
Organ damage in Atlantic herring larvae as a result of ocean acidification

INVESTIGATOR GROUP EXPEDITION 2006:FLEXIBLE FORAGING ECOLOGY OF A TEMPERATE HERBIVORE,
THE HERRING CALE, OLISTHOPS CYANOMELAS, IN SOUTH AUSTRALIA
S.A. SHEPHERD1 & J.L. BAKER2
1. South Australian Research and Development Institute,PO Box 120 Henley Beach, South Australia 5022
Email: [email protected]. Present address: 8 Fairfield Ave., Somerton Park, South Australia 5044
Abstract
The odacine fish, herring cale (HC), Olisthops cyanomelas, is a facultative browser, feeding mainlyon the secondary laminae of Ecklonia radiata, and the apical receptacles of the fucoids, Cystophoraspp. and Sargassum spp. Diet, daily feeding rates and time budgets of HC were examined at twoSouth Australian sites >1000 km apart and in different water movement conditions, with additionaldietary data from an exposed site. The diet and feeding behaviour were habitat-dependent. At onepartly-sheltered site with abundant Ecklonia, HC was most active immediately after dawn and towarddusk, whereas at the other more sheltered site, dominated by fucoid algae, it was inactive at dawnincreasing to a moderate level of activity in mid-morning, and peaking toward sunset. Feeding biterates were variable and mean rates ranged from 8 – 14.5 min-1 according to site. HC characteristicallyengaged in intermittent foraging bouts, interspersed with periods of inactivity, which averaged ~55% of daylight hours. Interactions with other species were few. Analysis showed no significantdifferences between small and large HC in bite rates, but significant between-site differences in biterates, and in the proportion of time spent foraging and swimming. In this study HC did not destroypatches of the kelp, Ecklonia, as it does in eastern Australia, even when the kelp comprises the maindiet, because it fed only on the distal parts of the kelp’s secondary laminae and meristem.
KEY WORDS: herbivory; herbivorous fish; Olisthops cyanomelas; Odax cyanomelas; Odacidae;Ecklonia radiata.
Introduction
Fish herbivory in temperate waters poses many questions for ecologists, such as optimal food choice,digestion and assimilation (reviewed by Choat & Clements 1998), and effect on biogenic reef structure(Andrew & Jones 1990). Some odacines, in particular, with 12 species in temperate Australasia, are knownto feed on laminarian and fucoid macro-algae, which they digest by microbial hindgut fermentation(Mountford et al. 2002). Of these 12 species, one of the largest, herring cale, Olisthops cyanomelas1
Richardson, occurs throughout southern Australia, spanning some 6 000 km from the Abrolhos Is, WesternAustralia to northern New South Wales. Yet its feeding ecology is known only from near the eastern end ofits range (Andrew & Jones 1990). These authors recorded exclusive foraging by the herring cale (HC) on thekelp, Ecklonia radiata, and the HC’s curious behaviour of clearing patches in Ecklonia forests, a behaviournever observed in southern Australia. Yet the HC lives in both kelp- and fucoid-dominated habitats, whichare widespread throughout southern Australia, and vary according to water movement (Shepherd &Womersley 1981; Edgar 1999; Edyvane & Shepherd 1999; Kendrick 1999; Steinberg & Kendrick 1999).
The purpose of this paper is to examine the generality of Andrew and Jones’ findings, by investigating theforaging and feeding behaviour of the HC, its feeding rates, selectivity, and activity patterns in differenthabitats. Such information is a first step toward a better understanding of the nutritional ecology of thisunusual herbivore and its effect on algal assemblages.
Materials and Methods
The study was carried out mainly at two South Australian sites: Site 1 ─ semi-sheltered Mooring Bay,Althorpe I. (34˚ 22΄ S; 136˚ 52΄ E.) during January and February, 2005 and 2006; Site 2 ─ sheltered Groper
147
Transactions of the Royal Society of South Australia (2008), 132(2): 147–162
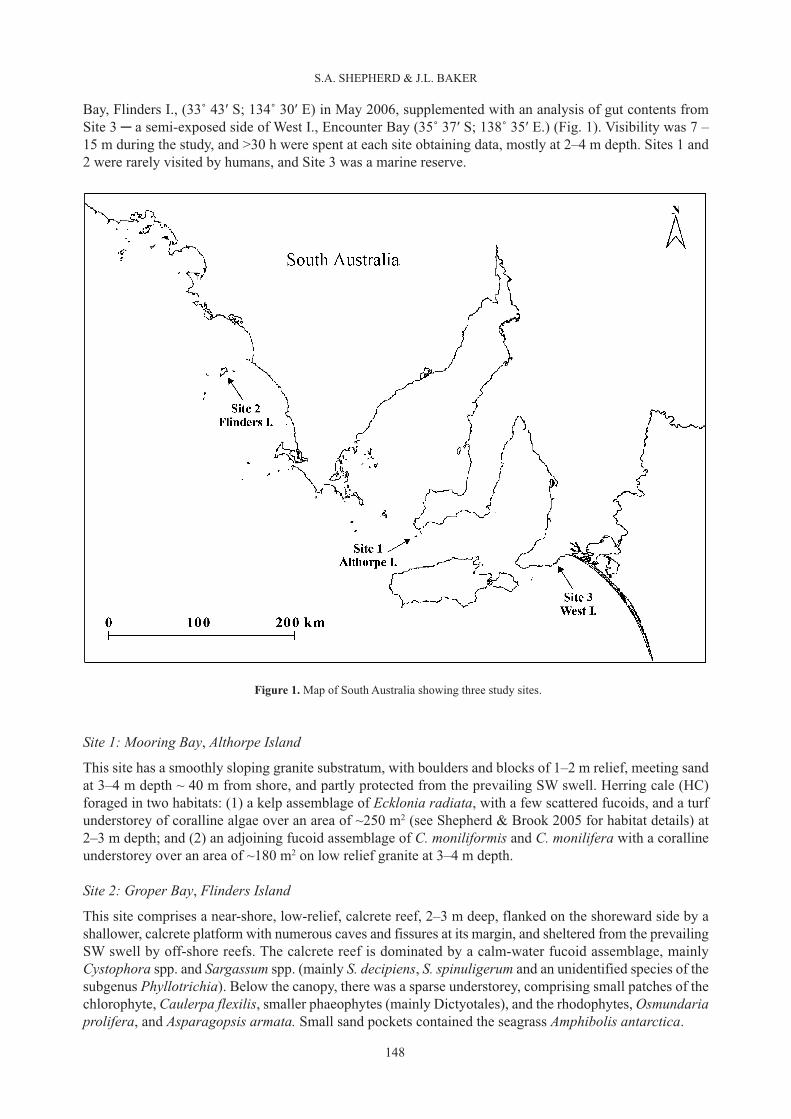
Bay, Flinders I., (33˚ 43′ S; 134˚ 30′ E) in May 2006, supplemented with an analysis of gut contents fromSite 3 ─ a semi-exposed side of West I., Encounter Bay (35˚ 37′ S; 138˚ 35′ E.) (Fig. 1). Visibility was 7 –15 m during the study, and >30 h were spent at each site obtaining data, mostly at 2–4 m depth. Sites 1 and2 were rarely visited by humans, and Site 3 was a marine reserve.
Site 1: Mooring Bay, Althorpe Island
This site has a smoothly sloping granite substratum, with boulders and blocks of 1–2 m relief, meeting sandat 3–4 m depth ~ 40 m from shore, and partly protected from the prevailing SW swell. Herring cale (HC)foraged in two habitats: (1) a kelp assemblage of Ecklonia radiata, with a few scattered fucoids, and a turfunderstorey of coralline algae over an area of ~250 m2 (see Shepherd & Brook 2005 for habitat details) at2–3 m depth; and (2) an adjoining fucoid assemblage of C. moniliformis and C. monilifera with a corallineunderstorey over an area of ~180 m2 on low relief granite at 3–4 m depth.
Site 2: Groper Bay, Flinders Island
This site comprises a near-shore, low-relief, calcrete reef, 2–3 m deep, flanked on the shoreward side by ashallower, calcrete platform with numerous caves and fissures at its margin, and sheltered from the prevailingSW swell by off-shore reefs. The calcrete reef is dominated by a calm-water fucoid assemblage, mainlyCystophora spp. and Sargassum spp. (mainly S. decipiens, S. spinuligerum and an unidentified species of thesubgenus Phyllotrichia). Below the canopy, there was a sparse understorey, comprising small patches of thechlorophyte, Caulerpa flexilis, smaller phaeophytes (mainly Dictyotales), and the rhodophytes, Osmundariaprolifera, and Asparagopsis armata. Small sand pockets contained the seagrass Amphibolis antarctica.
148
S.A. SHEPHERD & J.L. BAKER
Figure 1. Map of South Australia showing three study sites.

Site 3: West Island, Encounter Bay
This site was on the semi-exposed NE corner of the island, on sloping granitic rock dominated by thelaminarian, Ecklonia radiata, to ~ 5 m depth, with a sparse understorey mainly of smaller phaeophytes andchlorophytes (Shepherd & Womersley 1970).
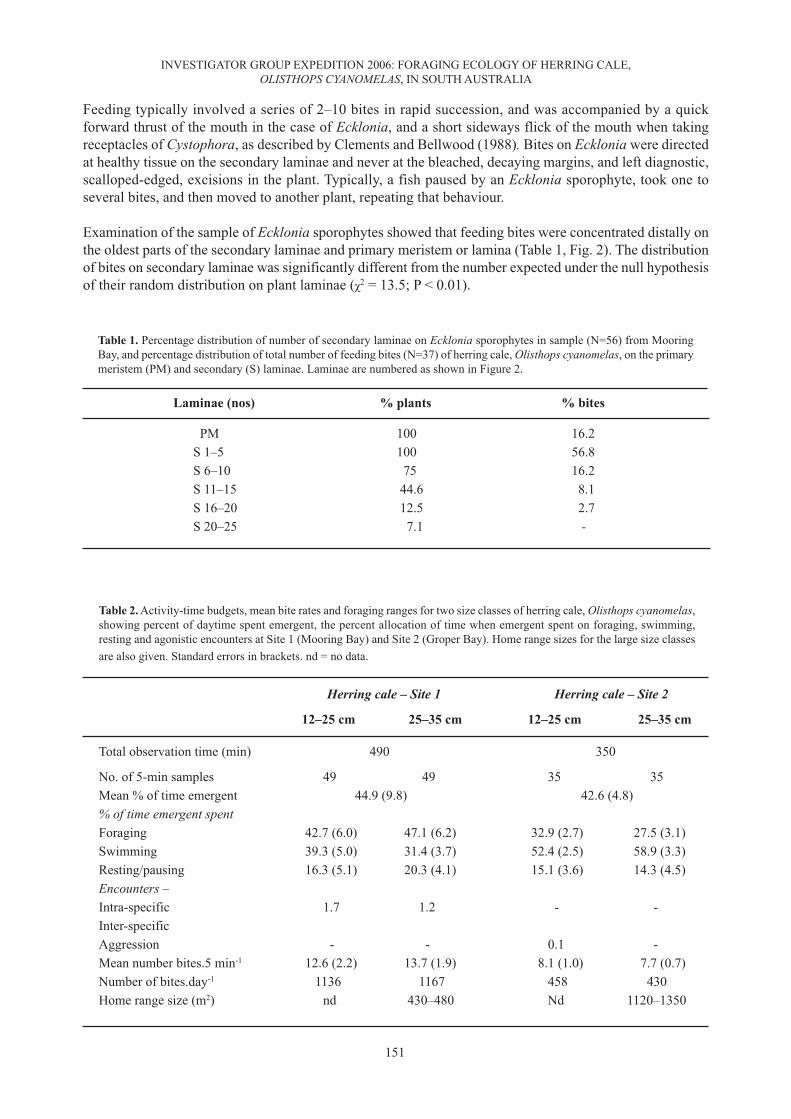
SamplingAt Site 1, 56 Ecklonia sporophytes were harvested haphazardly from the kelp forest where foraging tookplace, and examined for HC bite marks (described below). The secondary laminae were numbered sequentiallyfrom the distal end of the primary lamina i.e. with increasing age (Fig. 2), and the number of bites taken onlaterals on one side of the plant, haphazardly chosen, and on the primary lamina was counted and summed foreach lamina group. We tested the randomness of distribution of bites on secondary laminae with a χ2 test.
To test the foraging preferences of HC, we used Ivlev’s (1961) electivity coefficient, E, which measures thedegree of selection for a particular species. The equation is: E = (ri – pi) / (ri + pi) where ri is the proportionof feeding bites on taxa/habitats i relative to the total number of bites, and pi is the relative availability of thosetaxa/habitats, estimated by the proportional cover of the taxa or habitat within the home range. At Site 2 wetested a preference for Ecklonia sporophytes, although none were living at the site, by attaching threesporophytes to pieces of rock, and placing them within the foraging range of the HC. Three other sporophyteswere similarly attached to small rocks and placed outside their ranges.
Activity-time budgetFour categories of activities were recognised within two size groups of fish: 10–25 cm total length (TL); and25–35 cm TL. These activities (see Pavlov & Kasumyan 1998) were: (a) foraging, which included swimmingslowly in or close to the algal canopy, and inspecting and biting pieces of algae; (b) swimming above the algalcanopy; (c) resting or pausing briefly, often in the algal canopy; and (d) interactions with other species.
A focal fish was followed for 5 minutes, and the proportion of time spent in each activity recorded. The daywas divided into seven equal time periods, and seven 5-min observational samples of fish behaviour in small
149
INVESTIGATOR GROUP EXPEDITION 2006: FORAGING ECOLOGY OF HERRING CALE,OLISTHOPS CYANOMELAS, IN SOUTH AUSTRALIA
Figure 2. Sporophyte of Ecklonia radiatashowing numbered secondary laminae.

(< 25 cm) and large (≥25 cm) size classes obtained in each time period. At Site 1 the periods were of 2 h each,from sunrise at ~0630 h until sunset at ~1830 h, and at Site 2 they were of 1 h 30 min., from sunrise at ~ 0700h until sunset at ~ 1730 h. At sunset the fish retreated into shelter. We noted the alga or substratum at whicheach bite was directed, counted the number of bites, and measured the time swimming or resting duringfeeding bouts, and interactions with other HC individuals or other species. The behaviour of the focal fishwas noted as submissive if it retreated and aggressive if it pursued. We chose 5-min sampling periods becauseHC were usually emergent for 5–10 min bouts before submerging under cover. Fish quickly behaved neutrallyin the presence of divers/snorkellers, and during this study ~ 5 min in the beginning of the day was sufficientacceptance time.
The data on feeding bite frequencies were analysed by 2-way ANOVAs with time period of day and size-classas fixed factors at each of the two sites (1, 2). Levene’s test was used to check for homogeneity of variances,which did not differ significantly between time periods (Site 1: P=0.21; Site 2: P= 0.53). We did not comparebetween-site differences in feeding rates with ANOVA , because of the disparities between the season ofsamplings: Site 1 was sampled in late summer, when the number of daylight hours was 14, and seatemperatures were ~18ºC, whereas at Site 2 the respective figures were 10.5 h, and 16ºC. However, we didcompare the proportions of time emergent spent foraging, swimming and resting with t-tests, althoughseasonal effects might have influenced those parameters.
MappingSites 1 and 2 were mapped on an underwater slate, by using a 100 m tape, marked at 1 m intervals, and laidalong defining topographic features to measure distances along and between them. Two 100 m transect lineswere laid down in the foraging habitat used by HC, and the proportional cover of each habitat-type measuredby recording the distance under the line covered by each habitat-type. The size of the foraging ranges of fourlarge fish was estimated by the polygon method i.e. by recording on the above map the maximum spatialextent of their foraging recorded during the observation period, and calculating the area between the extremepoints. Extensive searches beyond the foraging ranges verified that fish did not make more distant excursions.
Population censusesAfter ~10 h of observations over several days, all HC at Sites 1 and 2 were recognisable individually bysize, colour and markings, and the total number present estimated. A census of the number of emergent HCwas carried out during each designated period of day by swimming twice throughout the known home ranges,so that foraging individuals missed on the first count would likely be included in the second. The methodestimates the mean proportion of the emergent population throughout the day (Shepherd 2006). From the dataon bite rates, and proportion of the population emergent for each time period, the daily number of feedingbites was calculated.
DietFollowing the study at each site, samples were taken by net or spear late in the day for gut content analysisin the laboratory. In this analysis gut contents were placed on a gridded Petri dish, examined under a binocularmicroscope, and algae identified to species where possible. The percentage volumetric abundance of algalcategories was estimated by the points method (Windell 1968).
Results
Foraging and feeding behaviour, feeding rates and selectivitySite 1―Mooring BayIn all, 16 female and 2 male HC were resident at this site (see below). The males were seldom seen, and nofeeding data were obtained for them. When emergent, feeding was highly selective. From dawn until lateafternoon HC fed mainly on Ecklonia sporophytes, and then, in the last 2 h of daylight, all HC left the kelpassemblage for the adjoining fucoid assemblage and fed on tips of the fucoids, Cystophora moniliformis andC. monilifera. At dusk they disappeared into crevices and holes.
150
S.A. SHEPHERD & J.L. BAKER
Feeding typically involved a series of 2–10 bites in rapid succession, and was accompanied by a quickforward thrust of the mouth in the case of Ecklonia, and a short sideways flick of the mouth when takingreceptacles of Cystophora, as described by Clements and Bellwood (1988). Bites on Ecklonia were directedat healthy tissue on the secondary laminae and never at the bleached, decaying margins, and left diagnostic,scalloped-edged, excisions in the plant. Typically, a fish paused by an Ecklonia sporophyte, took one toseveral bites, and then moved to another plant, repeating that behaviour.
Examination of the sample of Ecklonia sporophytes showed that feeding bites were concentrated distally onthe oldest parts of the secondary laminae and primary meristem or lamina (Table 1, Fig. 2). The distributionof bites on secondary laminae was significantly different from the number expected under the null hypothesisof their random distribution on plant laminae (χ2 = 13.5; P < 0.01).
151
INVESTIGATOR GROUP EXPEDITION 2006: FORAGING ECOLOGY OF HERRING CALE,OLISTHOPS CYANOMELAS, IN SOUTH AUSTRALIA
Table 1. Percentage distribution of number of secondary laminae on Ecklonia sporophytes in sample (N=56) from MooringBay, and percentage distribution of total number of feeding bites (N=37) of herring cale, Olisthops cyanomelas, on the primarymeristem (PM) and secondary (S) laminae. Laminae are numbered as shown in Figure 2.
Laminae (nos) % plants % bites
PM 100 16.2S 1–5 100 56.8S 6–10 75 16.2S 11–15 44.6 8.1S 16–20 12.5 2.7S 20–25 7.1 -
Table 2. Activity-time budgets, mean bite rates and foraging ranges for two size classes of herring cale, Olisthops cyanomelas,showing percent of daytime spent emergent, the percent allocation of time when emergent spent on foraging, swimming,resting and agonistic encounters at Site 1 (Mooring Bay) and Site 2 (Groper Bay). Home range sizes for the large size classesare also given. Standard errors in brackets. nd = no data.
Herring cale – Site 1 Herring cale – Site 2
12–25 cm 25–35 cm 12–25 cm 25–35 cm
Total observation time (min) 490 350
No. of 5-min samples 49 49 35 35Mean % of time emergent 44.9 (9.8) 42.6 (4.8)% of time emergent spentForaging 42.7 (6.0) 47.1 (6.2) 32.9 (2.7) 27.5 (3.1)Swimming 39.3 (5.0) 31.4 (3.7) 52.4 (2.5) 58.9 (3.3)Resting/pausing 16.3 (5.1) 20.3 (4.1) 15.1 (3.6) 14.3 (4.5)Encounters –Intra-specific 1.7 1.2 - -Inter-specificAggression - - 0.1 -Mean number bites.5 min-1 12.6 (2.2) 13.7 (1.9) 8.1 (1.0) 7.7 (0.7)Number of bites.day-1 1136 1167 458 430Home range size (m2) nd 430–480 Nd 1120–1350
152
S.A. SHEPHERD & J.L. BAKER
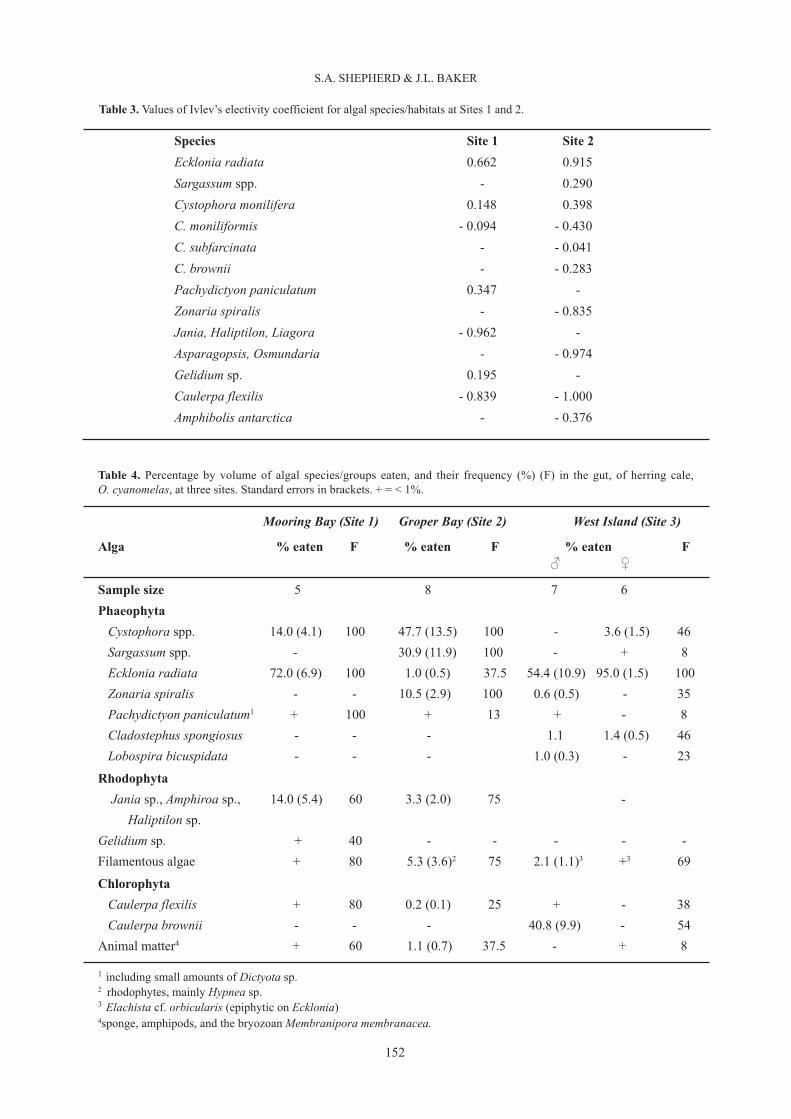
Table 3. Values of Ivlev’s electivity coefficient for algal species/habitats at Sites 1 and 2.
Species Site 1 Site 2Ecklonia radiata 0.662 0.915Sargassum spp. - 0.290Cystophora monilifera 0.148 0.398C. moniliformis - 0.094 - 0.430C. subfarcinata - - 0.041C. brownii - - 0.283Pachydictyon paniculatum 0.347 -Zonaria spiralis - - 0.835Jania, Haliptilon, Liagora - 0.962 -Asparagopsis, Osmundaria - - 0.974Gelidium sp. 0.195 -Caulerpa flexilis - 0.839 - 1.000Amphibolis antarctica - - 0.376
Table 4. Percentage by volume of algal species/groups eaten, and their frequency (%) (F) in the gut, of herring cale,O. cyanomelas, at three sites. Standard errors in brackets. + = < 1%.
Mooring Bay (Site 1) Groper Bay (Site 2) West Island (Site 3)
Alga % eaten F % eaten F % eaten F♂ ♀
Sample size 5 8 7 6Phaeophyta
Cystophora spp. 14.0 (4.1) 100 47.7 (13.5) 100 - 3.6 (1.5) 46Sargassum spp. - 30.9 (11.9) 100 - + 8Ecklonia radiata 72.0 (6.9) 100 1.0 (0.5) 37.5 54.4 (10.9) 95.0 (1.5) 100Zonaria spiralis - - 10.5 (2.9) 100 0.6 (0.5) - 35Pachydictyon paniculatum1 + 100 + 13 + - 8Cladostephus spongiosus - - - 1.1 1.4 (0.5) 46Lobospira bicuspidata - - - 1.0 (0.3) - 23
RhodophytaJania sp., Amphiroa sp., 14.0 (5.4) 60 3.3 (2.0) 75 -
Haliptilon sp.Gelidium sp. + 40 - - - - -Filamentous algae + 80 5.3 (3.6)2 75 2.1 (1.1)3 +3 69
ChlorophytaCaulerpa flexilis + 80 0.2 (0.1) 25 + - 38Caulerpa brownii - - - 40.8 (9.9) - 54
Animal matter4 + 60 1.1 (0.7) 37.5 - + 8
1 including small amounts of Dictyota sp.2 rhodophytes, mainly Hypnea sp.3 Elachista cf. orbicularis (epiphytic on Ecklonia)4sponge, amphipods, and the bryozoan Membranipora membranacea.
153
INVESTIGATOR GROUP EXPEDITION 2006: FORAGING ECOLOGY OF HERRING CALE,OLISTHOPS CYANOMELAS, IN SOUTH AUSTRALIA
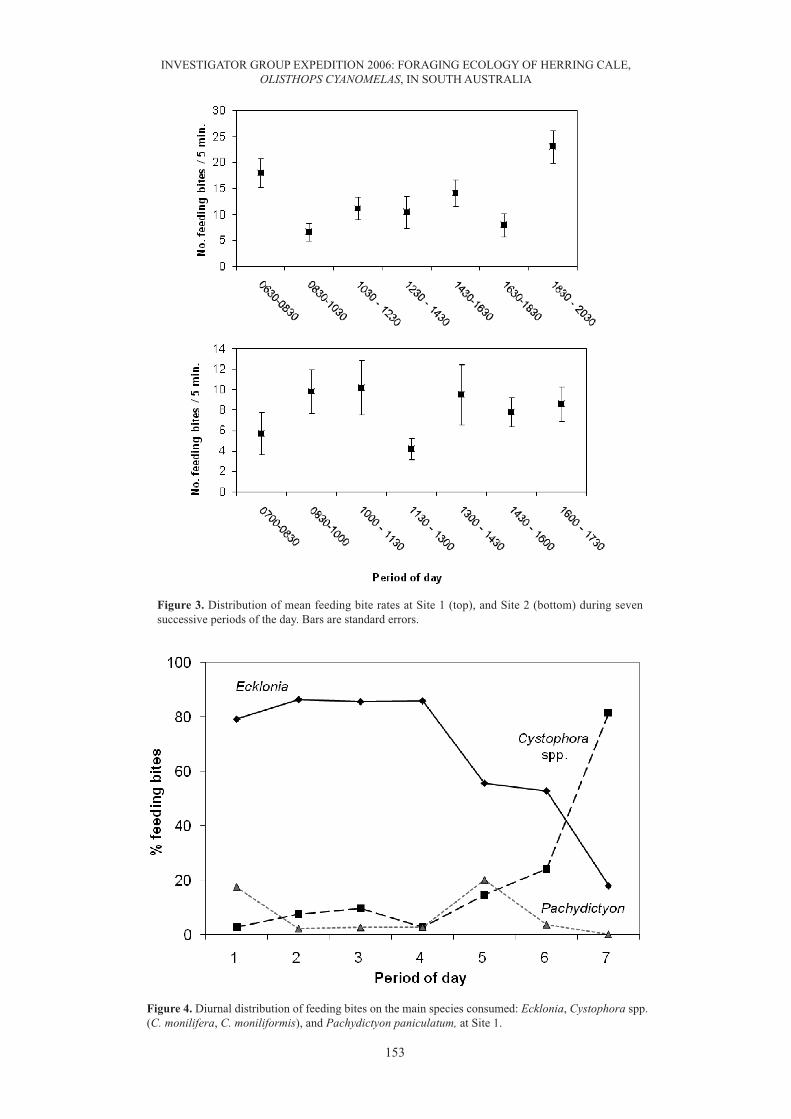
Figure 3. Distribution of mean feeding bite rates at Site 1 (top), and Site 2 (bottom) during sevensuccessive periods of the day. Bars are standard errors.
Figure 4. Diurnal distribution of feeding bites on the main species consumed: Ecklonia, Cystophora spp.(C. monilifera, C. moniliformis), and Pachydictyon paniculatum, at Site 1.
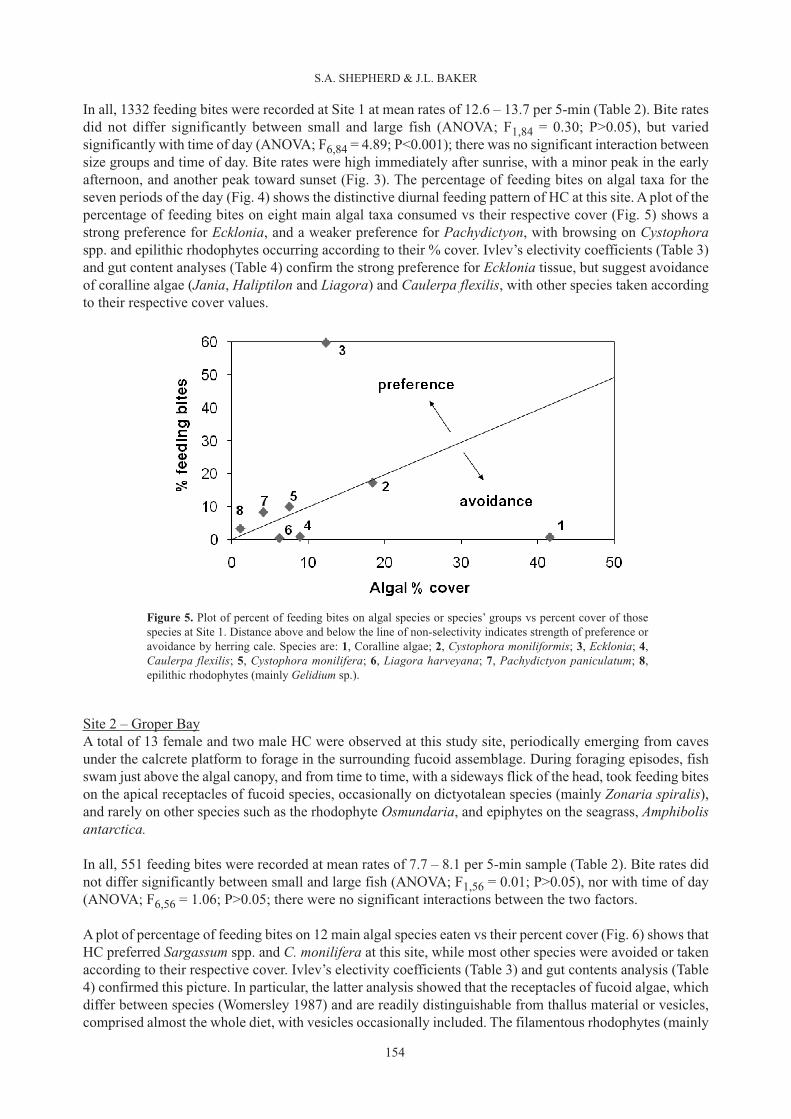
In all, 1332 feeding bites were recorded at Site 1 at mean rates of 12.6 – 13.7 per 5-min (Table 2). Bite ratesdid not differ significantly between small and large fish (ANOVA; F1,84 = 0.30; P>0.05), but variedsignificantly with time of day (ANOVA; F6,84 = 4.89; P<0.001); there was no significant interaction betweensize groups and time of day. Bite rates were high immediately after sunrise, with a minor peak in the earlyafternoon, and another peak toward sunset (Fig. 3). The percentage of feeding bites on algal taxa for theseven periods of the day (Fig. 4) shows the distinctive diurnal feeding pattern of HC at this site. A plot of thepercentage of feeding bites on eight main algal taxa consumed vs their respective cover (Fig. 5) shows astrong preference for Ecklonia, and a weaker preference for Pachydictyon, with browsing on Cystophoraspp. and epilithic rhodophytes occurring according to their % cover. Ivlev’s electivity coefficients (Table 3)and gut content analyses (Table 4) confirm the strong preference for Ecklonia tissue, but suggest avoidanceof coralline algae (Jania, Haliptilon and Liagora) and Caulerpa flexilis, with other species taken accordingto their respective cover values.
Site 2 – Groper BayA total of 13 female and two male HC were observed at this study site, periodically emerging from cavesunder the calcrete platform to forage in the surrounding fucoid assemblage. During foraging episodes, fishswam just above the algal canopy, and from time to time, with a sideways flick of the head, took feeding biteson the apical receptacles of fucoid species, occasionally on dictyotalean species (mainly Zonaria spiralis),and rarely on other species such as the rhodophyte Osmundaria, and epiphytes on the seagrass, Amphibolisantarctica.
In all, 551 feeding bites were recorded at mean rates of 7.7 – 8.1 per 5-min sample (Table 2). Bite rates didnot differ significantly between small and large fish (ANOVA; F1,56 = 0.01; P>0.05), nor with time of day(ANOVA; F6,56 = 1.06; P>0.05; there were no significant interactions between the two factors.
A plot of percentage of feeding bites on 12 main algal species eaten vs their percent cover (Fig. 6) shows thatHC preferred Sargassum spp. and C. monilifera at this site, while most other species were avoided or takenaccording to their respective cover. Ivlev’s electivity coefficients (Table 3) and gut contents analysis (Table4) confirmed this picture. In particular, the latter analysis showed that the receptacles of fucoid algae, whichdiffer between species (Womersley 1987) and are readily distinguishable from thallus material or vesicles,comprised almost the whole diet, with vesicles occasionally included. The filamentous rhodophytes (mainly
154
S.A. SHEPHERD & J.L. BAKER
Figure 5. Plot of percent of feeding bites on algal species or species’ groups vs percent cover of thosespecies at Site 1. Distance above and below the line of non-selectivity indicates strength of preference oravoidance by herring cale. Species are: 1, Coralline algae; 2, Cystophora moniliformis; 3, Ecklonia; 4,Caulerpa flexilis; 5, Cystophora monilifera; 6, Liagora harveyana; 7, Pachydictyon paniculatum; 8,epilithic rhodophytes (mainly Gelidium sp.).
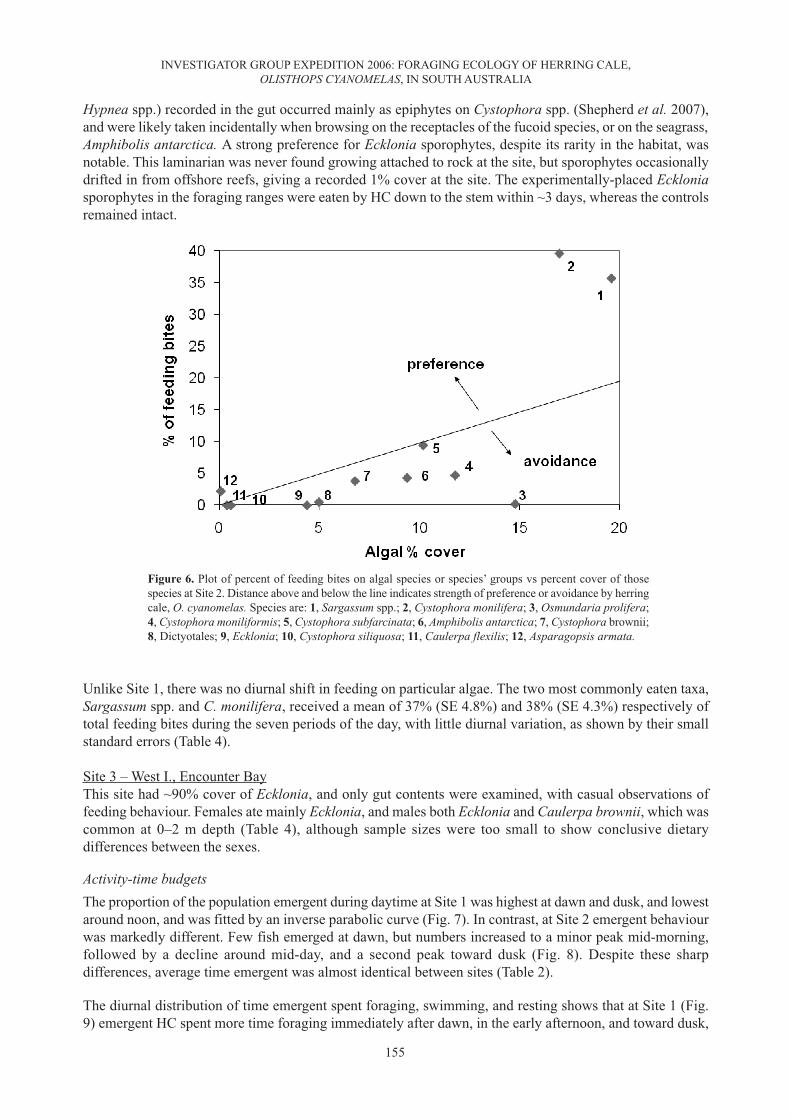
Hypnea spp.) recorded in the gut occurred mainly as epiphytes on Cystophora spp. (Shepherd et al. 2007),and were likely taken incidentally when browsing on the receptacles of the fucoid species, or on the seagrass,Amphibolis antarctica. A strong preference for Ecklonia sporophytes, despite its rarity in the habitat, wasnotable. This laminarian was never found growing attached to rock at the site, but sporophytes occasionallydrifted in from offshore reefs, giving a recorded 1% cover at the site. The experimentally-placed Eckloniasporophytes in the foraging ranges were eaten by HC down to the stem within ~3 days, whereas the controlsremained intact.
Unlike Site 1, there was no diurnal shift in feeding on particular algae. The two most commonly eaten taxa,Sargassum spp. and C. monilifera, received a mean of 37% (SE 4.8%) and 38% (SE 4.3%) respectively oftotal feeding bites during the seven periods of the day, with little diurnal variation, as shown by their smallstandard errors (Table 4).
Site 3 – West I., Encounter BayThis site had ~90% cover of Ecklonia, and only gut contents were examined, with casual observations offeeding behaviour. Females ate mainly Ecklonia, and males both Ecklonia and Caulerpa brownii, which wascommon at 0–2 m depth (Table 4), although sample sizes were too small to show conclusive dietarydifferences between the sexes.
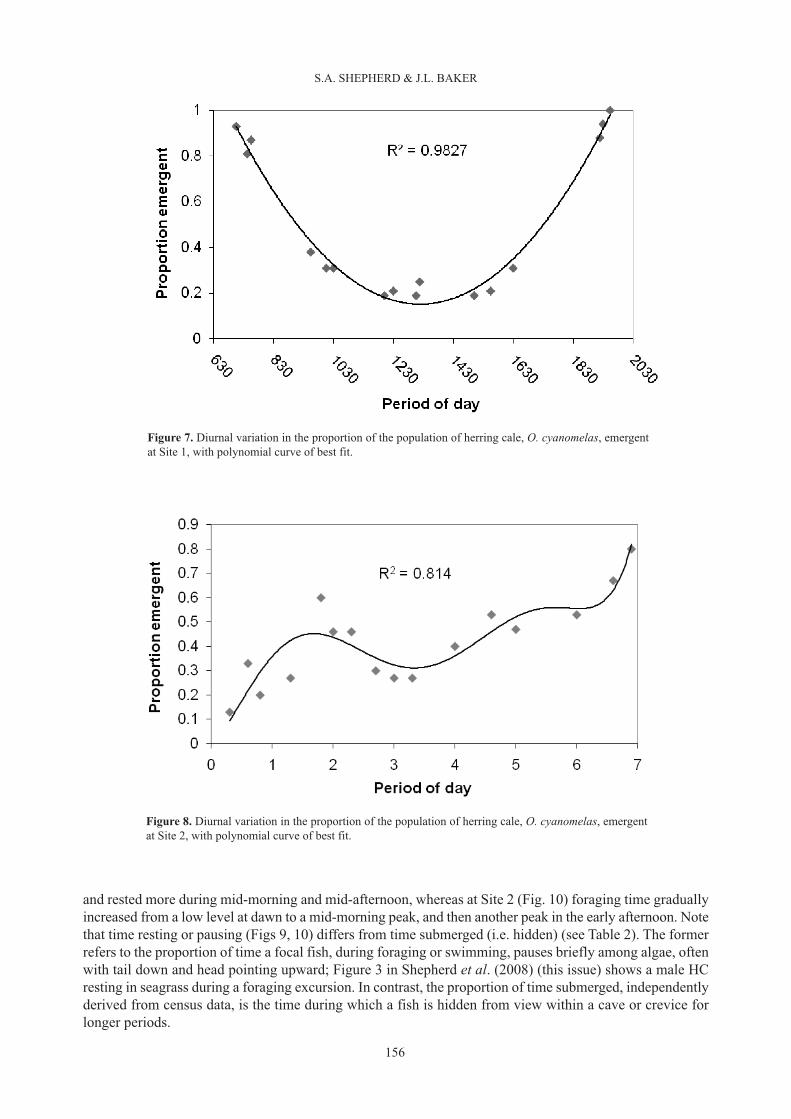
Activity-time budgetsThe proportion of the population emergent during daytime at Site 1 was highest at dawn and dusk, and lowestaround noon, and was fitted by an inverse parabolic curve (Fig. 7). In contrast, at Site 2 emergent behaviourwas markedly different. Few fish emerged at dawn, but numbers increased to a minor peak mid-morning,followed by a decline around mid-day, and a second peak toward dusk (Fig. 8). Despite these sharpdifferences, average time emergent was almost identical between sites (Table 2).
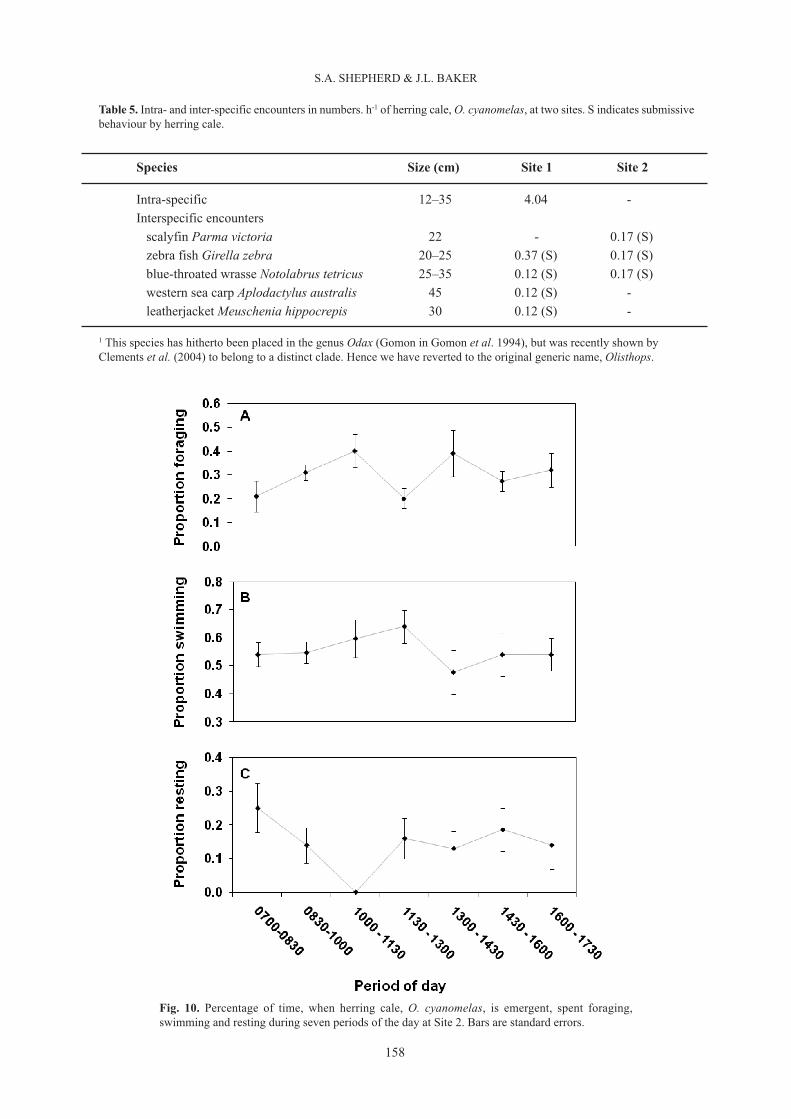
The diurnal distribution of time emergent spent foraging, swimming, and resting shows that at Site 1 (Fig.9) emergent HC spent more time foraging immediately after dawn, in the early afternoon, and toward dusk,
155
INVESTIGATOR GROUP EXPEDITION 2006: FORAGING ECOLOGY OF HERRING CALE,OLISTHOPS CYANOMELAS, IN SOUTH AUSTRALIA
Figure 6. Plot of percent of feeding bites on algal species or species’ groups vs percent cover of thosespecies at Site 2. Distance above and below the line indicates strength of preference or avoidance by herringcale, O. cyanomelas. Species are: 1, Sargassum spp.; 2, Cystophora monilifera; 3, Osmundaria prolifera;4, Cystophora moniliformis; 5, Cystophora subfarcinata; 6, Amphibolis antarctica; 7, Cystophora brownii;8, Dictyotales; 9, Ecklonia; 10, Cystophora siliquosa; 11, Caulerpa flexilis; 12, Asparagopsis armata.
and rested more during mid-morning and mid-afternoon, whereas at Site 2 (Fig. 10) foraging time graduallyincreased from a low level at dawn to a mid-morning peak, and then another peak in the early afternoon. Notethat time resting or pausing (Figs 9, 10) differs from time submerged (i.e. hidden) (see Table 2). The formerrefers to the proportion of time a focal fish, during foraging or swimming, pauses briefly among algae, oftenwith tail down and head pointing upward; Figure 3 in Shepherd et al. (2008) (this issue) shows a male HCresting in seagrass during a foraging excursion. In contrast, the proportion of time submerged, independentlyderived from census data, is the time during which a fish is hidden from view within a cave or crevice forlonger periods.
156
S.A. SHEPHERD & J.L. BAKER
Figure 7. Diurnal variation in the proportion of the population of herring cale, O. cyanomelas, emergentat Site 1, with polynomial curve of best fit.
Figure 8. Diurnal variation in the proportion of the population of herring cale, O. cyanomelas, emergentat Site 2, with polynomial curve of best fit.
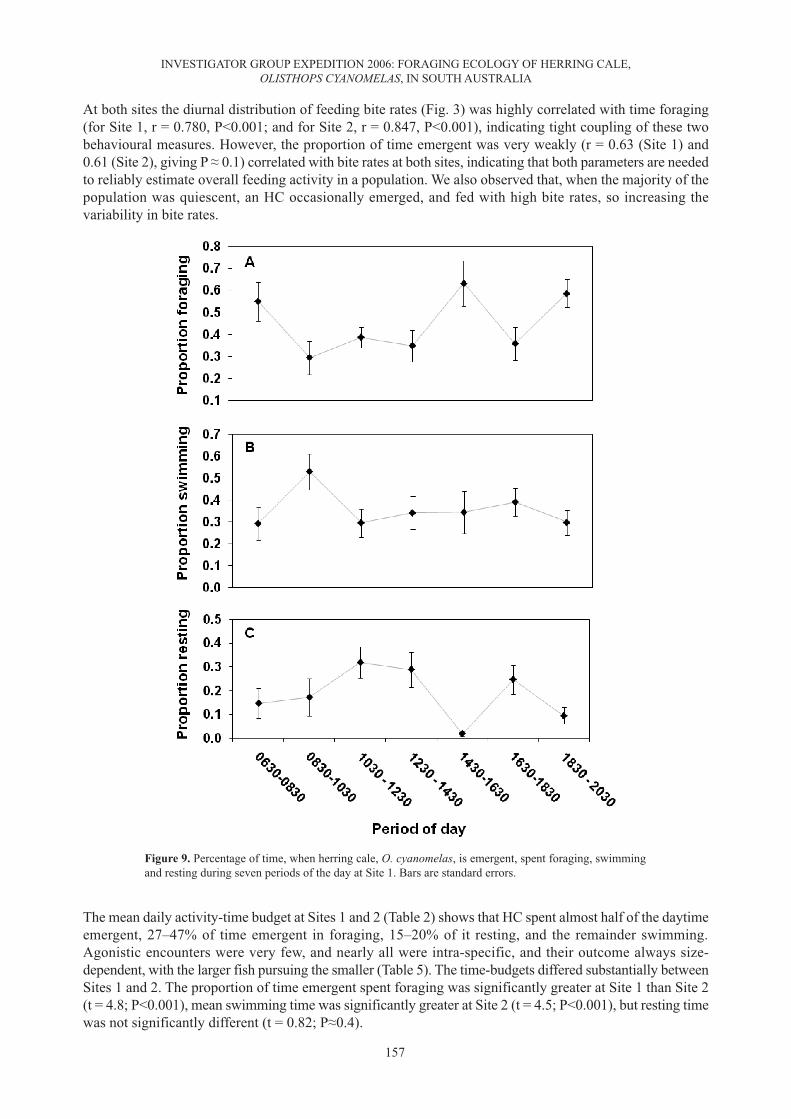
At both sites the diurnal distribution of feeding bite rates (Fig. 3) was highly correlated with time foraging(for Site 1, r = 0.780, P<0.001; and for Site 2, r = 0.847, P<0.001), indicating tight coupling of these twobehavioural measures. However, the proportion of time emergent was very weakly (r = 0.63 (Site 1) and0.61 (Site 2), giving P ≈ 0.1) correlated with bite rates at both sites, indicating that both parameters are neededto reliably estimate overall feeding activity in a population. We also observed that, when the majority of thepopulation was quiescent, an HC occasionally emerged, and fed with high bite rates, so increasing thevariability in bite rates.
The mean daily activity-time budget at Sites 1 and 2 (Table 2) shows that HC spent almost half of the daytimeemergent, 27–47% of time emergent in foraging, 15–20% of it resting, and the remainder swimming.Agonistic encounters were very few, and nearly all were intra-specific, and their outcome always size-dependent, with the larger fish pursuing the smaller (Table 5). The time-budgets differed substantially betweenSites 1 and 2. The proportion of time emergent spent foraging was significantly greater at Site 1 than Site 2(t = 4.8; P<0.001), mean swimming time was significantly greater at Site 2 (t = 4.5; P<0.001), but resting timewas not significantly different (t = 0.82; P≈0.4).
157
INVESTIGATOR GROUP EXPEDITION 2006: FORAGING ECOLOGY OF HERRING CALE,OLISTHOPS CYANOMELAS, IN SOUTH AUSTRALIA
Figure 9. Percentage of time, when herring cale, O. cyanomelas, is emergent, spent foraging, swimmingand resting during seven periods of the day at Site 1. Bars are standard errors.
158
S.A. SHEPHERD & J.L. BAKER
Table 5. Intra- and inter-specific encounters in numbers. h-1 of herring cale, O. cyanomelas, at two sites. S indicates submissivebehaviour by herring cale.
Species Size (cm) Site 1 Site 2
Intra-specific 12–35 4.04 -Interspecific encounters
scalyfin Parma victoria 22 - 0.17 (S)zebra fish Girella zebra 20–25 0.37 (S) 0.17 (S)blue-throated wrasse Notolabrus tetricus 25–35 0.12 (S) 0.17 (S)western sea carp Aplodactylus australis 45 0.12 (S) -leatherjacket Meuschenia hippocrepis 30 0.12 (S) -
1 This species has hitherto been placed in the genus Odax (Gomon in Gomon et al. 1994), but was recently shown byClements et al. (2004) to belong to a distinct clade. Hence we have reverted to the original generic name, Olisthops.
Fig. 10. Percentage of time, when herring cale, O. cyanomelas, is emergent, spent foraging,swimming and resting during seven periods of the day at Site 2. Bars are standard errors.

At Site 1 the foraging range sizes of large HC were small (Table 2), and foraging excursions rarely exceeded7–8 m, except toward dusk, when fish swam up to ~20 m into the deeper fucoid assemblage. At Site 2, theforaging ranges were about three times as large, with foraging excursions ranging up to 30 m.
Discussion
Algal dietHitherto HC has been considered a monophage, feeding almost exclusively on the kelp, Ecklonia (Andrew& Jones 1990; Jones 1999), whereas other odacines, notably Odax pullus and O. cyanoallix from NewZealand, are facultative herbivores, consuming a range of phaeophytes and rhodophytes according to theirabundance (Choat & Clements 1993; Clements & Choat 1993; Zemke-White & Clements 2004). This studyshows that, at a within-plant level, HC is a selective browser (sensu Horn 1989), with marked preferencesfor the margins of secondary laminae of Ecklonia, where sori are located, and for fucoid receptacles (cf.Clements & Choat 1993). At a plant assemblage level, HC is a facultative herbivore, but with well-definedalgal preferences. In the semi-exposed habitat with Ecklonia dominant (Site 3), that species was preferred,whereas, in the sheltered habitat without Ecklonia (Site 2) there was little selectivity among fucoid species.A common pattern of canopy algal species’ distributions on southern Australian coasts is the replacement ofEcklonia by various species of Cystophora and Sargassum sequentially on a decreasing wave exposuregradient (Shepherd & Womersley 1970, 1981; Collings & Cheshire 1998). This algal distribution patternexplains the shift in diet of this site-attached species from Ecklonia to fucoid algae with increasing shelteras reported here for three sites, and also the well attested correlation between Ecklonia biomass or cover andHC abundance on open coasts (e.g. Clements & Bellwood 1988; Andrew & Jones 1990; Jones 1999; Harmanet al. 2003; Shepherd & Brook 2003; Shepherd et al. 2005).
High levels of secondary metabolites, e.g. polyphenolics (Steinberg 1989), such as those occurring inCystophora spp., Sargassum spp. and Ecklonia (with highest within-plant levels in the secondary laminae ―see Andrew & Jones 1990) have previously been considered to be strong deterrents to herbivory. However,the digestive processes of odacines are now known to cope with them, and HC, in particular, has a highlyalkaline gut and a microbial gut flora that can ferment mannitol at rates comparable with herbivorous reptilesand mammals (Seeto et al. 1996; Choat & Clements 1998; Mountford et al. 2002).
Emergence and time budgetsEstimation of the true feeding activity of fish, which spend a portion of their time submerged (i.e. inactive),requires information on the proportion of total time submerged, as well as the feeding bite rate whenemergent. This study is the first to attempt to obtain both types of data for an odacine, although provided forother species e.g. Shepherd (2006). Overall, our study lends only limited support to the hypothesis of Choat& Clements (1993), who suggested, in the case of O. pullus, that foraging early in the day served to fill upthe gut, after which the fish merely “topped up” the anterior gut over the remainder of the day. The data forSite 1, with high early morning foraging, support the hypothesis, whereas those for Site 2, wherefeeding activity increased over several hours, does not. We speculate that the different behaviour atSite 2 may be due to a slower assimilation rate of fucoid algae by HC, as suggested also by themuch lower number of daily feeding bites at that site.
Other herbivorous fish show a gradual increase in feeding activity throughout the day (Polunin & Klumpp1989; Zemke-White et al. 2002), a pattern, which has given currency to the diel feeding hypothesis. Thisposits that under optimal foraging theory a feeding peak should occur late in the day as algaeaccumulate photosynthates. The diurnal variation in photosynthate production of the laminarian,Ecklonia, or of fucoids is not well known (Fairhead 2001), and is, in any event, not tightly coupledwith storage of photo-assimilates such as the polysaccharide, mannitol (Stewart et al. 1961), so thatsupport for the diel feeding hypothesis remains speculative. However, the bulk of receptacle tissuein Cystophora spp. comprises photosynthate (Nizamuddin 1962), and hence is highly nutritious.
159
INVESTIGATOR GROUP EXPEDITION 2006: FORAGING ECOLOGY OF HERRING CALE,OLISTHOPS CYANOMELAS, IN SOUTH AUSTRALIA

In this study HC differed substantially in foraging activity from O. pullus, which swam actively throughoutthe day (Choat & Clements 1993). At Sites 1 and 2, HC was submerged for 50–55% of the time, and evenwhen active frequently rested or paused for a further 10–20% of the time (Table 2), thus giving an overall75% level of inactivity during daylight hours. The occurrence of intra-specific interactions at Site 1, but notat Site 2, may simply reflect the fact that the density of HC at the former site was >5 times higher than at thelatter (Table 5).
While the different seasons of data collection might affect activity patterns, the very large differences betweensites in emergence patterns, activity and bite rates, and species consumed, pose complex questions beyondthe scope of this study. Their resolution must take into account assimilation rates, gut retention times, whichvary with diet and temperature, and physiological constraints of an ectothermic metabolism (Targett & Targett1990; Choat & Clements 1998). The tight relation between time foraging and bite rates suggests that foodavailability and quality are key determinants of foraging time and the size of the foraging range. In turn, thiswill influence the amount of time swimming, as shown by the greater amount of time spent swimming at Site2, with much larger home ranges, than at Site 1.
Lastly, the extensive clearing of kelp patches by HC in eastern Australia (Andrew & Jones 1990), comparedwith the absence of such behaviour at our study sites, raises intriguing questions about HC’s behaviouralflexibility. Jones (1999) noted that the impact of HC on Ecklonia is seasonal, and restricted to the spawningseason, when males defend territories and spawn with females within them. The HC preferentially feed onthe basal primary laminae of patches of the kelp, so destroying the plants and creating clearings in the forest,and making space for young plants to grow. Jones (1999) speculated that HC may prefer to eat one-year oldplants, but experiments suggested that the clearings more likely were focal points that facilitated theaggregative and mating behaviour of the species. Such clearing behaviour would be far more difficult if notimpossible on southern coasts, given the diversity and density of algal canopy species. The variable behaviourof this herbivore also indirectly illustrates the point made by students of food webs (McCann et al. 1998;Montoya et al. 2006) that trophic specialization (e.g. monophagy) can cause more variable web dynamics thanmore generalist consumption (polyphagy).
Acknowledgements
We thank Rob Lewis and Anthony Cheshire for sponsoring the expeditions to Althorpe I. and the InvestigatorGroup, the Friends of Althorpe I. for accommodation at Althorpe I., and James Teale for field assistance atAlthorpe I. Bryan Womersley, and Bob Baldock kindly assisted with algal determinations, and Ib Svane,Greg Collings and anonymous reviewers helpfully criticised the manuscript.
References
Andrew, N.L. & Jones, G.P. (1990). Patch formation by herbivorous fish in a temperate Australian kelp forest. Oecologia, 85: 57–86.
Choat, J.H. & Clements, K.D. (1993). Daily feeding rates in herbivorous labroid fishes. Marine Biology, 117: 205–211.
Choat, J.H. & Clements, K.D. (1998). Vertebrate herbivores in marine and terrestrial environments: A nutritional perspective. AnnualReview in Ecology and Systematics, 29: 375–403.
Clements, K.D. & Bellwood, D.R. (1988). A comparison of the feeding mechanisms of two herbivorous labroid fishes, the temperateOdax pullus and the tropical Scarus rubroviolaceus. Australian Journal of Marine and Freshwater Research, 39: 87–107.
Clements, K.D. & Choat, J.H. (1993). Influence of season, ontogeny and tide on the diet of the temperate marine herbivorous fishOdax pullus (Odacidae). Marine Biology, 117: 213–220.
Clements, K.D., Alfaro, M.E., Fessler, J.L. & Westneat, M.W. (2004). Relationships of the temperate Australasian labrid fish TribeOdacini (Perciformes; Teleostei). Molecular Phylogenetics and Evolution, 32: 575–587.
Collings, G.J. & Cheshire, A.C. (1998). Composition of the subtidal macroalgal communities of the lower gulf waters of SouthAustralia, with reference to water movement and geographical separation. Australian Journal of Botany, 46: 657–669.
160
S.A. SHEPHERD & J.L. BAKER

Edgar, G.J. (1999). Tasmania. In ‘Under Southern Seas: The Ecology of Australia’s Rocky Reefs’ (Ed. N. Andrew) pp. 30–39.(University of New South Wales Press, Sydney).
Edyvane, K. & Shepherd, S.A. (1999). South Australia. In ‘Under Southern Seas: The Ecology of Australia’s Rocky Reefs’. (Ed. N.Andrew) pp. 40–49. (University of New South Wales Press, Sydney).
Fairhead, V.A. (2001). ‘The ecophysiology and production ecology of the kelp Ecklonia radiata (C. Agardh) J.Agardh, at WestIsland, South Australia’. Ph.D. thesis (University of Adelaide, Adelaide).
Gomon, M.F., Glover, J.C.M. & Kuiter R.H. (1994). ‘Fishes of Australia’s South Coast’ (South Australian Government Printer,Adelaide).
Harman, N., Harvey, E.S. & Kendrick, G.A. (2003). Differences in fish assemblages from different reef habitats at Hamelin Bay,South-western Australia. Marine Freshwater Research, 54: 177–184.
Horn, M.H. (1989). Biology of marine herbivorous fishes. Oceanography and Marine Biology Annual Review, 27: 167–272.
Jones, G.P. (1999). Herbivorous fishes. In ‘Under Southern Seas: The Ecology of Australia’s Rocky Reefs’ (Ed. N. Andrew) pp.202–209. (University of New South Wales Press, Sydney).
Kendrick, G. (1999). Western Australia. In ‘Under Southern Seas: The Ecology of Australia’s Rocky Reefs’ (Ed. N. Andrew) pp.50–59 (University of South Wales Press, Sydney).
McCann, Hastings, K. A. & Huxel, G.R. (1998). Weak trophic interactions and the balance of nature. Nature, 393: 794–798.
Montoya, J.M., Pimm, S.L. & Solé, R.V. (2006). Ecological networks and their fragility. Nature, 442: 259–264.
Mountford, D.O., Campbell, J. & Clements, K.L. (2002). Hindgut fermentation in three species of marine herbivorous Fish. AppliedEnvironmental Microbiology, 68: 1374–1380.
Nizamuddin, M. (1962). The anatomy and life history of Cystophora, Acrocarpia, and Caulocystis. Botanica Marina, 7: 42–63.
Pavlov, D.S. & Kasumyan, A.O. (1998). The structure of the feeding behaviour of fishes. Journal of Ichthyology, 38: 116–128.
Polunin, N.V.C. & Klumpp, D.W. (1989). Ecological correlates of foraging periodicity in herbivorous reef fishes of the Coral Sea.Journal of Experimental Marine Biology and Ecology, 126: 1–20.
Seeto, G.S., Veivers P.C., Clements, K.D. & Slaytor, M. (1996). Carbohydrate utilization by microbial symbionts in the marineherbivorous fishes Odax cyanomelas and Crinodon lophodon. Journal of Comparative Physiology B, 165: 571–579.
Shepherd, S.A. (2006). Ontogenetic changes in diet, feeding behaviour and activity of the western blue groper, Achoerodus gouldii.In ‘The Marine Flora and Fauna of Esperance, Western Australia’ (Eds F.E. Wells, D.I. Walker, & G.A. Kendrick). pp 477–494(Western Australian Museum, Perth).
Shepherd, S.A. & Brook J.B. (2003). Encounter 2002 expedition to the Isles of St Francis, South Australia: Reef fishes. Transactionsof the Royal Society of South Australia, 127: 269–279.
Shepherd, S.A. & Brook, J.B. (2005). Foraging ecology of the western blue groper, Achoerodus gouldii, at Althorpe Islands, SouthAustralia. Transactions of the Royal Society of South Australia, 129: 202–208.
Shepherd, S.A. & Womersley, H.B.S. (1970). The sublittoral ecology of West Island, South Australia.1. Environmental factors andalgal ecology. Transactions of the Royal Society of South Australia, 94: 105–138.
Shepherd, S.A. & Womersley, H.B.S. (1981). The algal and seagrass ecology of Waterloo Bay, South Australia. Aquatic Botany, 11:305–371.
Shepherd, S.A., Edgar, G.J. & Barrett, N.S. (2005). Reef fishes of the Althorpe Islands and Adjacent Coasts of South Australia.Transactions of the Royal Society of South Australia, 129: 183–192.
Shepherd, S.A., Clark, M. & Ferguson, G. (2008). Investigator Group expedition 2006: Habitat-dependent-foraging behaviour anddiet of the scalyfin, Parma victoriae, in South Australia. Transactions of the Royal Society of South Australia, 132(2): 134–146.
Steinberg, P.D. (1989). Biogeographic variation in brown algal polyphenolics and other secondary metabolites: Comparison betweentemperate Australasia and North America. Oecologia, 78: 373–382.
161
INVESTIGATOR GROUP EXPEDITION 2006: FORAGING ECOLOGY OF HERRING CALE,OLISTHOPS CYANOMELAS, IN SOUTH AUSTRALIA

Steinberg, P. & Kendrick, G.A. (1999). Kelp forests. In ‘Under Southern Seas: The Ecology of Australia’s Rocky Reefs’ (Ed. N.Andrew) pp 60–71. (University of New South Wales Press, Sydney).
Stewart, C.M., Higgins, H.G. & Austin, S. (1961). Seasonal variation in Alginic Acid, Mannitol, Laminarin and Fucoidin in thebrown alga, Ecklonia radiata. Nature, 192: 1208.
Targett, T.E. & Targett, N.M. (1990). Energetics of food selection by the herbivorous parrotfish Sparisoma radians: Roles ofassimilation efficiency, gut evacuation rate, and algal secondary metabolites. Marine Ecology Progress Series, 66: 13–21.
Windell, J.T. (1968). Food analysis and rate of digestion. In ‘Methods for Assessment of Fish Production in Fresh Waters’ (Ed. W.E.Ricker) pp 197–203. (Blackwell, Oxford).
Womersley, H.B.S. (1987). ‘The Marine Benthic Flora of Southern Australia’ Part II (Fauna and Flora Handbooks Committee,Adelaide).
Zemke-White, W.L., Choat, J.H. & Clements, K.D. (2002). A re-evaluation of the diel feeding hypothesis for marine herbivorousfishes. Marine Biology, 141: 571–579.
Zemke-White, W.L. & Clements K.D. (2004). Relationship between long-term changes in algal community structure and herbivorediet at the Three Kings Islands, New Zealand. New Zealand Journal of Marine and Freshwater Research, 38: 837–844.
162
S.A. SHEPHERD & J.L. BAKER
Copyright © 2022 FDOKUMEN




















