
Isolation and characterization of a 28kDa major allergen from blackgram (Phaseolus mungo)
10
Immunobiology 217 (2012) 895–904 Contents lists available at SciVerse ScienceDirect Immunobiology j ourna l homepage: www.elsevier.de/imbio Isolation and characterization of a 28 kDa major allergen from blackgram (Phaseolus mungo) Dolly Kumari a,1 , Naveen Arora a , Ramkrashan Kasera a,b , Susheela Sridhara a , Raj Kumar c , Bhanu Pratap Singh a,∗ a Allergy and Immunology Lab, CSIR-Institute of Genomics and Integrative Biology, Delhi 110007, India b Department of Biotechnology, University of Pune, Ganeshkhind, Pune 411007, India c Department of Respiratory Allergy and Applied Immunology, V.P. Chest Institute, Delhi 110007, India a r t i c l e i n f o Article history: Received 11 January 2011 Received in revised form 30 December 2011 Accepted 30 December 2011 Keywords: Blackgram Allergy 28 kDa protein Skin prick tests Histamine a b s t r a c t Legumes are the major elicitors of IgE-mediated food allergy in many countries of the world. Purified major allergens are prerequisite for component resolved diagnosis of allergy. The present study was aimed to isolate and characterize a major allergenic protein from blackgram (Phaseolus mungo). Res- piratory allergy patients with history of blackgram allergy were skin prick tested (SPT) and sera were collected from SPT positive patients. The blackgram extract was fractionated using a combination of anion exchange and hydrophobic interaction chromatography. The purified protein was characterized by indirect ELISA, immunoblot, ELISA inhibition, SPTs, stripped basophil histamine release, lymphopro- liferation assay and digestibility assay. The purified protein separated at 28 kDa on 12% gel and showed IgE binding with 81% of blackgram hypersensitive patients’ sera on immunoblot indicating it to be a major allergen. Periodic Acid Schiff’s and meta-periodate treatment staining detected it to be a glyco- protein. The 28 kDa protein recognized 7/9 (77.8%) of blackgram positive patients by SPT, where as all 9 patients showed significant histamine release on stimulation with protein as compared to controls. The 28 kDa protein remained stable up to 15 min on incubation with SGF. Bands of 14–16 kDa appeared after 15 min of pepsin digestion that remained stable up to 60 min of incubation. However, purified protein degraded within 5 min after incubation with SIF. The N-terminus-12 residues sequence of 28 kDa protein was GRREDDYDNLQL. A stretch of residues ‘DDYDNLQL’ showed homology with Rho-specific inhibitor of transcription termination (E = 0.42, Identity = 87%) and NBS-LRR type disease resistant protein from peanut (Arachis hypogaea) (E = 2, Identity = 77%). In conclusion, the purified 28 kDa protein is a potent major allergen that may have implication in diagnosis of blackgram allergy. © 2012 Elsevier GmbH. All rights reserved. Introduction Legumes such as peanut, soybean, lentil and chickpea have been recognized as important triggers of food allergy in the US, Europe and Asian countries (Ibanez Sandin et al. 1999; Lin et al. Abbreviations: SPT, skin prick test; CRD, component-resolved diagnosis; SDS- PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; CBB, Coomassie brilliant blue; NCM, nitrocellulose membrane; PBMCs, peripheral blood mononu- clear cells; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; SGF, simulated gastric fluid; SIF, simulated intestinal fluid; BSA, bovine serum albu- min; PAS, Periodic Acid Schiff’s; PVDF, polyvinylidene difluoride membrane. ∗ Corresponding author at: Allergy and Immunology Section, Institute of Genomics and Integrative Biology, Mall Road, Delhi University Campus, Delhi 110007, India. Tel.: +91 11 2766156. E-mail address: [email protected] (B.P. Singh). 1 Present address: Department of Microbiology and Biotechnology, Rajasthan University of Veterinary and Animal Science, Bikaner 334001, India. 2006; Maleki et al. 2010; Patil et al. 2001). Ingestion or inhala- tion of vapours during cooking of legumes had provoked diarrhea, angioedema, or severe anaphylactic reactions in predisposed indi- viduals (Burks et al. 1999; Kalogeromitros et al. 1996; Martin et al. 1992; Niphadkar et al. 1997; Sicherer et al. 2000; Wassenberg and Hofer 2007). Blackgram (Phaseolus mungo), a protein rich legume is extensively used for preparing many delicious dishes worldwide and is identified as a major food allergen in asthma and rhinitis patients (Kumari et al. 2006). Many food allergens from legume family have been isolated and characterized in the past decades. These include Ara h 1–Ara h 8 from peanut, Len c 1 and Len c 2 from lentil and Gly m 1–Gly m 4 from soybean and Pis s 1 and Pis s 2 from pea (Breiteneder and Ebner 2000; Burks et al. 1991, 1992, 1998; Lopez-Torrejon et al. 2003; Mittag et al. 2004a). The availability of purified allergen components, makes it possible to delineate the patient’s individual IgE reactivity profile and to identify the disease-causing allergen molecules i.e. component-resolved diagnosis (CRD) (Heiss et al. 0171-2985/$ – see front matter © 2012 Elsevier GmbH. All rights reserved. doi:10.1016/j.imbio.2011.12.011
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Isolation and characterization of a 28kDa major allergen from blackgram (Phaseolus mungo)

I(
DRa
b
c
a
ARR3A
KBA2SH
I
bE
PbcSm
G1
U
0d
Immunobiology 217 (2012) 895– 904
Contents lists available at SciVerse ScienceDirect
Immunobiology
j ourna l homepage: www.elsev ier .de / imbio
solation and characterization of a 28 kDa major allergen from blackgramPhaseolus mungo)
olly Kumaria,1, Naveen Aroraa, Ramkrashan Kaseraa,b, Susheela Sridharaa,aj Kumarc, Bhanu Pratap Singha,∗
Allergy and Immunology Lab, CSIR-Institute of Genomics and Integrative Biology, Delhi 110007, IndiaDepartment of Biotechnology, University of Pune, Ganeshkhind, Pune 411007, IndiaDepartment of Respiratory Allergy and Applied Immunology, V.P. Chest Institute, Delhi 110007, India
r t i c l e i n f o
rticle history:eceived 11 January 2011eceived in revised form0 December 2011ccepted 30 December 2011
eywords:lackgramllergy8 kDa proteinkin prick testsistamine
a b s t r a c t
Legumes are the major elicitors of IgE-mediated food allergy in many countries of the world. Purifiedmajor allergens are prerequisite for component resolved diagnosis of allergy. The present study wasaimed to isolate and characterize a major allergenic protein from blackgram (Phaseolus mungo). Res-piratory allergy patients with history of blackgram allergy were skin prick tested (SPT) and sera werecollected from SPT positive patients. The blackgram extract was fractionated using a combination ofanion exchange and hydrophobic interaction chromatography. The purified protein was characterizedby indirect ELISA, immunoblot, ELISA inhibition, SPTs, stripped basophil histamine release, lymphopro-liferation assay and digestibility assay. The purified protein separated at 28 kDa on 12% gel and showedIgE binding with 81% of blackgram hypersensitive patients’ sera on immunoblot indicating it to be amajor allergen. Periodic Acid Schiff’s and meta-periodate treatment staining detected it to be a glyco-protein. The 28 kDa protein recognized 7/9 (77.8%) of blackgram positive patients by SPT, where as all 9patients showed significant histamine release on stimulation with protein as compared to controls. The28 kDa protein remained stable up to 15 min on incubation with SGF. Bands of 14–16 kDa appeared after
15 min of pepsin digestion that remained stable up to 60 min of incubation. However, purified proteindegraded within 5 min after incubation with SIF. The N-terminus-12 residues sequence of 28 kDa proteinwas GRREDDYDNLQL. A stretch of residues ‘DDYDNLQL’ showed homology with Rho-specific inhibitorof transcription termination (E = 0.42, Identity = 87%) and NBS-LRR type disease resistant protein frompeanut (Arachis hypogaea) (E = 2, Identity = 77%). In conclusion, the purified 28 kDa protein is a potentmajor allergen that may have implication in diagnosis of blackgram allergy.ntroduction
Legumes such as peanut, soybean, lentil and chickpea haveeen recognized as important triggers of food allergy in the US,urope and Asian countries (Ibanez Sandin et al. 1999; Lin et al.
Abbreviations: SPT, skin prick test; CRD, component-resolved diagnosis; SDS-AGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; CBB, Coomassierilliant blue; NCM, nitrocellulose membrane; PBMCs, peripheral blood mononu-lear cells; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide;GF, simulated gastric fluid; SIF, simulated intestinal fluid; BSA, bovine serum albu-in; PAS, Periodic Acid Schiff’s; PVDF, polyvinylidene difluoride membrane.∗ Corresponding author at: Allergy and Immunology Section, Institute ofenomics and Integrative Biology, Mall Road, Delhi University Campus, Delhi10007, India. Tel.: +91 11 2766156.
E-mail address: [email protected] (B.P. Singh).1 Present address: Department of Microbiology and Biotechnology, Rajasthanniversity of Veterinary and Animal Science, Bikaner 334001, India.
171-2985/$ – see front matter © 2012 Elsevier GmbH. All rights reserved.oi:10.1016/j.imbio.2011.12.011
© 2012 Elsevier GmbH. All rights reserved.
2006; Maleki et al. 2010; Patil et al. 2001). Ingestion or inhala-tion of vapours during cooking of legumes had provoked diarrhea,angioedema, or severe anaphylactic reactions in predisposed indi-viduals (Burks et al. 1999; Kalogeromitros et al. 1996; Martin et al.1992; Niphadkar et al. 1997; Sicherer et al. 2000; Wassenberg andHofer 2007). Blackgram (Phaseolus mungo), a protein rich legumeis extensively used for preparing many delicious dishes worldwideand is identified as a major food allergen in asthma and rhinitispatients (Kumari et al. 2006).
Many food allergens from legume family have been isolated andcharacterized in the past decades. These include Ara h 1–Ara h8 from peanut, Len c 1 and Len c 2 from lentil and Gly m 1–Glym 4 from soybean and Pis s 1 and Pis s 2 from pea (Breitenederand Ebner 2000; Burks et al. 1991, 1992, 1998; Lopez-Torrejon
et al. 2003; Mittag et al. 2004a). The availability of purified allergencomponents, makes it possible to delineate the patient’s individualIgE reactivity profile and to identify the disease-causing allergenmolecules i.e. component-resolved diagnosis (CRD) (Heiss et al.
8 obiolo
1uttede
lawnicc
M
P
t(e1ae
S
sOpsitgfs
P
erTuilrPuofe(opwt7uat
96 D. Kumari et al. / Immun
999; Hage-Hamsten et al. 2002). Purified allergens can also besed to supplement the allergen-extract with the relevant allergenshat are under-represented (spiking of extracts), thereby enhancinghe sensitivity of the extract (Ballmer-Weber et al. 2007). Soybeanxtract spiked by purified recombinant Gly m 4 showed higheriagnostic sensitivity than that of native extract-based test (Mittagt al. 2004b).
Previously, IgE mediated food allergy to blackgram was estab-ished by double blind placebo controlled food challenge test insthma and rhinitis patients (Kumari et al. 2006). Immunoblottingith hypersensitive patients’ sera detected 14 IgE-binding compo-ents with 8 major allergens of 78, 56, 47, 43, 40, 30, 28 and 16 kDa
n blackgram extract. The present study was aimed to isolate andharacterize a major allergenic protein of blackgram using columnhromatography and standard immunobiochemical methods.
ethods
reparation of extracts
Raw blackgram seeds were crushed, powdered and then defat-ed in diethyl ether at 4 ◦C. The extraction was carried out in 1:20w/v) ammonium bicarbonate buffer (50 mM, pH 8) as reportedarlier (Kumari et al. 2006). The extracts were centrifuged at0,000 × g, supernatant was filtered using a 0.22 �m membrane,nd freeze dried in small aliquots. Protein content of extracts wasstimated by Lowry’s method (Lowry et al. 1951).
tudy subjects
Respiratory allergy patients with history of food allergy werekin prick tested (SPT) with black gram (Vigna mungo) at theut Patients Department, V.P. Chest Institute, Delhi. Sera fromatients (n = 22) with marked positive skin reactions and elevatedpecific IgE to black gram extract were collected for immunolog-cal experiments. The present study protocol was approved byhe Human Ethics Committee of Institute of Genomics and Inte-rative Biology, Delhi. Informed written consent was obtainedrom patients and nonallergic volunteers for participation in thetudy.
urification of an allergenic protein from blackgram
The whole seed black gram extract was subjected to an ionxchange chromatography using fast protein liquid chromatog-aphy. Lyophilized protein (50 mg) was reconstituted in 20 mMris buffer (pH 7.9) and loaded onto 5 ml Q-Sepharose gel col-mn equilibrated with same buffer. After washing unbound protein
n equilibration buffer, the bound proteins were eluted with ainear salt gradient of 0–600 mM NaCl. All the fractions of eachesolved peak were pooled, dialyzed, lyophilized and run on SDS-AGE. The fractions were tested for IgE-binding on immunoblotsing pooled patients’ sera. Fractions containing a distinct bandf 28 kDa and two faint bands of 47- and 16 kDa were subjectedurther to hydrophobic interaction chromatography. The 28 kDanriched fractions were dialyzed and equilibrated with Tris buffer20 mM, pH 7.9) containing 1 M ammonium sulfate. This was loadednto Phenyl Sepharose column equilibrated with 20 mM Tris buffer,H 7.9 containing 1 M ammonium sulfate. The unbound proteinsere collected by washing the column with the same equilibra-
ion buffer. Bound proteins were eluted with 20 mM Tris buffer, pH.9. The unbound and eluted fractions were run on SDS-PAGE. Thenbound fraction containing 28 kDa protein was collected, dialyzedgainst PBS, filtered sterilized and finally lyophilized and reconsti-uted to protein concentration of 10 mg/ml.
gy 217 (2012) 895– 904
Specific IgE
Specific IgE binding against purified protein/whole seed black-gram extract were determined by indirect ELISA (Kumari et al.2006). Briefly, 200 ng of purified protein and/or 1 �g of black-gram extract in 100 �l of carbonate-bicarbonate buffer (pH 9.6)was coated per well, overnight at 4 ◦C (Nunc maxisorp plates,Nalge Nunc International, Rosklide, Denmark). After washing withPBS–Tween 20 (0.05%), non specific sites were blocked with 3%defatted milk for 3 h at 37 ◦C. The plates were washed and incu-bated with 1:10 (v/v) sera of blackgram hypersensitive patients’ at4 ◦C. Serum from healthy volunteers were used as negative control.The plates were washed and incubated with anti-human IgE-horseradish peroxidase 1:1000 (v/v) (Sigma) for 3 h at 37 ◦C. Color wasdeveloped with orthophenylene diamine. The reaction was stoppedafter 20 min by adding 5 N H2SO4 and the absorbance was read at492 nm.
ELISA inhibition
The allergenic potency of purified 28 kDa blackgram proteinwas determined by ELISA inhibition. Inhibition of IgE bindingwas assessed with serum pools of blackgram positive patients’preincubated with 0.1, 1, 10, 100, 1000 and 10,000 ng of 28 kDapurified protein and raw blackgram extract as inhibitors. The mix-ture was added to the solid phase bound raw blackgram extract inELISA plate. Here blackgram positive patients’ pooled sera withoutinhibitor was taken as a positive control. ELISA inhibition was alsocarried out using Curvularia lunata extract as an inhibitor. Percent-age inhibition was calculated as described below:
1 − OD of sample with inhibitorOD of sample without inhibitor
× 100
Sodium dodecyl polyacrylamide gel electrophoresis (SDS-PAGE)
Black gram extract (80 �g per lane) and purified protein (20 �gper lane) was loaded on a 12% reducing gel following proceduredescribed by Laemmeli (1970). The proteins were stained withCoomassie brilliant blue (CBB) or silver stained.
IgE immunoblot
SDS-PAGE resolved purified protein/blackgram extract wastransferred on to nitrocellulose membrane (NCM) as described ear-lier (Towbin et al. 1979). The unbound sites were blocked with 3%defatted milk for 3 h at 37 ◦C. The NCM strips were washed andincubated with 1:10 (v/v) blackgram hypersensitive patients’ serain PBS, overnight at 4 ◦C. A pool of normal human sera (n = 3) wasused as negative control. The strips were washed and incubatedwith 1:1000 (v/v) anti-human IgE peroxidase (Sigma). The IgE bind-ing was detected by adding 3,3-diaminobenzedene with hydrogenperoxidase in sodium acetate buffer (pH-5.0)
Skin test with blackgram extract and purified protein
Skin prick tests (SPTs) were carried out with blackgram extract(1: 500 (w/v) in PBS) and purified protein (100 �g/ml protein inPBS) at V.P. Chest Institute, Delhi on respiratory allergy patientswith history of food allergy. SPT was also performed with a panelof common inhalant allergens such as pollen, fungi and insects.Glycerinated (50%) phosphate buffer saline was used as a negative
control and histamine diphosphate (5 mg/ml) as a positive control.The skin tests were graded after 20 min in comparison to the whealdiameter of positive control. Wheal-diameter equal to that of pos-itive control or more (≥3 mm) were considered as marked positive
obiolo
sm
S
ow1Es
bstep
Lc
f(wsnpdvpipctcwmsop
Ek
D
i(mN1wwpybPmad
flm(
D. Kumari et al. / Immun
kin reactions. Blood was collected from patients (n = 9) showingarked positive reaction to blackgram extract and purified protein.
tripped basophil histamine release assay
The leukocyte suspension was prepared and basophils strippedff their IgE as described by Kleine Budde et al. (2001). The cellsere then passively sensitized by incubation (37 ◦C, 90 min) with
50 �l of hypersensitive individual patient’s serum (n = 9) in 4 mMDTA and 10 mg/ml heparin. As a control, basophils were also sen-itized with serum pool from healthy individuals (n = 3).
After sensitization, cells were incubated with purified protein orlackgram extract (10 ng/ml) diluted in HEPES buffer. Reaction wastopped by the addition of 750 �l of ice cold 0.9% NaCl (w/v). His-amine released was determined as described earlier (Kleine Buddet al. 2001). A histamine release of more than 10% was consideredositive.
ymphocyte proliferation and cytokine analysis on allergenhallenge
Peripheral blood mononuclear cells (PBMCs) were isolatedrom heparinized blood of blackgram hypersensitive patients’n = 9) by Ficoll Hypaque centrifugation (Sigma Chemical Co.). Cellsere washed 3 times with PBS and resuspended in RPMI-1640
upplemented with 23 mM sodium bicarbonate, 25 mM HEPES,on-essential amino acids and vitamins, streptomycin (100 U/ml),enicillin (100 �g/ml) and 10% fetal calf serum. Cell viability wasetermined by adding 20 �l of trypan blue to 20 �l of cells and theiable numbers of cells were adjusted to 106 cells/ml. For lympho-roliferation assay, 100 �l of cells (105 cells/well) were cultured
n RPMI media and challenged with 10 �g of blackgram extract orurified protein. Phytohaemagglutinin (10 �g) was used as positiveontrol and no stimulant was added to negative control. The cul-ure plates were incubated for 72 h and supernatant collected forytokine assay. Cells were washed with RPMI and incubated for 2 hith 3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bro-ide (MTT, 100 �g/ml). The cells were lysed in acidic isopropanol
olution and the absorbance read at 570 nm. OD values more thanr equal to three times of the negative control were considered asositive stimulation index.
IL-4 and IFN-� levels in culture supernatants were estimated byLISA following manufacturer’s protocol (BD Pharmingen OptEIAits).
igestibility of protein
The digestibility of purified protein and blackgram was exam-ned in simulated gastric fluid (SGF), as described by Astwood et al.1996). SGF was prepared as described in the United States Phar-
acopoea (Singh et al. 2006; Sharma et al. 2011) containing 0.03 MaCl, 0.32 (w/v) percentage of pepsin A (Sigma Chemical Co.), pH.2. Briefly, 28 kDa purified protein (680 �g) was treated separatelyith 200 �L of prewarmed SGF. Digestion was proceeded at 37 ◦Cith continuous shaking, and an aliquot (20 �L) of this digest waseriodically withdrawn (at 0.5, 1, 5, 15, 30, 45, and 60 min) for anal-sis. These aliquots were quickly mixed with 26 �L of a sampleuffer (containing 2% �-mercaptoethanol and 4% SDS) for SDS-AGE together with 6.0 �L of Na2CO3 solution (200 mM/L). Theixture was then boiled for 5 min and stored at −20 ◦C until further
nalyses. As control, each protein sample was treated with SGF thatid not contain pepsin A and then processed as described above.
The purified protein was also digested in simulated intestinaluid (SIF). SIF was prepared as described in the United States Phar-acopoea (Singh et al. 2006) containing 10 mg/ml of pancreatin
Sigma Chemical Co.) in 0.05 M KH2PO, pH 7.5. Purified protein was
gy 217 (2012) 895– 904 897
digested in SIF by incubation at 37 ◦C for 0.25, 1, 4, 8, 16 and 24 h. Theratio of pancreatin to test protein was about 13:1 (w/w). The reac-tion was immediately stopped by placing the tubes in boiling waterbath for 10 min. The end products were visualized by SDS-PAGE(Laemmeli 1970).
Protease activity
To determine the protease activity, 50 �g purified protein andblackgram extract (5 �g/�l in PBS) were incubated overnight at37 ◦C in the wells perforated in 0.1% agarose gel containing 50 �g ofbovine serum albumin (BSA), casein hydrolysate or gelatin individ-ually. A serine protease (Epi p 1) from mold Epicoccum purpurascenswas used as a positive control and PBS was used as a negative con-trol. The plates were stained with CBB. The lightly stained areaaround the wells indicates the proteolytic activity of protein.
Periodic Acid Schiff’s (PAS) staining
Periodic Acid Schiff’s (PAS) staining was used for detection ofcarbohydrate in purified protein and blackgram extract. For PASstaining, approximately 30 �g of purified protein or blackgramextract was run on SDS-PAGE. The gel was incubated in 1% peri-odic acid in 3% acetic acid solution for 15 min and stained in Schiff’sreagent for 15 min in dark. After staining, the gel was washed with5% sodium metabisulfite solution for 5 min followed by distilledwater to visualize the protein bands.
Effect of periodate oxidation on IgE binding
After electro-transfer of proteins onto NCM strip, it was incu-bated in dark with 20 mM sodium metaperiodate, overnight at 4 ◦C.Periodate was inactivated for 5–10 min by adding ethylene gly-col. To this, 1 mg/ml of sodium borohydride was added and keptovernight at 4 ◦C. The reaction was stopped by adding a drop ofacetic acid. The strip was washed with water and PBS and proteinfree sites were blocked with 3% defatted milk in PBS. The remainingsteps of western blot were carried out as described earlier (Towbinet al. 1979).
N-terminal amino acid sequencing
The purified protein was electrotransfered onto a polyvinyli-dene difluoride membrane (PVDF). After staining with 0.1% CBBand destaining, the protein band was cut and used for N-terminalamino acid sequencing on Protein Sequence Prosize 491 (PerkinElmer). The homology of the sequence obtained was determinedwith known proteins in the data bank using BLAST.
Statistical analysis
Correlation coefficients were determined by means of linearregression using the Graph pad programme. P value ≥ 0.05 wasconsidered significant.
Results
Purification of 28 kDa protein from blackgram extract
The blackgram extract was fractionated into five major peakson Q-sepharose gel (Fig. 1a). All fractions of each resolved peak
were pooled, dialyzed, lyophilized and run on SDS-PAGE. The pro-tein profile of unbound and eluted protein fractions is presented inFig. 1b. Peak 1 showed a distinct band of 28 kDa and two faint bandsof 47 and 16 kDa. In order to purify the proteins further, fractions of
898 D. Kumari et al. / Immunobiology 217 (2012) 895– 904
Fig. 1. (a) Elution profile of blackgram proteins on Q-sepharose anion exchange column. Unbound proteins were eluted with 60 ml of Tris buffer (20 mM, pH 7.9) at a flowrate of 2 ml/min. Bound proteins were eluted with a salt gradient from 0 to 0.6 M sodium chloride in Tris buffer (20 mM, pH 7.9) at 1 ml/min. OD at 280 nm (Left-Y-axis);N fractio1 f peak
ptasw
Ai
I
gp(s(OcIP
iv
in sera from all 9 skin test positive patients (ODs 0.404–0.918)
aCl gradient (0–0.6 M) (Right Y-axis). (b) Protein profile of anion exchange eluted
; lane 2, pool of peak 2; lane 3, pool of peak 3; lane 4, pool of peak 4; lane 5, pool o
eak 1 were loaded onto phenyl Sepharose column. The 28 kDa pro-ein appeared in the unbound fraction as a single band on SDS-PAGEnd was recognized by blackgram hypersensitive pooled patients’era on immunoblot (Fig. 2a and b). The yield of this 28 kDa proteinas 1.12 mg/g of blackgram protein.
llergenicity assessment of purified protein (28 kDa) byn vitro and in vivo methods
mmunoassays
Specific IgE binding of purified protein (28 kDa), and black-ram extract was determined by ELISA using 22 blackgram positiveatients’ sera. All 22 sera samples showed elevated IgE levelsODs 0.39–0.92) against blackgram extract whereas 18/22 of serahowed high IgE titer (ODs 0.29–0.63) against purified proteinTable 1). Specific IgE values ≥3 times of control (blackgram extractD 0.130, purified protein OD 0.088) were considered positiveut off for ELISA. There was a significant correlation of specificgE reactivity of purified protein and blackgram extract (r = 0.5626,
= 0.0064).IgE binding of 28 kDa protein was further assessed by
mmunoblotting with individual patients’ sera (n = 22) having ele-ated specific IgE levels to blackgram. Of these (n = 22), 18 (81.8%)
ns. MW, molecular weight marker; UB, unbound fraction; lane 1, fractions of peak 5. The protein bands were visualized by silver staining.
sera showed IgE binding with purified protein demonstrating itto be a major allergen (Fig. 3a). Four patients’ sera that did notshow IgE binding to this protein may be sensitized to other proteinfractions of blackgram.
Biopotency of 28 kDa protein
Out of 39 respiratory allergy patients skin prick tested with dif-ferent food extracts, 14 showed marked positive skin reactionswith one or more legumes. Of these (n = 14), 9 patients showedmarked positive skin reaction to blackgram extract while 7 patientswith purified 28 kDa protein. There was a significant correlationbetween skin reactivity of purified protein and blackgram extract(r = 0.72, P = 0.0035). Non allergic subjects did not demonstrate pos-itive skin reaction with any of the allergen extract. One patientwith positive SPT to purified protein showed negative skin reac-tion to the blackgram extract may be specifically sensitive to thisprotein.
Specific IgE levels against blackgram extract were elevated
whereas 7 patients demonstrated raised specific IgE titer (OD0.311–0.547) with purified protein in ELISA. A significant corre-lation was observed between the IgE response to purified proteinand blackgram extract (r = 0.68, P = 0.0071).
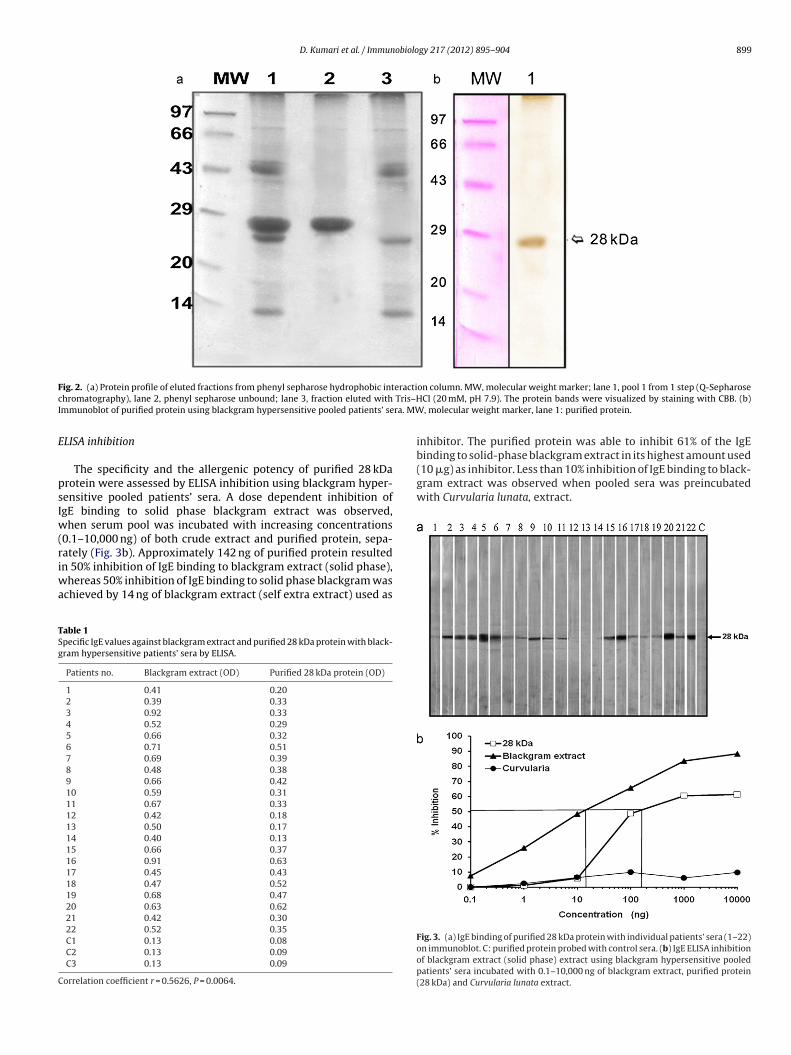
D. Kumari et al. / Immunobiology 217 (2012) 895– 904 899
F eractic Tris–HI ra. MW
E
psIw(riwa
TSg
C
binding to solid-phase blackgram extract in its highest amount used(10 �g) as inhibitor. Less than 10% inhibition of IgE binding to black-gram extract was observed when pooled sera was preincubatedwith Curvularia lunata, extract.
ig. 2. (a) Protein profile of eluted fractions from phenyl sepharose hydrophobic inthromatography), lane 2, phenyl sepharose unbound; lane 3, fraction eluted withmmunoblot of purified protein using blackgram hypersensitive pooled patients’ se
LISA inhibition
The specificity and the allergenic potency of purified 28 kDarotein were assessed by ELISA inhibition using blackgram hyper-ensitive pooled patients’ sera. A dose dependent inhibition ofgE binding to solid phase blackgram extract was observed,
hen serum pool was incubated with increasing concentrations0.1–10,000 ng) of both crude extract and purified protein, sepa-
ately (Fig. 3b). Approximately 142 ng of purified protein resultedn 50% inhibition of IgE binding to blackgram extract (solid phase),hereas 50% inhibition of IgE binding to solid phase blackgram waschieved by 14 ng of blackgram extract (self extra extract) used as
able 1pecific IgE values against blackgram extract and purified 28 kDa protein with black-ram hypersensitive patients’ sera by ELISA.
Patients no. Blackgram extract (OD) Purified 28 kDa protein (OD)
1 0.41 0.202 0.39 0.333 0.92 0.334 0.52 0.295 0.66 0.326 0.71 0.517 0.69 0.398 0.48 0.389 0.66 0.4210 0.59 0.3111 0.67 0.3312 0.42 0.1813 0.50 0.1714 0.40 0.1315 0.66 0.3716 0.91 0.6317 0.45 0.4318 0.47 0.5219 0.68 0.4720 0.63 0.6221 0.42 0.3022 0.52 0.35C1 0.13 0.08C2 0.13 0.09C3 0.13 0.09
orrelation coefficient r = 0.5626, P = 0.0064.
on column. MW, molecular weight marker; lane 1, pool 1 from 1 step (Q-SepharoseCl (20 mM, pH 7.9). The protein bands were visualized by staining with CBB. (b), molecular weight marker, lane 1: purified protein.
inhibitor. The purified protein was able to inhibit 61% of the IgE
Fig. 3. (a) IgE binding of purified 28 kDa protein with individual patients’ sera (1–22)on immunoblot. C: purified protein probed with control sera. (b) IgE ELISA inhibitionof blackgram extract (solid phase) extract using blackgram hypersensitive pooledpatients’ sera incubated with 0.1–10,000 ng of blackgram extract, purified protein(28 kDa) and Curvularia lunata extract.

900 D. Kumari et al. / Immunobiology 217 (2012) 895– 904
Fig. 4. (a) Histamine released from stripped basophils re-sensitized with individual patients’ sera (n = 9) on challenge with blackgram extract (�) and 28 kDa purified protein(�), separately. C1–C3: controls. (b) Lymphoproliferation assay. PBMCs from blackgram hypersensitive patients demonstrated significant proliferation when challenged with5 onalle5 egativw
S
heTfir
�g of blackgram (�) extract or purified 28 kDa protein (�), whereas PBMCs from n �g phytohemagglutinin (�) and without stimulation were taken as positive and nith blackgram extract (�) and 28 kDa purified protein (�). C1–C3: controls.
tripped basophil histamine release
The blackgram hypersensitive patients (n = 9) showed positiveistamine release upon challenge with 10 ng each of blackgram
xtract (31.41–65.98%) or purified protein (14.00–59.62%) (Fig. 4a).he controls (n = 3) with negative skin reactivity to both the puri-ed protein and blackgram extract demonstrated <10% histamineelease. Statistically significant correlation was observed betweenrgic controls (C1–C3) did not show significant proliferation. PBMCs stimulated withe controls (©). (c) TH2 cytokine IL-4 titer in the culture supernatant on stimulation
histamine release with purified protein and blackgram extract(r = 0.67, P = 0.048).
Lymphoproliferation assay
Lymphocyte proliferation was investigated in blackgram sensi-tized patients’ PBMCs by stimulation with 5 �g of purified proteinor blackgram extract. Purified 28 kDa protein induced significant

D. Kumari et al. / Immunobiology 217 (2012) 895– 904 901
Fig. 5. (a) Simulated gastric fluid (SGF) digestion of 28 kDa protein. MW, molecular weight marker; lane 1, pepsin; lane 2, 28 kDa protein (undigested); lanes 3–9, 28 kDa.Protein (34 �g) was incubated in SGF for 0.5, 1, 5, 10, 15, 30 and 60 min. The protein (digest) was electrophoresed on SDS-PAGE and visualized by silver staining. (b) Westernblot analysis of 28 kDa protein digested in SGF with serum pool of blackgram hypersensitive patients. Lane 1, pepsin (ponceau stained); lane 2, 28 kDa protein; lanes 3–9,purified protein incubated with SGF for 0.5, 1, 5, 10, 15, 30 and 60 min. (c) Simulated intestinal fluid (SIF) degradation of 28 kDa protein of blackgram. After electrophoresis, theprotein (digest) was visualized by silver staining. Mw, molecular weight marker; lane 1, pancreatin; lane 2, 28 kDa protein (untreated); lanes 3–9, purified protein incubatedwith SIF for 0.5, 1, 5, 10, 15, 30 and 60 min. (d) Western blot analysis of 28 kDa protein of blackgram digested with SIF with serum pool of blackgram hypersensitive patients.L cubat
ctOt(P
Iprw
B
D
wwm(ta(fs
ane 1, pancreatin; lane 2, 28 kDa protein (untreated); lanes 3–9, purified protein in
ellular proliferation (≥3 times of control) in 8 patients sensitizedo blackgram (Fig. 4b). OD values ranged from 0.088 to 0.577 (meanD 0.421 ± 0.09) on stimulation with purified protein as compared
o 0.323–0.699 with blackgram extract. However, no stimulationmean OD 0.06 ± 0.01) was observed in non allergic individuals’BMCs.
The purified 28 kDa protein also induced significantly higherL-4 release (mean 285 ± 78 pg/ml; range 121–413 pg/ml) as com-ared to controls (34–72 pg/ml) in cell culture supernatant. IL-4eleased on incubation with blackgram extract or purified proteinas comparable (r = 0.94, P < 0.05) (Fig. 4c).
iochemical characterization
igestion in SGF and SIF
The digestibility of blackgram and purified 28 kDa proteinas assessed by SGF and SIF. The 28 kDa protein was digestedith in 15 min treatment in SGF. After 15 min, 14–16 kDa frag-ents/polypeptides appeared that remained stable for 60 min
Fig. 5a). SDS-PAGE separated proteins (digests) were immunoblot-
ed with hypersensitive pooled patients’ sera. The 28 kDa proteinnd its fragments (14–16 kDa) showed IgE binding on immunoblotFig. 5b). The purified protein on digestion with SIF remained visibleor 15 min. (Fig. 5c) and showed IgE-reactivity with pooled patients’era on immunoblotting (Fig. 5d).ed with SIF for 0.5, 1, 5, 10, 15, 30 and 60 min.
Periodate treatment
The purified 28 kDa protein appeared to be a glycoprotein byPAS staining (Fig. 6a). On periodate treatment, the 28 kDa protein,showed a slight decrease in IgE binding with blackgram hypersen-sitive pooled patients’ sera on immunoblot (Fig. 6b).
The purified protein and blackgram extract did not show pro-teolytic activity on defatted milk (data not shown). Similar resultswere obtained with BSA, gelatin and casein hydrolysate. Purifiedprotein got cleaved by trypsin into ∼22 kDa fragment as visualizedon SDS-PAGE (data not shown).
N-terminal sequencing
N-terminal sequence of the 28 kDa purified protein showed 12residues GRREDDYDNLQL. In BLAST database, a stretch of residues‘DDYDNLQL’ showed homology with Rho-specific inhibitor of tran-scription termination (E = 0.42, Identity = 87%) and NBS-LRR typedisease resistant protein from peanut (Arachis hypogaea) (E = 2,Identity = 77%).
Discussion
The knowledge of biochemical and immunologic properties ofdisease-eliciting proteins is essential for the appropriate diagnosisand therapy of type I allergic disorders. Diagnosis of allergy with

902 D. Kumari et al. / Immunobiology 217 (2012) 895– 904
Fig. 6. (a) Glycoprotein detection by Periodic Acid Schiff’s (PAS) staining. Lane 1, blackgram extract; lane 2, purified protein. (b) Immunoblot analysis of purified protein afterperiodate treatment. The 28 kDa protein was electro-transfered onto NCM strip. It was incubated in dark with 20 mM sodium metaperiodate, overnight at 4 ◦C. The periodatew orohy ◦
d unobp
pett(aa1csbpas
aihwh(ampclbc2gso
tassh
as inactivated for 5–10 min by adding ethylene glycol. To this, 1 mg/ml of sodium brop of acetic acid. The strips were washed, blocked with 3% defatted milk and immrotein after periodate treatment.
urified allergens offers high sensitivity and specificity (Gamboat al. 2009). Also purified well characterized allergens can facili-ate in deciphering the mechanism underlying immunotherapy ando develop patient-tailored therapeutic and prophylactic strategiesValenta and Niederberger 2007). Legumes are widely consumed as
rich source of dietary proteins but they are implicated in IgE medi-ted food allergy in predisposed individuals (Ibanez Sandin et al.999; Bock et al. 2001; Rougé et al. 2010). Blackgram, a commonlyonsumed legume has been recognized as one of the important sen-itizers among allergic patients in India. IgE binding components oflackgram were identified by immunoblotting with hypersensitiveatients’ sera (Kumari et al. 2006). In the present study, a 28 kDallergen was isolated from blackgram extract and characterized bytandard in vivo and in vitro methods.
Over the years, allergenic proteins have been isolated and char-cterized from food sources using different chromatography andmmuno-biochemical techniques. In peanut, 8 allergenic proteinsave been identified namely, Ara h 1–Ara h 8. Ara h 1 (63 kDa)as purified on a MonoQ column (Burks et al. 1991) whereas Ara
2 (19 kDa) and Ara h 3 were purified by High Q anion columnBurks et al. 1992; Koppelman et al. 2003). Gly m Bd 28K – a majorllergen from soy, was purified by immunoaffinity column usingAb as a ligand (Tsuji et al. 1997). Anion-exchange chromatogra-
hy on a Mono Q column and reverse-phase HPLC on a Vydac-C4olumn were used to purify Len c 1.01 – a major allergen fromentil. Another 12–16 kDa allergen (Len c 1.02) was purified fromoiled lentil by a combination of gel filtration, cation exchangehromatography and reverse-phase HPLC (Sanchez-Monge et al.000). In the present study, we have purified a protein of black-ram using a combination of Q-sepharose column and phenylepharose column. The protein appeared as a single band of 28 kDan 12% SDS-PAGE.
An allergen is defined by its ability to exhibit IgE binding ando cross-link IgE/Fc�RI on the surface of mast cells and basophils
nd subsequent mediator release (Kay 2001, 2002). In the presenttudy, allergenicity of the purified 28 kDa protein was assessed bykin prick test, specific IgE, ELISA inhibition, immunoblotting andistamine release assay. Of the 9 patients SPT positive to blackgram,dride was added and kept overnight at 40 C. The reaction was stopped by adding alotted with pooled patients’ sera. Lane 1, 28 kDa protein (untreated); lane 2, 28 kDa
7 showed marked positive reactions to purified protein. Purifiedprotein showed IgE binding with more than 80% of hypersensitivepatients’ sera to blackgram on immunoblot indicating it to be amajor allergen. A dose dependent inhibition of IgE binding to theblackgram extract was observed by purified protein in competitiveELISA. One hundred and forty two ng of the purified protein wasrequired to achieve 50% inhibition as compared to 14 ng of the self-extract. A major peanut allergen Ara h 1 caused 50% inhibition at5.5 ng as compared to 1.5 ng of the self-extract. Up to 61% inhibitionwas observed with purified 28 kDa protein while up to 88% inhi-bition could be achieved by preincubaion of sera with blackgramextract (at 10 �g/mL). A major lentil allergen Len c 1 inhibited upto63% IgE binding to crude lentil extract (Sanchez-Monge et al. 2000).The 28 kDa protein demonstrated significant histamine release in9 patients’ samples hypersensitive to blackgram/purified protein.A previous study had shown that mediator release from basophilsignificantly correlated with airway reactivity in asthmatic patientshighlighting the effector function of basophils in inflammatory dis-eases and the pathologic significance of mediator release (Gaddyand Busse 1986).
Resistance to proteolytic enzymes and heat is a property asso-ciated with many food allergens. These produce stable allergenfragments that resist conditions of the gastrointestinal tract, andhave the potential to sensitize the immune system. Astwood et al.(1996) showed that food allergens were either resistant to pepsinin SGF or formed stable fragments, whereas the nonallergens werecompletely digested within 15 s. Stability of the whole protein orfragments from the allergens tested ranged from 8 to 60 min inSGF. Digestion of Ara h 1 by pepsin showed IgE reactive peptidefragments of 16–29 kDa that were stable till 30 min to 3 h whereastreatment with trypsin and chymotrypsin resulted in IgE reactivefragments ranging from 16 to 59 kDa (Maleki et al. 2000). In thepresent study, treatment of 28 kDa protein, with pepsin resulted instable IgE reactive fragments. The 28 kDa protein however digested
within 15 min upon treatment with pancreatin. Pancreatin hasbeen a mixture of amylase, trypsin and lipase with trypsin beingthe only proteolytic component which is assumed to digest restof the constituent proteins (amylase and lipase). This explains the
obiolo
r2
iwmffTs(m
tAh2fgec2psTi(ts
th(m
apc
A
lDfi
R
A
B
B
B
B
B
B
B
BG
D. Kumari et al. / Immun
eduction in number of bands in the molecular weight range of9–27 kDa with time.
The 28 kDa protein was detected as a glycoprotein by PAS stain-ng. On periodate treatment, there was a decrease in IgE binding
ith blackgram hypersensitive patients’ pooled sera. Ara h 1, aajor allergen of peanut is a glycoprotein belonging to the vicilin
amily (Shin et al. 1998). Len c 1 from lentil also showed 3 iso-orms varying in their N-glycosylation (Lopez-Torrejon et al. 2003).he major food allergens identified as class 1 allergens are water-oluble 10–70 kDa glycoproteins such as caseins (milk), ovomucoidegg), nonspecific lipid transfer proteins like Mal d 3 (apple) and Zea
14 (corn) (Breiteneder and Mills 2005).IL-4 cytokine is essential for initiation and maintenance of
he allergic disease (Seder et al. 1991; Scott-Taylor et al. 2005).llergen-dependent production of TH2 cytokines (IL-4, IL-5, IL-13)ave already been reported in many previous studies (Kurup et al.002; Kikuchi et al. 2006; Schwab et al. 2003) which are responsibleor inflammation and other immunopathological events. Food aller-enicity is characterized by abnormal IgE production, peripheralosinophilia, mast cell activation, and induction of TH2 lympho-ytes expressing cytokines such as IL-4 and IL-10 (Wong et al.001; Wierenga et al. 1991). In the present study, the purifiedrotein induced significant lymphocyte proliferation and increasedecretion of IL-4 than controls. The data indicates the predominantH2 response on exposure to purified protein further suggest-ng its allergenicity. IFN-� levels were however observed low20–50 pg/ml) as compared to non-allergic controls on stimula-ion with both blackgram extract and or purified protein (data nothown).
The N-terminus-12 residues sequence of 28 kDa purified pro-ein obtained was GRREDDYDNLQL. N-terminal sequence showedomology with NBS-LRR type disease resistant protein from peanutArachis hypogaea) and Rho-specific inhibitor of transcription ter-
ination.In conclusion, a 28 kDa protein was purified and characterized
s a major allergen from blackgram. The purified protein seems aotent allergen and may be implicated in possible strategies foromponent resolved diagnosis and therapy.
cknowledgements
One of the author (Dolly Kumari) received Senior Research Fel-owship from Council of Scientific and Industrial Research, Newelhi. Thanks are due to Dept. of Biotechnology, New Delhi fornancial support to this study.
eferences
stwood, J.D., Leach, J.N., Fuchs, R.L., 1996. Stability of food allergens to digestion invitro. Nat. Biotechnol. 14, 1269–1273.
allmer-Weber, B.K., Lidholm, J., Vieths, S., 2007. Recombinant allergens in the diag-nosis of food allergy: which are the advantages? Allergy 47, 126–128.
ock, S.A., Munoz-Furlong, A., Sampsom, H.A., 2001. Fatalities due to anaphylacticreactions to foods. J. Allergy Clin. Immunol. 107 (1), 191–193.
reiteneder, H., Ebner, C., 2000. Molecular and biochemical classification of plant-derived food allergens. J. Allergy Clin. Immunol. 106, 27–36.
reiteneder, H., Mills, E.N.C., 2005. Molecular properties of food allergens. J. AllergyClin. Immunol. 115, 14–23.
urks, A.W., William, L.W., Helm, R.M., Connaughton, C., Cockrell, G., Oı̌Brien, T.,1991. Identification of a major peanut allergen Ara h 1, in patients with atopicdermatitis and positive peanut challenges. J. Allergy Clin. Immunol. 88, 172–179.
urks, A.W., Williams, L.W., Connaughton, C., Cockrell, G., O’Brien, T.J., Helm, R.M.,1992. Identification and characterization of a second major peanut allergen Arah II, with use of the sera of patients with atopic dermatitis and positive peanutchallenge. J. Allergy Clin. Immunol. 90, 962–969.
urks, W., Bannon, G.A., Sicherer, S., Sampson, H.A., 1999. Peanut-induced anaphy-lactic reactions. Int. Arch. Allergy Immunol. 119, 165–172.
urks, W., Sampson, H.A., Bannon, G., 1998. Peanut allergens. Allergy 53, 725–730.addy, J.N., Busse, W.W., 1986. Enhanced IgE-dependent basophil histamine release
and airway reactivity in asthma. Am. Rev. Respir. Dis. 134 (5), 969–974.
gy 217 (2012) 895– 904 903
Gamboa, P.M., Sanz, M.L., Lombardero, M., Barber, D., Sánchez-Monje, R., Goikoetxea,M.J., Antépara, I., Ferrer, M., Salcedo, G., 2009. Component-resolved in vitrodiagnosis in peach-allergic patients. J. Investig. Allergol. Clin. Immunol. 19 (1),13–20.
Hage-Hamsten, M., Johansson, E., Kronqvist, M., Loughry, A., Cookson, W.O.,Moffatt, M.F., 2002. Associations of Fc epsilon R1-beta polymorphisms withimmunoglobin E antibody responses to common inhalant allergens in a ruralpopulation. Clin. Exp. Allergy 32, 838–842.
Heiss, S., Mahler, V., Steiner, R., Spitzauer, S., Schweiger, C., Kraft, D., Valenta, R., 1999.Component-resolved diagnosis (CRD) of type I allergy with recombinant grassand tree pollen allergens by skin testing. J. Invest. Dermatol. 113 (5), 830–837.
Ibanez Sandin, D., Martinez San Ireneo, M., Maranon Lizana, F., Fernandez-Caldas, E.,Alonso Lebrero, E., Laso Borrego, T., 1999. Specific IgE determinations to crudeand boiled lentil (Lens culinaris) extracts in lentil-sensitive children and controls.Allergy 54, 1209–1214.
Kalogeromitros, D., Armenaka, M., Galatas, I., Capellou, O., Katsarou, A., 1996. Ana-phylaxis induced by lentils. Ann. Allergy Asthma Immunol. 77, 480–482.
Kay, A.B., 2001. Allergy and allergic diseases. First of two parts. N. Engl. J. Med. 344(1), 30–37.
Kay, A.B., 2002. The cells and mediators of allergic inflammation. Clin. Exp. All. Rev.2, 8–12.
Kikuchi, Y., Takai, T., Kuhara, T., Ota, M., Kato, T., Hatanaka, H., Ichikawa, S., Tokura,T., Akiba, H., Mitsuishi, K., Ikeda, S., Okumura, K., Ogawa, H., 2006. Crucial com-mitment of proteolytic activity of a purified recombinant major house dust miteallergen Der p1 to sensitization toward IgE and IgG responses. J. Immunol. 177,1609–1617.
Kleine Budde, I., de Heer, P.G., van der Zee, J.S., Aalberse, R.C., 2001. The strippedbasophil histamine release bioassay as a tool for the detection of allergen-specific IgE in serum. Int. Arch. Allergy Immunol. 126 (4), 277–285.
Koppelman, S.J., Knol, E.F., Vlooswijk, R.A.A., Wensing, M., Knulst, A.C., Hefle, S.L.,Gruppen, H., Piersma, S., 2003. Peanut allergen Ara h 3: isolation from peanutsand biochemical characterization. Allergy 58, 1144–1151.
Kumari, D., Kumar, R., Sridhara, S., Arora, N., Gaur, S.N., Singh, B.P., 2006. Sensitizationto blackgram in patients with bronchial asthma and rhinitis: clinical evaluationand characterization of allergens. Allergy 61, 104–110.
Kurup, V.P., Xia, J.Q., Shen, H.D., Rickaby, D.A., Henderson Jr., J.D., Fink, J.N., Chou,H., Kelly, K.J., Dawson, C.A., 2002. Alkaline serine proteinase from Aspergillusfumigatus has synergistic effects on Asp-f-2-induced immune response in mice.Int. Arch. Allergy Immunol. 129, 129–137.
Laemmeli, U.K., 1970. Cleavage of structural proteins during the assembly of thehead of the bacteriophage T 4. Nature 227, 680–685.
Lin, J., Shewry, P.R., Archer, D.B., Beyer, K., Niggemann, B., Haas, H., Wilson, P., Alco-cer, M.J., 2006. The potential allergenicity of two 2S albumins from soybean(Glycine max): a protein microarray approach. Int. Arch. Allergy Immunol. 141(2), 91–102.
Lopez-Torrejon, G., Salcedo, G., Martin-Esteban, M., Diaz-Perales, A., Pascual, C.Y.,Sanchez-Monge, R., 2003. Len c 1, a major allergen and vicilin from lentil seeds:protein isolation and cDNA cloning. J. Allergy Clin. Immunol. 112, 1208–1215.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith the Folin–Phenol reagent. J. Biol. Chem. 193, 265–275.
Maleki, S.J., Casillas, A.M., Kaza, U., Wilson, B.A., Nesbit, J.B., Reimoneqnue, C.,Cheng, H., Bahna, S.L., 2010. Differences among heat-treated, raw, and commer-cial peanut extracts by skin testing and immunoblotting. Ann. Allergy AsthmaImmunol. 105 (6), 451–457.
Maleki, S.J., Kopper, R.A., Shin, D.S., Park, C.W., Compadre, C.M., Sampson, H.A., Burks,A.W., Bannon, G.A., 2000. Structure of the major peanut allergen Ara h 1 mayprotect IgE-binding epitopes from degradation. J. Immunol. 164, 5844–5849.
Martin, J.A., Compaired, J.A., de la Hoz, B., Quirce, S., Alonso, M.D., Igea, J.M., Losada,E., 1992. Bronchial asthma induced by chickpea and lentil. Allergy 47, 185–187.
Mittag, D., Akkerdaas, J., Ballmer-Weber, B.K., Vogel, L., Wensing, M., Becker, W.M.,Koppelman, S.J., Knulst, A.C., Helbling, A., Hefle, S.L., Van Ree, R., Vieths, S., 2004a.Ara h 8, a Bet v 1-homologous allergen from peanut, is a major allergen inpatients with combined birch pollen and peanut allergy. J. Allergy Clin. Immunol.114, 1410–1417.
Mittag, D., Vieths, S., Vogel, L., Becker, W.M., Rihs, H.P., Helbling, A., Wüthrich, B.,Ballmer-Weber, B.K., 2004b. Soybean allergy in patients allergic to birch pollen:clinical investigation and molecular characterization of allergens. J. Allergy Clin.Immunol. 113, 148–154.
Niphadkar, P.V., Patil, S.P., Bapat, M.M., 1997. Chickpea-induced anaphylaxis. Allergy52, 115–116.
Patil, S.P., Niphadkar, P.V., Bapat, M.M., 2001. Chickpea: a major food allergen inthe Indian subcontinent and its clinical and immunochemical correlation. Ann.Allergy Asthma Immunol. 87 (2), 140–145.
Rougé, P., Culerrier, R., Thibau, F., Didier, A., Barre, A., 2010. A case of severe ana-phylaxis to blackgram: phaseolin (vicilin) and PHA (lectin) identified as putativeallergens. Allergy [Epub ahead of print].
Sanchez-Monge, R., Pascual, C.Y., Diaz-Perales, A., Fernandez-Crespo, J., Martin-Esteban, M., Salcedo, G., 2000. Isolation and characterization of relevantallergens from boiled lentils. J. Allergy Clin. Immunol. 106, 955–961.
Schwab, C.J., Cooley, J.D., Brasel, T., Jumper, C.A., Graham, S.C., Straus, D.C., 2003.Characterization of exposure to low levels of viable Penicillium chrysogenum
conidia and allergic sensitization induced by a protease allergen extract fromviable P. chrysogenum conidia in mice. Int. Arch. Allergy Immunol. 130, 200–208.Scott-Taylor, T.H., Hourihane, J.B., Harper, J., Strobel, S., 2005. Patterns of foodallergen-specific cytokine production by T lymphocytes of children with multi-ple allergies. Clin. Exp. Allergy 35, 1473–1480.

9 obiolo
S
S
S
S
S
04 D. Kumari et al. / Immun
eder, R.A., Paul, W.E., Ben-Sasson, S.Z., LeGros, G.S., Kagey-Sobotka, A., Finkelman,F.D., Pierce, J.H., Plaut, M., 1991. Production of interleukin-4 and other cytokinesfollowing stimulation of mast cell lines and in vivo mast cells/basophils. Int.Arch. Allergy Appl. Immunol. 94, 137–140.
harma, P., Singh, A.K., Singh, B.P., Gaur, S.N., Arora, N., 2011. Allergenicity assess-ment of osmotin, a pathogenesis-related protein, used for transgenic crops. J.Agric. Food Chem. 59, 9990–9995.
hin, D.S., Compadre, C.M., Maleki, S.J., Kopper, R.A., Sampson, H., Huang, S.K., Burks,A.W., Bannon, G.A., 1998. Biochemical and structural analysis of the IgE bindingsites on Ara h 1, an abundant and highly allergenic peanut protein. J. Biol. Chem.273, 13753–13759.
icherer, S.H., Morrow, E.H., Sampson, H.A., 2000. Dose–response in double-blind,
placebo-controlled oral food challenges in children with atopic dermatitis. J.Allergy Clin. Immunol. 105, 582–586.ingh, A.K., Mehta, A.K., Sridhara, S., Gaur, S.N., Singh, B.P., Sarma, P.U., Arora, N.,2006. Allergenicity assessment of transgenic mustard (Brassica juncea) express-ing bacterial codA gene. Allergy 61, 91–97.
gy 217 (2012) 895– 904
Towbin, H.K., Staehelin, T.H., Gordon, J., 1979. Electrophoretic transfer of proteinsfrom polyacrylamide gels to nitrocellulose sheets. Proc. Natl. Acad. Sci. U.S.A. 76,4350.
Tsuji, H., Bando, N., Hiemori, M., Yamanishi, R., Kimoto, M., Nishikawa, K., Ogawa, T.,1997. Purification of characterization of soybean allergen Gly m Bd 28K. Biosci.Biotechnol. Biochem. 61 (6), 942–947.
Valenta, R., Niederberger, V., 2007. Recombinant allergens for immunotherapy. J.Allergy Clin. Immunol. 119, 826–830.
Wassenberg, J., Hofer, M., 2007. Lupine-induced anaphylaxis in a child withoutknown food allergy. Ann. Allergy Asthma Immunol. 98, 589–590.
Wierenga, E.A., Snoek, M., Jansen, H.M., Bos, J., Van Lier, R.A., Kapsenberg, M.L.,1991. Human atopen-specific types 1 and 2 T helper cell clones. J. Immunol.
147, 2942–2949.Wong, C.K., Ho, C.Y., Ko, F.W., Chan, C.H., Ho, A.S., Hui, D.S., Lam, C.W., 2001. Proin-flammatory cytokines (IL-17, IL-6, IL-18 and IL-12) and Th cytokines (IFN-�,IL-4 IL-10 and IL-13) in patients with allergic asthma. Clin. Exp. Immunol. 125,177–183.




















