
Is the Gibraltar Strait a barrier to gene flow for the bat Myotis myotis (Chiroptera:...
12
Molecular Ecology (2000) 9 , 1761–1772 © 2000 Blackwell Science Ltd Blackwell Science, Ltd Is the Gibraltar Strait a barrier to gene flow for the bat Myotis myotis (Chiroptera: Vespertilionidae)? V. CASTELLA,* M. RUEDI,*† L. EXCOFFIER,‡ C. IBÁÑEZ,§ R. ARLETTAZ* and J. HAUSSER* * Laboratoire de Zoologie, Institut d’Ecologie, Bâtiment de Biologie, Université de Lausanne, CH-1015 Lausanne, Switzerland, ‡ Laboratoire de Génétique et Biométrie, Département d’Anthropologie, Université de Genève, 1211 Genève 24, Switzerland, § Estación Biológica de Doñana, CSIC, Apartado 1056, 41080 Sevilla, Spain Abstract Because of their role in limiting gene flow, geographical barriers like mountains or seas often coincide with intraspecific genetic discontinuities. Although the Strait of Gibraltar represents such a potential barrier for both plants and animals, few studies have been conducted on its impact on gene flow. Here we test this effect on a bat species ( Myotis myotis ) which is apparently distributed on both sides of the strait. Six colonies of 20 Myotis myotis each were sampled in southern Spain and northern Morocco along a linear transect of 1350 km. Results based on six nuclear microsatellite loci reveal no significant population structure within regions, but a complete isolation between bats sampled on each side of the strait. Variability at 600 bp of a mitochondrial gene (cytochrome b ) confirms the existence of two genetically distinct and perfectly segregating clades, which diverged several million years ago. Despite the narrowness of the Gibraltar Strait (14 km), these molecular data suggest that neither males, nor females from either region have ever reproduced on the opposite side of the strait. Comparisons of molecular divergence with bats from a closely related species ( M. blythii ) suggest that the North African clade is possibly a distinct taxon warranting full species rank. We provisionally refer to it as Myotis cf punicus Felten 1977, but a definitive systematic understanding of the whole Mouse-eared bat species complex awaits further genetic sampling, especially in the Eastern Mediterranean areas. Keywords : Chiroptera, cytochrome b , Gibraltar, microsatellite, Myotis , population structure Received 27 February 2000; revision received 5 June 2000; accepted 5 June 2000 Introduction Recent reviews of intraspecific genetic variation (Taberlet et al . 1998; Avise & Walker 1999; Hewitt 1999) have shown that many animal or plant species present important genetic discontinuities when they are sampled over wide ranges. These discontinuities often result from recent admixture of populations, which diverged in allopatry (e.g. in glacial refugia) or coincide with current or historical barriers to gene flow (e.g. mountain ranges or ancient glaciers). The Gibraltar Strait, which separates the Iberian Peninsula from the Maghreb (Fig. 1) by a minimum gap of 14 km of open sea, could thus represent a barrier to gene flow. Indeed, since the last Messinian crisis (about 5.5 Ma) when most of the present-day Mediterranean Sea dried-up, no land bridge existed between these two land masses (see, e.g. Steininger et al . 1985). Thus, dispersal of both plants and animals across this Strait must have been severely limited since the Pliocene. Restricted over-water dispersal is suggested by Valdés (1991) who estimated that as much as 25% of the 3500 species of plants distributed in southern Spain and northern Morocco only occur in one side of the Strait. For mammals, even highly vagile species like the wolf ( Canis lupus ) or the jackal ( Canis aureus ) are currently found in Iberia and the Maghreb, respectively, but neither apparently ever crossed the Gibraltar Strait successfully. Genetic studies in Wild mice ( Mus spretus ) and Wood mice ( Apodemus sylvaticus ) have also suggested that the Gibraltar Strait is an effective barrier to gene flow (Boursot et al . 1985; Filippucci 1992). More generally, Dobson (1998) Correspondence: M. Ruedi: †Present address: Muséum d’histoire naturelle, CP 6434, CH-1211 Genève 6, Switzwerland. Fax: + 41 22 4186301; E-mail: [email protected]
Transcript of Is the Gibraltar Strait a barrier to gene flow for the bat Myotis myotis (Chiroptera:...

Molecular Ecology (2000)
9
, 1761–1772
© 2000 Blackwell Science Ltd
Blackwell Science, Ltd
Is the Gibraltar Strait a barrier to gene flow for the bat
Myotis myotis
(Chiroptera: Vespertilionidae)?
V. CASTELLA,* M. RUEDI , *† L . EXCOFFIER,‡ C . IBÁÑEZ,§ R . ARLETTAZ* and J . HAUSSER**
Laboratoire de Zoologie, Institut d’Ecologie, Bâtiment de Biologie, Université de Lausanne, CH-1015 Lausanne, Switzerland,
‡
Laboratoire de Génétique et Biométrie, Département d’Anthropologie, Université de Genève, 1211 Genève 24, Switzerland,
§
Estación Biológica de Doñana, CSIC, Apartado 1056, 41080 Sevilla, Spain
Abstract
Because of their role in limiting gene flow, geographical barriers like mountains or seasoften coincide with intraspecific genetic discontinuities. Although the Strait of Gibraltarrepresents such a potential barrier for both plants and animals, few studies have beenconducted on its impact on gene flow. Here we test this effect on a bat species (
Myotis myotis
)which is apparently distributed on both sides of the strait. Six colonies of 20
Myotis myotis
each were sampled in southern Spain and northern Morocco along a linear transect of1350 km. Results based on six nuclear microsatellite loci reveal no significant populationstructure within regions, but a complete isolation between bats sampled on each side of thestrait. Variability at 600 bp of a mitochondrial gene (cytochrome
b
) confirms the existence oftwo genetically distinct and perfectly segregating clades, which diverged several million yearsago. Despite the narrowness of the Gibraltar Strait (14 km), these molecular data suggest thatneither males, nor females from either region have ever reproduced on the opposite sideof the strait. Comparisons of molecular divergence with bats from a closely related species(
M. blythii
) suggest that the North African clade is possibly a distinct taxon warrantingfull species rank. We provisionally refer to it as
Myotis
cf
punicus
Felten 1977, but a definitivesystematic understanding of the whole Mouse-eared bat species complex awaits furthergenetic sampling, especially in the Eastern Mediterranean areas.
Keywords
: Chiroptera, cytochrome
b
, Gibraltar, microsatellite,
Myotis
, population structure
Received 27 February 2000; revision received 5 June 2000; accepted 5 June 2000
Introduction
Recent reviews of intraspecific genetic variation (Taberlet
et al
. 1998; Avise & Walker 1999; Hewitt 1999) have shownthat many animal or plant species present important geneticdiscontinuities when they are sampled over wide ranges.These discontinuities often result from recent admixtureof populations, which diverged in allopatry (e.g. in glacialrefugia) or coincide with current or historical barriers togene flow (e.g. mountain ranges or ancient glaciers). TheGibraltar Strait, which separates the Iberian Peninsulafrom the Maghreb (Fig. 1) by a minimum gap of 14 km ofopen sea, could thus represent a barrier to gene flow. Indeed,
since the last Messinian crisis (about 5.5 Ma) when mostof the present-day Mediterranean Sea dried-up, no landbridge existed between these two land masses (see, e.g.Steininger
et al
. 1985). Thus, dispersal of both plants andanimals across this Strait must have been severely limitedsince the Pliocene. Restricted over-water dispersal issuggested by Valdés (1991) who estimated that as muchas 25% of the 3500 species of plants distributed in southernSpain and northern Morocco only occur in one side of theStrait. For mammals, even highly vagile species like thewolf (
Canis lupus
) or the jackal (
Canis aureus
) are currentlyfound in Iberia and the Maghreb, respectively, but neitherapparently ever crossed the Gibraltar Strait successfully.Genetic studies in Wild mice (
Mus spretus
) and Wood mice(
Apodemus sylvaticus
) have also suggested that the GibraltarStrait is an effective barrier to gene flow (Boursot
et al
.1985; Filippucci 1992). More generally, Dobson (1998)
Correspondence: M. Ruedi: †Present address: Muséum d’histoirenaturelle, CP 6434, CH-1211 Genève 6, Switzwerland. Fax: + 4122 4186301; E-mail: [email protected]
MEC1069.fm Page 1761 Tuesday, October 10, 2000 10:18 AM

1762
V. C A S T E L L A
E T A L
.
© 2000 Blackwell Science Ltd,
Molecular Ecology
, 9, 1761–1772
considers that only four of the 17 nonflying mammalspecies occurring wild in both sides of the Strait were nottransported by man. In contrast, H. Dobson stresses thatmost bat species found in north-west Africa (26 species)and in Iberia (25 species) are naturally occurring in bothregions, and are thus much better over-water dispersers.In fact, several other taxa are distributed on both sides ofthe Strait, but its impact as a geographical barrier torestrict gene flow between populations has rarely beenaddressed specifically. For instance, a genetic survey ofrare plants distributed in southern Spain and Morocco(genus
Androcymbium
, Colchicaceae) revealed that popu-lation differentiation was similar when comparisons weremade within countries or across Gibraltar, suggesting thatthis strait was not a strong barrier for these species (Pedrola-Monfort & Caujapé-Castells (1994). More recently, Burban
et al
. (1999) found that both the maritime pine and itshomopteran parasite
Matsococcus feytaudi
shared commonhaplotypes in southern Spain and extreme northernMorocco, while populations from central Morocco werecompletely distinct.
The Greater Mouse-eared bat (
Myotis myotis
) is awidespread species, which is apparently distributed fromMorocco to Europe and the Middle east, including severalMediterranean islands (reviewed in Benda & Horacek1995; Arlettaz
et al
. 1997b). It also probably occurs in theremote Azore Islands, more than 1400 km west from theIberian Peninsula (Palmeirim 1979). Ringing studies haveshown that this strong flier is able to cover annually severalhundreds of kilometers between summer and winter
roosts (Horacek 1985; Paz
et al
. 1986). Moreover, femalesmay commute daily up to 25 km between their nurserycolonies and the feeding territories during lactation(Arlettaz 1996, 1999). Owing to such behavioural charac-teristics, we can anticipate that populations of the GreaterMouse-eared bat found on both sides of the GibraltarStrait may exchange migrants across the channel. We,therefore, assessed the impact of the strait as a barrier bycomparing levels of gene flow within southern Spain andwithin North Africa, and across Gibraltar, using a com-bination of nuclear and mitochondrial DNA (mtDNA)markers. These two classes of genetic markers enabledus to estimate in which proportion males and femalescontributed to the migrant pool, and provided a phylo-genetic perspective of their evolution.
Materials and methods
Taxonomic and geographical sampling
As European populations of Mouse-eared bats areprotected in most countries, animals were caught andsampled noninvasively under appropriate license (seeAcknowledgements). Upon capture, individuals wereidentified according to their lengths of forearm, ear andupper toothrow, (Arlettaz
et al
. 1997b) and sexed. Atotal of 119 bats from six colonies were sampled with amedical biopsy punch on the membrane of each wing(Worthington Wilmer & Barratt 1996). Animals werereleased back to their colony within one hour of capture.
Fig. 1 Distribution of the Greater Mouse-eared bat (Myotis myotis) in the westernMediterranean area (in black) accordingto Arlettaz et al. (1997b). The broken linedelineates the limits of the North Africanform, referred here as M. cf punicus Felten1977. The inset provides an enlargement ofthe area around the Gibraltar Strait. Numbersindicate the position of the following colonies(locality, province, country): Boumahden,Agadir, Morocco (1); Azrou, Meknes, Morocco(2); Mina del Agua, Ceuta, Spain (3); Cuevadel Agua, Cadiz, Spain (4); Canillas deAceituno, Malaga, Spain (5); Denia, Alicante,Spain (6); Inca, Mallorca, Spain (7); Oleta,Corsica, France (8); Ulassai, Sardinia, Italy(9). See also the Appendix for locations ofother examined specimens and for thoseof M. blythii.
MEC1069.fm Page 1762 Tuesday, October 10, 2000 10:18 AM

I S G I B R A LTA R A B A R R I E R TO G E N E F L O W F O R B AT S ?
1763
© 2000 Blackwell Science Ltd,
Molecular Ecology
, 9, 1761–1772
Three colonies of Mouse-eared bats were sampled oneach side of the Strait (Fig. 1). A distance of 270–770 kmseparated the North African colonies (nos 1–3; geographicaldistances were measured on a map and rounded to thenearest 10 km), while those from southern Spain (nos 4–6)were sampled at intervals of 130–580 km. These sixcolonies represent animals living along a nearly lineartransect of 1350 km. The colony of Ceuta (referred herecollectively with the Moroccan colonies, though is politicallya province of Spain) is located just in front of the Iberiancolony of Cadiz. They are separated by only 60 km ofland and 14 km of sea (see inset of Fig. 1). Because thereis a long lasting controversy about the taxonomic posi-tion of North African Mouse-eared bats (Felten
et al
. 1977;Arlettaz
et al
. 1997b), 18 Lesser Mouse-eared bats (
Myotisblythii
) coming from various places in Europe and Asiawere analysed for comparative purposes. We also includedseveral Mouse-eared bats from Mallorca, Corsica, andSardinia (see Fig. 1 and Appendix).
Myotis nattereri
, asmaller species closely related to the large
Myotis
(Ruedi& Mayer submitted), was used as an outgroup to root thephylogenetic trees.
DNA amplification
DNA was extracted from half wing punches using a saltprotocol (Miller
et al
. 1988). For the purpose of measuringthe genetic structure of bat populations at the nuclearlevel, six microsatellite loci developed specifically forthe Greater Mouse-eared bat were analysed (Table 1). TheGenBank accession nos and polymerase chain reaction(PCR) conditions used to amplify these loci are detailedin Castella & Ruedi (2000). The female contribution togene flow was evaluated by sequencing the first 600 bp ofa mtDNA marker, the cytochrome
b
gene. This gene wasamplified with primers L14724 (Kocher
et al
. 1989) and
MVZ16 (Smith & Patton 1993), and sequenced with anautomated DNA sequencer (Applied Biosystems model373XL) following standard methods. This mitochondrialgene was sequenced in the following subsample ofanimals from the transect: 14
M. myotis
from Spain (10from Cadiz, two each from Malaga and Alicante), and 14from Morocco (10 from Ceuta, and two each from Azrouand Agadir). The same gene was also sequenced in theother
Myotis
sampled elsewhere in Europe and Asia (seeAppendix). Sequences were aligned and edited visuallyusing Sequencher 3.0 (Gene Codes Corp.). All differenthaplotypes are deposited in GenBank under accessionnos AF246241–AF246261. Abbreviation of haplotypes orpopulations refer to Morocco (MA), Spain (ES), Sardinia(SA), Corsica (CO), Switzerland (CH), Greece (GR), theCzech Republic (TS), and Kirghizstan (KZ).
Microsatellite analysis
The genetic variability found within the six populationswas quantified by the number of alleles and by theobserved (
H
O
) and expected (
H
E
, Nei 1987) heterozygosity.Means and standard deviations of these variables werecalculated separately for both the three pooled Spanishand the three pooled Moroccan colonies. To determinewhether the six surveyed loci represent independentsamples of the Mouse-eared bats’ genome, genotypiclinkage disequilibrium was investigated for each pairof loci using the program
genepop
3.1
c
(Raymond &Rousset 1995). Genotypic proportions were tested fordeparture from Hardy–Weinberg (H–W) expectationsusing randomization tests available in the
fstat
2.9package (Goudet 1999a). For H–W expectations withinpopulations, alleles were permuted 1000 times amongindividuals, while alleles were permuted among popu-lations for H–W expectations within regions. The same
Table 1 Observed (HO) and expected heterozygosity (HE), number of alleles (A) and inbreeding coefficient (FIS) of Myotis myotispopulations calculated for each locus and over all loci. The mean values and standard deviations (SD) were calculated among threenursery colonies within each country. Twenty bats were sampled in each population, except in Ceuta (Moroccan side) where only 19individuals were captured
Spanish populations Moroccan populations
HO HE A FIS HO HE A FIS
B11 0.87 ± 0.06 0.83 ± 0.02 8.3 ± 0.6 –0.038 0.54 ± 0.14 0.56 ± 0.10 5.0 ± 1.0 0.033C113 0.12 ± 0.03 0.11 ± 0.03 2.0 ± 0.0 –0.039 0.03 ± 0.03 0.03 ± 0.03 1.7 ± 0.6 0.000D9 0.92 ± 0.06 0.90 ± 0.03 12.7 ± 0.6 –0.017 0.80 ± 0.08 0.88 ± 0.06 11.0 ± 2.0 0.092E24 0.83 ± 0.08 0.85 ± 0.08 13.7 ± 1.2 0.021 0.98 ± 0.03 0.92 ± 0.01 14.3 ± 1.2 –0.069G25 0.55 ± 0.09 0.62 ± 0.01 3.7 ± 1.2 0.114 0.03 ± 0.06 0.07 ± 0.06 1.7 ± 0.6 0.493H29 0.90 ± 0.05 0.91 ± 0.01 11.7 ± 1.5 0.013 0.83 ± 0.08 0.88 ± 0.04 10.3 ± 0.6 0.054Overall 0.67 ± 0.03 0.71 ± 0.02 8.7 ± 0.3 0.012 0.49 ± 0.04 0.56 ± 0.02 7.3 ± 0.6 0.035
MEC1069.fm Page 1763 Tuesday, October 10, 2000 10:18 AM

1764
V. C A S T E L L A
E T A L
.
© 2000 Blackwell Science Ltd,
Molecular Ecology
, 9, 1761–1772
program was used to test whether inbreeding coefficients(
F
IS
) deviated significantly from zero. In order to getmean
F
IS
values for each region, computations were runindependently for the two sets of three colonies. In bothcases,
P
-values were obtained following 1800 permuta-tions of alleles within populations with the specificalternative hypothesis of heterozygote deficiency. SequentialBonferroni corrections were used to compute the criticalsignificance levels for all simultaneous statistical tests(Rice 1989).
Nuclear population structure was quantified byestimating the differentiation between colonies with
F
-statistics (Wright 1978). Pairwise
F
ST
were computedaccording to Weir & Cockerham (1984) and populationdifferentiation was tested by randomizing (1500 permuta-tions) multilocus genotypes between each pair ofsamples with
fstat
2.9. A hierarchical design (Schneider
et al
. 1996), including the colony, the regional and thewhole transect levels, was also used to test for theeffect of the Gibraltar Strait on the apportionment ofmolecular variance. To fully exploit genetic informationat the individual level, the multilocus genotype of eachbat was introduced in a principal component analysisusing the program
pca-gen
1.2 (Goudet 1999b). As withassignment tests (Paetkau
et al
. 1995, 1997), this methodallows the identification of alien genotypes within localpopulation.
Haplotype analysis and phylogenetic reconstructions
The small number of bats sequenced in most populationsprecluded any test of a hierarchical design of populationdifferentiation. Contrary to the microsatellites, we onlycompared haplotypic variability within regions (i.e. Spainor Morocco) and among regions. Thus, the mitochondrialpopulation structure was analysed by estimating thecorrelation of haplotypes drawn from the same side ofthe Gibraltar Strait to haplotypes drawn randomly fromthe total sample in terms of
φ
ST
(Excoffier
et al
. 1992).Patterns of DNA evolution among haplotypes and allphylogenetic reconstructions were done with
paup
4.0
b
4(Swofford 1996). Phylogenetic relationships were recoveredwith a uniform parsimony analysis (branch-and-boundsearch), and reliability of nodes ascertained with 500bootstrap resampling. Other methods of phylogeneticinference (i.e neighbour-joining or maximum likelihoodmethods) gave similar results. Allozyme data previouslyused to differentiate
M. myotis
and
M. blythii
(Arlettaz
et al
. 1997b) were reanalysed here in a phylogenetic contextbased on a complete data set of 35 loci (see Ruedi
et al
.1990). Populations analysed here included
M. myotis
fromSpain (
n
= 12), and Switzerland (
n
= 5),
M. blythii
fromSpain (
n
= 9), Switzerland (
n
= 5) and Kighizstan (
n
= 7),Moroccan bats (
n
= 7) and a composite population of
three bats from Sardinia and two from Corsica. Allozymeallele frequencies were transformed into Nei’s geneticdistance (Nei 1987) and phylogenetic relationships werereconstructed with the
neighbour
subroutine in theprogram
phylip
3.5
c
(Felsenstein 1993). The resulting treewas also bootstrapped 500 times with
phylip
.
Results
Microsatellite variability
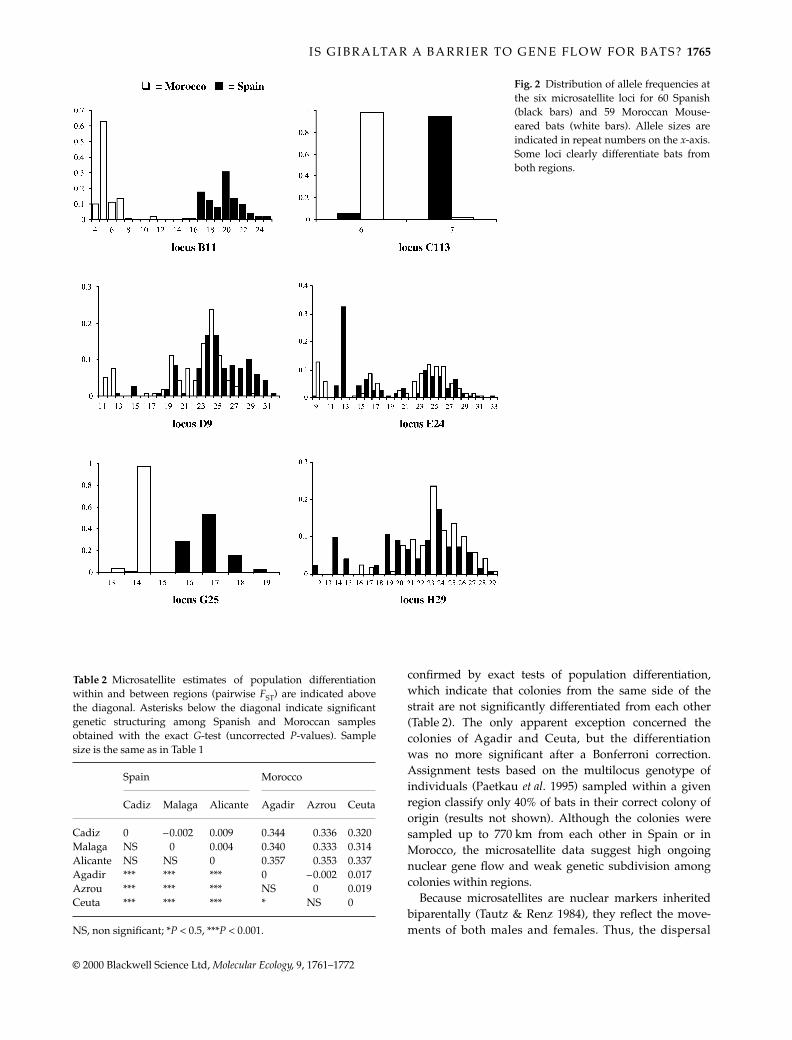
Within all colonies, bats showed considerable geneticvariability at the six microsatellite loci with total numberof alleles ranging from 41 to 54 and gene diversity from0.54 to 0.72 (Table 1; individual genotypes are availableat http://www.unil.ch/izea/research.html#mmyotis). Theloci C113 and G25 were the least variable as they werenearly fixed in one or both regions (Table 1 and Fig. 2).One test of linkage disequilibrium out of the 15 possiblewas highly significant in the colony of Ceuta (
P
< 0.001)and involved loci D9 and H29. This association remainedsignificant even after a sequential Bonferroni procedure(Rice 1989). In all other populations, no evidence of associa-tion was detected for any locus combination. Because onesignificant test out of 15 would be expected by chancealone at the population level (
P
> 0.1; binomial test with
α
= 0.05), there was no evidence for strong dependencebetween loci. All colonies, either considered independentlyor grouped by region, were in Hardy–Weinberg equilibriumfor each locus and over all loci. The only population whichhad an inbreeding coefficient that differed significantlyfrom zero (
F
IS
= 0.105, Azrou) was no more deviating fromrandom mating when sequential Bonferroni correctionwas applied. The high (but not significant)
F
IS
value forthe locus G25 in Moroccan populations (Table 1) was dueto the occurrence of one particular individual in thecolony of Ceuta. This bat was homozygous for a rareMoroccan allele (13 repeats; Fig. 2), while all remainingbats were either homozygous for the most common allele(56 bats) or heterozygous for both alleles (2 bats).
Patterns of genetic variation within regions
A total of 69 and 56 alleles were scored in the threeSpanish and three Moroccan colonies, respectively. Mostof these alleles were widespread among colonies withinregions, except 19 and 10 of them which were limited toone Iberian or one Moroccan colony, respectively. Themajority of these private alleles only occurred at very lowfrequencies (mostly one single copy). The rather uniformnumber of alleles and heterozygosity calculated amongcolonies (low standard deviations in Table 1) also suggeststhat the microsatellite variability is high but evenly dis-tributed among populations from the same region. This is
MEC1069.fm Page 1764 Tuesday, October 10, 2000 10:18 AM
I S G I B R A LTA R A B A R R I E R TO G E N E F L O W F O R B AT S ?
1765
© 2000 Blackwell Science Ltd,
Molecular Ecology
, 9, 1761–1772
confirmed by exact tests of population differentiation,which indicate that colonies from the same side of thestrait are not significantly differentiated from each other(Table 2). The only apparent exception concerned thecolonies of Agadir and Ceuta, but the differentiationwas no more significant after a Bonferroni correction.Assignment tests based on the multilocus genotype ofindividuals (Paetkau et al. 1995) sampled within a givenregion classify only 40% of bats in their correct colony oforigin (results not shown). Although the colonies weresampled up to 770 km from each other in Spain or inMorocco, the microsatellite data suggest high ongoingnuclear gene flow and weak genetic subdivision amongcolonies within regions.
Because microsatellites are nuclear markers inheritedbiparentally (Tautz & Renz 1984), they reflect the move-ments of both males and females. Thus, the dispersal
Fig. 2 Distribution of allele frequencies atthe six microsatellite loci for 60 Spanish(black bars) and 59 Moroccan Mouse-eared bats (white bars). Allele sizes areindicated in repeat numbers on the x-axis.Some loci clearly differentiate bats fromboth regions.
Table 2 Microsatellite estimates of population differentiationwithin and between regions (pairwise FST) are indicated abovethe diagonal. Asterisks below the diagonal indicate significantgenetic structuring among Spanish and Moroccan samplesobtained with the exact G-test (uncorrected P-values). Samplesize is the same as in Table 1
Spain Morocco
Cadiz Malaga Alicante Agadir Azrou Ceuta
Cadiz 0 –0.002 0.009 0.344 0.336 0.320Malaga NS 0 0.004 0.340 0.333 0.314Alicante NS NS 0 0.357 0.353 0.337Agadir *** *** *** 0 –0.002 0.017Azrou *** *** *** NS 0 0.019Ceuta *** *** *** * NS 0
NS, non significant; *P < 0.5, ***P < 0.001.
MEC1069.fm Page 1765 Tuesday, October 10, 2000 10:18 AM

1766 V. C A S T E L L A E T A L .
© 2000 Blackwell Science Ltd, Molecular Ecology, 9, 1761–1772
of one sex, e.g. the males only, would be sufficient tohomogenize populations, even in the absence of femaledispersal. We, therefore, measured patterns of geneflow through mtDNA variation which is transmitted byfemales only (Avise 1994). Twenty-eight individuals weresequenced for 600 bp of the cytochrome b gene (seeAppendix). The 14 Greater Mouse-eared bats from Spaindiffered at only two positions (one transition and onetransversion substitution), determining three differenthaplotypes. The most common haplotype (ES1) was foundin nine bats from Cadiz and two from Malaga. Similarly,only three positions were variable (all transitions) in thecytochrome b of the 14 Moroccan bats, determining fourclosely related haplotypes. The most common Moroccanhaplotype (MA1) was found in all individuals fromCeuta, and one bat from Azrou. Some haplotypes werefound in one colony alone (e.g. MA3 from Agadir, or ES3from Alicante) suggesting that females may be morephilopatric than males to their natal colonies. The lownumber of sequenced individuals in most colonies pre-cludes any formal testing of this hypothesis, but a similarbias of sex dispersal was already suggested in a populationof M. myotis from Germany (Petri et al. 1997).
Patterns of genetic variation across the Gibraltar Strait
In sharp contrast to the genetic homogeneity andhigh levels of nuclear gene flow inferred within regions,comparisons of bats sampled across the GibraltarStrait yielded strong levels of genetic differentiation. Forthe microsatellite data set, three loci showed contrastingpatterns of variation. In the B11 locus, none of the 17different alleles was distributed on both sides of the Strait.All alleles comprising 4–11 repeats were found exclusivelyin Morocco, while all those comprising 15–25 repeatswere typical of Spanish bats (Fig. 2). In the G25 locus, asingle copy (out of 120 sampled) of the 14-repeat allelewas found in the Spanish population of Cadiz, whilethis allele was nearly fixed (mean frequency 0.97) in allMoroccan colonies (Fig. 2). Finally, the locus C113 wasnearly fixed for the 7-repeat allele in Spain (averagefrequency = 0.94) and for the 6-repeat allele in Morocco
(mean frequency 0.98; Fig. 2). For both the G25 and C113loci, results from a broader population survey involving420 M. myotis from western Europe (V. Castella & M. Ruedi,unpublished data) showed that rare ‘Moroccan’ allelescan be found sporadically in several European populationssampled as far away as Switzerland or Poland. The threeremaining loci showed more overlapping allele distributionacross the strait (Fig. 2).
Contrary to the situation observed within regions,all pairs of populations were significantly differentiatedwhen comparisons were made across the Gibraltar Strait,even after Bonferroni adjustment (Table 2). Moreover,classical FST values were one or two orders of magnitudelarger when pairs of populations were compared betweenregions rather than within regions (Table 2). As a con-sequence, assignment tests among regions classified allbats in their correct country of origin (results not shown). Ahierarchical analysis of molecular variance showed thatmost of the microsatellite variability (66%) was distributedwithin colonies. However, along the whole transect of1350 km, only a negligible part of the genetic variance(0.5%) was due to differences among colonies, while 33%was explained by the effect of the Gibraltar Strait (Table 3).
In order to make comparisons with a closely relatedspecies, we analysed the same six microsatellite loci in 18M. blythii sampled in various locations in Europe andAsia (see Appendix). This heterogeneous sample included,in particular, two individuals from a nursery colony inTarifa (Spain) located just on the Iberian border of theGibraltar Strait (Fig. 1). Among the 52 alleles that werescored in this new sample, 20 were exclusively found inM. blythii. In particular, most alleles (15 out of 17) of thedinucleotide locus E24 showed single nucleotide shiftswhen compared to Iberian and Moroccan alleles. Thissuggests that an insertion or deletion unique to M. blythiihas occurred within this locus. In the diallelic C113 locus,both the 6-repeat (typical of Moroccan bats) and the 7-repeat allele (typical of Iberian bats) were present in thisspecies. The 6-repeat allele was the most frequent(frequency of 0.72) while the other allele was found in 10heterozygotes. All M. blythii tested were fixed for the17-repeat allele at the G25 locus; this allele is otherwise
Microsatellites mtDNA
% Φ % Φ
Among regions 33.3 0.333 99.1 0.991Among populations within region 0.5 0.007 — * — *Within populations 66.2 0.338 0.9 —
*hierarchical level not included for mtDNA.
Table 3 Analysis of microsatellite andmtDNA variance across the GibraltarStrait. The percentage of variance (%) andthe Φ-statistic are given at each hierarchicallevel (see Excoffier et al. 1992). Microsatelliteresults are based on the same sample sizeas in Table 1, but for mtDNA only 14 batswere analysed for Spanish and for Moroccanpopulations (see text)
MEC1069.fm Page 1766 Tuesday, October 10, 2000 10:18 AM

I S G I B R A LTA R A B A R R I E R TO G E N E F L O W F O R B AT S ? 1767
© 2000 Blackwell Science Ltd, Molecular Ecology, 9, 1761–1772
typical of M. myotis from the Iberian region. As shown bya principal component analysis, multilocus genotypes ofthese 18 M. blythii appear to be more closely related to theIberian M. myotis samples, than to the Moroccan bats(Fig. 3).
To come back to our main species, patterns of mtDNAvariation across the Gibraltar Strait supports the sameconclusion as the nuclear markers. Nearly all the molecu-lar variance was observed for comparisons made acrossthe Strait (99.1%; Table 3). Thus, all haplotypes could beunambiguously assigned to their correct region of origin.The two groups of haplotypes differed by a mean (± SD)genetic distance of 10.6 ± 0.2% (Kimura 2-parameter cor-rection). This represents 54–59 observed substitutionsover the 600 bp sequenced. To expand the geographicalcoverage of Mouse-eared bats from the Mediterraneanregion, we also sequenced animals from other parts ofEurope and from the islands of Mallorca (Spain), Corsica(France), and Sardinia (Italy), and also compared sequencesfrom M. blythii (see Appendix). M. myotis from Spain(including bats from Mallorca) and Switzerland were allvery similar to each other (within 1% of sequence diver-gence). Surprisingly, four Spanish M. blythii from Tarifa,Cadiz and Alicante had the same haplotype (ES1b) that isfound in most Iberian M. myotis. M. blythii from the Czech
Republic and Greece were slightly more divergent (up to2.8% sequence divergence), while the two central AsianLesser Mouse-eared bats differed by a mean of 6.2 ± 0.4%from European samples. By contrast, sequences fromCorsica and Sardinia were all closely related to the Moroccanhaplotypes. Thus, European M. myotis and M. blythii aremuch more closely related to each other than to theMoroccan sample. Accordingly, a phylogenetic analysis ofall different haplotypes places the Moroccan, Corsican,and Sardinian samples in a single, well-supported clade(Fig. 4A). All other haplotypes form a distinct clade, alsohighly supported by bootstrap resampling. This cladeinclude two Kirghiz M. blythii which emerge first as anearly offshoot, while sequences of European M. blythiiand M. myotis constitute a closely related crown-groupwhich did not attain reciprocal monophyly.
Phylogenetic analysis of Mouse-eared bat populationsusing 35 allozyme loci and M. nattereri as an outgroupconfirms the close relationships of Moroccan bats withthose of Corsica and Sardinia (93% bootstrap support;Fig. 4B). However, contrary to results of microsatellites(Fig. 3) and DNA sequencing (Fig. 4A), this allozyme treesuggests a closer phylogenetic relationship of this NorthAfrican taxon with M. myotis, but this node is not firmlyestablished (63% bootstrap support).
Fig. 3 Factor map of the two main axis of the Principal Component Analysis (PCA) carried out with individual microsatellitegenotypes. The x-axis represents 30% inertia (FST = 0.19), while the y-axis has 7% inertia (FST = 0.04). All Spanish bats (represented byblack triangles) display positive values on the first axis, while negative values characterize Moroccan bats (represented by whitesquares). No individual of these two groups lay in an intermediate position. European Myotis blythii, represented by grey circles,appear more closely related to Iberian rather than to Moroccan bats.
MEC1069.fm Page 1767 Tuesday, October 10, 2000 10:18 AM

1768 V. C A S T E L L A E T A L .
© 2000 Blackwell Science Ltd, Molecular Ecology, 9, 1761–1772
Discussion
Greater Mouse-eared bats are strong fliers able to coverseveral hundreds of kilometers annually (Horacek 1985;Paz et al. 1986). Consistent with this high dispersal ability,microsatellite data show that colonies within Spain, orwithin Morocco are only weakly differentiated (Table 2).This suggests that high levels of gene flow prevail amongcontinental colonies over large geographical distances (atleast up to 770 km). Accordingly, all mtDNA haplotypessequenced within Spain or within Morocco were identicalor very similar to each other. These patterns of microsatelliteand mtDNA variation within each region are consistentwith recent genetic data obtained for another Palaearcticspecies of bat, Nyctalus noctula. This true migratory speciescovers several hundreds of kilometers every year betweenbreeding and wintering grounds, and exhibits no significantnuclear genetic differentiation within a core area spawningmore than 3000 km (Petit & Mayer 1999).
In sharp contrast to the weak differentiation of popula-tions observed within Spain or within Morocco, the same
molecular markers revealed a clear dichotomy whencomparisons were made across the Gibraltar Strait. Forinstance, the Moroccan and Iberian populations share nomtDNA haplotypes in common, and at the microsatellitelocus B11, all short alleles are exclusively found in NorthAfrica, while longer repeat alleles are all Iberian (Fig. 2).Thus, haplotypes and multilocus genotypes of all sampledindividuals can be easily assigned to their correct regionof origin (Figs 3 and 4A). The 119 bats analysed from onlysix populations may represent a small sample to detectdirect migrants. This is certainly true for first generationmigrants which could occur in the area without repro-ducing, and which could have remained unsampled.However, unlike direct methods (e.g. ringing studies),which provide punctual information on the movementsof individuals, genetic data are representative of themovement of genes accumulated during many genera-tions (Slatkin 1994). In a principal component analysisbased on the multilocus genotype of each individual,recent hybrid genotypes are expected to occur in an inter-mediate position between pure-bred genotypes (but seeLugon-Moulin et al. 1999). As shown in Fig. 3, all Iberianand Moroccan bats appear to be pure-bred, with noapparent sign of recent admixture of both groups. BecausemtDNA markers are inherited clonally through the mother,they could introgress into a new population through asingle reproducing, alien female. Yet haplotypes segre-gate in two clades endemic to each region. Thus, theevidence supports that no Spanish migrant contributed toany significant gene flow into the Moroccan populationsor vice-versa. Moreover, the amount of genetic divergenceamong cytochrome b haplotypes measured betweenEuropean Myotis myotis and African bats (10.6 ± 0.2% forKimura 2-parameter corrected distance) suggests thatthey became separated long ago. Comparable amountsof genetic divergence were measured between the twocryptic species of pipistrelle bats (11% Barratt et al. 1997)or among congeneric mammals in general (reviewed inJohns & Avise 1998). To maintain such high levels of geneticdifference between conspecific populations, a strong,persistent and ancient barrier preventing any significantgene flow would be needed. Fossils of typical M. myotisare known at least since the Pleistocene in the IberianPeninsula (Sevilla 1989). Thus, owing to their vagility, thesebats should have had ample time to exchange migrantswith North Africa. Moreover, Mouse-eared bats havesuccessfully colonized all major islands of the MediterraneanSea, suggesting that they can be very effective colonizers.For instance, bats on Mallorca, which is about 200 km awayfrom mainland populations (Fig. 1), have haplotypes ident-ical or very similar to Spanish ones (Fig. 4A). Both thesetemporal and physical arguments suggest that the 14 kmof open sea, which separates Spain from Morocco (Fig. 1),is certainly not sufficient per se to prevent gene flow.
Fig. 4 Phylogenetic relationships of various Mouse-eared batsdeduced from mtDNA haplotypes (gene tree A), or from allozymedata (population tree B). The haplotype tree is a consensus of allequally parsimonious trees (score 154 steps) obtained from a branch-and-bound search using Myotis nattereri as an outgroup. Noticethat haplotypes of M. myotis and M. blythii are not reciprocallymonophyletic. The allozyme tree is a neighbour-joining tree basedon Nei’s (Nei 1987) genetic distances calculated among popu-lations. Nodes with bootstrap proportions above 50% are indicated.Abbreviation of haplotypes or populations refer to samplesfrom Morocco (MA), Spain (ES), Sardinia (SA), Corsica (CO),Switzerland (CH), Greece (GR), the Czech Republic (TS), andKirghizstan (KZ).
MEC1069.fm Page 1768 Tuesday, October 10, 2000 10:18 AM

I S G I B R A LTA R A B A R R I E R TO G E N E F L O W F O R B AT S ? 1769
© 2000 Blackwell Science Ltd, Molecular Ecology, 9, 1761–1772
We excluded the possibility that the North African batswere conspecific with another known species, namely M.blythii, as formerly hypothesized (Bogan et al. 1978).According to the microsatellite data (Fig. 3) and to themolecular tree of Fig. 4(A), M. blythii and M. myotisappear to be closely related to each other (mean haplo-typic divergence 2.1 ± 0.3%), but both differ considerablyfrom the North African form (mean 11.8% and 10.9%,respectively). In a previous review of ecological, morpho-logical and allozymic variation among M. myotis andM. blythii, Arlettaz et al. (1997b) analysed basically thesame populations as those presented in this paper. Theseauthors showed that the morphologically intermediatebats from North Africa, Corsica and Sardinia representeda homogeneous, monotypic assemblage (Figs 1 and 2 inArlettaz et al. 1997b) and have the same allozyme allelesas M. myotis at two discriminant loci (Arlettaz 1995; Arlettazet al. 1997b). Accordingly, a reanalysis of this data set using35 allozyme loci and M. nattereri as an outgroup, suggests acloser phylogenetic relationship between the Africantaxon and M. myotis rather than with M. blythii (Fig. 4B).However, these results depend on few variable allozymeloci (11 out of 35 essayed), most of which are di- or triallelicwithin Mouse-eared bats. For instance, the ADA andGOT-1 loci are fixed for alternative alleles in continentalM. myotis and M. blythii (Arlettaz 1995; Arlettaz et al. 1997b),and were used to identify bats from Morocco, Sardiniaand Corsica. Yet, the same alleles are found in otherspecies (e.g. in M. nattereri or M. daubentonii), suggestingthat retention of plesiomorphic alleles, homoplasy, paral-lelism or undetected mutations could be common in theseallozyme loci. According to the highest bootstrap valuesobtained for the mtDNA tree (Fig. 4A) and to the largernumber of individuals and informative loci used for themicrosatellite data set, we are more confident in thephylogenetic relationships obtained in the presentmolecular data.
Thus, our most likely interpretation of the availabledata is that the Moroccan and Iberian bats collected alongthe transect represent distinct biological species, whichdo not interbreed because occasional migrants acrossthe strait, if any, do not seem to transmit their genes to theresident populations. Unlike M. myotis and M. blythii, whichare found in strict sympatry over most of their Europeanrange, the North African form does not apparently coexistwith either species, at least in Morocco, Sardinia andCorsica. Interestingly, the niche of the latter is very similarto that of European continental M. myotis as they bothfeed essentially on ground-dwelling arthropods (carabidbeetles, ground crickets, scorpions, etc.) (Arlettaz et al. 1997a;Arlettaz 1999). Competitive exclusion would, therefore,be more likely between M. myotis and bats of the NorthAfrican clade. In contrast, M. blythii exploits a quitedistinct niche throughout its range, it’s diet consisting
mostly of grass-dwelling prey, namely bush crickets.Whether the current distribution of these three siblingspecies is due entirely to historical processes of coloniza-tion, to competitive exclusion, or a combination of bothremains open to debate. If one applies the ‘standard’ rateof 2% per million years for cytochrome b divergence inmammals (Johns & Avise 1998), the difference observedbetween both European Mouse-eared bats and the NorthAfrican taxon (about 11%) would support a divergencedating back to the Pliocene epoch. This very ancientdate does not mean that these taxa face each other atGibraltar since several million years. Rather, they prob-ably diverged from a common ancestor somewhere else,while the descendents colonized the present distributionarea more recently.
As the North African taxon is morphologically inter-mediate in size between M. myotis and M. blythii (Feltenet al. 1977; Benda & Horacek 1995), there has been consider-able debate concerning its systematic position: it was eithertreated as a smaller form of M. myotis (e.g. Ellerman &Morrison-Scott 1966; Benda & Horacek 1995), or as alarger form of M. blythii (Felten et al. 1977; Bogan et al.1978; Corbet 1978). Based on samples from the Maghreb,Sardinia, Corsica and Malta, Felten (in Felten et al. 1977)described them under the new name M. blythii punicusFelten 1977. Thus, we provisionally refer to the NorthAfrican, Corsican, and Sardinian ‘intermediate’ Mouse-eared bats as Myotis cf punicus Felten 1977. Of course, thedefinitive systematic position and distribution of thisinteresting complex of species awaits further morpholo-gical and molecular comparisons especially with batsfrom the eastern Mediterranean area and from elsewherein the Middle east.
In conclusion, even if the narrow Gibraltar Strait doesn’tseem to represent an insuperable obstacle for Mouse-earedbats (they can cross far broader stretches of open sea), itmay still prevent gene flow. For these bats, it materializesthe borders of distribution of sibling species, with two ofthem (M. myotis and M. blythii) apparently not coexistingwith the third one (M. cf punicus). It remains to be shownwhether other species of bats actually exchange migrantsacross Gibraltar, or whether they colonized southern Spainand northern Morocco via other routes like the majorityof other wild mammals (Dobson 1998).
Acknowledgements
We thank Javier Juste, Juan Quetglas, Elena Migens, AssociationSportive de Speleologie de Agadir, Asociación Roncadell, Juan R.Boyero, Pepe Ganfornina and many other bat workers for assist-ance in the field. Authorizations to take biopsy samples in Spainwere provided by the Consejeria de Medio Ambiente de la Juntade Andalucia and Consellería de Agricultura y Medio Ambientede la Generalitat Valenciana. Nelly diMarco provided help withDNA extraction. We thank also Eric Petit and François Balloux
MEC1069.fm Page 1769 Tuesday, October 10, 2000 10:18 AM

1770 V. C A S T E L L A E T A L .
© 2000 Blackwell Science Ltd, Molecular Ecology, 9, 1761–1772
for helpful comments on early versions of the manuscript. Finan-cial support was provided by the following institutions: SwissNational Funds for Scientific Research (grant # 3100–04 9245.96to MR), DGICYT (project PB 90–0143 to CI), Fondation du 450e
anniversaire (University of Lausanne), Société Académique Suisse,Société Suisse de Zoologie, Natural History Museum of Geneva,and Institut Menorquí d’Estudis.
References
Arlettaz R (1995) Ecology of the Sibling Mouse-Eared Bats (MyotisMyotis and Myotis Blythii): Zoogeography, Niche, Competition, andForaging. Horus Publishers, Martigny, Switzerland.
Arlettaz R (1996) Feeding behaviour and foraging strategy offree-living Mouse-eared bats, Myotis myotis and Myotis blythii.Animal Behaviour, 51, 1–11.
Arlettaz R (1999) Habitat selection as a major resource partition-ing mechanism between the two sympatric sibling bat speciesMyotis myotis and Myotis blythii. Journal of Animal Ecology, 68,460–471.
Arlettaz R, Perrin N, Hausser J (1997a) Trophic resource parti-tioning and competition between the two sibling bat speciesMyotis myotis and Myotis blythii. Journal of Animal Ecology, 66,897–911.
Arlettaz R, Ruedi M, Ibañes C, Palmeirim J, Hausser J (1997b) Anew perspective on the zoogeography of the sibling mouse-eared bat species Myotis myotis and M. blythii: morphological,genetical and ecological evidence. Journal of Zoology (London),242, 45–62.
Avise J, Walker D (1999) Species realities and numbers in sexualvertebrates: Perspectives from an asexually transmitted genome.Proceedings of the National Academy of Sciences of the USA, 96,992–995.
Avise JC (1994) Molecular markers, natural history and evolution.Chapman & Hall, New York.
Barratt EM, Deaville R, Burland TM et al. (1997) DNA answersthe call of pipistrelle bat species. Nature, 387, 138–139.
Benda P, Horacek I (1995) Biometrics of Myotis myotis and Myotisblythi. Myotis, 32-33, 45–55.
Bogan MA, Setzer HW, Findley JS, Wilson DE (1978) Phenetics ofMyotis blythi in Morocco. In: Proceedings of the Fourth InternationalBat Research Conference, Nairobi, pp. 217–230.
Boursot P, Jacquart T, Bonhomme F, Britton-Davidian J, Thaler L(1985) Différenciation géographique du génôme mitochondrialchez Mus spretus Lataste. Comptes Rendus de l’Académie desSciences de Paris, 301, 161–166.
Burban C, Petit RJ, Carcreff E, Jactel H (1999) Rangewide vari-ation of the maritime pine bast scale Matsucoccus feytaudi Duc.(Homoptera: Matsucoccidae) in relation to the genetic structureof its horst. Molecular Ecology, 8, 1593–1602.
Castella V, Ruedi M (2000) Characterisation of highly variablemicrosatellite loci in the bat Myotis myotis (Chiroptera: Vesper-tilionidae). Molecular Ecology, 9, 1000–1002.
Corbet GB (1978) The Mammals of the Palaearctic Region: a taxo-nomic review. Cornell University Press, London.
Dobson M (1998) Mammal distributions in the western Mediter-ranean: the role of human intervention. Mammal Review, 28,77–88.
Ellerman JR, Morrison-Scott TCS (1966) Checklist of Palaearctic andIndian mammals, 1758–1946. Alden Press, Oxford.
Excoffier L, Smouse PE, Quattro JM (1992) Analysis of molecular
variance inferred from metric distances among DNA haplotypes:application to human mitochondrial DNA restriction data.Genetics, 131, 479–491.
Felsenstein J (1993) PHYLIP (Phylogeny Inference Package). Univer-sity of Washington, Seattle, WA.
Felten H, Spitzenberger F, Storch G (1977) Zur KleinsäugerfaunaWest-Anatoliens. Teil IIIa. Senckenbergiana Biologica, 58, 1–44.
Filippucci MG (1992) Allozyme variation and divergence amongEuropean, Middle Eastern, and North African species of thegenus Apodemus (Rodentia, Muridae). Israel Journal of Zoology,38, 193–218.
Goudet J (1999a) FSTAT 2.9, a program to estimate and test genediversities and fixation indices (updated from Goudet 1995).Lausanne, Switzerland. http://www.unil.ch/izea/softwares/fstat.html.
Goudet J (1999b) PCA-GEN, Version 1.2. Lausanne, Switzerland.http://www.unil.ch/izea/softwares/pcagen.html.
Hewitt GM (1999) Post-glacial re-colonization of European biota.Biological Journal of the Linnean Society, 68, 87–112.
Horacek I (1985) Population ecology of Myotis myotis in CentralBohemia (Mammalia: Chiroptera). Acta Universitas Carolinae —Biologica, 8, 161–267.
Johns GC, Avise JC (1998) A comparative summary of geneticdistances in the vertebrates from the mitochondrial Cytochromeb gene. Molecular Biology and Evolution, 15, 1481–1490.
Kocher TD, Thomas WK, Meyer A et al. (1989) Dynamics ofmitochondrial DNA evolution in animals: amplification andsequencing with conserved primers. Proceedings of the NationalAcademy of Sciences of the USA, 86, 6196–6200.
Lugon-Moulin N, Brünner H, Wyttenbach A, Hausser J, Goudet J(1999) Hierarchical analyses of genetic differentiation in a hybridzone of Sorex araneus (Insectivora: Soricidae). Molecular Ecology,8, 419–431.
Miller SA, Dykes DD, Polesky HF (1988) A simple salting proce-dure for extracting DNA from human nucleated cells. NucleicAcids Research, 16, 215.
Nei M (1987) Molecular Evolutionary Genetics. Columbia UniversityPress, New York, USA.
Paetkau D, Calvert W, Stirling I, Strobeck C (1995) Microsatelliteanalysis of population structure in Canadian polar bears.Molecular Ecology, 4, 347–354.
Paetkau D, Waits L, Clarkson P, Craighead L, Strobeck C (1997)An empirical evaluation of genetic distance statistics usingmicrosatellite data from bear (Ursidae) populations. Genetics,147, 1943–1957.
Palmeirim J (1979) First record of Myotis myotis on the AzoresIslands (Chiroptera: Vespertilionidae). Arquivos do Museu Bocage,notas e suplementos, 46, 1–2.
Paz Od, Fernandez R, Benzal J (1986) El anillamiento de Quiro-pteros en el centro de la peninsula iberica durante el periodo1977-86. Boletin de la Estacion Central de Ecologia, 30, 113–138.
Pedrola-Monfort J, Caujapé-Castells J (1994) Allozymic andmorphological relationships among Androcymbium gramineum,A. europaeum, and A. psammophilum (Colchicaceae). PlantSystematics and Evolution, 191, 111–126.
Petit E, Mayer F (1999) Male dispersal in the noctule bat (Nyctalusnoctula): where are the limits? Proceedings of the Royal Society ofLondon, Series B, 266, 1717–1722.
Petri B, Pääbo S, Von Haeseler A, Tautz D (1997) Paternity assess-ment and population subdivision in a natural population ofthe Larger Mouse-eared bat Myotis myotis. Molecular Ecology, 6,235–242.
MEC1069.fm Page 1770 Tuesday, October 10, 2000 10:18 AM

I S G I B R A LTA R A B A R R I E R TO G E N E F L O W F O R B AT S ? 1771
© 2000 Blackwell Science Ltd, Molecular Ecology, 9, 1761–1772
Raymond M, Rousset F (1995) An exact test for populationdifferentiation. Evolution, 49, 1280–1283.
Rice WR (1989) Analysis tables of statistical tests. Evolution, 43,223–225.
Ruedi M, Arlettaz R, Maddalena T (1990) Distinction morphologiqueet biochimique de deux espèces jumelles de chauves-souris:Myotis myotis (Bork.) et Myotis blythi (Tomes) (Mammalia;Vespertilionidae). Mammalia, 54, 415–429.
Schneider S, Kueffer J-M, Roessli D, Excoffier L (1996) Arlequin: aSoftware Package for Population Genetics. Genetics and BiometryLab, Dept. of Anthropology, University of Geneva, Geneva.
Sevilla P (1989) Quaternary fauna of bats in Spain: Paleoecologicand biogeographic interest. In: European Bat Research 1987 (edsHanak V, Horacek I, Gaisler J), pp. 349–355. Charles UniversityPress, Praha, Tchechia.
Slatkin M (1994) Gene flow and population structure. In: Ecologi-cal Genetics (ed. Real LA), pp. 3–17. Princeton Universty Press,Princeton.
Smith MF, Patton JL (1993) The diversification of South Americanmurid rodents: evidence from mitochondrial DNA sequencedata for the akodontine tribe. Biological Journal of the LinneanSociety, 50, 149–177.
Steininger FF, Rabeder G, Rögl F (1985) Land mammal distributionin the Mediterranean Neogene: A consequence of geokinematicand climatic events. In: Geological Evolution of the MediterraneanBasin (eds Stanley DJ, Wezel F-C), pp. 559–571. Springer-Verlag,New York.
Swofford DL (1996) PAUP*: Phylogenetic analysis using parsimony(* And Other Methods), version 4.0. Sinauer Associates, Sunderland,Masachusetts.
Taberlet P, Fumagalli LW, ¸st-Saucy A-G, Cosson J-F (1998) Com-parative phylogeography and postglacial colonization routesin Europe. Molecular Ecology, 7, 453–464.
Tautz D, Renz M (1984) Simple sequences are ubiquitous repeti-tive components of eukaryotic genomes. Nucleic Acids Research,12, 4127–4138.
Valdés B (1991) Andalucia and the Rif. Floristic links and a com-mon flora. Botanika Chronika, 10, 117–124.
Weir BS, Cockerham CC (1984) Estimating F-statistics for theanalysis of population structure. Evolution, 38, 1358–1370.
Worthington Wilmer J, Barratt E (1996) A non-lethal method oftissue sampling for genetic studies of Chiropterans. Bat ResearchNews, 37, 1–3.
Wright S (1978) Variability within and among natural populations.Evolution and the genetics of populations. University of ChicagoPress, Chicago.
VC’s PhD project focuses on phylogeography and populationgenetics of the greater Mouse-eared bat. MR is conducting variousstudies on the molecular systematics and phylogeny of bats. LEis a human population geneticist with interest in inferring popu-lation history and demography from molecular markers in variousspecies. CI is interested in the phylogeography of bats from Iberia,Maghreb and Macaronesia. RA works on community and popu-lation ecology, behavioural ecology, evolutionary ecology, andconservation biology of bats and birds. JH’s prime researchinterests are population genetics and the evolutionary history ofsoricine shrews.
MEC1069.fm Page 1771 Tuesday, October 10, 2000 10:18 AM

1772 V. C A S T E L L A E T A L .
© 2000 Blackwell Science Ltd, Molecular Ecology, 9, 1761–1772
Appendix
List of material and locations used in the genetic analyses. The number of females (F) and males (M) genotyped at all microsatellite lociand the number of the sequenced haplotypes (see Fig. 4A) appear in parentheses
Myotis myotis — SPAIN: Cueva del Agua, Cadiz (15 F and 5 m; 9 × ES1, 1 × ES2); Canillas de Aceituno, Malaga (19 F and 1 m; 2 × ES1); Denia, Alicante (12 F and 8 m; 2 × ES4); Inca, Mallorca (2 F and 1 m; 1 × ES1, 2 × ES5). SWITZERLAND: Perreux, Neuchâtel (1 F and 1 m; 2 × CH1).
Myotis blythii — SPAIN: Tarifa, Cadiz (2 F; 2 × ES1b); Cueva del Agua, Cadiz (3 F; 1 × ES1b); Canillas, Malaga (1 F and 1 m); Denia, Alicante (1 m; 1 × ES1b); Castellon (1 m and 1 F); Malgrat, Barcelona (1 F). PORTUGAL: Loucal, Querença (1 indet.). FRANCE: Montredon, Narbonne (1 F and 1 m); Nice (2 F and 2 m). CZECH REPUBLIC: Bohemia (1 indet.; 1 × TS1). GREECE: Macedonia (1 indet.; 1 × GR1). KIRGHIZSTAN: Adzidar cave, Os (1 F and 1 m; 1 × KZ1, 1 × KZ2). Note that the latter four bats were not included in the microsatellite analyses.
Myotis cf punicus — MOROCCO: Azrou, Meknes (16 F and 4 m; 1 × MA1, 1 × MA2); Boumahden, Agadir (5 F and 15 m; 1 × MA3, 1 × MA4). SPAIN: Mina del Agua, Ceuta (6 F and 13 m; 10 × MA1). FRANCE: Oleta, Corsica (2 F; 1 × CO1, 1 × CO2). ITALY: Mt Maiore, Ulassai, Sardinia (1 F and 2 m; 2 × SA1).
Myotis nattereri — GREECE: Macedonia (not sexed).
MEC1069.fm Page 1772 Tuesday, October 10, 2000 10:18 AM




















