
Interspecific competition impacts on the morphology and distribution of fine roots in European beech...
12
ORIGINAL PAPER A. Bolte I. Villanueva Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.) Received: 1 December 2004 / Accepted: 28 July 2005 / Published online: 27 September 2005 ȑ Springer-Verlag 2005 Abstract Morphology and vertical distribution patterns of spruce and beech live fine roots (diameter £ 2 mm) were studied using a soil core method in three com- parable mature stands in the Solling: (1) pure beech, (2) pure spruce and (3) mixed spruce–beech. This study was aimed at determining the effects of inter- specific competition on fine root structure and spatial fine root distribution of both species. A vertical stratification of beech and spruce fine root systems was found in the mixed stand due to a shift in beech fine roots from upper to lower soil layers. Moreover, compared to pure beech, a significantly higher specific root length (SRL, P<0.05) and specific surface area (SSA, P<0.05) were found for beech admixed with spruce (pure beech/mixed beech SRL 16.1–23.4 m g 1 , SSA 286–367 cm 2 g 1 ). Both indicate a flexible ‘for- aging’ strategy of beech tending to increase soil exploitation and space sequestration efficiency in soil layers less occupied by competitors. Spruce, in con- trast, followed a more conservative strategy keeping the shallow vertical rooting and the root morphology quite constant in both pure and mixed stands (pure spruce/mixed spruce SRL 9.6/7.7 m g 1 , P>0.10; SSA 225/212 cm 2 g 1 , P>0.10). Symmetric competition belowground between mixed beech and spruce was observed since live fine roots of both species were under-represented compared to pure stand. However, the higher space sequestration efficiency suggests a higher competitive ability of beech belowground. Keywords Belowground competition Soil cores Live fine roots Vertical fine root distribution Fine root biomass Fine root length Root area index Specific root length Specific surface area Introduction In the low mountain ranges of north Germany, the areas of mixed forests comprising European beech (Fagus sylvatica) and Norway spruce (Picea abies) are increas- ing through current silvicultural practices favouring close-to-nature forest management (Otto 1992). Mixed forests are considered to be less susceptible to abiotic hazards like wind throw and snow break than pure stands (Otto 1990; Ro¨hrig and Gussone 1990; Burschel and Huss 1997; Duchiron 2000; Schu¨tz 2001). Comple- mentary resource exploitation by mixed tree species due to their different tree height, crown form, rooting depth and/or phenology is believed to reduce competition but also increase stability and often productivity in mixed forests (Kelty 1992). In addition to interactions above- ground, the complementary rooting patterns of spruce and beech are believed to produce positive effects on mechanical stand stability and resource exploitation in mixed spruce–beech stands due to the deeper root pen- etration of soils (Ko¨stler et al. 1968; Otto 1986). How- ever, few studies have quantified interspecific competition or the competitive ability of spruce and beech in mixed stands using root structure data (Grams et al. 2002; Schmid 2002). Competition between plant individuals, brought about by a shared requirement for a limited resource, influences individuals’ performance, i.e. survival, growth and reproduction (Begon et al. 1996). Root systems are the link between the plant, soil water, and nutrient re- sources (Atkinson 2000). Thus belowground competi- tion in forests is influenced by tree root system characteristics. The uptake of water and nutrients occurs primarily in the finest unsuberized tree roots with a diameter of 2 mm or less (defined in Bolte et al. 2003). Communicated by Hans Pretzsch A. Bolte (&) I. Villanueva Institute of Silviculture, Dept. I, Georg-August-University Go¨ttingen, Bu¨sgenweg 1, 37077 Go¨ttingen, Germany E-mail: [email protected] Tel.: +49-551-393650 Fax: +49-551-393270 Eur J Forest Res (2006) 125: 15–26 DOI 10.1007/s10342-005-0075-5
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Interspecific competition impacts on the morphology and distribution of fine roots in European beech...

ORIGINAL PAPER
A. Bolte Æ I. Villanueva
Interspecific competition impacts on the morphology and distributionof fine roots in European beech (Fagus sylvatica L.) and Norwayspruce (Picea abies (L.) Karst.)
Received: 1 December 2004 / Accepted: 28 July 2005 / Published online: 27 September 2005� Springer-Verlag 2005
Abstract Morphology and vertical distribution patternsof spruce and beech live fine roots (diameter £ 2 mm)were studied using a soil core method in three com-parable mature stands in the Solling: (1) pure beech,(2) pure spruce and (3) mixed spruce–beech. Thisstudy was aimed at determining the effects of inter-specific competition on fine root structure and spatialfine root distribution of both species. A verticalstratification of beech and spruce fine root systemswas found in the mixed stand due to a shift in beechfine roots from upper to lower soil layers. Moreover,compared to pure beech, a significantly higher specificroot length (SRL, P<0.05) and specific surface area(SSA, P<0.05) were found for beech admixed withspruce (pure beech/mixed beech SRL 16.1–23.4 m g�1,SSA 286–367 cm2 g�1). Both indicate a flexible ‘for-aging’ strategy of beech tending to increase soilexploitation and space sequestration efficiency in soillayers less occupied by competitors. Spruce, in con-trast, followed a more conservative strategy keepingthe shallow vertical rooting and the root morphologyquite constant in both pure and mixed stands (purespruce/mixed spruce SRL 9.6/7.7 m g�1, P>0.10; SSA225/212 cm2 g�1, P>0.10). Symmetric competitionbelowground between mixed beech and spruce wasobserved since live fine roots of both species wereunder-represented compared to pure stand. However,the higher space sequestration efficiency suggests ahigher competitive ability of beech belowground.
Keywords Belowground competition Æ Soil cores Æ Livefine roots Æ Vertical fine root distribution Æ Fine rootbiomass Æ Fine root length Æ Root area index Æ Specificroot length Æ Specific surface area
Introduction
In the low mountain ranges of north Germany, the areasof mixed forests comprising European beech (Fagussylvatica) and Norway spruce (Picea abies) are increas-ing through current silvicultural practices favouringclose-to-nature forest management (Otto 1992). Mixedforests are considered to be less susceptible to abiotichazards like wind throw and snow break than purestands (Otto 1990; Rohrig and Gussone 1990; Burscheland Huss 1997; Duchiron 2000; Schutz 2001). Comple-mentary resource exploitation by mixed tree species dueto their different tree height, crown form, rooting depthand/or phenology is believed to reduce competition butalso increase stability and often productivity in mixedforests (Kelty 1992). In addition to interactions above-ground, the complementary rooting patterns of spruceand beech are believed to produce positive effects onmechanical stand stability and resource exploitation inmixed spruce–beech stands due to the deeper root pen-etration of soils (Kostler et al. 1968; Otto 1986). How-ever, few studies have quantified interspecificcompetition or the competitive ability of spruce andbeech in mixed stands using root structure data (Gramset al. 2002; Schmid 2002).
Competition between plant individuals, broughtabout by a shared requirement for a limited resource,influences individuals’ performance, i.e. survival, growthand reproduction (Begon et al. 1996). Root systems arethe link between the plant, soil water, and nutrient re-sources (Atkinson 2000). Thus belowground competi-tion in forests is influenced by tree root systemcharacteristics. The uptake of water and nutrients occursprimarily in the finest unsuberized tree roots with adiameter of 2 mm or less (defined in Bolte et al. 2003).
Communicated by Hans Pretzsch
A. Bolte (&) Æ I. VillanuevaInstitute of Silviculture, Dept. I, Georg-August-UniversityGottingen, Busgenweg 1, 37077 Gottingen,GermanyE-mail: [email protected].: +49-551-393650Fax: +49-551-393270
Eur J Forest Res (2006) 125: 15–26DOI 10.1007/s10342-005-0075-5

Attributes describing the functional fine root architec-ture for space sequestration efficiency and soil resourceexploitation (e.g. surface area and specific root length) aswell as fine root biomass and distribution are thereforeassumed to indicate tree competitive ability below-ground (Fitter 1987; Bauhus and Messier 1999a, Bauhuset al. 2000; Grams et al. 2002).
Recent studies on the vertical fine root distribution ofmature spruce and beech belowground are rare and fo-cus on root mass and density mainly. Two studies inBavaria and Austria comparing mixed and pure standsboth found a vertical stratification of fine root biomassin mixtures, indicating species complementarity: spruceroots were more superficially distributed and beech rootsshifted down to deeper mineral soil layers. However,while Rothe (1997) claimed spruce was more competitivebelowground Schmid (2002) maintained beech was moreso. For beech, a vertical stratification of rooting zone isalso reported in admixes with other tree species (Mac-Queen 1968; Buttner and Leuschner 1994; Rysavy andRoloff 1994; Hendriks and Bianchi 1995; Rust and Savill2000; Leuschner et al. 2001). However, contrastingfindings that vertical root stratification is less pro-nounced, or that beech roots mainly coexist in the samesoil layers as the competing tree species have also beenreported (Ehwald et al. 1961; Curt and Prevosto 2003b,Horn and Murach 2003).
A suppression of roots, indicated by reduced totalfine root biomass in mixtures, was found for spruceadmixed with beech (Schmid and Kazda 2002), silverbirch Betula pendula (Brandtberg et al. 2000), and forboth spruce and beech in mixed stands (Nielsen andMackenthun 1991). Several authors stated that beechdevelops a more dynamic and adaptive root systemcompared to competitive species in mixed stands (Butt-ner and Leuschner 1994; Hertel 1999; Curt and Prevosto2003b, Bolte et al. 2004). All things considered, there areseveral indications for different rooting adaptability andcompetitive ability of spruce and beech, and for a speciescomplementarity belowground.
The present comparative investigation was carriedout in three adjacent mature stands (pure beech, purespruce and mixed spruce–beech) with comparable cli-mate and site properties. The objective of the study wasto: (1) determine how interspecific competition affectedfine root morphology and the vertical fine root distri-bution of spruce and beech, and (2) evaluate the com-petition direction and competitive ability of bothadmixed species belowground.
Materials and methods
Site and stand description
The research area was located in the Solling Mountainsin southern Lower Saxony (latitude 09�39¢E, longitude51�43¢N). Our three study sites represented the block A3‘‘Eschershausen’’ within a block design comprising 24
beech, spruce, and mixed forest stands, combined ineight blocks in three forest age classes (Y: 40–60 years,M: 60–90 years, A: >90 years). This block design wasdeveloped for comparative studies in the cooperativeproject ‘‘Indicators and strategies of a sustainable,multi-functional forestry—case study Solling’’ of theForest Ecosystems Research Centre at Gottingen Uni-versity (Beese et al. 2001).
The sites were situated in the growth district ‘‘HoherSolling’’ at 400 to 450 m a. s. l. on N toNW facing slopeswith 0–5� inclination (pure spruce, pure beech) and on aNE to E facing slope (spruce–beech). The montane cli-mate is classified as humid and sub-continental with alow mean annual temperature (6.5�C), and high meanannual values of precipitation (1,050 mm) and relativehumidity (83.5%) (Otto 1991; Wolff et al. 2003).
At all sites, the soils have developed from the sameparent soil material, a loess (eolic silt cover) over Tri-assic sandstone sediments (Ellenberg et al. 1986), whichis typical of the Solling region. Soil texture was domi-nated by silt (40–65%) and sand (35–50%). The clayproportion is somewhat low (10–15%). In the purebeech and the mixed stand the humus layer is a typicalmor-type humus, whereas in the pure spruce stand, moreorganic material has accumulated and it tends towards araw humus type. All soils are well-drained dystric cam-bisols (podzolic brown earths) with a high moisturestatus. Chemical properties of the soils are given inTable 1. All soils are acidic, occurring in the aluminiumbuffer range usually with limited exchangeable base ca-tions. Liming has not been reported in the stands since1987 (Weckesser 2003). The soil chemical status of allstands is comparable, although in the uppermost min-eral soil layer (0–5 cm) of the pure beech stand thecontribution of K, Ca and Mg to the effective exchangecapacity (CEC) was somewhat higher. This was partlybalanced by a lower CEC. However, this small variationin soil acidity is not expected to influence the fine rootsystem structure of beech as Leuschner et al. (2004)found weak acidification and nutrition effects on fineroot system size and morphology only when comparingbeech forest along a large site gradient from a rendzinasoil (base saturation 98.9%) to a dystric cambisol onglacial sandy deposits (base saturation 3.9%).
The natural forest community can be classified asLuzulo-Fagetum (Oberdorfer 1992) at all sites as spruceis not part of the natural vegetation at Solling (e.g.Firbas 1952). The ground vegetation in the pure beechstand was dominated by Luzula luzoloides and Oxalisacetosella, whereas mainly Deschampsia flexuosa andGalium saxatile covered the forest floor in the purespruce stand. All four species can be found in the mixedspruce and beech sites together with Agrostis capillariesand Rubus idaeus agg., which dominate in some parts ofthe stand (Weckesser 2003).
Spruce was planted in the study area, while the beechoriginated primarily from natural regeneration. How-ever, planted beech sometimes occurs in the pure beechstand. The growth stages of all stands were classified as
16

‘optimum phase’, characterized by a dense overstoreytrees and high wood volume (Walter and Breckle 1991).Due to the different growth dynamics of beech andspruce, mean age of the stands compared ranged from93 years (pure spruce) to 152 years (pure beech) in the2001 sampling year. In the mixed stand, spruce rangedfrom 110 to 125 years old and beech ranged from 127 to135 years old on average, arranged predominantly ingroupings of each species. More stand structure data arepresented in Table 2.
Field sampling
For the field sampling, five circular plots were estab-lished in each pure stand and nine circular plots in themixed stand, where a higher heterogeneity in standstructure was expected (Fig. 1). In all stands, the plotscovered a surface area of 0.05 ha (r @ 12.62 m). Thecircles were distributed within selected tree groups con-sidered representative of average stem density. In themixed stands, we placed the circles in the transitionzones between spruce and beech groups, therebyexcluding stand areas with natural regeneration to avoidthe need to identify roots of mature trees. This proce-
dure resulted in comparable mean stocking degrees ofpure and mixed stand plots (0.8–1.0, Table 2) that weretypical for managed stands in the Solling region. Forfine roots sampling, square plots with 25 m2 surface areawere placed at the centre of all circular plots so that thecorners of the squares were oriented north, east, south,and west. All together five fine root sample points weresituated at the edges and at the centre of the square plot(Fig. 1).
Stem position (azimuth angle, distance to plot cen-tre), stem diameter at breast height (DBH, 1.3 maboveground), and total height of all trees (stem diam-eter > 7 cm) were measured using a girth tape, a com-pass with dioter (Suunto) and a digital hypsometerVertex III (Haglof) respectively. We recorded all treeswithin each circular plot in addition to all adjacent treeswith crowns reaching into the plot. Thus, abovegrounddata sets for trees both within the circular plots, and at adistance of less than 10 m from any of the root samplingpoints could be generated for subsequent analyses.
Fine root sampling was carried out from the end ofSeptember to the beginning of October 2001. Five soilcores with a surface area of 50 cm2 were taken from theplot centre and plot corners (Fig. 1) in each square plotusing a soil auger (d � 8 cm, Murach 1984). The maxi-
Table 2 Structural characteristics of the investigated pure and mixed stand plots
Pure stand Mixed stand
Spruce Beech Spruce Beech Total
Stand age 2001 (a) 93 152 110–125 127–135 110–135Tree number (n ha�1) 272 108 151 76 228Tree height (m) 31.3 28.9 32.4 25.3 30.1DBH (cm) 42.6 53.7 48.9 37.0 44.9Basal area (m2 ha�1) 39.4 25.2 28.8 8.7 37.5Stocking degree 0.8 0.8 0.7 0.3 1.0
Mean values of circular plots (surface 0.05 ha, pure stands: n = 5, mixed stands: n = 9) are shown. DBH is the diameter at breast height(1.3 m above ground), stocking degree refers to yield tables of Schober (1987) for moderate thinning
Table 1 Selected chemical properties of the three stands
Pure spruce stand Mixed stand Pure beech stand
Humus layer n = 5 n = 9 n = 5Thickness (cm) 6 4 – 5 3 – 4pH (KCl) 2.9 – 3.4 2.8 – 4.2 3.4 – 4.7C/N ratio 22.2 – 25.9 20.7 – 27.1 17.9 – 24.6Mineral soil at 0–5 cm (Ahe horizona)pH (KCl) 3.0 – 3.3 3.0 – 3.4 3.3 – 4.1CEC (lmolc g
�1) 104 – 125 76 – 107 87 – 93Al (% CEC) 59.8 – 73.5 53.3 – 68.3 43.3 – 43.6K + Ca + Mg (% CEC) 3.5 – 11.3 4.7 – 12.6 27.0 – 33.0
Mineral soil at 5–40 cm (Bv horizona)pH (KCl) 3.7 – 4.2 3.5 – 4.2 4.0 – 4.2CEC (lmolc g
�1) 44 – 69 45 – 78 43 – 48Al (% CEC) 91.9 – 95.5 85.4 – 95.6 88.0 – 94.6K + Ca + Mg (% CEC) 1.7 – 4.9 2.6 – 6.7 3.4 – 7.7
a Classification according to AK Standortskartierung (1996)The ranges between minimum and maximum values of n combined samples consisting of four single samples taken at every subplot aredisplayed. CEC: effective cation exchange capacity
17

mum soil depth sampled was 40 cm, below which a solidbedrock layer was often found. However, much of thefine root biomass (estimated >80%) could be sampledwithin this profile (Murach 1984; Rapp 1991; Buttnerand Leuschner 1994; Puhe 1994; Rothe 1997; Hertel1999).
Within a radius of 10 m around the root samplepoints, the mean distances between the sample point andneighbouring trees did not differ significantly betweenthe stands (pure beech, pure spruce, mixed; H Test,H=4.53, P=0.10). Moreover, no significant variation intree distances could be detected between spruce andbeech in the mixed stand (U Test, Z=0.109, P=0.91).Thus, biased sampling due to different tree densitiesaround the sampling points in the different stands wasunlikely.
The extracted soil cores were divided into six soillayers or depth classes: humus layer Of/Oh, and mineralsoil layers 0 – 5 cm, 5 – 10 cm, 10 – 20 cm, 20 – 30 cmand 30 – 40 cm. Sub-samples of 2 cm width were cut outcentrally from the 5-cm layers and 4-cm sub-samplesfrom the 10-cm layers of the mineral soil. The humuslayer samples were evaluated entirely. According toMurach (1984), this sub-sampling method producesrepresentative results for root biomass and distributionin the upper mineral soil layers with intensive rooting.After extraction, all sub-samples were refrigerated (4�C)and stored for further processing not later than twoweeks after sampling.
Processing and sorting of roots
Mineral soil and humus layer samples were soakedovernight. Roots of the mineral soil were then separated
from soil material using the floating method (Bohm1979). Watered samples were poured into trays androots floating on top of the water were sieved (1-mmmesh). This procedure was repeated until only rockswere left in the soil sample. Root material was extractedfrom fine soil organic matter in the Of/Oh samples bygentle washing. Roots and the remaining soil organiccompounds, dispersed in water filled containers, wereseparated manually. Washed root samples were kept indeionised water at 4�C until sorting.
All root parts less than or equal to 2 mm in diam-eter were classified as fine roots (Bolte et al. 2003) andseparated. Root fractions greater than 2 mm in diam-eter were not investigated. All fine roots were thensorted according to vitality (live, dead) and species(spruce, beech, shrubs/herbs). To assess vitality, themorphological criteria of dark discolouration of thecentral cylinder and a decreased flexibility of rootsegments were used to identify dead root parts (e.g.Murach 1984; Bauhus and Messier 1999a). Live rootswere differentiated by species. Spruce and beech rootswere identified visually according to root elasticity androot cortex properties: spruce roots are elastic with arelatively thick and irregularly structured brownishcortex, whereas beech roots are less elastic and the red-brown cortex is thin with lines along the longitudinalaxis (Schmid 2002).
Assessment of fine root attributes
Selected fine root attributes of spruce and beech indi-cating soil exploitation and competitive ability, such asspecific root length SRL (Bauhus et al. 2000) wereanalyzed using the digital image analysis system ‘‘Win-RHIZO V3.10‘‘ (Regents Instruments Inc., Canada).The roots were placed in a transparent water filled tray(10 cm·15 cm) on a scanner to facilitate root spreading.The system scanned all fine root fragments and theprogram calculated architectural traits like root lengthand root surface for all fine roots and 0.2-mm rootsections. Tests by Bauhus and Messier (1999b) using thesame technique showed negligible errors for fine rootstructure measurements through root overlapping andabutment.
After scanning, all root fragments were dried for 48 hat 40�C and weighed.
Based on the results, specific root length SRL andspecific root surface area SSA could be determined fromthe ratio of fine root length, and fine root surface arearespectively, and dry weight of fine root biomass.
Data analysis
In order to compare fine root attributes (biomass, sur-face area and length) of pure and mixed stand plots withvariable species abundance aboveground, total corevalues of spruce and beech were adjusted to one unit of
Fig. 1 Plot design for stand assessment (circular plot, 0.05 ha) andfine root sampling (central square, 25 m2) in a mixed stand plot (no.708); crosses at the corners of the central plots indicate soil coresampling locations
18
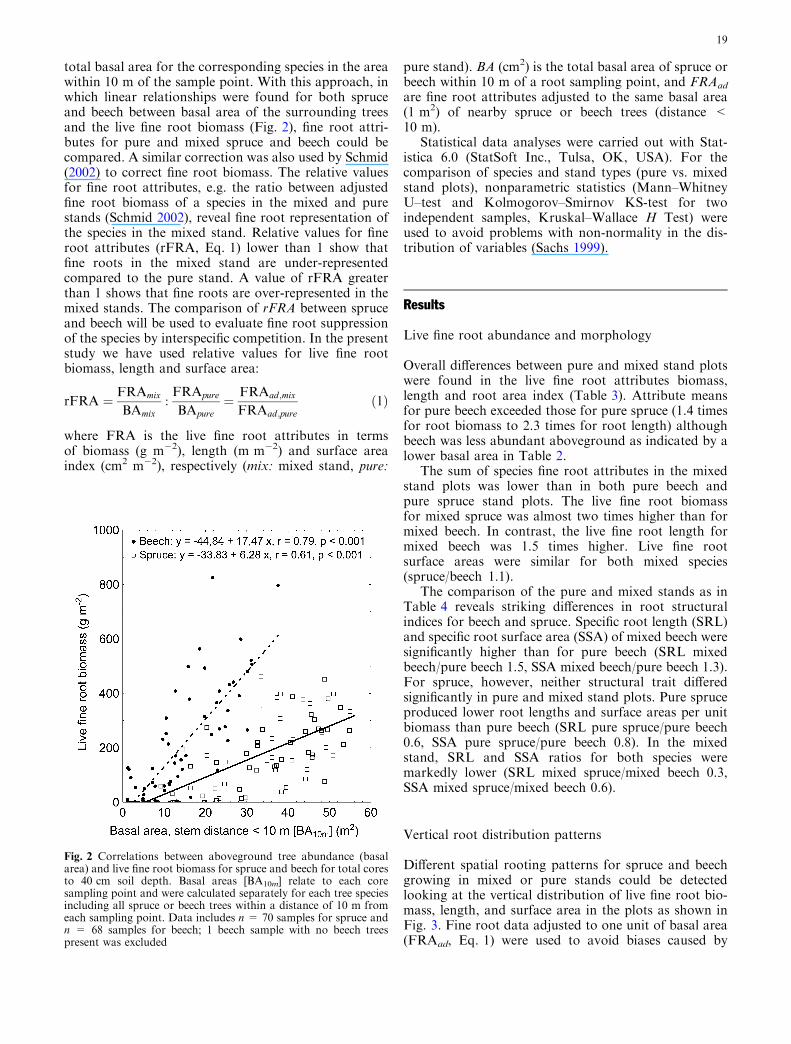
total basal area for the corresponding species in the areawithin 10 m of the sample point. With this approach, inwhich linear relationships were found for both spruceand beech between basal area of the surrounding treesand the live fine root biomass (Fig. 2), fine root attri-butes for pure and mixed spruce and beech could becompared. A similar correction was also used by Schmid(2002) to correct fine root biomass. The relative valuesfor fine root attributes, e.g. the ratio between adjustedfine root biomass of a species in the mixed and purestands (Schmid 2002), reveal fine root representation ofthe species in the mixed stand. Relative values for fineroot attributes (rFRA, Eq. 1) lower than 1 show thatfine roots in the mixed stand are under-representedcompared to the pure stand. A value of rFRA greaterthan 1 shows that fine roots are over-represented in themixed stands. The comparison of rFRA between spruceand beech will be used to evaluate fine root suppressionof the species by interspecific competition. In the presentstudy we have used relative values for live fine rootbiomass, length and surface area:
rFRA ¼ FRAmix
BAmix:FRApure
BApure¼ FRAad;mix
FRAad;pureð1Þ
where FRA is the live fine root attributes in termsof biomass (g m�2), length (m m�2) and surface areaindex (cm2 m�2), respectively (mix: mixed stand, pure:
pure stand). BA (cm2) is the total basal area of spruce orbeech within 10 m of a root sampling point, and FRAad
are fine root attributes adjusted to the same basal area(1 m2) of nearby spruce or beech trees (distance <10 m).
Statistical data analyses were carried out with Stat-istica 6.0 (StatSoft Inc., Tulsa, OK, USA). For thecomparison of species and stand types (pure vs. mixedstand plots), nonparametric statistics (Mann–WhitneyU–test and Kolmogorov–Smirnov KS-test for twoindependent samples, Kruskal–Wallace H Test) wereused to avoid problems with non-normality in the dis-tribution of variables (Sachs 1999).
Results
Live fine root abundance and morphology
Overall differences between pure and mixed stand plotswere found in the live fine root attributes biomass,length and root area index (Table 3). Attribute meansfor pure beech exceeded those for pure spruce (1.4 timesfor root biomass to 2.3 times for root length) althoughbeech was less abundant aboveground as indicated by alower basal area in Table 2.
The sum of species fine root attributes in the mixedstand plots was lower than in both pure beech andpure spruce stand plots. The live fine root biomassfor mixed spruce was almost two times higher than formixed beech. In contrast, the live fine root length formixed beech was 1.5 times higher. Live fine rootsurface areas were similar for both mixed species(spruce/beech 1.1).
The comparison of the pure and mixed stands as inTable 4 reveals striking differences in root structuralindices for beech and spruce. Specific root length (SRL)and specific root surface area (SSA) of mixed beech weresignificantly higher than for pure beech (SRL mixedbeech/pure beech 1.5, SSA mixed beech/pure beech 1.3).For spruce, however, neither structural trait differedsignificantly in pure and mixed stand plots. Pure spruceproduced lower root lengths and surface areas per unitbiomass than pure beech (SRL pure spruce/pure beech0.6, SSA pure spruce/pure beech 0.8). In the mixedstand, SRL and SSA ratios for both species weremarkedly lower (SRL mixed spruce/mixed beech 0.3,SSA mixed spruce/mixed beech 0.6).
Vertical root distribution patterns
Different spatial rooting patterns for spruce and beechgrowing in mixed or pure stands could be detectedlooking at the vertical distribution of live fine root bio-mass, length, and surface area in the plots as shown inFig. 3. Fine root data adjusted to one unit of basal area(FRAad, Eq. 1) were used to avoid biases caused by
Fig. 2 Correlations between aboveground tree abundance (basalarea) and live fine root biomass for spruce and beech for total coresto 40 cm soil depth. Basal areas [BA10m] relate to each coresampling point and were calculated separately for each tree speciesincluding all spruce or beech trees within a distance of 10 m fromeach sampling point. Data includes n = 70 samples for spruce andn = 68 samples for beech; 1 beech sample with no beech treespresent was excluded
19

different stem abundance in mixed and pure stand plots(see Table 2). Due to an asymmetric distribution of data,medians and percentiles were used to display their cen-tral tendency and dispersion (compare with Lorenz1996).
For spruce, adjusted fine root biomass, length andsurface area in mixed stands were significantly lowerthan in the pure stands at all soil depths except the topsoil layer (0–5 cm depth, see Figs. 3.1a to 3.3a). Mixedspruce fine roots were predominantly concentrated inthe humus layer and the upper mineral soil down to5 cm soil depth. According to the displayed medians, atleast half the mixed spruce samples did not contain anylive fine roots at mineral soil depths of 10 cm or below.Fine roots of pure spruce were also mostly abundant inthe humus layer, although they were also present inmineral soil down to 20 cm soil depth. Further thebiomass, length and surface area was higher for spruceat depths 20–40 cm than for mixed spruce in mostsamples.
For beech, significant differences in all fine structuraltraits occurred between the pure and mixed stand plotsin the humus layer and the upper mineral soil down to10 cm depth (Figs. 3(1b), 3(3b)). Fine root abundance inmixed beech was remarkably low in the upper soil layers;
most samples did not contain any live fine roots, similarto the results for mixed spruce in the deeper soil layers.In deeper soil layers from 10–40 cm soil depth, beechfine root abundance, particularly fine root length andsurface area, increased noticeably due to high specificfine root length and surface area (SRL, SSA). For purebeech, in contrast, fine roots in the top soil down to20 cm soil depth were more evenly distributed; themaximum root biomass, length and surface area oc-curred in the humus layer. However, root abundancedown to 20 cm soil depth was high, declining in thedeeper horizons.
The comparison of beech and spruce in the mixedstand plots (see solid lines in Fig. 3) revealed a nearlycomplete spatial stratification of vertical fine root dis-tribution and soil resource exploitation. Spruce fineroots were concentrated in the upper soil horizons andthe humus layer with minimal beech fine roots, whilebeech roots occurred predominantly in deeper soil layerswhere spruce roots were virtually absent.
Total fine roots (spruce plus beech) in the mixedstand were clearly more abundant in the deeper soilhorizons (10–40 cm depth) compared to the purespruce stand; yet little difference was observed whenthey were compared to the pure beech stand. In theuppermost soil layers (humus layer, mineral soil to5 cm depth) the total root abundance in the mixedstand was similar, and somewhat lower to that foundin the pure spruce stand, and the pure beech stand,respectively.
Relative fine root attributes
Clear differences in spruce and beech fine root structurecan be observed comparing structural traits adjusted toone unit basal area (BA, Table 5). In both pure andmixed stand plots, the abundance of spruce live fineroots amounted to only 40% or less than that of beech.All adjusted fine root architectural traits for spruce andbeech were lower in the mixed stand compared to thepure stands. This indicated a markedly lower below-ground representation of both species in the mixed
Table 4 Comparison of live fine root structural traits of beech andspruce in pure and mixed stand plots
Parameters Pure stand Mixed stand
Spruce n=25 n=42SRL (m g�1) 9.6 (3.4)a 7.7 (2.7)aSSA (cm2 g�1) 225 (46)a 212 (54)aBeech n=24 n=34SRL (m g�1) 16.1 (4.4)a 23.4 (14.2)bSSA (cm2 g�1) 286 (57)a 367 (166)b
Parameters were specific root length (SRL) and specific root sur-face area (SSA). Means and standard deviation (in parentheses) aredisplayed. Means followed by different letters are significantlydifferent at P < 0.05 (Kolmogorov–Smirnov KS two-sample-test).Reduced sample numbers of mixed stand plots are caused bysamples without any fine roots
Table 3 Live fine root (d £ 2 mm) attributes of spruce and beech in pure and mixed stand plots (25 m2)
Plot type Spruce Beech Total
Live fine root biomass FRB, dry weight (g m�2)Pure stand 290 (80) 412 (174)Mixed stand 134 (114) 72 (97) 206 (104)Live fine root length FRL (km m�2)Pure stand 2.8 (1.2) 6.5 (2.9)Mixed stand 1.0 (1.0) 1.5 (2.3) 2.5 (2.1)Live fine root area index RAI (m2 m�2)Pure stand 6.5 (2.0) 11.5 (4.6)Mixed stand 2.7 (2.3) 2.4 (3.5) 5.1 (3.2)
Mean values and standard deviations (in parentheses) of total cores to max. 40 cm soil depth are displayed; n=25 samples for pure spruce,n=24 samples for pure beech, n=45 samples for mixed spruce and beech
20

stand. Accordingly, relative fine root attributes (rFRA),i.e. the ratio of adjusted mixed and pure stand traits(compare Eq. 1), had values less than 1 and ranged from0.53 to 0.67 for the different traits.
For spruce fine roots, relative biomass exceeded thevalues of relative surface area and length. For beech, incontrast, relative surface area and length of the live fineroots were higher than their biomass.
Fig. 3 Vertical distribution ofthe adjusted live fine rootparameters (FRAad) (1) rootbiomass, (2) root length and (3)root surface area for a Norwayspruce and b European beechgrowing in pure and mixedstands. Whisker plots (median,10/90 percentile) are shownusing root structure data fromeach sample point with rootdata adjusted by the basal areasums of all trees less than 10 mdistant (BA 10 m). Starsindicate significant differencesbetween pure and mixed standvalues at P < 0.05 (Mann–Whitney U test)
21

Discussion
Fine root morphology and distribution are fundamentalroot system traits that control plant soil water andnutrient exploitation. By altering these architecturaltraits, plants adapt their nutrient and water exploitationto the spatially and temporally varying distribution ofresources in soil (Fitter 2002). Limited information ex-ists about the correlation of fine root architecture andcompetitive interactions of different tree species (com-pare with Coomes and Grubb 2000).
Live fine root abundance and morphology
Fine root abundance of spruce and beech in pure andmixed stands has commonly been analyzed using rootmass data at a stand level only (e.g. Gottsche 1972;Nielsen and Mackenthun 1991; Murach 1991; Rothe1997; Schmid 2002). Our mean total live fine root bio-mass for pure stand plots (Table 3) is consistent with theresults of previous investigations for stands in the Sol-ling taken from literature. For pure spruce, the mean livefine root biomass of 290 g m�2 corresponds to datafrom Murach (1991) who found biomass ranged from190 to 330 g m�2 in two pure spruce stands over a 2-yearperiod. For pure beech, the high-live fine root biomass inthis study (412 g m�2) lies centrally within the biomassdata range from the literature (99–617 g m�2; Gottsche1972; Rapp 1991; Bauhus and Bartsch 1996; Hertel1999; Kehong 2000). The high beech fine root biomass ata stand level points to the ability of this tree species toform a dense fine root system (compare with Kostleret al. 1968). The mean total fine root biomass in themixed stand plots (206 g m�2) in our study was lowcompared to values of 435 to 535 g m�2 for two Aus-trian mixed spruce beech stands (Schmid 2002). Thismay be due to the different location, climate, site, andstand characteristics.
In comparison with biomass, live fine root length(FRL) and surface root area (RAI, Table 3) data aremore important in explaining resource exploitation oftrees, and providing additional information on fine root
morphology and exploration (Fitter 2002). For bothparameters, pure spruce and beech values are compa-rable to the few values available for spruce and beech inpure and mixed stands from the literature, with theexception of the lower FRL for beech (spruce: RAI 5.4beech: FRL 18.4, RAI 3–11, Van Praag et al. 1988;Hendriks and Bianchi 1995; Leuschner et al. 2001;2004). The low mixed stand values correspond to thefindings for fine root biomass. However, due to aremarkably high FRL and RAI of mixed beech com-pared to its fine root biomass (Table 3), total mixedstand FRL and RAI approach the pure spruce standlevel. This clearly indicates changed root morphology ofbeech growing in mixed stands.
Considering root system cost/gain ratios for carbonand nutrients for soil exploration and resource uptake inaccordance with the Optimality theory (Bloom et al.1985), exploitation efficiency of trees may increase withthe rise of absorbing root surface area and root lengthper unit biomass (specific surface area SSA, specific rootlength SRL, Bauhus and Messier 1999a, Leuschner et al.2004). In this respect, beech in this study exploited waterand nutrient resources more efficiently when growingadmixed to spruce as displayed by significant higherspecific root lengths (SRL, P < 0.05, Table 4) andspecific root surface areas (SSA, P < 0.05, Table 4)compared to pure beech values. For spruce in contrast,fine root morphology was rather similar in pure andmixed stands. These findings are in agreement with thoseof Grams et al. (2002) who compared pure and mixedstands of spruce and beech in the ‘‘Kranzberg Forst’’near Freising (Bavaria). Curt and Prevosto (2003b) alsoobserved a significant SRL increase in beech regenera-tion with rising competition intensity under a Scots pine(Pinus sylvestris) or silver birch (Betula pendula) canopy,while SRL of the overstorey species decreased or re-mained unchanged. However, one should be aware thatthe use of SRL and SSA as exploitation indicatorssimplifies the complex resource acquisition processes ofroots e.g. assuming a constant area-related uptake rateand neglecting the effects of mycorrhiza (Leuschneret al. 2004). Moreover, increased resource exploitationefficiency of one species indicated by increased SSA andSRL may be a reaction to resource depletion of the
Table 5 Total means, standard deviation (in parentheses) and relative mixed stand values (rFRA, Eq. 1) for different adjusted fine rootattributes (adjusted to one unit basal area BA)
Pure stand Mixed stand rFRA
Spruce n=25 n=45Live fine root biomass (g m�2 m�2 BA) 6.5 (2.0)a 4.4 (3.6)b 0.67Live fine root length (m m�2 m�2 BA) 62.0 (27.0)a 33.1 (30.8)b 0.53Live fine root surface area (cm2 m�2 m�2 BA) 1461.4 (737.6)a 888.5 (497.1)b 0.61Beech n=24 n=44Live fine root biomass (g m�2 m�2 BA) 18.7 (8.0)a 11.1 (20.0)b 0.59Live fine root length (m m�2 m�2 BA) 296.9 (142.7)a 200.1 (322.5)b 0.67Live fine root surface area (cm2 m�2 m�2 BA) 5267.6 (2242.2)a 3375.6 (5632.0)b 0.64
Means followed by different letters are significantly different at P < 0.05 (Kolmogorov–Smirnov KS two-sample-test)
22

competing species (cf. Bauhus et al. 2000). When com-paring beech and spruce seedlings, Wang et al. (2001,2003) found lower nutrient acquisition and accumula-tion efficiencies for beech, although beech SRL wassignificantly higher than spruce SRL.
A slightly different approach introduced by Gramset al. (2002) defines SRL as a measure of space seques-tration efficiency of competing tree species, which relatesdirectly to the competitiveness of different species inoccupying space belowground. On the basis of thisdefinition, beech is the superior ‘belowground competi-tor’ over spruce in our mixed stand plots.
Vertical fine rooting patterns
Beech and spruce exhibited different vertical fine rootdistribution patterns when growing in pure or mixedstand (Fig. 3a). The fine root profile of both pure andmixed stand spruce followed a similar pattern in theuppermost layers, often found for sinker root systems(Kostler et al. 1968; Polomski and Kuhn 1998; Bolteet al. 2003), where fine roots are concentrated. Howeverthis is followed by a modest to sharp decrease in fineroot concentration in pure and mixed stands (cf. Puhe2003).
The completely different vertical fine rooting profilesfor pure and mixed beech (Fig. 3b) corresponds to pre-vious findings (Rothe 1997; Schmid 2002). The quiteequal vertical beech fine root distribution in pure stands,typical for heartroot systems (Kostler et al. 1968; Pol-omski and Kuhn 1998), was contrasted by a shift ofbeech fine roots downwards to deeper soil horizons inresponse to interspecific competition, leading to verticalstratification of beech and spruce fine rooting zones(compare with the solid lines in Fig. 3). Vertical rootingstratification in mixed beech stands, mostly due to anexpansion of beech roots in deeper layers and a super-ficial root concentration of the co-existing species, wasalso found with the admixed species common ash(Fraxinus excelsior, Rysavy and Roloff 1994; Rust andSavill 2000), Sessile oak (Quercus petraea, Buttner andLeuschner 1994; Leuschner et al. 2001), Douglas fir(Pseudotsuga menziesii, Hendriks and Bianchi 1995) andScots pine (Pinus sylvestris, MacQueen 1968; Curt andPrevosto 2003b). Spatial separation of fine root systemsis thought to reduce interspecific competition amongtree species (Nambiar and Sands 1993; Casper andJackson 1997) by the complementary resource exploi-tation in different soil layers (Bennett et al. 2002). In thisstudy, belowground ‘niche partitioning’ (Parrish andBazzaz 1976) was predominantly produced by rootingvariations of beech.
This morphological plasticity of beech fine rootsystems in mixed stands was also stressed by Curt andPrevosto (2003b) with the demonstration of a variablearchitecture in adaptation to different competitionintensities. At low-competition level under silver birch(Betula pendula), fine roots of both competing species
co-existed in the same soil layers, and beech fine rootswere rather coarse and short (beech SRL 8.3 m g�1).However, under Scots pine (Pinus sylvetris) at high-competition level, a vertical stratification of fine rootsystems occurred, and beech fine roots were elongatedand more ramified (beech SRL 13.4 m g�1). Underhigh interspecific competition pressure, beech seems tofollow a ‘root foraging strategy’ that is reported forsubordinate species in order to access soil resourcesless exploited by the dominant species (Grime 1979;Bauhus and Messier 1999a). This may be an elementof beech’s late-successional colonization strategy(Grime 1979), supporting the invasion of existingstands of early- and mid-successional species. Thisflexible rooting and soil exploitation strategy of beechcontrasts with the conservative one of spruce, whichmay be an adaptation to limited C and nutrient sup-ply, thereby avoiding unprofitable fine root develop-ment in deeper soil horizons (Bauhus and Messier1999a). In this respect, the spruce–beech complemen-tarity belowground found in our study may be closelylinked to the species-specific soil exploitation strategiesthat are obviously genetically inherited (Curt andPrevosto 2003a) due to different colonization and siteadaptation strategies.
Relative fine root abundance and competition direction
Relative fine root attributes (RFA), when comparedwith BA adjusted live fine root attributes of mixedwith pure stands (Table 5), indicated negative inter-specific competition effects on fine root abundancesfor both spruce and beech. These results point to aquite symmetrical interspecific competition of maturespruce and beech at montane Solling sites and are inaccordance with the suggestions of Nielsen and Mac-kenthun (1991) for spruce–beech stands at the near-bywestern edges of the Harz Mountains, where theclimate is comparable. In contrast, the findings ofSchmid (2002) for mixed spruce–beech woods indi-cated an asymmetric interspecific root competition infavour of beech on stagnic cambisol sites in LowerAustria. However, the latter study was conductedunder comparatively warmer climate conditions thanfound at our Solling sites with 200 mm lower annualprecipitation (843 mm) and 1.9�K higher mean annualtemperature (8.4�C), which is supposed to favour thecompetitive ability of beech over spruce (Otto 1994;Ellenberg 1996).
The RFA value of mixed beech for root length andsurface area exceeded relative fine root biomass, whereasthe converse was true of spruce RFA (Table 5). Thehigher relative fine root length and surface area of beechcompared to spruce may reveal a better adaptability offine root architecture characteristics which have leadother authors to attribute beech with a dynamic andcompetitive rooting system (Buttner and Leuschner1994; Schmid 2002; Curt and Prevosto 2003a).
23

Conclusions
The present study confirms previous findings thatmixed spruce and beech exhibit a vertical stratificationof fine root systems due to interspecific competition(Rothe 1997; Schmid 2002). The conservative strategyof spruce of keeping its shallow vertical fine root dis-tribution patterns and morphology in both pure andmixed stands contrasts with the flexible ‘foraging’strategy of mixed beech which shifts fine roots fromupper to lower soil layers and increases SRL, SSA andthereby soil exploitation efficiency (Bauhus et al. 2000;Grams et al. 2002). This points to a complementaryexploitation of soil resources in mixed spruce–beechstands compared to pure spruce stands if the spatialrooting and resource uptake patterns coincide, whichis, however, not always the case (George and Mars-chner 1996). Positive tree nutrition effects in the mixedstand facilitated by nutrient uptake of the shallow-rooting spruce in the upper soil, and the deeper rootingbeech in the lower subsoil (‘nutrient pump effect’,Rothe and Binkley 2001) cannot be expected at oursites since no nutrient-rich subsoil layers could bereached by the roots. The shallower fine root systemsof mixed spruce compared to pure spruce may increasedrought vulnerability (Schmid 2002), which is incon-sistent with the commonly held view that stand stabilityis higher in mixtures formed by complementary treespecies (Kelty 1992; Burschel and Huss 1997).
A symmetric competition and similar competitiveability belowground, found in our mature spruce–beechstand, may provide one indication for co-dominance ofboth species in mixture. However, this must be consid-ered with caution because we lack knowledge of theinterspecific competition dynamics of spruce and beechbelowground brought about by their different growthdynamics (e.g. Schober 1987; Kramer 1988). Superiornutrient acquisition efficiencies of spruce fine roots at theregeneration stage (e.g. Wang et al. 2001; 2003) contrastwith higher fine root abundances and space sequestra-tion efficiency in mature beech stands (Grams et al.2002), the latter also found in the present study. Thismay indicate a temporal shift of interspecific root com-petition direction favouring beech with increasing standage.
Acknowledgements This study was supported by the GermanFederal Ministry of Education, Science, Research and Technol-ogy within the cooperative project ‘Indicators and strategies of asustainable, multi-functional forestry—case study Solling’(BMBF FKZ 033947C/3) conducted by the Forest EcosystemResearch Centre at Gottingen University. The authors aregrateful to Andreas Parth, Institute of Silviculture at GottingenUniversity, for his skilful help in sampling, sorting and pro-cessing the fine roots and Dr. Steffen Rust, Dresden Universityof Technology, Institute of Forest Botany and Forest Zoology atTharandt and Dr. Matts Lindbladh, SLU, Southern Forest Re-search Centre at Alnarp (Sweden) for comments on a previousdraft of the manuscript.
References
Atkinson D (2000) Root characteristics: why and what to measure.In: Smit AL, Bengough AG, Engels C, Van Noordwijk M,Pellerin S, Van De Geijn SC (eds) Root methods: a hand book.Springer, Berlin Heidelberg New York, pp 2–32
Bauhus J, Bartsch N (1996) Fine-root growth in beech (Fagussylvatica) forest gaps. Can J For Res 26:2153–2159
Bauhus J, Messier C (1999a) Soil exploitation strategies of fineroots in different tree species of the southern boreal forest ofeastern Canada. Can J For Res 29:260–273
Bauhus J, Messier C (1999b) Evaluation of fine root length anddiameter measurements obtained using RHIZO image analysis.Agron J 91:142–147
Bauhus J, Khanna PK, Menden N (2000) Aboveground andbelowground interactions in mixed plantations of Eucalyptusglobulus and Acacia mearnsii. Can J For Res 30:1886–1894
Beese F (ed) (2001) Indikatoren und Strategien fur eine nachhal-tige, multifunktionelle Waldnutzung – Fallstudie Waldlands-chaft Solling. Zwischenbericht 1999/2000 zum BMBF-Verbundforschungsvorhaben, Teil 1: Gesamtbericht und Ku-rzfassungen. Ber. Forschungsz. Waldokosysteme Univ. Got-tingen (FZW) B 63, pp 1–65
Begon ME, Harper JL, Townsend CR (1996) Ecology: individuals,populations and communities, 3rd edn. Blackwell, Oxford
Bennett JN, Andrew B, Prescott CE (2002) Vertical fine root dis-tribution of western redcedar, western hemlock, and salal inold-growth cedar-hemlock forests on Northern Vancouver Is-land. Can J For Res 32: 1208–1216. DOI 10.1139/X02–034
Bloom AJ, Chapin FS, Mooney HA (1985) Resource limitation onplants – an economic analogy. Annu Rev Ecol Syst 16:363–392.DOI 10.1146/annurev.es.16.110185.002051
Bohm W (1979) Methods of studying root systems. Ecol. Stud. 33.Springer, Berlin, Heidelberg, New York
Bolte A, Hertel D, Ammer C, Schmid I, Norr R, Kuhr M, Redde N(2003) Freilandmethoden zur Untersuchung von Baumwurzeln.Forstarchiv 74:240–262
Bolte A, Rahmann T, Kuhr M, Pogoda P, Murach D, Gadow VK(2004) Relationships between tree dimension and coarse rootbiomass in mixed stands of European beech (Fagus sylvatica L.)and Norway spruce (Picea abies [L.] Karst.). Plant Soil 264: 1-11. DOI 10.1023/B:PLSO.0000047777.23344.a3
Brandtberg P-O, Lundkvist H, Bengtsson J (2000) Changes inforest-floor chemistry caused by a birch admixture in Norwayspruce stands. For Ecol Manage 130: 253–264. DOI 10.1016/S0378-1127(99)00183-8
Burschel P, Huss J (1997) Grundriß des Waldbaus. Parey, BerlinButtner V, Leuschner C (1994) Spatial and temporal patterns of
fine root abundance in a mixed oak beech forest. For EcolManage 70: 11-21. DOI 10.1016/0378-1127(94)90071-X
Casper BB, Jackson RB (1997) Plant competition underground.Annu Rev Ecol Syst 28:545–570. DOI 10.1146/annurev.ecol-sys.28.1.545
Coomes DA, Grubb PJ (2000) Impacts of root competition inforests and woodlands: a theoretical framework and review ofexperiments. Ecol Monogr 70(2):171–207
Curt T, Prevosto B (2003a) Root biomass and rooting profile ofnaturally regenerated beech in mid-elevation Scots pine wood-lands. Plant Ecol 167:269–282. DOI 10.1023/A:1023904912712
Curt T, Prevosto B (2003b) Rooting strategy of naturally regen-erated beech in Silver birch and Scots pine woodlands. PlantSoil 255:265–279. DOI 10.1023/A:1026132021506
Duchiron M-S (2000) Strukturierte Mischwalder: eine Heraus-forderung fur den Waldbau unserer Zeit. Parey, Berlin
Ehwald E, Grunert F, Schulz W, Vetterlein E (1961) Zur Okologievon Kiefern-Buchen-Mischbestanden. Arch Forstwesen10:397–415
Ellenberg H (1996) Vegetation Mitteleuropas mit den Alpen, 5thedn. Eugen Ulmer, Stuttgart
24

Ellenberg H, Mayer R, Schauermann J (1986) Okosystemforschung– Ergebnisse der Sollingprojektes. Eugen Ulmer, Stuttgart
Firbas F (1952) Spat- und nacheiszeitliche Waldgeschichte Mittel-europas nordlich der Alpen, Bd. II: Waldgeschichte der ein-zelnen Landschaften. G. Fischer, Jena
Fitter AH (1987) An architectural approach to the comparariveecology of plant root systems. New Phytol 206(Suppl):61–77
Fitter AH (2002) Characteristics and functions of root systems. In:Waisel Y, Eshel A, Kafkafi U (eds) Plant roots—the hiddenhalf, 3rd edn. Dekker, New York, pp 15–32
George E, Marschner H (1996) Nutrient and water uptake by rootsof forest trees. J Plant Nutr Soil Sci 159:11–21
Gottsche D (1972) Verteilung von Feinwurzeln und Mykorrhizenim Bodenprofil eines Buchen- und Fichtenbestandes im Solling.Mitt. Bundesforschungsanst. Forst- u. Holzwirtschaft 88, MaxWiedebusch, Hamburg, pp 1–102
Grams TEE, Kozovits AR, Winkler JB, Sommerkorn M, BlaschkeH, Haberle K-H, Matyssek R (2002) Quantifying competitive-ness in woody plants. Plant Biol 4: 153–158. DOI 10.1055/s-2002–25729
Grime JP (1979) Plant strategies and vegetation processes. Wiley,Chichester
Hendriks CMA, Bianchi FJJA (1995) Root density and root bio-mass in pure and mixed forests of Douglas fir and beech. Neth JAgric Sci 43:321–331
Hertel D (1999) Das Feinwurzelsystem von Rein- und Mis-chbestanden der Rotbuche: Struktur, Dynamik und interspez-ifische Konkurrenz. Diss Bot 317, J Cramer, Berlin, pp 1–206
Horn A, Murach D (2003) Vertikale Feinwurzelverteilung undHinweise auf interspezifische Wurzelkonkurrenz in Eschen/Buchen-Naturverjungungen. Forstarchiv 74:46–52
Kehong Wu (2000) Fine root production and turnover and itscontribution to nutrient cycling in two beech (Fagus sylvaticaL.) forest ecosystems. Ber. Forschungsz. WaldokosystemeUniv. Gottingen (FZW) A 170, pp 1–130
Kelty MJ (1992) Comparative productivity of monocultures andmixed-species stands. In: Kelty MJ, Larson BC, Oliver CD (eds)The ecology and silviculture of mixed-species forests. Kluwer,Dordrecht, pp 125–141
Keyes MR, Grier CC (1981) Above- and belowground net pro-duction in 40-year-old Douglas fir stand on low and high pro-ductivity sites. Can J For Res 11:599–605
Kostler JN, Bruckner E, Bibelriether H (1968) Die Wurzel derWaldbaume. Parey, Hamburg
Kramer H (1988) Waldwachstumslehre. Parey, HamburgLeuschner C, Hertel D, Coners H, Buttner V (2001) Root com-
petition between beech and oak: a hypothesis. Oecologia 126:276–284. DOI 10.1007/s004420000507
Leuschner C, Hertel, D, Schmid I, Koch O, Muhs A, Holscher D(2004) Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soilfertility. Plant Soil 258: 43–56. DOI 10.1023/B:PLSO.00000-16508.20173.80
Lorenz R L (1996) Grundbegriffe der Biometrie. G. Fischer,Stuttgart
MacQueen DR (1968) The quantitative distribution of absorbingroots of Pinus sylvestris and Fagus sylvatica in a forest suc-cession. Oecol Plantarum 3: 83–99
Murach D (1984) Die Reaktion der Feinwurzeln von Fichten aufzunehmende Bodenversauerung. Gottinger Bodenkundl Ber 77,pp 1–127
Murach D (1991) Feinwurzelumsatze auf bodensauren Fichtenst-andorten. Forstarchiv 62:12–17
Nambiar EKS, Sands R (1993) Competition for water and nutri-ents in forests. Can J For Res 23:1955–1968
Newton M, Cole E (1991) Root development in planted Douglas-fir under varying competitive stress. Can J For Res 21:25–31
Nielsen CCN, Mackenthun G (1991) Die horizontale Variation derFeinwurzelintensitat in Waldboden in Abhangigkeit von derBestockungsdichte. Eine rechnerische Methode zur Bestim-mung der ‘‘Wurzelintensitatsglocke‘‘ an Einzelbaumen. AllgForst- u J Ztg 162:112–119
Oberdorfer E (1992) Suddeutsche Pflanzengesellschaften. Teil IV:Walder und Gebusche, 2nd edn. G. Fischer, Jena
Otto H-J (1986) Standortliche Voraussetzungen, Ziele und Wald-bautechnik in Fichten-Buchen-Mischbestanden des Harzes.Allg. Forst- u. J.Ztg. 157:214–222
Otto H-J (1990) Die forstliche Entwicklung und Ziele einer nat-urnahen Waldpflege. Allg. Forstzeitschr 45:723–726
Otto H-J (1991) Langfristige, okologische Waldbauplanung fur dieNiedersachsischen Landesforsten Bd. 2: Das niedersachsischeBergland und Gesamtauswertung. Aus demWalde 43, pp 1–527
Otto H-J (1992) Rahmenbedingungen und Moglichkeiten zurVerwirklichung der okologischen Waldentwicklung in denniedersachsischen Landesforsten. Forst und Holz 47:75–78
Otto H-J (1994) Waldokologie. Eugen Ulmer, StuttgartParrish JAD, Bazzaz FA (1976) Underground niche separation in
successional plants. Ecology 57:1281–1288Polomski J, Kuhn N (1998) Wurzelsysteme. Haupt, Bern Stuttgart
WienPuhe J (2003) Growth and development of the root system of
Norway spruce (Picea abies) in forest stands—a review. ForEcol Manage 175: 253–273. DOI 10.1016/S0378-1127(02)00134-2
Puhe, J (1994) Die Wurzelentwicklung der Fichte (Picea abies) beiunterschiedlichen chemischen Bodenbedingungen. Ber. For-schungsz. Waldokosysteme Univ. Gottingen (FZW) A 108, pp1–128
Rapp C (1991) Untersuchungen zum Einfluß von Kalkung undAmmoniumsulfat-Dungung auf Feinwurzeln und Ek-tomykorrhizen eines Buchenaltbestandes im Solling. Ber. For-schungsz. Waldokosysteme Univ. Gottingen (FZW) A 108, pp1–293
Rohrig E, Gussone HA (1990) Waldbau auf okologischer Grund-lage, Bd. 2: Baumartenwahl, Bestandesbegrundung und Best-andespflege. Parey, Hamburg Berlin
Rothe A (1997) Einfluss des Baumartenanteils auf Durchwurze-lung, Wasserhaushalt, Stoffhaushalt und Zuwachsleitung einesFichten-Buchen-Mischbestandes am Standort Hoglwald. For-stl. Forschungsber. Munchen 163, pp. 1–213
Rothe A, Binkley D (2001) Nutritional interactions in mixed spe-cies forests: a synthesis. Can J For Res 31:1855–1870. DOI10.1139/cjfr-31-11-1855
Rust S, Savill PS (2000) The root systems of Fraxinus excelsior andFagus sylvatica and their competitive relationships. Forestry73(5): 499–508. DOI 10.1093/forestry/73.5.499
Rysavy T, Roloff A (1994) Ursachen der Vereschung in Mis-chbestanden und Vorschlage zu ihrer Vermeidung. Forst u Holz49:392–395
Sachs L (1999) Angewandte Statistik, 9th ed. Springer, BerlinHeidelberg New York
Schmid I (2002) The influence of soil type and interspecific com-petition on the fine root system of Norway spruce and Euro-pean beech. Basic Appl Ecol 3(4): 339–355. DOI 10.1078/1439-1791-00116
Schmid I, Kazda M (2002) Root distribution of Norway spruce inmonospecific and mixed stands on different soils. For EcolManage 159: 37–47. DOI 10.1016/S0378-1127(01)00708-3
Schober R (1987) Ertragstafeln wichtiger Baumarten, 3rd edn. J.D.Sauerlander’s Verlag, Frankfurt/M
Schutz J-P (2001) Der Plenterwald und weitere Formen strukturi-erter und gemischter Walder. Parey, Berlin
Van Praag HJ, Sougnez-Remy S, Weissen F, Carletti G (1988)Root turnover in a beech and a spruce stand of the BelgianArdennes. Plant Soil 105: 87–103
Walter H, Breckle S-W (1991) Okologie der Erde Bd. 1: Okologi-sche Grundlagen in globaler Sicht. 2nd ed. Spektrum G. Fi-scher, Stuttgart
Wang Z, Gottlein A, Bartonek G (2001) Effects of growing roots ofNorway spruce (Picea abies [L.] Karst.] and European beech(Fagus sylvatica L.) on rhizosphere nutrient chemistry in aforest soil. J Plant Nutr Soil Sci 164:35–41. DOI 10.1002/1522-2624(200102)164:1<35
25

Wang Z, Gottlein A, Rodenkirchen H (2003) Soil exploration,nutrient partitioning, and nutrient efficiency of beech andspruce seedlings in a rhizotrone experiment. J Plant Nutr SoilSci 166:390–391. DOI 10.1002/jpln.200390060
Weckesser M (2003) Die Bodenvegetation von Buchen-Fichten-Mischbestanden im Solling – Struktur, Diversitat und Stoff-haushalt. Cuvillier, Gottingen
Wolff B, Erhard M, Holzhausen M, Kuhlow T (2003) Das Klima inden forstlichen Wuchsgebieten und Wuchsbezirken Deutsch-lands. Mitt Bundesforschungsanst Forst- u Holzwirtschaft 211,Max Wiedebusch, Hamburg, pp 1–29, 97 p (supplement)
26




















