
164-Burger and Co. Stock, Part III: U.S. Covers, Confederate ...
Upload
independentCategory
view
1download
0
ARTICLE IN PRESS
0958-6946/$ - se
doi:10.1016/j.id
�Correspond3250.
E-mail addr
(M.J. Coventry
International Dairy Journal 15 (2005) 133–143
www.elsevier.com/locate/idairyj
Isolation and characterisation of antibacterial peptides derived fromthe f(164–207) region of bovine aS2-casein
K.B. McCanna, B.J. Shiellb, W.P. Michalskib, A. Leec, J. Wanc,H. Roginskia, M.J. Coventryc,�
aInstitute of Land and Food Resources, Gilbert Chandler College, The University of Melbourne, Sneydes Road, Werribee, Vic. 3030, AustraliabAustralian Animal Health Laboratory, CSIRO Livestock Industries, Private Bag 24, Geelong, Vic. 3220, Australia
cFood Science Australia, Private Bag 16, Sneydes Road, Werribee, Vic. 3030, Australia
Received 23 May 2003; accepted 24 June 2004
Abstract
A chymosin digest of sodium caseinate, which showed antibacterial activity against Listeria innocua, was fractionated using
reverse phase high performance liquid chromatography and the purified antibacterial peptides were characterised by mass
spectrometry, N-terminal amino acid sequencing and comparison to peptide masses of theoretical enzymic digests of milk proteins.
Five antibacterial peptides, Cr1, Cr3, Cr4, Cr5 and Cr7 corresponding to amino acid residues 181–207, 180–207, 175–207, 164–207
and 172–207 of bovine aS2-casein, respectively, were isolated. The minimal inhibitory concentration of peptides Cr1, Cr4 and Cr5
was determined against a range of Gram-positive and Gram-negative bacteria and showed similar activities to those of the
bacteriocin peptide, nisin and the antibacterial peptide, lactoferricin B against certain Gram-positive bacteria. A partially purified
chymosin digest of sodium caseinate (CrMIX) was prepared and observed to be heat stable for up to 15min on exposure to 121 1C.
Although CrMIX showed bactericidal activity against Salmonella Typhimurium in 0.1% (w/v) peptone medium, no antibacterial
activity was observed when tested in skim milk at the same concentration.
r 2004 Elsevier Ltd. All rights reserved.
Keywords: Antibacterial; Peptide; aS2-Casein
1. Introduction
There has been a continuing interest in naturallyoccurring bioactive molecules and in their potential useas aids in food and health-care product preservation andhuman and veterinary therapeutic treatment (Bellamy,Takase, Wakabayashi, Kawase, & Tomita, 1992a;Gould, 1996). Jones and Simms (1930) reported theantibacterial activity of milk in vitro as far back as 1930,and it has long been recognised that breast-feeding ofinfants reduces the rate of enteric and respiratoryinfections (Welsh, Skurrie, & May, 1978; Portelli,
e front matter r 2004 Elsevier Ltd. All rights reserved.
airyj.2004.06.008
ing author. Tel.: +61-3-9731-3447; fax: +61-3-9731-
ess: [email protected]
).
Gordon, & May, 1998). The antibacterial activity ofmilk is attributed mainly to the immunoglobulins, butthe non-immune proteins, lactoferrin, lactoperoxidaseand lysozyme also exhibit distinct antibacterial activities(Pakkanen & Aalto, 1997).
In addition to the naturally occurring antimicrobialproteins present in milk, there are also a variety ofantibacterial peptides encrypted within the sequence ofmilk proteins that are released upon suitable hydrolysis ofthe precursor protein. To date, antibacterial peptides havebeen derived from the bovine milk proteins, lactoferrin(Bellamy et al., 1992b), aS1-casein (Lahov & Regelson,1996), aS2-casein (Zucht, Forssmann, Raida, & Ader-mann, 1996; Recio & Visser, 1999), a-lactalbumin(Pellegrini, Thomas, Bramaz, Hunziker, & von Fellenberg,1999), b-lactoglobulin (Pellegrini, Dettling, Thomas, &Hunziker, 2001) and k-casein (Malkoski et al., 2001).

ARTICLE IN PRESSK.B. McCann et al. / International Dairy Journal 15 (2005) 133–143134
Casocidin-I, isolated by Zucht, Raida, Adermann,Magert, and Forssman (1995) after boiling milk for5min in the presence of 10% glacial acetic acid,corresponds to residues 150–188 of bovine aS2-casein.Casocidin-I is a highly cationic peptide (calculatedisoelectric point (pI)¼ 10:08), with 10 of the total 39-amino acid residues being basic (Zucht et al., 1996) andreported to display strong inhibition against Escherichia
coli and Staphylococcus carnosus (Zucht et al., 1995).Two antibacterial peptides derived from aS2-casein byRecio and Visser (1999) were also highly basic, withf(164–179) having a calculated pI of 10.17 andf(183–207) having a calculated pI of 10.12. Peptidef(164–179) corresponded exactly to a region within thesequence of casocidin-I previously reported by Zucht etal. (1995). However, f(183–207) had only six of the total25-amino acid residues overlapping with the aminoacid sequence of the casocidin-I peptide. Of the twopeptides, f(183–207) showed greater antibacterialactivity against a selection of Gram-positive andGram-negative bacteria.
The aim of the current study was to isolate andcharacterise antibacterial peptides from chymosin di-gests of bovine casein and further characterise andcompare the antibacterial activity of the overlappingsequences of aS2-casein with those reported by Zuchtet al. (1995) and Recio and Visser (1999).
2. Materials and methods
2.1. Materials and bacterial cultures
The bovine milk proteins, sodium caseinate and aS-casein and chymosin (rennin) (EC 3.4.23.4) wereobtained from the Sigma Chemical Company (St. Louis,MO). A purified lactoferricin B sample (greater than90% purity as determined by reverse phase highperformance liquid chromatography (RP-HPLC) andmass spectrometry (MS) analysis) was kindly donatedby Dr. Paul Grieve, Centre for Food Technology,Hamilton, Qld., Australia. A purified nisin sample(greater than 90% w/v purity as determined by RP-HPLC analysis) was kindly donated by Dr. EgonHansen (Chr. Hansen, Denmark). Bacteriological pep-tone, yeast extract and dextrose were supplied by OxoidPty. Ltd. (West Heidelberg, Vic., Australia). Cultures ofBacillus subtilis (FSAW 0320), Bacillus cereus (FSAW0303), E. coli (NCTC 8196), Listeria innocua (FSAW2305), Listeria monocytogenes (FSAW 2310), L. mono-
cytogenes (FSAW 2325), Salmonella enterica subspeciesenterica serovars Agona (FSAW 3416), Bovismorbifi-cans (FSAW 3417), Dublin (FSAW 3421), Enteritidis(FSAW 3420), Salford (FSAW 3414), and Typhimurium(FSAW 3412) were obtained from the culture collectionat Food Science Australia, Werribee, Australia. All
reagents were analytical grade and obtained fromreputable suppliers.
2.2. Digestion of milk proteins
Bovine milk proteins were prepared in deionisedwater at a concentration of 50mgmL�1 and chymosinadded at a concentration of 1.5mgmL�1, followed byadjustment of the pH to 3.0 using 1mol L�1 HCl. Alldigestions were performed at 37 1C for 4 h andterminated by heat inactivation at 80 1C for 15min.After cooling, the pH of the digest was adjusted to 7.0using 1mol L�1 NaOH, centrifuged at 10,000g for 5minto remove any insoluble material, and filter sterilised(0.22 mm, Minisart, Sartorius, Germany).
2.3. Antibacterial assay
All milk protein digests were screened for antibacter-ial activity against L. innocua using a modification of thecritical dilution method of Wan, Hickey, and Coventry(1995). Milk protein digests (10 mL) were applied to PEPagar (containing 10 g bacteriological peptone, 10 gbacteriological agar per litre, pH 6.8) plates and overlaidwith 7mL of PYG soft agar medium (containing 10 gbacteriological peptone, 0.5 g yeast extract, 10 gdextrose, 6 g bacteriological agar per litre, pH 6.8) at50 1C, seeded with L. innocua (approximately1� 106CFUmL�1). The seeded plates were incubatedat 30 1C for 18–20 h and examined for zones of growthinhibition on the resultant bacterial lawn.
2.4. Capillary zone electrophoresis
Capillary zone electrophoresis (CZE) analysis wascarried out with a Beckman (Palo Alto, CA) P/ACE5510 instrument equipped with automated GOLDsoftware. Sodium caseinate and aS-casein samples(20mgmL�1) were diluted (1:5) in sample buffer(5mmol L�1 tri-sodium citrate, 10mmol L�1 dithiothre-tol, 6mol L�1 urea, pH 8.0) containing an internalstandard (2.2mmol L�1 glycyl-L-tyrosine). UncoatedCElect FS25 capillaries (27 cm� 25 mm i.d.; Supelco,Bellefonte, PA) were used. The samples and standardwere loaded under pressure by applying 3.8 kPa for 30 s.Separation occurred in the running buffer (20mmol L�1
citrate buffer containing 6mol L�1 urea, 0.5mgmL�1
methylhydroxyethyl cellulose, pH 3.0) and separationswere performed at 110V cm�1 with a typical current of90 mA. Ultraviolet absorbance was monitored at 214 nm.
2.5. Purification of the chymosin digest of sodium
caseinate
Initial fractionation of the chymosin digest of sodiumcaseinate was achieved using preparative reverse phase

ARTICLE IN PRESSK.B. McCann et al. / International Dairy Journal 15 (2005) 133–143 135
chromatography (RP-HPLC). A preparative C-18 col-umn (25 cm� 1.0 cm i.d.; Vydac, Hesperia, CA) wasused with a Hewlett-Packard 1050 series HPLC(Waldbronn, Germany) equipped with an HP auto-mated Chemstation and diode array detector; all eluateswere monitored at 210, 254 and 280 nm. The solventsconsisted of: (A) 0.1% (v/v) trifluoroacetic acid (TFA)(Merck Pty. Ltd., Kilsyth, Vic., Australia) in deionisedwater and (B) 0.1% (v/v) TFA in acetonitrile. Afterinjection of the samples, the HPLC system (flow rate of2mLmin�1) was equilibrated with 95% solvent A for5min, followed by a linear gradient of 5–45% (v/v)solvent B over 80min to elute peptides, and a 5min re-equilibration with 95% (v/v) solvent A.
Further fractionation of the chymosin digest ofsodium caseinate and re-chromatography of purifiedpeptides was performed using an analytical C-18 column(25 cm� 0.46 cm i.d., Vydac, Hesperia, CA) compatiblewith MS detection and connected to the HPLC. Thecolumn temperature was maintained at 45 1C using aBio-Rad HPLC column heater (Bio-Rad Laboratories,Hercules, CA). The solvents consisted of: (A) 0.02%(v/v) TFA in deionised water and (B) 0.02% (v/v) TFAin acetonitrile. After injection of samples onto thecolumn, the HPLC system was operated at a flow rate of1mLmin�1 under the linear gradient and equilibrationconditions described above.
The fractions from RP-HPLC fractionation of thechymosin digest of sodium caseinate were collected atprogrammed time intervals using a fraction collector(Foxy Jr., Isco Inc., Lincoln, USA). Fractions wereflushed with nitrogen gas for 30min to evaporatethe acetonitrile, transferred to sterile microcentri-fuge tubes, frozen (�20 1C) and lyophilised (FD-5freeze drier; Dynavac, Melbourne, Australia). Thelyophilised fractions from RP-HPLC were dissolved inappropriate volumes of sterile deionised water tofacilitate sample concentration and were assessed forantibacterial activity against L. innocua as describedabove.
2.6. Peptide identification by mass spectrometry
Purified antibacterial peptides were analysed using aFinnigan LCQ Classic quadrupole ion-trap massspectrometer (San Jose, CA, USA). Peptide samplescollected from RP-HPLC fractionation were preparedby mixing at a 1:1 weight ratio with a solutioncontaining 50% (v/v) methanol and 1% (v/v) aceticacid (both from Ajax Chemicals, Auburn, NSW,Australia) and were directly infused at a flow rate of5 mLmin�1 to the electrospray ion (ESI) source of themass spectrometer. Data were acquired over a mass/charge (m=z) range of 400–2000 and the zoom scanfunction permitted high mass accuracy for determina-tion of the mass to charge ratio. The Qual Browser
software (Finnigan, San Jose, CA, USA) was used tocalculate the mass of the peptides. ESI source settingswere as follows: ion spray voltage 5.5 kV, capillarytemperature 175 1C, capillary voltage 30V and tube lensoffset 10V.
The determined molecular masses of the antibacterialpeptides were compared with those of peptides gener-ated by theoretical enzymatic digests of the original milkproteins. The sequences were obtained from theNational Centre for Biotechnology Information (NCBI)non-redundant database, which included those of aS1-casein (Accession No. P02662, Stewart, Willis, &Mackinlay, 1984), aS2-casein (Accession No. P02663,Stewart et al., 1987), b-casein (Accession No. P02666,Stewart et al., 1987) and k-casein (Accession No.P02668, Stewart et al., 1984). A set of theoreticalpeptide masses was generated using the ProteinInfo-enzymatic cleavage tool available at http://prowl1.rock-efeller.edu.prowl-cgi/enzyme.exe (ProteoMetrics LLC,New York, NY). Theoretical chymosin digests of thebovine caseins were generated using the expectedchymosin cleavage sites provided by Fox and McSwee-ney (1996). Where an exact match between the experi-mental and theoretical molecular masses could not befound, the peptide samples were analysed by N-terminalamino acid sequencing.
2.7. Calculation of theoretical isoelectric point of peptides
The theoretical pI of peptides was calculated using theCompute pI/MW tool available at http://expasy.ch/tools/pi_tool.html (Bjellqvist et al., 1993).
2.8. Edman degradation N-terminal amino acid
sequencing
RP-HPLC purified antibacterial peptides weresubjected to N-terminal amino acid sequence analysiswith pulse-liquid delivery of critical reagents in anautomated sequenator (Procise, Applied Biosystems,Foster City, CA) in conjunction with a phenyl-thiohydantoin (PTH)–amino acid separation system(Procise, PTH Analyser, Applied Biosystems, FosterCity, CA).
2.9. Amino acid analysis of peptides
The antibacterial peptides were quantified usingamino acid analysis according to the method of Mooreand Steine (1963). Peptides were dissolved in 6M HClcontaining 1% (v/v) phenol and hydrolysed by heatingat 108 1C for 24 h. Hydrolysates were subsequentlyanalysed by cation exchange HPLC with post-columnaddition of ninhydrin.

ARTICLE IN PRESSK.B. McCann et al. / International Dairy Journal 15 (2005) 133–143136
2.10. Minimum inhibitory concentration determination of
antibacterial peptides
Minimum inhibitory concentration (MIC) assayswere performed in sterile 96-well microtitre plates(Nunc, Roskilde, Denmark) by the method of Hoek,Milne, Grieve, Dionysius, and Smith (1997). PYG broth(containing 10 g bacteriological peptone, 0.5 g yeastextract, 10 g dextrose per litre, pH 6.8) was used forgrowth of L. innocua, B. subtilis, B. cereus and L.
monocytogenes, and PEP broth (containing 10 g bacter-iological peptone per litre, pH 6.8) for the growth of E.
coli, S. Agona, S. Bovismorbificans, S. Dublin, S.
Salford, S. Enteritidis and S. Typhimurium. The testmixture comprised growth medium (190 mL), peptidetest solution (50 mL of two-fold dilutions in steriledistilled water) and bacterial inoculum (10 mL ofovernight broth culture diluted to approximately1� 104CFUmL�1 in sterile 0.1% (w/v) peptone).Controls containing sterile water in place of thepeptide solution and the bacterial inoculum werealso included and four replicates were tested for eachsample.
The MIC of the antibacterial peptide was defined asthe lowest concentration of peptide (expressed asmgmL�1 and mmolL�1) that resulted in no increase ofabsorbance at 650 nm (Thermomax microtitre platereader (Molecular Devices, USA)) after incubation at30 1C for 18 h without shaking.
2.11. Large-scale production and crude purification of
chymosin digest of bovine sodium caseinate
Sodium caseinate (15 g) was digested with chymosinat pH 3.0 as described in Section 2.2 previously. Thechymosin digest of sodium caseinate was initiallypurified by centrifugation at low pH, followed by cationexchange chromatography using an ion-exchange syr-inge fit filtration unit (Sartobind-S, Sartorius, Ger-many). Cation exchange chromatography wasperformed at a flow rate of 5mLmin�1 and the unitwas equilibrated with 10mL of 10mmol L�1 sodiumphosphate buffer (pH 7.0). The chymosin digest ofsodium caseinate (50mgmL�1) was diluted (1:5 v/v) in10mmol L�1 sodium phosphate buffer (pH 7.0). Asample of the diluted digest (5mL) was loaded ontothe ion-exchange unit and washed with 10mL of10mmol L�1 sodium phosphate buffer (pH 7.0). Thepeptides bound to the ion-exchange membrane wereeluted by washing with 10mL of 10mmol L�1 sodiumphosphate buffer (pH 7.0) containing 2mol L�1 NaCl.The absorbance of the effluent was monitored at 280 nmusing an Econo UV monitor (Bio-Rad Laboratories,Hercules, CA). The fraction eluted from the cationexchange resin using NaCl showed antibacterial activityagainst L. innocua and was desalted by dialysis against
deionised water at 4 1C for 24 h, using a Spectra/Por 71000Da molecular mass cut-off membrane (SpectrumLaboratories Inc., Rancho Dominguez, USA). Thisfraction was referred to as CrMIX.
2.12. Effect of CrMIX on the viable count of S.
Typhimurium in peptone solution and skim milk
The determination of the effect of the antibacterialpeptides on bacterial cells was performed by the methoddescribed by Wan, Wilcock, and Coventry (1998) in0.1% (w/v) peptone and ultra-heat treated (UHT) skimmilk. An S. Typhimurium culture was grown in TSYEmedium (containing 30 g tryptone soya broth, 6 g yeastextract per litre, pH 7) at 30 1C for 24 h and seriallydiluted in sterile 0.1% (w/v) peptone. Cell suspensions(1mL) of approximately 105CFUmL�1 of S. Typhi-murium were centrifuged (10,000g, 5min) and re-suspended in either an equal volume of sterile 0.1%(w/v) peptone or UHT skim milk. Volumes of CrMIX
(1mL) were prepared in 0.1% (w/v) peptone or in UHTskim milk at a concentration of 10mgmL�1 and addedto S. Typhimurium cells (1mL) in either peptone orskim milk (to achieve a final test concentration of5mgmL�1 CrMIX), respectively, and incubated at 30 1Cfor 2, 6 or 24 h. As controls, S. Typhimurium in 0.1%(w/v) peptone or UHT skim milk were incubated at30 1C diluted 1:1 with either peptone or UHT skim milkwithout addition of CrMIX for the same periods. Afterincubation the samples (1mL) were washed, centrifuged(10,000g, 5min) and the pellets were re-suspended in anequal volume of sterile 0.1% (w/v) peptone; viable cellcounts were performed on the re-suspended pellet.Viable counts were determined in three replicate samplesplated (1mL pour plates) onto TSYE agar (containing10 g bacteriological agar per litre) and incubated at30 1C for 48 h.
2.13. Heat stability of CrMIX
CrMIX (50mgmL�1) was transferred to sterile boil-resistant microcentrifuge tubes, heated at either 100 1Cfor 15min or 121 1C for 15min, and cooled on ice. Theantibacterial activity of the heated and non-heatedCrMIX samples were assessed by MIC against B. subtilis,as described in Section 2.10.
3. Results
3.1. Purification and identification of antibacterial
peptides
Preliminary inspection showed that the chymosindigests of both sodium caseinate and aS-casein had anti-L. innocua activity. Almost identical CZE profiles with
ARTICLE IN PRESSK.B. McCann et al. / International Dairy Journal 15 (2005) 133–143 137
respect to the types and relative proportion of caseins(data not shown) were observed for the undigestedpreparations of casein and aS-casein. Further studieswere performed on digests of sodium caseinate due to itsavailability in larger quantities.
The chymosin digest of sodium caseinate contained ahigh amount of insoluble material after 4 h digestion at37 1C. Initial purification of the chymosin digest ofsodium caseinate by centrifugation at pH 3.0 removed theinsoluble protein and peptide material without affectingthe antibacterial activity (data not shown), indicating thatantibacterial peptides were soluble at pH 3.0.
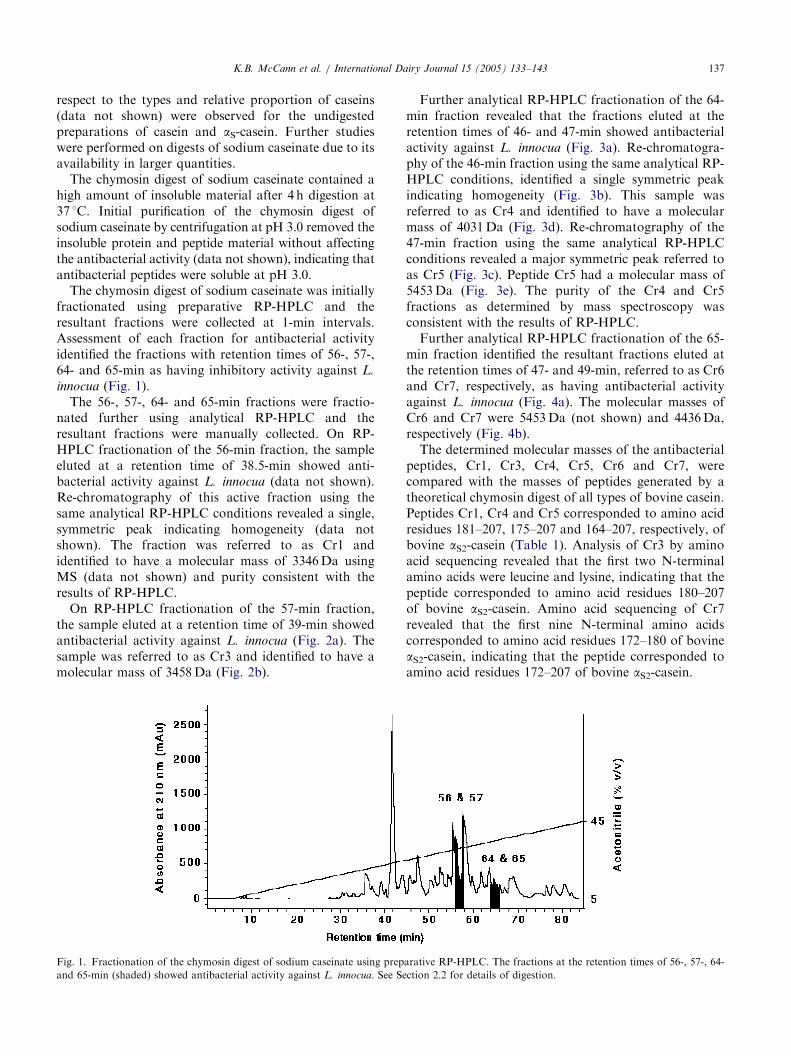
The chymosin digest of sodium caseinate was initiallyfractionated using preparative RP-HPLC and theresultant fractions were collected at 1-min intervals.Assessment of each fraction for antibacterial activityidentified the fractions with retention times of 56-, 57-,64- and 65-min as having inhibitory activity against L.
innocua (Fig. 1).The 56-, 57-, 64- and 65-min fractions were fractio-
nated further using analytical RP-HPLC and theresultant fractions were manually collected. On RP-HPLC fractionation of the 56-min fraction, the sampleeluted at a retention time of 38.5-min showed anti-bacterial activity against L. innocua (data not shown).Re-chromatography of this active fraction using thesame analytical RP-HPLC conditions revealed a single,symmetric peak indicating homogeneity (data notshown). The fraction was referred to as Cr1 andidentified to have a molecular mass of 3346Da usingMS (data not shown) and purity consistent with theresults of RP-HPLC.
On RP-HPLC fractionation of the 57-min fraction,the sample eluted at a retention time of 39-min showedantibacterial activity against L. innocua (Fig. 2a). Thesample was referred to as Cr3 and identified to have amolecular mass of 3458Da (Fig. 2b).
Fig. 1. Fractionation of the chymosin digest of sodium caseinate using prep
and 65-min (shaded) showed antibacterial activity against L. innocua. See Se
Further analytical RP-HPLC fractionation of the 64-min fraction revealed that the fractions eluted at theretention times of 46- and 47-min showed antibacterialactivity against L. innocua (Fig. 3a). Re-chromatogra-phy of the 46-min fraction using the same analytical RP-HPLC conditions, identified a single symmetric peakindicating homogeneity (Fig. 3b). This sample wasreferred to as Cr4 and identified to have a molecularmass of 4031Da (Fig. 3d). Re-chromatography of the47-min fraction using the same analytical RP-HPLCconditions revealed a major symmetric peak referred toas Cr5 (Fig. 3c). Peptide Cr5 had a molecular mass of5453Da (Fig. 3e). The purity of the Cr4 and Cr5fractions as determined by mass spectroscopy wasconsistent with the results of RP-HPLC.
Further analytical RP-HPLC fractionation of the 65-min fraction identified the resultant fractions eluted atthe retention times of 47- and 49-min, referred to as Cr6and Cr7, respectively, as having antibacterial activityagainst L. innocua (Fig. 4a). The molecular masses ofCr6 and Cr7 were 5453Da (not shown) and 4436Da,respectively (Fig. 4b).
The determined molecular masses of the antibacterialpeptides, Cr1, Cr3, Cr4, Cr5, Cr6 and Cr7, werecompared with the masses of peptides generated by atheoretical chymosin digest of all types of bovine casein.Peptides Cr1, Cr4 and Cr5 corresponded to amino acidresidues 181–207, 175–207 and 164–207, respectively, ofbovine aS2-casein (Table 1). Analysis of Cr3 by aminoacid sequencing revealed that the first two N-terminalamino acids were leucine and lysine, indicating that thepeptide corresponded to amino acid residues 180–207of bovine aS2-casein. Amino acid sequencing of Cr7revealed that the first nine N-terminal amino acidscorresponded to amino acid residues 172–180 of bovineaS2-casein, indicating that the peptide corresponded toamino acid residues 172–207 of bovine aS2-casein.
arative RP-HPLC. The fractions at the retention times of 56-, 57-, 64-
ction 2.2 for details of digestion.

ARTICLE IN PRESS
Fig. 2. (a) Fractionation of the 57-min retention time fraction, obtained by preparative RP-HPLC of sodium caseinate digest (see Fig. 1). Using
analytical RP-HPLC yielded a fraction (Cr3) with a retention time of 39-min (shaded area) which showed antibacterial activity against L. innocua; (b)
molecular mass of Cr3.
K.B. McCann et al. / International Dairy Journal 15 (2005) 133–143138
The amino acid sequences of the antibacterial peptides,Cr1, Cr3, Cr4, Cr5, and Cr7, are aligned and comparedwith the antibacterial peptide sequences reported byZucht et al. (1995) and Recio and Visser (1999) in Table1. The theoretical pI values of Cr1, Cr3, Cr4, Cr5 andCr7 were all high, ranging from 10.10 to 10.35 (Table 1).
3.2. Minimum inhibitory concentration of antibacterial
peptides
The antibacterial peptides, Cr1, Cr4 and Cr5, were themajor peptides derived from the chymosin digest of
sodium caseinate and the concentrations of the peptideswere determined by amino acid analysis to be 0.84, 0.86and 0.38mgmL�1, respectively. The MIC of Cr1, Cr4and Cr5 was determined against a range of bacterialcultures and compared with the bacteriocin peptide,nisin, and the antibacterial peptide, lactoferricin B. Inaddition, a chymosin digest of sodium caseinate thathad been partially purified using cation exchangechromatography (referred to as CrMIX) was alsoassessed for its MIC.
Similar to nisin, the MIC of Cr1, Cr4 and Cr5 againstthe Gram-positive bacteria, L. innocua, B. subtilis and

ARTICLE IN PRESS
Fig. 3. (a) Further fractionation of the 64-min fraction using analytical RP-HPLC—46-min (Cr4) and 47-min (Cr5) fractions showed antibacterial
effects against L. innocua; (b) re-chromatography of Cr4 using analytical RP-HPLC; (c) re-chromatography of Cr5 using analytical RP-HPLC; (d)
molecular mass of Cr4; and (e) molecular mass of Cr5.
K.B. McCann et al. / International Dairy Journal 15 (2005) 133–143 139
L. monocytogenes (ranging from 4.8 to 21 mgmL�1)was considerably lower than the MIC against theGram-negative bacteria, E. coli and S. Enteritidis(476 mgmL�1) (Table 2).
The MIC of the partially purified chymosin digestof sodium caseinate (CrMIX) was very high againstall bacteria tested and was most active againstL. innocua (625 mgmL�1), B. subtilis (312.5 mgmL�1),

ARTICLE IN PRESS
Fig. 4. (a) Fractionation of the 65-min retention time fraction, obtained by preparative RP-HPLC of sodium caseinate digest (see Fig. 1). Using
analytical RP-HPLC yielded two fractions (Cr6 and Cr7) with retention times of 47- and 49-min, respectively (shaded areas) that showed
antibacterial activity against L. innocua; (b) molecular mass of Cr7.
K.B. McCann et al. / International Dairy Journal 15 (2005) 133–143140
S. Typhiumurium (625mgmL�1), S. Salford (625mgmL�1)and the two strains of L. monocytogenes (625mgmL�1)(Table 2).
In contrast, lactoferricin B showed relatively lowMICs ranging from 6.3 to 28 mgmL�1, for Gram-positive bacteria and some Gram-negative bacteria.
3.3. Antibacterial activity of peptides against
S. Typhimurium in peptone solution and reconstituted
skim milk
The antibacterial activity of CrMIX was determinedagainst S. Typhiumurium (challenged at 105CFU
mL�1) at a concentration of 5mgmL�1 (eight-foldMIC) in 0.1% w/v peptone and skim milk. In 0.1% w/vpeptone, CrMIX (at a concentration of 5mgmL�1)reduced the viable counts of S. Typhiumurium by5-log cycles after 24 h exposure at 30 1C comparedwith the S. Typhiumurium cells in the 0.1% w/vpeptone control which reached 108CFUmL�1 after24 h (Fig. 5). When CrMIX was added to skim milk at aconcentration of 5mgmL�1, no antibacterial activitywas observed against S. Typhiumurium, with similarviable counts as S. Typhiumurium in the skimmilk control being observed after 24 h incubation at30 1C (Fig. 5).

ARTICLE IN PRESS
Table
1
Aminoacidsequence
alignmentofantibacterialpeptides
derived
from
bovinea S
2-casein
(inthecurrentstudy)andcomparisonwithcasocidin-I,f(164–179),f(183–207)andf(181–207)
Peptide
Aminoacidsequence
Reference
pIb
Theoretical
molecular
mass
(Da)
Determined
molecular
mass
(Da)
Casocidin-I
a150KTKLTEEEKNRLNFLKKISQRYQKFALPQYLKTVYQHQK
188a
Zuchtet
al.(1995)
10.08
f(164–179)
164LKKISQRYQKFALPQY
179
Recio
andVisser(1999)
10.17
f(183–207)
183VYQHQKAMKPWIQ
PKTKVIPYVRYL207
Recio
andVisser(1999)
10.12
f(181–207)
181KTVYQHQKAMKPWIQ
PKTKVIPYVRYL207
Recio
andVisser(1999)
10.22
Cr1
181KTVYQHQKAMKPWIQ
PKTKVIPYVRYL207
Thisstudy
10.22
3345
3346
Cr3
180LKTVYQHQKAMKPWIQ
PKTKVIPYVRYL207
Thisstudy
10.22
3458
3458
Cr4
175ALPQYLKTVYQHQKAMKPWIQ
PKTKVIPYVRYL207
Thisstudy
10.10
4031
4031
Cr7
172QKFALPQYLKTVYQHQKAMKPWIQ
PKTKVIPYVRYL207
Thisstudy
10.35
4434
4436
Cr5/C
r6164LKKISQRYQKFALPQYLKTVYQHQKAMKPWIQ
PKTKVIPYVRYL207
Thisstudy
10.17
5452
5453
aNumbersonleft
andrightofsequence
correspondto
aminoacidresiduenumber.
bpI,isoelectric
point.
0
2
4
6
8
10
0 5 10 15 20 25S. T
yphi
mur
ium
(lo
g C
FU
mL-1
)
Exposure time at 30°C (h)
Fig. 5. Antibacterial activity of CrMIX (5mg mL�1) in 0.1% w/v
peptone and reconstituted skim milk (standard deviation did not
exceed 6% for the mean of triplicate samples) against S. Typhimurium:
(J) skim milk control; (n) skim milk+CrMIX; (&) 0.1% w/v
peptone+CrMIX; and (K) 0.1% w/v peptone control.
K.B. McCann et al. / International Dairy Journal 15 (2005) 133–143 141
The antibacterial activity of CrMIX was assessed afterexposure to 100 1C for 15min and 121 1C for 15min.The MIC of CrMIX against B. subtilis after 15minexposure to 100 or 121 1C (312.5 mgmL�1) was identicalto the MIC of the unheated control (312.5 mgmL�1)indicating that the constituent antibacterial peptides ofCrMIX were heat stable.
4. Discussion
In the current study, five antibacterial peptides wereisolated from a chymosin digest of bovine sodiumcaseinate. The peptides referred to as Cr1, Cr3, Cr4, Cr5and Cr7 all originated from the C-terminal of bovineaS2-casein and all five were different from the casocidin-Isequence reported by Zucht et al. (1995). A peptidefraction Cr6 was also isolated but sequence analysisdetermined that it was identical with Cr7. The aminoacid sequence of Cr1 corresponded exactly with theantibacterial peptide, f(181–207) reported by Recio andVisser (1999). The amino acid sequences of peptidesCr3, Cr4, Cr5 and Cr7 have not been previouslyreported, but all four peptides share the identical 27-amino acid residue region as the f(181–207) sequencereported by Recio and Visser (1999). Compared withCr1, the peptides Cr3, Cr4, Cr5 and Cr7 containedbetween 1 and 17 additional amino acid residues at theirN-terminals.
Peptides Cr1, Cr4 and Cr5 had MICs against Gram-positive bacteria which were similar to that of nisin andlactoferricin B. Although peptides Cr1, Cr4 and Cr5share an identical 27-amino acid residue region, theyexhibited considerable differences in their activityagainst Gram-positive bacteria. Peptide Cr4, whichcontained an additional six amino acids compared withCr1, had MIC values against Gram-positive bacteriathat were 2.3 times lower than those of Cr1 on a molar

ARTICLE IN PRESS
Table 2
Minimum inhibitory concentration (MIC) of antibacterial peptides derived by chymosin hydrolysis of aS2-casein against selected bacteria incubated
at 30 1C for 18 h
Bacterial culture Minimum inhibitory concentration (MIC) of peptidea
Cr1 Cr4 Cr5 CrMIX Lactoferricin B Nisin
mgmL�1b mmolL�1c mgmL�1 mmolL�1 mgmL�1 mmolL�1 mgmL�1 mmolL�1 mgmL�1 mmolL�1 mgmL�1 mmolL�1
Bacillus cereus 484 425.1 — — — — — — 400 125 63 17.8
Bacillus subtilis 21 6.3 10.7 2.7 4.8 0.9 312.5 N/A 6.3 2.0 7.8 2.2
Listeria innocua 21 6.3 10.7 2.7 4.8 0.9 625 N/A 6.3 2.0 3.9 1.1
Listeria monocytogenes 2310 21 6.3 10.7 2.7 4.8 0.9 625 N/A 6.3 2.0 3.9 1.1
Listeria monocytogenes 2325 — — — — — — 625 N/A 6.3 2.0 3.9 1.1
Escherichia coli 4168 450.3 4171.2 442.4 476.2 4 14 5000 N/A 12.5 3.9 400 114
Salmonella Agona — — — — — 410,000 N/A — — — —
Salmonella Bovismorbificans — — — — — — 410,000 N/A — — — —
Salmonella Dublin — — — — — — 5000 N/A — — — —
Salmonella Enteritidis 484 425.1 — — — — 5000 N/A 25 7.8 200 57
Salmonella Salford — — — — — — 625 N/A — — — —
Salmonella Typhimurium 21 6.3 21.4 5.2 — — 625 N/A 6.3 2.0 50 14.2
Note: N/A indicates not applicable as sample contains multiple peptides;—indicates not tested.aMIC determined as mean of four replicates.bMIC expressed as mgmL�1 of each peptide.cMIC expressed as mmolL�1 of each peptide.
K.B. McCann et al. / International Dairy Journal 15 (2005) 133–143142
basis. Peptide Cr5, which contained an additional 17amino acids compared with Cr1, had MIC values thatwere generally seven times lower than those of Cr1 on amolar basis against the Gram-positive bacteria.
While it was identified that lactoferricin B andpeptides Cr1, Cr4 and Cr5 had similar MIC valuesagainst the Gram-positive bacteria, lactoferricin B was,however, more effective against the Gram-negativebacteria. Yamauchi, Tomita, Giehl, and Ellison III(1993) reported that lactoferricin B binds to thelipopolysaccharide molecules of the outer membraneof Gram-negative bacteria causing an increase in itspermeability, thereby allowing lactoferricin B to act onthe bacterial cytoplasmic membrane. Conversely, theaS2-casein derived peptides (Cr1, Cr4 and Cr5) may lackthe ability to permeabilise the outer membrane of Gram-negative bacteria, as effectively as lactoferricin B.
The partially purified chymosin digest of sodiumcaseinate (CrMIX), containing the antibacterial peptidesCr1, Cr4 and Cr5, also possessed high amounts of otherpeptides, resulting in the MIC value of the sample beingvery high. The CrMIX sample was shown to retain fullantibacterial activity against B. subtilis after heating at121 1C for 15min, suggesting that the constituentantibacterial peptides were heat resistant. The heatstabilities of peptides encrypted within milk proteinshave not been widely reported. The heat stability of theconstituent antibacterial peptides of CrMIX was greaterthan that reported for pediocin PO2 by Coventry,Muirhead, and Hickey (1995), who found that theantibacterial activity of pediocin PO2 (a hydrophobicpeptide bacteriocin) was not affected by heating at100 1C for 15min.
The CrMIX preparation showed bactericidal activityagainst S. Typhiumurium in 0.1% w/v peptone medium(5-log cycle reduction in cell count at 5mgmL�1).However, addition of CrMIX to skim milk at the sameconcentration did not restrict the growth or reduce theviable count of S. Typhimurium. This is consistent withthe studies of Jones, Smart, Bloomberg, Burgess, andMillar (1994), Appendini and Hotchkiss (2000) andChantaysakorn and Richter (2000), all of whomdemonstrated bactericidal activity of cationic peptidesin minimal media, but observed a substantial loss ofactivity when the peptides were added to foods includingmilk and carrot juice. Further studies by Chantaysakornand Richter (2000) identified the presence of metalcations as the major reason for the reduced activity ofcationic antibacterial peptides in certain foods. It hasbeen proposed that the metal cations stabilise negativelycharged target sites such as the lipopolysaccharides ofthe outer membrane of Gram-negative bacteria therebyreducing the affinity of the peptides for these sites(Ellison III & Giehl, 1991; Stevens, Klapes, Sheldon, &Klaenhammer 1992). The successful application ofcationic antibacterial peptides as antimicrobials inbiological systems where metal cations are likely tointerfere will require the development of deliverystrategies and approaches to minimise the adverseeffects of metal cations on their biological activity.
Acknowledgements
K.B. McCann is the recipient of the Australian DairyResearch and Development Corporation Postgraduate

ARTICLE IN PRESSK.B. McCann et al. / International Dairy Journal 15 (2005) 133–143 143
Research Scholarship (UM075). The assistance of Dr.Xinqing Xu (Food Science Australia, Werribee, Aus-tralia; CZE), Gary Beddome (CSIRO Livestock Indus-tries, AAHL, Geelong, Australia; N-terminal aminoacid sequencing) and Nick Bartone (CSIRO HealthSciences and Nutrition, Parkville, Australia; amino acidanalysis) is gratefully acknowledged.
References
Appendini, P., & Hotchkiss, J. H. (2000). Antimicrobial activity of a
14-residue synthetic peptide against foodborne microorganisms.
Journal of Food Protection, 63, 889–893.
Bellamy, W., Takase, M., Wakabayashi, H., Kawase, K., & Tomita,
M. (1992a). Antibacterial spectrum of lactoferricin B, a potent
bactericidal peptide derived from the N-terminal region of bovine
lactoferrin. Journal of Applied Bacteriology, 73, 472–479.
Bellamy, W., Takase, M., Yamauchi, K., Wakabayashi, H., Kawase,
K., & Tomita, M. (1992b). Identification of the bactericidal domain
of lactoferrin. Biochimica et Biophysica Acta, 1121, 130–136.
Bjellqvist, B., Hughes, G. J., Pasquali, Ch., Paquet, N., Ravier, F.,
Sanchez, J. Ch., Frutiger, S., & Hochstrasser, D. F. (1993). The
focusing positions of polypeptides in immobilized pH gradients can
be predicted from their amino acid sequences. Electrophoresis, 14,
1023–1031.
Chantaysakorn, P., & Richter, R. L. (2000). Antimicrobial properties
of pepsin-digested lactoferrin added to carrot juice and filtrates of
carrot juice. Journal of Food Protection, 63, 376–380.
Coventry, M. J., Muirhead, K., & Hickey, M. W. (1995). Partial
characterisation of pediocin PO2 and comparison with nisin for
biopreservation of meat products. International Journal of Food
Microbiology, 26, 133–145.
Ellison, R. T., III, & Giehl, T. J. (1991). Killing of gram-negative
bacteria by lactoferrin and lysozyme. Journal of Clinical Investiga-
tion, 88, 1080–1091.
Fox, P. F., & McSweeney, P. L. H. (1996). Proteolysis in cheese during
ripening. Food Reviews International, 12, 457–509.
Gould, G. W., 1996. Industry perspectives on the use of natural
antimicrobials and inhibitors for food applications. Journal of Food
Protection (Suppl.), 82–86.
Hoek, K. S., Milne, J. M., Grieve, P. A., Dionysius, D. A., & Smith, R.
(1997). Antibacterial activity of bovine lactoferrin-derived peptides.
Antimicrobial Agents and Chemotherapy, 41, 54–59.
Jones, F. S., & Simms, H. S. (1930). The bacterial growth inhibitor
(lactenin) of milk. Journal of Experimental Medicine, 51, 327–339.
Jones, E. M., Smart, A., Bloomberg, G., Burgess, L., & Millar, M. R.
(1994). Lactoferricin, a new antimicrobial peptide. Journal of
Applied Bacteriology, 77, 208–214.
Lahov, E., & Regelson, W. (1996). Antibacterial and immunostimulat-
ing casein-derived substances from milk: casecidin, isracidin
peptides. Food and Chemical Toxicology, 34, 131–145.
Malkoski, M., Dashper, S. G., O’Brien-Simpson, N. M., Talbo, G. H.,
Marcis, M., Cross, K. J., & Reynolds, E. C. (2001). Kappacin, a
novel antibacterial peptide from bovine milk. Antimicrobial Agents
and Chemotherapy, 45, 2309–2315.
Moore, S., & Steine, W. H. (1963). Methods in enzymology, Vol. VI.
New York: Academic Press, p. 820.
Pakkanen, R., & Aalto, J. (1997). Growth factors and antimicrobial
factors of bovine colostrum. International Dairy Journal, 7,
285–297.
Pellegrini, A., Dettling, C., Thomas, U., & Hunziker, P. (2001).
Isolation and characterization of four bactericidal domains
in the b-lactoglobulin. Biochimica et Biophysica Acta, 1526,
131–140.
Pellegrini, A., Thomas, U., Bramaz, N., Hunziker, P., & von
Fellenberg, R. (1999). Isolation and identification of three
bactericidal domains in the bovine a-lactalbumin molecule.
Biochimica et Biophysica Acta, 1426, 439–448.
Portelli, J., Gordon, A., & May, J. T. (1998). Effect of compounds with
antibacterial activities in human milk on respiratory syncytial virus
and cytomegalovirus in vitro. Journal of Medical Microbiology, 47,
1015–1018.
Recio, I., & Visser, S. (1999). Identification of two distinct
antibacterial domains within the sequence of bovine aS2-casein.Biochimica et Biophysica Acta, 1428, 314–326.
Stevens, K. A., Klapes, N. A., Sheldon, B. W., & Klaenhammer, T. R.
(1992). Antimicrobial action of nisin against Salmonella typhimur-
ium lipopolysaccharide mutants. Applied and Environmental Micro-
biology, 58, 1786–1788.
Stewart, A. F., Bonsing, J., Beattie, C. W., Shah, F., Willis, I. M., &
Mackinlay, A. G. (1987). Complete nucleotide sequences of bovine
alpha S2-and beta-casein cDNA’s: comparisons with related
sequences in other species. Molecular Biology and Evolution, 4,
231–241.
Stewart, A. F., Willis, I. M., & Mackinlay, A. G. (1984). Nucleotide
sequences of bovine alpha S1-and kappa-casein cDNA’s. Nucleic
Acids Research, 12, 3895–3907.
Wan, J., Hickey, M. W., & Coventry, M. J. (1995). Continuous
production of bacteriocins, brevicin, nisin, and pediocin, using
calcium alginate-immobilized bacteria. Journal of Applied Bacter-
iology, 79, 671–676.
Wan, J., Wilcock, A., & Coventry, M. J. (1998). The effect of essential
oils of basil on the growth of Aeromonas hydrophila and
Pseudomonas fluorescens. Journal of Applied Microbiology, 84,
152–158.
Welsh, J. K., Skurrie, I. J., & May, J. T. (1978). Use of Semliki forest
virus to identify lipid-mediated antiviral activity and anti-alpha-
virus immunoglobulin A in human milk. Infection and Immunity,
19, 395–401.
Yamauchi, K., Tomita, M., Giehl, T. J., & Ellison, R. T., III (1993).
Antibacterial activity of lactoferrin and a pepsin-derived lactoferrin
peptide fragment. Infection and Immunity, 61, 719–728.
Zucht, H. -D., Forssmann, W. -G., Raida, M., & Adermann, K.
(1996). Procedure for extraction of an antibiotically active
preparation from bovine milk and for its synthesis. German Patent
Application DE 44 44 753 A1.
Zucht, H. D., Raida, M., Adermann, K., Magert, H. J., & Forssman,
W. G. (1995). Casocidin-I: a casein-aS2 derived peptide exhibits
antibacterial activity. FEBS Letters, 372, 185–188.
Copyright © 2022 FDOKUMEN




















