). Aunque el patrón geográ-fico de deuterio en precipitación para Amé-rica del Sur ha sido desarrollado de un modopreliminar (Bowen et al. 2005, Fig. 2), la corre-lación entre la firma isotópica en precipita-ción y en plumas no ha sido aún generadapara este continente. Existen buenas oportu-nidades para estudiar la migración usandoisótopos estables en América del Sur. La selva
FIG 2. Patrón de composición isotópica del agua de lluvia en Sudamérica corregida por altitud (producidapor Gabriel Bowen, también ver Bowen et al. 2005). Los datos son resultado de mediciones promedio deisótopos en agua de lluvia generados por la Red Global de Isótopos en Lluvia (GNIP) durante la época decrecimiento máximo (Septiembre a Marzo) por aproximadamente 40 años. La Red Global de Isótopos enLluvia es administrada por la Associación Internacional de Energía Atómica (IAEA) y la OrganizaciónMeteorológica Mundial (WMO) (Bowen et al. 2005).

103
ISÓTOPOS ESTABLES Y AVES
Amazónica, el cono sur de América de Sur yla Cordillera de los Andes poseen isopaisajesdramáticos y dignos de uso en investigación(Fig. 2). El potencial de examinar la variaciónisotópica de una variedad de elementos queconstituyen las fuentes alimenticias locales delas aves sudamericanas para rastrear sus movi-mientos migratorios está completamenteabierto.
ESTUDIOS DE NICHO TRÓFICO
Señales isotópicas indicadoras de trama trófica y susaplicaciones. Una gran variedad de fenómenoslleva a cocientes isotópicos característicos enlos materiales bióticos y abióticos que usan losanimales. Estos cocientes isotópicos puedenllamarse “señales isotópicas” y, debido a quelos animales las incorporan en sus tejidos,podemos usarlas para determinar sitios de ori-gen, como mostramos en la sección anterior,o para determinar el uso de recursos. Quizásel proceso de fraccionamiento isotópicomejor estudiado en biología es la fotosíntesis(Farquhar et al. 1989). Los isótopos de car-bono pueden ser usados para diferenciar entreplantas que utilizan los modos C3, C4 y CAMde fotosíntesis. Las plantas C3 discriminan encontra de CO2 que contiene 13C en dos pasosdel proceso de fijación del carbono: durante ladifusión/disolución y durante la carboxilaciónllevada a cabo por la enzima RUBISCO en elciclo de Calvin (Farquhar et al. 1989). La difu-sión a través de los estomas y la disolución enel agua del tejido fotosintético produce unfraccionamiento contra el 13C de aproximada-mente 2‰ (Farquhar et al. 1989). El segundopaso que discrimina contra el 13C, la carboxila-ción por RUBISCO, tiene un efecto mayor (de≈19‰, Farquhar et al. 1989). El resultado deestos dos procesos es que los tejidos de lasplantas C3 están empobrecidos en 13C, convalores de δ13C de entre -35‰ a -22‰, mien-tras que el CO2 atmosférico tiene un valor deaproximadamente -8‰. Los valores extre-
mos de δ13C dentro de este rango son de plan-tas que crecen en bosques y bajo doselescerrados, o en hábitats áridos, respectiva-mente. Norris et al. (2004) mostraron que dosindicadores de éxito reproductivo en el avemigratoria Neotropical Setophaga ruticilla, elnúmero de polluelos producidos y la fecha dellegada al área de reproducción, están correla-cionadas con el valor de δ13C de sus tejidos.Individuos que pasaron el invierno en zonashúmedas, y que por tanto tienen tejidos convalores de δ13C mas negativos, producen maspolluelos y llegan al sitio de reproducción mastemprano, que individuos con valores de δ13Cmas positivos y que por tanto pasaron elinvierno en sitios mas secos (Marra et al.1998).
Los procesos fotosintéticos C4 y CAM sonacompañados de menores fraccionamientosque la fotosíntesis C3. La razón de esta dife-rencia es que en estos sistemas, la actividad deRUBISCO tiene lugar bajo condiciones mas omenos cerradas en las que el CO2 enriquecidoen 13C no puede ser liberado a la atmósfera ytiene que ser utilizado. En ambos sistemas C4y CAM, el CO2 disuelto en el agua del tejidofotosintético es fijado en ácidos orgánicos y elfraccionamiento de este proceso es pequeño(≈ 2‰, Farquhar et al. 1989, Dawson et al.2002). Estos ácidos orgánicos son subsecuen-temente decarboxilados y el CO2 resultante esusado por RUBISCO en la reducción fotosin-tética del carbono. Los tejidos de las plantasC4 y CAM obligadas son isotópicamente massimilares a la atmósfera (entre -14 y -10 ‰)que los de las plantas C3 (Farquhar et al. 1989,Dawson et al. 2002). Wolf & Martínez del Rio(2000) and Wolf et al. (2002) han usado la dife-rencia en valores de δ13C entre plantas CAM yC3 para medir la dependencia de dos especiesde palomas de desierto (Zenaida asiatica y Z.macroura) por una planta clave, el saguaro (Car-negeia gigantea). Estos autores tomaron ventajade que el agua en el néctar y la pulpa de losfrutos del saguaro están fuertemente enrique-

104
PÉREZ ET AL.
cidas en D para investigar la dependencia deestas palomas, no solamente de la energíaprovista por el saguaro, sino además del aguaque esta planta ofrece. Encontraron que en Z.asiatica, δ13C y δD están positivamente corre-lacionadas. En esta especie, la adquisición decarbono de saguaro está acompañada deadquisición de agua. En cambio, en Z.macroura δ13C y δD son independientes. Estaespecie solamente obtiene energía delsaguaro.
Las plantas marinas y dulceacuícolas pre-sentan un rango muy amplio de valores deδ13C (Tabla 2). Esta amplitud de rango sedebe a que además de la variación en el modode fotosíntesis, otros factores influyen en elgrado en que sus tejidos discriminan en con-tra del 13C. La difusión del bicarbonato a tra-vés de una barrera de agua estancada puedeprevenir la discriminación contra el 13C. Sitodo el CO2 es usado, no existe la posibilidadde que RUBISCO discrimine en contra del13C y en consecuencia, los tejidos de las plan-tas reflejan el valor isotópico del CO2 disponi-ble. Finlay et al. (1999) encontraron que losvalores de δ13C de los productores primarios y
los herbívoros de ríos y arroyos dependen delflujo del agua. En arroyos rápidos y, por tanto,con abundante CO2, el valor de δ13C es masnegativo que en arroyos de bajo flujo. Elmismo proceso tiene lugar a una escala mayoren el mar. France (1995) encontró diferenciasentre el valor de δ13C entre algas bentónicas yplanctónicas, y Hobson et al. (1994) usaronestas diferencias para distinguir entre avesmarinas que comen de fuentes cercanas y leja-nas a la costa. A pesar de que las plantas acuá-ticas muestran variación significativa en δ13C,existen suficientes diferencias en la composi-ción de isótopos de carbono entre plantasterrestres y acuáticas para distinguir la contri-bución relativa de fuentes acuáticas/marinas yterrestres a la dieta de aves (Harding & Ste-vens 2004).
Tal como es el caso con el carbono, el usode los isótopos de nitrógeno (15N y 14N) enornitología depende de su distribución en elalimento. Así como en el caso del carbono, lavariación isotópica en el nitrógeno (i.e., en elvalor de δ15N) se debe a una gran variedad deprocesos (revisados por Robinson 2001).Debido a que muchos de estos procesos no
TABLA 2. La composición isotópica de carbono de los productores primarios en los ecosistemas costerosvaría dramáticamente según la fuente. La información de esta tabla fue compilada por Fry & Sherr (1989)de diversas fuentes.
Fuente Rango usual en δ13C en ‰Plantas C3 terrestres -23 a -30Plantas C4 terrestres -10 a -14Seston ripario (Partículas orgánicas suspendidas) -25 a -27Depósitos de turba -12 a -28Plantas C3 de marismas -23 a -26Pastos C4 de marismas -12 a -14Pastos marinos -3 a -15Macroalgas -8 a -27Algas unicelulares bentónicas -10 a -20Fitoplancton de mares templados -18 a -24Fitoplancton ripario-estuarino -24 a -30Bacterias autotróficas dependientes de azufre -20 a -38Bacterias oxidadoras de metano -62

105
ISÓTOPOS ESTABLES Y AVES
han sido tan bien estudiados como la fotosín-tesis, el uso de los isótopos de nitrógeno esmas fenomenológico que el de carbono (sinque esto los haga menos útiles para describirvariables bionómicas). Las plantas varían enδ15N por al menos cuatro razones (revisadaspor Dawson et al. 2002): 1) los suelos varíanen δ15N, 2) las plantas con raíces profundastienden a estar mas enriquecidas en 15N quelas plantas con raíces someras, 3) las plantasasociadas a bacterias fijadoras de nitrógenotienden a estar empobrecidas entre 2 y 4 ‰ en15N con relación a las plantas que no presen-tan la capacidad de fijar nitrógeno, y 4) el fito-plancton (y las plantas marinas en general)tienden a estar mas enriquecidas en 15N quelas plantas terrestres (Kelly 1999). El rango enδ15N generado por estas cuatro causas esamplio. Abarca aproximadamente 26‰ y,potencialmente, provee de una gran variedadde señales isotópicas que pueden ser usadaspor ornitólogos y otros ecólogos animales.
δ15N como indicador de nivel trófico. Quizás el usomas frecuente de δ15N en ecología animal yornitología es la determinación de nivel tró-fico. En un artículo pionero, DeNiro & Eps-tein (1981) documentaron que el valor de∆15Ntejidos-dieta era en promedio de 3.4 ‰. Estaobservación es potencialmente muy útil. Sieste valor se mantiene constante entre espe-cies y niveles tróficos, podemos fácilmenteestimar el nivel trófico de un animal con lasiguiente formula:
λ = 1 + δ15Nconsu. + (δ15Nconsu.– δ15Nbase)/∆15N
ecuación 5
en donde λ es la posición trófica del orga-nismo usado como base (λ = 1 si se usanproductores primarios), y ∆15N = 3,4‰ (Post2002). Desafortunadamente, la realidad esmas complicada. Existe abundante evidenciaque apoya la idea de que existe un enriqueci-
miento en 15N entre un consumidor y su dieta.Sin embargo, la magnitud de este enriqueci-miento es muy variable (entre -1 y 6‰, Peter-son & Fry 1987, Post 2002). En un meta-análisis, Vander Klift & Ponsard (2003)encontraron diferencias consistentes entretaxa y tipos de tejido. Muchos factores puedendeterminar la variación en ∆15N, incluyendo latasa de crecimiento y la calidad de la dieta, y lamayoría de estos no han sido investigados(Martínez del Rio & Wolf 2004). Robbins et al.(2005) demostraron que ∆15N decrece con lacalidad de la proteína en la dieta de muchostaxa de vertebrados, y Trueman et al. (2005)encontraron que 1∆15N esta negativamentecorrelacionado con la tasa de crecimiento. Apesar del uso generalizado de ∆15N = 3,4 ‰,nos parece prudente sugerir cautela en su uso.En el mejor de los casos el valor de 1∆15Ndebe ser determinado experimentalmente, ocon datos de campo, en lugar de usar unaconstante de generalidad dudosa.
Una complicación adicional es la variaciónpotencial en la composición isotópica en labase de cadenas tróficas. Vander Zanden &Rasmussen (1999) reportan que los producto-res primarios en lagos varían en δ15N entre -2y 9‰. Herrera et al. (2003 y 2006) midieronδ15N en los frutos consumidos por aves enuna selva tropical húmeda y una seca, yencontraron diferencias notables en δ15N. Enla selva húmeda (Los Tuxtlas, México), losfrutos de plantas C3 y CAM tienen valores deδ15N de aproximadamente 1,5 y -4‰, res-pectivamente. En la selva tropical seca (Cha-mela, México), estos frutos tiene valores de4,6 y 2,6‰, respectivamente. Estos ejemplosdemuestran que los valores de δ15Nbase y 1∆15Nen la ecuación 5, no pueden ser consideradosconstantes.
Isótopos estables como covariables en estudios compa-rativos. La biología comparada depende de unmodo esencial del conocimiento de la historianatural de los organismos. La dieta ha sido
106
PÉREZ ET AL.
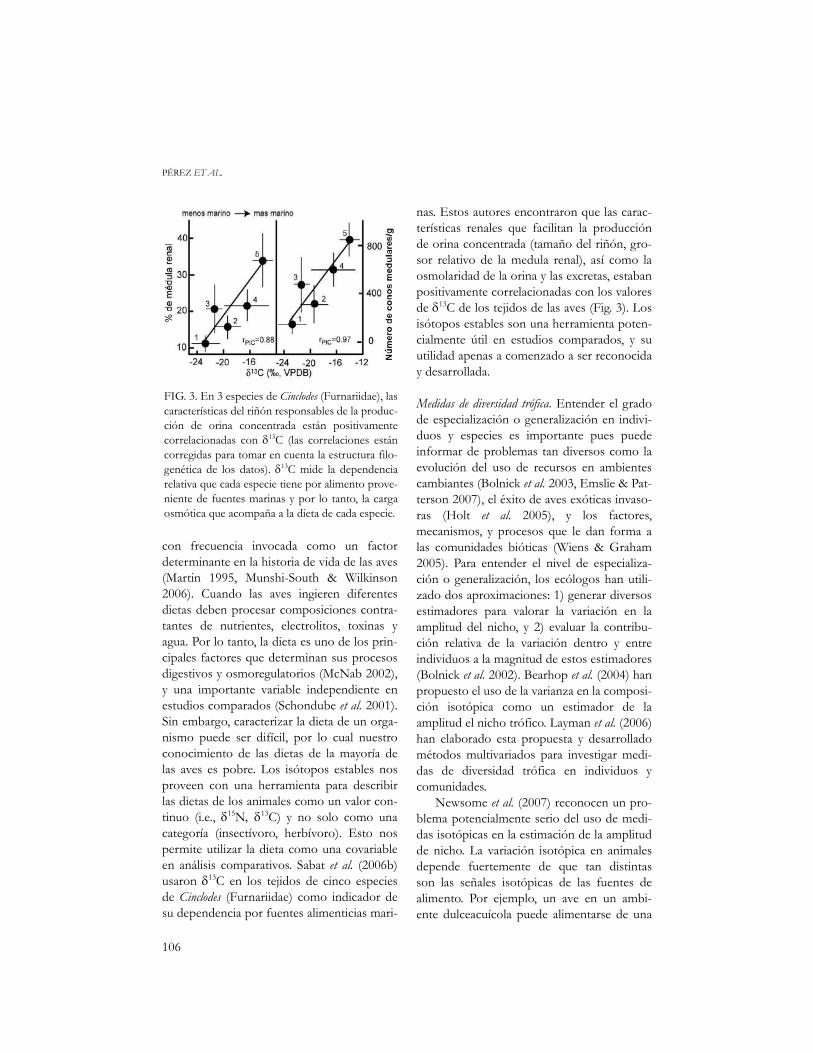
con frecuencia invocada como un factordeterminante en la historia de vida de las aves(Martin 1995, Munshi-South & Wilkinson2006). Cuando las aves ingieren diferentesdietas deben procesar composiciones contra-tantes de nutrientes, electrolitos, toxinas yagua. Por lo tanto, la dieta es uno de los prin-cipales factores que determinan sus procesosdigestivos y osmoregulatorios (McNab 2002),y una importante variable independiente enestudios comparados (Schondube et al. 2001).Sin embargo, caracterizar la dieta de un orga-nismo puede ser difícil, por lo cual nuestroconocimiento de las dietas de la mayoría delas aves es pobre. Los isótopos estables nosproveen con una herramienta para describirlas dietas de los animales como un valor con-tinuo (i.e., δ15N, δ13C) y no solo como unacategoría (insectívoro, herbívoro). Esto nospermite utilizar la dieta como una covariableen análisis comparativos. Sabat et al. (2006b)usaron δ13C en los tejidos de cinco especiesde Cinclodes (Furnariidae) como indicador desu dependencia por fuentes alimenticias mari-
nas. Estos autores encontraron que las carac-terísticas renales que facilitan la producciónde orina concentrada (tamaño del riñón, gro-sor relativo de la medula renal), así como laosmolaridad de la orina y las excretas, estabanpositivamente correlacionadas con los valoresde δ13C de los tejidos de las aves (Fig. 3). Losisótopos estables son una herramienta poten-cialmente útil en estudios comparados, y suutilidad apenas a comenzado a ser reconociday desarrollada.
Medidas de diversidad trófica. Entender el gradode especialización o generalización en indivi-duos y especies es importante pues puedeinformar de problemas tan diversos como laevolución del uso de recursos en ambientescambiantes (Bolnick et al. 2003, Emslie & Pat-terson 2007), el éxito de aves exóticas invaso-ras (Holt et al. 2005), y los factores,mecanismos, y procesos que le dan forma alas comunidades bióticas (Wiens & Graham2005). Para entender el nivel de especializa-ción o generalización, los ecólogos han utili-zado dos aproximaciones: 1) generar diversosestimadores para valorar la variación en laamplitud del nicho, y 2) evaluar la contribu-ción relativa de la variación dentro y entreindividuos a la magnitud de estos estimadores(Bolnick et al. 2002). Bearhop et al. (2004) hanpropuesto el uso de la varianza en la composi-ción isotópica como un estimador de laamplitud el nicho trófico. Layman et al. (2006)han elaborado esta propuesta y desarrolladométodos multivariados para investigar medi-das de diversidad trófica en individuos ycomunidades.
Newsome et al. (2007) reconocen un pro-blema potencialmente serio del uso de medi-das isotópicas en la estimación de la amplitudde nicho. La variación isotópica en animalesdepende fuertemente de que tan distintasson las señales isotópicas de las fuentes dealimento. Por ejemplo, un ave en un ambi-ente dulceacuícola puede alimentarse de una
FIG. 3. En 3 especies de Cinclodes (Furnariidae), lascaracterísticas del riñón responsables de la produc-ción de orina concentrada están positivamentecorrelacionadas con δ13C (las correlaciones estáncorregidas para tomar en cuenta la estructura filo-genética de los datos). δ13C mide la dependenciarelativa que cada especie tiene por alimento prove-niente de fuentes marinas y por lo tanto, la cargaosmótica que acompaña a la dieta de cada especie.

107
ISÓTOPOS ESTABLES Y AVES
combinación de invertebrados que depen-den de una trama trófica basada en me-tano (δ 13C ≈ -60‰, Kohzu et al. 2004) o unabase trófica basada en detritus terrestre(δ 13C ≈ -25‰). La varianza isotópica en lostejidos de esta ave será siempre mayor quela de un ave terrestre que usa insectosherbívoros que se alimentan de plantas C3(δ 13C ≈ -25‰) y C4 (δ
13C ≈ -12‰). A pesarde que ambas especies usan solamente dostipos de recursos isotópicamente distintos,una de las especies siempre tendrá varianzasmayores en virtud de la diferencia mayor entrela composición de las fuentes. Newsome et al.(2006) proponen una solución a este pro-blema.
En lugar de usar valores de δ per se paradefinir nichos isotópicos, estos autores propo-nen usar modelos de mezclado para transfor-mar estos valores a proporciones (p) de cadafuente istópicamente distinta. Resumiendo,uno puede determinar la contribución de n +1 fuentes isotópicamente distintas resolviendoun sistema de n + 1 ecuaciones lineales (verPhillips & Gregg 2001). Esta transformaciónde espacio-δ a espacio-p, resuelve las discre-pancias en la escala que acabamos de descri-bir, y permite usar las medidas de amplitud denicho mas comúnmente usadas por los ecólo-gos (Bolnick et al. 2002, Layman et al. 2006).Enfatizamos que las descripciones del “nichoisotópico” en espacio-p y espacio-δ son com-plementarias, y no alternativas excluyentes. Altransformar los datos de valores de δ a valoresde p, ganamos la capacidad de construirmétricas que son independientes del valorabsoluto de las señales isotópicas de los recur-sos, y que son comparables a otras formula-ciones de nicho, pero perdemos informaciónsobre los tipos de recursos y la posición en losisopaisajes que revelan los espacios δ.
La cinética de la incorporación isotópica. El análisisde isótopos estables es especialmente apro-piado para investigar los componentes intra- e
inter-individuales de variación en el nicho.Debido a que diferentes tipos de tejidosincorporan señales isotópicas a distintas tasas,la información que nos aportan representadiferentes escalas temporales. Esta es unaventaja significativa sobre los métodos tradi-cionales de análisis de dieta, como las obser-vaciones de forrajeo, o el análisis decontenidos estomacales (Dalerum & Angerb-jörn 2005). Por ejemplo, las proteínas en elplasma incorporan la señal isotópica de ladieta mas rápidamente que las células de lasangre. En aves, la vida media (τ) de unamolécula de N o C en plasma es del orden dedías, la de las células en sangre es del orden dedecenas de días, mientras que la del colágenode los huesos es del orden de meses (Hobson& Clark 1992a, Pearson et al. 2003, Podlesak etal. 2005). Los valores exactos de la vida mediade C y N en estos tejidos están determinados,en buena medida, por la masa corporal de lasaves (Carleton & Martínez del Rio 2005). Lasplumas, son generadas durante la muda ymantienen la señal isotópica de los recursosusados durante su crecimiento. El uso devarios tejidos, medidos en un solo momento,puede proporcionar información muy ricasobre los cambios temporales en el uso derecursos de un animal. Langin et al. (2006)midieron la composición isotópica del plasmay células de la sangre en S. ruticilla reciente-mente llegados a Canada desde sus áreas deinvernación en Jamaica. El plasma de estosanimales adquirió la señal isotópica de losrecursos de la zona de reproducción, mientrasque las células de la sangre mantuvieron laseñal de los recursos usados en la zona deinvernación. Norris et al. (2005) usaron losdatos de δ13C en células sanguíneas para mos-trar que, durante el invierno, las hembras de S.ruticilla tienden a usar hábitats mas secos quelos machos.
Comparar la composición isotópica detejidos “lentos” y “rápidos” puede tambiéngenerar datos sobre el “tamaño de grano”

108
PÉREZ ET AL.
del patrón de forrageo (en el sentido deMacArthur & Levins 1964). Los forrajeadoresde grano fino usan recursos isotópicamentedistintos en rápida sucesión y, por tanto,tejidos con tasas de incorporación contrastan-tes deben tener la misma composición isotó-pica. En cambio, los forrajeadores de granogrueso se especializan temporalmente en unsolo recurso y, por lo tanto, esperamosdiferencias entre los tejidos rápidos y loslentos que incorporan la señal promediopor un intervalo temporal largo. A nuestroentender, estas ideas no han sido puestas aprueba.
AGRADECIMIENTOS
Agradecemos a Gabriel Bowen por producirla Figura 2. CMR fue apoyado por un grant deNSF (IBN 0421738). CMR agradece a losorganizadores del VIII NOC la invitación adar la plenaria que dio origen a este docu-mento.
REFERENCIAS
Arneson, L. S., & S. E. MacAcoy. 2005. Carbon,nitrogen, and sulfur diet-tissue discriminationin mouse tissues. Can. J. Zool. 83: 989–995.
Austin, M. P. 2002. Spatial prediction of speciesdistribution: an interface between ecologicaltheory and statistical modeling. Ecol. Mod.157: 101–108.
Bearhop S., C. E. Adams, S. Waldron, R. A. Fuller,& H. Macleod. 2004. Determining trophicniche width: a novel approach using stable iso-tope analysis. J. Anim. Ecol. 73: 1007–1012.
Begon, M., C. Townsend, & J. L. Harper. 2006.Ecology: from individuals to ecosystems.Blackwell Publishing, Oxford, UK.
Bolnick D. I., L. H. Yang, J. A. Fordyce, J. M. Davis& R. Svanbäck. 2002. Measuring individual-level resource specialization. Ecology 83:2936–2941.
Bolnick D. I., R. Svanback, J. A. Fordyce, L. H.Yang, J. M. Davis, C. D. Hulsey, & M. L. Foris-ter. 2003. The ecology of individuals: Incidence
and implications of individual specialization.Am. Nat. 161: 1–28.
Bowen, G. J., L. I. Wassenaar, & K. A. Hobson.2005. Global application of stable hydrogenand oxygen isotopes to wildlife forensics.Oecologia 143: 337–348.
Carleton, S. A., & C. Martínez del Rio. 2005. Theeffect of cold-induced increased metabolic rateon the rate of 13C and 15N incorporation inHouse Sparrows (Passer domesticus). Oecologia144: 226–232.
Cerling, T. E., & J. M. Harris. 1999. Carbon isotopefractionation between diet and bioapatite inungulate mammals and implications for ecolog-ical and paleoecological studies. Oecologia 120:347–363.
Chamberlain, C. P., J. D. Blum, R. T. Holmes, X.Feng, T. W. Sherry, & G. R. Graves. 1997. Theuse of isotope tracers for identifying popula-tions of migratory birds. Oecologia 109: 132–141.
Chase, J. M., & M. A. Liebold. 2003. Ecologicalniches: linking classical and contemporaryapproaches. Univ. of Chicago Press, Chicago,Illinois.
Dalerum F., & A. Angerbjörn. 2005. Resolvingtemporal variation in vertebrate diets using nat-urally occurring stable isotopes. Oecologia 144:647–658.
Dawson, T. E., S. Mambelli, A. H. Plamboeck, P.H. Templer, & K. P. Tu. 2002. Stable isotopesin plant ecology. Ann. Rev. Ecol. Syst. 33: 507–559.
DeNiro, M. S., & S. Epstein. 1978. Influence ofdiet on the distribution of carbon isotopes inanimals. Geochim. Cosmochim. Acta 42: 495–506.
DeNiro, M. S., & S. Epstein. 1981. Influence ofdiet on the distribution of nitrogen isotopes inanimals. Geochim. Cosmochim. Acta: 45: 341–351.
Elith J., C. H. Graham, R. P. Anderson, M. Dudík,S. Ferrier, A. Guisan, R. J. Hijmans, F. Huett-mann, J. R. Leathwick, A. Lehmann, J. Li, L. G.Lohmann, B. A. Loiselle, G. Manion, C. Moritz,M. Nakamura, Y. Nakazawa, J. M. Overton, A.T. Peterson, S. J. Phillips, K. S. Richardson, R.Scachetti-Pereira, R. E. Schapire, J. Soberón, S.Williams, M. S. Wisz, & N. E. Zimmermann.

109
ISÓTOPOS ESTABLES Y AVES
2006. Novel methods improve prediction ofspecies’ distributions from occurrence data.Ecography 29: 129–151.
Emslie, S. D., & W. P. Patterson. 2007. Abruptrecent shift in d13C and d15N values in AdéliePenguin eggshell in Antarctica. Proc. Natl.Acad. Sci. 104: 11666–11669.
Farmer, A., R. Rye, G. Landis, C. Bern, C. Kester,& I. Ridley. 2003. Tracing the pathways of Neo-tropical migratory songbirds using stable iso-topes: a pilot study. Isot. Environ. Health Stud.39: 169–177.
Farquhar, G. D., J. R. Ehleringer, & K. T. Kubick.1989. Carbon isotope discrimination and pho-tosynthesis. Ann. Rev. Plant Physiol. Plant Mol.Biol. 40: 503–537.
Finlay, J. C., M. E. Power, & G. Cabana. 1999.Effects of water velocity on algal carbon iso-tope ratios: implications for river food webstudies. Limnol. Oceanogr.44: 1198–1203.
France, R. L. 1995. Differentiation between littoraland pelagic food webs in lakes using stable iso-topes. Limnol. Oceanogr. 40: 1310–1313.
Fry, B. 1988. Food web structure on George Bankfrom stable C, N, and S isotopic compositions.Limnol. Oceanogr. 33: 1182–1190.
Fry, B. 2006. Stable isotope ecology. Springer-Ver-lag Press, New York, New York.
Fry, B., & B. E. Sherr. 1989. 13C measurements asindicators of carbon flow in marine and fresh-water ecosystems. Pp. 196–229 in Rundel, P. W.,J. R. Ehleringer, & K. A. Nagy (eds.). Stable iso-topes in ecological research. Springer-Verlag,Berlin, Germany.
Fry, B., R. K. Anderson, L. Entzeroth, J. L. Bird, &P. L. Parker. 1984. 13C enrichment and oceanicfood web structure in the northwest Gulf ofMexico. Contrib. Mar. Sci. 27: 49–63.
Grinnell, J. 1917. The niche-relationships of theCaliforia thrasher. Auk 34: 427–433.
Harding, E. K., & E. Stevens. 2004. Using stableisotopes to assess seasonal patterns of avianpredation across terrestrial-marine landscape.Oecologia 129: 436–444.
Herrera, M. L. G., K. A. Hobson, M. Rodríguez, &P. Hernandez. 2003. Trophic partitioning intropical rain forest birds: insight from stableisotope analysis. Oecologia 136: 439–444.
Herrera, M. L. G., K. A. Hobson, J. C. Martínez, &
G. Mendez. 2006. Tracing origins of dietaryprotein in tropical dry forest birds. Biotropica38: 735–742.
Hobson, K. A. 1999. Tracing origins and migrationof wildlife using stable isotopes: a review.Oecologia 120: 314–326.
Hobson, K. A. 2003. Making migratory connec-tions with stable isotopes. Pp. 379–391 in Ber-thold, P., E. Gwinner, & E. Sonnenschein(eds.). Avian migration. Springer-Verlag, Ber-lin, Germany.
Hobson, K. A. 2005. Stable isotopes and the deter-mination of avian migratory connectivity andseasonal interactions. Auk 122: 1037–1048.
Hobson, K. A., & R. G. Clark. 1992a. Assessingavian diets using stable isotopes I: Turnover of13C in tissues. Condor 94: 181–188.
Hobson, K. A., & R. G. Clark. 1992b. Assessingavian diets using stable isotopes II: factorsinfluencing diet-tissue fractionation. Condor94: 189–197.
Hobson, K. A., & L. I. Wassenaar. 1997. Linkingbreeding and wintering grounds of Neotropicalmigrant songbirds using stable hydrogen isoto-pic analysis of feathers. Oecologia 109: 142–148.
Hobson, K. A., & L. I. Wassenaar. 2001. Isotopicdelineation of North American migratory wild-life populations: Loggerhead Shrikes. Ecol.Appl. 11: 1545–1553.
Hobson, K. A., J. F. Piatt, & J. Pitocchelli. 1994.Using stable isotopes to determine seabirdtrophic relationships. J. Anim. Ecol. 63: 786–798.
Hobson, K. A., L. I. Wassenaar, B. Milá, I. Lovette,C. Dingle, & T. B. Smith. 2003. Stable isotopesas indicators of altitudinal distribution andmovements in an Ecuadorian hummingbirdcommunity. Oecologia. 134: 302–308.
Hobson, K. A., G. J. Bowen, L. I. Wassenaar, Y.Ferrand, & H. Lormee. 2004. Using stablehydrogen and oxygen isotope measurements offeathers to infer geographical origins of migrat-ing European birds. Oecologia 141: 477–488.
Hoeffs, J. 2004. Stable isotope geochemistry.Springer-Verlag, New York, New York.
Holt, R. D., M. Barfield, & R. Gomulkiewicz. 2005.Theories of niche conservatism and evolution:could exotic species be potential tests? Pp. 259–

110
PÉREZ ET AL.
290 in Sax, D., J. Stachowicz, & S. D. Gaines(eds.). Species invasions: insights into ecology,evolution, and biogeography. Sinauer Associ-ates, Sunderland, Massachusetts.
Hutchinson G. E. 1957. Concluding remarks. ColdSpring Harbor Symp. Quant. Biol. 22: 415–427.
Hutchinson G. E. 1978. An introduction to popu-lation biology. Yale Univ. Press, New Haven,Connecticut.
Karasov, W. H., & C. Martínez del Rio. 2007. Phys-iological ecology: How animals process energy,nutrients, and toxins. Princeton Univ. Press,Princeton, New Jersey.
Kearney, M. 2006. Habitat, environment, andniche: What are we modeling? Oikos 115: 186–191.
Kelly, J. F. 1999. Stable isotopes of carbon andnitrogen in the study of avian and mammaliantrophic ecology. Can. J. Zool. 78: 1–27.
Koch, P. L., J. Heisinger, C. Moss, R. W. Carlson, &M. L. Fogel. 1995. Isotopic tracking of changein diet and habitat use in African elephants. Sci-ence 267: 1340–1343.
Kohzu, A., C. Kato, T. Iwata, D. Kishi, M.Murakami, N. Nakano, & E. 2004. Stream foodweb fueled by methane-derived carbon. Aquat.Microb. Ecol. 36: 189–194.
Lajtha, K., & J. D. Marshall. 1994. Sources of varia-tion in the stable isotope composition ofplants. Pp. 1–21 in Lajtha, K., & R. H.Michener (eds.). Stable isotopes in ecology andenvironmental sciences. Blackwell Scientific,Oxford, UK.
Langin, K. M., D. R. Norris, T. K. Kyser, P. P.Marra, & L. M. Ratcliffe. 2006. Capital versusincome breeding in a migratory passerine bird:evidence from stable-carbon isotopes. Can. J.Zool. 87: 947–953.
Langin, K. M., M. W. Reudink, P. P. Marra, D. R.Norris, T. K. Kyser, & L. M. Ratcliffe. 2007.Hydrogen isotopic variation in migratory birdtissues of known origin: implications for geo-graphic assignment. Oecologia 152: 449–457.
Layman, C. A., D. A. Arrington, C. G. Montaña, &D. M. Post. 2006. Can stable isotope ratios pro-vide for community-wide measures of trophicstructure. Ecology 88: 42–48.
MacArthur, R., & R. Levins. 1964. Competitionhabitat selection and character displacement in
a patchy environment. Proc. Natl. Acad. Sci.51: 1207–1210.
Martin, K. 1995. Patterns and mechanisms for age-dependent reproduction and survival in birds.Am. Zool. 38: 340–348.
Martínez del Rio, C., & B. O. Wolf. 2004. Mass-bal-ance models for animal-isotopic ecology. Pp.141–174 in Stack, M., & T. Wang (eds.). Physio-logical and ecological adaptations to feeding invertebrates. Science Publishers, Enfield, NewHampshire.
Marra, P. P., K. A. Hobson, & R. T. Holmes. 1998.Linking winter and summer events in migra-tory bird by using stable-carbon isotopes. Sci-ence 282: 1884–1886.
McKechnie, A. E., B. O. Wolf, & C. Martínez delRio. 2004. Deuterium stable isotope ratios astracers of water resource use: an experimentaltest with Rock Doves. Oecologia 140: 191–200.
McNab, B. K. 2002. Short-tern energy conserva-tion in endotherms in relation to body mass,habits and environment. J. Thermal Biol. 27:459–466.
Meehan, T. D., R. N. Rosenfield, V. N. Atudorei, J.Bielefeldt, L. J. Rosenfield, A. C. Stewart, W. E.Stout, W. E. Stout, & M. A. Bozek. 2003. Varia-tion in hydrogen stable-isotope ratios betweenadult and nestling Cooper's Hawks. Condor105: 567–572.
Meehan, T. D., J. T. Giermakowski, & P. M. Cryan.2004. A GIS-based model od stable hydrogenisotope ratios in North American growing-sea-son precipitation for the use in animal move-ment studies. Isot. Environ. Health Stud. 40:291–300.
Mirón, L. L., L. G. Herrera, N. Ramírez, & K. A.Hobson. 2006. Effect of diet quality on carbonand nitrogen turnover and isotopic discrimina-tion in blood of a New World nectavivorous. J.Exp. Biol. 209: 541–548.
Munshi-South, J., & G. S. Wilkinson. 2006. Dietinfluences life span in parrots (Psittaciformes).Auk 123: 108–118.
Newsome, S. D., M. A. Ethier, & D. Aurioles-Gambao. 2006. Using carbon and nitrogen iso-tope values to investigate maternal strategies innortheast Pacific Otariids. Mar. Mammal Sci. 2:556–572.
Newsome, S. D, C. Martínez del Rio, D. L. Phillips,

111
ISÓTOPOS ESTABLES Y AVES
& S. Bearhop. 2007. A niche for isotopic ecol-ogy. Front. Ecol. Environ. (in press).
Norris, D. R., P. P. Marra, T. K. Kyser, T. W. Sherry,& T. M. Ratcliffe. 2004. Tropical winter habitatlimits reproductive success on the temperatebreeding grounds in a migratory bird. Proc. R.Soc. Lond. B 271: 59–64.
Norris, D. R., P. P. Marra, T. K. Kyser, & L. M. Rat-cliffe. 2005. Tracking habitat use of a long-dis-tance migratory bird, the American RedstartSetophaga ruticilla, using stable-carbon isotopesin cellular blood. J. Avian Biol. 36: 164–170.
Norris, D. R., M. B. Wunder, & M. Boulet. 2006.Perspectives in migratory connectivity. Orni-thol. Monogr. 61: 79–88.
Pearson, R. G., & T. P. Dawson. 2003. Predictingthe impacts of climate change on the distribu-tion of species: are bioclimate envelope modelsuseful? Global Ecol. Biogeogr. 12: 361–371.
Pearson, S. F., D. J. Levey, C. H. Greenberg, & C.Martínez del Rio. 2003. Effects of elementalcomposition on the incorporation of dietarynitrogen and carbon isotopic signatures in anomnivorous songbird. Oecologia 135: 516–523.
Peterson, B. J., & B. Fry. 1987. Stable isotopes inecosystem studies. Ann. Rev. Ecol. Syst. 18:293–320.
Pérez, G. E., & K. A. Hobson. 2006. Isotopic eval-uation of interrupted molt in northern breed-ing populations of the Loggerhead Shrike.Condor 108: 877–886.
Pérez, G. E., & K. A. Hobson. 2007. Feather deu-terium measurements reveal origins of migra-tory western Loggerhead Shrikes (Laniusludovicianus) wintering in Mexico. Divers. Dis-trib. 13: 166–171.
Phillips, D., & J. W. Gregg. 2001. Uncertainty insource partitioning using stable isotopes. Oeco-logia 127: 171–179.
Phillips, D., & J. W. Gregg. 2003. Source partition-ing using stable isotopes: coping with too manysources. Oecologia 136: 261–269.
Phillips, D., & P. L. Koch. 2002. Incorporating con-centration dependence in stable isotopes mix-ing models. Oecologia 130: 114–125.
Podlesak, D. W., S. R. Williams, & K. A. Hatch.2005. Stable isotopes in breath, blood, fecesand feathers can indicate intra-individualchanges in the diet of migratory songbirds.
Oecologia 142: 501–512.Post D. M. 2002. Using stable isotopes to estimate
trophic position: models, methods, andassumptions. Ecology 83: 703–718.
Robbins, C. T., L. A. Felicetti, & M. Sponheimer.2005. The effect of dietary protein quality onnitrogen isotope discrimination in mammalsand birds. Oecologia 144: 534–540
Robinson, D. 2001. Š15N as an integrator of thenitrogen cycle. Trend Ecol. Evol. 16: 153–162.
Rubenstein, D. R., & K. A. Hobson. 2004. Frombirds to butterflies: animal movement patternsand stable isotopes. Trend Ecol. Evol. 19: 256–263.
Rubenstein, D. R., C. P. Chamberlain, R. T.Holmes, M. P. Ayres, J. R. Waldbauer, G. R.Graves, & N. C. Tuross. 2002. Linking breedingand wintering ranges of a migratory songbirdusing stable isotopes. Science 295: 1062–1065.
Rundel, P. W., J. R. Ehleringer, & K. R. Nagy. 1989.Stable isotopes in ecological research. Springer-Verlag, Berlin, Germany.
Sabat, P., K. Maldonado, & C. Martínez del Rio.2006a. Osmoregulatory capacity and the abilityto use marine food sources in two coastal song-birds (Cinclodes: Furnariidae) along a latitudinalgradient. Oecologia 148: 250–257.
Sabat, P., K. Maldonado, M. Canals, & C. Martínezdel Rio. 2006b. Osmoregulation and adaptiveradiation in the ovenbird genus Cinclodes (Passe-riformes: Furnariidae). Funct. Ecol. 20: 799–805.
Schondube, J. E., L. G. Herrera-M., & C. Martínezdel Rio. 2001. Diet and the evolution of diges-tion and renal function in phyllostomid bats.Zoology 104: 59–73.
Soberón, J. M. 2007. The Grinnelian and Eltonianniches and geographic distribution of species.Ecol. Lett. (in press).
Speakman, J. R. 2005. The role of technology in thepast and future development of the doublylabeled water method. Isot. Environ. HealthStud. 41: 335–343.
Sponheimer, M., J. A. Lee-Thorp, D. J. DeRuiter, J.M. Smith, N. J. van der Merwe, K. Reed, C. C.Grant, L. K. Ayliffe, T. F. Robison, C. Heidel-berger, & W. Marcus. 2003. Diets of southernAfrican Bovidae: stable isotope evidence. J.Mammal. 84: 471–479.

112
PÉREZ ET AL.
Stapp, P., & G. A. Polis. 2003. Marine resourcessubsidize insular rodent populations in theGulf of California, Mexico. Oecologia 134:496–504.
Tilman, D. 1988. Plant strategies and the dynamicsand structure of plant communities. Princeton,Princeton Univ. Press, Princeton, New Jersey.
Trueman, C. N., R. A. R. McGill, & P. H. Guyard.2005. The effect of growth rate on tissue dietisotopic spacing in rapidly growing animals. Anexperimental study on Atlantic salmon (Salmosalar). Rapid Commun. Mass Spectrom. 19:3239–3247.
Vander Klift, M. A., & S. Ponsard. 2003. Sourcesof variation in consumer-diet delta 15-Nenrichment: a meta-analysis. Oecologia 136:161–182.
Vander Zanden, J., & J. B. Rasmussen. 1999. Pri-mary consumer δ13C and δ15N and the trophicposition of aquatic consumers. Ecology 80:1395–1408.
Wassenaar, L. I., & K. A. Hobson. 2000. Improvedmethod for determining the stable hydrogenisotopic composition (δD) of complex organicmaterials of organic interest. Environ. Sci.Technol. 34: 2354–2360.
Webster, M. S., P. P. Marra, S. M. Haig, S. Bensch,& R. T. Holmes. 2002. Links between worlds:unraveling migratory connectivity. Trend Ecol.Evol. 17: 76–83.
Werner, R. A., & W. A. Brand. 2001. Referencingstrategies and techniques in stable isotope anal-ysis. Rapid Commun. Mass Spectrom. 15: 501–
519.West, J. B., G. J. Bowen, T. E. Cerling, & J. R.
Eheringer. 2006. Stable isotopes as one ofnature’s recorders. Trend Ecol. Evol. 21: 408–414.
Wiens, J. J., & C. H. Graham. 2005. Niche conser-vatism: integrating evolution, ecology, and con-servation biology. Ann. Rev. Ecol. Evol. Syst.36: 519–539.
Wolf, B., & C. Martínez del Rio. 2000. Use ofsaguaro fruit by White-winged Doves: Isotopicevidence of a tight ecological association.Oecologia 124: 536–543.
Wolf, B. O., & C. Martínez del Rio. 2004. Howimportant are CAM succulents as sources ofwater and nutrients for desert consumers? Areview. Isot. Environ. Health Stud. 39: 53–67.
Wolf, B. O., J. Babson, & C. Martínez del Rio. 2002.Saguaros, doves, and isotopes: differentialwater and carbon acquisition by animals feed-ing on a single resource. Ecology 83: 1286–1293.
Yapp, C. J., & S. Epstein. 1982. A re-examinationof cellulose carbon-bound hydrogen D mea-surements and some factors affecting plant-water D/H relationships. Geochim. Cosmo-chim. Acta 46: 955–965.
Yohannes, E., K. A. Hobson, & D. J. Pearson.2007. Feather stable-isotope profiles revealstopover habitat selection and site fidelity innine migratory species moving through sub-Saharan Africa. J. Avian Biol. 38: 347–355.




















