
Invalidación de autapomorfias putativas mediante análisis de filogenia molecular en Anopheles...
18
37 BOLETÍN DE MALARIOLOGÍA Y SALUD AMBIENTAL Vol. XLIX, Nº 2, Agosto-Diciembre, 2009 Invalidación de autapomorfias putativas mediante análisis de filogenia molecular en Anopheles subgenero Nyssorhynchus Juan-Carlos Navarro 1 *, Camila Hernández 1 , Gabriela Rangel-Díaz 1 , Edmundo Guerrero 1 , Yadira Rangel 1 & Jazzmin Arrivillaga 2 1 Instituto de Zoología Tropical, Laboratorio Biología de Vectores, Universidad Central de Venezuela, Apdo. 47058, Caracas 1041-A, Venezuela. 2 Departamento de Estudios Ambientales, Laboratorio de Genética de Poblaciones, Sección Invertebrados, Sartenejas, Baruta, Apdo. 89000. Universidad Simón Bolívar, Caracas - Venezuela. *Autor de correspondencia: [email protected] Colectas e identificaciones realizadas en un área de transmisión malárica en el estado Sucre, Venezuela, mostraron mediante el uso de las claves ad hoc, la posible presencia en simpatría de poblaciones de Anopheles aquasalis Curry y An. benarrochi Gabaldón, Cova-García & López. Adicionalmente, se detectaron poblaciones de individuos que mostraron mezcla de los caracteres diagnósticos señalados en las claves. Debido a la importancia epidemiológica que representan ambas especies en dicha zona malárica, este estudio se orientó hacia la determinación de las tres entidades señaladas, mediante filogenia molecular con las secuencias de ADN mitocondrial (region Citocromo Oxidasa 1). En este sentido, el concepto de especie filogenética fue evaluada utilizando análisis de parsimonia máxima que mostraron politomías (nodos no resueltos) fuertemente apoyadas entre las secuencias inclusive la del haplotipo proveniente de un macho identificado como aquasalis, mostrando hipótesis (árboles) totales sin resolución ni apoyo de grupos internos con los tres morfotipos como un grupo natural único. Las distancias genéticas (Kimura-2P) mostraron que la variablidad inter-morfotipos están dentro de la varianza intra sugiriendo que los haplotipos problema son una única entidad polimórfica y con plasticidad fenotípica. Se concluye que An. aquasalis es la entidad única de las poblaciones estudiadas y que los caracteres morfológicos como: 1) vena Medial con escamas negras y 2) palpos maxilares con superficie ventral cubierta de escamas blancas, que son propuestas en la mayoría de las claves no son caracteres válidos para la identificación de hembras de An. benarrochi y que tales caracteres forman parte de la variación intraespecífica en An. aquasalis. Esta conclusión apoya la revisión de Faran (1980), quién no propone caracteres diferenciales para hembras de ambas especies, siendo indistinguibles por medio de los caracteres morfológicos tradicionalmente utilizados. Palabras clave: aquasalis, ADN mitocondrial, benarrochi, cladística, claves morfológicas, especie filogenética, malaria, Venezuela. INTRODUCCIÓN Anopheles aquasalis Curry, es un vector de orígen Neotropical, importante en la transmisión de malaria por Plasmodium vivax en América del sur. Es una de las 33 especies pertenecientes al subgénero Nyssorhynchus (Harbach, 2004). La distribución de An. aquasalis en el continente es básicamente costera, y se extiende en la costa Atlántica desde Nicaragua hasta Brasil, y en la costa Pacífica hasta Ecuador, incluyendo también su presencia en varias islas del Caribe (Faran, 1980; Guimaraes, 1997). En Venezuela, se encuentra en toda la costa del país desde el Delta Amacuro hasta el estado Zulia (Sutil, 1980; Rubio-Palis, 2000). Los inmaduros se desarrollan en una amplia variedad de ambientes desde cuerpos de agua dulce hasta manglares en aguas salobres, incluyendo canales de irrigación y bosques inundados (Berti et al., 1993; Grillet, 2000). Epidemiologicamente, An. aquasalis ha sido incriminado como vector de P. vivax en Venezuela, Trinidad, Guyana y Brasil (Berti et al., 1993, Chadee & Kitron, 1999; Laubach et al., 2001; Deane et al., 1948; Povoa et al., 2003 y Da Silva et al., 2006).
Transcript of Invalidación de autapomorfias putativas mediante análisis de filogenia molecular en Anopheles...

37
BOLETÍNDE MALARIOLOGÍAY SALUD AMBIENTALVol. XLIX, Nº 2, Agosto-Diciembre, 2009
Invalidación de autapomorfias putativas mediante análisis de filogenia molecular en Anopheles subgenero NyssorhynchusJuan-Carlos Navarro1*, Camila Hernández1, Gabriela Rangel-Díaz1, Edmundo Guerrero1, Yadira Rangel1 & Jazzmin Arrivillaga2
1 Instituto de Zoología Tropical, Laboratorio Biología de Vectores, Universidad Central de Venezuela, Apdo. 47058, Caracas 1041-A, Venezuela.2 Departamento de Estudios Ambientales, Laboratorio de Genética de Poblaciones, Sección Invertebrados, Sartenejas, Baruta, Apdo. 89000. Universidad Simón Bolívar, Caracas - Venezuela.
*Autor de correspondencia: [email protected]
Colectase identificaciones realizadasenunáreade transmisiónmaláricaenelestadoSucre,Venezuela,mostraronmedianteelusodelasclavesadhoc,laposiblepresenciaensimpatríadepoblacionesdeAnopheles aquasalisCurryyAn. benarrochiGabaldón,Cova-García&López.Adicionalmente,sedetectaronpoblacionesde individuosquemostraronmezclade loscaracteresdiagnósticos señalados en las claves. Debido a la importancia epidemiológica que representanambas especies en dicha zona malárica, este estudio se orientó hacia la determinación de lastres entidades señaladas,mediante filogeniamolecular con las secuencias deADNmitocondrial(regionCitocromoOxidasa1).Enestesentido,el conceptodeespeciefilogenética fueevaluadautilizandoanálisisdeparsimoniamáximaquemostraronpolitomías(nodosnoresueltos)fuertementeapoyadasentrelassecuenciasinclusiveladelhaplotipoprovenientedeunmachoidentificadocomoaquasalis,mostrandohipótesis(árboles)totalessinresoluciónniapoyodegruposinternosconlostresmorfotiposcomoungruponaturalúnico.Lasdistanciasgenéticas(Kimura-2P)mostraronquelavariablidadinter-morfotiposestándentrodelavarianzaintrasugiriendoqueloshaplotiposproblemasonunaúnicaentidadpolimórficayconplasticidadfenotípica.SeconcluyequeAn. aquasalis es la entidadúnicadelaspoblacionesestudiadasyqueloscaracteresmorfológicoscomo:1)venaMedialcon escamas negras y 2) palposmaxilares con superficie ventral cubierta de escamas blancas,que sonpropuestas en lamayoría de las clavesno son caracteres válidospara la identificacióndehembrasdeAn. benarrochi yquetalescaracteresformanpartede lavariación intraespecíficaen An. aquasalis.EstaconclusiónapoyalarevisióndeFaran(1980),quiénnoproponecaracteresdiferencialesparahembrasdeambasespecies,siendoindistinguiblespormediodeloscaracteresmorfológicostradicionalmenteutilizados.
Palabras clave: aquasalis, ADN mitocondrial, benarrochi, cladística, claves morfológicas, especie filogenética, malaria, Venezuela.
INTRODUCCIÓN
Anopheles aquasalis Curry, es un vector de orígen Neotropical, importante en la transmisión de malaria por Plasmodium vivax en América del sur. Es una de las 33 especies pertenecientes al subgénero Nyssorhynchus (Harbach, 2004). La distribución de An. aquasalis en el continente es básicamente costera, y
se extiende en la costa Atlántica desde Nicaragua hasta Brasil, y en la costa Pacífica hasta Ecuador, incluyendo también su presencia en varias islas del Caribe (Faran, 1980; Guimaraes, 1997). En Venezuela, se encuentra en toda la costa del país desde el Delta Amacuro hasta el estado Zulia (Sutil, 1980; Rubio-Palis, 2000). Los inmaduros se desarrollan en una amplia variedad de ambientes desde cuerpos de agua dulce hasta manglares en aguas salobres, incluyendo canales de irrigación y bosques inundados (Berti et al., 1993; Grillet, 2000).Epidemiologicamente, An. aquasalis ha sido incriminado como vector de P. vivax en Venezuela, Trinidad, Guyana y Brasil (Berti et al., 1993, Chadee & Kitron, 1999; Laubach et al., 2001; Deane et al., 1948; Povoa et al., 2003 y Da Silva et al., 2006).

Bol. Mal. Salud Amb.38
Invalidación de autapomorfias putativas en Anopheles
Desde su descripción por Curry en 1932, la morfología tanto en adultos como en sus fases inmaduras ha dejado un resultado de cinco sinonimias: emilianus, guarujaensis, guarauno, deltaorinoquensis y delta (Faran, 1980; Portal del Walter Reed Biosystematics Unit, Smithsonian Institution: http://www.mosquitocatalog.org/species/species_profile.asp?ID=293). Estas sinonimias sugieren la dificultad de encontrar caracteres diagnósticos en una especie que parece mostrar una variabilidad morfológica mayor al promedio. Este hecho, promovió la búsqueda de caracteres no tradicionales como la taxonomía de huevos mediante microscopía electrónica de barrido y diferentes abordajes bajo técnicas genéticas desde las isoenzimas, cromosomas hasta el uso de secuencias ribosomales y mitocondriales del ADN, proponiéndose hipótesis de la presencia de un complejo de especies, hasta una especie polimorfica con alta variabilidad intraespecífica (Lounibos & Conn, 2000). Autores como Linley et al. (1993) y Maldonado et al. (1997), demuestran bajo microscopía electrónica de barrido que los huevos de An. aquasalis entre Brasil y Venezuela muestran una variación correspondiente a una especie simple. Las muestras analizadas de Venezuela que cubren la costa en sus extremos y muestran una variación clinal sin discontinuidades. Mas recientemente, Sallum et al. (2000) en un estudio filogenético con caracteres morfológicos de las diferentes fases de vida sugieren que An. aquasalis es una especie simple.
Los estudios genéticos han mostrado resultados conflictivos, así, mientras los estudios con cromosomas sugieren la presencia de una especie simple (Kitzmiller & Chow, 1971; Moncada & Conn, 1992) los estudios moleculares parecen apoyar la presencia de un complejo de especies (Steiner et al., 1982; Conn et al., 1993 con Isoenzimas y RFLP-ADN mitocondrial respectivamente). Recientemente, Fairley et al. (2002), demuestra mediante análisis con ADN mitochondrial (Citocromo Oxidasa 1) que el río Amazonas no es una barrera definitiva para diferenciar las poblaciones de Brasil, y Farley et al. (2005) con ADN ribosomal (ITS1 e ITS2) muestran heterogeneidad intragenómica para An. aquasalis, pero ambos estudios concluyen en definirla como una especie simple.
Bajo la hipótesis que, las poblaciones de Venezuela representan una especie simple, de acuerdo a los resultados de Linley et al., 1993; Maldonado et al.,
1997 y Fairley et al., 2005, mas el intervalo ecólogico que sus inmaduros pueden ocupar (Berti et al., 1993; Grillet, 2000), en un estudio eco-epidemiológico que se desarrolla en el estado Sucre, al oriente de Venezuela (Fig. 1), en el Municipio Cajigal, se colectaron ejemplares anofelinos que fueron identificados con las claves mas utilizadas en Venezuela: Cova-Garcia et al. (1966, 1975) y Rubio-Palis (2000). Las primeras identificaciones llevaron a la conclusión preliminar que estabamos en presencia de al menos dos especies, cuyas hembras capturadas, eran identificadas como Anopheles aquasalis Curry y An. benarrochi Gabaldón, Cova-García & López, con base en los caracteres diagnósticos de las escamas en la vena medial y los palpos. Adicionalmente, otro morfotipo con una mezcla de caracteres diagnósticos fue identificado y separado como Anopheles “Mixto”. En vista de la importancia que representa en un area malárica, identificar correctamente las especies potencialmente involucradas en la transmisión, como primer paso para poder estructurar medidas de control y seguimiento adecuados, nuestro objetivo fue verificar y validar los caracteres morfológicos diagnósticos que muestran las claves referidas, así como también la clave de Consoli & de Oliverira (1994) y González & Carrejo (2007), las cuales contrastan con la clave propuesta por Faran (1980: 28) quién en la identificación de hembras no muestra ningún caracter diagnóstico, finalizando la identificación con ambas especies (aquasalis y benarrochi) sin diferenciación posible.
No obstante, la diferenciación en machos es inobjetable debido a que los genitales masculinos muestran diferencias notables, no es la fase adulta mas capturada en muestreos epidemiológicos, siendo la identificación de la fase adulta transmisora (hembra) la que en su mayoria es colectada por diferentes métodos (reposo, trampas de luz, cebo animal y alimentándose sobre humanos) siendo fundamental para una captura y definición rápida de las especies posiblemente involucradas.
De esta forma, el presente trabajo muestra la validación de los caracteres diagnósticos putativos en dichas claves mediante análisis de las secuencias del gen citocromo oxidasa 1 (CO1), del ADN mitocondrial (ADNmt) extraido y amplificado por PCR, de los ejemplares (hembras y machos) colectados e identificados, y análizados explícitamente mediante métodos cladísticos.

Vol. XLIX, N° 2, Agosto-Diciembre, 2009 39
Navarro J. C. et al.
El ADN mitocondrial muestra un patrón de evolución rápida con genes básicos como el CO1 o CO2 que participan en funciones muy conservadas como la cadena respiratoria, ha sido usado como indicador de estructura poblacional, determinación de flujo génico, para definir patrones históricos poblacionales, relaciones filoegenéticas, resolver problemas taxonómicos y análisis filogeografícos (Moritz et al., 1987; Simon et al., 1994, 2006).
En nuestro caso, los análisis de filogenéticos de las secuencias tanto de hembras y machos de los diferentes ejemplares-especies-tipos identificados morfológicamente y que representarán nuestro grupo problema, nos mostrarán mediante un análisis cladístico si pertenecen o no a un mismo linaje evolutivo, si pueden o no estos linajes ser considerados una misma o diferentes unidades taxonómicas (concepto de especie filogenética) y, si a su vez, son concordantes con los caracteres morfologícos bajo las cuales fueron identificadas buscando una correlación genotipo-fenotipo.
Para correlacionar los caracteres moleculares con los morfológicos que diferencian -según las claves referidas- a An. benarrochi y An. aquasalis, se realizaron análisis moleculares utilizando la región de ADN mitocondrial Citocromo Oxidasa C subunidad I (CO1), tasa mutacional rápida, por lo que está región mitocondrial se ha empleado en el estudio de relaciones filogenéticas de especies muy relacionadas, crípticas, complejo de especies, y a nivel infra-específico, y en donde las diferencias y divergencias genéticas detectadas representan un buen carácter a nivel taxonómico (Simon et al., 1994; 2006).
MATERIALES Y MÉTODOS
Mosquitos, colecta e identificación
El material de trabajo consistió en muestras de adultos del género Anopheles (Anopheles aquasalis y Anopheles benarrochi) colectadas mediante trampas de luz tipo CDC, y en reposo en diferentes
Fig 1. Ubicación geográfica en Venezuela de las localidades de colecta.
Bol. Mal. Salud Amb.40
Invalidación de autapomorfias putativas en Anopheles
Genitales de machos adultos
Los individuos machos provenientes de crias asociadas fueron evaluados e identificados morfológicamente por medio del montaje y observación de los genitales utilizándo el protocolo original de Cova-García (1961). La observación de estructuras se realizó por medio de un microscopio óptico (Leitz Wetzlar LABORLUX S, modelo 513591) y se tomaron fotografías de las mismas con una cámara SONY F828 de 8 megapixel.
Extracción de ADN
Se realizaron extracciones del ADN genómico de las dos patas para cada individuo, siguiendo el protocolo de Bender et al. (1983) modificado por Arrivillaga et al. (2002). Las extracciones se realizaron partiendo de un total de 62 individuos, clasificados en tres grupos taxonómicos-tipológicos bajo nuestro criterio: aquellos que según ambas claves taxonómicas utilizadas poseían los caracteres de An. aquasalis, de An. benarrochi y una combinación de ambos tipos morfológicos a los cuales denominamos Anopheles “mixto”. Adicionalmente, se tomaron 10 individuos de Anopheles albimanus, el cual fue considerado como grupo externo en los análisis filogenéticos.
localidades dentro del Municipio Cajigal del Estado Sucre, Venezuela (Fig. 1), utilizando en el último caso aspiradores manuales a bateria. Adicionalmente, se obtuvieron crías asociadas desde larvas y pupas colectadas directamente de los criaderos (manglares, prados inundados, pantano herbáceo dulce y salobre). Adicionalmente, se emplearon muestras de Anopheles albimanus, las cuales fueron colectadas en el lago de Valencia, estado Carabobo como taxón de referencia para los análisis filogenéticos de acuerdo con Sallum et al. (2000) como especie hermana. Los ejemplares (adultos) colectados fueron montados en forma convencional (Belkin et al., 1965) y utilizados como “voucher taxonómico” para comprobaciones morfológicas posteriores. Los “vouchers” fueron depositados en la Colección del Laboratorio de Biología de Vectores, Instituto de Zoología Tropical, Universidad Central de Venezuela (LBV sensu Guimaraes, 1997). Las identificaciones se realizaron mediante el uso de claves morfológicas tradicionales y descripciones, en particular: Cova-Garcia et al. (1961), Cova-Garcia & Sutil (1975) y Rubio-Palis (2000), sin embargo fueron consultados Curry (1932), Gabaldón et al. (1941); Faran (1980); Faran & Linthicum (1981), de Consoli & de Oliveira (1994). Cada ejemplar montado se le disecaron dos patas (media y anterior derecha) que fueron preservadas en etanol al 95% a -80°C para su posterior uso en análisis de ADN (Navarro & Weaver, 2004).
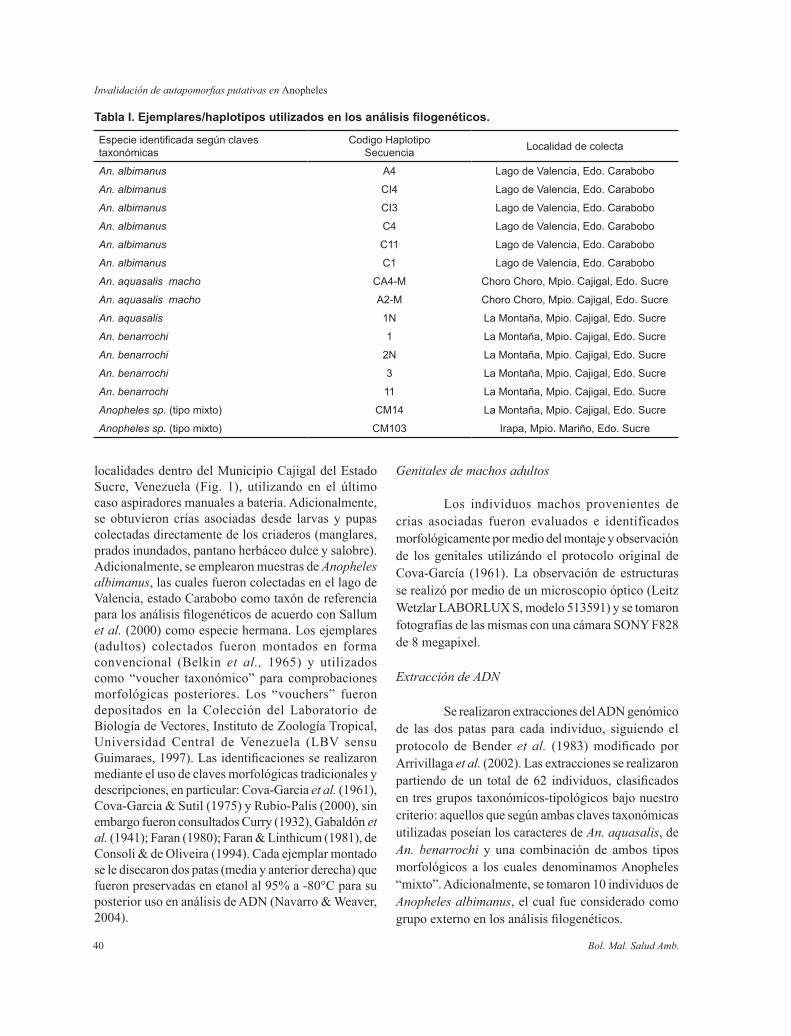
Especieidentificadasegúnclavestaxonómicas
CodigoHaplotipoSecuencia Localidaddecolecta
An. albimanus A4 LagodeValencia,Edo.Carabobo
An. albimanus CI4 LagodeValencia,Edo.Carabobo
An. albimanus CI3 LagodeValencia,Edo.Carabobo
An. albimanus C4 LagodeValencia,Edo.Carabobo
An. albimanus C11 LagodeValencia,Edo.Carabobo
An. albimanus C1 LagodeValencia,Edo.Carabobo
An. aquasalis macho CA4-M ChoroChoro,Mpio.Cajigal,Edo.Sucre
An. aquasalis macho A2-M ChoroChoro,Mpio.Cajigal,Edo.Sucre
An. aquasalis 1N LaMontaña,Mpio.Cajigal,Edo.Sucre
An. benarrochi 1 LaMontaña,Mpio.Cajigal,Edo.Sucre
An. benarrochi 2N LaMontaña,Mpio.Cajigal,Edo.Sucre
An. benarrochi 3 LaMontaña,Mpio.Cajigal,Edo.Sucre
An. benarrochi 11 LaMontaña,Mpio.Cajigal,Edo.Sucre
Anopheles sp.(tipomixto) CM14 LaMontaña,Mpio.Cajigal,Edo.Sucre
Anopheles sp.(tipomixto) CM103 Irapa,Mpio.Mariño,Edo.Sucre
Tabla I. Ejemplares/haplotipos utilizados en los análisis filogenéticos.

Vol. XLIX, N° 2, Agosto-Diciembre, 2009 41
Navarro J. C. et al.
Amplificación de COI via PCR, y secuenciación
Se amplificó una región del COI de 537pb entre las posiciones 1632 y 2188, utilizando los cebadores: C1J1632 (+): 5’ – TGATCAAATTTATAAT - 3’ de Kambhampati & Smith (1995) y C1N2191 (-): 5’ – GGTAAAATTAAAATATAAACTTC- 3’ de Simon et al. (1994). La amplificación de COI se realizó de forma automatizada en termociclador PT-200 MJ Research, siguiendo el programa de Kambhampati & Smith (1995).
Finalmente los productos amplificados fueron visualizados con UV en geles de agarosa al 1%, 0.5X TBE, teñidos con Bromuro de Etidio (Sigma, 10 mg/ml). Las amplificaciones positivas, fueron secuenciadas directamente en ambos sentidos empleando los mismos cebadores de PCR, utilizando un secuenciador automatico Applied Biosystems (Foster City, CA) ABI 377 bajo las especificaciones del fabricante.
Distancia genética
Las distancias genéticas (Kimura- 2 parametros) intra e inter morfotipos problema fueron calculadas con el programa Mega 4 (Tamura et al., 2007) a fin de evaluar y definir los límites de la varianza intra-morfotipos, intra-específica e inter-especifica.
Análisis filogenéticos
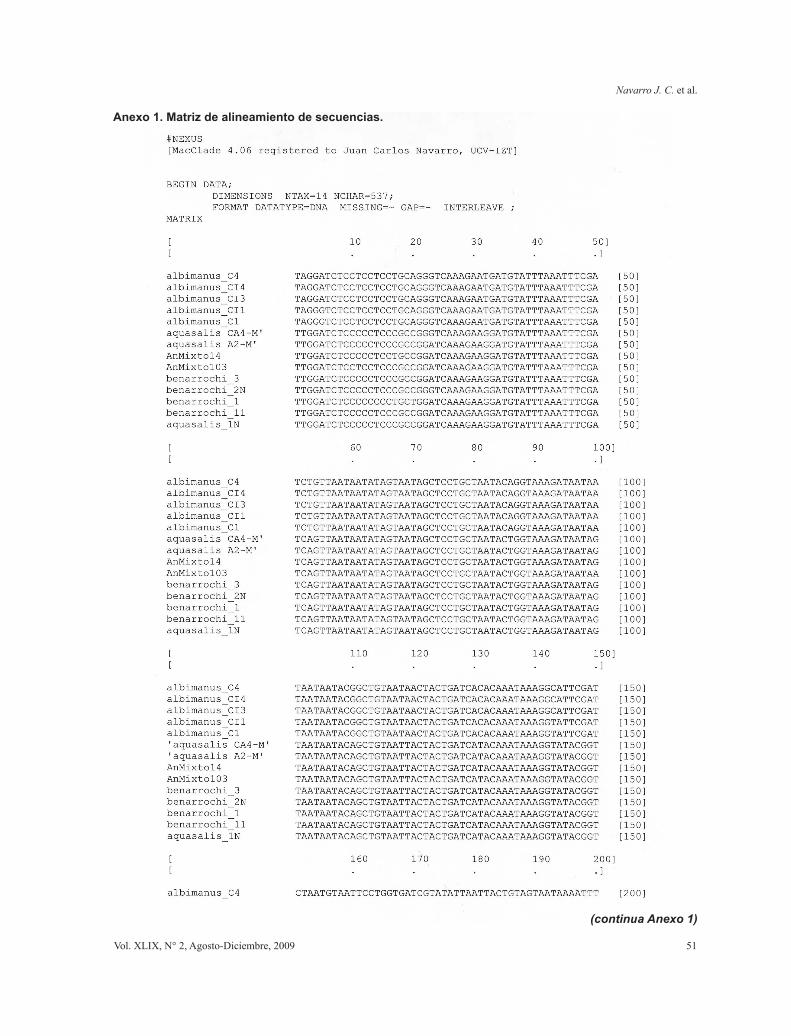
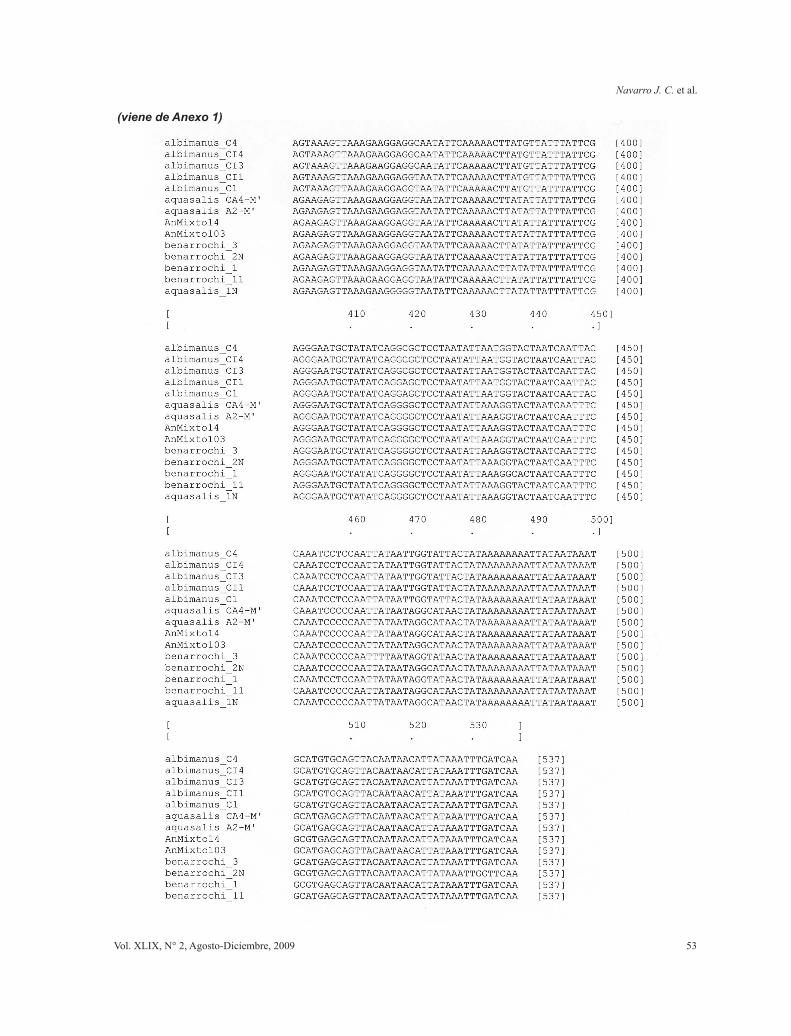
Las secuencias obtenidas fueron procesadas con Sequencer 4.2.2 (Gene Codes, Ann Arbor, MI) para la constrastación de secuencias directas y reversas y producción de un “contig” o secuencia única para cada haplotipo-ejemplar. Luego se alinearon en búsqueda de homologías posicionales mediante alineamiento múltiple local con ClustalW mediante MacVector 7.2 (Accelrys, Madison, WI). Las secuencias finales se contrastaron por búsquedas en Gen Bank NCBI (Blast) para el alineamiento global y la verificación de la válidez de las secuencias mediante su identidad con secuencias de otros anofelinos y/o especies cercanas.
Con las secuencias obtenidas (procesadas y validadas) (Tabla I) se realizaron alineamientos múltiples producto de secuenciales alineamientos pareados entre secuencias y clusters (agrupamientos) empleando en el programa Clustal W [en MacVector 7.2; Accelrys, Madison, WI].
Las secuencias alineadas y en formato Nexus fueron chequedas bajo MacClade 4.06 (Maddison & Maddison, 2003) para corregir errores de alineamiento y escoger aquellos haplotipos únicos entre todas las secuencias obtenidas de todos los ejemplares de cada morfotipo. La matriz final se introdujo en PAUP 4.010b (Swofford, 2002) para los análisis filogenéticos (La matriz de alineamiento de las secuencias se muestra en el Anexo 1). La búsqueda de hipótesis de relaciones evolutivas se realizó mediante análisis de parsimonia considerando los caracteres como desordenados, colapsando ramas cuando la máxima longitud es cero (amb-), con optimización de caracteres-estados mediante ACCTRAN (accelerated transformation) para buscar apoyo en las bases de las ramas y tratando los multiestados como inciertos. Los taxa fueron adicionados al azar con 1000 replicas por cada adición. Se utilizó el algoritmo de recorte y reconexión de árboles (TBR - Tree Bisection Reconnection) de barridos de ramas con adición de pasos al azar usando 1000 replicaciones, guardando sólo los mejores árboles (más cortos) obtenidos en cada réplica para la búsqueda de islas de arreglos. El repesado de caracteres se realizó por pesado sucesivo (Farris, 1969) mediante el índice de consistencia recalculado (RC). Se consideraron las secuencias de los ejemplares de Anopheles albimanus como grupo externo.
La búsqueda de “robustez” o apoyo de cada rama, clado o nodo fue estimada por remuestreo de la matriz por Bootstrapping (Felsenstein, 1985) con 1000 replicaciones por cada 10 replicaciones heurísticas después de eliminar aquellos caracteres no informativos (Carpenter, 1996). Los valores mostrados son aquellos clados o grupos con mas de un 50% de aparición en las búsquedas de árboles óptimos.
RESULTADOS
Identificación morfológica
De acuerdo con la identificación de las claves arriba señaladas, del total de individuos analizados, un 60% correspondió a ejemplares identificados como Anopheles aquasalis, mientras que un 30% a Anopheles benarrochi y un 10% al grupo denominado como Anopheles “Mixto”. La mayor dificultad se presentó al tratar de identificar las diferencias entre los individuos con los caracteres empleados en las claves como son las escamas blancas en la superficie inferior de los segmentos 4º y 5º de los palpos y la proporción de escamas claras y oscuras presentes en la porción no bifurcada de la vena media (M) alar.

Bol. Mal. Salud Amb.42
Invalidación de autapomorfias putativas en Anopheles
Un total de 46 amplificaciones fueron secuenciadas, pertenecientes a los tres morfotipos identificados: 25 de An. aquasalis (entre ellos 8 machos producto de crias asociadas), 10 de An. benarrochi y todos los 11 ejemplares identificados con caracteres diagnósticos mezclados de ambas especies, rotulados como Anopheles “Mixtos”. Un total de 9 haplotipos diferentes fueron identificados de los cuales tres pertenecen a An. aquasalis (CA4-M, A2-M, 1N), cuatro a An. benarrochi (1, 3, 2N,11) y dos a An “Mixto” (14, 103) (Tabla I). Las secuencias de los haplotipos se muestran alineadas en el Anexo 1.
Los genitales de machos fueron montados en láminas e identificadas en su totalidad (8 ejemplares) como An. aquasalis basándose en la descripción original de Curry (1932), y en las claves taxonómicas de Cova-García (1961), Cova –García & Sutil (1977), Faran (1980) y Faran & Linthicum (1981), los cuales produjeron uno de los dos haplotipos análizados filogenéticamente (aquasalis A2-M y CA4-M).
Características de las secuencias
Las secuencias presentaron una longitud de 537 pb. El contenido de Guanina+Citosina (G+C) en las secuencias obtenidas fue de 31,9% similar al porcentaje señalado para otras especies de dípteros (Anopheles 31,4%; Drosophila 30,2%; Herbeck & Novembre, 2003). El intervalo de variación estuvo entre 15% y 16%, sin una variación significativa entre los porcentajes de citosina y guanina entre morfotipos. (aquasalis, benarrochi y Mixto). Adicionalmente, no se observó ninguna relación directa entre la longitud de pares de base del fragmento de COI amplificado en las secuencias con la variación del %G+C.
No obstante, se observaron diferencias en los porcentajes de Adenina y Timina en las secuencias de las especies analizadas de Anopheles, siendo más altos los valores de % A (38,5-38,9 %) en relación con los % T (29,0-30,9 %). Por otro lado, el contenido de A+T fue de 67,9 %, indicando una preferencia en la terminación A o T en los tripletes (uso de codones); resultado que se asemeja con el obtenido por Morlais et al. (2002) para Aedes aegypti.
Distancia genética
Las distancias genéticas, indicaron valores de K2P intra-morfotipos entre 0,0037- 0,0168, siendo los
valores por morfotipo de 0,0037 (aquasalis), 0,0168 (benarrochi), 0,0151 (“Mixto”) y 0,0087 para el grupo referencia (albimanus). Las distancias inter-morfotipos arrojaron los siguientes valores: 0,0094 (aquasalis vs “Mixto”), 0,0103 (benarrochi Vs aquasalis) y 0,0142 (benarrochi Vs “Mixto”), mostrándo valores de distancia genética inter-morfotipos dentro del intervalo de valores K2P intra-morfotipos. Los valores entre albimanus y los morfotipos oscila entre 0,0756 a 0,0796.
Análisis filogenéticos
La matriz nexus para los análisis de parsimonia muestra un total de 14 taxa (14 haplotipos, 5 albimanus y 9 del grupo problema), con 537 caracteres (pares de base) de los cuales 482 son constantes, 9 no informativos y 46 son informativos para parsimonia.Los análisis con los caracteres de igual peso (Fig. 2) y con caracteres homólogos repesados (Fig. 3) arrojaron los siguientes resultados: 4 y 1 árboles o soluciones más parsimoniosas, longitud (L) de 66 y 57.4, Indice de Consistencia (IC)= 0,85 y 0,91, Indice de Retención (IR)= 0,86 y 0,93 respectivamente. El análisis con pesos iguales arrojó 2 topologias una de las cuales ubicó a “An Mixto” 103 externamente en algunas soluciones (Fig. 2).
El análisis con repesado de caracteres mostró una única solución (Fig. 4) similar a una de las topologías sin pesado. En todas las topologías y consensos las secuencias de albimanus se ubicaron externamente y separadas del grupo interno de haplotipos problema, quienes no mostraron una diferenciación marcada en clados diferentes. El análisis de re-muestreo o boostrapping realizado con 1000 replicas (Fig. 5) que busca apoyo estadístico de grupos, muestra las secuencias de albimanus como grupo basal, y diferenciadas de todas las secuencias/haplotipos problema derivadas internamente con un valor de apoyo de 100%. Internamente, las secuencias/haplotipos problema muestran una gran una politomía (“peine”) con todas las secuencias restantes a excepción de la secuencia de “Mixto 103” con muy bajo apoyo (63%), mostrando ninguna relación de ancestro-descendiente que pudiese indicar diferenciación entre ellas. Solo dos ramas internas con 89% y otra con 64% muestran secuencias del morfotipo benarrochi y “mixto” y la otra con secuencias de benarrochi y aquasalis. La topología del bootstrap con la politomia interna, muestra concordancia con las distancias geneticas por Kimura-2P entre el grupo problema.

Vol. XLIX, N° 2, Agosto-Diciembre, 2009 43
Navarro J. C. et al.
DISCUSIÓN
La identificación correcta de posibles vectores en un área determinada es de vital importancia para el diseño de estrategias y aplicación de control de vectores con el objetivo de reducir la tasa de transmisión y la incidencia malárica. Sin embargo, las dificultades para identificar hembras adultas de mosquitos del subgénero Nyssorhynchus han sido expuestas y discutidas en diversos trabajos (Conn & Lounibos, 2002) y reuniones de taxónomos de anofelinos. Problemas en la identificación en especies como nuneztovari (Conn et al., 1993b, 1998; Rangel et al., 1998; Sierra et al., 2004 y otros), oswaldoi (Marrelli et al., 1999; Scarpassa, 2005; Ruiz et al., 2005; Scarpassa & Conn, 2006 y otros) y aquasalis (Steiner et al., 1981, Moncada & Conn 1992, Conn et al.,1993a y otros) entre las mas estudiadas, han sido ampliamente referidos.
Variaciones en la morfología de cualquiera de las fases de desarrollo, en el comportamiento de picada o de reposo, constiuyen la base para el
Fig. 2. Las dos topologías resultantes (4 árboles) del Análisis de Parsimonia (PAUP 4.010b) sin pesado de caracteres y con 1000 replicaciones al azar y TBR. L=66; IC= 0,86; IR= 0,95.
Fig. 3. La única solución parsimoniosa con caracteres re-pesados obtenido por análisis de parsimonia (PAUP 4.010b) con 1000 replicaciones al azar y TBR. En gris se muestran las secuencias pertenecientes a los machos identificados por genitales. L= 54,7; IC= 0.93; IR= 0,97
Fig. 4. Arbol de consenso de mayoría producido por análisis de parsimonia y remuestreo por Boostrap (1000 replicas) por PAUP 4.010b. Lo valores arriba de las ramas representan el valor de apoyo entre 50 y 100% para cada clado. Aquellos no mostrados produjeron valores menores al 50% colapsando las ramas (politomias).

Bol. Mal. Salud Amb.44
Invalidación de autapomorfias putativas en Anopheles
estudio de posibles complejos de especies crípticas por medio de diferentes métodos, entre los cuales los métodos moleculares han sido los mas recientemente abordados. La detección de multiples sinonímias en la historia taxonómica de una especie, demuestra problemas intrínsecos en su identificación incluso por personal muy experimentado. Esto se traduce en la estructuración de claves morfológicas de identificación que no muestran las referidas variaciones e incluso se muestran tipológicamente caracteres diagnósticos (autapomorfias) putativos muy subjetivos, confusos o que se solapan entre especies muy similares.
Anopheles aquasalis Curry, un importante vector de malaria en América del Sur y el Caribe, es un caso en el cual las variaciones morfológicas han promovido el estudio de sus poblaciones por diferentes técnicas y abordajes, y en el cual no se ha llegado a una única conclusión entre las dos vertientes: una población que representa una única entidad taxonómica polimorfica o un complejo de especies crípticas (Conn & Lounibos, 2002; Fairley et al., 2005 entre otros ).
En nuestro caso, la identificación de ejemplares en Municipio Cajigal, estado Sucre, un área malárica al este de Venezuela, no representaba mayor problema de identificación hasta su análisis detallado por medio de las claves mas utilizadas local o regionalmente. Estas claves (Cova-Garcia & Sutil, 1977; Rubio-Palis, 2000) muestran caracteres diagnósticos que llevaron a la identificación y posible presencia de dos especies que podrían ser simpátridas en la región y en el área malárica (Sutil, 1980). Adicionalmente, ambas especies son referidas como vectores en Venezuela y Brasil (aquasalis, Berti et al., 1993; Povoa et al., 2003; Da Silva et al., 2006) y Perú (benarrochi, Flores-Mendoza et al., 2004) pudiendo ser benarrochi la especie mas abundante en zonas internas de agua dulce y aquasalis como la preponderante en la costa en aguas salobres, y complicando las posibles medidas de control.
Mientras las claves morfológicas para hembras arrojan la identificación de dos especies y un tercer morfotipo que muestra caracteres mezclados de ambas, la morfología de los genitales del los machos obtenidos por cría asociada señalan la presencia solo de An. aquasalis. Sin embargo, las formas de muestreo mas usuales resultan en la captura de una proporción de hembras cercana a un 100%, obteniendo solo machos por medio de crias asociadas y muy pocos en reposo,
es imperativo confirmar genética y filogenéticamente que los ejemplares hembra colectados corresponden a las especies identificadas por las claves (validación de los caracteres morfológicos diagnósticos) o representan una identidad única (invalidación de dichos caracteres).
Las distancias genéticas evidencian que la varianza inter se ubica dentro de la varianza intra-mortipos, sugiriendo que los morfotipos son parte de la varianza genética intraespecífica (0,0037-0,0168 Vs 0,0094-0,0142 respectivamente), lo que sugiere cierto grado de polimorfismo genético dentro de una entidad discreta definida como aquasalis + benarrochi + “Mixto” (0,0117) en contraste con An. albimanus (0,0087), lo que podría explicar la plasticidad fenotípica dentro de esta especie. Adicionalmente, las distancia genética entre los tres morfotipos de problema y albimanus, oscila entre K2P= 0,0756 a 0,0796. La varianza inter dentro de los valores intra -morfotipos sugiere la presencia de una población genética.
Los análisis cladísticos o filogenéticos mostraron tanto en la matriz con caracteres con pesos iguales (Fig. 2) y con caracteres repesados (Fig. 3), árboles igualmente parsimoniosos con topologías que muestran politomías y sin diferenciación de ramas. El análisis de apoyo de grupos (boostrap) y el consenso obtenido (Fig. 4) muestra que las secuencias pertenecientes a los tres morfotipos (aquasalis, benarrochi y “An Mixto”) están localizadas en una politomia fuertemente apoyada (100%), ubicándose en forma indistinta en las diferentes ramas internas sin una estructura filegenética definida y apoyada, lo cual indica que todas pertenecen a una misma entidad filogenética, contrario a lo esperado en caso de representar dos o tres entidades diferentes, en el cual se mostrarían dos o tres clados monofiléticos con un fuerte apoyo de grupo y de distancia genética en las topologias resultantes. En este caso la monofilia está representada y apoyada en un 100% por todas las secuencias problema sin diferenciación por clados o grupos.
Adicionalmente, las secuencia/haplotipo A2-M y CA4-M de machos identificados por genitales como An. aquasalis se ubicaron en el clado interno de la politomia, confirmando así, que el total de las secuencias/haplotipos en el ánalisis pertenecen a un mismo clado filogenético y monofilético (especie filogenética) concordante con An. aquasalis. Por tanto, los caracteres

Vol. XLIX, N° 2, Agosto-Diciembre, 2009 45
Navarro J. C. et al.
utilizados como autapomorfias (carácter diagnóstico) putativas en estas claves para la identificación de ambas especies (benarrochi y aquasalis) representan simple variabilidad intraespecífica de An. aquasalis Curry, no siendo caracteres válidos para la identificación y diferenciación (en hembras) de la especie An. benarrochi Gabaldón, Cova-García & López.
Esto demuestra que la correcta identificación de las variantes de An. aquasalis es problemática empleando las claves taxonómicas existentes, implicando que ambas especies (benarrochi y aquasalis) pueden ser comúnmente mal identificadas a lo largo de Latinoamérica como también lo señalan Ruiz et al. (2005).
Todas las claves morfológicas actualmente utilizadas (Faran, 1980; Faran & Linthicum, 1981; Consoli & de Oliveira, 1994; Rubio-Palis, 2000; González & Carrejo, 2007) para identificar hembras adultas anofelinas de las especies aquasalis y benarrochi concuerdan en que éstas no pueden ser fácilmente diferenciadas, sin embargo toman en cuenta los siguientes caracteres para separarlas (Fig. 2); en el caso de An. aquasalis presenta el segundo segmento de los tarsos posteriores con la mitad o más de la mitad negra, palpos con los segmentos 4º y 5º separados por un anillo negro y tallo de la vena media con escamas claras y oscuras, por su parte An. benarrochi presenta el segundo segmento de los tarsos con menos de la mitad negra, palpos con los segmento 4º y 5º separados por un anillo negro y con escamas oscuras en la parte ventral o lateral entre las escamas claras y tallo de la vena media con escamas oscuras. A pesar de esto, Faran (1981: 28) considera que las hembras adultas de estas especies no pueden ser separadas, lo cual es evidente en su clave. Según las claves de Cova-García (1961), Gorham et al., (1967), Consoli & de Oliveira (1994), Rubio-Palis (2000) y González & Carrejo (2007), el carácter diagnóstico de aquasalis son las escamas oscuras y claras presentes en la porción no bifurcada de la vena media (M) alar y benarrochi con escamas predominantemente oscuras, mientras que Faran (1981) describe a benarrochi con la presencia o ausencia de una mancha sectoral oscura y escamas predominantemente claras en la porción no bifurcada de la vena media (M); pudiéndose inferir que el patrón del color de las escamas del ala puede variar dependiendo de las condiciones ambientales tal como ha sido demostrado para otro Nyssorhynchus, An. nuneztovari (Hribar, 1995; 1997).
En relación a los resultados de la identificación de la genitalia masculina se obtuvo que todos los ejemplares analizados presentaron las características morfológicas correspondientes a la especie An. aquasalis y concordó filogenéticamente los haplotipos A2-M y CA4-M con la misma entidad de todas las secuencias. La gran similaridad entre hembras de An. aquasalis y An. benarrochi, conduce hasta ahora hacia sólo poder ser identificadas como adultos a través de los caracteres presentes en la genitalia masculina. An. aquasalis se identifica por: 1.- ápice del claspete ventral moderadamente estrecho, ancho en el ápice sobre 0.3 del largo del claspete; 2.- lóbulo basal del claspete ventral ancho con espículas distribuidas uniformemente, espículas a lo largo del margen basal igual o levemente más largo que la anchura del aedeagus y usualmente no curveadas; 3.- superficie lateral y ventral del claspete ventral con espículas cortas 0.5 o menos que el ancho del aedeagus y 4.- placa preapical del claspete ventral moderadamente pequeño, ovalado a circular y débil a fuertemente esclerotizado (Faran, 1980; Cova-García & Sutil, 1977).
Genéticamente, con la información aportada por las secuencias obtenidas podemos sugerir que COI del ADN mitocondrial es un gen funcional para definir relaciones en niveles infragenéricos y además es un buen marcador de las relaciones evolutivas entre poblaciones de una misma especie y entre especies cercanas, lo cual lo hace de importancia filogenética. Es por ello, que este gen evidenció diferencias entre especies An. aquasalis y An. albimanus, y nos proporcionó certeza que el clado politómico encontrado corresponde a una única especie, An.aquasalis, corroborando lo obtenido con la identificación de la genitalia masculina.
Especial referencia se merece el caso de An. benarrochi, el cual se mantiene como una entidad taxonómica confusa. Esta especie tiene como localidad tipo La Ceiba, edo. Trujillo, Venezuela, sin embargo, debido a la inexistencia del holotipo, las colectas e identificaciones realizadas recientemente por diferentes autores carecen de verificación morfológica confiable (Ruiz et al., 2005; Quiñones et al., 2006). Algunas identificaciones parecen coincidir con una variante del subgrupo Oswaldoi, sin embargo no es posible asignar una denominación taxonómica hasta comparar con ejemplares colectados con topotipos, lo cual ha sido infructuoso de obtener en par de ocasiones (R Wilkerson de WRBU y los autores CH, JCN) debido

Bol. Mal. Salud Amb.46
Invalidación de autapomorfias putativas en Anopheles
muy posiblemente a los grandes cambios ecológicos que presenta la localidad tipo. Faran (1980) revisó 286 ejemplares provenientes de crias asociadas de Villavicencio (Meta, Colombia) y 7 ejemplares de Loreto (Iquitos, Perú), pero no revisó material de la localidad tipo, no obstante, señala que se debe ser cuidadoso con las comparaciones morfológicas con ejemplares de otras localidades debido a la gran similaridad que existe entre las hembras de benarrochi con aquasalis y en menor grado con strodei. Faran (1980) y Rubio-Palis (2000) resaltan la gran similaridad entre estas especies, asi como también es discutido por González & Carrejo (2007). Esta similaridad hace posible que los ejemplares identificados como An. aquasalis por Brown et al., (2001) en La Ceiba, hayan sido en realidad An. benarrochi, debido particularmente a que esta localidad está muy alejada de zonas costeras.
Quiñones et al. (2001) y Calle et al. (2008) señalan una variante morfológica de An. benarrochi B de Putumayo en Colombia que posee caracteres que se solapan con An. oswaldoi, no obstante la descripción original de benarrochi, las claves y descripciones de Faran (1980) no parecen asociar morfológicamente ambas especies. Por su parte, Ruiz et al. (2005) señalan que una secuencia de benarrochi de Colombia resultó idéntica con una rotulada como parte del complejo oswaldoi de Perú de Marrelli et al. (1999), lo cual según Ruiz y colaboradores sugieren la dificultad y posibilidad de cometer errores de identificación en latinoamérica al usar las claves existentes. Tanto Ruiz et al. (2005) como Quiñones et al. (2006) sugieren la existencia de un complejo de especies en benarrochi y en el subgrupo Oswaldoi, e incluso secuencias de benarrochi de Putumayo y de Perú, son distintas genéticamente con secuencias de benarrochi de Rondonia, Brasil, mientras que una comparación con una secuencia rotulada An. benarrochi (U92325 Danoff-Burg & Conn, en GenBank, 1997) resultó ser An albimanus (Ruiz et al., 2005).
Los resultados obtenidos en el presente trabajo, demuestran que los caracteres autapomórficos putativos de las claves morfológicas existentes para la identificación taxonómica de hembras adultas de Anopheles no permiten diferenciar entre An. aquasalis y An. benarrochi, apoyando así la revisión de Faran (1980). Por esta razón se requiere de la clarificación de la identidad taxonómica “An. benarrochi” por medio de un topotipo/neotipo y el estudio detallado
de nuevos caracteres morfológicos en la hembras para la identificación correcta de estas especies, y su corroboración con caracteres moleculares y análisis explícitos como es mostrado en esta trabajo.
AGRADECIMIENTOS
A Nelson Moncada, Napoleón León, Alexander Suárez y Víctor Hugo Aguilar por su colaboración en la logística y trabajo de campo. Al personal de Gerencia de Saneamiento Ambiental y Malariología, Región XI, Carúpano, Sucre, Al Lic. Juan Martínez por la elaboración del mapa y Lic Adriana Zorrilla por apoyo en el laboratorio. Financiamiento: FONACIT-LAB No. 2000001593, Grupo CNRA-Fonacit (José Clavijo A. y JC Navarro). G2 000001541 GIM-Fonacit, Subproyecto Ecogenética (Yadira Rangel).
Invalidation of putative autapomorphic characters using molecular phylogenetic a n a l y s e s i n A n o p h e l e s s u b g e n u s Nyssorhynchus
SUMMARY
Collected specimens from a malaric area in Sucre State, Venezuela, using the ad hoc Keys, were identified as Anopheles aquasalis Curry and An. benarrochi Gabaldón, Cova-García and López populations. Additionally, other individuals have shown a mixture of diagnostics characters of both species. As consequence of the epidemiological importance that both species represents in this malaric area and its correct identification for vector control aims, we addressed the research to the determination of both entities by mean of phylogenetic methods using mitochondrial DNA (Cytochrome Oxidase 1) from these three entities. In order to evaluate the phylogenetic species concept the Maximum Parsimony analyses showed strong supported politomies (unresolved nodes), yielded solutions with no supported groups correlated with two or three morphological entities including the male haplotype identified as An. aquasalis. The genetic distances (K-2P) showed that the variability inter-morphotype is in the variance intra, suggesting that the haplotypes belongs to the three morphotypes represents a unique polymorphic species with phenotypic plasticity. We conclude that An. aquasalis is the unique taxonomic entity in the populations studied and also that the

Vol. XLIX, N° 2, Agosto-Diciembre, 2009 47
Navarro J. C. et al.
morphological characters as: 1) Medial vein with black scales and 2) maxillary palpus with ventral surface covered with white scales, proposed in most of keys used are not valid characters to distinguish females of An. benarrochi from An. aquasalis but also such characters belongs to the intra-specific variation of An. aquasalis. This conclusion support the Faran (1980) key, which did not report any morphological characters, showing undistinguished females under classical characters used.
Key words: aquasalis, benarrochi, cladistics, malaria, mitochondrial DNA, morphological keys, phylogenetic species, Venezuela.
REFERENCIAS
Arrivillaga J. C., Norris D. E., Feliciangeli M. D. & Lanzaro G. C. (2002). Phylogeography of the Neotropical sand fly Lutzomyia longipalpis inferred from mitochondrial DNA sequences. Infection, Genetics and Evolution. 2: 83-95.
Belkin J. N., Hogue C. L., Galindo P., Aitken T. H. G., Schick R. X. & Powder W. A. (1965). Mosquito studies (Diptera, Culicidae). II. Methods for the collection, rearing and preservation of mosquitoes. Contrib. Am. Entomol. Inst. 1: 19-78.
Berti J., Zimmerman R. & Amarista J. (1993). Spatial and temporal distribution of anopheline larvae in two malarious areas in Sucre state, Venezuela. Mem. Inst. Oswaldo Cruz. 88: 353-362.
Bender W., Spierer P. & Hogness D. S. (1983). Chromosomal walking and jumping to isolate DNA from the ace and rosy loci and the bithorax complex in Drosophila melanogaster. J. Mol. Biol. 168: 17-33.
Brown E., Villegas C. & Araujo R. (2001) Abundancia y diversidad de la fauna anofelina en areas vulnerables a la transmisión malárica del estado Trujillo, Venezuela. Bol. Malarial. Saneam. Amb. 41: 1-10.
Carpenter J. M. (1996). Uninformative bootstrapping. Cladistics. 12: 177-181.
Calle D. A., Quiñones M. L., Erazo H. F. & Jaramillo N. (2008). Discriminación por morfometría geométrica de once especies de Anopheles
(Nyssorhynchus) presentes en Colombia. Biomedica. 28: 371-85
Chadee D. D. & Kitron U. (1999). Spatial and temporal patterns of imported malaria cases and local transmission in Trinidad. Am. J. Trop. Med. Hyg. 61: 513-517.
Conn J. E., Cockburn A. F. & Mitchell S. E. (1993)a. Population differentiation of the malaria vector Anopheles aquasalis using mitochondrial DNA. J. Heredity. 84: 248-253.
Conn J., Rangel Y. & Seawright J. A. (1993)b. A New Cytotype of Anopheles nuneztovari from Western Venezuela and Colombia. J. Am. Mosq. Control Assoc. 9: 294-301.
Consoli Rotraut A. G. B. & Lourenço de Oliveira R. (1994). Principais Mosquitos do Importancia Sanitaria no Brasil. Editora FIOCRUZ, Rio do Janeiro, Brasil 225 pp.
Cova-García P. (1961). Notas sobre los Anofelinos de Venezuela y su identificación. 2ª Edición. Editora Grafos. Caracas. 213 pp.
Cova-García P. & Sutil E. (1977). Claves gráficas para la identificación de anofelinos de Venezuela. Publicación de la división de endemias Rurales, Dirección de Malariologia y Saneamiento Ambiental. Ministerio de Sanidad y Asistencia Social.
Curry D. P. (1932). Some observations of the Nyssorhynchus group of the Anopheles (Culicidae) of Panama. Am. J. Hyg. 15: 566-572.
Da Silva A. N. M., Santos C. B., Lacerda R. N., Machado R. L. D. & Póvoa M. M. (2006). Susceptibility of Anopheles aquasalis and An. darlingi to Plasmodium vivax VK210 and VK247. Mem. Inst. Oswaldo Cruz, Rio de Janeiro. 101: 547-550.
Deane L. M., Causey O. R. & Deane M. P. (1948). Notas sobre a distribuicao e a biologia dos anofelinos das Regiones Nordestina e Amazonica do Brasil. Rev. Serv. Esp. Saude Publ. 2: 793-808.
Faran M. E. (1980). Mosquito studies (Diptera, Culicidae) XXXIV. A revision of the Albimanus

Bol. Mal. Salud Amb.48
Invalidación de autapomorfias putativas en Anopheles
section of the subgenus Nyssorhynchus of Anopheles. Contrib. Am. Entomol. Inst. 15: 1-215.
Faran M. E. & Linthicum K. J. (1981). A handbook of the Amazonian species of Anopheles (Nyssorhynchus) (Diptera: Culicidae). Mosq. Syst. 13: 1-81.
Fairley T. L., Povoa M. M. & Conn J. E. (2002). Evaluation of the Amazon River delta as a barrier to gene flow for the regional malaria vector, Anopheles aquasalis (Diptera: Culicidae) in northeastern Brazil. J. Med. Entomol. 39: 861-869.
Fairley T. L., Kilpatrick C. W. & Conn J. E. (2005). Intragenomic Heterogeneity of Internal Transcribed Spacer rDNA in Neotropical Malaria Vector Anopheles aquasalis (Diptera: Culicidae) J. Med. Entomol. 42: 795-800.
Farris J. S. (1969). A successive approximations approach to character weighting. Syst. Zool. 26: 269-276.
Felsenstein J. (1985). Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 39: 783-791.
Gabaldón A., Cova-García P. & López J. A. (1941). Estudio sobre Anophelinos. Serie II. Anopheles (Nyssorhynchus) benarrochi, una especie de la subserie Triannulatus. Publ. Div. Malariol. Caracas. 7: 3-24.
González R. & Carrejo N. (2007). Introducción al estudio taxonómico de Anopheles de Colombia claves y notas de distribución. Publicación de Grupo de Investigaciones Entomológicas, Fac Ciencias Naturales y Exactas, Universidad del Valle, Cali, Colombia. 237 pp.
Gorham J. R., Stojanovich C. J. & Scott H. G. (1967). Clave Ilustrada para los Mosquitos Anofelinos de Sudamerica Oriental. National Communicable Disease Center, Atlanta, 62 pp.
Grillet M. E. (2000). Environmental factors associated with the spatial and temporal distribution of Anopheles aquasalis and Anopheles oswaldoi in
wetlands of an endemic malaric area in northeastern Venezuela. J. Med. Entomol. 37: 231-238.
Guimarães J. H. (1997). Systematic Database of Diptera of the Americas South of the United States, Family Culicidae. São Pablo: Fundação de Amparo a Pesquisa do Estado de São Paulo/Ed. Pleide. 286 p.
Harbach R. E. (2004). The classification of genus Anopheles (Diptera: Culicidae): a working hypothesis of phylogenetic relationships. Bull. Entomol. Res. 94: 537-553.
Herbeck J. T. & Novembre J. (2003). Codon usage patterns in Citochrome Oxidase I across multiplee insects orders. J. Molecular Evolution. 56: 692-701.
Hribar L. (1995). Costal wing spot variation within and among progeny of single female Anopheles nuneztovari (Diptera: Culicidae). Mosq. Syst. 27: 1-15.
Hribar L. (1997). Effect of larval rearing temperatures on adult female morphology of Anopheles (Nyssorhynchus) albimanus (Diptera: Culicidae). Invert. Reprod. Develop. 32: 85-88.
Kambhampati S. & Smith P. T. (1995). PCR primers for the amplifcation of four insect mitochondrial gene fragments. Insect. Mol. Biol. 4: 223-236.
Kitzmiller J. B. & Chow G. W. (1971). The salivary gland chromosomes of Anopheles aquasalis. Rev. Bras. Malariol. Doencas Trop. 23: 65-85.
Laubach H. E., Validum L., Bonilla J. A., Agar A., Cummings R., Mitchell C., Cuadrado R. R. & Palmer C. J. (2001).Identifcation of Anopheles aquasalis as a possible vector of malaria in Guyana, South America. West Indian Med. J. 50: 319-21.
Latorre A., Moya A. & Ayala F. J. (1986). Evolution of mitochondrial DNA in Drosophila subobscura. Proc. Natl. Acad. Sci. USA. 83: 8649-8653.
Linley J. R., Lounibos L. P. & Conn J. E. (1993). A description and morphometric analysis of the eggs of four South American populations of Anopheles (Nyssorhynchus) aquasalis (Diptera: Culicidae). Mosq. Syst. 25: 198-214.

Vol. XLIX, N° 2, Agosto-Diciembre, 2009 49
Navarro J. C. et al.
Lounibos L. P. & Conn J. E. (2000). Malaria vector heterogeneity in South America. Am. Entomol. 46: 238-249.
Maddison W. P. & Maddison D. R. (2003). MacClade: analysis of phylogeny and character evolution. Version 4.06. Sinauer Associates, Massachusetts.
Maldonado V., Finol H. J. & Navarro J. C. (1997). A comparison of the Anopheles aquasalis Curry 1932 eggs in two Venezuelan localities by means of scanning electron microscopy. Mem. Inst. Oswaldo Cruz. Brasil. 92: 487-491.
Marrelli M. T., Malafronte R. S., Flores-Mendoza C., Lourenco-De-Oliveira R., Kloetzel J. K. & Marinotti O. (1999). Sequence analysis of the second internal transcribed spacer of ribosomal DNA in Anopheles oswaldoi (Diptera: Culicidae). J. Med. Entomol. 36: 679-684.
Moncada Perez A. & Conn J. E. (1992). The larval polytene chromosomes of Anopheles aquasalis (Diptera: Culicidae) from Venezuela. Genome. 35: 327-331.
Moritz C., Dowling T. E. & Brown W. M. (1987). Evolution of animal mitochondrial DNA: relevance for population biology and systematics. Ann. Rev. Ecol. Syst. 18: 269-292.
Morlais, I. & Severson D. W. (2002). Complete mitochondrial DNA sequence and amino acid analysis of the cytochrome C oxidase subunit I (COI) from Aedes aegypti. DNA Seq. 13: 123-7.
Navarro J. C. & Weaver S. C. (2004). Molecular Phylogeny of the Vomerifer and Pedroi Groups in the Spissipes Section of Culex (Melanoconion). J. Med. Entomol. 41: 575-581.
Póvoa M. M., Conn J. E., Schlichting C. D., Amaral J. C. O. F., Segura M. N. O., Da Silva A. N. M., Dos Santos C. C. B., Lacerda R. N. L., De Souza R. T. L., Galiza D., Santa Rosa E. P. & Wirtz R. A. (2003). Malaria vectors, epidemiology and the re-emergence of Anopheles darlingi in Belém, Pará, Brazil. J. Med. Entomol. 40: 379-86.
Quiñones M. L., Harbach R. E., Calle D. A., Ruiz F., Erazo H. F. & Linton Y-M. (2001). Variante
morfológica de adultos hembras de Anopheles benarrochi (Diptera: Culicidae) en Putumayo, Colombia. Biomédica. 21: 351-359.
Rubio-Palis Y. (2000). Anopheles (Nyssorhynchus) de Venezuela: Taxonomía, Bionomía, Ecología e Importancia Médica. Escuela de Malariologia y Saneamiento Ambiental, Maracay, Venezuela, 120 pp.
Rangel Y., Vele G., Chadee D & Berti J. (1998). Estudios biosistemáticos de Anopheles aquasalis y sus implicaciones para el control de la malaria en el Estado Sucre. Bol. Malariol. y Saneam. Amb. 38: 31-37
Ruiz F., Quiñones M. L., Erazo H. F., Calle D. A., Alzate J. F. & Linton Y-M. (2005). Molecular differentiation of Anopheles (Nyssorhynchus) benarrochi and An. (N.) oswaldoi from Southern Colombia. Mem. Inst. Oswaldo Cruz. 100: 155-160.
Sallum M. A. M., Shultz T. R. & Wilkerson R. C. (2000). Phylogeny of Anophelinae (Diptera Culicidae) Based on Morphological Characters. Ann. Entomol. Soc. Am. 93: 745-775.
Sallum M. A. M., Schultz T. R., Foster P. G., Aronstein K., Wirtz R. A. & Wilkerson R. C. (2002). Phylogeny of Anophelinae (Diptera: Culicidae) based on nuclear ribosomal and mitochondrial DNA sequences. Syst. Entomol. 27: 361-382.
Scarpassa V. M. (2005). Isozyme similarity in Anopheles oswaldoi sensu lato (Diptera: Culicidae) from the Amazon Region, Brazil. J. Med. Entomol. 42: 319-326.
Sequencher 4.4.2 (2004). Gene Codes Corp., Ann Arbor, MI.
Sierra D. M., Velez I. D. & Linton Y-M. (2004). The malaria vector Anopheles (Nyssorhynchus) nuneztovari Gabaldón comprises one genetic species in Colombia based on homogeneity of nuclear ITS2 rDNA. J. Med. Entomol. 41: 302-307.
Simon C., Frati F., Beckenbach A., Crespi B., Liu H. & Flook P. (1994). Evolution, weighting,

Bol. Mal. Salud Amb.50
Invalidación de autapomorfias putativas en Anopheles
and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved PCR primers. Ann. Entomol. Soc. Am. 87: 651-701.
Simon C., Buckley T. R., Frati F., Stewart J. B. & Beckenbach A. T. (2006). Incorporating Molecular Evolution into Phylogenetic Analysis, and a New Compilation of Conserved Polymerase Chain Reaction Primers for Animal Mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 37: 545–79
Steiner W. W. M., Narang S., Kitzmiller J. B. & Swofford D. L. (1982). Genetic divergence and evolution in Neotropical Anopheles (Subgenus Nyssorhnchus), pp. 523-550. In Recent developmentsin the genetics of insect disease vectors. Steiner W. W. M., Tabachnik W. J., Rai K. S., and Narang S. [eds.]. Stipes Pub Co., Champaign, IL.
Swofford D. L. (2002). “Phylogenetic Analysis Using Parsimony”. PAUP version 4.010b. Sinauer Assoc., Inc., Sunderland, MA.
Sutil O. E. (1980) Enumeración histórica y geográfica de las especies de Culicidae de Venezuela ordenadas según su taxonomía. Bol. Dir. Malariol. San. Amb. 20: 1-32.
Tamura K., Dudley J., Nei M. & Kumar S. (2007) MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Molecular Biology and Evolution. 24: 1596-1599.
WRBU. http://www.mosquitocatalog.org/species/species_profile.asp?ID=293 [Consultado, 16 Febrero, 2009).
RecibidoelXX/XX/2008AceptadoelXX/XX/2009
Vol. XLIX, N° 2, Agosto-Diciembre, 2009 51
Navarro J. C. et al.
Anexo 1. Matriz de alineamiento de secuencias.
(continua Anexo 1)

Bol. Mal. Salud Amb.52
Invalidación de autapomorfias putativas en Anopheles
(viene de Anexo 1)
Vol. XLIX, N° 2, Agosto-Diciembre, 2009 53
Navarro J. C. et al.
(viene de Anexo 1)





















