
Activity and shelter selection by Myotis myotis and Rhinolophus hipposideros hibernating in the...
11
www.elsevier.de/mambio Original investigation Activity and shelter selection by Myotis myotis and Rhinolophus hipposideros hibernating in the Kater ˇinska ´ cave (Czech Republic) By J. Zukal, Hana Berkova ´ and Z. R ˘ eha ´k Institute of Vertebrate Biology, Academy of Sciences of the Czech Republic, Brno und Department of Zoology and Ecology, Faculty of Science, Masaryk University, Brno, Czech Republic Receipt of Ms. 21.4.2004 Acceptance of Ms. 9.3.2005 Abstract In 1992–1993, the bat species Myotis myotis and Rhinolophus hipposideros hibernating in the Kater ˇinska ´ cave were investigated by means of regular censuses without any handling and marking of the animals. Three basic parameters of their shelters were recorded (position in cave, type and relative height). In total, during 26 checks we registered 1141 findings of nine bat species. Movement activity, expressed as percentage of new findings during a particular visit, was registered during the whole winter season. Its level fluctuated in different ways and the hibernation period of R. hipposideros could be divided into three different parts, while the level of M. myotis movement activity was relatively high during all season. The shelter selection of R. hipposideros was not dependent on the part of cave where the bats were hibernating, and it did not change during the season. Hibernating specimens of R. hipposideros most frequently used exposed places, in which they were always hanging free. Myotis myotis was registered in all types of shelter with one exception. Rhinolophus hipposideros used mainly the middle part of the cave at a distance between 121 and 180m from the entrance. The most preferred part of the cave by M. myotis was a small segment of Corridor (between 21 and 30m), i.e., the entrance part of the cave. Rhinolophus hipposideros is a highly specialized species which prefers parts of the cave with very stable microclimatic conditions and, on the contrary, M. myotis appears to be indifferent to all parameters studied, and it uses the shelters indiscriminately. r 2005 Deutsche Gesellschaft fu ¨r Sa ¨ugetierkunde. Published by Elsevier GmbH. All rights reserved. Key words: Bats, shelter selection, activity, hibernation Introduction With its more than 1100 caves, the Moravian Karst is among the most important regions for the hibernation of bats in Europe. Even when the first data on bats originate from the half of the 19th century, systematic research was initiated there by Gaisler as late as 1957. Some caves in the southern part of the Moravian Karst are hibernaculas which have been monitored for the longest time in central Europe (R ˘ eha´k 1997). Attention has mainly been given to the monitoring of long-term changes in bat communities (Zima et al. ARTICLE IN PRESS 1616-5047/$ - see front matter r 2005 Deutsche Gesellschaft fu ¨r Sa ¨ugetierkunde. Published by Elsevier GmbH. All rights reserved. doi:10.1016/j.mambio.2005.03.003 Mamm. biol. 70 (2005) 5 271–281
Transcript of Activity and shelter selection by Myotis myotis and Rhinolophus hipposideros hibernating in the...

ARTICLE IN PRESS
www.elsevier.de/mambio
1616-5047/$ - see
doi:10.1016/j.mam
Original investigation
Activity and shelter selection by Myotis myotis andRhinolophus hipposideros hibernating in the Katerinskacave (Czech Republic)
By J. Zukal, Hana Berkova and Z. Rehak
Institute of Vertebrate Biology, Academy of Sciences of the Czech Republic, Brno und Department of Zoology andEcology, Faculty of Science, Masaryk University, Brno, Czech Republic
Receipt of Ms. 21.4.2004Acceptance of Ms. 9.3.2005
Abstract
In 1992–1993, the bat species Myotis myotis and Rhinolophus hipposideros hibernating in theKaterinska cave were investigated by means of regular censuses without any handling and markingof the animals. Three basic parameters of their shelters were recorded (position in cave, type andrelative height). In total, during 26 checks we registered 1141 findings of nine bat species.Movement activity, expressed as percentage of new findings during a particular visit, was registeredduring the whole winter season. Its level fluctuated in different ways and the hibernation period ofR. hipposideros could be divided into three different parts, while the level of M. myotis movementactivity was relatively high during all season. The shelter selection of R. hipposideros was notdependent on the part of cave where the bats were hibernating, and it did not change during theseason. Hibernating specimens of R. hipposideros most frequently used exposed places, in whichthey were always hanging free. Myotis myotis was registered in all types of shelter with oneexception. Rhinolophus hipposideros used mainly the middle part of the cave at a distance between121 and 180 m from the entrance. The most preferred part of the cave by M. myotis was a smallsegment of Corridor (between 21 and 30 m), i.e., the entrance part of the cave. Rhinolophushipposideros is a highly specialized species which prefers parts of the cave with very stablemicroclimatic conditions and, on the contrary, M. myotis appears to be indifferent to all parametersstudied, and it uses the shelters indiscriminately.r 2005 Deutsche Gesellschaft fur Saugetierkunde. Published by Elsevier GmbH. All rights reserved.
Key words: Bats, shelter selection, activity, hibernation
Introduction
With its more than 1100 caves, the MoravianKarst is among the most important regionsfor the hibernation of bats in Europe. Evenwhen the first data on bats originate from thehalf of the 19th century, systematic researchwas initiated there by Gaisler as late as 1957.
front matter r 2005 Deutsche Gesellschaft fur S
bio.2005.03.003
Some caves in the southern part of theMoravian Karst are hibernaculas which havebeen monitored for the longest time in centralEurope (Rehak 1997). Attention has mainlybeen given to the monitoring of long-termchanges in bat communities (Zima et al.
augetierkunde. Published by Elsevier GmbH. All rights reserved.
Mamm. biol. 70 (2005) 5 � 271–281

ARTICLE IN PRESS
t6
Dome of Chaos
Dome of the Witch
Not checked spaces
t7
t8
272 J. Zukal et al.
1994). Studies of bat ecology during hiberna-tion are very rare and are mostly concernedwith the thermopreferendum of bats (Gaisler1970; Bauerova and Zima 1988).The aim of the present study is to evaluatethe selection of different places with specificparameters by two most abundant bat speciesin a natural cave and the level of theirmovement activity. We tested the hypothesisthat shelter selection by bats depends on theinternal characteristics of cave and, therefore,it differs in the particular parts of the caveand changes in the course of a year.
0 50 m
N
Path
Entrance
Corridor
Main Dome
t1
t2
t3
t4
t5
Fig. 1. Division of the Katerinska Cave into four basicparts. Explanations: t1..8–points of air temperaturemeasurement.
Material and methods
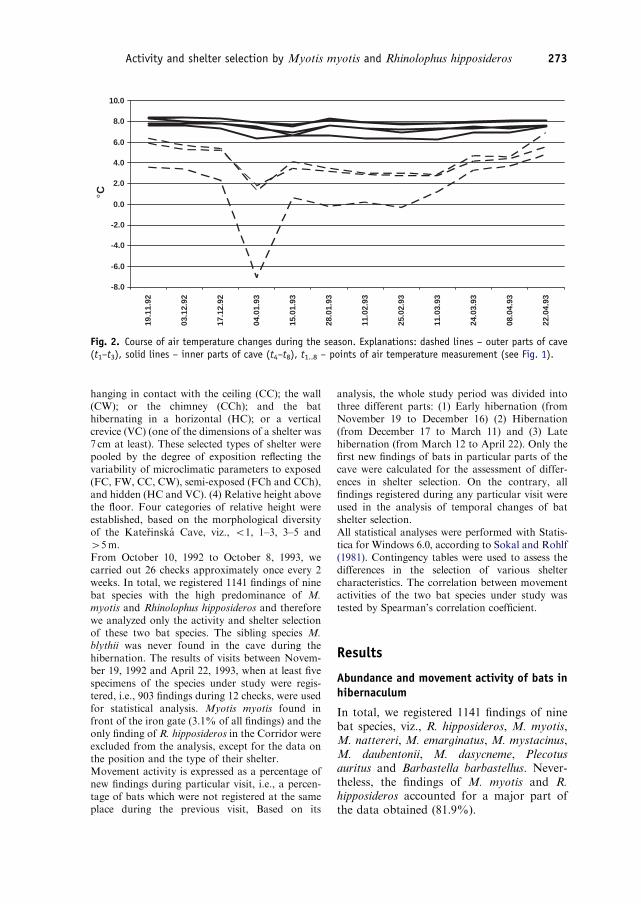
Research was carried out in a natural limestonecave (Katerinska Cave) situated in the eastern partof the Czech Republic. The total length of the caveis about 300m and it has only one entrance whichis closed by an iron gate with a vertical hole in itsupper part (Rehak et al. 1994). The cave can bedivided into a dynamic and a static part by thedifferences in temperature and airflow. The ap-proximately 50m long outer part (Corridor) isdynamic from the microclimatic point of view, asthe air temperature in this part of the cave isinfluenced by the changes in ambient temperature(Figs. 1 and 2). The main inner part of the caveconsists of three large domes (Main dome, Domeof the Witch and Dome of Chaos) which areconsidered as microclimatically stable parts. Theyare characterized by relatively stable temperatures,ranging between 6.3 and 8.8 1C in the course of theyear (Fig. 2). Two small and inaccessible parts werenot checked regularly as there were no hibernatingbats, and thus these parts were excluded from themonitoring (Fig. 1). Except for 2 months (Decem-ber and January) when only the employees of theAgency for Nature Conservation and LandscapeProtection of the Czech Republic were permitted toenter the cave, tourists visited the cave.We used the visual census method without anyhandling and marking of animals (Zima et al.1994), as one of the main requirements of ourresearch was to avoid any disturbance of thehibernating bats. Air temperature was measuredby the digital thermometer DT-20 during thecensus of hibernating bats and it was left at least5min at each point. We registered the followingcharacters for each bat or their cluster (i.e., groupof two or more specimens in close contact): (1)Species, number of specimens and their clusters.Small species of Myotis spp. were recorded as M.sp. if we were not able to determine them exactly.
(2) Position of a shelter in the cave, with accuratelyplotting it in a map. The exact position of ahibernating bat with information whether it was anew finding or the same specimen and/or clusterwas registered again during the subsequent visit.The relative distance of the bat shelter from thecave entrance (the iron gate with an exit hole) wassubsequently ascertained from the cave map. Forthis purpose, the position of the iron gate wasconsidered to be the center point from whichconcentric circles were drawn at distances of 10m.(3) Type of shelter. Eight types of shelters weredistinguished, viz., the bat hanging free on theceiling (i.e., a slope o451) (FC); on the wall (i.e., aslope 4451) (FW); in the chimney (FCh); the bat
ARTICLE IN PRESS
-8.0
-6.0
-4.0
-2.0
0.0
2.0
4.0
6.0
8.0
10.0
19.1
1.92
03.1
2.92
17.1
2.92
04.0
1.93
15.0
1.93
28.0
1.93
11.0
2.93
25.0
2.93
11.0
3.93
24.0
3.93
08.0
4.93
22.0
4.93
°C
Fig. 2. Course of air temperature changes during the season. Explanations: dashed lines – outer parts of cave(t1–t3), solid lines – inner parts of cave (t4–t8), t1..8 – points of air temperature measurement (see Fig. 1).
Activity and shelter selection by Myotis myotis and Rhinolophus hipposideros 273
hanging in contact with the ceiling (CC); the wall(CW); or the chimney (CCh); and the bathibernating in a horizontal (HC); or a verticalcrevice (VC) (one of the dimensions of a shelter was7 cm at least). These selected types of shelter werepooled by the degree of exposition reflecting thevariability of microclimatic parameters to exposed(FC, FW, CC, CW), semi-exposed (FCh and CCh),and hidden (HC and VC). (4) Relative height abovethe floor. Four categories of relative height wereestablished, based on the morphological diversityof the Katerinska Cave, viz., o1, 1–3, 3–5 and45m.From October 10, 1992 to October 8, 1993, wecarried out 26 checks approximately once every 2weeks. In total, we registered 1141 findings of ninebat species with the high predominance of M.
myotis and Rhinolophus hipposideros and thereforewe analyzed only the activity and shelter selectionof these two bat species. The sibling species M.
blythii was never found in the cave during thehibernation. The results of visits between Novem-ber 19, 1992 and April 22, 1993, when at least fivespecimens of the species under study were regis-tered, i.e., 903 findings during 12 checks, were usedfor statistical analysis. Myotis myotis found infront of the iron gate (3.1% of all findings) and theonly finding of R. hipposideros in the Corridor wereexcluded from the analysis, except for the data onthe position and the type of their shelter.Movement activity is expressed as a percentage ofnew findings during particular visit, i.e., a percen-tage of bats which were not registered at the sameplace during the previous visit, Based on its
analysis, the whole study period was divided intothree different parts: (1) Early hibernation (fromNovember 19 to December 16) (2) Hibernation(from December 17 to March 11) and (3) Latehibernation (from March 12 to April 22). Only thefirst new findings of bats in particular parts of thecave were calculated for the assessment of differ-ences in shelter selection. On the contrary, allfindings registered during any particular visit wereused in the analysis of temporal changes of batshelter selection.All statistical analyses were performed with Statis-tica for Windows 6.0, according to Sokal and Rohlf(1981). Contingency tables were used to assess thedifferences in the selection of various sheltercharacteristics. The correlation between movementactivities of the two bat species under study wastested by Spearman’s correlation coefficient.
Results
Abundance and movement activity of bats inhibernaculum
In total, we registered 1141 findings of ninebat species, viz., R. hipposideros, M. myotis,M. nattereri, M. emarginatus, M. mystacinus,M. daubentonii, M. dasycneme, Plecotusauritus and Barbastella barbastellus. Never-theless, the findings of M. myotis and R.hipposideros accounted for a major part ofthe data obtained (81.9%).

ARTICLE IN PRESS
Fig. 3. Changes in the level of movement activity. Explanations: dotted line – the dividing of separate parts ofseason.
274 J. Zukal et al.
On the basis of the activity level the hiberna-tion period of R. hipposideros can be dividedinto three different parts (Fig. 3). During thefirst one (‘‘early hibernation’’) the number ofbats considerably increased and this increasewas correlated with the high level of move-ment activity. The number of hibernating R.hipposideros was subsequently stabilized (ca.20 specimens) and only occasional move-ments were registered (‘‘hibernation’’). Sincethe middle of March the abundance hasdropped and the bats continuously left thehibernaculum (‘‘late hibernation’’). This de-crease in the abundance of bats correlatedagain with the increase in their movementactivity. There was a significant difference inthe level of movement activity in the succes-sive periods (w2, DF=2, Po0:001).The level of M. myotis movement activity wasrelatively high during all season and the‘‘deep hibernation’’ period with decreasedactivity could not be clearly separated. Incontrast to R. hipposideros, the total numberof bats of this species was increasing gradu-ally since October. The peak was recorded atthe beginning of April (112 individuals). Thehighest increase in abundance was registeredin the clusters formed in Corridor. Unfortu-
nately, we could not assess the exactnumber of new bats in clusters by thevisual method of census without disturbingthe bats but only the change in the totalnumber of hibernating bats, i.e., the differ-ence between the previous and presentnumbers. Nevertheless, the movement activ-ity of M. myotis and R. hipposideros wassignificantly correlated (rS ¼ 0:60, P ¼ 0:040,N ¼ 12), therefore the hibernation periodwas divided in the same way for both species(Fig. 3).
Shelter selection and its changes during theseason
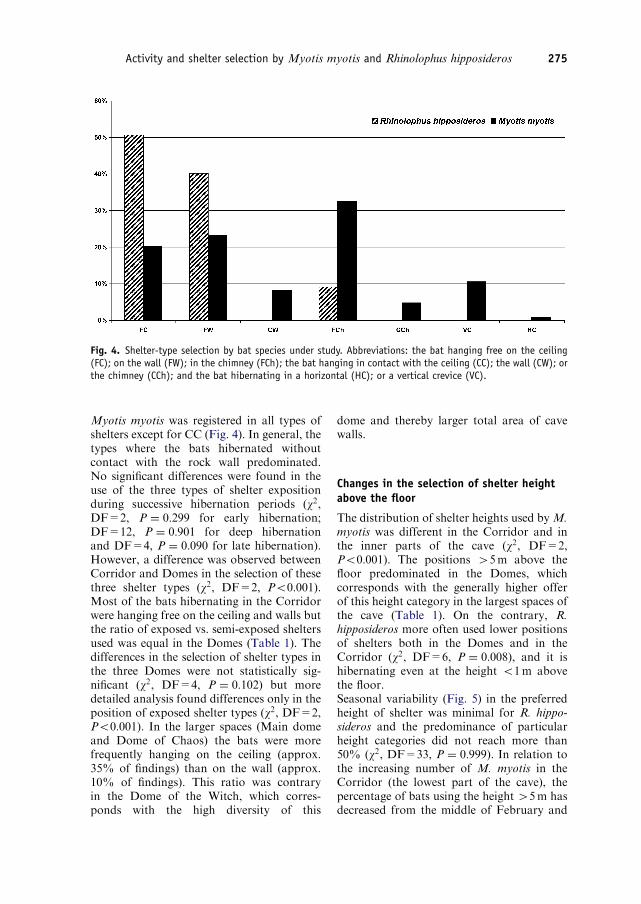
The shelter selection of R. hipposideros wasnot dependent on the part of the cave wherethe bats were hibernating (w2, DF=4,P ¼ 0:609) and it did not change during theseason (w2, DF=22, P ¼ 0:642). HibernatingR. hipposideros most frequently used ex-posed places (FC, FW), where they werealways hanging free (Fig. 4). The onlyindividual R. hipposideros found in theCorridor (22.4.1993) was also hanging freeon the wall of the cave at the height of 3–5mabove the floor.
ARTICLE IN PRESS
Fig. 4. Shelter-type selection by bat species under study. Abbreviations: the bat hanging free on the ceiling(FC); on the wall (FW); in the chimney (FCh); the bat hanging in contact with the ceiling (CC); the wall (CW); orthe chimney (CCh); and the bat hibernating in a horizontal (HC); or a vertical crevice (VC).
Activity and shelter selection by Myotis myotis and Rhinolophus hipposideros 275
Myotis myotis was registered in all types ofshelters except for CC (Fig. 4). In general, thetypes where the bats hibernated withoutcontact with the rock wall predominated.No significant differences were found in theuse of the three types of shelter expositionduring successive hibernation periods (w2,DF=2, P ¼ 0:299 for early hibernation;DF=12, P ¼ 0:901 for deep hibernationand DF=4, P ¼ 0:090 for late hibernation).However, a difference was observed betweenCorridor and Domes in the selection of thesethree shelter types (w2, DF=2, Po0:001).Most of the bats hibernating in the Corridorwere hanging free on the ceiling and walls butthe ratio of exposed vs. semi-exposed sheltersused was equal in the Domes (Table 1). Thedifferences in the selection of shelter types inthe three Domes were not statistically sig-nificant (w2, DF=4, P ¼ 0:102) but moredetailed analysis found differences only in theposition of exposed shelter types (w2, DF=2,Po0:001). In the larger spaces (Main domeand Dome of Chaos) the bats were morefrequently hanging on the ceiling (approx.35% of findings) than on the wall (approx.10% of findings). This ratio was contraryin the Dome of the Witch, which corres-ponds with the high diversity of this
dome and thereby larger total area of cavewalls.
Changes in the selection of shelter heightabove the floor
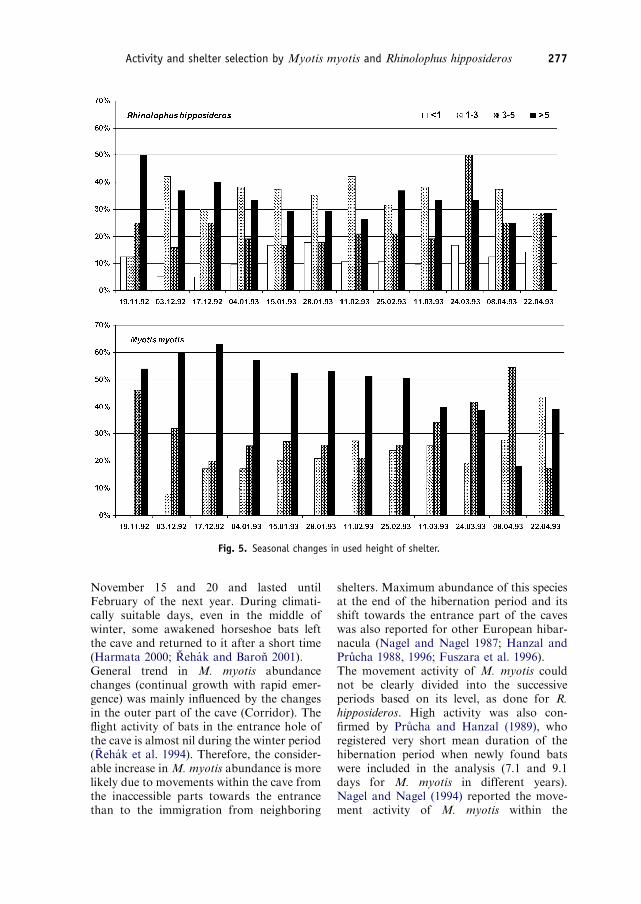
The distribution of shelter heights used by M.myotis was different in the Corridor and inthe inner parts of the cave (w2, DF=2,Po0:001). The positions 45m above thefloor predominated in the Domes, whichcorresponds with the generally higher offerof this height category in the largest spaces ofthe cave (Table 1). On the contrary, R.hipposideros more often used lower positionsof shelters both in the Domes and in theCorridor (w2, DF=6, P ¼ 0:008), and it ishibernating even at the height o1m abovethe floor.Seasonal variability (Fig. 5) in the preferredheight of shelter was minimal for R. hippo-sideros and the predominance of particularheight categories did not reach more than50% (w2, DF=33, P ¼ 0:999). In relation tothe increasing number of M. myotis in theCorridor (the lowest part of the cave), thepercentage of bats using the height 45m hasdecreased from the middle of February and
ARTICLE IN PRESS
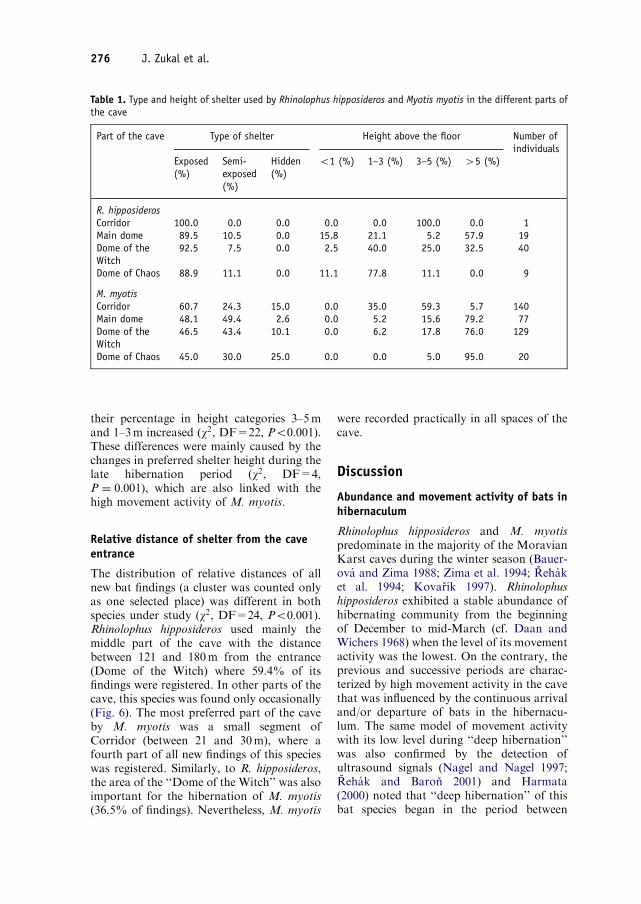
Table 1. Type and height of shelter used by Rhinolophus hipposideros and Myotis myotis in the different parts ofthe cave
Part of the cave Type of shelter Height above the floor Number ofindividuals
Exposed(%)
Semi-exposed(%)
Hidden(%)
o1 (%) 1–3 (%) 3–5 (%) 45 (%)
R. hipposiderosCorridor 100.0 0.0 0.0 0.0 0.0 100.0 0.0 1Main dome 89.5 10.5 0.0 15.8 21.1 5.2 57.9 19Dome of theWitch
92.5 7.5 0.0 2.5 40.0 25.0 32.5 40
Dome of Chaos 88.9 11.1 0.0 11.1 77.8 11.1 0.0 9
M. myotisCorridor 60.7 24.3 15.0 0.0 35.0 59.3 5.7 140Main dome 48.1 49.4 2.6 0.0 5.2 15.6 79.2 77Dome of theWitch
46.5 43.4 10.1 0.0 6.2 17.8 76.0 129
Dome of Chaos 45.0 30.0 25.0 0.0 0.0 5.0 95.0 20
276 J. Zukal et al.
their percentage in height categories 3–5mand 1–3m increased (w2, DF=22, Po0:001).These differences were mainly caused by thechanges in preferred shelter height during thelate hibernation period (w2, DF=4,P ¼ 0:001), which are also linked with thehigh movement activity of M. myotis.
Relative distance of shelter from the caveentrance
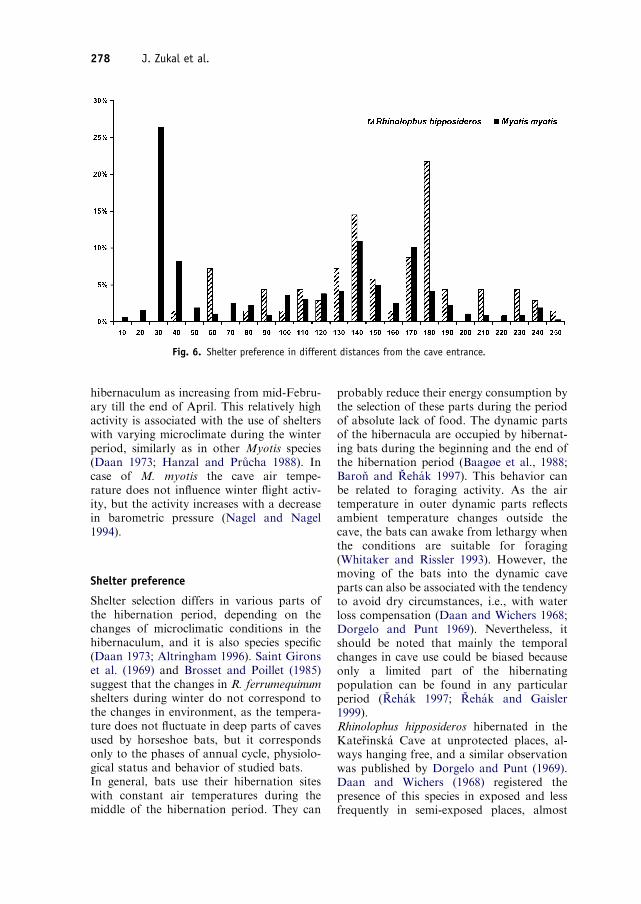
The distribution of relative distances of allnew bat findings (a cluster was counted onlyas one selected place) was different in bothspecies under study (w2, DF=24, Po0:001).Rhinolophus hipposideros used mainly themiddle part of the cave with the distancebetween 121 and 180m from the entrance(Dome of the Witch) where 59.4% of itsfindings were registered. In other parts of thecave, this species was found only occasionally(Fig. 6). The most preferred part of the caveby M. myotis was a small segment ofCorridor (between 21 and 30m), where afourth part of all new findings of this specieswas registered. Similarly, to R. hipposideros,the area of the ‘‘Dome of the Witch’’ was alsoimportant for the hibernation of M. myotis(36.5% of findings). Nevertheless, M. myotis
were recorded practically in all spaces of thecave.
Discussion
Abundance and movement activity of bats inhibernaculum
Rhinolophus hipposideros and M. myotispredominate in the majority of the MoravianKarst caves during the winter season (Bauer-ova and Zima 1988; Zima et al. 1994; Rehaket al. 1994; Kovarık 1997). Rhinolophushipposideros exhibited a stable abundance ofhibernating community from the beginningof December to mid-March (cf. Daan andWichers 1968) when the level of its movementactivity was the lowest. On the contrary, theprevious and successive periods are charac-terized by high movement activity in the cavethat was influenced by the continuous arrivaland/or departure of bats in the hibernacu-lum. The same model of movement activitywith its low level during ‘‘deep hibernation’’was also confirmed by the detection ofultrasound signals (Nagel and Nagel 1997;Rehak and Baron 2001) and Harmata(2000) noted that ‘‘deep hibernation’’ of thisbat species began in the period between
ARTICLE IN PRESS
Fig. 5. Seasonal changes in used height of shelter.
Activity and shelter selection by Myotis myotis and Rhinolophus hipposideros 277
November 15 and 20 and lasted untilFebruary of the next year. During climati-cally suitable days, even in the middle ofwinter, some awakened horseshoe bats leftthe cave and returned to it after a short time(Harmata 2000; Rehak and Baron 2001).General trend in M. myotis abundancechanges (continual growth with rapid emer-gence) was mainly influenced by the changesin the outer part of the cave (Corridor). Theflight activity of bats in the entrance hole ofthe cave is almost nil during the winter period(Rehak et al. 1994). Therefore, the consider-able increase in M. myotis abundance is morelikely due to movements within the cave fromthe inaccessible parts towards the entrancethan to the immigration from neighboring
shelters. Maximum abundance of this speciesat the end of the hibernation period and itsshift towards the entrance part of the caveswas also reported for other European hibar-nacula (Nagel and Nagel 1987; Hanzal andPrucha 1988, 1996; Fuszara et al. 1996).The movement activity of M. myotis couldnot be clearly divided into the successiveperiods based on its level, as done for R.hipposideros. High activity was also con-firmed by Prucha and Hanzal (1989), whoregistered very short mean duration of thehibernation period when newly found batswere included in the analysis (7.1 and 9.1days for M. myotis in different years).Nagel and Nagel (1994) reported the move-ment activity of M. myotis within the
ARTICLE IN PRESS
Fig. 6. Shelter preference in different distances from the cave entrance.
278 J. Zukal et al.
hibernaculum as increasing from mid-Febru-ary till the end of April. This relatively highactivity is associated with the use of shelterswith varying microclimate during the winterperiod, similarly as in other Myotis species(Daan 1973; Hanzal and Prucha 1988). Incase of M. myotis the cave air tempe-rature does not influence winter flight activ-ity, but the activity increases with a decreasein barometric pressure (Nagel and Nagel1994).
Shelter preference
Shelter selection differs in various parts ofthe hibernation period, depending on thechanges of microclimatic conditions in thehibernaculum, and it is also species specific(Daan 1973; Altringham 1996). Saint Gironset al. (1969) and Brosset and Poillet (1985)suggest that the changes in R. ferrumequinumshelters during winter do not correspond tothe changes in environment, as the tempera-ture does not fluctuate in deep parts of cavesused by horseshoe bats, but it correspondsonly to the phases of annual cycle, physiolo-gical status and behavior of studied bats.In general, bats use their hibernation siteswith constant air temperatures during themiddle of the hibernation period. They can
probably reduce their energy consumption bythe selection of these parts during the periodof absolute lack of food. The dynamic partsof the hibernacula are occupied by hibernat-ing bats during the beginning and the end ofthe hibernation period (Baagøe et al., 1988;Baron and Rehak 1997). This behavior canbe related to foraging activity. As the airtemperature in outer dynamic parts reflectsambient temperature changes outside thecave, the bats can awake from lethargy whenthe conditions are suitable for foraging(Whitaker and Rissler 1993). However, themoving of the bats into the dynamic caveparts can also be associated with the tendencyto avoid dry circumstances, i.e., with waterloss compensation (Daan and Wichers 1968;Dorgelo and Punt 1969). Nevertheless, itshould be noted that mainly the temporalchanges in cave use could be biased becauseonly a limited part of the hibernatingpopulation can be found in any particularperiod (Rehak 1997; Rehak and Gaisler1999).Rhinolophus hipposideros hibernated in theKaterinska Cave at unprotected places, al-ways hanging free, and a similar observationwas published by Dorgelo and Punt (1969).Daan and Wichers (1968) registered thepresence of this species in exposed and lessfrequently in semi-exposed places, almost

ARTICLE IN PRESS
Activity and shelter selection by Myotis myotis and Rhinolophus hipposideros 279
always in the rear of an artificial limestonecave system. Moreover, R. hipposideros wasusing almost absolutely the inner parts of theKaterinska Cave (Domes) (with exception ofone individual found in the Corridor),similarly as in the limestone cavern in Polandwhere 68.6% of the hibernating bats werefound in its deepest part (Harmata 2000) andin the pseudokarstic sandstone crevice-typecaves in Northern Moravia (Baron andRehak 1997).The seasonal dynamic in the use of particularparts of the cave was characterized for M.myotis by declining percentages of thisspecies in the Domes (inner parts) andincreasing abundance of bats in the Corridorfrom mid-November to the beginning ofApril. A similar tendency was also registeredfor M. myotis by Dorgelo and Punt (1969)and Hanzal and Prucha (1988). The increas-ing number of bats during winter period wasmost probably caused by the movements ofhibernating bats from highly protected shel-ters and/or places which are not accessible forpeople (high chimneys, etc.) to more exposedplaces. The increase could not be influencedby immigration of bats from neighboringshelters, as the flight activity at the entranceof the Katerinska Cave (netting) was practi-cally nil during winter months (Rehak et al.1994). The mentioned type of movementbehavior was also reported in some otherspecies of Myotis (Daan and Wichers 1968;Daan 1973; Degn 1987; Baagøe et al., 1988).Myotis myotis hibernated mostly exposed,hanging free on the ceiling, free on the wall orfree in the chimney. Nevertheless, they weresometimes found in crevices too (Bezem et al.1964; Hanzal and Prucha 1988). Lesinski(1986) recorded this bat species hibernatingin forts near Warszawa, mainly in ventilationshafts, i.e., in protected type of shelter, andless frequently in crevices and exposed places.Daan and Wichers (1968) observed that M.myotis were using highly protected places inthe entrance section of a limestone cave,similarly as did Hanzal and Prucha (1988)who noted that M. myotis selected ratherprotected (in crevices deeper than 10 cm) orsemi-protected (in crevices up to 10 cm)places after having moved into the colderparts of hibernacula. On the other hand, in
the Katerinska Cave the preference of ex-posed places (ceiling, wall) was higher in theCorridor (60%) than in the Domes (45%).Bezem et al. (1964) also registered the heightwhere the hibernating bats were hanging, therelation between shelter type and heightabove the floor, and the relative height ofsite (height of hibernating bat divided bycorresponding height of corridor). They didnot find any statistically significant correla-tion of these parameters and consequentlythey classified M. myotis as a species usingthe highest located shelters and, on thecontrary, R. hipposideros using the lowestlocation of shelters. There was no significantcorrelation between the type of shelter and itsheight in both M. myotis and R. hipposideroshibernating in Katerinska Cave. Harmata(2000) noted that the height of hanging R.hipposideros did not change during hiberna-tion. The horseshoe bats were hibernatingmostly at height between 1 and 3m but oftenthey used very low lying shelters (20–30 cmabove the floor).Generally, the range of temperatures detectedduring hibernation of both species understudy overlaps in wide extent (R. hipposideros2.0–13.0 1C and M. myotis �4.0 to 12.0 1C,Webb et al. 1996) but both species differ inthe time of arrival in the hibernaculum,development of abundance changes, level ofinternal migration and shelter preference inthe caves. These differences should reflect thelife history and body size of bat species understudy. The cave temperatures selected by batsin a cold climate are negatively correlatedwith their body weight (McNab 1974) andthe clusters of bats select even coldertemperatures. Small non-clustering R. hippo-sideros (5–9 g) developed specific style ofhibernation, i.e., hanging free with wingmembrane covering all the body. It is capableof hibernating at higher air temperature but itneeds the stability of microclimate that isensured by use of inner parts of caves. Myotismyotis, a large species (20–35 g), is able tohibernate in clusters within colder parts ofcave close to the entrance. The clusteringbehavior and large weight determinatealso its ability to have a high movementactivity during the hibernation period(McNab 1974).

ARTICLE IN PRESS
280 J. Zukal et al.
Acknowledgements
The authors wish to thank all colleagues whoparticipated in the field works. We are alsomuch obliged to Prof. Jirı Gaisler, Dr. Sc.,for kindly providing inaccessible literatureand valuable comments on the manuscript,
and RNDr. Miroslav Kovarık, for long-term help and cooperation in the field.The work was supported by a grant ofThe Ministry of Education of the CzechRepublic No. 143100010 and a grant ofGrant Agency of the Czech Republic No.206/01/1555.
Zusammenfassung
Aktivitat und Ruheplatzwahl von winterschlafenden Myotis myotis und Rhinolophushipposideros in der Katerinska Hohle (Tschechien)
In 1992–1993 wurden die in der Katerinska Hohle uberwinternden Individuen von Myotis myotis undRhinolophus hipposideros untersucht. Die Untersuchung bestand aus einer regelmaXigen Zahlung,bei der die Tiere nicht beruhrt bzw. markiert wurden. Es wurden drei grundlegende Parameter ihrerHangplatze aufgenommen (Lage innerhalb der Hohle, Typ und relative Hohe). Insgesamt wurden imLaufe von 26 Kontrollgangen 1141 Einzelbeobachtungen von neun Fledermausarten erfaXt.Bewegungstatigkeit, ausgedruckt als prozentueller Anteil der Neufunde wahrend eines bestimmtenKontrollgangs, wurde wahrend der gesamten Wintersaison registriert. Ihr Niveau schwankteunterschiedlich, wobei die Uberwinterungsperiode von R. hipposideros in drei unterschiedlicheAbschnitte aufgeteilt werden konnte, wahrend das Niveau der Bewegungsaktivitat von M. myotis imVerlauf der Saison standig relativ hoch blieb. Die Ruheplatzwahl von R. hipposideros war unabhangigvon dem Hohlenteil in dem die Fledermause uberwinterten und unterlag im Saisonverlauf keinenAnderungen. Uberwinternde Exemplare von R. hipposideros nutzten vor allem exponierte Platze andenen sie immer frei hingen. Myotis myotis wurde an allen Hangplatztypen bis auf einen festgestellt.Rhinolophus hipposideros nutzte vor allem den mittleren Hohlenteil in einer Entfernung von 121 bis180 m vom Hohleneingang. Von M. myotis wurde ein kurzer Abschnitt des Hohlenganges (zwischen21 und 30 m), d.h. nahe dem Eingang, am meisten bevorzugt. Rhinolophus hipposideros ist einehochspezialisierte Art, welche Hohlenteile mit sehr stabilen mikroklimatischen Bedingungenbevorzugt, wahrend M. myotis den erfaXten Parametern gegenuber als vollig indifferent erscheint.r 2005 Deutsche Gesellschaft fur Saugetierkunde. Published by Elsevier GmbH. All rights reserved.
References
Altringham, J. D. (1996): Bats: biology andbehaviour. New York: Oxford University Press.
Baagøe, H.; Degn, H. J.; Nielsen, P. (1988):Departure dynamics of Myotis daubentonii
(Chiroptera) leaving a large hibernaculum.Vidensk. Meddr dansk Naturh. Foren. 147,
7–24.Baron, I.; Rehak, Z. (1997): Contribution to the
hibernation of lesser horseshoe bats (Rhinolo-
phus hipposideros) in the Vsetın region. Zprav.Okr. Vlastived. Muzea ve Vsetıne 1997, 3–12 (inCzech).
Bauerova, Z.; Zima, J. (1988): Seasonal changes invisits to a cave by bats. Folia Zool. 37, 97–111.
Bezem, J. J.; Sluiter, J. W.; Van Heerdt, P. F.(1964): Some characteristics of the hibernating
locations of various species of bats in southLimburg. II. Proc. Koninkl. Nederl. Akademievan Wetenschappen, Ser. C 67, 337–350.
Brosset, A.; Poillet, A. (1985): Structure d’unepopulation hibernante de grands rhinolophesRhinolophus ferrumequinum dans l’est de laFrance. Mammalia 49, 221–233.
Daan, S. (1973): Activity during natural hiberna-tion in three species of vespertilionid bats.Netherlands J. Zool. 23, 1–71.
Daan, S.; Wichers, H. J. (1968): Habitat selectionof bats hibernating in a limestone cave. Z.Saugetierkd. 33, 262–287.
Degn, H. J. (1987): Bat counts in Mønstedlimestone cave during the year. Myotis 25,
85–90.

ARTICLE IN PRESS
Activity and shelter selection by Myotis myotis and Rhinolophus hipposideros 281
Dorgelo, J.; Punt, A. (1969): Abundance and‘‘internal migration’’ of hibernating bats in anartificial limestone cave (‘‘Sibbergroeve’’). Lynx(n.s.) 10, 101–125.
Fuszara, E.; Kowalski, M.; Lesinski, G.; Cygan, J.P. (1996): Hibernation of bats in undergroundshelters of central and northeastern Poland.Bonn. Zool. Beitr. 46, 349–38.
Gaisler, J. (1970): Remarks on the thermoprefer-endum of palearctic bats in their naturalhabitats. Bijdr. Dierk. (Amsterdam) 40, 33–35.
Hanzal, V.; Prucha, M. (1988): Seasonal dynamicsof bat communities at winter shelters of Ceskykras (Central Bohemia) during 1984–1986.Lynx (n.s.) 24, 15–35 (in Czech).
Hanzal, V.; Prucha, M. (1996): Annual course ofcave visitation by bats (Mammalia: Chiroptera)in the Bohemian Karst (Czech Republic). ActaSoc. Zool. Bohem. 60, 25–30.
Harmata, W. (2000): Phenological dynamics ofhibernating lesser horseshoe bat Rhinolophus
hipposideros (Bechst.) (Chiroptera, Rhinolophi-dae) in a limestone cavern. Studia Chiropterol.1, 13–28 (in Polish).
Kovarık, M. (1997). Research and protection ofRhinolophus hipposideros in protected landscapearea Moravian Karst during hibernation. In:Zur Situation der Hufeisennasen in Europa.Ed. by Arbeitskreisirs Fledermause Sachsen-Anhalt e.V. Berlin: IFA Verlag. Pp. 85 – 87.
Lesinski, G. (1986): Ecology of bats hibernatingunderground in central Poland. Acta Theriol.31, 507–521.
McNab, B. K. (1974): The behavior of temperatecave bats in a subtropical environment. Ecol-ogy 55, 943–958.
Nagel, A.; Nagel, R. (1987): Veranderungen desBestandes winterschlafender Fledermause imWinter 1982/83 in Wurttemberg. Myotis 25,
91–94.Nagel, A.; Nagel, R. (1994): Activity of hibernating
bats in their natural habitat recorded byautomatic data logging. Bat Res. News 35,
37.Nagel, A., Nagel, R. (1997). Nutzung eines
Untertagequartieres durch die Kleine Hufeisen-nase (Rhinolophus hipposideros). In: Zur Situa-tion der Hufeisennasen in Europa. Ed. by
Arbeitskreisirs Fledermause Sachsen-Anhalte.V. Berlin: IFA Verlag. Pp. 97 – 108.
Prucha, M.; Hanzal, V. (1989): Some aspects ofhibernation of bats wintering in the Bohemiankarst (Central Bohemia, Czechoslovakia). ActaUniv. Carol. Biol. 33, 315–333.
Rehak, Z. (1997): Long-term changes in the size ofbat populations in Central Europe. Vespertilio2, 81–96 (in Czech).
Rehak, Z.; Baron, I. (2001): Detectoring of flightactivity of Lesser horseshoe bats, Rhinolophus
hipposideros, at a cave entrance over a year.Nietoperze 2, 202–208.
Rehak, Z.; Gaisler, J. (1999): Long-term changes inthe number of bats in the largest man-madehibernaculum of the Czech Republic. ActaChiropterol. 1, 113–123.
Rehak, Z.; Zukal, J.; Kovarık, M. (1994): Long-and short-term changes in the bat communityof the Katerinska cave (Moravian karst)—fundamental assessment. Folia Zool. 43,
425–436.Saint Girons, H.; Brosset, A.; Saint Girons, M. C.
(1969): Contribution a la connaissance du cycleannuel de la chauve-souris Rhinolophus ferru-
mequinum (Schreber, 1774). Mammalia 33,
357–374.Sokal, R. R.; Rohlf, F. J. (1981): Biometry. New
York: Freeman and Co.Webb, P. I.; Speakman, J. R.; Racey, P. A. (1996):
How hot is a hibernaculum? A review of thetemperatures at which bats hibernate. Can. J.Zool. 74, 761–765.
Whitaker Jr., J. O.; Rissler, L. J. (1993): Do batsfeed in winter? Am. Midl. Nat. 129, 200–203.
Zima, J.; Kovarık, M.; Gaisler, J.; ak, Z.; Zukal, J.(1994): Dynamics of the number of batshibernating in the Moravan karst in 1983 to1992. Folia Zool. 43, 109–119.
Authors’ addresses:
Jan Zukal and Hana Berkova, Institute of VertebrateBiology Academy of Sciences of the Czech Republic,Kvetna 8, 603 65 Brno, Czech Republic(e-mail: [email protected])Zdenek Rehak, Department of Zoology and Ecology,Faculty of Science, Masaryk University Brno, Ko-tlarska 2, 611 37 Brno, Czech Republic


![Yap - Shelter for All - An Enabling or Empowering Strategy [1995]](https://static.fdokumen.com/doc/165x107/631a1dc720bd5bb1740c2432/yap-shelter-for-all-an-enabling-or-empowering-strategy-1995.jpg)

















