
Interactions between invading benthivorous fish and native whitefish in subarctic lakes
17
Interactions between invading benthivorous fish and native whitefish in subarctic lakes BRIAN HAYDEN* ,† , TIINA HOLOPAINEN ‡ , PER-ARNE AMUNDSEN § , ANTTI P. ELORANTA ¶ , RUNE KNUDSEN § , KIM PRÆBEL §,1 AND KIMMO K. KAHILAINEN* ,† *Kilpisj€ arvi Biological Station, University of Helsinki, Kilpisj€ arvi, Finland † Department of Environmental Sciences, University of Helsinki, Helsinki, Finland ‡ Department of Biology, Faculty of Science and Forestry, University of Eastern Finland, Joensuu, Finland § Department of Arctic and Marine Biology, Faculty of Biosciences, Fisheries and Economics, University of Tromsø, Tromsø, Norway ¶ Department of Biological and Environmental Sciences, University of Jyv€ askyl€ a, Jyv€ askyl€ a, Finland SUMMARY 1. Many species are expanding their distribution towards higher latitudes and altitudes in response to climate change. These range shifts are expected to change fish community structure and alter food-web dynamics in subarctic lakes. However, the impacts of invading species on native fish and invertebrate prey communities remain understudied. 2. The trophic ecology of invasive species determines the likelihood of direct resource competition with native taxa. In Northern Europe, perch (Perca fluviatilis), a trophic generalist, and ruffe (Gymno- cephalus cernuus), a benthic specialist, are expanding their distribution ranges northwards, colonising lakes inhabited by a native generalist, whitefish (Coregonus lavaretus). We predicted that increased fish diversity and density would deplete the invertebrate community and increase resource competi- tion between native and invasive species. 3. To assess the degree of resource competition between native and invasive species, we compared (i) fish and invertebrate community structure; (ii) diet and stable carbon and nitrogen ratios of white- fish, ruffe and perch and (iii) growth, condition and relative population size of whitefish in two non- invaded lakes with two lakes containing one and two lakes containing both invasive species. Each lake was sampled on a single occasion between August and September. 4. Benthic macroinvertebrate density and community structure were unaffected by increased con- sumer diversity, while top-down control of pelagic zooplankton density and size was evident in lakes with increased fish diversity. 5. Differences in diet and stable isotope ratios were evident between all whitefish populations, although these were not directly related to the presence of invasive species. Specialised adaptations of invasive species may confer a competitive advantage in invaded lakes; ruffe dominated the pro- fundal niche, while perch displayed an ontogenetic shift to piscivory, reducing niche overlap with native whitefish. 6. Growth rate and population density of whitefish were largely independent of fish community structure and were governed by local variations in lake productivity. However, there was a sign of lowered condition of whitefish in invaded systems. Shallow and more productive lakes with higher food availability supported populations of native and invasive species. 7. Our findings indicate that trophic specialisations of invasive species play a key role in determining their impacts on the systems they invade. This study focussed on early stages of invasion, and the outcome of species interactions may change following the establishment of new species. In addition, these impacts will not be uniform across the invaded landscape as lake-specific variations in Correspondence: Brian Hayden, Department of Environmental Sciences, University of Helsinki, P.O. Box 65, FI-00014, Finland. E-mail: brian.hayden@helsinki.fi 1 Present address: Kim Præbel, Department of Biology, Centre for Ecological and Evolutionary Synthesis (CEES), University of Oslo, P.O. Box 1066, Blindern, N-0315, Oslo, Norway. 1234 © 2013 Blackwell Publishing Ltd Freshwater Biology (2013) 58, 1234–1250 doi:10.1111/fwb.12123
Transcript of Interactions between invading benthivorous fish and native whitefish in subarctic lakes

Interactions between invading benthivorous fish and nativewhitefish in subarctic lakes
BRIAN HAYDEN* ,† , TI INA HOLOPAINEN‡, PER-ARNE AMUNDSEN§, ANTTI P. ELORANTA¶, RUNE
KNUDSEN§, KIM PRÆBEL§ , 1 AND KIMMO K. KAHILAINEN*,†
*Kilpisj€arvi Biological Station, University of Helsinki, Kilpisj€arvi, Finland†Department of Environmental Sciences, University of Helsinki, Helsinki, Finland‡Department of Biology, Faculty of Science and Forestry, University of Eastern Finland, Joensuu, Finland§Department of Arctic and Marine Biology, Faculty of Biosciences, Fisheries and Economics, University of Tromsø, Tromsø, Norway¶Department of Biological and Environmental Sciences, University of Jyv€askyl€a, Jyv€askyl€a, Finland
SUMMARY
1. Many species are expanding their distribution towards higher latitudes and altitudes in response
to climate change. These range shifts are expected to change fish community structure and alter
food-web dynamics in subarctic lakes. However, the impacts of invading species on native fish and
invertebrate prey communities remain understudied.
2. The trophic ecology of invasive species determines the likelihood of direct resource competition
with native taxa. In Northern Europe, perch (Perca fluviatilis), a trophic generalist, and ruffe (Gymno-
cephalus cernuus), a benthic specialist, are expanding their distribution ranges northwards, colonising
lakes inhabited by a native generalist, whitefish (Coregonus lavaretus). We predicted that increased
fish diversity and density would deplete the invertebrate community and increase resource competi-
tion between native and invasive species.
3. To assess the degree of resource competition between native and invasive species, we compared
(i) fish and invertebrate community structure; (ii) diet and stable carbon and nitrogen ratios of white-
fish, ruffe and perch and (iii) growth, condition and relative population size of whitefish in two non-
invaded lakes with two lakes containing one and two lakes containing both invasive species. Each
lake was sampled on a single occasion between August and September.
4. Benthic macroinvertebrate density and community structure were unaffected by increased con-
sumer diversity, while top-down control of pelagic zooplankton density and size was evident in
lakes with increased fish diversity.
5. Differences in diet and stable isotope ratios were evident between all whitefish populations,
although these were not directly related to the presence of invasive species. Specialised adaptations
of invasive species may confer a competitive advantage in invaded lakes; ruffe dominated the pro-
fundal niche, while perch displayed an ontogenetic shift to piscivory, reducing niche overlap with
native whitefish.
6. Growth rate and population density of whitefish were largely independent of fish community
structure and were governed by local variations in lake productivity. However, there was a sign of
lowered condition of whitefish in invaded systems. Shallow and more productive lakes with higher
food availability supported populations of native and invasive species.
7. Our findings indicate that trophic specialisations of invasive species play a key role in determining
their impacts on the systems they invade. This study focussed on early stages of invasion, and the
outcome of species interactions may change following the establishment of new species. In addition,
these impacts will not be uniform across the invaded landscape as lake-specific variations in
Correspondence: Brian Hayden, Department of Environmental Sciences, University of Helsinki, P.O. Box 65, FI-00014, Finland.
E-mail: [email protected] Present address: Kim Præbel, Department of Biology, Centre for Ecological and Evolutionary Synthesis (CEES), University of Oslo, P.O.
Box 1066, Blindern, N-0315, Oslo, Norway.
1234 © 2013 Blackwell Publishing Ltd
Freshwater Biology (2013) 58, 1234–1250 doi:10.1111/fwb.12123

morphometry and resource availability will alter the competitive balance between native and inva-
sive species.
Keywords: climate change, competition, fish, food webs, invasive species, invertebrates, lakes, stable iso-topes
Introduction
Shifts in distribution driven by climate change have
been recorded in numerous terrestrial, marine and fresh-
water species (Mack et al., 2000; Graham & Harrod,
2009; Jeppesen et al., 2012). Such range shifts may lead
to novel species assemblages (Pereira et al., 2010; Comte
et al., 2013) and altered food-web structures (Vander
Zanden, Casselman & Rasmussen, 1999; Simon & Town-
send, 2003), which can have detrimental effects on
native species and ecosystem functioning (Parmesan,
2006). Investigations of trophic interactions between
native and invasive species at their current distribution
range boundaries can provide an insight into the likely
repercussions of range expansion.
Bioclimate envelope models (i.e. direct extrapolation
of species range with changing temperature) are helpful
to detail how species distributions may change in accor-
dance with increasing ambient temperature (Pearson &
Dawson, 2003). As fish can be classified into ‘warm’,
‘cool’ and ‘cold water’ thermal guilds, depending on
their preferred temperature range (Fry, 1971; Magnuson,
Crowder & Medvick, 1979), they make ideal candidates
for bioclimatic modelling. However, such models
exclude the interspecific ecological interactions that
determine species distributions at a local level (Hampe,
2004). Here, ecological interactions (i.e. predation and
resource competition) with native species determine not
only the degree to which invading species will succeed
in novel environments, but also the impacts of invasions
upon the receiving ecosystem (Bystr€om et al., 2007;
Vitule, Freire & Simberloff, 2009). The ecological strate-
gies of native and invasive species are important in this
regard (Marchetti, Moyle & Levine, 2004; Sax et al.,
2007). For example, an introduced specialist may be able
to dominate its preferred resource and exclude native
competitors (Bøhn & Amundsen, 2001; Bøhn, Amundsen
& Sparrow, 2008). Conversely, an invasive generalist
may be able to utilise a variety of resources in the novel
habitat to alleviate direct competition with native taxa.
In addition to competitive interactions, the carrying
capacity of the system is crucial in determining whether
additional species will be able to establish and have a
deleterious effect upon the receiving ecosystem (Lock-
wood, Hoopes & Marchetti, 2007). When the system
approaches carrying capacity, consumers will exhibit a
controlling effect on the abundance of prey items, and
consequently, instances of resource competition between
native and invasive species will increase (Bøhn &
Amundsen, 2001). In addition, climate-driven range
shifts are not limited to one species, as multiple species
will be likely to migrate into new habitats during a simi-
lar timeframe (Graham & Harrod, 2009). This poses
questions regarding the impacts, not only of increased
consumer numbers, but also novel assemblages of spe-
cies and foraging strategies which affect both resident
prey and fish communities.
Subarctic lakes provide ideal ecosystems to study these
questions as they maintain relatively simple food webs.
The dominant native fish species in subarctic Fennoscan-
dian lakes is the monomorphic large sparsely rakered
(LSR) whitefish (Coregonus lavaretus), which is an ecologi-
cal generalist utilising both pelagic and benthic prey
(Kahilainen et al., 2007; Harrod, Mallela & Kahilainen,
2010). In some large and deep lakes, whitefish have
undergone adaptive radiations into specialist pelagic and
benthic morphs, but most lakes are inhabited by mono-
morphic LSR whitefish (Østbye et al., 2006; Siwertsson
et al., 2010; Kahilainen et al., 2011). This region represents
the northern range limit for percid species; perch
(Perca fluviatilis) and ruffe (Gymnocephalus cernuus)
(Tammi et al., 2003), both of which have potential to
expand their range northwards in parallel with increas-
ing ambient water temperatures. Perch is an ecological
generalist utilising ontogenetic dietary shifts from zoo-
plankton to benthic macroinvertebrates before finally
becoming piscivorous (Persson, 1983; Amundsen et al.,
2003). Ruffe is a benthic specialist with a diet dominated
by chironomid larvae and a special preference for egg
predation (Ogle, 1998). Invasive populations of ruffe have
been recorded outside the native range of the species
(Winfield, Dodge & R€osch, 1998). As an efficient benthi-
vore, ruffe is able to dominate benthic resources and
potentially to cause detrimental trophic shifts in native
species, particularly generalists such as perch and white-
fish (Bergman & Greenberg, 1994; Adams & Maitland,
1998). Range expansion by ruffe and perch into lakes
containing resident whitefish provides an opportunity to
examine the ecological processes of early stage invasion
in fish communities (Lockwood et al., 2007).
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
Benthivorous fish invasions in subarctic lakes 1235

We investigated the trophic ecology of native white-
fish in non-invaded (NI) lakes and in lakes containing
one or both invasive percids, highlighting the effects of
increased consumer diversity on invertebrate community
structure and in turn on growth, condition, length and
population size of native whitefish. We tested three prin-
cipal hypotheses: first, that increased fish diversity and
density would alter the community structure of benthic
macroinvertebrates and zooplankton towards smaller
species and body sizes via selective predation (Brooks &
Dodson, 1965; Blumenshine, Lodge & Hodgson, 2000;
Amundsen et al., 2009); second, that perch and ruffe, as
more efficient benthivores (Ogle, 1998; Amundsen et al.,
2003), would dominate the benthic prey resources result-
ing in a dietary shift towards pelagic feeding by general-
ist whitefish; third, that whitefish would exhibit reduced
levels of somatic growth and condition in invaded sys-
tems due to resource competition with the invasive spe-
cies (Bøhn & Amundsen, 2004).
Methods
The study was conducted on six lakes in north-western
Fennoscandia (Fig. 1), comprising two replicates of (i)
whitefish-dominated NI lakes; (ii) lakes containing
whitefish and ruffe (WR) and (iii) lakes containing
whitefish, ruffe and perch (WRP). Numbers of fish spe-
cies in each lake varied from 4 to 12, of which the core-
gonids [LSR whitefish, densely rakered (DR) whitefish
and vendace Coregonus albula] and invasive percids
(ruffe and perch) were the most abundant (Table 1).
Additional fish species, most notably brown trout (Salmo
trutta), grayling (Thymallus thymallus), pike (Esox lucius)
and burbot (Lota lota), were present in some lakes
(Table 1). Sampling was conducted on one occasion
between August and September 2011, except in Lake
Palo, which was sampled in August 2009. With the
exception of Lake Palo, which is classed as mesotrophic,
all lakes were oligotrophic as is typical of this region
(Table 1).
Field sampling
Bathymetric maps were created for each lake using
hydroacoustics. Vertical light profiles were measured
so that compensation depths (depth at which
light = 1% of surface light) could be calculated (Lam-
pert & Sommer, 2007). Littoral (benthic areas situated
in waters shallower than the compensation depth),
profundal (benthic areas deeper than compensation
depth) and pelagic (open water above the profundal)
zones were subsequently identified and quantified in
each lake prior to sampling.
Benthic macroinvertebrates were collected from a tran-
sect using an Ekman grab (sampling area of 234 cm2)
following a previously established methodology (Kahilai-
nen, Lehtonen & K€on€onen, 2003). Lake Palo was sampled
using a larger Ekman grab (sampling area of 272 cm2).
Three replicates at each depth were taken along a depth
contour transect at depths of 1, 2, 3, 5, 10, 15 and 20 m
(when lake depth permitted) to represent a continuum
from littoral to profundal habitats. Samples were sieved
through a 0.5-mm mesh, and individuals were subse-
quently identified to family level and macroinvertebrate
density (individuals m�2) calculated for each depth. Blot-
ted mass (g) of each group was weighed (accuracy
0.0001 g), and biomass values (g m�2) were calculated for
littoral and profundal macroinvertebrate communities in
each lake. Mean individual mass was obtained by divid-
ing total invertebrate biomass by the number of individu-
als. Biomass data were not available for Lake Palo due to a
restricted sampling procedure in place at that time (2009).
Pelagic zooplankton were sampled using a zooplank-
ton net (25 cm diameter, 50-lm mesh size) hauled verti-
cally through the entire water column. Individuals were
identified to family level, and zooplankton density (indi-
viduals L�1) was determined for each lake. Body length
(accuracy of 0.01 mm) of a random subsample (n = 30,
when available) of individuals of each family was mea-
sured using an Olympus CK30-F200 microscope (Olym-
pus Optical Co., Postfach Hamburg, Germany). Body
length of all small benthic crustaceans (i.e. Ostracoda,
Eurycercus spp. and Megacyclops spp.) sampled was
recorded in each lake except Lake Palo.
Fish were sampled using gill nets. Selection bias associ-
ated with gill netting is well established (Carol & Garc�ıa-
Berthou, 2007); however, the methodology remains the
most effective and widely employed way of sampling fish
in lakes (CEN, 2005). To overcome the inherent bias, a
wide selection of mesh sizes were used and sampling
effort was standardised between systems. Gill net series
consisting of eight nets, each 30 9 1.8 m with knot-to-
knot mesh sizes of 12, 15, 20, 25, 30, 35, 45 and 60 mm
(Kahilainen et al., 2004), and a 30 9 1.5 m Nordic multi-
mesh net, which was comprised of 12 equidistant 2.5 m
panels with mesh sizes ranging from 5 to 55 mm (Appel-
berg, 2000). Nets were set overnight in littoral, profundal
and pelagic zones of each lake. The number of nets set
was determined by the size of the lake, but at least three
replicates were conducted in each habitat type (littoral,
profundal and pelagic). The shallow littoral nature of
Lakes Kolta and Palo precluded pelagic/profundal sam-
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
1236 B. Hayden et al.

pling. Consequently, fish density analysis was limited to
data collected from benthic (combined littoral and pro-
fundal) gill nets, which were comparable across all sys-
tems. Density and biomass catch per unit effort (CPUE)
values were calculated as number of fish and grams of
fish per gill net series per hour.
All fish captured were identified to species level in
the field and whitefish were identified to morph accord-
ing to head and gill raker morphology (Kahilainen et al.,
2011). Total length (�1 mm) and blotted mass (�0.1 g)
were recorded for each individual, and representative
subsamples of fish were frozen (�20 °C) for subsequent
69°
70°
71°
68°Latit
ude
(°N
)
16° 28°20° 24°
(a)
FinlandSweden
Norway
Russia
L. Kuohkima (NI)
L. Kolta (NI)
L. Oiko(WR)
L. Kivi(WRP)
L. Ropi(WR)
L.Palo(WRP)
(b)
20° 23°21° 22°
68°
68.5°
69°
Longitude (°E)19°
(b)
Fig. 1 Location of study lakes in the (a) north–western Fennoscandia and (b) the upper reaches of the Tornio–Muoniojoki catchment.
Table 1 Biotic and abiotic characteristics of the non-invaded (NI), whitefish and ruffe (WR), and whitefish, ruffe and perch (WRP) lakes
Parameter L. Kolta L. Kuohkima L. Oiko L. Ropi L. Kivi L. Palo
Lake type NI NI WR WR WRP WRP
Latitude (°N) 69°03′ 69°03′ 68°50′ 68°41′ 68°49′ 68°34′Longitude (°E) 20°30′ 20°33′ 21°13′ 21°35′ 21°15′ 23°21′Surface area (km2) 1.3 0.3 1.2 1.3 3.5 3.5
Altitude (m a.s.l.) 490 488 448 398 445 346
Max depth (m) 3 10 10 20 10 2
Mean depth (m) 1.1 2.6 3.1 6.9 2.8 0.8
Secchi depth (m) 3 4.5 2.5 2.5 3 2
Compensation depth (m) >3 8 5 5 5 >2Total P (lg L�1) – – 7* 10* 7* 21†
Total N (lg l�1) – – 215* 273* 201* 470†
Species composition a, b, c, d a, b, c, d, e, f a, b, c, d, e, g, h a, b, c, d, e, f, g,
h, i, j, k, l
a, b, c, e, g, i, l a, b, c, f, g, h,
i, l, m
LSR whitefish density 2.1 (1.1) 4.4 (1.7) 8.6 (4.3) 1.1 (1.2) 9.5 (4.6) 0.6 (0.4)
LSR whitefish biomass 322 (282) 677 (152) 280 (101) 91 (99) 316 (154) 372 (224)
Ruffe biomass – – 23 (6) 8 (3) 34 (32) 50 (44)
Perch biomass – – – – 3089 (4985) 43 (37)
Pike biomass 82 (89) 74 (48) 93 (100) 64 (112) 135 (177) 67 (65)
Vendace biomass – – – – – 42 (28)
DR whitefish biomass – – – 198 (134) – –Other species biomass 1 (1) 15 (18) 10 (15) 84 (65) 54 (127) 1 (1)
Species composition is designated by letters; a, pike; b, large sparsely rakered (LSR) whitefish; c, burbot; d, minnow (Phoxinus phoxinus); e,
grayling; f, Alpine bullhead (Cottus poecilopus); g, ruffe; h, roach (Rutilus rutilus); i, perch; j, ide (Leuciscus idus); k, densely rakered (DR)
whitefish; l, brown trout; m, vendace. Mean (�SD) density (n net series�1 h�1) and biomass (g net series�1 h�1) of key fish species recorded
during the study are presented.
*Water quality data courtesy of S. Taipale.
†Data from Lapland Centre for Economic Development, Transport and Environment.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
Benthivorous fish invasions in subarctic lakes 1237

analysis of diet, growth and carbon (d13C) and nitrogen
(d15N) stable isotope ratios.
Laboratory analysis
Piscivory by perch is common in the study area (Amund-
sen et al., 2003). However, the focus of this study was
ecological interactions among whitefish, ruffe and perch,
which may use similar invertebrate prey resources. As
piscivory is rare in whitefish and ruffe, perch whose
stomachs contained fish were considered piscivorous
and analysed independently of the rest of the sample.
Individual stomach fullness was visually estimated
using the points method (Hynes, 1950) on a scale of
0–10 (0 = empty, 10 = extended full stomach). Relative
volumetric proportions of different prey categories were
estimated. Stomach contents were divided into eight
categories: chironomid larvae, trichoptera larvae, small
benthic crustaceans, large benthic crustaceans (Asellus
aquaticus and Gammarus lacustris), Mollusca (Valvata sp.,
Lymnaea sp. and Pisidium sp.), pelagic zooplankton
(Bosmina sp., Daphnia sp., Calanoida, Cyclopoida), sur-
face insects (insect pupae & adults) and other rare prey
items (macrophytes, Hydracarina, Polyphemus pediculus,
Hirudinea, biofilm and decomposed material). The rela-
tive volume of each category was calculated for each fish
population. Body size of zooplankton and diameter of
molluscs (10 random individuals per fish, when avail-
able) were subsequently measured to identify variations
in prey size selectivity between native and invasive fish
species (Kahilainen et al., 2007).
A small piece of muscle tissue was excised from the dor-
sal flank of a subsample of each species for stable isotope
analysis. Sample sizes of whitefish, ruffe and perch varied
between 44 and 75 individuals per species. In addition, all
pike captured (n = 15–28 per lake) were analysed. All iso-
tope samples were freeze-dried, ground to a fine powder
and weighed (0.5–0.6 mg) in aluminium foil cups. d13Cand d15N values were recorded using FlashEA 1112 ele-
mental analyser coupled to Thermo Finnigan DELTAPLUS
Advantage mass spectrometer (for details see Fry & Sherr,
1984). Fish d13C values were arithmetically lipid norma-
lised to remove the influence of variable lipid concentra-
tions prior to data analysis (Kiljunen et al., 2006). d13C and
d15N values were obtained for each macroinvertebrate
family recorded at each depth along the transect. In
instances where too few individuals were obtained to
provide an effective sample size (0.5–0.6 mg), samples
from neighbouring depths were pooled. Mean littoral and
profundal values, based on the light compensation point
in each lake, were subsequently calculated from the
macroinvertebrate family values at the relevant depths.
Zooplankton samples from multiple vertical tows were
pooled to obtain sufficient sample weight (0.5–0.6 mg).
Data analysis
Growth rates of whitefish were calculated from length-
at-age data determined from burnt and cracked otoliths
(Bagenal & Tesch, 1978). As the data were not normally
distributed, mean length-at-age values between systems
were compared using pairwise Welch’s t-test, an ana-
logue of the Student’s t-test without the requirements
for similarities of variance. Welch’s t-tests were per-
formed in R (R Development Core Team, 2012). Fulton
condition factor (k) of each whitefish was determined
according to Bagenal & Tesch (1978):
k ¼ M
TL3
� �� 100
where M is the blotted fish mass (g) and TL is the total
fish length (cm). Variation in condition factors between
lakes and fish communities was assessed using Welch’s
t-test.
Variation in fish and invertebrate community struc-
ture and in the stomach content of fish was examined
using PERMANOVA (PRIMER 6.1.13; PRIMER-E, Plym-
outh, U.K.), a nonparametric permutation-based ana-
logue of analysis of variance between two or more
groups based on a distance measure (Anderson, 2001;
McArdle & Anderson, 2001). In each case, a Bray–
Curtis similarity matrix (Bray & Curtis, 1957) was created
from non-transformed abundance data. Two factor
PERMANOVAS were performed on the fish, macroinverte-
brate and zooplankton community structure similarity
matrices to test the effect of ‘lake’ (six levels, random)
and ‘fish community’ (three levels, fixed), with ‘lake’
nested within ‘fish community’, on variation within the
data set. A third factor, ‘species’ (three levels, fixed), was
included in the analysis of stomach content. In invaded
lakes, levels of dietary overlap were explored using pair-
wise PERMANOVA analysis. When significant variation
(P < 0.05) was observed between groups, percentage sim-
ilarity analysis (SIMPER) was used to determine which
prey items contributed most to the difference (Clarke,
1993). Comparisons of mean length of zooplankton and
small benthic crustaceans, and mean mass of macroinver-
tebrates between fish communities were performed using
Welch’s t-test.
The dietary niche of each species was calculated using a
standardised Levins’ index (Levins, 1968). As the value of
Levins’ index increases with sample size, niche width was
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
1238 B. Hayden et al.

calculated based on the diet of a randomly selected sub-
sample of individuals (n = 30). Ontogenetic shift in perch
diet was examined using a Spearman’s rank correlation of
the proportion of fish in the stomach against total length
of perch. In addition, stomach content of the top con-
sumer, pike (n = 15–28 per lake), was examined to test the
degree to which they fed on native and invasive species.
Isotope values of consumers are largely defined by the
isotopic values of their prey, which may vary both spa-
tially and temporally (Syv€aranta, H€am€al€ainen & Jones,
2006). As such, between-lake analyses may be biased by
underlying variation and were not conducted in this
case. Levels of overlap between species stable isotope
ratios were determined using pairwise PERMANOVA analy-
sis of d13C-d15N centroids, based on a Euclidean distance
matrix created from the d13C and d15N values of white-
fish, ruffe and perch in each lake. Resource use of fish
was subsequently determined using stable isotope mix-
ing models (Boecklen et al., 2011). The relative impor-
tance of littoral, pelagic and profundal sources to each
species was calculated using the Bayesian mixing model
‘Stable Isotope Analysis in R’ (SIAR; Parnell et al., 2010).
Standard trophic fractionation values for muscle tissue
(D13C = 1.3 � 0.3, D15N = 2.9 � 0.3) were used in all
cases (McCutchan et al., 2003). Comparison of resource
use between species and systems was conducted by com-
paring the 95% credibility limits of each prey source, if
credibility limits did not overlap species were deemed to
be utilising the resource at significantly different levels.
The isotopic niche breadth of each species was calcu-
lated based on the standard ellipse of d13C-d15N space
(Jackson et al., 2011). To overcome the disparity in sam-
ple sizes, the area of a small-sample-size-corrected
ellipse (SEAc), calculated using ‘Stable Isotope Bayesian
Ellipses in R’ (SIBER; Jackson et al., 2011), was used to
determine isotopic niche size. Variation in niche size
between species was subsequently calculated using the
likelihood test in SIBER (Jackson et al., 2011). SIAR and
SIBER analyses were performed using the ‘SIAR pack-
age, version 4.1.3’, in R (R Development Core Team,
2012). Ontogenetic shifts in isotopic position of fish were
tested using a Spearman’s rank correlation of d13C and
d15N against total length of fish.
Results
Invertebrate community
Benthic macroinvertebrate abundance (n m�2) varied
between lakes (pseudo F3,45 = 5.8, P < 0.01), but variation
was not associated with fish community structure Tab
le2
Mean(�
SD)den
sity
ofben
thic
invertebrates(n
m�2)an
dpelag
iczo
oplankton(n
L�1)observed
innon-invad
ed(N
I),whitefi
shan
druffe(W
R)an
dwhitefi
shruffean
dperch
(WRP)lakes
Community
Lak
eHab
itat
Chiro
Tric
SBC
LBC
Moll
Oligo
Insecta
Hydra
Other
Cladocera
Copep
oda
NI
L.Kolta
Littoral
2464
(116
0)11
(21)
627(402
)11
(31)
222(215
)12
65(160
0)11
(21)
83(73)
83(73)
0.22
(0.31)
2.01
(1.18)
Profundal
––
––
––
––
––
–
L.Kuohkim
aLittoral
833(483
)20
(23)
30(89)
10(20)
168(213
)59
(86)
10(20)
10(30)
5(15)
0.46
(0.24)
4.21
(1.51)
Profundal
747(105
7)–
141(323
)67
(163
)81
(139
)22
(37)
–15
(36)
––
–
WR
L.Oiko
Littoral
1554
(491)
44(102)
705(621)
–30
6(165
)11
3(97)
25(32)
44(44)
5(15)
0.13
(0.08)
1.29
(0.77)
Profundal
969(103
0)1(18)
318(477
)–
59(61)
200(156
)–
81(114
)22
(24)
––
L.Ropi
Littoral
557(279
)49
(72)
508(242
)23
7(480
)10
9(74)
666(449
)10
(30)
123(125
)25
(32)
0.10
(0.01)
0.19
(0.03)
Profundal
170(190
)–
159(174
)–
41(61)
111(75)
––
––
–
WRP
L.Kivi
Littoral
2033
(1029)
59(104)
286(261)
5(15)
405(277)
409(464)
158(71)
44(54)
39(47)
0.19
(0.2)
0.13
(0.15)
Profundal
1066
(452
)–
311(348
)–
111(88)
178(225
)7(18)
44(40)
7(18)
––
L.Palo
Littoral
1374
(650
)55
(45)
3367
(352
6)13
5(177
)27
6(354
)43
5(191
)27
0(209
)14
1(133
)86
(50)
0.59
(0.48)
0.97
(0.23)
Profundal
––
––
––
––
––
–
Abbreviationsofben
thic
invertebratesreferto
chironomid
larvae
(Chiro),trichoptera
larvae
(Tric),sm
allben
thic
crustaceans(SBC;Ostracoda,
Eurycercussp
p.,Megacyclops
spp.),large
ben
thic
crustaceans(LBC;Asellusaquaticus,Gam
maruslacustris),molluscs(M
oll;Valvata
sp.,Lym
naeasp
.,Pisidium
sp.),Oligoch
aeta
(Oligo),Insecta(Ephem
eroptera,Sialisan
dPlecop-
tera)an
dHydracarina(H
ydra).In
lakes
containingaprofundal
zone,
ben
thic
invertebratesaresu
bdivided
into
littoralan
dprofundal.Pelag
iczo
oplanktonaresu
bdivided
into
clad
o-
cera
andcopep
oda.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
Benthivorous fish invasions in subarctic lakes 1239

(pseudo F2,45 = 0.7, P = 0.8; Table 2). Overall macroinver-
tebrate density was greater in WRP lakes than in WR
lakes (t = �3.9, d.f. = 34, P < 0.01). High variation
between both NI lakes (L. Kolta contained the highest
density recorded in the study and L. Kuohkima con-
tained the lowest density; Fig. 2a) obscured any differ-
ence between these and WR (t = 1.6, d.f. = 27.2,
P = 0.11) or WRP lakes (t = �0.77, d.f. = 34.7, P = 0.45).
Within-lake variation was evident as littoral macroinver-
tebrate density was higher than its profundal counter-
part in all lakes (pseudo F1,45 = 6.5, P < 0.01).
Zooplankton community structure (ind L�1) varied
between lakes (pseudo F3,17 = 6.2, P < 0.01; Table 2), but
this was unrelated to fish community structure (pseudo
F2,17 = 1.6, P = 0.24). Mean density of pelagic zooplank-
ton (Fig. 2a) was higher in NI lakes than in with WR
lakes (t = 3.1, d.f. = 6.8, P = 0.02) or WRP lakes (t = 3,
d.f. = 6.5, P = 0.02), although no variation was evident
between invaded lakes (t = �0.2, d.f. = 9.9, P = 0.85).
Mean zooplankton size decreased relative to fish diver-
sity (Fig. 2b). Zooplankton were smaller in WR lakes
than in NI lakes (t = 10.7, d.f. = 525.3, P < 0.01). Simi-
larly, zooplankton in WRP were smaller than in WR
lakes (t = 8.6, d.f. = 450.1, P < 0.01). Measurements were
not available for small benthic crustaceans in Lake Palo
but the populations in Lake Kivi contained smaller indi-
viduals than those in NI (t = 2.6, d.f. = 0.01, P < 0.01)
and WR lakes (t = 3.4, d.f. = 164.5, P < 0.01). No
variation in mean size was evident between small ben-
thic crustaceans in NI and WR lakes (t = 3.4, d.f. =
149.6, P = 0.28; Fig. 2b). Mean macroinvertebrate mass
did not vary between fish communities (P > 0.05 in all
cases; Fig. 2b).
Fish community
Two perch were recorded from Lake Ropi, indicating that
the species may be beginning to establish in this lake;
however, this was not deemed sufficient to reclassify the
lake. Significant variation in fish community structure
(CPUE) was evident between lake types (pseudo
F3,37 = 2.7, P < 0.04), principally due to variation between
NI and WRP lakes (Table 1; t = 2.3, d.f. = 2, P = 0.02).
Proportional composition of whitefish decreased from c.
90% in NI lakes (mean � SD: L. Kolta: 88 � 9%; L. Ku-
ohkima: 89 � 8%) to a lower proportion in WR lakes (L.
Oiko: 74 � 14%; L. Ropi: 16 � 20%) and WRP lakes (L.
Kivi: 42 � 15%; L. Palo: 10 � 7%). Whitefish density var-
ied between lakes but this variation was not associated
with fish community structure (Fig. 3a). Whitefish
2000
4000
6000
8000 BMI
0
0.025
0.05
0.075
0.01
Mea
n in
divi
dual
mas
s (g
)L. Kolta L. Kuohkima L. Oiko L. Ropi L. Kivi L. Palo L. Kolta L. Kuohkima L. Oiko L. Ropi L. Kivi L. Palo
2000
4000
6000
8000 SBC SBC
Leng
th (m
m)
2
4
6
8
Den
sity
(n l–
1 )D
ensi
ty (n
m–2
)D
ensi
ty (n
m–2
)10 ZPL ZPL
0
0.5
1
1.5
2
Leng
th (m
m)
BMI
LittoralProfundal
0
0.5
1
1.5
2
(a) (b)
Fig. 2 (a) Density and (b) mean size of zooplankton (ZPL; Bosmina sp., Daphnia sp., Calanoida, Cyclopoida), small benthic crustaceans (SBC;
Ostracoda, Eurycercus spp., Megacyclops spp.) and benthic macroinvertebrates (BMI) in non-invaded (L. Kolta, L. Kuohkima), whitefish and
ruffe (L. Oiko, L. Ropi), and whitefish, ruffe and perch (L. Kivi, L. Palo) lakes. Error bars indicate 95% confidence limits.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
1240 B. Hayden et al.

biomass (g net series�1 h�1) was indistinguishable
between NI and WRP lakes (t = 1.2, d.f. = 12.3, P = 0.27),
but reduced biomass was observed in WR lakes relative
to both NI (t = 2.8, d.f. = 10.7, P = 0.01) and WRP
(t = �2.6, d.f. = 26.9, P = 0.01) lakes (Table 1; Fig. 3b).
Mean total length of whitefish also varied between fish
communities; whitefish in NI lakes (mean � SD:
25.3 � 7.4 cm) were larger (t = 10.7, d.f. = 362.2,
P < 0.01) than those in WR (14.8 � 4.7 cm) lakes and WRP
(17.3 � 7.8 cm) lakes (t = 10.5, d.f. = 287.9, P < 0.01). Mean
size of whitefish in WRP lakes was larger than in WR
lakes (t = �6.7, d.f. = 675.5, P < 0.01).
A decrease in mean condition factor (k) of whitefish was
observed between NI and WR lakes (Fig. 3c; t = 3.2,
d.f. = 239.8, P < 0.01). Condition of whitefish was lower in
WRP lakes than inWR lakes (t = �8.8, d.f. = 187, P < 0.01).
However, significant variation was evident between the
two study lakes in the latter category (t = �23.5,
d.f. = 129.6, P < 0.01); whitefish in Lake Kivi exhibited the
lowest condition (mean � SD: 0.7 � 0.1), while the highest
condition factor was observed in Lake Palo (1.2 � 0.15).
The highest growth rates of whitefish were observed
in Lakes Palo and Kolta (Table S2; Fig. 3d). Growth of
whitefish was unrelated to fish community structure.
Significant variation was observed between pairwise
length-at-age values of whitefish in the replicate lakes in
NI (t = 3.5, d.f. = 5, P = 0.02), WR (t = �2.9, d.f. = 7,
P = 0.02) and WRP (t = �7.7, d.f. = 6, P < 0.01) systems.
L. Kolta L. Kuoh L. Oiko L. Ropi L. Kivi L. Palo0.5
0.75
1
1.5
1.25
Lake
Con
ditio
n fa
ctor
(k)
(b)
Age1 2 3 4 5 6 7 8
Tota
l len
gth
(mm
)
0
100
200
300
400
500
(c)
L. Kolta L. Kuoh L. Oiko L. Ropi L. Kivi L. Palo
10
20
30
0
Den
sity
(n n
et s
erie
s–1
h–1 )
(a)
L. Kolta L. Kuoh L. Oiko L. Ropi L. Kivi L. Palo
(d)
Bio
mas
s (1
000
g ne
t ser
ies–
1 h
–1)
0
1
2
3
4
Lake Lake
WhitefishRuffePerchOther
L. KoltaL. KuohL. OikoL. RopiL. KiviL. Palo
Fig. 3 Characteristics of fish populations in non-invaded (L. Kolta & L. Kuohkima), whitefish and ruffe (L. Oiko & L. Ropi) and whitefish,
ruffe and perch (L. Kivi & L. Palo) lakes. (a) Mean density (catch per unit effort) and (b) biomass of large sparsely rakered whitefish, ruffe,
perch and combined other species. (c) Boxplots of condition factor (k) of whitefish. Median value is denoted by a horizontal line, box out-
lines upper and lower quartiles, and outliers are represented as circles. (d) Mean total length-at-age of whitefish. L. Kuohkima is abbreviated
to L. Kuoh for illustrative purposes.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
Benthivorous fish invasions in subarctic lakes 1241

Stomach contents
Whitefish and percids utilised distinct prey resources
across all systems (pseudo F1,2 = 6.2, P < 0.01). In each
lake, the diet of ruffe was distinguished from that of
whitefish and perch by a higher proportion of chirono-
mid larvae and large crustaceans (Table 3; Fig. 4). Varia-
tion between whitefish and perch was largely due to an
increased abundance of small benthic crustaceans in
whitefish diet relative to perch.
The diet of whitefish did not differ between fish com-
munities (pseudo F2,486 = 0.99, P = 0.53), but variation in
diet between lakes within each community type was
observed (pseudo F3,486 = 15.9, P < 0.01). Similarly, the
diet of ruffe was unaffected by the presence of perch
(pseudo F1,420 = 2.1, P = 0.19), although lake-specific vari-
ations were evident (pseudo F2,420 = 24.8, P < 0.01).
Mean prey size of whitefish (1.95 mm) was larger than
that of ruffe (1.74 mm), at a close to significant level
(t = 2.04, d.f. = 13, P = 0.06). Prey size did not vary sig-
nificantly between whitefish and perch (t = 0.84, d.f. = 3,
P = 0.46) or between perch and ruffe (t = 0.17, d.f. = 4,
P = 0.87). No variation in niche width was evident
between whitefish and ruffe (t = �0.65, d.f. = 3,
P = 0.56). Where present, perch utilised a broader niche
than whitefish (t = �28, d.f. = 1, P = 0.02) and had a sim-
ilar niche width to ruffe (t = �1.91, d.f. = 1, P = 0.31).
An ontogenetic shift to piscivory was evident in the
diet of perch. The proportion of fish in the stomach was
correlated with total length in Lake Kivi (rs = 0.53,
n = 99, P < 0.01) and Lake Palo (rs = 0.65, n = 40,
P < 0.01). The minimum size of piscivorous perch (i.e.
perch with fish prey in their stomach) was 162 mm in
Lake Kivi and 150 mm in Lake Palo. In perch over
150 mm, fish prey comprised 70 � 44% of stomach con-
tent in Lake Kivi and 77 � 42% in Lake Palo. Whitefish
was the most common prey (43 � 48%) in the stomach
content of piscivorous perch, considerably outnumbering
ruffe (6 � 23%). Whitefish was the most common prey
source of pike in Lakes Oiko (61 � 49%; n pike = 27),
Ropi (54 � 55%; n = 22), Kivi (45 � 49%; n = 18) and
Palo (48 � 50%; n = 14). Ruffe were recorded in pike
stomachs in Lakes Oiko (6 � 17%) and Kivi (12 � 35%),
while perch were not found in any pike stomachs. No
fish prey was found from ruffe or whitefish stomachs.
Stable isotope analysis
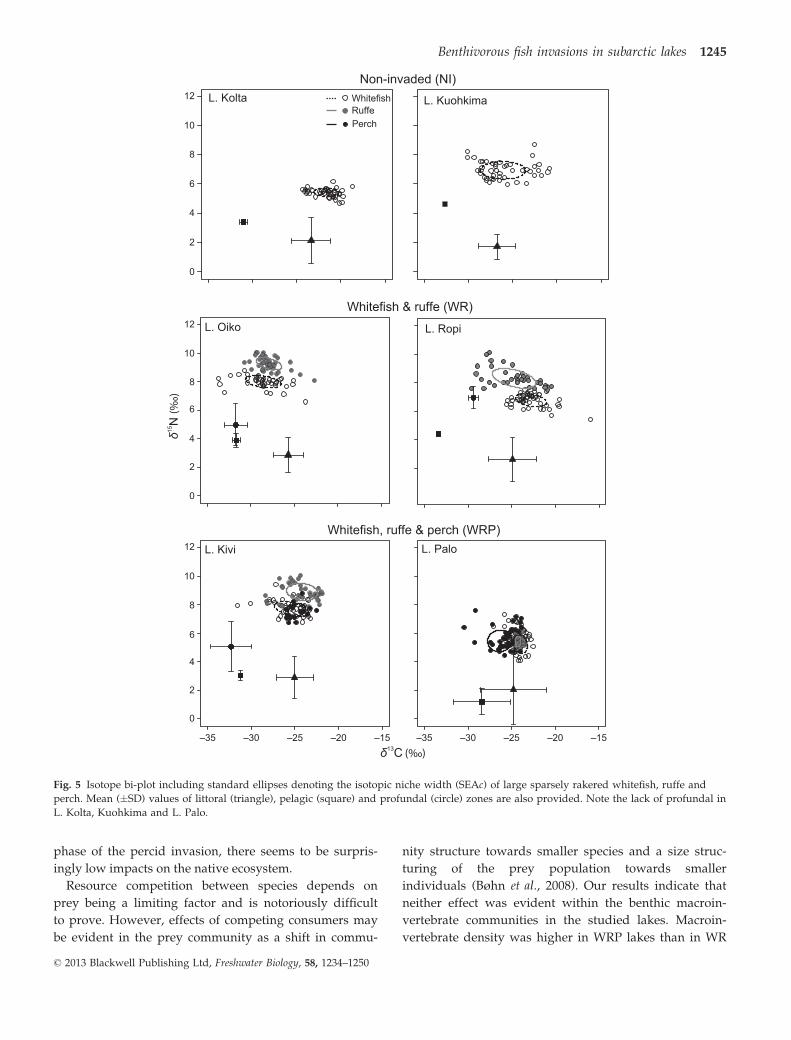
Pairwise PERMANOVA revealed trophic segregation
between species in most cases (Table 4; Fig. 5). In both
WR lakes, ruffe displayed a higher d15N value than
Tab
le3
Mean(�
SD)relativeproportion(%
)ofpreygroupsin
thestomachcontents
ofwhitefi
sh(W
),ruffe(R)an
dperch
(P)
Community
Lak
eSpecies
nChiro
Tric
SBC
LBC
Moll
Insecta
Other
ZPL
T.insect
Fullness
Lev
ins
PERMANOVA
NI
L.Kolta
Whitefi
sh80
6(10)
7(21)
16(26)
<1(1)
4(15)
4(15)
1(15)
51(44)
10(23)
4.3
0.45
L.Kuohkim
aW
hitefi
sh11
99(19)
5(19)
42(40)
–16
(25)
2(12)
–11
(28)
15(29)
3.7
0.48
WR
L.Oiko
Whitefi
sh15
03(9)
–52
(39)
<1(<1)
25(34)
<1(<1)
<1(<1)
16(32)
3(13)
3.7
0.27
W*R
P<0.01
Ruffe
171
11(15)
17(23)
57(31)
6(17)
3(8)
2(7)
<1(<1)
4(16)
–4.5
0.24
L.Ropi
Whitefi
sh77
4(14)
6(21)
35(39)
5(15)
30(35)
<1(<1)
2(1)
11(30)
7(23)
2.4
0.43
W*R
P<0.01
Ruffe
7415
(27)
2(10)
50(37)
23(33)
6(13)
–2(12)
2(7)
–3.9
0.31
WRP
L.Kivi
Whitefi
sh57
3(8)
4(16)
50(42)
–17
(29)
2(8)
4(16)
19(37)
1(6)
2.8
0.26
W*R
P<0.01
Ruffe
9121
(24)
8(18)
15(21)
32(38)
5(12)
1(11)
1(1)
17(19)
–4.3
0.39
W*P
P<0.01
Perch
150
–40
(45)
9(27)
4(17)
11(28)
9(25)
16(37)
11(29)
–2.7
0.55
P*R
P<0.01
L.Palo
Whitefi
sh75
<1(2)
<1(2)
76(22)
3(11)
19(18)
<1(1)
–1(6)
–6.3
0.08
W*R
P<0.01
Ruffe
155
18(18)
4(1)
36(35)
37(29)
<1(1)
3(7)
–2(11)
–5.7
0.30
W*P
P<0.01
Perch
471(6)
10(27)
41(37)
––
6(23)
20(32)
21(33)
<1(2)
3.7
0.35
P*R
P<0.01
Abbreviationsreferto
chironomid
larvae
(Chiro),trichoptera
larvae
(Tric),sm
allben
thic
crustaceans(SBC),largeben
thic
crustaceans(LBC)an
dmolluscs(M
oll),zo
oplankton(ZPL)
andterrestrialinsects(T.insect).Other
includes
macrophytes,
Hydracarina,
Polyphemuspediculus,Hirudinea,biofilm
anddecomposedmaterial.Estim
ationofstomachfullness(1–10),
stan
dardised
Lev
insindex
ofnichewidth
and
PERMANOVAcomparisonsofdietary
overlapareincluded
,instan
cesofsignificantvariationin
dietarehighlightedin
bold.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
1242 B. Hayden et al.

whitefish (Table 4). This segregation in d15N was also
evident in Lake Kivi, whereas in Lake Palo, the differ-
ence was marginally significant (Table 4). Ruffe also dis-
played elevated d15N relative to perch in Lake Kivi, but
not in Lake Palo (Fig. 5). Variation observed in Lake
Palo was primarily due to differences in d13C (Table 4).
The isotopic values of perch and whitefish were statisti-
cally indistinguishable in Lake Kivi, while in Lake Palo
perch were depleted in d13C relative to whitefish
(Table 4). SIBER ellipses revealed increased levels of
overlap between the isotopic niches of native and inva-
sive species in lakes with increased species diversity,
especially in Lake Kivi (Fig. 5). Isotopic niche size varied
considerably between species, although no species
employed a characteristically smaller or larger niche
than any other across all lakes (Table 4). Stable isotope
values revealed pike to be the apex predator in all sys-
tems; mean d13C values of pike were intermediate
between perch, ruffe and whitefish, while d15N was ele-
vated relative to all other species except in Lake Ropi,
where ruffe had the highest d15N values.
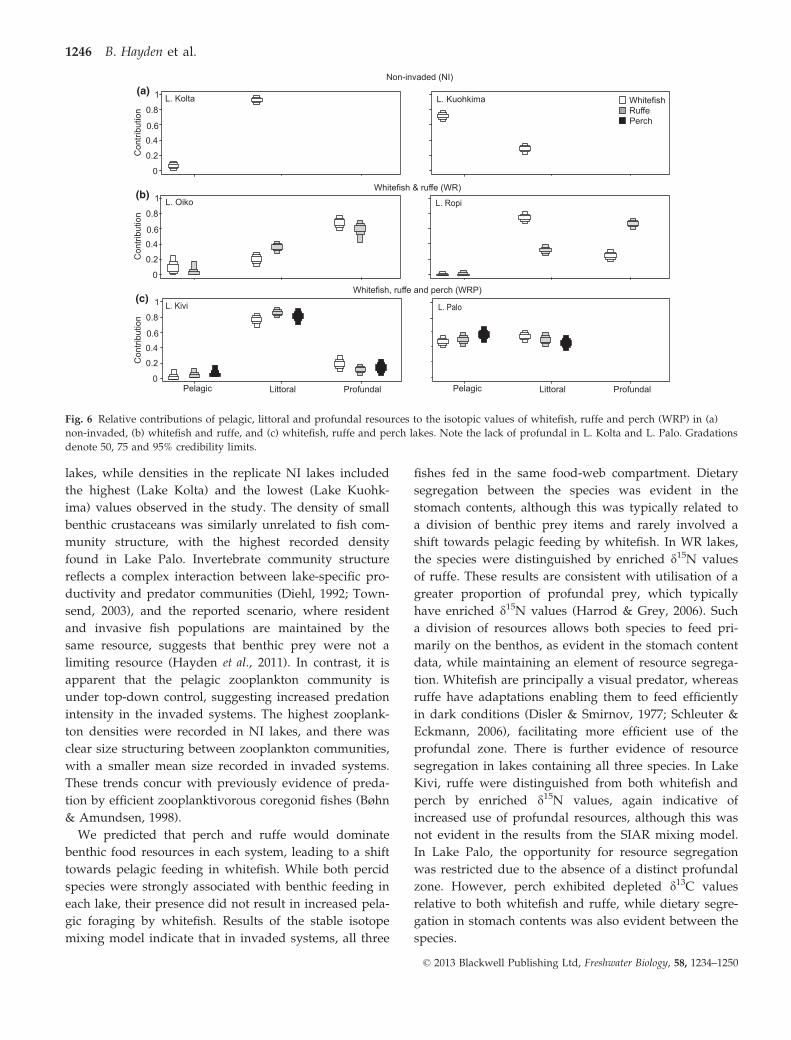
The isotope mixing model identified overlapping
resource use between species in most invaded lakes
(Table 4, Fig. 6). Resource segregation was most evident
in WR lakes. In Lake Oiko, levels of littoral resource use
differed between whitefish and ruffe, however profun-
dal and pelagic resource use of either species were
within the 95% credibility limits of the other (Table 4,
Fig. 6). In Lake Ropi, whitefish predominantly utilised
littoral resources, while ruffe were more closely aligned
with the profundal (Table 4, Fig. 6). Resource segrega-
tion was less evident in WRP lakes as resource use of all
species was within the 95% credibility limits (Fig. 6).
Ontogenetic variation in isotopic position was not evi-
dent in the majority of cases (Table 4). In whitefish, d13Cvalues increased over ontogeny in Lake Kolta, while an
increase in d15N was evident in Lake Palo, both probably
associated with the diet shift from zooplankton to ben-
thic macroinvertebrates. d15N values were strongly cor-
related with total length in both perch populations,
indicating a diet shift from invertebrates to fish.
Discussion
The invasion of percids did not alter the benthic macroin-
vertebrate community structure or density directly, but
indirectly lowered zooplankton density and size. This
apparently was not due to a direct niche shift of whitefish
into the pelagic habitat. There was some evidence of low-
ered condition, mean length and population size of white-
fish in invaded lakes, suggesting that whitefish may face
increased levels of resource competition following inva-
sion. In general however, the invading percids showed
niche segregation with native whitefish. Thus, in an early
Chiro LBC SBC ZPL Other
L. Palo
0
L. Kivi
50
100
Abu
ndan
ce (%
)
L. RopiL. Oiko
L. KuohkimaL. Kolta
Chiro LBC SBC ZPL Other
0
50
100
0
50
100
0
50
100
0
50
100
0
50
100WhitefishRuffePerch
Non-invaded (NI)
Whitefish & ruffe (WR)
Whitefish, ruffe & perch (WRP)
Fig. 4 Proportional abundance of key prey groups in the stomach contents of whitefish, ruffe and perch in non-invaded, whitefish and
ruffe, and whitefish, ruffe and perch lakes. Prey groups are summarised as chironomid larvae (Chiro), large benthic crustaceans (LBC;
Asellus aquaticus, Gammarus lacustris), small benthic crustaceans (SBC; Ostracoda, Eurycercus spp. & Megacyclops spp.), zooplankton (ZPL;
Bosmina sp., Daphnia sp., Calanoida, Cyclopoida) and other prey (other; Trichoptera, Valvata sp., Lymnaea sp., Pisidium sp., Hydracarina,
Polyphemus pediculus, Hirudinea, insect larvae, pupae and adults). Error bars represent 95% confidence limits.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
Benthivorous fish invasions in subarctic lakes 1243

Table
4Mean(�
SD)stab
leisotopevalues
ofwhitefi
sh(W
),ruffe(R),perch
(P)an
dpike.
Community
Lak
eSpecies
nd1
3C‰
d15N‰
d13C
r sd1
5N
r sPERMANOVA
Littoral(%
)Pelag
ic(%
)Profundal
(%)
SEAc
Nichesize
NI
L.Kolta
Whitefi
sh50
�21.6(1.4)
5.4(0.3)
0.86
�0.12
93(87–99
)7(1–13)
–1.3
Pike
16�2
2.3(1.1)
6.9(0.9)
L.Kuohkim
aW
hitefi
sh50
�26.1(2.5)
6.9(0.6)
0.12
�0.27
29(22–36
)71
(64–78
)–
4.8
Pike
15�2
4.4(1.0)
8.0(0.7)
WR
L.Oiko
Whitefi
sh50
�28.5(2.2)
8.0(0.4)
0.3
�0.14
W*R
P<0.01
22(11–29
)9(0–26)
69(57–78
)2.1
W*R
P=0.03
Ruffe
44�2
7.9(1.5)
9.3(0.4)
0.41
0.3
37(28–44
)2(0–17)
61(43–71
)1.7
Pike
2827
.4(0.8)
9.7(0.8)
L.Ropi
Whitefi
sh50
�22.8(1.7)
6.8(0.5)
0.21
�0.16
W*R
P<0.01
70(63–76
)1(0–4)
29(23–36
)2.4
W*R
P<0.01
Ruffe
44�2
4.8(2.5)
8.3(0.7)
0.37
�0.46
35(30–40
)2(0–7)
63(57–68
)4.7
Pike
22�2
4.0(1.6)
7.9(0.8)
WRP
L.Kivi
Whitefi
sh44
�25.4(1.9)
7.7(0.5)
�0.17
0.47
W*R
P<0.01
77(67–86
)2(0–9)
21(10–30
)2.8
W*R
P=0.72
Ruffe
44�2
4.4(1.8)
8.9(0.6)
�0.08
�0.27
W*P
P=0.22
89(81–96
)2(0–8)
9(1–16)
3.4
W*P
P=0.01
Perch
50�2
5.0(0.9)
7.5(0.5)
0.44
0.93
R*P
P<0.01
86(72–95
)3(0–14)
11(1–23)
1.4
R*P
P=0.01
Pike
18�2
5.0(0.7)
9.7(0.7)
L.Palo
Whitefi
sh75
�24.4(0.8)
5.7(0.6)
�0.1
0.86
W*R
P=0.06
54(46–62
)46
(38–54
)–
1.6
W*R
P=0.01
Ruffe
60�2
4.2(0.3)
5.5(0.4)
0.1
0.39
W*P
P<0.01
50(40–59
)50
(41–59
)–
0.4
W*P
P=0.01
Perch
46�2
5.8(1.4)
5.6(0.7)
�0.1
0.7
R*P
P<0.01
45(34 –54
)55
(46–66
)–
3.2
R*P
P<0.01
Pike
15�2
5.5(1.4)
6.9(1.2)
Spearm
an’s
rankcorrelations(rs)ofisotopic
positionan
dfish
totallength,PERMANOVA
comparisonsofcentroid
isotopic
positionareprovided
forwhitefi
sh,perch
andruffe.Mean
(95%
Bay
esiancred
ibilityintervalsin
paren
theses)relativeproportionoflittoral,pelag
ican
dprofundal
preyresourceas
determined
bytheStable
IsotopeAnalysisin
Rmixingmodel
aresh
own,note
theab
sence
ofaprofundal
zonein
Lak
esKolta,
Kuohkim
aan
dPalo.A
measu
reofisotopic
nichewidth
(SEAc)
andapairw
isecomparisonofnichewidth
arealso
displayed
.Statistically
significantvalues
(P<0.05)arehighlightedin
bold.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
1244 B. Hayden et al.
phase of the percid invasion, there seems to be surpris-
ingly low impacts on the native ecosystem.
Resource competition between species depends on
prey being a limiting factor and is notoriously difficult
to prove. However, effects of competing consumers may
be evident in the prey community as a shift in commu-
nity structure towards smaller species and a size struc-
turing of the prey population towards smaller
individuals (Bøhn et al., 2008). Our results indicate that
neither effect was evident within the benthic macroin-
vertebrate communities in the studied lakes. Macroin-
vertebrate density was higher in WRP lakes than in WR
δ15N
0
2
4
6
8
10
12 L. Oiko
0
2
4
6
8
10
12
–35 –30 –25 –20 –15 –35 –30 –25 –20 –15
δ13C
0
2
4
6
8
10
12 L. Kolta L. Kuohkima
L. Ropi
L. Kivi L. Palo
WhitefishRuffePerch
Non-invaded (NI)
Whitefish & ruffe (WR)
Whitefish, ruffe & perch (WRP)
Fig. 5 Isotope bi-plot including standard ellipses denoting the isotopic niche width (SEAc) of large sparsely rakered whitefish, ruffe and
perch. Mean (�SD) values of littoral (triangle), pelagic (square) and profundal (circle) zones are also provided. Note the lack of profundal in
L. Kolta, Kuohkima and L. Palo.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
Benthivorous fish invasions in subarctic lakes 1245
lakes, while densities in the replicate NI lakes included
the highest (Lake Kolta) and the lowest (Lake Kuohk-
ima) values observed in the study. The density of small
benthic crustaceans was similarly unrelated to fish com-
munity structure, with the highest recorded density
found in Lake Palo. Invertebrate community structure
reflects a complex interaction between lake-specific pro-
ductivity and predator communities (Diehl, 1992; Town-
send, 2003), and the reported scenario, where resident
and invasive fish populations are maintained by the
same resource, suggests that benthic prey were not a
limiting resource (Hayden et al., 2011). In contrast, it is
apparent that the pelagic zooplankton community is
under top-down control, suggesting increased predation
intensity in the invaded systems. The highest zooplank-
ton densities were recorded in NI lakes, and there was
clear size structuring between zooplankton communities,
with a smaller mean size recorded in invaded systems.
These trends concur with previously evidence of preda-
tion by efficient zooplanktivorous coregonid fishes (Bøhn
& Amundsen, 1998).
We predicted that perch and ruffe would dominate
benthic food resources in each system, leading to a shift
towards pelagic feeding in whitefish. While both percid
species were strongly associated with benthic feeding in
each lake, their presence did not result in increased pela-
gic foraging by whitefish. Results of the stable isotope
mixing model indicate that in invaded systems, all three
fishes fed in the same food-web compartment. Dietary
segregation between the species was evident in the
stomach contents, although this was typically related to
a division of benthic prey items and rarely involved a
shift towards pelagic feeding by whitefish. In WR lakes,
the species were distinguished by enriched d15N values
of ruffe. These results are consistent with utilisation of a
greater proportion of profundal prey, which typically
have enriched d15N values (Harrod & Grey, 2006). Such
a division of resources allows both species to feed pri-
marily on the benthos, as evident in the stomach content
data, while maintaining an element of resource segrega-
tion. Whitefish are principally a visual predator, whereas
ruffe have adaptations enabling them to feed efficiently
in dark conditions (Disler & Smirnov, 1977; Schleuter &
Eckmann, 2006), facilitating more efficient use of the
profundal zone. There is further evidence of resource
segregation in lakes containing all three species. In Lake
Kivi, ruffe were distinguished from both whitefish and
perch by enriched d15N values, again indicative of
increased use of profundal resources, although this was
not evident in the results from the SIAR mixing model.
In Lake Palo, the opportunity for resource segregation
was restricted due to the absence of a distinct profundal
zone. However, perch exhibited depleted d13C values
relative to both whitefish and ruffe, while dietary segre-
gation in stomach contents was also evident between the
species.
L. Kivi L. Palo
L. KuohkimaL. Kolta
Pelagic Littoral Profundal Pelagic Littoral Profundal
0
0.6
0.2
0.4
0.8
1
L. RopiL. Oiko
(a)
(b)
(c)
noitubirtnoC
noitubirtnoC
noitubirtnoC
0
0.6
0.2
0.4
0.8
1
0
0.6
0.2
0.4
0.8
1
Non-invaded (NI)
Whitefish & ruffe (WR)
Whitefish, ruffe and perch (WRP)
WhitefishRuffePerch
Fig. 6 Relative contributions of pelagic, littoral and profundal resources to the isotopic values of whitefish, ruffe and perch (WRP) in (a)
non-invaded, (b) whitefish and ruffe, and (c) whitefish, ruffe and perch lakes. Note the lack of profundal in L. Kolta and L. Palo. Gradations
denote 50, 75 and 95% credibility limits.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
1246 B. Hayden et al.

Lake-specific factors influenced resource availability in
the invaded systems, potentially dictating the opportuni-
ties for resource segregation. Two of the invaded lakes
contained a pelagic specialist, the DR morph of white-
fish in Lake Ropi and vendace in Lake Palo. Both DR
whitefish and vendace are more efficient zooplankti-
vores than LSR whitefish and may exclude them from
pelagic feeding (Bøhn & Amundsen, 2001; Harrod et al.,
2010). As such, pelagic resources were not available to
LSR whitefish in these lakes and could not be used to
avoid resource competition with percids. The bathymet-
ric differences between lakes also influenced niche avail-
ability for the entire fish fauna. The lack of pelagic zone
in Lakes Kolta, Kuohkima and Palo forced all the fish
species to forage in the littoral, while the low amount of
pelagic habitat in Lakes Oiko and Kivi limited LSR
whitefish’s ability to shift to planktivory.
We predicted that effects of resource competition
would be evident in the life-history characteristics of
native whitefish in invaded lakes. However, lake-specific
factors such as bathymetry and productivity appear to
govern the effect of increased competitor diversity on
the population size, growth and condition of native
whitefish. Whitefish density was unrelated to fish com-
munity structure. While the proportion of whitefish stea-
dily declined in parallel with invader densities, invaded
lakes exhibited both the highest and the lowest whitefish
densities recorded in the study. Biomass of whitefish
also displayed evidence of site-specific variation. While
reduced biomass was evident in WR lakes, the biomass
of whitefish in WRP lakes was similar to that found in
NI systems. Whitefish growth was fastest in Lakes Kolta
and Palo, which are both shallow and contained high
benthic macroinvertebrate densities. Condition factor of
whitefish showed clear variation according to fish com-
munity structure. Increased competitor diversity corre-
sponded with a decrease in the relative condition of
whitefish, although the highest condition factor was
observed in mesotrophic Lake Palo.
The effect of this lake-specific variation is difficult to
thoroughly determine due to differences between repli-
cate lakes. Lake Palo is an outlier in some respects as it
is considerably more productive than the other lakes.
Increased nutrient load facilitates a greater density of
invertebrate prey items, probably accounting for the ele-
vated growth and condition of whitefish in the presence
of numerous competitors. In addition, Lake Palo was
sampled in 2009, two years earlier than the other lakes.
While this scenario is not ideal, previous studies of
whitefish in this region revealed a high comparability of
dietary results from different years (Kahilainen et al.,
2004; Harrod et al., 2010; Hayden, Harrod & Kahilainen,
2013). In addition, our samples were obtained from each
lake during late summer; consequently, we cannot
account for potential seasonal variation in diet and asso-
ciated dietary overlap. Lakes in this region are ice cov-
ered for six months per year, limiting primary
production and the density of zooplankton (Forsstr€om
et al., 2005). Under these conditions, the diet of whitefish
switches from a broad generalist during the open-water
season to one dominated by benthic prey in winter
(Hayden et al., 2013). This may increase the likelihood of
dietary overlap with benthivorous percids during the
winter period. Further research is currently underway to
address this issue; however, based on the data presented
here, it is evident that dietary overlap between native
and invasive species is not the case in invaded lakes
during the summer growing season.
Length-at-age data indicates that percid species have
been present in these lakes throughout the lifespan of the
current population of whitefish. However, the current sit-
uation is likely to present the early phase of invasion. The
invasion time of percids to these lakes is unknown, but
reports from fishermen in Lake Kivi suggest that percid
species are recent arrivals to this previously whitefish-
dominated system. As such, both species can be consid-
ered as still establishing in the region, and competition
for resources will probably increase as they become more
numerous (Colautti & Macisaac, 2004). The high overlap
in resource use between perch and whitefish, in conjunc-
tion with the highest density of perch observed in Lake
Kivi, indicates that increased percid density may result in
increased competition between native and invasive
fishes. Additionally, higher ruffe densities may have an
impact on native whitefish via egg predation (Winfield
et al., 1998), while increased predation on juvenile white-
fish by perch may have an impact on whitefish popula-
tion densities. Stable isotope and stomach content data of
pike indicate a greater reliance on whitefish than percids
as a prey source. Previous studies of selective predation
by pike in this region indicated strong preference for
whitefish (Amundsen et al., 2003; Kahilainen & Lehtonen,
2003), suggesting that densities of invasive percids in the
study lakes are not enough to change pike prey selection
from native whitefish, although this may change as the
invasion progresses. Ruffe is known to reach very high
population densities after establishment (Adams &
Maitland, 1998), and further investigations in the lower
reaches of the Tornio–Muoniojoki catchment, which con-
tains higher densities of both percid species (K. K. Kahi-
lainen unpublished data), would give better
understanding of the likely future scenario.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
Benthivorous fish invasions in subarctic lakes 1247

Data presented here suggest that specialist adaptations
allow moderate densities of invading species to be
subsumed into novel ecosystems before direct resource
competition with native taxa occurs. Determining the
point at which ecosystems become saturated, resulting
in resource competition deleterious to the native popula-
tions, is of key importance to understanding the ecologi-
cal impact of invasions. Future monitoring of these
populations may reveal how potentially changing inva-
der densities affect the interactions between native and
invasive taxa and any associated impacts upon the
native species (Simon & Townsend, 2003). Furthermore,
sampling of lower-latitude lakes in this catchment may
help explain how different densities of invasive species
influence the interactions with native species. Our find-
ings are particularly pertinent to the role of climate
change in determining future species range boundaries.
The carrying capacity of the lake ecosystem appears to
play a key role in regulating the impact of range expan-
sion on resident taxa. Moderate warming will probably
increase productivity levels in subarctic latitudes (Elliott,
Jones & Thackeray, 2006), and in addition, a longer
growing season and increased terrestrial vegetation will
indirectly increase the nutrient load in subarctic lakes
(Karlsson, Jonsson & Jansson, 2005). As such, a trade-off
may ensue whereby increased consumer density is med-
iated by increased resource availability. In effect, lakes
in this region represent a large biological experiment,
testing the interactions between resident and native spe-
cies in a habitat which is increasingly novel to both. This
provides a unique opportunity to examine the role
played by trophic ecology and resource competition as
the ecosystem reaches a new equilibrium.
In the systems, we assessed site-specific variations in
resource availability determine the outcomes of inva-
sions, defining the point where overlap in resource use
becomes resource competition. An assumption that
invading populations will de facto limit the resources
available to native species appears naive and should be
treated with caution. Rather the species interactions will
more likely be governed by variations in lake morphol-
ogy, productivity and community structure, leading to
an increasingly complex scenario of potential outcomes.
Acknowledgments
This manuscript was significantly enhanced by insight-
ful comments from two anonymous reviewers and the
journal editor. The study was financed by European
Regional Developmental Fund (A30205), Academy of
Finland (140903) and the Norwegian research council
(186320/V40). Authors thank C. Lien, K. Mankinen, P.
Nieminen, O. Saari and M. Sujala for the help in field
and laboratory work. Kilpisj€arvi Biological Station kindly
provided good facilities during the field work. KKK was
personally financed by Emil Aaltonen Foundation.
References
Adams C.E. & Maitland P.S. (1998) The ruffe population of
Loch Lomond, Scotland: its introduction, population
expansion, and interaction with native species. Journal of
Great Lakes Research, 24, 249–262.
Amundsen P.-A., Bøhn T., Popova O.A., Staldvik F.J., Res-
hetnikov Y.S., Kashulin N.A. et al. (2003) Ontogenetic
niche shifts and resource partitioning in a subarctic pisci-
vore fish guild. Hydrobiologia, 497, 109–119.
Amundsen P.-A., Siwertsson A., Primicerio R. & Bøhn T.
(2009) Long-term responses of zooplankton to invasion
by a planktivorous fish in a subarctic watercourse. Fresh-
water Biology, 54, 24–34.
Anderson M.J. (2001) A new method for non-parametric
multivariate analysis of variance.Austral Ecology, 26, 32–46.
Appelberg M. (2000) Swedish standard methods for sam-
pling freshwater fish with multi-mesh gillnets. Fiskeriver-
ket Information, 1, 3–31.
Bagenal T.B. & Tesch F.W. (1978) Age and growth. In: Meth-
ods for Assessment of Fish Production in Fresh Waters (Ed.
T.B. Bagenal), pp. 101–136. Blackwell Scientific Publica-
tions Ltd, London.
Bergman E. & Greenberg L.A. (1994) Competition between
a planktivore, a benthivore, and a species with ontoge-
netic diet shifts. Ecology, 75, 1233–1245.
Blumenshine S.C., Lodge D.M. & Hodgson J.R. (2000) Gra-
dient of fish predation alters body size distributions of
lake benthos. Ecology, 81, 374–386.
Boecklen W.J., Yarnes C.T., Cook B.A. & James A.C. (2011)
On the use of stable isotopes in trophic ecology. Annual
Review of Ecology, Evolution and Systematics, 42, 411–440.
Bøhn T. & Amundsen P.-A. (1998) Effects of invading vend-
ace (Coregonus albula L.) on species composition and body
size in two zooplankton communities of the Pasvik River
System, northern Norway. Journal of Plankton Research, 20,
243–256.
Bøhn T. & Amundsen P.-A. (2001) The competitive edge of
an invading specialist. Ecology, 82, 2150–2163.
Bøhn T. & Amundsen P.-A. (2004) Invasion-mediated
changes in the population biology of a dimorphic white-
fish Coregonus lavaretus population. Annales Zoologici Fen-
nici, 41, 125–136.
Bøhn T., Amundsen P.-A. & Sparrow A. (2008) Competitive
exclusion after invasion? Biological Invasions, 10, 359–368.
Bray J.R. & Curtis J.T. (1957) An ordination of the upland
forest communities of southern Wisconsin. Ecological
Monographs, 27, 326–349.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
1248 B. Hayden et al.

Brooks J.L. & Dodson S.I. (1965) Predation, body size and
composition of plankton. Science, 150, 28–35.
Bystr€om P., Karlsson J., Nilsson P.A., Van Kooten T., Ask J.
& Olofsson F. (2007) Substitution of top predators: effects
of pike invasion in a subarctic lake. Freshwater Biology, 52,
1271–1280.
Carol J. & Garc�ıa-Berthou E. (2007) Gillnet selectivity and
its relationship with body shape for eight freshwater fish
species. Journal of Applied Ichthyology, 23, 654–660.
CEN (2005) Water quality sampling of fish with multi-mesh
gillnets. EN 14757. (E.C.F. Standardization). CEN,
Brussels.
Clarke K.R. (1993) Non-parametric multivariate analysis of
changes in community structure. Australian Journal of
Ecology, 18, 117–143.
Colautti R.I. & MacIsaac H.J. (2004) A neutral terminology
to define ‘invasive’ species. Diversity and Distributions, 10,
135–141.
Comte L., Buisson L., Daufresne M. & Grenouillet G. (2013)
Climate-induced changes in the distribution of freshwater
fish: observed and predicted trends. Freshwater Biology,
DOI: 10.1111/fwb.12081.
Diehl S. (1992) Fish predation and benthic communtiy
structure – the role of omnivory and habitat complexity.
Ecology, 73, 1646–1661.
Disler N.N. & Smirnov S.A. (1977) Sensory organs of the
lateral-line canal system in two percids and their impor-
tance in behavior. Journal of the Fisheries Research Board of
Canada, 34, 1492–1503.
Elliott J.A., Jones I.D. & Thackeray S.J. (2006) Testing the
sensitivity of phytoplankton communities to changes in
water temperature and nutrient load, in a temperate lake.
Hydrobiologia, 559, 401–411.
Forsstr€om L., Sorvari S., Korhola A. & Rautio M. (2005) Sea-
sonality of phytoplankton in subarctic Lake Saanaj€arvi in
NW Finnish Lapland. Polar Biology, 28, 846–861.
Fry B. & Sherr E.B. (1984) d13C Measurements as indicators
of carbon flow in marine and fresh-water ecosystems.
Contributions in Marine Science, 27, 13–47.
Fry F.E.J.. (1971) The effect of environmental factors on
the physiology of fish. In: Fish Physiology (Ed. W.S. Hoar
& D.J. Randall), pp. 1–87. Academic Press, New York.
Graham C.T. & Harrod C. (2009) Implications of climate
change for the fishes of the British Isles. Journal of Fish
Biology, 74, 1143–1205.
Hampe A. (2004) Bioclimate envelope models: what they
detect and what they hide. Global Ecology and Biogeogra-
phy, 13, 469–471.
Harrod C. & Grey J. (2006) Isotopic variation complicates
analysis of trophic relations within the fish community of
Plußsee: a small, deep, stratifying lake. Archiv Fur Hydro-
biologie, 167, 281–299.
Harrod C., Mallela J. & Kahilainen K.K. (2010) Phenotype-
environment correlations in a putative whitefish adaptive
radiation. Journal of Animal Ecology, 79, 1057–1068.
Hayden B., Harrod C. & Kahilainen K.K. (2013) The effects
of winter ice cover on the trophic ecology of whitefish
(Coregonus lavaretus L.) in subarctic lakes. Ecology of Fresh-
water Fish, 22, 192–201.
Hayden B., Massa-Gallucci A., Caffrey J., Harrod C., Mari-
ani S., O’Grady M. et al. (2011) Trophic dynamics within
a hybrid zone – interactions between an abundant cypri-
nid hybrid and sympatric parental species. Freshwater
Biology, 56, 1723–1735.
Hynes H.B.N. (1950) The food of freshwater sticklebacks
(Gasterosteus aculeatus and Pygosteus pungitius), with a
review of methods used in studies of the food of fishes.
Journal of Animal Ecology, 19, 36–58.
Jackson A.L., Inger R., Parnell A.C. & Bearhop S. (2011)
Comparing isotopic niche widths among and within com-
munities: SIBER – Stable Isotope Bayesian Ellipses in R.
Journal of Animal Ecology, 80, 595–602.
Jeppesen E., Mehner T., Winfield I.J., Kangur K., Sarvala J.,
Gerdeaux D. et al. (2012) Impacts of climate warming on
the long-term dynamics of key fish species in 24 Euro-
pean lakes. Hydrobiologia, 694, 1–39.
Kahilainen K. & Lehtonen H. (2003) Piscivory and prey
selection of four predator species in a whitefish domi-
nated subarctic lake. Journal of Fish Biology, 63, 659–672.
Kahilainen K., Lehtonen H. & K€on€onen K. (2003) Conse-
quence of habitat segregation to growth rate of two spar-
sely rakered whitefish (Coregonus lavaretus (L.)) forms in a
subarctic lake. Ecology of Freshwater Fish, 12, 275–285.
Kahilainen K., Malinen T., Tuomaala A. & Lehtonen H.
(2004) Diel and seasonal habitat and food segregation of
three sympatric Coregonus lavaretus forms in a subarctic
lake. Journal of Fish Biology, 64, 418–434.
Kahilainen K.K., Malinen T., Tuomaala A., Alaj€arvi E., Tolo-
nen A. & Lehtonen H. (2007) Empirical evaluation of phe-
notype–environment correlation and trait utility with
allopatric and sympatric whitefish, Coregonus lavaretus
(L.), populations in subarctic lakes. Biological Journal of the
Linnean Society, 92, 561–572.
Kahilainen K.K., Siwertsson A., Gjelland K.Ø., Knudsen R.,
Bøhn T. & Amundsen P.-A. (2011) The role of gill raker
number variability in adaptive radiation of coregonid
fish. Evolutionary Ecology, 25, 573–588.
Karlsson J., Jonsson A. & Jansson M. (2005) Productivity of
high-latitude lakes: climate effect inferred from altitude
gradient. Global Change Biology, 11, 710–715.
Kiljunen M., Grey J., Sinisalo T., Harrod C., Immonen H. &
Jones R.I. (2006) A revised model for lipid-normalizing
delta C-13 values from aquatic organisms, with implica-
tions for isotope mixing models. Journal of Applied Ecology,
43, 1213–1222.
Lampert W. & Sommer U. (2007) Limnoecology: The Ecology
of Lakes and Streams. Oxford University Press, Oxford.
Levins R. (1968) Evolution in Changing Environments: Some
Theoretical Explorations. Princeton University Press, Prince-
ton, NJ.
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
Benthivorous fish invasions in subarctic lakes 1249

Lockwood J.L., Hoopes M.F. & Marchetti M.P. (2007) Inva-
sion Ecology. Plackwell Publishing, Oxford.
Mack R.N., Simberloff D., Lonsdale W.M., Evans H., Clout
M. & Bazzaz F.A. (2000) Biotic invasions: causes, epide-
miology, global consequences, and control. Ecological
Applications, 10, 689–710.
Magnuson J.J., Crowder L.B. & Medvick P.A. (1979) Tem-
perature as an ecological resource. American Zoologist, 19,
331–343.
Marchetti M.P., Moyle P.B. & Levine R. (2004) Alien fishes
in California watersheds: characteristics of successful and
failed invaders. Ecological Applications, 14, 587–596.
McArdle B.H. & Anderson M.J. (2001) Fitting multivariate
models to community data: a comment on distance-based
redundancy analysis. Ecology, 82, 290–297.
McCutchan J.H., Lewis W.M., Kendall C. & Mcgrath C.C.
(2003) Variation in trophic shift for stable isotope ratios
of carbon, nitrogen, and sulfur. Oikos, 102, 378–390.
Ogle D.H. (1998) A synopsis of the biology and life history
of ruffe. Journal of Great Lakes Research, 24, 170–185.
Østbye K., Amundsen P.-A., Bernatchez L., Klemetsen A.,
Knudsen R., Kristoffersen R. et al. (2006) Parallel evolu-
tion of ecomorphological traits in the European whitefish
Coregonus lavaretus (L.) species complex during postgla-
cial times. Molecular Ecology, 15, 3983–4001.
Parmesan C. (2006) Ecological and evolutionary responses
to recent climate change. Annual Review of Ecology, Evolu-
tion, and Systematics, 37, 637–669.
Parnell A.C., Inger R., Bearhop S. & Jackson A.L. (2010)
Source partitioning using stable isotopes: coping with too
much variation. PLoS One, 5, e9672.
Pearson R.G. & Dawson T.P. (2003) Predicting the impacts
of climate change on the distribution of species: are
bioclimate envelope models useful? Global Ecology and
Biogeography, 12, 361–371.
Pereira H.M., Leadley P.W., Proenca V., Alkemade R.,
Scharlemann J.P.W., Fernandez-Manjarres J.F. et al. (2010)
Scenarios for global biodiversity in the 21st century. Sci-
ence, 330, 1496–1501.
Persson L. (1983) Effects of intraspecific and interspecific
competition on dynamics and size structure of a perch
Perca fluviatilis and a roach Rutilus rutilus population. Oi-
kos, 41, 126–132.
R Development Core Team (2012) R: A Language and Envi-
ronment for Statistical Computing. R Foundation for Statisti-
cal Computing, Vienna, Austria.
Sax D.F., Stachowicz J.J., Brown J.H., Bruno J.F., Dawson
M.N., Gaines S.D. et al. (2007) Ecological and evolution-
ary insights from species invasions. Trends in Ecology and
Evolution, 22, 465–471.
Schleuter D. & Eckmann R. (2006) Competition between
perch (Perca fluviatilis) and ruffe (Gymnocephalus cernuus):
the advantage of turning night into day. Freshwater Biol-
ogy, 51, 287–297.
Simon K.S. & Townsend C.R. (2003) Impacts of freshwater
invaders at different levels of ecological organisation,
with emphasis on salmonids and ecosystem conse-
quences. Freshwater Biology, 48, 982–994.
Siwertsson A., Knudsen R., Kahilainen K.K., Praebel K., Pri-
micerio R. & Amundsen P.-A. (2010) Sympatric diversifi-
cation as influenced by ecological opportunity and
historical contingency in a young species lineage of
whitefish. Evolutionary Ecology Research, 12, 929–947.
Syv€aranta J., H€am€al€ainen H. & Jones R.I. (2006) Within-lake
variability in carbon and nitrogen stable isotope signa-
tures. Freshwater Biology, 51, 1090–1102.
Tammi J., Appelberg M., Beier U., Hesthagen T., Lappalainen
A. & Rask M. (2003) Fish status survey of nordic lakes:
effects of acidification, eutrophication and stocking activity
on present fish species composition. Ambio, 32, 98–105.
Townsend C.R. (2003) Individual, population, community,
and ecosystem consequences of a fish invader in New
Zealand streams. Conservation Biology, 17, 38–47.
Vander Zanden M.J., Casselman J.M. & Rasmussen J.B.
(1999) Stable isotope evidence for the food web
consequences of species invasions in lakes. Nature, 401,
464–467.
Vitule J.R.S., Freire C.A. & Simberloff D. (2009) Introduction
of non-native freshwater fish can certainly be bad. Fish
and Fisheries, 10, 98–108.
Winfield I.J., Dodge D.P. & R€osch R. (1998) Introductions of
the ruffe, Gymnocephalus cernuus, to new areas of Europe
and to North America: history, the present situation and
management implications. In: Stocking and Introduction of
Fish (Ed. I. Cowx), pp. 191–200. Fishing News Books.
Blackwell Science Ltd, Oxford.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Table S1. Mean (�SD) total length-at-age values (mm)
of whitefish, ruffe and perch.
Table S2. Mean (�SD) d13C and d15N isotope values (&)
of baseline invertebrates, including zooplankton (ZPL)
and benthic macroinvertebrates (BMI).
(Manuscript accepted 29 January 2013)
© 2013 Blackwell Publishing Ltd, Freshwater Biology, 58, 1234–1250
1250 B. Hayden et al.




















