
Involvement of CRH-R2 receptor in eating behavior and in the response of the HPT axis in rats...
15
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Involvement of CRH-R2 receptor in eating behavior and in the response of the HPT axis in rats...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Involvement of CRH-R2 receptor in eating behaviorand in the response of the HPT axis in rats subjectedto dehydration-induced anorexia
Patricia de Gortari a,b,*, Karen Mancera a, Antonieta Cote-Velez c, Marıa IsabelAmaya a, Adrian Martınez a, Lorraine Jaimes-Hoy a,b, Patricia Joseph-Bravo c
aDivision de Investigaciones en Neurociencias, Instituto Nacional de Psiquiatrıa RFM, Calzada Mexico-Xochimilco 101,Col. San Lorenzo Huipulco, 14370 Mexico, D.F., Mexicob Escuela de Dietetica y Nutricion del ISSSTE, Callejon Vıa San Fernando 12, Col. San Pedro Apostol, 14070 Mexico, D.F., Mexicoc Instituto de Biotecnologıa, Universidad Nacional Autonoma de Mexico (UNAM), A.P. 510-3, Mor 62210, Mexico
Received 24 April 2008; received in revised form 9 September 2008; accepted 10 September 2008
Psychoneuroendocrinology (2009) 34, 259—272
KEYWORDSTRH;Thyroid axis;Anorexia nervosa;Antisauvagine-30;PVN
Summary Wistar rats subjected to dehydration-induced anorexia (DIA), with 2.5% NaCl solu-tion as drinking water for 7 days, decrease by 80% their food intake and present some changescommon to pair-fed food restricted rats (FFR) such as: weight loss, decreased serum leptin andexpression of orexigenic arcuate peptides, increasing the anorexigenic ones and serum corti-costerone levels. In contrast, the response of the HPTaxis differs: DIA animals have increased TRHexpression in PVN and present primary as opposed to the tertiary hypothyroidism of the FFR.Exclusive to DIA is the activation of CRHergic neurons in the lateral hypothalamus (LH) thatproject to PVN. Since TRH neurons of the PVN contain CRH receptors, we hypothesized that thedifferences in the response of the HPT axis to DIA could be due to CRH regulating TRHergicneurons. CRH effect was first evaluated on TRH expression of cultured hypothalamic cells whereTRH mRNA levels increased after 1 h with 0.1 nM of CRH. We then measured the mRNA levels ofCRH receptors in the PVN of male and female rats subjected to DIA; only those of CRH-R2 weremodulated (down-regulated). The CRH-R2 antagonist antisauvagine-30 was therefore injectedinto the PVN of male rats, during the 7 days of DIA. Antisauvagine-30 induced a higher food intakethan controls, and impeded the changes produced by DIA on the HPT axis: PVN TRH mRNA, andserum TH and TSH levels were decreased to similar values of FFR animals. Results corroborate theanorexigenic effect of CRH and show its role, acting through CRH-R2 receptors, in the activationof TRHergic PVN neurons caused by DIA. These new data further supports clinical trials with CRH-R2 antagonists in anorexia nervosa patients.# 2008 Elsevier Ltd. All rights reserved.
* Corresponding author at: Division de Investigaciones en Neurociencias, Instituto Nacional de Psiquiatrıa RFM, Calzada Mexico-Xochimilco101, Col. San Lorenzo Huipulco, 14370 Mexico, D.F., Mexico. Tel.: +52 555 655 3232; fax: +52 555 655 9980.
E-mail address: [email protected] (P. de Gortari).
ava i lab le at www.sc ienced i rect .com
journa l homepage: www.e l sev ie r.com/locate/psyneuen
0306-4530/$ — see front matter # 2008 Elsevier Ltd. All rights reserved.doi:10.1016/j.psyneuen.2008.09.010

Author's personal copy
1. Introduction
Thyrotropin-releasing hormone (TRH), synthesized in thehypophysiotropic neurons of the hypothalamic paraventricu-lar nucleus (PVN) and released from the middle eminence tothe pituitary, regulates the synthesis and release of thyro-tropin (TSH) that controls thyroid function. The pituitary—thyroid axis (HPT) plays an important role in energy home-ostasis (Lechan and Fekete, 2006a). TRH synthesis andrelease are modified according to the energy status; situa-tions of food deprivation, or restriction, decrease TRH mRNAlevels in the PVN and also, those of TSH and thyroid hormones(TH) in serum (condition recognized as tertiary hypothyroid-ism) (Blake et al., 1991; van Haasteren et al., 1995, 1996).Metabolic signals are first sensed by neurons of the arcuatenucleus that express orexigenic (NPY, AgRP) and anorexigenicpeptides (CART, a-MSH) which in turn, affect PVN TRH andcorticotrophin-releasing hormone (CRH) mRNA levels (Bradyet al., 1990; Leibowitz and Wortley, 2004; Lechan andFekete, 2006b). Among the peripheral signals involved, lep-tin secreted form the adipose tissue exerts an importantregulatory function as its injection into fasted animalsimpedes TRH mRNA down-regulation (Legradi et al., 1997).
Diminished expression of PVN-TRH and HPT function isobserved after acute psychological stress as immobilization(Pacak and Palkovits, 2001), diabetes (van Haasteren et al.,1997), or anorectic conditions as those caused by lipopoly-saccharide (LPS) or interleukin-1beta (IL-1 beta) injection(Kakucska et al., 1994). An exception is the paradigm ofdehydration-induced anorexia (DIA), where rats voluntarilyreduce their food intake (Watts et al., 1999) and lose moreweight than the pair-fed forced food restricted (FFR) group.DIA provokes similar hormonal changes (leptin, corticoster-one and insulin) as those observed in FFR male (Watts et al.,1999) or female rats (Jaimes-Hoy et al., 2008); in the latter,estradiol serum concentrations are reduced (Jaimes-Hoyet al., 2008). In contrast to the tertiary hypothyroidismpresent in FFR, the HPTaxis of female DIA rats is differentiallyaltered: PVN pro-TRH mRNA and TSH serum levels areincreased while TRH content in middle eminence and TRH-R1 expression in the anterior pituitary (AP), decreased(Jaimes-Hoy et al., 2008). The changes in TRH found in DIAare not likely due to arcuate NPYor POMC signals, neither toincreased glucocorticoids, or decreased insulin and leptinserum content, since all these peptides and hormones aresimilarly modified in both experimental conditions (Wattset al., 1999; Jaimes-Hoy et al., 2008). The HPTaxis of DIA ratsthus fails, to adapt to low food intake.
A particular change caused by DIA is the increased expres-sion of CRH in the lateral hypothalamus (LH), which occurs ina proportional manner to the intensity of anorexia (Wattset al., 1999). This is relevant due to the behavioral anorexi-genic effects of CRH injected centrally or directly into the LH(Arase et al., 1988; Cullen et al., 2001). Furthermore, whenadministered into the PVN, CRH inhibits the orexigeniceffects of NPY (Heinrichs et al., 1993; Menzaghi et al.,1993). CRHergic neurons of the LH project to the PVN, mostabundantly to the parvicellular region where TRH and CRHare expressed (Watts et al., 1999). CRH signal is de-codifiedby the two receptors CRH-R1 and CRH-R2 expressed in brain,the anterior pituitary (AP), and peripheral tissues (Aguileraet al., 1987; Chalmers et al., 1995; Lovenberg et al., 1995;
Kageyama et al., 2003). CRH-R2 is present in higher concen-trations than CRH-R1 in the PVN where it is involved in theanorexigenic effects of CRH (Chalmers et al., 1995; Baleet al., 2000; Hillhouse and Grammatopoulos, 2006). In theanterior pituitary (AP) it is CRH-R1 that mediates CRH effectson corticotrophin (ACTH) release from corticotrophs afterstress (Aguilera et al., 1987; DeSouza and Grigoriadis, 1990).
Given the particular changes provoked by DIA on CRH inthe LH, and as PVN TRHergic neurons receive projections ofCRH-synthesizing cells (Hisano et al., 1993), we sought tostudy the role of CRH in the response of TRH neurons and theactivity of HPT axis. We first analyzed the in vitro effect ofCRH onmodulating TRHmRNA levels in hypothalamic primarycultures (Joseph-Bravo et al., 2002; Cote-Velez et al., 2005).As the work of Watts et al. (1999) was performed on malesand ours on females (Jaimes-Hoy et al., 2008), and theresponse of the HPT axis to starvation and fasting showssexual dimorphism (Rondeel et al., 1992; van Haasterenet al., 1996), we evaluated whether there was a sex differ-ence in the HPTresponse to DIA, as well as, on the expressionof CRH receptors in the PVN. Down-regulation of CRH-R2expression was observed only in DIA animals. We thereforetested the effect of its selective antagonist (antisauvagine-30) (Ruhmann et al., 1998) into the PVN, on the HPTresponseto DIA.
2. Methods
The Ethics Committee and the Project Commission of theInstituto Nacional de Psiquiatrıa (INPRFM) approved all dietsand treatments of animals following the Guide for the Careand Use of Laboratory Animals of the U.S. National Institutesof Health. Wistar female and male rats from the Institute’s(INPRFM) animal house with an average body weight (b.w.) of237 � 2 g for females and 245 � 2 g for male animals, weremaintained in a 12 h light—dark cycle (07:00—19:00 h), at22 8C with water and food offered (Lab rodent diet # 5001,PMI feeds; Brentwood, MO, USA) ad libitum.
2.1. Dehydration-induced anorexia
One week before the experiment, animals from the samelot were placed in individual cages and randomly selectedto form three groups. We followed the protocol previouslydescribed (Watts et al., 1999; Jaimes-Hoy et al., 2008).Briefly, one group, naive (N) received water and food adlibitum; the dehydrated group (DIA) received a 2.5% NaCl(SIGMA, St. Louis, MO, USA) solution as their sole drinkingliquid; the forced food restricted group (FFR) was providedwith tap water ad libitum and the same amount of foodconsumed by DIA animals. Total energy of solid foodingested corresponded to the metabolizable energy inPMI food (3.04 kcal/g). Body weight and solid food intakewere recorded daily at 18:00 h before the end of the lightcycle, when food and fresh solution were supplied(18:30 h).
On days 3, 5 and 7, 1/3 of the animals of each group weresacrificed by decapitation between 11:00 and 12:00 h; trunkblood was collected to obtain serum and the brain removedcarefully cutting the optic chiasm to avoid damaging themedian eminence. Brains were stored at �70 8C until ana-lyzed. The PVN and LH were punch-dissected from a coronal
260 P. de Gortari et al.

Author's personal copy
slice of the frozen brain (�0.6 to�3.6 mm from bregma) witha 1.0 mm diameter sample corer (Paxinos and Watson, 2005).
2.2. Injection of CRH-R2 antagonist,antisauvagine-30 into the PVN
A guide cannula of 12 mm long (outer diameter: 0.47 mm andthe inner: 0.18 mm) was inserted stereotaxically in theanterior parvicellular area of the paraventricular nucleusof the hypothalamus [AP = �1.08; L = 0.2; V = 7.2 (Paxinosand Watson, 2005)] of anesthetized male rats with Ketamine(AnestekTM) 73 U/kg and Xylazine 60 U/kg of b.w. (Rom-punTM); the site of injection of CRH antagonist was observedin some control animals by infusing 1 ml of pontamine blue(4% in distilled water and Tween 20 (Sigma, St. Louis MO,USA)), and taking a digital picture of a scanned (Scanner HP5550) coronal frozen slice (100 mm width, cryostat Zeiss,Microm) (Figure 1C). Animals were allowed to recover for1 week in individual cages with water and food ad libitum.DIA was then induced in all animals as stated above. Theywere randomly divided in four groups and injected since 1stday, and during the 7 days of DIA, a volume of 1 ml of 0.9%saline to sham animals (control group: S, n = 6), or with a doseof antisauvagine-30 (ASG-30) (Sigma, St. Louis, MO, USA)dissolved in saline solution. Since the dose of ASG-30 of30 nmol/1 ml attenuates the anorectic effect of CRH injectedinto the LH (Cullen et al., 2001), we used this dose (as a
medium one: MD, n = 6), a lower one (LD: 15 nmol/1 ml,n = 3), and a higher one (HD: 60 nmol/1 ml, n = 3). Twenty-four hours after the 7th injection, animals were decapitated,and trunk blood collected; AP and brains were excised andkept at �70 8C. PVN, and right lateral hypothalamus (LH)were dissected from frozen coronal slices as explained above(Paxinos and Watson, 2005).
2.3. mRNA semi-quantification
Total RNA was extracted from frozen tissues homogenized in4 M guanidine thyocianate (ICN, Aurora, OH, USA), asdescribed (Chomczynski and Sacchi, 1987). RNA quality ofall samples was verified by the ratio of O.D. absorbancies260/280 nm and 260/230 nm considered appropriate whenvalue was >1.8, and by electrophoresis quantifying 28S/18Sratio and discarded if lower than 1.8 or, when evidence ofdegradation was observed by increased staining at the end ofthe gel (Aguilar-Valles et al., 2005; de Gortari et al., 2006).mRNA levels of pro-TRH, pro-CRH, TRH-R1, CRH-R1 and CRH-R2 were semi-quantified by reverse transcriptase polymerasechain reaction (RT-PCR); cyclophilin or glyceraldehyde 3-phos-phate dehydrogenase (G3PDH) were used as control tran-scripts, as previously reported (Aguilar-Valles et al., 2005;Jaimes-Hoy et al., 2008); cDNA was prepared from 1.5 mg ofRNA (M-MLV reverse transcriptase (Carlsbad, CA, USA), andoligo-dT, followed by the PCR reaction: 6 ml of cDNA with
Figure 1 Methodological optimization. Number of cycles for probes. Data are optical densities of the signal of the PCR product atdifferent cycles of every probe of the (A) PVN and (B) lateral hypothalamus subjected to electrophoresis in a 2% agarose gel. (C) Digitalpicture of a scanned (Scanner HP 5550) coronal fresh slice (100 mmwidth, cryostat Zeiss, microm) of PVN showing the site of injectionof 1 ml of pontamine blue.
CRH activates PVN TRH expression during dehydration 261

Author's personal copy
25 pmol of specific oligonucleotides (except 50 pmol for themarkergenes cyclophilin andG3PDH) (synthesizedat theUNAMBiotechnology Institute’s facilities) and 0.5 ml Taq DNA poly-merase (5 U/ml) (Biotecnologıas Universitarias, UNAM, DF,Mexico), as described (Aguilar-Valles et al., 2005; Jaimes-Hoy et al., 2008). Probes for CRH-R2 were: (sense: 50-TTGGCAAGGAACCTGGTGAC-30, antisense: 50-ACCCAGG-GAAGGCTGTGAAG-30) (Lovenberg et al., 1995) for CRH-R1:(sense: 50-TCCACTACATCTGAGACCATTCAGTACA-30, antisense:50-TCCTGCCACCGGCGCCACCTCTTCCGGA-30). The number ofcycles of various probes were previously optimized to assurelinear conditions (Jaimes-Hoy et al., 2008); for CRH-R2, CRH-R1 or other tissues such as, lateral hypothalamus, each probewas tested for different cycles (Figure 1). Final conditions inPVNwere: 32cycles forCRH-R1 andCRH-R2; for LH samples: 28cycles for pro-TRH, 30 for pro-CRH and 20 cycles; for the rest,conditionswere as described (Jaimes-Hoy et al., 2008); for AP:26 for TRH-R1. Each cycle consisted of: 1 min at 94 8C, fol-lowed by 1 min at 64 8C for pro-TRH, TRH-R1, CRH-R2, cyclo-philin and G3PDH; 55 8C for CRH-R1; 60 8C for pro-CRH and afinal min at 72 8C. All cDNAs had a final extension of 10 min at72 8C. Several cDNAs were semi-quantified from the same RTreaction (Figure 1). PCR products (10 ml of each DNA, and 5 mlof either G3PDH or cyclophilin) were separated by 2% ofagarose (or 2.5% for CRH) (Ultra-pure Bio-Rad, Hercules CA,USA) gel electrophoresis, stained with ethidium bromide(1 mg/l) and density measured with the Advanced AmericanBiotech Imaging software (American-Applied Biotechnology,Fullerton, CA, USA). The relative amounts of the cDNAs understudy were calculated as the ratio of each cDNA over cyclo-philin or G3PDH cDNA. Care was taken to include in the samegel, samples of N, FFR and DIA.
2.4. Radioimmunoassays
Rat serum TSH levels were quantified with NIDDK (NationalHormone and Pituitary Program) protocol and materials using50 ml of serum diluted 1:3 with RIA buffer [limit of detection:5 pg, 13% inter-assay, 6% intra-assay variability]. Corticoster-one was quantified using ICN Biomedicals (Aurora, OH, USA)kit, with antiserum, diluted 1:1000 [limit of detection: 5 pg;8% inter-assay, 7% intra-assay variability]. 100 and 25 ml ofserumwere used to measure T3 and T4, respectively, with theRIA kit from Diagnostic Products Corporation (Los Angeles,CA, USA) [limit of detection: T3 (7 ng/dl), T4 (0.25 ml/dl);inter-assay variability (T3, T4 < 10%), intra-assay variabilityT3 or T4 < 7%)]. Serial dilutions of normal serum were used asinternal standard in all assays. Leptin was measured only inmales at the 7th day, with a colorimetric assay using theenzyme immunometric kit for rat leptin Titerzyme (AssayDesigns, Ann Arbor, MI, USA) using 100 ml of serum diluted 1:2with assay buffer (detection limit: 47 pg/ml; 3.3% inter-assay, 4.1% intra-assay variability).
2.5. Cell culture
Primary cultures of hypothalamic cells were performed asreported (Perez-Martinez et al., 1998; Joseph-Bravo et al.,2002). Briefly, on the 17th day of gestation, dams wereanesthetized with 0.45 ml Sedalphorte (63 mg sodium pen-tobarbital) and the embryos removed. The head was cutbetween the eye and superior edge of the ear, the brain
was kept in the skull to avoid tissue distortion, and thehypothalamus dissected by cutting the area limited by theoptic chiasm and lateral sulcus including the mammillarybodies, to a 2—3 mm depth. 6 � 105 cells were plated in16 mm dishes pre-coated with 1.5 mg/ml poly-D-lysine; cul-tures incubated with DMEM supplemented with 10% fetal calfserum, 0.25% glucose, 2 mM glutamine, 3.3 mg/ml insulin, 1%antibiotic-antimycotic and 1% vitamin solution. Cultureswere maintained at 90% humidity, 7% CO2, 93% air, 37 8C.On the 4th day in vitro (DIV) 10�5 M cytosine arabinofurano-side was added; after 2 days, half the incubation mediumwasreplaced with fresh supplemented DMEM every second day, asdescribed (Charli et al., 1995). Cultures were kept for 18 DIVand observed under an inverted microscope (Zeiss Axioskopmicroscope) at every medium change. CRH (Sigma, St. Louis,MO, USA) was dissolved in DMEM and added at the concen-trations and for the time stated in results.
2.6. Data analysis
Most results were analyzed by a three-way ANOVA. A two-wayANOVAwas performed to analyze data of in vitro studies, thecomparison of PVN CRH expression between males andfemales at day 7, food intake and body weight changes ofantagonist injection experiment; a one-way ANOVA was usedto analyze data of leptin content in males, of HPT axischanges of the antagonist injected animals, and pro-TRHexpression in cultured cells; when significant ( p < 0.05),analyses were followed by Fisher’s post hoc test.
3. Results
3.1. In vitro effect of CRH on TRH mRNA levels
CRH (10 nM) increased pro-TRH mRNA levels in hypothalamiccell culture by 30 min of incubation (one-way ANOVA:F(4,15) = 3.153; p < 0.05) (Figure 2A). A dose response curvewas performed for 1 and 2 h incubations; the lowest dose,0.1 nM, significantly augmented pro-TRH mRNA levels at 1 h.Two-way ANOVA showed differences between doses (F(1,31)= 4.192, p < 0.01) and, incubation time (F(1,31) = 10.758,p < 0.01) (Figure 2B).
3.2. Comparison of gender response to DIA
The effect of 7 days of DIAwas studied in male and female ratsfrom the same lot. Males of the naıve group ate more thanfemales (80 � 3 g/kg vs. 73� 2 g/kg b.w.; sex: F(1,282)
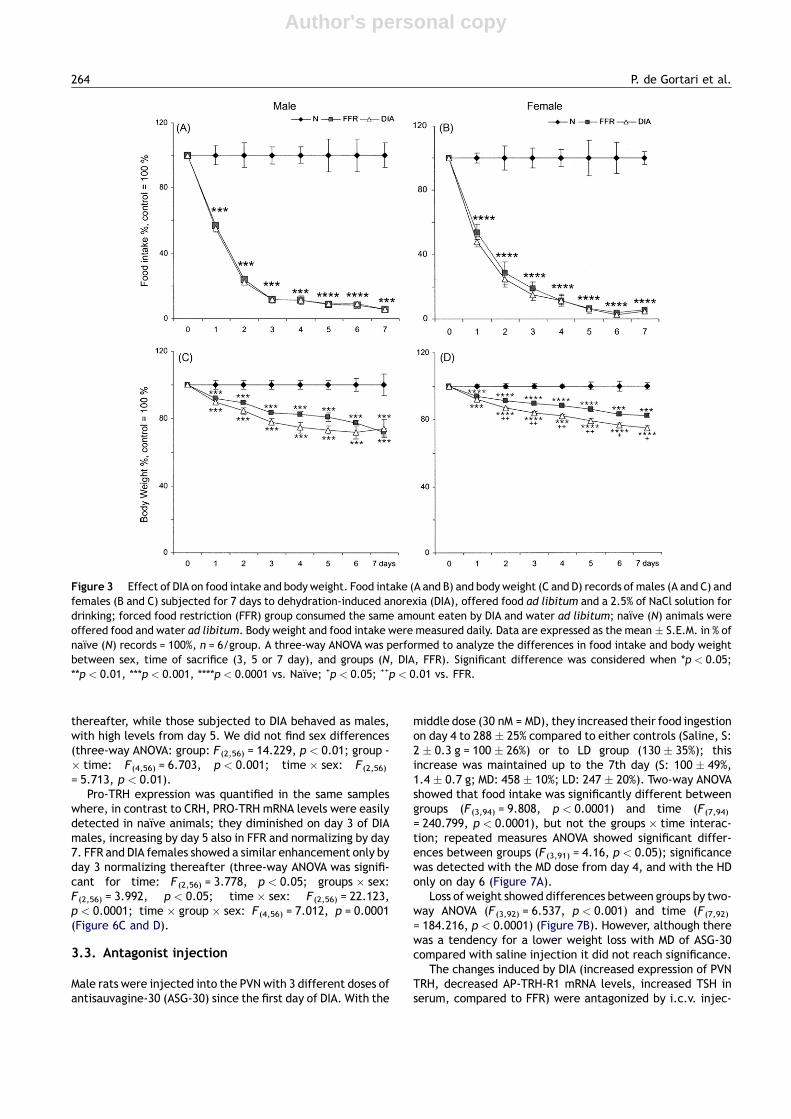
= 30.75, p < 0.0001). DIA induced a drastic decrease in foodintake of males from day 1 to 4, maintaining steady intakethereafter in contrast to females, whose decreased consump-tion continued until the 6th day (time: F(7,282) = 205,p < 0.0001; groups: F(2,282) = 1277, p < 0.0001); group-s � time: F(14,282) = 50, p < 0.0001; groups � sex: F(2,282)
= 3.2, p < 0.05; time � sex: F(7,282) = 2.7, p < 0.05) (Figure3A and B). Animals started losing body weight from the firstday, achieving a b.w. loss of almost 30% by the 7th day inmales(N = 100%, FFR = 73.4%, DIA = 67.7%), higher than that offemales (N = 100%, FFR = 82.4%, DIA = 74.9%). Three-wayANOVA showed differences in body weight between groups(F(2,282) = 365.179, p < 0.0001); time (F(7,282) = 21.328,
262 P. de Gortari et al.

Author's personal copy
p < 0.0001); groups and time (F(14,282) = 16.43, p < 0.0001);sex: (F(1,282) = 16.8, p < 0.0001); groups � sex: (F(2,282)
= 10.4, p < 0.0001) (Figure 3C and D). Normalizing food intakeby b.w., the relation gave a significant effect of sex, showingthat females ate less (i.e., on day 4males: 12.2 � 2 g/kg b.w.,females: 11.2 � 3 g/kg b.w.; three way ANOVA: F(1,282)
= 30.775, p < 0.0001). Despite that the FFR groups receivedtheexactamountof food ingestedbyDIAanimals,b.w. losswasmore pronounced in DIA (two-way ANOVA males: F(2,156)
= 173.1, p < 0.0001; time F(7,156) = 7, p < 0.0001; group -� time F(14,156) = 7.6 p < 0.0001; females: groupsF(2,126) = 454.6, p < 0.0001; times F(7,126) = 71.7,p < 0.0001; group � time F(14,126) = 21 p < 0.0001) (Figure3C and D).
HPT axis activity was analyzed measuring mRNA levels ofpro-TRH in PVN and of TRH-R1 in anterior pituitary, as well asTSH and TH serum content. Basal levels of TSH were lower, ofT4 similar, and of T3 higher in females than inmales. Comparedto naıve animals, FFR or DIA induced changes in females aspreviously reported (Jaimes-Hoy et al., 2008): PVN-TRHexpression increased from day 5 in DIA but decreased in FFR(three-way ANOVA showed significance for groups: F(2,52)
= 107.102, p < 0.0001; time: F(2,52) = 4.109, p < 0.05; sex:
F(1,52) = 24.191, p < 0.0001; groups � sex: F(2,52) = 9.668,p < 0.001; time � sex: F(2,52) = 31.397, p < 0.0001; time -� sex � groups: F(4,52) = 9.752, p < 0.0001) (Figure 4A and B).
An opposite response between DIA and FFR was alsoobserved in TRH-R1 expression in the AP, decreasing in DIAbut increasing in FFR of both sexes albeit stronger in females(Figure 4C and D) (three-way ANOVA: groups � sex:F(2,49) = 6.469, p < 0.01; time: F(2,49) = 19.567, p < 0.0001;groups: F(2,49) = 125.157, p < 0.0001; groups � time: F(4,49)
= 5.914, p < 0.001; time � sex: F(2,49) = 15.994, p < 0.0001;time � groups � sex: F(4,49) = 5.583, p < 0.001).
TSH values increased in DIA compared toN or FFR since day3 on males and until day 7 on females (three-way ANOVAshowed differences only between groups: F(2,56) = 11.864,p < 0.0001) (Figure 4E and F). Serum T4 levels displayedthe greatest sex difference. In DIA-males it was increasedon day 3 (as was TSH) and day 5, normalizing by day 7. In FFRthe increase was seen only on day 5 (Figure 4G); in contrast,T4 values were decreased in females relative to N only on day5 and in both the FFR and DIA groups (three-way ANOVAshowed for groups: F(2,51) = 14.351, p < 0.0001, time:F(2,51) = 5.981, p < 0.01, sex: F (1,51) = 44.583, p < 0.01,groups � sex: F(2,51) = 17.38, p < 0.0001, time � sex:F(2,51) = 20.073, p0.0001, time � groups � sex: F (4,51)
= 7.206, p < 0.0001) (Figure 4H). T3 concentrations in theserum of FFR and DIA were lower than N to a similar extent inmales and females (three-way ANOVA for groups: F(2,60)
= 32.340, p < 0.0001) (Figure 4I and J).Leptin levels ofmales after 7 days of DIA (63 � 18 pg/ml) or
FFR (191� 18 pg/ml) were reduced compared to N(292� 42 pg/ml) (one-way ANOVA: F(2,20) = 7.98, p < 0.01),as reported (Watts et al., 1999) and similar to the observeddecrease in females (Jaimes-Hoy et al., 2008).
Basal corticosterone concentration was higher in femalethan male rats (73.2 � 0.3 ng/ml vs. 33.4 � 4 ng/ml). Levelsincreased since day 3 in FFR of both sexes (albeit higher inmales 283� 18% vs. 188 � 29% in females), and by day 5in females (161� 18% males, 198 � 11% in females;naıve = 100%). DIA induced changes only from day 5 onwards(males: 196 � 14, females 207 � 3%). Highest increase wasproducedatday7 (Figure5AandB); (three-wayANOVA showedsignificant differences between groups (F(2,60) = 108.9,p < 0.0001), time (F(2,60) = 35.5, p < 0.0001); time � groupsF(4,60) = 15.6, p < 0.0001).
As for corticosterone, CRH mRNA levels in the PVN werehigher in the females (not shown); CRH mRNA levels dimin-ished with food restriction or DIA to a similar extent in malesof both groups while in females, the DIA group lowered itsvalues more than the FFR did (two-way ANOVA on day 7:F(2,17) = 3.621, p < 0.05) (Figure 5A and B). mRNA levels ofCRH receptors, evaluated in the same samples, showed novariation in CRH-R1 expression either by sex or treatment.CRH-R2 mRNA levels decreased by DIA in both males andfemales, and only slightly increased in FFR males (three-wayANOVA, groups: F(2,51) = 18.006, p < 0.0001); time: (F(2,51)
= 5.53, p < 0.01; sex: (F (1,51) = 22.761, p < 0.0001 group-s � sex: F(2,51) = 5.963, p < 0.01; time � sex: F(2,51)
= 4.539, p < 0.05) (Figure 5A and B).CRH mRNA levels were also measured in lateral hypotha-
lamus where, as reported (Watts et al., 1999) increased onlywith DIA since the 5th day in males (Figure 6A and B). FFRfemale rats showed a transient increase on day 3 normalizing
Figure 2 Effect of CRH on pro-TRH mRNA levels in culturedhypothalamic cells. (A) Kinetics of pro-TRH mRNA levelsincreases, measured by RT-PCR, by the addition of CRH(10 nM) to primary cultured hypothalamic cells at 18 DIV (n = 8dishes/point). (B) Dose response of TRH expression changes at 1and 2 h after the addition of CRH. Data are the mean � S.E.M. ofn = 4 dishes/point; data are expressed as percentage of TRHexpression in control cultures = 100%. Post hoc analysis per-formed when ANOVA was significant: when *p < 0.05,**p < 0.01 vs. time 0 or vehicle (dose 0).
CRH activates PVN TRH expression during dehydration 263
Author's personal copy
thereafter, while those subjected to DIA behaved as males,with high levels from day 5. We did not find sex differences(three-way ANOVA: group: F (2,56) = 14.229, p < 0.01; group -� time: F(4,56) = 6.703, p < 0.001; time � sex: F (2,56)
= 5.713, p < 0.01).Pro-TRH expression was quantified in the same samples
where, in contrast to CRH, PRO-TRH mRNA levels were easilydetected in naıve animals; they diminished on day 3 of DIAmales, increasing by day 5 also in FFR and normalizing by day7. FFR and DIA females showed a similar enhancement only byday 3 normalizing thereafter (three-way ANOVA was signifi-cant for time: F (2,56) = 3.778, p < 0.05; groups � sex:F(2,56) = 3.992, p < 0.05; time � sex: F (2,56) = 22.123,p < 0.0001; time � group � sex: F(4,56) = 7.012, p = 0.0001(Figure 6C and D).
3.3. Antagonist injection
Male rats were injected into the PVN with 3 different doses ofantisauvagine-30 (ASG-30) since the first day of DIA. With the
middle dose (30 nM = MD), they increased their food ingestionon day 4 to 288 � 25% compared to either controls (Saline, S:2 � 0.3 g = 100 � 26%) or to LD group (130 � 35%); thisincrease was maintained up to the 7th day (S: 100 � 49%,1.4 � 0.7 g; MD: 458 � 10%; LD: 247 � 20%). Two-way ANOVAshowed that food intake was significantly different betweengroups (F(3,94) = 9.808, p < 0.0001) and time (F (7,94)
= 240.799, p < 0.0001), but not the groups � time interac-tion; repeated measures ANOVA showed significant differ-ences between groups (F (3,91) = 4.16, p < 0.05); significancewas detected with the MD dose from day 4, and with the HDonly on day 6 (Figure 7A).
Loss of weight showed differences between groups by two-way ANOVA (F (3,92) = 6.537, p < 0.001) and time (F(7,92)
= 184.216, p < 0.0001) (Figure 7B). However, although therewas a tendency for a lower weight loss with MD of ASG-30compared with saline injection it did not reach significance.
The changes induced by DIA (increased expression of PVNTRH, decreased AP-TRH-R1 mRNA levels, increased TSH inserum, compared to FFR) were antagonized by i.c.v. injec-
Figure 3 Effect of DIA on food intake and body weight. Food intake (A and B) and body weight (C and D) records of males (A and C) andfemales (B and C) subjected for 7 days to dehydration-induced anorexia (DIA), offered food ad libitum and a 2.5% of NaCl solution fordrinking; forced food restriction (FFR) group consumed the same amount eaten by DIA and water ad libitum; naıve (N) animals wereoffered food and water ad libitum. Body weight and food intake were measured daily. Data are expressed as the mean � S.E.M. in % ofnaıve (N) records = 100%, n = 6/group. A three-way ANOVA was performed to analyze the differences in food intake and body weightbetween sex, time of sacrifice (3, 5 or 7 day), and groups (N, DIA, FFR). Significant difference was considered when *p < 0.05;**p < 0.01, ***p < 0.001, ****p < 0.0001 vs. Naıve; +p < 0.05; ++p < 0.01 vs. FFR.
264 P. de Gortari et al.

Author's personal copy
tions of antisauvagine-30. pro-TRH mRNA levels decreased(most with MD) compared to saline injected DIA (one-wayANOVA: F3,13) = 10.102, p = 0.001), correlating negativelywith food intake (insert of Figure 8; F(1,11) = 16, p < 0.005,
r = 0.784). TRH-R1 expression in anterior pituitary increasedand serum levels of TSH and T4 decreased with themedial andhigh doses (Figure 8) (One-way ANOVA for TRH-R1:F(3,8) = 54.058, p < 0.0001; serum TSH, F(3,10) = 4.218,
Figure 4 Variations of HPT axis function parameters. PVN mRNA levels of TRH (A and B) and AP TRH-R1 (C and D); serumconcentrations of TSH (E and F), T4 (G and H), T3 (I and J) of male (A, C, E, G and I) or female (B, D, F, H and J) rats sacrificedat days 3, 5 or 7 after dehydration-induced anorexia (DIA), forced food restriction (FFR) and naıve (N). Values of TRH and TRH-R1 mRNAlevels are the mean � S.E.M. of the ratio of each cDNA/cyclophilin cDNA signals (arbitrary units). All data are expressed as percentageof N values considered as 100%, n = 6/point. Naıve values of males: TSH = 1.7 � 0.2 ng/ml, T4 = 2.5 � 0.3 mg/dl; T3 = 74 � 2.7 ng/dl;females: TSH = 1.58 � 0.09 ng/ml; T4 = 4 � 0.9 mg/dl; T3 = 74 � 5 ng/dl. when *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs.Naıve; +p < 0.05, ++p < 0.01, +++p < 0.001, ++++p < 0.0001 vs. FFR.
CRH activates PVN TRH expression during dehydration 265

Author's personal copy
p < 0.05; T4, F (3,11) = 4.035, p < 0.05). For comparison,values of saline injected DIA animals: TSH = 3.75 � 0.3 ng/ml; T4: 2 � 0.3 mg/dl; T3: 46 � 9 ng/dl, vs. those of control(Naıve) values for TSH = 1.4 � 0.19 ng/ml; T4: 2.2 � 0.6 mg/dl; T3: 74 � 2 ng/dl.
4. Discussion
In this work we corroborated the effect of dehydration-induced anorexia on the increase in CRH expression of lateralhypothalamus, and the lack of adaptation of the HPT axis tofood restriction, more prominently in females. As this adap-tation is essential for survival in conditions of negative energybalance (Wiersinga, 2005), we pursued the study on theeffect of CRH on TRH metabolism. We demonstrated thatCRH can induce a fast increase in pro-TRH mRNA levels inhypothalamic cultured cells. In vivo, we showed that the DIA-induced CRH expression of the lateral hypothalamus wasaccompanied by a preferential down-regulation of CRH-R2expression in the PVNwhich justified a study of the effect of aselective CRH-R2 antagonist, antisauvagine-30 (ASG-30), on
the response of the HPT axis to DIA, and showed that itsinjection into the PVN allowed adaptation of the HPT axis tofood restriction.
4.1. Sex difference in the response to DIA of theHPT axis
The reported sexual dimorphism in several parameters of theHPT and HPA axis was confirmed: female rats show higher T3serum levels and lower TSH release than males, but nodifference in basal pro-TRH mRNA levels (Donda et al.,1987; Cizza et al., 1996; Doufas and Mastorakos, 2000;Marassi et al., 2007). As previously reported (Lunga andHerbert, 2004; Barna et al., 2003), basal corticosteronelevels were higher in female than male rats of the samelot, while the relative CRH mRNA levels were lower (notshown). Fasting or food restriction are conditions that down-regulate pro-TRHmRNA levels in the PVN (Blake et al., 1991),in spite of the negative feedback that the low TH serum levelsshould be exerting in the hypothalamus (Lechan and Fekete,2006a). The tertiary hypothyroidism caused by conditions ofnegative energy balance has been considered as an evolu-tionary advantage that leads to a reduced metabolic rate andin consequence, to a slower degradation of energy depositsthat favours survival. Sex differences in energy balance occurin response to fasting or to food restriction; males are moresusceptible than females, altering their expression of anor-exigenic or orexigenic peptides in arcuate and paraventri-cular nuclei (NPY, POMC, Gal, CRH) and decreasing theresponse of those peripheral hormones involved in metabolicbalance (leptin, thyroid hormones) that account for thegreater body and organ-weight loss of males (Brady et al.,1990; Valle et al., 2005; Gayle et al., 2006). In conditions offood restriction (100 days of 40% caloric restriction) forexample, female rats lose less body and organ-weight, andpresent lower VO2 (indirect measure of energy expenditure)(Valle et al., 2005). This could be related to a decreasedmetabolic rate due to differences in HPT activity as femalesshow a greater decrease than males in TRH release, and intotal and free T4 or T3, in response to either fasting or foodrestriction (Rondeel et al., 1992; van Haasteren et al., 1996).A direct comparison of food restriction (4 days) vs. 2 week-food deprivation shows the former to be more stressful andcause more changes than the latter (Brady et al., 1990). Inour paradigm, pair-fed animals were food restricted onlyduring 7 days but more drastically (50% on day 1, 85—90% byday 3) and at the end (90—95%) the low intake was almost likefasting. DIA and FFR females showed a higher susceptibility ofthe HPTaxis than males, in particular on the levels of T4 thatwere decreased only in females from the 5th day, in contrastto the increased levels in DIA males on day 3 and 5. The highserum T4 levels could produce a differential tissue conversionto T3 (Coppola et al., 2005; Bianco et al., 2005). Despite thissex difference, T3 levels in serum were decreased similarly inboth sexes by day 7 and cannot be explained be a differentialdeiodinase I activity reported to be higher in male’s liver,pituitary and thyroid (Marassi et al., 2007). Several para-meters of the HPT axis are influenced by estrogens (Dondaet al., 1987; Cizza et al., 1996; Marassi et al., 2007), includ-ing the expression of thyroid binding globulin (TBG) thatcould limit the amount of free T4 available to deiodinases.
Figure 5 Variations of HPA axis function parameters. Naıve(N), dehydrated (DIA) or food restricted (FFR) for a weekmale (A)or female (B) rats, were sacrificed at day 7; values were calcu-lated as % of N group (=100%) and expressed as mean � S.E.M..Corticosterone control values of males: 33.4 � 4 ng/ml;females: 73.2 � 0.3 ng/ml. Data of mRNA levels in PVN arethe ratio of cDNA of CRH/G3PDH, CRH-R1 or CRH-R2/cyclophilin(arbitrary units). *p < 0.05, **p < 0.01, ***p < 0.001,****p < 0.0001 vs. Naıve, +p < 0.05, +++p < 0.001 vs. FFR.
266 P. de Gortari et al.

Author's personal copy
There is controversy over the effects of estrogens on thelevels of hormone binding proteins. Some authors attributeincreased TGB levels to elevated levels of estradiol (Arafah,2001). Others suggest that ovariectomy or replacement withestradiol does not have any effect (Emerson et al., 1990).Corticosterone in contrast, decreases TGB levels (adrenalec-tomized female rats have increased TBG synthesis and serumconcentration than males) (Emerson et al., 1993). As thelevels of 17b-estradiol decrease after 7 days of FR or DIA to8—10 pg/ml (similar or lower values than those of diestrous Ifrom 26 � 4 pg/ml of controls (similar to levels attained inthe morning of proestrous (Nequin et al., 1979; Tohei et al.,1998; Jaimes-Hoy et al., 2008))). Testosterone levels are notmodified by food restriction (Leal and Moreira, 1997) theexpression and activity of deiodinases thus requires to bestudied.
These new data show that the HPT axis of males, aspreviously shown for females (Jaimes-Hoy et al., 2008),
responds differently to DIA compared to equivalent foodrestriction: greater levels of TRH expression in the PVN,TRH release (inferred by decreased TRH-R1 in the AP (Yangand Tashjian, 1993), and TSH in serum, than their pair-fedfood restricted group. Sex differences were howeverobserved; pro-TRH mRNA levels, TSH and T4 were increasedearlier in DIA males than in females. Although T3 serumconcentrations were diminished, doubt remains whether thisalso occurs in other tissues such as muscle or adipose tissue,since T4 is increased in DIA male rats and a differentialactivity of the deiodinase type II (DII) could therefore accountfor the greater weight reduction of DIA compared to FFRmalerats or, to DIA female rats. Whether this relates, for example,to a greater ghrelin content increase induced by 12 h fast infemales compared to males (Gayle et al., 2006) or, the sexdifferences in the expression of peptides of the lateral andarcuate nuclei involved in feeding behavior, or in gustatoryperception of salt or motivational arousal (Brady et al., 1990;
Figure 6 Changes in TRH and CRH gene expression in the lateral hypothalamus. Dehydrated (DIA), food restricted (FFR), and naıve(N) male (A and C) and female (B and D) animals were sacrificed after 3, 5 and 7 days on each paradigm. CRH (A and B) and TRH (C and D)expression was measured in the right lateral hypothalamus. Values are the mean � S.E.M. of the ratio of CRH cDNA/G3PDH cDNA signalor TRH cDNA/cyclophilin cDNA signals (arbitrary units). All data are expressed as percentage of N values considered as 100%, n = 4.Three-way ANOVAwas performed and when p < 0.05, followed by a Fisher’s post hoc analysis. *p < 0.05, **p < 0.01, ****p < 0.0001 vs.Naıve; +p < 0.05, ++++p < 0.0001 vs. FFR. The insert depicts representative gels of cDNAs from naıve (N, two first lanes), dehydration-induced anorexic (DIA, 2middle lanes) and, forced food restricted (FFR, 2 last lanes) animals; superior band = G3PDH, lower band = CRHcDNA signal.
CRH activates PVN TRH expression during dehydration 267

Author's personal copy
Curtis et al., 2004; Mogi et al., 2005; Shelley et al., 2007) thatmay affect TRHergic cells of the PVN (Mountjoy et al., 1994;Fekete et al., 2001, 2002), awaits resolution.
4.2. Response of the CRHergic hypothalamicpathways
CRH and its receptors regulate in concert, the autonomic,behavioral, immunologic and endocrine responses of theorganism allowing it to face a changing environment. Starva-tion, body weight reduction, and negative energy balance aresome of the stressful conditions that induce differentialchanges in the HPA axis, as well as in hypothalamic and centralCRH metabolism, enabling the organism to adapt to situationsof low energy availability. CRH also participates in the regula-tion of energy intake and utilization through its actions on theSympathic Nervous System, activating thermogenesis in thebrown adipose tissue (LeFeuvre et al., 1987; Arase et al.,1988). As previously reported, CRH expression in the PVNwas decreased after 7 days of DIA or FFR in female and malerats (Brady et al., 1990; Isse et al., 1999; Watts et al., 1999;Jaimes-Hoyet al., 2008) but toa greater extent inDIA females;this sex-difference was not observed for the increases in
corticosterone. Even though, CRH-R1 expression in the PVNis up-regulated in several stressful conditions supposedly byCRH released into the PVNwhich in turn up-regulates PVN-CRHexpression (Mansi et al., 1996; Imaki et al., 1996; Turnbull andRivier, 1997), the mRNA levels of this receptor wereunchanged. In contrast, CRH-R2 mRNA levels were down-regulated by DIA, but not by FFR; the lack of response ofFFR animals coincided with that reported after starvation(Makino et al., 1998). CRH-R2 is the receptor involved inthe anorexigenic effects of administered CRH (Morley andLevine, 1982; Arase et al., 1988; Krahn et al., 1990). Knockdown of CRH-R2, but not of CRH-R1, attenuates appetite andfood intake reduction (Smagin et al., 1998) as well as theanorexigenic response to urocortin (Ucn) injection in the LH(Cullen et al., 2001). Ucn is a CRH-like peptide that shows alower anorexigenic effect than CRH despite having 20� higheraffinity than CRH for CRH-R2 (Vaughan et al., 1995).
CRH mRNA levels are increased, exclusively by DIA, inneurons of the lateral hypothalamus of male rats (theseresults and Watts et al., 1999), as well as, females’. TheseCRHergic neurons project to the PVN and have been proposedas important targets for osmosensitive afferents from theforebrain, the middle preoptic nucleus and subfornical organand from the fusiform nucleus of the bed nucleus of the striaterminallis; projections mediating part of the response ofpeptidergic mRNAs in the lateral hypothalamus during dehy-dration (Kelly and Watts, 1996). Increased CRH expression inDIA animals is not dependent of corticosterone levels, sinceafter adrenalectomy and dehydration, rats still present
Figure 8 Changes of HPT axis parameters after ASG-30 injec-tion. PVN TRH, AP TRH-R1 expression and TSH, T4 and T3 serumlevels were measured in male animals subjected to DIA paradigmand injected daily with either 1 ml of 0.9% saline solution (S),15 nM (LD), 30 nM (MD) or 60 nM (HD) of ASG-30 and sacrificedafter 7 days of treatment. Values of PVN TRH and AP TRH-R1expression are the means � S.E.M. of the ratio of each cDNA/cyclophilin signal (arbitrary units); all data are expressed aspercentage of S values considered as 100% (n = 3 for LD, HD,n = 6 for S and MD). One-way ANOVA was performed to analyzechanges in PVN TRH expression followed by a Fisher’s post hocanalysis; when *p < 0.05, **p < 0.01, ***p < 0.001,****p < 0.0001 vs. Saline; +p < 0.05, vs. ++p < 0.01 vs. LD,#p < 0.05, ##p < 0.01, ###p < 0.001 vs. MD injected animals.
Figure 7 Daily food intake and body weight records of animalsinjected with antisauvagine-30 in the PVN. Male animals sub-jected to dehydration-induced anorexia (DIA) were injecteddaily directly into the PVN with either 1 ml of 0.9% saline solution(S), 15 nM (LD), 30 nM (MD) or 60 nM (HD) of ASG-30. Food intakeand body weight were recorded daily. Values are the means(g) � S.E.M. of the records (n = 3 for LD, HD, n = 6 for S andMD). Repeated measures ANOVA showed significant differencesbetween groups (F(3,91) = 3.92, p < 0.05).
268 P. de Gortari et al.

Author's personal copy
higher mRNA levels than FFR group (Watts et al., 1999). It istempting to suggest that an increased release of CRH fromthe lateral hypothalamus impinges on CRH-R2 receptors ofthe PVN, causing a receptor down-regulation, as shown forthe hypophysial homologous down-regulation of CRH recep-tor (Rabadan-Diehl et al., 1996). Whether the receptiveneurons express TRH, is currently investigated.
As TRH has also been suggested to have anorexigeniceffects (Vijayan and McCann, 1977; Karydis and Tolis,1998; Choi et al., 2002) its mRNA levels were also measuredin lateral hypothalamus. Early changes of pro-TRH expressionoccurred differently in DIA or FFR and by animals’ sex, but didnot relate to changes in HPT axis. Pro-TRH expression in theLH is not modified by fasting (Legradi et al., 1997) or thyroidstatus (Segerson et al., 1987) although recent data showedincreased immunoreactive TRH precursor by inducedhypothyroidism (Espinosa et al., 2007). In contrast to thecharacterization and function of TRHergic neurons of the PVN(Lechan and Fekete, 2006a), little is known for those of theLH. Some TRH expressing neurons in the lateral hypothalamusproject to the lateral septum where TRH induces arousal andmodulates the cholinergic septo-hippocampal pathways(Ishikawa et al., 1986; Prokai, 2002) but, they do not havehypophysiotropic functions. Destiny of other afferents and inparticular if they make intra-hypothalamic contacts remainsto be studied.
4.3. Effect of CRH-R2 antagonist on the HPTresponse to DIA
As male rats lost more weight and showed greater activationof the HPT after DIA than females, and estrogens indirectlydown-regulate PVN-TRH expression (unpublished) we pre-ferred to study the role of CRH-R2 receptor in male rats.ASG-30 injection enhanced feeding supporting its antagonis-tic effects on endogenous CRH (Brown et al., 1982; Rohner-Jeanrenaud et al., 1989; Rivest et al., 1989; Buwalda et al.,1997; Bornstein et al., 1998; Cullen et al., 2001). The loss ofappetite induced by CRH seemed thus to involve CRH-R2 ofthe PVN which antagonism also diminished DIA’s effects onHPTaxis. Our results did not show an effect of ASG-30 on bodyweight, the tendency to diminish weight loss did not reachsignificance probably due to the short times used, comparedto other reports (Cullen et al., 2001).
We did not compare the effect of CRH-R2 antagonist onDIA-induced alterations in naıve animals as CRH expressionincreases in the LH only after DIA and, ASG-30 antagonism ofCRH anorexigenic effects has been shown when co-injectedsimultaneously (Cullen et al., 2001). However, compared to Sanimals (dehydrated and injected with saline solution) ASG-30 administration induced the expected adaptation of theHPT axis of animals subjected to food restriction: decreasedlevels of TRHmRNA in the PVN, as well as TSH and T4 in serum(this study and Jaimes-Hoy et al., 2008). The middle dose ofASG-30 was the most effective (as Cullen et al., 2001),probably due to the partial agonist activity at high doses(Ruhmann et al., 1998).
Antagonizing CRH-R2 in the PVN of animals subjected toDIA was able to attenuate the anorexigenic effects inducedby dehydration, and restored the ability of the HPT axis toadjust to the negative energy situation. These results,together with the in vitro stimulatory effect of CRH on
pro-TRH expression, suggest that the DIA-induced activationof the HPTaxis is affected at the PVN level by CRH through theCRH-R2 receptor. It however remains to be shown that theDIA-activated CRHergic neurons of the LH, which project tothe PVN (Watts et al., 1999), target TRH synthesizing cellsand that these latter cells do express CRH-R2. The negativecorrelation found between PVN-pro-TRH mRNA levels andfood intake gives further support for TRH anorectic effects(Choi et al., 2002; Schuhler et al., 2007).
This animal model of anorexia is induced by dehydrationas a homeostatic adaptative response to the hydric imbal-ance, thus avoiding an increase in osmolality levels, thatwould be provoked by further feeding. Afferents from regionsinvolved in the sensory perception of salt may also influencethis (Watts et al., 2007). This paradigm differs from humanetiology of anorexia nervosa (Watts et al., 2007). However,some of the altered parameters of HPT axis are similar tothose described in anorexic patients (i.e., normal TSH and T4levels despite the reduced food intake). The previous sugges-tions of a central deregulation of TRH-TSH signaling (Douyonand Schteingart, 2002) seem supported by our results. It isdifficult to mimic human psychobehavioral disease such asanorexia in animal models; for example, restraint stress orLPS injection diminish food intake but also, TRH expressionand TSH release, in contrast to anxiety-producing paradigmswhere pro-TRHmRNA levels are increased in the PVN (Gutier-rez-Mariscal et al., 2008). The failure of HPT adaptationduring anorexia becomes deleterious to the subject asgreater body weight loss is observed; our results providesome hope for the use of CRH-R2 antagonists in anorexicpatients as this disorder constitutes a growing health problemwhere the excessive weight loss without apparent decreasein metabolic rate is potentially lethal.
Role of the funding source
Funding for this study was provided by CONACYT 61410(PdeG), 43503Q (PJB) and DGAPA-UNAM IN215507 (PJB). Noneof them had any further role in study design; in the collec-tion, analysis and interpretation of data; in the writing of thereport or in the decision to submit the paper for publication.
Conflict of interest
None declared.
Acknowledgements
The authors thank the technical support of QFB MiguelCisneros and to Raul Cardoso and Jose Luis Calderon for theirphotographic and drawing work.
References
Aguilar-Valles, A., Sanchez, E., de Gortari, P., Balderas, I., Ramirez-Amaya, V., Bermudez-Rattoni, F., Joseph-Bravo, P., 2005. Analysisof the stress response in rats trained in the water-maze: differ-ential expression of corticotropin-releasing hormone, CRH-R1,glucocorticoid receptors and brain-derived neurotrophic factor inlimbic regions. Neuroendocrinology 82, 306—319.
Aguilera, G., Millan, M.A., Hauger, R.L., Catt, K.J., 1987. Cortico-tropin-releasing factor receptors: distribution and regulation in
CRH activates PVN TRH expression during dehydration 269

Author's personal copy
brain, pituitary, and peripheral tissues. Ann. N. Y. Acad. Sci. 512,48—66.
Arafah, B.M., 2001. Increased need for thyroxine in women withhypothyroidism during estrogen therapy. N. Engl. J. Med. 344,1743—1749.
Arase, K., York, D.A., Shimizu, H., Shargill, N., Bray, G.A., 1988.Effects of corticotropin-releasing factor on food intake and brownadipose tissue thermogenesis in rats. Am. J. Physiol. 255, E255—E259.
Bale, T.L., Contarino, A., Smith, G.W., Chan, R., Gold, L.H., Saw-chenko, P.E., Koob, G.F., Vale, W.W., Lee, K.F., 2000. Micedeficient for corticotropin-releasing hormone receptor-2 displayanxiety-like behaviour and are hypersensitive to stress. Nat.Genet. 24, 410—414.
Barna, I., Balint, E., Baranyi, J., Bakos, N., Makara, G.B., Haller, J.,2003. Gender-specific effect of maternal deprivation on anxietyand corticotropin-releasing hormone mRNA expression in rats.Brain Res. Bull. 62, 85—91.
Bianco, A.C., Maia, A.L., da Silva, W.S., Christoffolete, M.A., 2005.Adaptive activation of thyroid hormone and energy expenditure.Biosci. Rep. 25, 191—208.
Blake, N.G., Eckland, D.J., Foster, O.J., Lightman, S.L., 1991.Inhibition of hypothalamic thyrotropin-releasing hormone mes-senger ribonucleic acid during food deprivation. Endocrinology129, 2714—2718.
Bornstein, S.R., Webster, E.L., Torpy, D.J., Richman, S.J., Mitsiades,N., Igel, M., Lewis, D.B., Rice, K.C., Joost, H.G., Tsokos, M.,Chrousos, G.P., 1998. Chronic effects of a nonpeptide cortico-tropin-releasing hormone type I receptor antagonist on pituitary—adrenal function, body weight, and metabolic regulation. Endo-crinology 139, 1546—1555.
Brady, L.S., Smith, M.A., Gold, P.W., Herkenham, M., 1990. Alteredexpression of hypothalamic neuropeptide mRNAs in food-restricted and food-deprived rats. Neuroendocrinology 52,441—447.
Brown, M.R., Fisher, L.A., Spiess, J., Rivier, J., Rivier, C., Vale, W.,1982. Comparison of the biologic actions of corticotropin-releas-ing factor and sauvagine. Regul. Pept. 4, 107—114.
Buwalda, B., de Boer, S.F., Van Kalkeren, A.A., Koolhaas, J.M., 1997.Physiological and behavioral effects of chronic intracerebroven-tricular infusion of corticotropin-releasing factor in the rat.Psychoneuroendocrinology 22, 297—309.
Chalmers, D.T., Lovenberg, T.W., De Souza, E.B., 1995. Localizationof novel corticotropin-releasing factor receptor (CRF2) mRNAexpression to specific subcortical nuclei in rat brain: comparisonwith CRF1 receptor mRNA expression. J. Neurosci. 15, 6340—6350.
Charli, J.L., Cruz, C., Redondo, J.L., Guerra, C., Joseph-Bravo, P.,1995. Homologous conditioned medium enhances expression ofTRH in hypothalamic neurons in primary culture. Dev. Brain Res.89, 155—160.
Choi, Y.H., Hartzell, D., Azain, M.J., Baile, C.A., 2002. TRH decreasesfood intake and increases water intake and body temperature inrats. Physiol. Behav. 77, 1—4.
Chomczynski, P., Sacchi, N., 1987. Single-step method of RNA isola-tion by acid guanidinium thiocyanate—phenol—chloroformextraction. Anal. Biochem. 162, 156—159.
Cizza, G., Brady, L.S., Esclapes, M.E., Blackman, M.R., Gold, P.W.,Chrousos, G.P., 1996. Age and gender influence basal and stress-modulated hypothalamic—pituitary—thyroidal function in Fischer344/N rats. Neuroendocrinology 64, 440—448.
Coppola, A., Hughes, J., Esposito, E., Schiavo, L., Meli, R., Diano, S.,2005. Suppression of hypothalamic deiodinase type II activityblunts TRH mRNA decline during fasting. FEBS Lett. 579, 4654—4658.
Cote-Velez, A., Perez-Martinez, L., Diaz-Gallardo, M.Y., Perez-Mon-ter, C., Carreon-Rodriguez, A., Charli, J.L., Joseph-Bravo, P.,2005. Dexamethasone represses cAMP rapid upregulation of
TRH gene transcription: identification of a composite glucocorti-coid response element and a cAMP response element in TRHpromoter. J. Mol. Endocrinol. 34, 177—197.
Cullen, M.J., Ling, N., Foster, A.C., Pelleymounter, M.A., 2001.Urocortin, corticotropin releasing factor-2 receptors and energybalance. Endocrinology 142, 992—999.
Curtis, K.S., Davis, L.M., Johnson, A.L., Therrien, K.L., Contreras,R.J., 2004. Sex differences in behavioral taste responses to andingestion of sucrose and NaCl solutions by rats. Physiol. Behav. 80,657—664.
de Gortari, P., Uribe, R.M., Garcia-Vazquez, A., Aguilar-Valles, A.,Martinez, A., Valdes, A., Charli, J.L., Fernandez-Guardiola, A.,Joseph-Bravo, P., 2006. Amygdala kindling differentially regu-lates the expression of the elements involved in TRH transmis-sion. Neurochem. Int. 48, 31—42.
DeSouza, E., Grigoriadis, D., 1990. Corticotropin releasing factor(CRF) receptors in the rat central nervous system: autoradio-graphic localization studies. In: DeSouza, E., Nemeroff, C.(Eds.), Corticotropin Releasing Factor: Basic and Clinical Stu-dies of a Neuropeptide. CRC Press, Boca Raton, pp. 69—90.
Donda, A., Reymond, M.J., Zurich, M.G., Lemarchand-Beraud, T.,1987. Influence of sex and age on T3 receptors and T3 concentra-tion in the pituitary gland of the rat: consequences on TSHsecretion. Mol. Cell. Endocrinol. 54, 29—34.
Doufas, A.G., Mastorakos, G., 2000. The hypothalamic—pituitary—thyroid axis and the female reproductive system. Ann. N. Y. Acad.Sci. 900, 65—76.
Douyon, L., Schteingart, D.E., 2002. Effect of obesity and starvationon thyroid hormone, growth hormone, and cortisol secretion.Endocrinol. Metab. Clin. North. Am. 31, 173—189.
Emerson, C.H., Cohen 3rd, J.H., Young, R.A., Alex, S., Fang, S.L.,1990. Gender-related differences of serum thyroxine-bindingproteins in the rat. Acta Endocrinol. (Copenh) 123, 72—78.
Emerson, C.H., Seiler, C.M., Alex, S., Fang, S.L., Mori, Y., DeVito,W.J., 1993. Gene expression and serum thyroxine-binding globu-lin are regulated by adrenal status and corticosterone in the rat.Endocrinology 133, 1192—1196.
Espinosa, V.P., Ferrini, M., Shen, X., Lutfy, K., Nillni, E.A., Fried-man, T.C., 2007. Cellular colocalization and coregulationbetween hypothalamic pro-TRH and prohormone convertasesin hypothyroidism. Am. J. Physiol. Endocrinol. Metab. 292,E175—E186.
Fekete, C., Kelly, J., Mihaly, E., Sarkar, S., Rand, W.M., Legradi, G.,Emerson, C.H., Lechan, R.M., 2001. Neuropeptide Y has a centralinhibitory action on the hypothalamic—pituitary—thyroid axis.Endocrinology 142, 2606—2613.
Fekete, C., Sarkar, S., Rand, W.M., Harney, J.W., Emerson, C.H.,Bianco, A.C., Beck-Sickinger, A., Lechan, R.M., 2002. Neuropep-tide Y1 and Y5 receptors mediate the effects of neuropeptide Yonthe hypothalamic—pituitary—thyroid axis. Endocrinology 143,4513—4519.
Gayle, D.A., Desai, M., Casillas, E., Beloosesky, R., Ross, M.G., 2006.Gender-specific orexigenic and anorexigenic mechanisms in rats.Life Sci. 79, 1531—1536.
Gutierrez-Mariscal, M., de Gortari, P., Lopez-Rubalcava, C., Marti-nez, A., Joseph-Bravo, P., 2008. Analysis of the anxiolytic-likeeffect of TRH and the response of amygdalar TRHergic neurons inanxiety. Psychoneuroendocrinology 33, 198—213.
Heinrichs, S.C., Menzaghi, F., Pich, E.M., Hauger, R.L., Koob, G.F.,1993. Corticotropin-releasing factor in the paraventricularnucleus modulates feeding induced by neuropeptide Y. BrainRes. 611, 18—24.
Hillhouse, E.W., Grammatopoulos, D.K., 2006. The molecularmechanisms underlying the regulation of the biological activityof corticotropin-releasing hormone receptors: implications forphysiology and pathophysiology. Endocr. Rev. 27, 260—286.
Hisano, S., Fukui, Y., Chikamori-Aoyama, M., Aizawa, T., Shibasaki, T.,1993. Reciprocal synaptic relations between CRF-immunoreactive-
270 P. de Gortari et al.

Author's personal copy
and TRH-immunoreactive neurons in the paraventricular nucleusof the rat hypothalamus. Brain Res. 620, 343—346.
Imaki, T., Naruse, M., Harada, S., Chikada, N., Imaki, J., Onodera,H., Demura, H., Vale,W., 1996. Corticotropin-releasing factor up-regulates its own receptor mRNA in the paraventricular nucleus ofthe hypothalamus. Mol. Brain Res. 38, 166—170.
Ishikawa, K., Taniguchi, Y., Kurosumi, K., Suzuki, M., 1986. Origin ofseptal thyrotropin-releasing hormone in the rat. Neuroendocri-nology 44, 54—58.
Isse, T., Ueta, Y., Serino, R., Noguchi, J., Yamamoto, Y., Nomura, M.,Shibuya, I., Lightman, S.L., Yamashita, H., 1999. Effects of leptinon fasting-induced inhibition of neuronal nitric oxide synthasemRNA in the paraventricular and supraoptic nuclei of rats. BrainRes. 846, 229—235.
Jaimes-Hoy, L., Joseph-Bravo, P., de Gortari, P., 2008. Differentialresponse of TRHergic neurons of the hypothalamic paraventricu-lar nucleus (PVN) in female animals submitted to food-restrictionor dehydration-induced anorexia and cold exposure. Horm.Behav. 53, 366—377.
Joseph-Bravo, P., Perez-Martinez, L., Lezama, L., Morales-Chapa, C.,Charli, J.L., 2002. An improved method for the expression of TRHin serum-supplemented primary cultures of fetal hypothalamiccells. Brain Res. Protoc. 9, 93—104.
Kageyama, K., Li, C., Vale, W.W., 2003. Corticotropin-releasingfactor receptor type 2messenger ribonucleic acid in rat pituitary:localization and regulation by immune challenge, restraint stress,and glucocorticoids. Endocrinology 144, 1524—1532.
Kakucska, I., Romero, L.I., Clark, B.D., Rondeel, J.M., Qi, Y., Alex, S.,Emerson, C.H., Lechan, R.M., 1994. Suppression of thyrotropin-releasing hormone gene expression by interleukin-1-beta in therat: implications for nonthyroidal illness. Neuroendocrinology 59,129—137.
Karydis, I., Tolis, G., 1998. Orexis, anorexia, and thyrotropin-releas-ing hormone. Thyroid 8, 947—950.
Kelly, A.B., Watts, A.G., 1996. Mediation of dehydration-inducedpeptidergic gene expression in the rat lateral hypothalamic areaby forebrain afferent projections. J. Comp. Neurol. 370, 231—246.
Krahn, D.D., Gosnell, B.A., Majchrzak, M.J., 1990. The anorecticeffects of CRH and restraint stress decrease with repeated expo-sures. Biol. Psychiatry 27, 1094—1102.
Leal, A.M., Moreira, A.C., 1997. Daily variation of plasma testoster-one, androstenedione, and corticosterone in rats under foodrestriction. Horm. Behav. 31, 97—100.
Lechan, R.M., Fekete, C., 2006a. The TRH neuron: a hypothalamicintegrator of energy metabolism. Prog. Brain Res. 153, 209—235.
Lechan, R.M., Fekete, C., 2006b. Role of melanocortin signaling inthe regulation of the hypothalamic—pituitary—thyroid (HPT) axis.Peptides 27, 310—325.
LeFeuvre, R.A., Rothwell, N.J., Stock, M.J., 1987. Activation ofbrown fat thermogenesis in response to central injection ofcorticotropin releasing hormone in the rat. Neuropharmacology26, 1217—1221.
Legradi, G., Emerson, C.H., Ahima, R.S., Flier, J.S., Lechan, R.M.,1997. Leptin prevents fasting-induced suppression of prothyro-tropin-releasing hormone messenger ribonucleic acid in neuronsof the hypothalamic paraventricular nucleus. Endocrinology 138,2569—2576.
Leibowitz, S.F., Wortley, K.E., 2004. Hypothalamic control of energybalance: different peptides, different functions. Peptides 25,473—504.
Lovenberg, T.W., Liaw, C.W., Grigoriadis, D.E., Clevenger, W., Chal-mers, D.T., De Souza, E.B., Oltersdorf, T., 1995. Cloning andcharacterization of a functionally distinct corticotropin-releasingfactor receptor subtype from rat brain. Proc. Natl. Acad. Sci.U.S.A. 92, 836—840.
Lunga, P., Herbert, J., 2004. 17Beta-oestradiol modulates glucocor-ticoid, neural and behavioural adaptations to repeated restraintstress in female rats. J. Neuroendocrinol. 16, 776—785.
Makino, S., Nishiyama, M., Asaba, K., Gold, P.W., Hashimoto, K., 1998.Altered expression of type 2 CRH receptor mRNA in the VMH byglucocorticoids and starvation. Am. J. Physiol. 275, R1138—R1145.
Mansi, J.A., Rivest, S., Drolet, G., 1996. Regulation of corticotropin-releasing factor type 1 (CRF1) receptor messenger ribonucleicacid in the paraventricular nucleus of rat hypothalamus by exo-genous CRF. Endocrinology 137, 4619—4629.
Marassi, M.P., Fortunato, R.S., da Silva, A.C., Pereira, V.S., Carvalho,D.P., Rosenthal, D., da Costa, V.M., 2007. Sexual dimorphism inthyroid function and type 1 iodothyronine deiodinase activity inpre-pubertal and adult rats. J. Endocrinol. 192, 121—130.
Menzaghi, F., Heinrichs, S.C., Pich, E.M., Tilders, F.J., Koob, G.F.,1993. Functional impairment of hypothalamic corticotropin-releasing factor neurons with immunotargeted toxins enhancesfood intake induced by neuropeptide Y. Brain Res. 618, 76—82.
Mogi, K., Funabashi, T., Mitsushima, D., Hagiwara, H., Kimura, F.,2005. Sex difference in the response of melanin-concentratinghormone neurons in the lateral hypothalamic area to glucose, asrevealed by the expression of phosphorylated cyclic adenosine30,50-monophosphate response element-binding protein. Endocri-nology 146, 3325—3333.
Morley, J.E., Levine, A.S., 1982. Corticotrophin releasing factor,grooming and ingestive behavior. Life Sci. 31, 1459—1464.
Mountjoy, K.G., Mortrud, M.T., Low, M.J., Simerly, R.B., Cone, R.D.,1994. Localization of the melanocortin-4 receptor (MC4-R) inneuroendocrine and autonomic control circuits in the brain.Mol. Endocrinol. 8, 1298—1308.
Nequin, L.G., Alvarez, J., Schwartz, N.B., 1979. Measurement ofserum steroid and gonadotropin levels and uterine and ovarianvariables throughout 4 day and 5 day estrous cycles in the rat.Biol. Reprod. 20, 659—670.
Pacak, K., Palkovits, M., 2001. Stressor specificity of central neu-roendocrine responses: implications for stress-related disorders.Endocr. Rev. 22, 502—548.
Paxinos, G., Watson, C., 2005. The Rat Brain in Stereotaxic Coordi-nates. Elsevier Academic Press, San Diego, CA.
Perez-Martinez, L., Carreon-Rodriguez, A., Gonzalez-Alzati, M.E.,Morales, C., Charli, J.L., Joseph-Bravo, P., 1998. Dexametha-sone rapidly regulates TRH mRNA levels in hypothalamic cellcultures: interaction with the cAMP pathway. Neuroendocrinol-ogy 68, 345—354.
Prokai, L., 2002. Central nervous system effects of thyrotropin-releasing hormone and its analogues: opportunities and perspec-tives for drug discovery and development. Prog. Drug Res. 59,133—169.
Rabadan-Diehl, C., Kiss, A., Camacho, C., Aguilera, G., 1996. Reg-ulation of messenger ribonucleic acid for corticotropin releasinghormone receptor in the pituitary during stress. Endocrinology137, 3808—3814.
Rivest, S., Deshaies, Y., Richard, D., 1989. Effects of corticotropin-releasing factor on energy balance in rats are sex dependent. Am.J. Physiol. 257, R1417—R1422.
Rohner-Jeanrenaud, F., Walker, C.D., Greco-Perotto, R., Jeanre-naud, B., 1989. Central corticotropin-releasing factor adminis-tration prevents the excessive body weight gain of geneticallyobese (fa/fa) rats. Endocrinology 124, 733—739.
Rondeel, J.M., Heide, R., de Greef, W.J., van Toor, H., van Haas-teren, G.A., Klootwijk, W., Visser, T.J., 1992. Effect of starvationand subsequent refeeding on thyroid function and release ofhypothalamic thyrotropin-releasing hormone. Neuroendocrinol-ogy 56, 348—353.
Ruhmann, A., Bonk, I., Lin, C.R., Rosenfeld, M.G., Spiess, J., 1998.Structural requirements for peptidic antagonists of the cortico-tropin-releasing factor receptor (CRFR): development ofCRFR2beta-selective antisauvagine-30. Proc. Natl. Acad. Sci.U.S.A. 95, 15264—15269.
Schuhler, S., Warner, A., Finney, N., Bennett, G.W., Ebling, F.J.,Brameld, J.M., 2007. Thyrotrophin-releasing hormone decreases
CRH activates PVN TRH expression during dehydration 271

Author's personal copy
feeding and increases body temperature, activity and oxygenconsumption in Siberian hamsters. J. Neuroendocrinol. 19,239—249.
Segerson, T.P., Kauer, J., Wolfe, H.C., Mobtaker, H., Wu, P., Jackson,I.M., Lechan, R.M., 1987. Thyroid hormone regulates TRH bio-synthesis in the paraventricular nucleus of the rat hypothalamus.Science 238, 78—80.
Shelley, D.N., Dwyer, E., Johnson, C., Wittkowski, K.M., Pfaff, D.W.,2007. Interactions between estrogen effects and hunger effects inovariectomized female mice. I. Measures of arousal. Horm.Behav. 52, 546—553.
Smagin, G.N., Howell, L.A., Ryan, D.H., De Souza, E.B., Harris, R.B.,1998. The role of CRF2 receptors in corticotropin-releasing fac-tor- and urocortin-induced anorexia. Neuroreport 9, 1601—1606.
Tohei, A., Imai, A., Watanabe, G., Taya, K., 1998. Influence ofthiouracil-induced hypothyroidism on adrenal and gonadal func-tions in adult female rats. J. Vet. Med. Sci. 60, 439—446.
Turnbull, A.V., Rivier, C., 1997. Corticotropin-releasing factor (CRF)and endocrine responses to stress: CRF receptors, binding pro-tein, and related peptides. Proc. Soc. Exp. Biol. Med. 215, 1—10.
Valle, A., Catala-Niell, A., Colom, B., Garcia-Palmer, F.J., Oliver, J.,Roca, P., 2005. Sex-related differences in energy balance inresponse to caloric restriction. Am. J. Physiol. Endocrinol. Metab.289, E15—E22.
van Haasteren, G.A., Linkels, E., Klootwijk, W., van Toor, H., Ron-deel, J.M., Themmen, A.P., de Jong, F.H., Valentijn, K., Vaudry,H., Bauer, K., et al., 1995. Starvation-induced changes in thehypothalamic content of prothyrotrophin-releasing hormone(proTRH) mRNA and the hypothalamic release of proTRH-derivedpeptides: role of the adrenal gland. J. Endocrinol. 145, 143—153.
van Haasteren, G.A., Linkels, E., van Toor, H., Klootwijk, W., Kap-tein, E., de Jong, F.H., Reymond, M.J., Visser, T.J., de Greef,W.J., 1996. Effects of long-term food reduction on the hypotha-lamus—pituitary—thyroid axis in male and female rats. J. Endo-crinol. 150, 169—178.
van Haasteren, G.A., Sleddens-Linkels, E., van Toor, H., Klootwijk,W., de Jong, F.H., Visser, T.J., de Greef, W.J., 1997. Possible roleof corticosterone in the down-regulation of the hypothalamo—hypophysial—thyroid axis in streptozotocin-induced diabetesmellitus in rats. J. Endocrinol. 153, 259—267.
Vaughan, J., Donaldson, C., Bittencourt, J., Perrin, M.H., Lewis, K.,Sutton, S., Chan, R., Turnbull, A.V., Lovejoy, D., Rivier, C., et al.,1995. Urocortin, a mammalian neuropeptide related to fish uroten-sin I and to corticotropin-releasing factor. Nature 378, 287—292.
Vijayan, E., McCann, S.M., 1977. Suppression of feeding and drinkingactivity in rats following intraventricular injection of thyrotropinreleasing hormone (TRH). Endocrinology 100, 1727—1730.
Watts, A.G., Salter, D.S., Neuner, C.M., 2007. Neural network inter-actions and ingestive behavior control during anorexia. Physiol.Behav. 91, 389—396.
Watts, A.G., Sanchez-Watts, G., Kelly, A.B., 1999. Distinct patternsof neuropeptide gene expression in the lateral hypothalamic areaand arcuate nucleus are associated with dehydration-inducedanorexia. J. Neurosci. 19, 6111—6121.
Wiersinga, W.M., 2005. Nonthyroidal illness. In: Braverman, L.E.,Utiger, R.D. (Eds.), The Thyroid. Lippincott, Philadelphia, pp.246—263.
Yang, J., Tashjian Jr., A.H., 1993. Regulation of endogenous thyro-tropin-releasing hormone (TRH) receptor messenger RNA by TRHin GH4C1 cells. Mol. Endocrinol. 7, 753—758.
272 P. de Gortari et al.




















