
Oligocene‐miocene palynomorph assemblages from eastern Venezuela
Upload
khangminh22Category
view
1download
0
729
Journal of Mammalogy, 84(2):729–752, 2003
A NEW BAT (CHIROPTERA: NATALIDAE) FROM THE EARLYMIOCENE OF FLORIDA, WITH COMMENTS ON
NATALID PHYLOGENY
GARY S. MORGAN* AND NICHOLAS J. CZAPLEWSKI
New Mexico Museum of Natural History, 1801 Mountain Road NW,Albuquerque, NM 87104, USA (GSM)
Oklahoma Museum of Natural History, University of Oklahoma, Norman, OK 73072, USA (NJC)
We describe a new extinct genus and species of bat belonging to the endemic Neotropicalfamily Natalidae (Chiroptera) from the Thomas Farm Local Fauna in northern peninsularFlorida of early Miocene age (18–19 million years old). The natalid sample from ThomasFarm consists of 32 fossils, including a maxillary fragment, periotics, partial dentaries,isolated teeth, humeri, and radii. A proximal radius of an indeterminate natalid is reportedfrom the I-75 Local Fauna of early Oligocene age (about 30 million years old), also fromnorthern Florida. These fossils from paleokarst deposits in Florida represent the 1st Tertiaryrecords of the Natalidae. Other extinct Tertiary genera previously referred to the Natalidae,including Ageina, Chadronycteris, Chamtwaria, Honrovits, and Stehlinia, may belong tothe superfamily Nataloidea but do not fit within our restricted definition of this family.Eight derived characters of the Natalidae sensu stricto are discussed, 5 of which are presentin the new Miocene genus. Intrafamilial phylogenetic analysis by parsimony of the Natal-idae suggests that the 3 living subgenera, Natalus (including N. major, N. stramineus, andN. tumidirostris), Chilonatalus (including C. micropus and C. tumidifrons), and Nyctiellus(including N. lepidus), deserve full generic rank. The Natalidae apparently evolved in NorthAmerica before the late Oligocene, went extinct in what is now the Nearctic region (i.e.,Florida) after the early Miocene, and survived in tropical Middle America during the re-mainder of the Tertiary. The presence of 2 endemic genera and 4 endemic species suggeststhat natalids reached the West Indies by overwater dispersal early in their history (Oligoceneor Miocene). The lack of a Tertiary fossil record, marginal distribution, and limited speciesrichness and endemism of natalids in South America are suggestive of a comparatively latearrival on that continent, possibly in the late Pliocene after the beginning of the GreatAmerican Faunal Interchange.
Key words: bat, biogeography, Chiroptera, Florida, fossil, Miocene, Natalidae, Nataloidea, paleo-karst, phylogeny
We describe a new extinct genus and spe-cies of bat in the endemic Neotropical fam-ily Natalidae from the early Miocene (earlyHemingfordian land mammal age) ThomasFarm Local Fauna in northern peninsularFlorida. The Natalidae are a small familycomposed of 6 living species restricted toMiddle America, South America, and the
* Correspondent: [email protected]
West Indies. The species described in thisstudy is the 1st known extinct member ofthe Natalidae, excluding 5 other extinct Ter-tiary genera (Ageina, Chadronycteris,Chamtwaria, Honrovits, and Stehlinia) thathave been considered natalids based on theconcept of the family proposed by Van Val-en (1979). These 5 genera are referred tothe superfamily Nataloidea following Sim-
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

730 Vol. 84, No. 2JOURNAL OF MAMMALOGY
mons and Geisler (1998) but are not con-sidered members of the Natalidae sensustricto.
We also report a proximal radius of a na-talid from the Oligocene I-75 Local Fauna,located near Gainesville, Alachua County,Florida, about 80 km southeast of ThomasFarm. The I-75 site is the oldest land-ver-tebrate locality in Florida (Patton 1969),dating to the Whitneyan land mammal age(early Oligocene, about 30 3 106 yearsago). The radius from the I-75 site cannotbe identified below the family level but isimportant because it represents the oldestfossil record of the Natalidae.
The discovery of a new genus and spe-cies of Natalidae from the early Miocene ofFlorida provides new information on thehistorical biogeography and phylogeny ofthis small and enigmatic group of NewWorld bats. The identification of a natalidnearly 20 million years older than any otherknown member of this family prompted usto examine the phylogenetic relationshipsof all species within the Natalidae. Al-though our primary aim was to determinethe systematic position of the new FloridaMiocene natalid, we also examined the re-lationships among the 6 extant species ofnatalids because no formal hypothesis of re-lationships has been proposed for the fam-ily.
MATERIALS AND METHODS
In the late 1950s and early 1960s, ClaytonRay of the Florida State Museum (now the Flor-ida Museum of Natural History [FLMNH]) andPierce Brodkorb of the University of Florida De-partment of Zoology began screenwashing sed-iments from Thomas Farm for small vertebrates.These early screenwashing efforts led to system-atic studies of frogs (Holman 1965, 1967), sal-amanders and lizards (Estes 1963), snakes (Auf-fenberg 1963), birds (Brodkorb 1956), and ro-dents (Black 1963). Bat fossils also were recov-ered during this early screenwashing program,but early finds did not include specimens of thenew natalid. The majority of bat fossils fromThomas Farm, including all natalid specimensdescribed here, were collected from about 2 met-
ric tons of sediment screenwashed between 1981and 1985 by Ann Pratt and Arthur Poyer as partof Pratt’s dissertation on the taphonomy and pa-leoecology of the Thomas Farm vertebrate fauna(Pratt 1989, 1990; Pratt and Morgan 1989). Theuse of fine-mesh screens (1-mm opening) re-sulted in the recovery of large numbers of iso-lated bat teeth that passed through the standardwindow screening used by earlier workers. Batfossils were recovered throughout the 3-m sec-tion of sediments excavated at Thomas Farm inthe early 1980s, but the greatest number of batsoccurred in a 1-m-thick unit of lime sand nearthe top of the section (unit 15; Pratt 1989,1990).
All fossils cited are from the vertebrate pale-ontology collection, FLMNH, University ofFlorida, Gainesville (UF). We follow the chirop-teran dental terminology of Legendre (1984) andMenu and Sige (1971). We use standard abbre-viations for tooth positions in mammals, withuppercase letters for upper teeth and lowercaseletters for lower teeth: I and i (upper and lowerincisors), C and c (upper and lower canines), Pand p (upper and lower premolars), and M andm (upper and lower molars). The terminologyfor chiropteran postcranial elements followsSmith (1972) and Vaughan (1959). All measure-ments of fossils are in millimeters.
We compared the Thomas Farm natalid fossilswith at least 2 skulls and 2 complete skeletonsof each of the 6 extant species of Natalidae andthe potentially related families Furipteridae,Thyropteridae, and Vespertilionidae (see Appen-dix I for comparative material examined).
Methods of phylogenetic analysis.—There isno previous intrafamilial phylogenetic analysisfor the Natalidae. We developed a hypothesis ofrelationships based on parsimony analysis (usingPAUP 4.0b10 software—Swofford 2000) of theFloridian fossils and all 5 extant species gener-ally recognized as members of the Natalidae(Koopman 1993, 1994; Nowak 1994; AppendixI) plus Natalus major, which we recognize as avalid species following Morgan (1989b). Mor-phological comparisons disclosed 50 charactersthat may be phylogenetically informative (Ap-pendix II). We scored 24 cranial, dental, and hu-meral characters that are present in the fragmen-tary fossils as well as in extant natalids (Appen-dix II, characters 1–24). The remaining 26 cra-nial and postcranial characters were availableonly in the extant natalids (with missing values
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 731
FIG. 1.—Location of the 2 sites in Florida thathave produced Tertiary fossils of Natalidae, I-75(Oligocene) and Thomas Farm (early Miocene).
coded for the fossil taxon). Characters and char-acter states are based on Dalquest (1950), Good-win (1959), Koopman (1994), Miller (1907), Ot-tenwalder and Genoways (1982), Silva Taboada(1979), Simmons (1998), Simmons and Geisler(1998), Van Valen (1979), and our personal ob-servations.
There has been controversy surrounding thehigher-level relationships of the Natalidae. In 1study by Simmons and Geisler (1998), the Na-talidae were part of a clade (Nataloidea) thatalso included the Furipteridae, Thyropteridae,and Myzopodidae. In their analysis (Simmonsand Geisler 1998), this clade was sister to theVespertilionoidea (Vespertilionidae and Molos-sidae) and farther removed from the Noctilion-oidea. In a contrasting analysis based on mito-chondrial DNA, Van Den Bussche and Hoofer(2001) found the nataloids (except Myzopodi-dae) strongly associated with the Noctilionoidea(Noctilionidae, Mystacinidae, Mormoopidae,and Phyllostomidae) and far removed from theVespertilionidae and Molossidae. More recently,using additional evidence from nuclear genes,Hoofer et al. (in press) reexamined yangochirop-teran bats, including all the aforementioned fam-ilies. In their analysis Nataloidea was found tobe polyphyletic; Natalidae clustered with Ves-pertilionidae and Molossidae, whereas Furipter-idae and Thyropteridae clustered with the Noc-tilionoidea. Based in part on the analysis ofHoofer et al. (in press) and in part on the tra-ditional association of Natalidae with Vesperti-lionidae from morphological data, we used 2vespertilionids (Kerivoula picta and Myotis lu-cifugus) as a monophyletic out-group in ourphylogenetic analysis. Moreover, in a recent ar-ticle, Horacek (in press) noted that among livingvespertilionids, Myotis and Kerivoula had themost primitive dentition.
A simple branch-and-bound search yielded asingle most parsimonious tree of 82 steps. Wethen used the bootstrap and jackknife methodswith branch-and-bound search to examine treesthat were 1 step to a few steps longer than themost parsimonious tree in order to calculate Bre-mer support values for branches in the consen-sus cladograms (Simmons 2000). Results frombootstrap and jackknife methods were very sim-ilar, so only the bootstrap results are reportedbelow. The number of bootstrap replicates wasset to 10,000 but was similar when set to 1,000.
Fossil localities.—The Thomas Farm site, lo-
cated 12 km north of Bell in Gilchrist County innorthern peninsular Florida (Fig. 1), has pro-duced the best known early Miocene (earlyHemingfordian land mammal age) vertebratefauna in eastern North America. There are de-tailed accounts of the discovery, excavation, ge-ology, and stratigraphy of Thomas Farm (Auf-fenberg 1963; Pratt 1989, 1990; Simpson 1932;White 1942), as well as faunal lists and bibli-ographies (Olsen 1962; Ray 1957; Webb 1981).
The Thomas Farm Local Fauna is composedof about 90 species of vertebrates (Webb 1981),primarily consisting of terrestrial forms, al-though freshwater taxa are present, includingfrogs, salamanders, pond turtles, alligators, andaquatic birds. There are 23 species of largemammals from Thomas Farm, the most abun-dant of which is the three-toed horse, Parahip-pus leonensis (Hulbert 1984). This site has anexceptionally rich small-vertebrate fauna ofnearly 70 species, including large and taxonom-ically diverse samples of frogs, lizards, snakes,birds, bats, and rodents. In addition to bats, othersmall mammals from Thomas Farm include adidelphid, a soricid, 3 sciurids, 2 heteromyids,and an eomyid (Pratt 1989; Webb 1981).
The fossiliferous Miocene sediments atThomas Farm consist of alternating layers ofclay and sand filling a 30-m-deep sinkhole de-veloped in Eocene marine limestone (Pratt 1989,1990). The fossil assemblage from the lime sand
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

732 Vol. 84, No. 2JOURNAL OF MAMMALOGY
appears to represent a cave deposit that formedthrough 2 processes: the natural accumulation ofbat carcasses on a cave floor and a coprocoe-nosis representing the fecal remains from smallmammalian carnivores or the regurgitation pel-lets of raptorial birds. Pratt (1989) concludedthat the Thomas Farm site consisted of a deepsinkhole surrounded by a forested habitat. Theabundance of bats suggests that caves probablyoccurred in the walls of the sinkhole.
The mammalian fauna from Thomas Farm, inparticular the carnivores, equids, artiodactyls,and sciurids (Forsten 1975; Maglio 1966; Prattand Morgan 1989; Tedford and Frailey 1976),indicates a late early Hemingfordian age (be-tween 18 3 106 and 19 3 106 years ago). Theearly Hemingfordian is defined in part by the 1stappearance of the immigrant carnivores Amphi-cyon, Hemicyon (Phoberocyon), and Leptarctus,the 1st occurrence of the canid Tomarctus andthe camelid Floridatragulus, and the last occur-rence of the small amphicyonid Cynelos and therhinocerotid Menoceras (Tedford et al. 1987), allof which are present at Thomas Farm. Other cor-relative early Hemingfordian faunas from thewestern United States are the Garvin Gully Fau-na of Texas, the faunas from the RunningwaterFormation of Nebraska, the Flint Hill Local Fau-na of South Dakota, and the Martin Canyon Lo-cal Fauna of Colorado (Tedford et al. 1987).
Fossil bats were initially reported from Thom-as Farm by Lawrence (1943), who described 2new genera and species of Vespertilionidae,Suaptenos whitei and Miomyotis floridanus.Czaplewski and Morgan (2000) recently de-scribed a 3rd extinct genus and species of ves-pertilionid from Thomas Farm, Karstala silva,characterized by its large size. All the natalidfossils from Thomas Farm were collected in theearly 1980s as part of an intensive screenwash-ing project (Pratt 1989, 1990). Thomas Farm hasthe largest known sample of bats from any pre-Pleistocene fossil deposit in North America,numbering almost 2,000 specimens. The chirop-teran fauna is composed of at least 8 species,including 1 species each in the Neotropical fam-ilies Emballonuridae, Natalidae, and Molossidaeand 5 species of Vespertilionidae (Czaplewskiand Morgan 2000; Morgan 1989a; Morgan andCzaplewski 2000). In addition to Suaptenos whi-tei, Miomyotis floridanus, and Karstala silva, theThomas Farm vespertilionids include an unde-scribed genus similar to Lasiurus and a 2nd un-
described genus similar to Corynorhinus. TheVespertilionidae dominate the Thomas Farm chi-ropteran fauna.
The I-75 Local Fauna is not nearly as welldocumented as Thomas Farm, with only 1 sum-mary paper on the locality (Patton 1969). Thesite was discovered in 1965 during the construc-tion of Interstate Highway 75 through Gaines-ville, Alachua County, in northern peninsularFlorida (Fig. 1). The I-75 site consists of fossil-iferous clays deposited in a small karst solutioncavity developed in marine Eocene limestone.The I-75 site has a diverse vertebrate fauna con-taining about 45 species (Patton 1969). Thelarge mammal fauna is composed of 2 carni-vores, including the amphicyonid Daphoenusand a mustelid, the horse Miohippus, a tayas-suid, an oreodont, and the small artiodactyl Lep-tomeryx. I-75 also has a rich microvertebratefauna, including frogs, salamanders, lizards,snakes, and small mammals (Holman 1999; Pat-ton 1969). Small mammals include the didelphidHerpetotherium; the insectivore Centetodon cf.C. wolffi; 6 species of bats (see below); the le-porid Palaeolagus; and at least 4 species of ro-dents, Eutypomys, a heteromyid, and 2 eomyids.
Previous workers (Emry et al. 1987; Patton1969) placed the I-75 Local Fauna in the Whit-neyan land mammal age (early Oligocene, 30 3106 to 32 3 106 years ago—Woodburne andSwisher 1995), making it the oldest land ver-tebrate fauna known from Florida. Whitneyanfaunas are otherwise unknown outside thenorthern Great Plains (Tedford et al. 1996).Most of the age-diagnostic mammals from I-75occur in the Whitneyan and early Arikareeanland mammal ages. The presence of Centeto-don, Eutypomys, Leptomeryx, Miohippus, andPalaeolagus in I-75 establishes an early Ari-kareean or older age (older than 24 3 106 yearsago—Tedford et al. 1987, 1996). The similarityof the bat faunas from I-75 and the early Ari-kareean (26 3 106 to 28 3 106 years ago)Brooksville 2 Local Fauna from central Florida(Hayes 2000) suggests that these 2 faunas areclose in age. The absence of mammals in theI-75 Local Fauna that are restricted to the Ari-kareean (e.g., the small artiodactyl Nanotra-gulus) indicates a late Whitneyan age (about 303 106 years ago), making the I-75 natalid spec-imen about 12 million years older than theThomas Farm natalid.
The I-75 site provides the earliest record of
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 733
FIG. 2.—Dentaries of Primonatalus prattae n.gen. et sp. A–C) Partial right dentary with m1–3 (UF 108641, holotype) in labial, occlusal, andlingual views. D and E) Posterior fragment ofright dentary with intact angular process (UF121145) in labial and lingual views. F–H) Eden-tulous left partial ramus with mandibular sym-physis and alveoli for i1–3, c1, and p2–4 (UF108647) in labial, occlusal, and lingual views.
the taxonomic diversity and community struc-ture of North American middle Cenozoic chi-ropteran faunas. In addition to the natalid re-ported in this study, 5 other species of bats areknown from I-75, including a new mormoopid,2 new emballonurids, a new possible phyllos-tomid, and a vespertilionid. Species belonging tofamilies now restricted to the Neotropics are themost abundant bats in the I-75 fauna, includinga large emballonurid and a small mormoopid.
SYSTEMATIC PALEONTOLOGY
Primonatalus, new genus
Type species.—Primonatalus prattae.Included species.—Only the type species
is known.Diagnosis.—Mandibular ramus is deep
below molars. Ventral margin of mandibleis straight between p4 and mandibular an-gle. Summit of coronoid process of man-dible is comparatively well developed withtriangular-shaped dorsal tip. Anterior edgeof coronoid process curves posterodorsallyfrom alveolar margin to dorsal tip of coro-noid at an angle of about 708. Coronoid pro-cess rises above level of articular condyle.Posterior portion of coronoid process slopesventrally from tip of coronoid down to con-dyle. Distinct mandibular angle is presentdirectly ventral to tip of coronoid. Base ofangular process is located halfway betweenventral edge of mandible and alveolar mar-gin in vertical dimension and about halfwaybetween coronoid and condyle in antero-posterior dimension. Angular process doesnot flare strongly laterally. Mandibular fo-ramen opens level with alveolar margin.Two features, the comparatively well-de-veloped triangular-shaped coronoid processthat is taller than the articular condyle andthe ventral position of the mandibular fo-ramen and angular process, reflect a lesserdegree of dorsal cranial flexion in Primon-atalus than in Natalus. Posterior mental fo-ramen located in deep concavity near al-veolar margin between roots of c1 and p2.The p3 is larger, longer, and more com-pressed with distinct concavity posterior tomain cusp. Metaconid and entoconid same
height on m1 and m2. Carnassial-likenotches on cristid obliqua and postcristid onlower molars (Figs. 2A, 2B, and 3C) pre-sent but weak. Lophid associated with car-nassial-like notch extending posteriorlyfrom cristid obliqua into talonid basin ab-sent. Talonid notch was deeper than in Re-
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

734 Vol. 84, No. 2JOURNAL OF MAMMALOGY
FIG. 3.—Lower teeth of Primonatalus prattaen. gen. et sp. A) Left dentary fragment with p3,alveoli for c1, p2, and p4, and anterior root ofm1 (UF 108642) in labial view. B) Same as A,in occlusal view. C) Right lower molar series(UF 108641, holotype) in occlusal view.
cent natalids because of taller metaconidand more ventral connection of entocristidto trigonid. Cristid obliqua also connects totrigonid more ventrally. Labial cingularcusp of P4 is low. Occlusal outline of P4has anterior indentation. Talon weakly de-veloped on M1–2. Lingual cingulum weakon upper molars. Humerus has broad tri-angular-shaped epitrochlea with no notchbetween weak medial process and distalspinous process. Distal spinous processsmall, projecting only to edge of trochlea orvery slightly distal to it, and separated fromtrochlea by narrow but distinct notch.
Etymology.—Primus (Latin), 1st, origi-nal, early, and Natalus (Latin), genericname for all currently recognized Recentspecies in the family Natalidae. The nameindicates that this is the earliest known rep-resentative of the Natalidae.
Primonatalus prattae, new species
Holotype.—UF 108641, partial right den-tary with m1–3 (Figs. 2A–C and 3C). Theholotype, paratypes, and all referred speci-mens are from the early Miocene ThomasFarm Local Fauna, Florida.
Paratypes.—UF 121145, ascending ra-mus of right dentary (Figs. 2D and 2E); UF108647, edentulous partial left dentary withsymphysis and alveoli for i1–p4 (Figs. 2F–H); UF 108642, partial left dentary with p3and alveoli for p2 and p4 (Figs. 3A and3B); UF 121136, right P4 (Figs. 4E–G); UF108638, partial left maxilla with M1–2(Figs. 4B–D); UF 121139, right M3 (Fig.4A); UF 121141, periotic (Figs. 5A–D); UF108650, distal half of left humerus (Fig. 6).
Referred specimens.—UF 121137, partialleft maxilla with M1; UF 108639, 108640,2 right M1; UF 121138, left M1; UF121140–121142, 3 periotics (2 left, 1 right);UF 121143, partial left dentary with m1–2;UF 108643, partial right dentary with m1and alveoli for p4 and m2; UF 108644, par-tial left dentary with m1; UF 108646, eden-tulous partial right dentary with alveoli form1–3; UF 121144, edentulous partial leftdentary with alveolus for m3; UF 108645,right m3; UF 108648, 108649, 108651,108652, 4 distal portions of humeri (3 right,1 left); UF 108653, 108654, 2 right proxi-mal portions of radii; UF 108655–108658,4 distal portions of radii (2 right, 2 left);UF 108660, right proximal metacarpal III;UF 108659, distal portion of femur.
Type locality and age.—Thomas FarmLocal Fauna (UF locality number GI01), 12km northeast of Bell, Gilchrist County,Florida, early Miocene, early Hemingfor-dian.
Diagnosis.—Same as for genus.Etymology.—Named for Ann E. Pratt in
recognition of her work on Miocene smallvertebrate faunas from Florida. Dr. Pratt’sscreenwashing efforts at Thomas Farm pro-duced all the natalid fossils described in thisstudy.
Descriptions and comparisons.—The
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 735
FIG. 4.—Upper teeth of Primonatalus prattaen. gen. et sp. A) Right M3 (UF 121139) in oc-clusal view. B–D) Left maxillary fragment withM1–2 and lateral alveoli for M3 (UF 108638) inocclusal, posterior, and labial views. E–G) RightP4 (UF 121136) in labial, occlusal, and lingualviews.
Thomas Farm fossils of Primonatalus arecompared with Recent Natalidae in consid-erable detail because of their importance ininterpreting the phylogenetic relationshipsamong all natalids. Most comparisons arewith the 3 subgenera of Natalus: Natalus(including the species major, stramineus,and tumidirostris), Chilonatalus (includingthe species micropus and tumidifrons), andNyctiellus (including the species lepidus).The subgenera of Natalus are discussed inthe context of genera, and our phylogeneticanalysis suggests that these 3 subgenera arebest recognized as genera (see below). In-dividual species are mentioned only wheresignificant differences exist between thespecies in a subgenus. Primonatalus is in-termediate in size for the family, smallerthan the species in the subgenus Natalus butlarger than N. (Chilonatalus) micropus andN. (Nyctiellus) lepidus. The fossils are mostsimilar in size to N. (Chilonatalus) tumidi-frons from the Bahamas.
The most complete fossil of Primonatal-us prattae from Thomas Farm is the holo-type, a partial right dentary with all 3 mo-lars (UF 108641; Figs. 2A–C and 3C). Theascending ramus of the type is nearly com-plete, but the horizontal ramus is missinganterior to m1. The coronoid process andarticular condyle are intact, and the angularprocess is missing only the tip. The hori-zontal ramus is comparatively deep belowthe molars and has a straight ventral edge.The ventral margin is nearly parallel to thealveolar margin of the toothrow from thep4 posteriorly to the mandibular angle. Thehorizontal ramus is also relatively deepwith nearly parallel ventral and alveolarmargins in Natalus. The horizontal ramusis more slender and curved ventrally belowthe m1 and m2 in Chilonatalus and Nyc-tiellus. The coronoid process of Primona-talus is triangular in shape and has a defi-nite dorsal tip. The tip of the coronoid istaller than the articular condyle so that theposterior portion of the coronoid processslopes posteroventrally from its tip to thecondyle. In all Recent natalids, the coronoid
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022
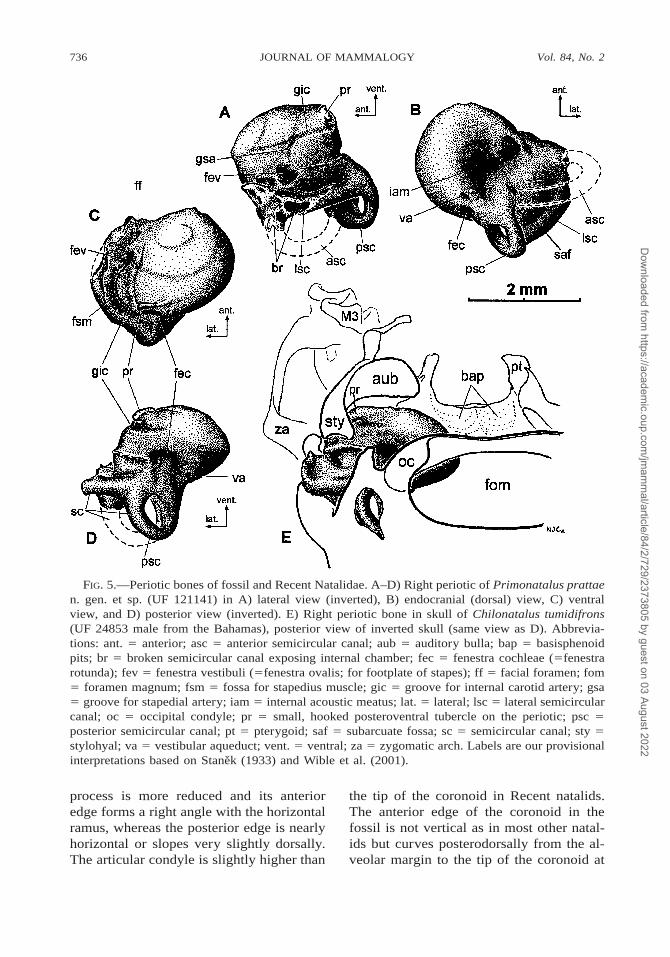
736 Vol. 84, No. 2JOURNAL OF MAMMALOGY
FIG. 5.—Periotic bones of fossil and Recent Natalidae. A–D) Right periotic of Primonatalus prattaen. gen. et sp. (UF 121141) in A) lateral view (inverted), B) endocranial (dorsal) view, C) ventralview, and D) posterior view (inverted). E) Right periotic bone in skull of Chilonatalus tumidifrons(UF 24853 male from the Bahamas), posterior view of inverted skull (same view as D). Abbrevia-tions: ant. 5 anterior; asc 5 anterior semicircular canal; aub 5 auditory bulla; bap 5 basisphenoidpits; br 5 broken semicircular canal exposing internal chamber; fec 5 fenestra cochleae (5fenestrarotunda); fev 5 fenestra vestibuli (5fenestra ovalis; for footplate of stapes); ff 5 facial foramen; fom5 foramen magnum; fsm 5 fossa for stapedius muscle; gic 5 groove for internal carotid artery; gsa5 groove for stapedial artery; iam 5 internal acoustic meatus; lat. 5 lateral; lsc 5 lateral semicircularcanal; oc 5 occipital condyle; pr 5 small, hooked posteroventral tubercle on the periotic; psc 5posterior semicircular canal; pt 5 pterygoid; saf 5 subarcuate fossa; sc 5 semicircular canal; sty 5stylohyal; va 5 vestibular aqueduct; vent. 5 ventral; za 5 zygomatic arch. Labels are our provisionalinterpretations based on Stanek (1933) and Wible et al. (2001).
process is more reduced and its anterioredge forms a right angle with the horizontalramus, whereas the posterior edge is nearlyhorizontal or slopes very slightly dorsally.The articular condyle is slightly higher than
the tip of the coronoid in Recent natalids.The anterior edge of the coronoid in thefossil is not vertical as in most other natal-ids but curves posterodorsally from the al-veolar margin to the tip of the coronoid at
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 737
FIG. 6.—Distal portion of left humerus (UF108650) of Primonatalus prattae n. gen. et sp.:A) anterior view; B) medial view; C) posteriorview; D) lateral view; and E) distal view. Ab-breviations: c 5 capitulum; le 5 lateral epicon-dyle; lrc 5 lateral ridge of capitulum; me 5 epi-trochlea or medial epicondyle; mp 5 medial pro-cess of epitrochlea; mrc 5 medial ridge of ca-pitulum; rf 5 radial fossa; sp 5 spinous processof epitrochlea (5distal spinous process); tr 5trochlea.
an angle of about 708. The presence of awell-developed coronoid with a distinct tri-angular dorsal portion is clearly a primitivefeature of Primonatalus, probably reflectingits lesser degree of dorsal cranial flexioncompared with modern natalids.
Primonatalus has a distinct mandibularangle located on the ventral edge of thedentary anterior to the angular process anddirectly below the dorsal tip of the coro-noid. There is a minor degree of dorsal flex-ion of the ascending ramus posterior to themandibular angle. A distinct mandibularangle is lacking in Natalus because of thesharp dorsal upturning of the mandible justposterior to m3. Chilonatalus has a weakmandibular angle located in a more dorsalposition and farther posteriorly than in thefossil, posterior to the tip of the coronoid.Nyctiellus has a distinct mandibular anglelocated slightly posterior to the dorsalmostprojection of the coronoid and dorsal to theventral margin of the horizontal ramus.
The angular process on the type dentaryof Primonatalus is damaged, but it does notflare as strongly laterally as in other natal-ids, particularly Natalus. In lateral aspect,the base of the angular process of the fossilis situated at a level halfway between thealveolar margin and the ventral edge of thedentary and is also located farther anteriorlythan in living natalids, approximately half-way between the articular condyle and thetip of the coronoid. The anteroventral po-sition of the angular process in Primona-talus also reflects the lesser dorsal flexionof the ascending ramus in the fossil species.Viewed laterally, the angular process is lo-cated directly ventral to the articular con-dyle in Natalus and is positioned well dor-sal to the alveolar margin of the toothrow.The angular process of Chilonatalus is lo-cated directly below, or very slightly ante-rior to, the articular condyle and is at thesame level or slightly dorsal to the alveolarmargin of the toothrow. In Nyctiellus, theangular process is located anterior to the ar-ticular condyle and slightly ventral to thealveolar margin of the toothrow. In dorsal
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

738 Vol. 84, No. 2JOURNAL OF MAMMALOGY
and posterior views, the angular process ofPrimonatalus does not flare laterally fromthe vertical plane of the ascending ramus.In Chilonatalus and Nyctiellus the angularprocess flares slightly, and in Natalus itflares widely. In lateral view, the angularprocess of Primonatalus is curved andbears a small transverse crest along the dor-sal edge of the tip. The angular process ismore curved with a sharper hook in Nyc-tiellus, strongly curved and hooked at thetip in Chilonatalus, and straight with a crestalong the lateral edge of the tip in Natalus.
In dorsal view, the ascending ramus ofthe dentary in Primonatalus is not inclinedas far laterally as in Natalus and Chilona-talus. The coronoid process of the fossil isnearly vertical and located in the sameplane as the labial edge of the toothrow.The coronoid process flares much fartherlaterally in Natalus and Chilonatalus. Nyc-tiellus is somewhat intermediate betweenPrimonatalus and the 2 other living sub-genera in the location of the coronoid pro-cess. The relative position of the mandibu-lar foramen also reflects the degree of dor-sal flexion of the braincase and ascendingramus. The mandibular foramen is locatedat the same level as the alveolar margin inPrimonatalus and Nyctiellus and dorsal tothe alveolar margin in Chilonatalus and Na-talus. Among the 3 living subgenera of Na-talidae, Natalus shows the greatest degreeof dorsal flexion of the skull and conse-quently the strongest dorsal upturning ofthe ascending ramus of the dentary. Nyc-tiellus shows the least dorsal flexion of thebraincase and ascending ramus, whereasChilonatalus is somewhat intermediate, al-though more similar to Natalus. All livingnatalids are more derived in this suite ofcharacters than is Primonatalus. Comparedwith living natalids, Primonatalus has a re-duced degree of dorsal upturning of the as-cending ramus of the dentary that is reflect-ed in the ventral position of the articularcondyle, angular process, and mandibularforamen and the well-developed coronoidprocess with a distinct dorsal process. The
fossil species is clearly the most primitivemember of the family in characters relatingto the dorsal flexion of the ascending ra-mus.
The lower molars of Primonatalus aremost similar in size to teeth of Chilonatalustumidifrons from the Bahamas. Measure-ments of P. prattae teeth are provided inTable 1. The cusps of the fossil teeth areless inflated or robust than in Natalus butare more inflated than in Chilonatalus, inwhich the crests are more slender and thecusps are sharp and spikelike. The talonidsare somewhat narrower in the fossil than inChilonatalus. The metaconid and entoconidare the same height on the m1 and m2 ofthe type dentary of P. prattae, whereas onm3 the metaconid is slightly taller than theentoconid. The entoconid is slightly higherthan the metaconid on m1–3 in all livingnatalids. The talonid notch is deeper in thefossil species owing to both the relativelyhigher metaconid and the more ventral con-nection of the entocristid on the trigonid be-low the metaconid. The cristid obliqua alsoconnects nearer the base of the trigonidventral to the protoconid in the fossils, thusforming a deeper notch than those in Na-talus and Chilonatalus. The small carnas-sial-like notches in the cristid obliqua andpostcristid (Figs. 2A, 2B, and 3C) tend toform a small ‘‘valley’’ connecting the 2crests in Primonatalus, but this feature isnot as well developed as in Recent natalids.The ridge trending posteriorly from thecristid obliqua, just lingual to the ‘‘valley’’connecting the carnassial-like notches, islacking in the fossils. Lower molars of Nyc-tiellus are distinctly different from those ofPrimonatalus, Natalus, and Chilonatalus.Talonids are very broad and compose abouttwo-thirds the length of the tooth on m1 andm2. In other natalids, the trigonid and tal-onid are approximately equal in size. Be-cause of the greater breadth of the talonidsin Nyctiellus compared with other natalids,the cristid obliqua forms a more acute anglewith the trigonid. The cristid obliqua attach-es to the trigonid ventrally in Nyctiellus, as
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 739
TABLE 1.—Measurements (in mm) of the teeth of Primonatalus prattae from the early Miocene(Hemingfordian) Thomas Farm Local Fauna, Florida.
Specimen number ToothAnteroposterior
lengthTransverse
widthTrigonid
widthTalonidwidth
UF 121136UF 108639UF 108640UF 121137UF 121138UF 108638UF 108638UF 121139UF 108642
P4M1M1M1M1M1M2M3p3
1.09
1.251.251.28
1.221.13a
1.16
1.091.381.281.341.441.251.311.31a
0.50UF 108641 (type)UF 108641 (type)UF 108641 (type)UF 108644UF 121143UF 121143UF 108645
m1m2m3m1m1m2m3
1.251.191.191.251.221.191.06
0.750.780.720.730.690.690.59
0.780.780.630.750.750.720.56
a Estimated measurement of a broken tooth.
in the fossil species. Both the paraconid andmetaconid are very small in Nyctiellus com-pared with other extant natalids and Pri-monatalus. Overall, the lower molars ofPrimonatalus are most similar to those ofNatalus.
One of the paratype dentaries of Primo-natalus prattae (UF 108642) possesses thep3 and the alveoli for p2 and p4 (Figs. 3Aand 3B), providing additional informationon the dental morphology not evident in thetype specimen. Like all living natalids, P.prattae has a single-rooted p2 and a double-rooted p3 and p4. The p3 of Primonatalusis relatively larger than the p3 of Natalus,even though the species of Natalus are larg-er than the fossil in most other dimensions.The p3 of the fossil species is comparative-ly longer and more laterally compressedthan in any living natalid. The p3 in thefossil possesses a distinct concavity be-tween 2 vertical crests posterior to the pri-mary cusp. This concavity is present inChilonatalus, but absent in Natalus, inwhich the p3 is symmetrical anterior andposterior to the main cusp with only 1 crestdescending posterior to the main cusp. Thep3 is comparatively shorter and broader in
Nyctiellus than in Primonatalus, but it hasa distinct concavity between 2 crests pos-terior to the main cusp.
A 2nd paratype dentary of Primonatalusprattae (UF 108647) is edentulous but con-tains the alveoli for the lower incisors, ca-nine, and p2–4 (Figs. 2F–H). This speci-men has a large mental foramen located ina deep concavity positioned high on thedentary between the roots of c and p2. InNatalus and Chilonatalus, the region be-tween the alveoli of c and p2 is not as con-cave as in the fossil, and the mental fora-men is more ventrally located, about half-way between the alveolar margin of thetoothrow and the ventral margin of the den-tary and ventral to the alveoli of both c andp2. Nyctiellus has a smaller mental foramenin a shallower concavity. The anterior men-tal foramen is comparatively large in Pri-monatalus and is located at the mandibularsymphysis, immediately ventral to i1 andextending posteriorly to the level of i2. Theanterior mental foramen is very small andinconspicuous in Natalus and Chilonatalusand extends posteriorly only to the middleof i1. The anterior mental foramen is welldeveloped in Nyctiellus and extends poste-
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

740 Vol. 84, No. 2JOURNAL OF MAMMALOGY
riorly to i2. The mandibular symphysis inthe fossil is a short, inclined oval with anearly flat articular surface. The ventraledge of the symphysis of Primonatalus pro-jects slightly below the lower margin of thehorizontal ramus. The symphysis in Chilon-atalus and most Natalus (except N. stra-mineus) has a narrower, more elongate ovaloutline and projects much more posteroven-trally than in Primonatalus, Nyctiellus, andNatalus stramineus. This process is smallin Nyctiellus, like the fossil. The fossil den-tary possesses alveoli for 3 lower incisors;the first 2 are coalesced and are separatedfrom the 3rd by a tiny ridge. This situationmatches that in Chilonatalus, in which i1and i2 are close to one another but are sep-arated from i3 by a minute diastema. InNyctiellus and Natalus the lower incisorsare equally spaced.
A partial left maxilla with M1 and M2(UF 108638) is the only portion of the skullof Primonatalus preserved (Figs. 4B–D).This specimen retains the anteriormost re-gion of the zygomatic arch and the anteriorand ventral margins of the orbit, as well asthe posterior opening for the infraorbital fo-ramen. The zygomatic arch is flared some-what laterally in the fossil and the anterioredge of the muscle scar for the massetermuscle is located at the level of the poste-rior root of the M3. The anterior edge ofthe orbit in Primonatalus is located abovethe mesostyle of M2 and is nearly vertical,inclined only slightly anteriorly. The por-tion of the maxilla between the ventral edgeof the orbit and the alveolar margin of thetoothrow above M1–3 is relatively deep inthe fossil. Primonatalus possesses a deep,funnel-shaped fossa at the anterior edge ofthe orbit that incorporates both the lachry-mal foramen and the posterior opening ofthe infraorbital foramen. The dorsal lach-rymal foramen and ventral infraorbital fo-ramen are both large in Primonatalus andare separated only by a thin splint of bone.The anterior orbital fossa of Natalus (ex-cluding N. tumidirostris) is similar in mor-phology to that of the fossil, being a deep
funnel-shaped cavity with large lachrymaland infraorbital foramina separated by athin ridge of bone. The overall size of theanterior orbital fossa and the 2 foramina arereduced in N. tumidirostris because of theinflation of the rostrum on the medial wallof the orbit. In Chilonatalus, the anteriororbital fossa is reduced in size comparedwith the fossil and most species of Natalus.The posterior opening of the infraorbital fo-ramen in Chilonatalus is large and roundand widely separated from the small dorsallachrymal foramen. The anterior orbital fos-sa and the openings of both foramina lo-cated within it are much reduced in Nyc-tiellus owing to the greatly inflated rostrum.The anterior edge of the orbit in Chilona-talus is inclined slightly anteriorly as in thefossil, and the maxilla below the orbits isbroader anteriorly and thinner posteriorlycompared with Primonatalus. The anteriorportion of the zygomatic arch above the M3is considerably thinner in Chilonatalus thanin the fossil. The anterior base of the zy-gomatic arch begins at the anterior edge ofM3 in Chilonatalus but posterior to the M3in the fossil. In Natalus, the anterior edgeof the orbit is vertical, the anterior portionof the zygomatic arch is thinner and moreparallel-sided than in Primonatalus, and themasseteric muscle scar extends anteriorly toabout the middle of M3. In Nyctiellus, theanterior edge of the orbit is oriented atabout a 458 angle to the toothrow, the max-illa below the orbit is very deep, and themasseteric muscle scar extends to the mid-dle of the M3. The anterior process of thezygomatic arch is more robust in Nyctiellusthan in any other natalid.
The upper molars of Primonatalus prat-tae are similar to those of living membersof the Natalidae. Apparently, dental mor-phology of both the upper and lower molarshas remained persistently conservative inthis family because only minor changes areobserved to have occurred since the earlyMiocene. The M1 and M2 of Primonatalushave a narrower trigon basin than in Recentnatalids owing to the reduced talon and
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 741
metaconule in the terminology of Legendre(1984). An ‘‘incipient hypocone’’ appearsas a small cusp on the cingulum of the tal-on. This cusp is present on M1, but not onM2, in Primonatalus, Natalus major, andN. stramineus but not in N. tumidirostris.The M1 talon lacks the ‘‘incipient hypo-cone’’ in Nyctiellus, Chilonatalus, and N.tumidirostris. The talon is very small onM1 and virtually absent on M2. The par-astyle is reduced on M2, and the protoconeis located closer to the paracone on bothM1 and M2. The lingual cingulum is re-duced in the fossil teeth, especially ventralto the protocone. In Natalus, M1 is squar-ish, and M2 is more rectangular in occlusaloutline. Both upper molars in Natalus aresquared off lingually because of the broadtrigon basin and the well-developed talon.Both teeth are nearly as broad lingually aslabially. In Primonatalus, the 2 upper mo-lars are noticeably narrower lingually thanlabially. The M1 is broader lingually thanthe M2 owing to the slight development ofa talon, whereas M2 is essentially triangu-lar. In Chilonatalus, M2 is more similar tothe fossil, with a very weak to nearly absenttalon. The parastyle and talon are both pre-sent on M2 in Nyctiellus but are very small.The M1 and M2 are both narrow antero-posteriorly in Nyctiellus as in the fossilsowing to the weakly developed talon. InPrimonatalus and Nyctiellus there is noconnection between the posterior end of thepostprotocrista and the metacingulum inM1 and M2. In Chilonatalus and Natalusthere is a weak-to-moderate connection ofthese 2 structures. The lingual cingulum inM1 and M2 is complete in Natalus, inter-rupted around the base of the protocone inPrimonatalus and Chilonatalus, and morewidely interrupted in Nyctiellus.
A dental synapomorphy of natalids thatis absent in other extant and extinct nata-loids is the presence in M1 and M2 of asmall curved crest extending forward fromthe mesostyle into the anteroexternal valley(Fig. 4B). In M1 this crest is straighter andmore prominent and has a tendency to be
rather cuspate, often giving a twinned ap-pearance to the mesostyle. In M2 the crestis less prominent with no tendency to ap-pear cuspate; instead, it is more smoothlycurved inward toward the postparacrista. InPrimonatalus this mesostylar crest is irreg-ularly cristate in M1 and is abruptly ter-minated at its anterior end by a small cuspseparating it from an anterior remnant ofthe ectocingulum that extends backwardfrom the parastyle. Often a miniscule stylarcuspule appears at the posterior end of thisanterior ectocingulum, across the small gapfrom the mesostylar crest. In M2 of Pri-monatalus the mesostylar crest is moresmoothly cristate and more strongly curvedinward (lingually). A small posterior rem-nant of the ectocingulum occurs in both M1and M2 of Primonatalus, extending for-ward from the metastyle halfway across theposteroexternal valley and terminating in avery small stylar cuspule. Attributes of themesostylar crest and stylar shelf of M1 andM2 in Chilonatalus and Natalus are similarto Primonatalus. Nyctiellus differs from theother genera in having mesostylar creststhat are very small and weak and in lackingthe remnants of the ectocingulum posteriorto the parastyle and anterior to the meta-style. M3 has a weak mesostylar crest inPrimonatalus and Chilonatalus. This ridgeis absent in M3 in Nyctiellus and Natalus.
Another dental synapomorphy uniting allNatalidae is the presence of a cingular (or‘‘stylar’’) cusp labial to the primary cusp onP4 (Figs. 4E and 4F). This cusp is absentin the extinct nataloids Honrovits, Chad-ronycteris, and Chamtwaria. Stehlinia hasP4 with a strong rise in the labial cingulum,but no cingular cusp occurs in that genuseither. In Primonatalus the tip of the cin-gular cusp is broken in the sole availablespecimen of P4 (UF 121136), but the cuspappears to have been low. In extant generathe labial cingular cusp is low in Nyctiellus,intermediate in height in Natalus, and high-est in Chilonatalus. The P4 in all natalidsis 3-rooted. The P4 of Primonatalus is dis-tinct from that of extant natalids in having
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

742 Vol. 84, No. 2JOURNAL OF MAMMALOGY
a narrow anterolabial projection in occlusalview; the portion of P4 supported by theanterolabial root is separated from the lin-gual portion of the tooth by an indentationin the cingulum. The lingual moiety of P4(‘‘talon’’) is broadly rounded in Primona-talus. In the extant natalids there is no an-terolingual indentation, and the anterolabialcorner of the P4 is rather broadly quadrate.The overall outline of P4 in occlusal viewis roughly squarish in the labial half andbroadly rounded in the lingual half in Chi-lonatalus and Natalus. Nyctiellus is distinctfrom the others in being relatively shorteranteroposteriorly and in having a narrowerand more pointed lingual half, giving thetooth a roughly triangular occlusal outline.In Chilonatalus the rounded lingual portiondoes not extend so far lingually as in Pri-monatalus so that the tooth is transverselynarrower.
Three periotics of Primonatalus (Figs.5A–D) are virtually identical to the periot-ics of modern natalids. All are broken, but1 of the fossil periotics preserves 2 of thesemicircular canals. The cochlea of Pri-monatalus makes about 2¼ turns. The co-chlea of Natalus stramineus appears tomake about 2¼ turns, but in all other livingspecies of natalids the cochlea makes about2½ turns. A small tubercle arises from thesmooth, rounded posteroventral surface ofthe cochlea of all natalids. This small tu-bercle is situated just ventral to the fenestracochleae. It bears a tiny hooklike process atits lateral end that partly encircles the in-ternal carotid artery in life and contacts partof the posterior end of the stylohyal. Thetubercle and hook are transversely short inPrimonatalus. The transverse width of thistubercle varies in Natalus; in N. stramineusit is short and very similar to that in Pri-monatalus, in N. tumidirostris it is longerand rather more flattened on its posteroven-tral surface, and in N. major it is still longerand flattened. In Chilonatalus the tubercleforms a taller, larger hook. In Nyctiellus thetubercle is transversely wide and distinctivein possessing a medial pointed projection as
well as the lateral hook. The shallowgrooves along which the internal carotid ar-tery and stapedial artery run are apparent inthe fossil periotics and follow paths acrossthe cochlear surface that are identical to thepaths in the modern natalids.
Five distal ends of the humerus referableto the Natalidae are known from ThomasFarm, but no proximal humeri are presentin the sample. The fossil humeri are readilyidentified as natalids and separated fromthose of vespertilionids by the laterally off-set distal articular surface and the broad, tri-angular-shaped epitrochlea (5medial epi-condyle; Fig. 6). The following range ofmeasurements (mm) were made of the dis-tal humeri of Primonatalus prattae fromThomas Farm (UF 108648–108652): max-imum distal width 2.8–2.9 (n 5 4); widthof distal articular surface 1.9–2.0 (n 5 4);and shaft diameter 1.0–1.1 (n 5 4). Thedistal humeri of P. prattae have a broadtriangular-shaped epitrochlea that has nonotch or concavity between the spinousprocess of the epitrochlea (5distal spinousprocess) and the medial process of the epi-trochlea. The medial process is broad andblunt in the fossil humeri and is barely sep-arable from the large epitrochlea of whichit is a part. In Natalus the medial processis better developed, primarily resultingfrom the stronger notch located on the me-dial margin of the epitrochlea halfway be-tween the medial process and the spinousprocess. The medial process is better de-veloped in Chilonatalus, in which it isprominent and elongated. The medial pro-cess is separated from the spinous processby a deep rounded notch in the epitrochleathat forms nearly a right angle. The entireepitrochlea is greatly reduced in Nyctiellus,and the medial process is absent. The spi-nous process is small in Primonatalus, pro-jects slightly laterally, and is somewhatpointed distally. In the fossils this processprojects distally only to the level of the dis-tal edge of the trochlea or slightly distal toit in several specimens. The spinous processis separated from the medial edge of the
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 743
trochlea by a narrow but rather deep notch.A small spinous process is also character-istic of Natalus, but this process is con-nected to the trochlea by a ridge, and it thuslacks the distinct notch found in the fossilhumeri. Because this notch is lacking inNatalus, the spinous process does not ap-pear to be as prominent as in Primonatalus,even though it projects the same distancedistally in both genera. The spinous processis very well developed in Chilonatalus, ex-tending noticeably distal to the medial edgeof the trochlea and separated from it by adeep notch. The spinous process of Chilo-natalus also bears a sharp laterally project-ing process. Nyctiellus has a very large spi-nous process that extends far distal to thearticular surface. In characters of the spi-nous process, Primonatalus is intermediatebetween Natalus, in which this process issmall, short, blunt distally, and connectedto the medial edge of the trochlea, and Chi-lonatalus, in which the process is very largeand broad, extends well distal to the artic-ular surface, projects noticeably laterally,and is separated from the medial edge ofthe trochlea by a deep notch.
In anterior aspect, the medial ridge of thecapitulum is bulbous and almost sphericalin Primonatalus and Natalus. In these 2genera, this rounded portion of the capitu-lum is separated from the lateral ridge ofthe capitulum and the trochlea by relativelydeep, broadly rounded grooves. The medialridge of the capitulum is more flattened inChilonatalus, and the grooves separating itfrom the trochlea and lateral capitulum areshallow. The medial ridge of the capitulumis reduced in Nyctiellus, and the entire distalarticular surface is relatively narrower com-pared with all other natalids.
In posterior aspect, the proximal marginof the lateral ridge of the capitulum is flat-tened in the fossil, Chilonatalus, and Nyc-tiellus and is separated from the medial ca-pitulum by a shallow groove. In Natalus thedorsal surface of the lateral capitulum is ex-panded into a large rounded process; con-sequently, there is a deep rounded notch be-
tween it and the medial surface of the ca-pitulum. The medial edge of the trochlea onthe posterior surface is a well-developedridge extending proximally to the flaredpart of the shaft in the fossils and Natalus.This ridge is weakly developed in Chilo-natalus and absent in Nyctiellus. In lateralview, the articular surface of the lateral ca-pitulum of the Primonatalus humeri is nar-row anteroposteriorly and is flattened dis-tally, and the lateral surface of the capitu-lum has a deep elliptically shaped concavitythat opens proximally. In other natalids, thearticular surface of the lateral capitulum ismore rounded, and the lateral concavity ispresent but shallower.
Two proximal radii from Thomas Farmare very similar in size and morphology tothe proximal radius of extant natalids. Theonly difference is that the process extendingproximal to the articular surface for the hu-merus is more rounded in the fossils. Thisproximal process is more pointed and tri-angular-shaped in living natalids. Despitethe slight difference in the proximal pro-cess, the fossil radii are more similar to theradius of natalids than to that of any otherbat present at Thomas Farm. Four distal ra-dii from Thomas Farm are essentially iden-tical to the distal radius of living natalids.As with most other elements of the ThomasFarm natalid, the fossil radii are most sim-ilar in size to the radius of Chilonatalustumidifrons.
The proximal end of a chiropteran radius(UF 121723) from the Oligocene I-75 LocalFauna is very similar to the 2 proximal radiifrom Thomas Farm in its morphology,small size, and delicate shaft. The tip of theproximal process is slightly damaged in theI-75 fossil; thus, it is not possible to deter-mine whether this specimen differs fromthe living natalids in the same feature asdoes Primonatalus. Although diagnostic atthe family level, the proximal end of theradius is not useful for distinguishingamong species in the Natalidae, except forsize. The I-75 radius is referred to the Na-talidae, but the genus and species are in-
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

744 Vol. 84, No. 2JOURNAL OF MAMMALOGY
FIG. 7.—Potential relationships within thefamily Natalidae, based on an analysis of 50morphological characters of the teeth and skel-eton. Topology represents the single most par-simonious tree from a branch-and-bound search(tree length 82 steps; consistency index 50.7439; retention index 5 0.6769). Numbersabove branches are decay values (Bremer sup-port values); numbers below branches are boot-strap values representing the percentage of10,000 bootstrap replicates in which each cladeappeared.
determinate. The identification of this spec-imen as a member of the Natalidae is sig-nificant because it represents the oldestknown record for the family, some 12 mil-lion years older than Primonatalus fromThomas Farm.
The distal end of a femur of Primona-talus from Thomas Farm is similar in shaftdiameter and distal articular width to thefemora of Chilonatalus micropus and Na-talus stramineus. No characters distinctiveat the genus or species level were found inthe fossil or in the distal femur in livingnatalids. The relative length of the femurvaries significantly among extant natalids,but it is not possible to determine length ofthe femur from the incomplete fossil. Thefemur is longer than the humerus (about105%) in Chilonatalus tumidifrons, slightlyshorter than the humerus (about 90–95%)in C. micropus and Natalus, and muchshorter than the humerus (about 80%) inNyctiellus. The variation in relative lengthof the humerus and femur among living na-talids indicates differences in size and, pos-sibly, function of the hind limb and tailmembrane.
PHYLOGENETIC ANALYSIS
We find strong support for the monophy-ly of the family Natalidae (sensu stricto),including Primonatalus prattae (Fig. 7).Among the extant taxa, the 3 species of Na-talus are monophyletic, and this clade isstrongly supported. Within this clade, Na-talus stramineus and Natalus tumidirostrisare sister taxa (not surprising, because the2 species are difficult to tell apart in handand are very similar in skeletal morpholo-gy), with strong support for the inclusion ofNatalus major. The 2 species of Chilona-talus are monophyletic, supporting the no-tion that Chilonatalus is distinct from Na-talus. Chilonatalus and Natalus cluster to-gether, but support for this clade is moder-ate. There is moderate support fordistinction of the traditional subgenera Nyc-tiellus, Chilonatalus, and Natalus. The fos-sil Primonatalus is the most primitive
member of the family and is distinct fromthe other natalids, but its membership in thefamily is strongly supported. These resultsare remarkably concordant with aspects ofthe traditional classification scheme for theliving natalids and suggest that there maybe a phylogenetic signal in the dental–cra-nial–postcranial morphology in this family.
All living members of the Natalidae typ-ically are placed in the genus Natalus(Koopman 1993). We suggest, however,that the 6 extant species should be separatedinto 3 genera, Natalus (including the spe-cies N. major, N. stramineus, and N. tumi-dirostris), Chilonatalus (including the spe-cies C. micropus and C. tumidifrons), andNyctiellus (including the species N. lepi-dus), each previously recognized as a sub-genus of Natalus (Dalquest 1950; Goodwin1959; Koopman 1993). Most workers con-sider N. major to be a junior synonym ofN. stramineus (Koopman 1993; Silva Ta-boada 1979; Varona 1974), but Morgan(1989b) noted several morphological differ-ences between the 2 species that he thoughtjustified recognition of N. major as a dis-tinct species, including large size (there isno overlap in measurements with the small-er N. stramineus); more inflated braincase
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 745
that meets the rostrum at a steeper angle;more strongly constricted interorbital re-gion; stronger sagittal crest; and less inflat-ed rostrum that is more tapered anteriorly,constricting the size of the narial opening.
Among the 3 living genera, Nyctiellus isthe most distinct. We recognize Nyctiellusas a separate genus on the basis of numer-ous cranial and dental characters that differsubstantially from other natalids (characternumbers [character states] from AppendixII), including small size; broader, deeper,and more inflated rostrum (character31 [state 1]); lower, less inflated braincase(25 [1]) and less dorsally upturned braincase(26 [0]); deeper, more robust zygomatic arch(34 [1]); deep midline (unpaired) basisphe-noidal pit (36 [2]); inflated auditory bullae(37 [1]); cleft between premaxillae reducedso that I1s nearly meet at midline (35 [1]);reduced upper canines (27 [1]); and greatlyreduced P2 (29 [1]) and p2 (50 [1]). Chilon-atalus is distinguished by having P4 with astrong labial cingular cusp (14 [2]) and ahighly derived degree of fusion in the axialskeleton, with only 1 free (unfused) poste-rior lumbar vertebrae (42 [2]) and ribs thatare greatly coalesced with each other andwith the sternum (44 [2]). Such axial fusionis found in varying degrees in several fam-ilies of bats and is probably related to theenergetics of echolocation (Lancaster et al.1995, 2001). Natalus sensu stricto is distin-guished by having a straight angular process(4 [0]) that flares strongly outward (5 [2])and a distal spinous process on the humerusthat is not separated from the medial edgeof the trochlea by a notch (23 [1]).
The fossil material of Primonatalus prat-tae is not nearly so complete as that avail-able for the 6 living species of natalids. Al-though many of the derived characters ofthe Natalidae are unknown in Primonatal-us, enough of the cranial and dental anat-omy of Primonatalus is preserved to con-firm that this taxon is unquestionably refer-able to the Natalidae. The combination ofthe derived character states listed belowunite all members of the Natalidae and dis-
tinguish them from other chiropteran fami-lies.
The derived features of the Natalidae areas follows (characters in our data set areidentified by their character number in pa-rentheses followed by the character state inbrackets, from Appendix II): elongated,dorsoventrally flattened rostrum; reducedorbit with anterior edge located far poste-rior above M2; deep funnel-shaped fossa atanterior edge of orbit enclosing lachrymalforamen and posterior opening of infraor-bital foramen (39[1]); greatly elongated in-fraorbital canal originating above M2 andopening anteriorly above P3 (38[1]); elon-gated styliform process of ectotympanicthat extends anteromedially and connectswith pterygoid process (33[1]); presence onP4 of cingular or stylar cusp labial to pri-mary cusp (14[1]); presence on M1 and M2of small curved crest extending anteriorlyfrom mesostyle into anteroexternal valley(17[1]); manubrium (presternum) verybroad with strong posteriorly oriented me-dian keel (43[1]); last thoracic and all lum-bar vertebrae except the last 1 or 2 fusedinto rigid, laterally compressed structurewith prominent dorsal and ventral ridges(41[1]); exceptionally elongated caudal ver-tebrae 3 through 6; femur and tibia ex-tremely elongated and slender; proximalend of femur strongly bent laterally (46[1]);proximal end of femur with greater andlesser trochanters reduced (48[1]); and pres-ence of natalid organ, a subcutaneousglandlike organ on forehead and muzzle ofmales. Simmons (1998) listed the presenceof the natalid organ as an apomorphy di-agnosing the Natalidae.
DISCUSSION
Five extinct genera have been referred tothe Natalidae, including Ageina from theearly Eocene of Europe (Russell et al.1973), Stehlinia from the Eocene and Oli-gocene of Europe (Sige 1974), Honrovitsfrom the early Eocene of Wyoming (Beardet al. 1992), Chadronycteris from the lateEocene of Nebraska (Ostrander 1983), and
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

746 Vol. 84, No. 2JOURNAL OF MAMMALOGY
Chamtwaria from the Miocene of Africa(Butler 1984). On considering these 5 ex-tinct genera to be members of the Natalidae,Beard et al. (1992) followed an expandeddefinition of the family proposed by VanValen (1979), which included, in additionto the Natalidae sensu stricto, the Furipter-idae, Thyropteridae, Myzopodidae, andKerivoulidae as subfamilies. Excluding theKerivoulidae, this is similar to the conceptof the superfamily Nataloidea proposed bySimmons (1998) and Simmons and Geisler(1998), consisting of the families Natalidae,Furipteridae, Thyropteridae, and Myzopod-idae. These authors considered the Kerivou-lidae to be a subfamily of the Vespertilion-idae. McKenna and Bell (1997) placedHonrovits in the Natalidae sensu stricto,Stehlinia and Chamtwaria in the Vesperti-lionidae, and Ageina and Chadronycteris inMicrochiroptera incertae sedis. Simmonsand Geisler (1998) regarded Ageina, Chad-ronycteris, Honrovits, and Stehlinia as in-certae sedis within the Nataloidea (theyconsidered only Eocene forms and thus didnot mention the Miocene Chamtwaria). All5 extinct Tertiary genera referred to the Na-talidae or Nataloidea by previous authors,including Ageina, Chadronycteris, Cham-twaria, Honrovits, and Stehlinia, fall out-side the strict definition of the Natalidae, asrecognized in this study (i.e., they lack thederived features of the Natalidae listedabove). Only the 6 extant species previous-ly placed in the genus Natalus and the Mio-cene Primonatalus, described in this study,belong to the Natalidae sensu stricto.
Many authors have discussed the closerelationship between the Natalidae, Furip-teridae, and Thyropteridae (Dalquest 1950;Miller 1899, 1907; Simmons and Geisler1998; Slaughter 1970). Miller (1899) orig-inally placed the Furipteridae and Thyrop-teridae within the Natalidae, but in his clas-sic study of the families and genera of bats,Miller (1907) assigned the genera Furipte-rus and Amorphochilus to the Furipteridaeand Thyroptera to the Thyropteridae. Mill-er’s (1907) arrangement of these 3 families
has been followed by most subsequent chi-ropteran systematists. An analysis of chi-ropteran dentitions supported Miller’s sug-gestion of a close relationship between theNatalidae, Furipteridae, and Thyropteridae(Slaughter 1970). Van Valen (1979) ex-panded the concept of the Natalidae by in-cluding not only the Furipteridae and Thy-ropteridae but also the Myzopodidae andKerivoulidae as subfamilies. Van Valen’s(1979) expanded concept of the Natalidaewas supposedly based on shared derivedcharacters; however, the supporting data forhis phylogenetic hypothesis were not pro-vided, and most subsequent authors havenot followed his arrangement. Recently,Van Den Bussche and Hoofer (2001) pro-vided molecular evidence that Nataloidea isnot a monophyletic lineage. Their mito-chondrial DNA sequence data suggested in-stead that Myzopodidae represents a basalmicrochiropteran lineage and that Furipter-idae, Thyropteridae, and Natalidae are suc-cessive out-groups to the Noctilionoidea.However, subsequent to that analysis, Hoof-er et al. (in press) extended their analysisby examining DNA sequences from a nu-clear gene in all families of Yangochirop-tera. Their goal specifically was to reconcilecontroversies surrounding 5 families whosephylogenetic positions have been unstable(Furipteridae, Mystacinidae, Myzopodidae,Natalidae, and Thyropteridae). On combin-ing their new nuclear sequence data withthe previous mitochondrial data, Hoofer etal. (in press) found support for an associa-tion of Natalidae with Vespertilionidae andMolossidae in the Vespertilionoidea (butFuripteridae and Thyropteridae remainedwith Noctilionoidea). Accordingly, andbased on our results, we adhere to the con-ventional arrangement of the Natalidae, in-cluding only 3 living genera (or subgenera)Natalus, Chilonatalus, and Nyctiellus, alongwith the extinct Miocene genus Primona-talus.
The presence of natalids in the middleCenozoic of Florida provides importantnew information pertaining to the evolu-
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 747
tionary history of this small family of bats.Before the discovery of Primonatalus inFlorida, the Natalidae sensu stricto had nopre–late Pleistocene fossil record. The fos-sils from the Oligocene I-75 Local Faunaand the early Miocene Thomas Farm LocalFauna extend the record of the Natalidaeback nearly 30 million years in NorthAmerica. Furthermore, the Florida fossilsoccur in what is now the Nearctic region,whereas no species in the Natalidae are cur-rently found outside the Neotropical region.Late Pleistocene fossils representing extantspecies of Natalidae are known from manyislands in the West Indies (Morgan 2001)and from Brazil (Czaplewski and Cartelle1998). Tertiary fossils of natalids are un-known from South America, although thefossil record of bats on this continent isvery sparse (Czaplewski 1997). Of the 2other Neotropical families supposedlyclosely related to the Natalidae, the Furip-teridae occur in Pleistocene deposits in Bra-zil (Czaplewski and Cartelle 1998) andPeru (Morgan and Czaplewski 1999) andthe Thyropteridae are known from the mid-dle Miocene of Colombia (Czaplewski1997).
The Natalidae are currently restricted tothe Neotropical region, although severalspecies occur at the northern limits of theNeotropics in northern Mexico, the Baha-mas, and Cuba. Natalus stramineus isknown from the states of Sonora and NuevoLeon in northern Mexico (Hall 1981), Chi-lonatalus tumidifrons occurs on Abaco andAndros in the Bahamas, and C. micropusand Nyctiellus lepidus are found in Cuba(Morgan 2001). The family attains its great-est species richness in the West Indies,where all 3 genera and 6 species of the fam-ily occur. Two of the genera (Chilonatalusand Nyctiellus) and 4 of the species (Na-talus major from Jamaica and Hispaniola;Chilonatalus micropus from Cuba, Jamaica,and Hispaniola; C. tumidifrons from the Ba-hamas; and Nyctiellus lepidus from Cubaand the Bahamas) are endemic to the WestIndian subregion of the Neotropical region.
Natalids are widespread throughout tropicalMiddle America (Hall 1981), but they havea restricted distribution in South America,where they occur primarily in the Carib-bean and Atlantic coastal regions in thenorthern and northeastern portions of thecontinent (Koopman 1982). Natalus stra-mineus is the most widely distributed spe-cies in the family, occurring in the mainlandNeotropics from northern Mexico, through-out Middle America, and as far south asBrazil, with a disjunct population in thenorthern Lesser Antilles (Hall 1981; Koop-man 1982, 1989). Natalus tumidirostris isfound in northern Colombia and Venezuelaand on the southern Caribbean islands ofTrinidad, Margarita, and Curacao (Koop-man 1982). Although these islands are lo-cated in the West Indies, they were exclud-ed from the West Indian subregion byKoopman (1958, 1989) because they lackendemic species of Antillean bats.
All living natalids are obligate cavedwellers. In the West Indies, most speciesin this family roost in chambers deep withinlarge caves that are characterized by a sta-ble microenvironment with high tempera-ture and relative humidity (Goodwin 1970;Silva Taboada 1979). The roosting ecologyof natalids suggests that their present (andpresumably past) distribution is limited bythe availability of extensive cave systems(Morgan 2001).
The Tertiary fossil record of the Natali-dae is limited to the early Oligocene andearly Miocene of Florida. The family pre-sumably survived in tropical North Ameri-ca (i.e., Middle America) between the earlyMiocene and the late Pleistocene, althoughnatalids have no fossil record during thattime period. The presence in the West In-dies of 2 endemic genera and 4 endemicspecies of natalids suggests this group prob-ably reached the Antilles by overwater dis-persal from tropical North America early intheir evolutionary history, presumably inthe Oligocene or Miocene. Conversely, inSouth America, the Natalidae have a mar-ginal distribution, limited species richness
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

748 Vol. 84, No. 2JOURNAL OF MAMMALOGY
(2 species of Natalus, 1 of which is endem-ic), and lack a Tertiary fossil record. Thesefactors suggest that natalids may not haveentered South America until comparativelylate in time, perhaps in the Pliocene afterthe formation of the Panamanian Isthmusand the beginning of the Great AmericanFaunal Interchange.
ACKNOWLEDGMENTS
We thank A. E. Pratt of Georgia SouthernUniversity and A. R. Poyer of the Florida Mu-seum of Natural History for their help in thefield and with screenwashing and sorting matrix.All the Miocene natalid fossils described in thisarticle were recovered through their efforts. Wethank S. D. Webb and B. J. MacFadden of theFlorida Museum of Natural History for loansand permission to study the Thomas Farm andI-75 bats. For access to Recent comparativespecimens, we thank L. Wilkins of the FloridaMuseum of Natural History, L. K. Gordon of theNational Museum of Natural History, Washing-ton, D.C., N. B. Simmons of the American Mu-seum of Natural History, New York, and W. L.Gannon of the Museum of Southwestern Biol-ogy, University of New Mexico, Albuquerque.We greatly appreciate the advice of R. L. Nydamand N. B. Simmons on the parsimony analyses.N. B. Simmons and an anonymous reviewer pro-vided helpful comments on the manuscript.Funding for this study was provided by a grantfrom the National Science Foundation (DEB9981512) to N. J. Czaplewski and G. S. Morgan.
LITERATURE CITED
AUFFENBERG, W. 1963. The fossil snakes of Florida.Tulane Studies in Zoology 10:131–216.
BEARD, K. C., B. SIGE, AND L. KRISHTALKA. 1992. Aprimitive vespertilionoid bat from the early Eoceneof central Wyoming. Comptes Rendus des Seancesde l’Academie des Sciences Paris 314:735–741.
BLACK, C. C. 1963. Miocene rodents from the ThomasFarm Local Fauna, Florida. Bulletin of the Museumof Comparative Zoology 128:483–501.
BRODKORB, P. 1956. Two new birds from the Mioceneof Florida. Condor 58:367–370.
BUTLER, P. M. 1984. Macroscelidea, Insectivora, andChiroptera from the Miocene of East Africa. Pa-laeovertebrata 14:117–200.
CZAPLEWSKI, N. J. 1997. Chiroptera. Pp. 410–431 inVertebrate paleontology in the Neotropics: the Mio-cene fauna of La Venta, Colombia (R. F. Kay, R. H.Madden, R. L. Cifelli, and J. J. Flynn, eds.). Smith-sonian Institution Press, Washington, D.C.
CZAPLEWSKI, N. J., AND C. CARTELLE. 1998. Pleisto-
cene bats from cave deposits in Bahia, Brazil. Jour-nal of Mammalogy 79:784–803.
CZAPLEWSKI, N. J., AND G. S. MORGAN. 2000. A newvespertilionid bat (Mammalia: Chiroptera) from theearly Miocene (Hemingfordian) of Florida, USA.Journal of Vertebrate Paleontology 20:736–742.
DALQUEST, W. W. 1950. The genera of the chiropteranfamily Natalidae. Journal of Mammalogy 31:436–443.
EMRY, R. J., P. R. BJORK, AND L. S. RUSSELL. 1987.The Chadronian, Orellan, and Whitneyan NorthAmerican land mammal ages. Pp. 118–152 in Ce-nozoic mammals of North America: geochronologyand biostratigraphy (M. O. Woodburne, ed.). Uni-versity of California Press, Berkeley.
ESTES, R. 1963. Early Miocene salamanders and liz-ards from Florida. Quarterly Journal of the FloridaAcademy of Sciences 26:234–256.
FORSTEN, A. 1975. The fossil horses of the Texas GulfCoastal Plain: a revision. Texas Memorial Museum,Pearce-Sellards Series 22:1–86.
GOODWIN, G. G. 1959. Bats of the subgenus Natalus.American Museum Novitates 1977:1–22.
GOODWIN, R. E. 1970. The ecology of Jamaican bats.Journal of Mammalogy 51:571–579.
HALL, E. R. 1981. The mammals of North America.2nd ed. John Wiley & Sons, Inc., New York 1:1–600 1 90.
HAYES, F. G. 2000. The Brooksville 2 Local Fauna(Arikareean, latest Oligocene): Hernando County,Florida. Bulletin of the Florida Museum of NaturalHistory 43:1–47.
HOLMAN, J. A. 1965. Early Miocene anurans fromFlorida. Quarterly Journal of the Florida Academyof Sciences 28:68–82.
HOLMAN, J. A. 1967. Additional Miocene anurans fromFlorida. Quarterly Journal of the Florida Academyof Sciences 30:121–140.
HOLMAN, J. A. 1999. Early Oligocene (Whitneyan)snakes from Florida (USA), the second oldest col-ubrid snakes in North America. Acta Zoologica Cra-coviensia 42:447–454.
HOOFER, S. R., S. A. REEDER, E. W. HANSEN, AND R.A. VAN DEN BUSSCHE. In press. Molecular phylo-genetics and taxonomic review of noctilionoid andvespertilionoid bats (Chiroptera: Yangochiroptera).Journal of Mammalogy.
HORACEK, I. In press. On the early history of vesper-tilionid bats in Europe: the lower Miocene recordfrom the Bohemian Massif. Myotis.
HULBERT, R. C., JR. 1984. Paleoecology and popula-tion dynamics of the early Miocene (Hemingfordian)horse Parahippus leonensis from the Thomas FarmSite, Florida. Journal of Vertebrate Paleontology 4:547–558.
KOOPMAN, K. F. 1958. Land bridges and ecology in batdistribution on islands off the northern coast ofSouth America. Evolution 12:429–439.
KOOPMAN, K. F. 1982. Biogeography of the bats ofSouth America. Pp. 273–302 in Mammalian biologyin South America (M. A. Mares and H. H. Geno-ways, eds.). Special Publication Series, PymatuningLaboratory of Ecology, University of Pittsburgh,Pittsburgh, Pennsylvania 6:1–539.
KOOPMAN, K. F. 1989. A review and analysis of the
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 749
bats of the West Indies. Pp. 635–644 in Biogeogra-phy of the West Indies: past, present, and future (C.A. Woods, ed.). Sandhill Crane Press, Gainesville,Florida.
KOOPMAN, K. F. 1993. Order Chiroptera. Pp. 137–241in Mammal species of the world: a taxonomic andgeographic reference. 2nd ed. (D. E. Wilson and D.M. Reeder, eds.). Smithsonian Institution Press,Washington, D.C.
KOOPMAN, K. F. 1994. Chiroptera: systematics. Hand-book of zoology: Mammalia. Walter de Gruyter,New York 8(60):1–217.
LANCASTER, W. C., M. B. FENTON, AND J. EGER. 2001.Morphology of the axial skeleton in relation to thestyle of biosonar. Bat Research News 41:25.
LANCASTER, W. C., O. W. HENSON, JR., AND A. W.KEATING. 1995. Respiratory muscle activity in rela-tion to vocalization in flying bats. Journal of Exper-imental Biology 198:175–191.
LAWRENCE, B. 1943. Miocene bat remains from Flor-ida, with notes on the generic characters of the hu-merus of bats. Journal of Mammalogy 24:356–369.
LEGENDRE, S. 1984. Etude odontologique des represen-tants actuels du groupe Tadarida (Chiroptera, Mo-lossidae). Implications phylogeniques, systema-tiques et zoogeographiques. Revue Suisse de Zool-ogie 91:399–442.
MAGLIO, V. J. 1966. A revision of the fossil selenodontartiodactyls from the middle Miocene Thomas Farm,Gilchrist County, Florida. Breviora, Museum ofComparative Zoology 255:1–27.
MCKENNA, M. C., AND S. K. BELL. 1997. Classificationof mammals above the species level. Columbia Uni-versity Press, New York.
MENU, H., AND B. SIGE. 1971. Nyctalodontie et my-otodontie, importants caracteres de grades evolutifschez les chiropteres entomophages. Comptes Rendusde Seances de l’Academie des Sciences 272:1735–1738.
MILLER, G. S., JR. 1899. History and characters of thefamily Natalidae. Bulletin of the American Museumof Natural History 12:245–253.
MILLER, G. S., JR. 1907. The families and genera ofbats. United States National Museum Bulletin 57:1–282.
MORGAN, G. S. 1989a. New bats from the Oligoceneand Miocene of Florida and the origins of the Neo-tropical chiropteran fauna. Journal of Vertebrate Pa-leontology 9(3, supplement):33A.
MORGAN, G. S. 1989b. Fossil Chiroptera and Rodentiafrom the Bahamas, and the historical biogeographyof the Bahamian mammal fauna. Pp. 685–740 inBiogeography of the West Indies: past, present, andfuture (C. A. Woods, ed.). Sandhill Crane Press,Gainesville, Florida.
MORGAN, G. S. 2001. Patterns of extinction in WestIndian bats. Pp. 369–407 in Biogeography of theWest Indies: patterns and perspectives. 2nd ed. (C.A. Woods and F. E. Sergile, eds.). CRC Press, BocaRaton, Florida.
MORGAN, G. S., AND N. J. CZAPLEWSKI. 1999. First fos-sil record of Amorphochilus schnablii (Chiroptera:Furipteridae), from the late Quaternary of Peru. ActaChiropterologica 1:75–79.
MORGAN, G. S., AND N. J. CZAPLEWSKI. 2000. A new
bat in the Neotropical family Natalidae from the ear-ly Miocene (Hemingfordian) Thomas Farm LocalFauna, Florida. Journal of Vertebrate Paleontology20(3, supplement):59A.
NOWAK, R. M. 1994. Walker’s bats of the world. JohnsHopkins University Press, Baltimore, Maryland.
OLSEN, S. J. 1962. The Thomas Farm fossil quarry.Quarterly Journal of the Florida Academy of Sci-ences 25:142–146.
OSTRANDER, G. E. 1983. New early Oligocene (Chad-ronian) mammals from the Raben Ranch Local Fau-na, northwest Nebraska. Journal of Paleontology 57:128–139.
OTTENWALDER, J. A., AND H. H. GENOWAYS. 1982. Sys-tematic review of the Antillean bats of the Natalusmicropus-complex (Chiroptera: Natalidae). Annalsof Carnegie Museum 51:17–38.
PATTON, T. H. 1969. An Oligocene land vertebrate fau-na from Florida. Journal of Paleontology 43:543–546.
PRATT, A. E. 1989. Taphonomy of the microvertebratefauna from the early Miocene Thomas Farm locality,Florida (U.S.A.). Palaeogeography, Palaeoclimatol-ogy, Palaeoecology 76:125–151.
PRATT, A. E. 1990. Taphonomy of the large vertebratefauna from the Thomas Farm locality (Miocene,Hemingfordian), Gilchrist County, Florida. Bulletinof the Florida Museum of Natural History, Biolog-ical Sciences 35:35–130.
PRATT, A. E., AND G. S. MORGAN. 1989. New Sciuridae(Mammalia: Rodentia) from the early MioceneThomas Farm local fauna, Florida. Journal of Ver-tebrate Paleontology 9:89–100.
RAY, C. E. 1957. A list, bibliography, and index of thefossil vertebrates of Florida. Florida Geological Sur-vey, Special Publication 3:1–175.
RUSSELL, D. E., P. LOUIS, AND D. E. SAVAGE. 1973.Chiroptera and Dermoptera of the French Early Eo-cene. University of California Publications in theGeological Sciences 95:1–55.
SIGE, B. 1974. Donees nouvelles sur le genre Stehlinia(Vespertilionoidea, Chiroptera) du Paleogened’Europe. Palaeovertebrata 6:253–272.
SILVA TABOADA, G. 1979. Los murcielagos de Cuba.Editorial Academia, La Habana, Cuba.
SIMMONS, N. B. 1998. A reappraisal of interfamilialrelationships of bats. Pp. 3–26 in Bat biology andconservation (T. H. Kunz and P. A. Racey, eds.).Smithsonian Institution Press, Washington, D.C.
SIMMONS, N. B. 2000. Bat phylogeny: an evolutionarycontext for comparative studies. Pp. 9–58 in Ontog-eny, functional ecology, and evolution of bats (R. A.Adams and S. C. Pedersen, eds.). Cambridge Uni-versity Press, Cambridge, United Kingdom.
SIMMONS, N. B., AND J. H. GEISLER. 1998. Phylogeneticrelationships of Icaronycteris, Archaeonycteris,Hassianycteris, and Palaeochiropteryx to extant batlineages, with comments on the evolution of echo-location and foraging strategies in Microchiroptera.Bulletin of the American Museum of Natural His-tory 235:1–182.
SIMPSON, G. G. 1932. Miocene land mammals fromFlorida. Florida Geological Survey Bulletin 10:7–41.
SLAUGHTER, B. H. 1970. Evolutionary trends of chi-
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

750 Vol. 84, No. 2JOURNAL OF MAMMALOGY
ropteran dentitions. Pp. 51–83 in About bats: a chi-ropteran symposium (B. H. Slaughter and D. W.Walton, eds.). Southern Methodist University Press,Dallas, Texas.
SMITH, J. D. 1972. Systematics of the chiropteran fam-ily Mormoopidae. University of Kansas Museum ofNatural History, Miscellaneous Publication 56:1–132.
STANEK, V. J. 1933. K topograficke a srovnavacı ana-tomii sluchoveho organu nasich chiropter (Toward atopographic and comparative anatomy of the audi-tory organ of bats). Nakladem Ceske Akademie Veda Umeni, Prague, Czech Republic (in Czech).
SWOFFORD, D. L. 2000. PAUP: phylogenetic analysisusing parsimony. Version 4.02b. Sinauer Associates,Inc., Publishers, Sunderland, Massachusetts.
TEDFORD, R. H., AND C. D. FRAILEY. 1976. Review ofsome Carnivora (Mammalia) from the Thomas FarmLocal Fauna (Hemingfordian, Gilchrist County,Florida). American Museum Novitates 2610:1–9.
TEDFORD, R. H., ET AL. 1987. Faunal succession andbiochronology of the Arikareean through Hemphil-lian interval (late Oligocene through earliest Plio-cene epochs) in North America. Pp. 153–210 in Ce-nozoic mammals of North America: geochronologyand biostratigraphy (M. O. Woodburne, ed.). Uni-versity of California Press, Berkeley.
TEDFORD, R. H., J. B. SWINEHART, C. C. SWISHER III,D. R. PROTHERO, S. A. KING, AND T. E. TIERNEY.1996. The Whitneyan-Arikareean transition in theHigh Plains. Pp. 312–334 in The terrestrial Eocene-Oligocene transition in North America (D. R. Proth-ero and R. J. Emry, eds.). Cambridge UniversityPress, Cambridge, United Kingdom.
VAN DEN BUSSCHE, R. A., AND S. R. HOOFER. 2001.Evaluating monophyly of Nataloidea (Chiroptera)with mitochondrial DNA sequences. Journal ofMammalogy 82:320–327.
VAN VALEN, L. 1979. The evolution of bats. Evolu-tionary Theory 4:103–121.
VARONA, L. S. 1974. Catalogo de los mamıferos vi-vientes y extinguidos de las Antillas. Academia deCiencias de Cuba, La Habana, Cuba.
VAUGHAN, T. A. 1959. Functional morphology of threebats: Eumops, Myotis, and Macrotus. University ofKansas Publications, Museum of Natural History 12:1–153.
WEBB, S. D. 1981. The Thomas Farm fossil site. Plas-ter Jacket 37:6–25.
WHITE, T. E. 1942. The lower Miocene mammal faunaof Florida. Bulletin of the Museum of ComparativeZoology 92:1–49.
WIBLE, J. R., G. W. ROUGIER, M. J. NOVACEK, AND M.C. MCKENNA. 2001. Earliest eutherian ear region: apetrosal referred to Prokennalestes from the earlyCretaceous of Mongolia. American Museum Novi-tates 3322:1–44.
WOODBURNE, M. O., AND C. C. SWISHER III. 1995.Land mammal high-resolution geochronology, inter-continental overland dispersals, sea level, climate,and vicariance. Geochronology, time scales, andglobal stratigraphic correlation. Society of EconomicPaleontologists and Mineralogists, Special Publica-tion 54:336–364.
Submitted 13 August 2001. Accepted 19 August 2002.
Associate Editor was Thomas J. O’Shea.
APPENDIX I
Modern comparative specimens of natalidsand other nataloids are from the mammal col-lections of the Florida Museum of Natural His-tory, University of Florida (UF), Gainesville,Florida; United States National Museum of Nat-ural History (USNM), Smithsonian Institution,Washington, D.C.; Museum of Southwestern Bi-ology (MSB), University of New Mexico, Al-buquerque, New Mexico; Field Museum of Nat-ural History (FMNH), Chicago, Illinois; andOklahoma Museum of Natural History(OMNH), Norman, Oklahoma.
Recent comparative specimens examined.—Natalidae: Natalus (Natalus) major, UF 13691(/), UF 13692 (?), St. Clair Cave, Jamaica; Na-talus (Natalus) stramineus, UF 9847 (?), RioFrio Caves, Belize, MSB 53771 (?), MSB53776 (/), La Aduana, Sonora, Mexico; Natalus(Natalus) tumidirostris, UF 22512 (/), PuebloNuevo, Venezuela, USNM 441670 (?), USNM441671 (?), Miranda, Venezuela; Natalus (Chi-lonatalus) micropus, UF 13698 (/), UF 19076(?), St. Clair Cave, Jamaica; Natalus (Chilon-atalus) tumidifrons, UF 24850 (?), UF 24853(?), Eight Mile Cave, Abaco, Bahamas; Natalus(Nyctiellus) lepidus, UF 12835 (?), Pinder’s Set-tlement, Long Island, Bahamas, USNM 300528(?), Havana, Cuba. Furipteridae: Furipterushorrens, USNM 315737 (?), Changuinola, Pan-ama, USNM 405785 (?), T. F. Amazonas, Ve-nezuela, USNM 549507 (?), Rio Xingu, Brazil;Amorphochilus schnablii, USNM 152261 (sexunknown), Dintorni di Lima, Peru, USNM269981 (/), Arequipa, Peru. Thyropteridae:Thyroptera discifera, USNM 102922 (?),USNM 105422 (/), San Julian, Venezuela,USNM 457965 (/), Barro Colorado Island, Pan-ama; Thyroptera tricolor, USNM 281200 (?),Colonia Agricola, Colombia, USNM 564337(/), Beni, Bolivia, USNM 582082 (?), BarroColorado Island, Panama; Thyroptera lavali,FMNH 89118 (?), FMNH 89120 (/), Loreto,Peru. Vespertilionidae: Kerivoula (Phoniscus)jagori, USNM 155837 (/), Java, Indonesia;Kerivoula hardwickii, USNM 17909 (/), para-type, Cambodia; Kerivoula picta, USNM356429 (/), Da Nang, Viet Nam; Myotis luci-fugus, OMNH uncatalogued (sex unknown),
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

May 2003 MORGAN AND CZAPLEWSKI—NEW MIOCENE NATALID FROM FLORIDA 751
Killdeer, North Dakota. Specimens consist ofskulls and mandibles, most with complete post-cranial skeletons.
APPENDIX II
Characters used in phylogenetic analysis,with the states for each character.
1. Coronoid process of dentary, height rela-tive to articular condyle: (0) higher than con-dyle; (1) same level as condyle; (2) lower thancondyle. 2. Angular process of dentary in lateralview projects: (0) at or below level of occlusalplane of toothrow; (1) above level of occlusalplane. 3. Base of angular process of dentary inlateral view is situated: (0) ventral to condyle;(1) at a level between condyle and tip of coro-noid process. 4. Shape of angular process: (0)straight; (1) slightly curved; (2) strongly hooked.5. Angular process in dorsal or posterior view:(0) in same vertical plane as condyle and as-cending ramus; (1) slightly flared laterally; (2)strongly flared laterally. 6. Position of mandib-ular foramen: (0) opens above level of alveolarmargin of lower toothrow; (1) opens at samelevel as alveolar margin. 7. Size of anterior men-tal foramen (ventral to lower incisors): (0) large,extending distally beneath i2; (1) small, extend-ing distally beneath i1. 8. Spacing of lower in-cisor alveoli: (0) alveoli of i1 and i2 nearly co-alesced, small gap between i2 and i3; (1) alveoliof i1, i2, and i3 evenly spaced. 9. Shape of ar-ticular surface of mandibular symphysis in me-dial view: (0) rounded oval with small ventralprojection; (1) narrow, elongate oval with longposteroventral projection; (2) elongate oval withno ventral projection. 10. Crown of p3: (0) hav-ing 2 posterior crests separated by a concavity;(1) having 1 posterior or lingual crest. 11. Oc-clusal outline of p3: (0) long and strongly lat-erally compressed (length divided by widthgreater than 1.7); (1) shorter and broader (lengthdivided by width less than 1.7). 12. Alignmentof paraconid of m1: (0) in line with other lingualcusps; (1) situated slightly labial in relation to aline drawn through metaconid and entoconid.13. Notches in cristid obliqua and postcristid oflower molars: (0) absent; (1) weakly developed;(2) strongly developed, carnassial-like. 14. La-bial cingular cusp of P4: (0) absent; (1) weak;(2) strong. 15. P4 occlusal outline: (0) indentedanterolingually; (1) not indented anterolingually.16. P4 lingual outline: (0) broadly rounded; (1)narrowed anteroposteriorly. 17. Mesostyle on
upper molars: (0) normal; (1) extended antero-medially as a curved mesostylar crest. 18. Ec-tocingulum of upper molars posterior to para-style and anterior to metastyle: (0) present butweak; (1) absent. 19. M1 postprotocrista: (0) ex-tends only to ‘‘metaconule’’ region (i.e., a truemetaconule is absent), does not reach metaconenor metacingulum; (1) becomes confluent withdistal cingulum (metacingulum); (2) joins lin-gual base of metacone; (3) extends as ridge ontorelatively well-developed talon. 20. Posteroven-tral tubercle of cochlea: (0) absent; (1) present,with lateral hook; (2) present, with lateral andmedial projections; (3) present but weakly de-veloped. 21. Cochlea: (0) makes about 2¼ turns;(1) makes about 2½ turns; (2) makes more than2½ turns. 22. Epitrochlea of humerus: (0) withno medial process; (1) with weak medial pro-cess; (2) with prominent medial process. 23.Distal spinous process of humerus: (0) separatedfrom medial edge of trochlea by small notch; (1)no notch. 24. Shape of spinous process of hu-merus: (0) short with blunt tip; (1) intermediatein length with lateral projection at tip; (2) longand curved laterad; (3) long and nearly straight.25. Braincase shape: (0) inflated; (1) relativelyuninflated. 26. Cranial flexion (rostral–foreheadangle): (0) forehead not sharply elevated aboverostrum (.1358); (1) more sharply elevatedabove rostrum (,1358). 27. Upper canine heightrelative to P4: (0) taller than P4; (1) about equalto P4. 28. Lower canine height relative to p4:(0) taller than p4; (1) about equal to p4. 29. P2size: (0) normal; (1) greatly reduced. 30. Ros-trum length: (0) longer than or equal to brain-case; (1) shorter than braincase. 31. Sides of ros-trum: (0) not markedly inflated; (1) markedly in-flated. 32. Posterior border of palate: (0) not orslightly emarginated; (1) deeply emarginated.33. Elongated styliform process of ectotympan-ic: (0) absent; (1) present, extends anteromedi-ally and contacts pterygoid process. 34. Anteriorportion (maxillary branch) of zygomatic arch:(0) shallow and gracile; (1) deeper and more ro-bust. 35. Cleft between premaxillae: (0) deep,widely separating I1s; (1) tiny, I1s nearly meetat midline. 36. Basisphenoid pits: (0) absent orshallow, double; (1) moderately deep, double;(2) deep and steep-sided, single. 37. Auditorybullae: (0) not enlarged, covers half or less ofthe periotic; (1) greatly enlarged, covers most ofthe periotic. 38. Infraorbital canal length: (0) rel-atively short, extending from level of P4 to M1;
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022

752 Vol. 84, No. 2JOURNAL OF MAMMALOGY
(1) elongated, extending from level of P2 to M2.39. Fossa at anterior edge of orbit (a deep,sharp-edged, funnel-shaped cavity that housesthe lacrimal foramen and posterior end of theinfraorbital canal): (0) absent; (1) present. 40.Fusion of 7th cervical and anterior thoracic ver-tebrae: (0) 7 C not fused to 1 T; (1) 7 C and 1T fused; (2) 7 C and 1–2 T fused; (3) 7 C and1–3 T fused. 41. Fusion of thoracic and lumbarvertebrae (to form what is often termed a lumbar‘‘shield’’ or ‘‘tube,’’ consisting of 10th and 11ththoracic and all but the last 1–2 lumbar verte-brae): (0) lumbar ‘‘shield’’ absent; (1) lumbar‘‘shield’’ present. 42. Number of unfused pos-terior lumbar vertebrae: (0) all lumbar vertebraefree and unfused; (1) only last 2 lumbar verte-brae unfused; (2) only last lumbar vertebra un-fused. 43. Shape of manubrium of sternum: (0)not expanded laterally; (1) expanded laterallyand posteriorly into platelike structure. 44.Shape of ribs: (0) normal, not expanded orfused; (1) broadly expanded but not fused to oneanother; (2) coalesced with each other and withsternum, especially ventrally. 45. Length of cau-dal vertebrae 3 through 6: (0) not elongated; (1)greatly elongated. 46. Proximal end of femur:(0) not sharply bent laterally; (1) sharply bentlaterally. 47. Shape of femur head: (0) rounded;(1) squarish. 48. Greater and lesser trochantersof femur: (0) unreduced, typical for bats; (1)
moderately reduced; (2) highly reduced. 49. P3size: (0) normal; (1) greatly reduced; (2) absent.50. p2 size: (0) normal, about equal to p3; (1)greatly reduced.
APPENDIX III
Data matrix used in the phylogenetic analysis.Numbers after each species name denote char-acter states given in Appendix II. Question markindicates that no data were available for a par-ticular character. Dash indicates that this char-acter is absent.
Primonatalus prattae.—00110, 10000, 01110,01101, 0101?, ?????, ???0?, ??11?, ?????, ????0.Chilonatalus micropus—10121, 01010, 01121,01101, 12010, 10000, 00100, 10113, 12121,11100. Chilonatalus tumidifrons—10121,11010, 00221, 01001, 12010, 10000, 00100,10113, 12121, 11100. Natalus major—10002,11110, 01211, 01001, 12100, 10000, 00100,00111, 11111, 11100. Natalus stramineus—10002, 01101, 01221, 01011, 02100, 10000,00100, 00111, 11111, 11100. Natalus tumidi-rostris—10002, 01111, 01221, 01011, 12100,10000, 11100, 00111, 11111, 11100. Nyctielluslepidus—10111, 11000, 11211, 11002, 10121,01010, 10111, 21113, 11111, 11101. Kerivoulapicta—01011, 01130, 10000, 00000, 10– – 0,00000, 00000, 00200, 00011, 000??. Myotis lu-cifugus—00121, 11130, 10100, 00100, 01001,00011, 00000, 10000, 00101, 00010.
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/84/2/729/2373805 by guest on 03 August 2022
Copyright © 2022 FDOKUMEN




















