
Hydroacoustic assessment of mono- and polymorphic Coregonus density and biomass in subarctic lakes
14
Hydroacoustic assessment of mono- and polymorphic Coregonus density and biomass in subarctic lakes Tommi Malinen 1 , Antti Tuomaala 1 , Hannu Lehtonen 1 , Kimmo K. Kahilainen 1,2 1 Department of Environmental Sciences, University of Helsinki, PO Box 65, FI-00014, Helsinki, Finland 2 Kilpisj€ arvi Biological Station, University of Helsinki, K€ asivarrentie 14622, FI-99490, Kilpisj€ arvi, Finland Accepted for publication August 5, 2013 Abstract – Density and biomass estimates of pelagic fish are essential to understand food web interactions and ecosystem functioning. We conducted surveys of six subarctic lakes for assessing both mono- and polymorphic whitefish Coregonus lavaretus (L.) populations. Monomorphic whitefish lakes were inhabited by a habitat and diet generalist, large sparsely rakered (LSR) morph, whereas polymorphic whitefish lakes had a littoral benthivorous LSR morph, a pelagic planktivorous densely rakered (DR) morph and in two cases a benthivorous small sparsely rakered (SSR) morph inhabiting the profundal zone. In addition, an introduced specialist zooplanktivore, vendace Coregonus albula (L.), inhabited one of the monomorphic lakes. Hydroacoustics was found to be an appropriate method for estimating coregonid densities and biomasses in large and deep polymorphic lakes occupied by the planktivorous DR morph or vendace, but only during dark nights in autumn. The suitability of hydroacoustic assessment for benthivorous LSR and SSR morphs was low, especially in polymorphic whitefish lakes due to their preference for near-bottom habitat or shallow areas not sampled with hydroacoustics. The pelagic density of DR morph varied from 330 to 1780 fishha 1 and biomass from 1.4 to 13.3 kgha 1 in polymorphic whitefish lakes, whereas corresponding estimates for LSR morph were 10–320 fishha 1 and 0.5–8.4 kgha 1 in monomorphic whitefish lakes. In general, polymorphism tended to increase the density and biomass of whitefish in the pelagic area compared with monomorphic systems. Key words: echosounding; habitat selection; trawling; vendace; vertical migration; whitefish morph Introduction Resource polymorphism in freshwater fish occurs widely in the northern hemisphere. Origin of resource polymorphism is related to either phenotypic plastic- ity or speciation events depending on the species position in the speciation continuum (Hendry 2009). The general pattern of fish divergence is along the littoral–pelagic resource axis, the two most frequently available habitat types in lakes. Pelagic morphs have typically a pointed head and fusiform body especially adapted to the utilisation of small zooplankton prey in open-water area, while littoral morphs have robust bodies shape and subterminal mouths adapted to uti- lise larger benthic macroinvertebrates (Schluter & McPhail 1993). Despite the considerable ecomorpho- logical research directed in polymorphic fish popula- tions, there have been few attempts to quantify densities or biomasses of sympatric morphs (Snorra- son et al. 1992a; Kahilainen et al. 2004). Whitefish Coregonus lavaretus (L.) is a highly polymorphic fish species widely distributed in the northern hemisphere. At the northern limits of its dis- tribution, in subarctic lakes below the treeline, it is often the dominant species. In most of these lakes, C. lavaretus occurs as a monomorphic population utilising both pelagic and littoral prey resources (Amundsen et al. 2004; Kahilainen et al. 2007; Harrod et al. 2010). However, in many of the north- ern Fennoscandian lakes, adaptive radiation of C. lavaretus from a single ancestor occurred rapidly after deglaciation some 10,000 years ago (Østbye Correspondence: T. Malinen, Department of Environmental Sciences, University of Helsinki, PO Box 65, FIN-00014 Helsinki, Finland. E-mail: Tommi. [email protected] doi: 10.1111/eff.12096 1 Ecology of Freshwater Fish 2013 Ó 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd ECOLOGY OF FRESHWATER FISH
Transcript of Hydroacoustic assessment of mono- and polymorphic Coregonus density and biomass in subarctic lakes

Hydroacoustic assessment of mono- andpolymorphic Coregonus density and biomass insubarctic lakesTommi Malinen1, Antti Tuomaala1, Hannu Lehtonen1, Kimmo K. Kahilainen1,21Department of Environmental Sciences, University of Helsinki, PO Box 65, FI-00014, Helsinki, Finland2Kilpisj€arvi Biological Station, University of Helsinki, K€asivarrentie 14622, FI-99490, Kilpisj€arvi, Finland
Accepted for publication August 5, 2013
Abstract – Density and biomass estimates of pelagic fish are essential to understand food web interactions andecosystem functioning. We conducted surveys of six subarctic lakes for assessing both mono- and polymorphicwhitefish Coregonus lavaretus (L.) populations. Monomorphic whitefish lakes were inhabited by a habitat and dietgeneralist, large sparsely rakered (LSR) morph, whereas polymorphic whitefish lakes had a littoral benthivorousLSR morph, a pelagic planktivorous densely rakered (DR) morph and in two cases a benthivorous small sparselyrakered (SSR) morph inhabiting the profundal zone. In addition, an introduced specialist zooplanktivore, vendaceCoregonus albula (L.), inhabited one of the monomorphic lakes. Hydroacoustics was found to be an appropriatemethod for estimating coregonid densities and biomasses in large and deep polymorphic lakes occupied by theplanktivorous DR morph or vendace, but only during dark nights in autumn. The suitability of hydroacousticassessment for benthivorous LSR and SSR morphs was low, especially in polymorphic whitefish lakes due to theirpreference for near-bottom habitat or shallow areas not sampled with hydroacoustics. The pelagic density of DRmorph varied from 330 to 1780 fish�ha�1 and biomass from 1.4 to 13.3 kg�ha�1 in polymorphic whitefish lakes,whereas corresponding estimates for LSR morph were 10–320 fish�ha�1 and 0.5–8.4 kg�ha�1 in monomorphicwhitefish lakes. In general, polymorphism tended to increase the density and biomass of whitefish in the pelagicarea compared with monomorphic systems.
Key words: echosounding; habitat selection; trawling; vendace; vertical migration; whitefish morph
Introduction
Resource polymorphism in freshwater fish occurswidely in the northern hemisphere. Origin of resourcepolymorphism is related to either phenotypic plastic-ity or speciation events depending on the speciesposition in the speciation continuum (Hendry 2009).The general pattern of fish divergence is along thelittoral–pelagic resource axis, the two most frequentlyavailable habitat types in lakes. Pelagic morphs havetypically a pointed head and fusiform body especiallyadapted to the utilisation of small zooplankton preyin open-water area, while littoral morphs have robustbodies shape and subterminal mouths adapted to uti-lise larger benthic macroinvertebrates (Schluter &McPhail 1993). Despite the considerable ecomorpho-
logical research directed in polymorphic fish popula-tions, there have been few attempts to quantifydensities or biomasses of sympatric morphs (Snorra-son et al. 1992a; Kahilainen et al. 2004).Whitefish Coregonus lavaretus (L.) is a highly
polymorphic fish species widely distributed in thenorthern hemisphere. At the northern limits of its dis-tribution, in subarctic lakes below the treeline, it isoften the dominant species. In most of these lakes,C. lavaretus occurs as a monomorphic populationutilising both pelagic and littoral prey resources(Amundsen et al. 2004; Kahilainen et al. 2007;Harrod et al. 2010). However, in many of the north-ern Fennoscandian lakes, adaptive radiation ofC. lavaretus from a single ancestor occurred rapidlyafter deglaciation some 10,000 years ago (Østbye
Correspondence: T. Malinen, Department of Environmental Sciences, University of Helsinki, PO Box 65, FIN-00014 Helsinki, Finland. E-mail: [email protected]
doi: 10.1111/eff.12096 1
Ecology of Freshwater Fish 2013 � 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd
ECOLOGY OFFRESHWATER FISH

et al. 2005, 2006). Divergence to sympatric morphshas occurred especially in large, deep and slightlymore productive lakes, in which C. lavaretus morphsmay dominate all available habitats (Harrod et al.2010; Siwertsson et al. 2010). Phenotype is corre-lated with niche usage in all the sympatric C. lavar-etus morphs: DR morph with the highest number ofgill rakers is a specialised zooplanktivore utilisingmainly pelagic habitat, the large sparsely rakered(LSR) morph having intermediate number of gill rak-ers utilises littoral benthic macroinvertebrate prey,and the small sparsely rakered (SSR) morph with thelowest gill raker counts dwells in profundal habitatand preys upon benthic macroinvertebrates (Kahilai-nen et al. 2004; Kahilainen & Østbye 2006; Harrodet al. 2010). In consequence of human introductions,vendace Coregonus albula (L.) also inhabits somenorthern Fennoscandian lakes (Amundsen et al.1999; Kahilainen et al. 2011a). This highly specia-lised pelagic zooplanktivore competes with nativeDR morph (Bøhn & Amundsen 2001; Bøhn et al.2008). Furthermore, C. albula can also hybridisewith native C. lavaretus (Kahilainen et al. 2011b). Insubarctic lakes, coregonids are generally the mainpredators of zooplankton (Kahilainen et al. 2005) andthe main prey for predatory fish; thus, they cansubstantially influence both lower and higher trophiclevels (Kahilainen & Lehtonen 2003; Gjelland 2008;Jensen et al. 2008; Kahilainen et al. 2009). There-fore, quantitative estimates for densities and biomas-ses of different coregonids are essential for modellingfood web dynamics.However, the estimation of coregonid density in
subarctic lakes is generally challenging, because thefishing pressure is relatively low, and typically, onlythe LSR morph is targeted. Hence, stock assessmentmethods based on catch statistics are practically use-less. The relative density of morphs in different habi-tats can be studied with gillnets, but combining theseindices to lake-wide estimates is difficult. In anycase, the estimation of fish density (fish�ha�1), bio-mass (kg�ha�1) or total abundance is not possibleusing only gillnet-based estimates. Hydroacoustics isprobably the most promising method for the assess-ment of coregonids in subarctic lakes excluding veryshallow ones. Coregonus albula is essentially anobligate pelagic species, of which population densi-ties have been successfully estimated with acousticmethods in many Fennoscandian lakes (Jurveliuset al. 1988; Marjom€aki & Huolila 1995; Nyberget al. 2001). Coregonus lavaretus densities have beenstudied especially in large Alpine lakes, which haveproved optimal for hydroacoustics because of thepelagic behaviour of C. lavaretus and its strongdominance in the fish assemblage (Eckmann 1991;Appenzeller 1995). In addition, in these lakes, the
horizontal distribution and vertical distribution ofC. lavaretus and other fish species are typically non-overlapping (Appenzeller 1995; Yule et al. 2013)favouring hydroacoustic estimation. In the southernFennoscandia, the presence of other abundant pelagicspecies, especially C. albula, can complicate C. lav-aretus assessments (Sandlund et al. 1985; Enderlein& Appelberg 1992; Linløkken 1995).While an estimate of pelagic coregonid density is
achievable in most cases, accurate population assess-ment using hydroacoustics is possible only when fishinhabit pelagic habitat and are absent from the acous-tic dead zone and blind zone (i.e., not close to thelake bed or lake surface, respectively). In subarcticlakes, the level of utilisation of the pelagic layer isstrongly influenced by the polar light regime(Kahilainen et al. 2004, 2009). In mid-summer, thewater column is well illuminated all the time,whereas during autumn, both day and night photope-riods occur. During daylight, predation risk inducedby the most abundant pelagic predator, brown troutSalmo trutta L. is dramatically higher than duringdarkness (Gjelland et al. 2009; Elliott 2011). Anotherspecial characteristic of subarctic lakes is large andrapid seasonal fluctuations in pelagic zooplanktonresources (Kahilainen et al. 2005), which influencesthe relative profitability of pelagic habitat. Because thevertical habitat choice of planktivorous fish is a trade-off between predation risk and feeding opportunitiesand/or temperature-related bioenergetic efficiency(Clark & Levy 1988; Scheuerell & Schindler 2003;Kahilainen et al. 2009; Mehner 2012), the pelagicoccurrence of coregonids in subarctic lakes is a subjectto large variations with season and diel period.Repeated surveys in two small lakes in the lower
part of subarctic Paatsjoki-Pasvik watercourse in thenorthern Norway suggested that hydroacoustics is anadequate method for C. albula population assessmentthroughout the open-water season (Gjelland 2008). Incontrast, the results from Lake Muddusj€arvi locatedin the same watercourse indicated that the DR morphpopulation can be assessed reliably with hydroacous-tics only during dark autumn nights (Kahilainen et al.2004). It is obvious that the habitat selection of core-gonids differs greatly among morphs (Kahilainenet al. 2004) and types of lakes (Ahrenstorff et al.2013). However, an extensive study concerning thesuitability of different coregonid populations forhydroacoustic assessment has not been undertaken.In the same vein, little is known about how diver-gence into morphs influences overall densities andbiomasses of coregonids.Our primary objective was to determine the appli-
cability of hydroacoustics in the estimation of densityand biomasses of different coregonid populations insix subarctic lakes. The suitability of two seasons
2
Malinen et al.

(mid-summer and autumn) and two diel periods (dayand night in autumn) were explored. Because a previ-ous case study (Kahilainen et al. 2009) indicated thestrongest utilisation of the pelagic habitat duringautumn nights, we predicted that this combination ofseason and photoperiod (hereafter occasion) wouldbe the best time for conducting surveys. Our secondobjective was to estimate the density and biomass ofthe different coregonid populations. We hypothesisedthat divergence into morphs increases the density andbiomass in pelagic habitat; thus, the highest overalllevels would occur in lakes having the DR morph.
Material and methods
Study lakes
Lake Kilpisj€arvi is located in north-western corner ofFinland, whereas other Lakes Vuontisj€arvi, Rahaj€arvi,Vastusj€arvi, Muddusj€arvi and Paadarj€arvi (hereaftertermed without Finnish word ‘j€arvi’) are located innorth-eastern Finnish Lapland (Fig. 1). Study lakes
are relatively deep, except L. Vastus (Table 1). All thestudy lakes are oligotrophic (tot P 4–8 lg�l�1 and totN 120–240 lg�l�1) and neutral (pH 7.0–7.2), and thewater transparency varies from very clear L. Kilpis(Secchi depth 10 m and colour 8 Pt�l�1) to slightlybrownish L. Vastus (Secchi depth 2 m and colour25 mg�Pt�l�1). The difference in water colour is lar-gely dependent on the proportion of peatland in thecatchment area. All study lakes are located in a sub-arctic forest zone, L. Kilpis in a birch Betula sp. forestand the other lakes in a pine Pinus silvestris L. forest.Coregonids are the dominant fish in all six lakes
where they exist as either monomorphic C. lavaretusin L. Kilpis and Vuontis, polymorphic C. lavaretusin L. Vastus, Muddus and Paadar, or monomorphicC. lavaretus and introduced C. albula in L. Raha(Table 2). Lakes Kilpis and Vuontis are inhabited byLSR morph having generalist diet and niche utilisa-tion (Harrod et al. 2010). Large and deep lakes, L.Muddus and L. Paadar, have trimorphic C. lavaretuspopulations consisting of pelagic zooplanktivorousDR morph, littoral benthivorous LSR morph and
Fig. 1. The map of northern Fennoscandia indicating location of study lakes. Shaded areas represent study area of hydroacoustics, solid linesindicate echosounding transects, and broken lines indicate boundaries between strata in lakes, where stratified sampling was applied.
3
Hydroacoustic assessment of mono- and polymorphic Coregonus

profundal benthivorous SSR morph (Harrod et al.2010). The small and shallow L. Vastus has only DRand LSR morphs (Kahilainen et al. 2011a). L. Rahais inhabited by the LSR morph and introduced C. al-bula both preferring a pelagic niche (Kahilainen et al.2011b). The most important pelagic piscivorous fishin all study lakes is S. trutta (Kahilainen & Lehtonen2002; Jensen et al. 2008; Kahilainen et al. 2009).Other piscivores include pike Esox lucius L., burbotLota lota (L.) and Arctic charr Salvelinus alpinus(L.). These species consume coregonids as their mainprey, but prefer benthic habitats (Kahilainen &Lehtonen 2003). The total number of fish species instudy lakes varies between 8 and 10.
Study strategy
Surveys were conducted in 2000–2005. In one analy-sis, the data set was complimented with newer data(2006 and 2010) from two lakes (Table 1). Each lakewas surveyed three times; in mid-summer, the surveywas conducted only once under the midnight sun,whereas in autumn, when a distinct day and nightprevail, sampling was carried out during bothphotoperiods. The suitability of two seasons (mid-summer and autumn) and two diel periods (day andnight in autumn) for population assessment was com-pared using hydroacoustic and bottom gillnet catch-per-unit-effort (CPUE) data. Species/morph-specificdensity and biomass estimates were computed for themost suitable study occasion using hydroacousticdata and simultaneous trawl and gillnet data on thefish assemblage. In addition, the accuracy of hydroa-coustic assessment was evaluated based on indicatorsof fish densities in the shallow areas outside thehydroacoustic sampling (gillnet CPUE), in thesurface blind zone (trawling) and in the acoustic deadzone close to the lake bed (gillnet CPUE).
Hydroacoustics
The study area was >6 m deep in L. Kilpis, Vuontis,Muddus and >5 m deep in L. Vastus, Paadar and Raha.As lakes were generally deep, the difference betweendepth contours of 5 and 6 m was negligible. The deepareas were sampled along 7–19 transects during eachsurvey (Fig. 1). However, in L. Raha, only five tran-sects recorded in summer were analysed due to techni-cal problems. Transects were equidistant within astratum, and the distance between them varied from0.1 to 1 km depending on surface area. The locationof the first transect in each lake was randomised.Hydroacoustic data were collected with a SIMRAD
EY-500 echosounder (Simrad AS, Horten, Norway)equipped with a split-beam transducer ES120-7F orES120-7C (operating frequency 120 kHz with a half-power beam angle of 7°). The transducer wasmounted on a towed body, which was lowered to adepth of 0.6 m on the left side of the boat. Pulseduration was set to 0.3 ms. The transducer was cali-brated using a standard copper sphere [target strength(TS) �40.4 dB].
Comparison of different survey occasions
The relative suitability of different survey occasionswas evaluated using an index of fish density in thepelagic mid-water layer sampled with hydroacoustics.The index used was the echo integral [mean area back-scattering coefficient, sa (m
2�ha�1)] (MacLennan et al.2002), which is linearly related to the biomass density.This index was used, because it was not possible tocompute reliable fish density estimates during alloccasions. In mid-summer and in autumn during theday, the low number of single fish targets ruled echo-counting out (MacLennan & Simmonds 1992), whilevery low trawl catches hampered the determination of
Table 1. Background data on lake morphometry and water quality of study lakes. Survey occasions are also indicated.
Kilpis Vuontis Raha Vastus Muddus Paadar
Location 69°00′N, 20°49′E 69°01′N, 27°04′E 68°45′N, 27°17′E 69°03′N, 27°07′E 69°00′N, 26°50′E 68°52′N, 26°35′EArea (km2) 37 11 23 4 48 21Altitude (m a.s.l.) 473 151 132 146 146 144Max depth (m) 57 31 46 15 73 56Mean depth (m) 19.4 6.5 14.1 2.7 8.5 11.7Secchi depth (m) 10 8 6.5 2 3 6Colour (mg�Pt�l�1)† 8 8 8 25 25 20pH† 7.1 7.2 7.2 7.0 7.2 7.1Tot P (lg�l�1)† 4 7 4 7 5 6Tot N (lg�l�1)† 120 170 100 240 160 160Area sampled by acoustics (%) 83 50 78 19 54 62Survey 1 July 2000 June 2004 July 2005 July 2005 June 2001 June 2004Survey 2 September 2002 September 2004 September 2005 September 2005 September 2000 September 2004Survey 3 September 2010 – – – September 2006 –
†Data from Lapland Centre for Economic Development, Transport and the Environment.
4
Malinen et al.

average cross-section from length distributions, andhence the use of echo integration. In the analysis, theaverage sa was computed for each lake and occasionin two layers: in the actual pelagic layer from 2 mbeneath the surface to 2 m above the bottom and in anear-bottom layer from 0.5 to 2 m above the bottom.High relative sa, especially in the actual pelagic layer,indicated high suitability of the survey occasion.
Density estimation
Species/morph-specific density estimates were com-puted only for autumn nights, because it was the only
time when the proportion of single fish targets washigh enough for reliable echo-counting. In the analy-sis, acoustic transects were used as sampling units.The fish densities of each transect were computed withEP 500 software (Lindem Data Acquisition, Norway),which uses 40 log R time-varied gain (TVG) functionfor estimating TS of single targets and 20 log R TVGfunction for summing up the echo integral from multi-ple targets (i.e., from fish shoals). The programcomputes fish density assuming the TS distributionsof fish in shoals, and fish detected as single targetswere identical. Based on TS distributions and species-specific length distributions from trawl catches, a TSthreshold was set to �60 to �53 dB depending on thelake. Smaller targets were considered as ninespinedsticklebacks Pungitius pungitius (L.) and were thusexcluded from density calculations. An integrationthreshold was set to �65 dB based on thresholdingwith different values (Eckmann 1998). The speciescomposition was estimated from the trawl catchesexcept in L. Kilpis, where gillnet data were usedbecause no trawling was conducted. In addition,approximate swept area density estimates (catch innumber divided by trawled area) were computed forthe surface blind zone based on surface trawl hauls.The species/morph-specific density estimates were
computed with poststratified sampling (Cochran1977), lake basins as strata in multibasin lakes (L.Kilpis, Vuontis, Raha, Muddus), whereas in one-basin lakes (L. Vastus and Paadar), a simple randomsampling was applied (Fig. 1). The variance of meanfish density in a stratum h VarðyhÞ was computedusing the equation (Shotton & Bazigos 1984):
Varð�yhÞ ¼Pni¼1
ðyi � �yÞ2 � lih i
Pni¼1
li � ðn� 1Þ; where
yi = fish density in ith transect�y = mean fish densityli = length of ith transectn = number of transectsThe variance of the mean density in the whole
study area, Varð�yÞ, was computed using the equation(Cochran 1977):
Varð�yÞ ¼XLh¼1
Ah
A
� �2
�Varð�yhÞ" #
; where
Ah = area of hth stratumA = study areaL = number of strataThe approximate 95% confidence limits for the
fish density estimate were calculated on the basis ofPoisson distribution (Jolly & Hampton 1990):
Table 2. Fish community information on the study lakes. The presence ofspecies/morph is denoted by ‘9’. Numbers refer to the footnoteidentification key of coregonids.
Kilpis Vuontis Raha Vastus Muddus Paadar
WhitefishCoregonus lavaretus (L.)
LSR morph1 9 9 9 9 9 9
DR morph2 9 9 9
SSR morph3 9 9
Vendace4
Coregonusalbula (L.)
9
Brown troutSalmo trutta L.
9 9 9 9 9 9
Arctic charrSalvelinusalpinus (L.)
9 9 9
GraylingThymallusthymallus (L.)
9 9 9 9 9 9
MinnowPhoxinusphoxinus (L.)
9 9 9 9 9 9
Three-spinedsticklebackGasterosteusaculeatus L.
9 9 9 9 9
Nine-spinedsticklebackPungitiuspungitius (L.)
9 9 9 9 9
PerchPerca fluviatilis L.
9 9 9 9 9
PikeEsox lucius L.
9 9 9 9 9 9
BurbotLota lota (L.)
9 9 9 9 9 9
Alpine bullheadCottus poecilopusHeckel
9
DR, densely rakered; LSR, large sparsely rakered; SSR, small sparselyrakered.120–30 short gill rakers, subterminal mouth, normal silvery whitefishcoloration.230–40 long gill rakers, slender body shape, terminal mouth, dark coloration.3<20 very short gill rakers, large head, very subterminal mouth, very largeeye, large fins.440–50 very long and fine gill rakers, up-pointing lower jaw, very pointedhead.
5
Hydroacoustic assessment of mono- and polymorphic Coregonus

The end points of confidence limits
¼ �y � 1� Varð�yÞ�y2
� �� 2 �
ffiffiffiffiffiffiffiffiffiffiffiffiffiVarð�yÞ
pSpecies/morph-specific biomass estimates were
computed by multiplying the density estimates withthe mean weight of each group in mid-water trawlcatches, excluding L. Kilpis and L. Vuontis, wheremean weights obtained from gillnet catches were used.
Fish sampling
The mid-water trawling stations and fishing depthswere selected based on the echosounding, in order togather samples in areas and layers where fish densitywas the highest. This was important, because it waspossible to conduct only 5–10 trawl hauls per studyoccasion, and the species/morph composition of thedensest aggregations has the strongest effect on den-sity estimates. The location of trawl stations in thesurface layer, however, was randomised to estimateapproximately the fish density in the blind zone ofechosounder.A small pair trawl (4–5 m high, 8 m wide and
cod-end mesh bar size 3 mm) was towed with twosmall (length 4–7 m) outboard motor boats (enginepower 25–40 hp). The trawl was lowered to thedesired depth (0–20 m) using iron weights, and thefishing depth as well as the opening of the trawlwas confirmed by echosounding. Each haul took 10–60 min, and the average towing speed was3.1 km�h�1. Benthic gillnet series combined of eight30-m-long and 1.8-m-high nets with mesh sizes of12, 15, 20, 25, 30, 35, 45 and 60 mm (knot to knot)
were used. The fishing depth of benthic gill net serieswas verified by echosounding. In L. Kilpis, where notrawling was conducted, the gill net series were usedalso in mid-water. The C. lavaretus morphs wereidentified and separated in the field according to dif-ferences in body shape, head shape and gill rakercounts (Table 2, Kahilainen & Østbye 2006). Lengthand weight of all captured fish were measured to anaccuracy of 1 mm and 0.1 g. The respective CPUEwas calculated as the number of fish captured withone gillnet series per hour.
Statistical testing
Only nonparametric statistics were applied, becauseassumptions of parametric statistics (normality andequal variances) were not met. Four differentquestions were tested with Mann–Whitney U-test(Conover 1980): differences in (1) density estimatesfor lake group A (monomorphic C. lavaretus lakes)and lake group B (polymorphic C. lavaretus lakes),(2) daytime and night-time bottom gillnet CPUEs, (3)echo integrals between two survey seasons (summervs. autumn night surveys) and (4) echo integralsbetween day and night surveys in autumn. On com-parisons 1, 2 and 4, specific hypotheses justified one-sided testing. On comparison 1, we hypothesised thatthe pelagic zooplankton resources are more activelyharvested in lakes where specialised zooplanktivoreDR morph exists (group B) resulting in higher pela-gic fish density (Kahilainen et al. 2011a). Oncomparisons 2 and 4, we hypothesised that fishascend from the bottom layer to the upper layer tofeed upon zooplankton at dusk (Kahilainen et al.
Fig. 2. The species/Coregonus lavaretus morph composition of the mid-water trawl catch at night in September. The number of trawl haulsand the total mid-water trawl catch in numbers (in parenthesis) are indicated above the columns. *In L. Kilpis, mid-water gillnet data arepresented because no trawling was conducted.
6
Malinen et al.
2004) resulting in lower CPUE in bottom gillnets andhigher echo integrals in mid-water at night. On thecomparison 3, a two-sided testing was applied. Allthese tests were carried out over all lakes (samplesize = 6). In addition, we tested the differencesbetween day and night echo integrals within eachlake with a one-sided sign test. In this analysis, oneobservation pair consisted of day and night integralsfrom one particular transect. Hence, the sample sizesvaried between lakes (Fig. 1). In all statistical tests,differences were considered significant if P < 0.05.
Results
The pelagic fish assemblage
Coregonus lavaretus was the dominant fish speciesin the pelagic habitat of all the study lakes (Fig. 2).
In lakes with polymorphic C. lavaretus (L. Vastus,Muddus, Paadar), the pelagic habitat was inhabitedmainly by DR morph. In the smallest lake (L. Vastus),both perch Perca fluviatilis and LSR morph usedpelagic habitat, whereas in the deep lakes (L. Muddusand Paadar), the dominance of the DR morph wasevident. In lakes with monomorphic C. lavaretus(L. Kilpis, Vuontis, Raha), the only morph (LSR)dominated pelagic habitat. In L. Raha, C. albula wasrelatively abundant. The catch of other species(mainly P. pungitius and S. trutta) was low (<10%)except in L. Vuontis. In this comparatively shallowand clear lake, the trawl catch was exceptionally low:58 fish of which only eight were caught from mid-water. In all lakes, very small-sized fish dominatedthe pelagic zone. The mean total length of differentcoregonid morphs/species varied from 8 to 19 cmacross the study (Table 3).
Table 3. Species/morph-specific mean total length (cm), mean weight (g) used in biomass calculations and number of fish measured.
LSR Coregonus lavaretus DR C. lavaretus C. albula Perca fluviatilis
Length Weight n Length Weight n Length Weight n Length Weight n
Kilpis 19.3 68.2 86 – – – – – – – – –Vuontis 16.9 41.3 626 – – – – – – † † †Raha 11.2 14.5 1350 – – – 14.1 24.5 555 † † †Vastus 13.3 27.2 251 8.2 4.3 455 – – – 11.3 22.0 378Muddus † † † 12.4 14.8 711 – – – † † †Paadar † † † 9.1 7.5 2190 – – – † † †
DR, densely rakered; LSR, large sparsely rakered; SSR, small sparsely rakered.Values are based on trawl catches, excluding L. Kilpis and L. Vuontis, where gillnet catches were used.–: species/morph does not exist in the lake.†Species/morph does not inhabit the pelagic habitat of the lake.
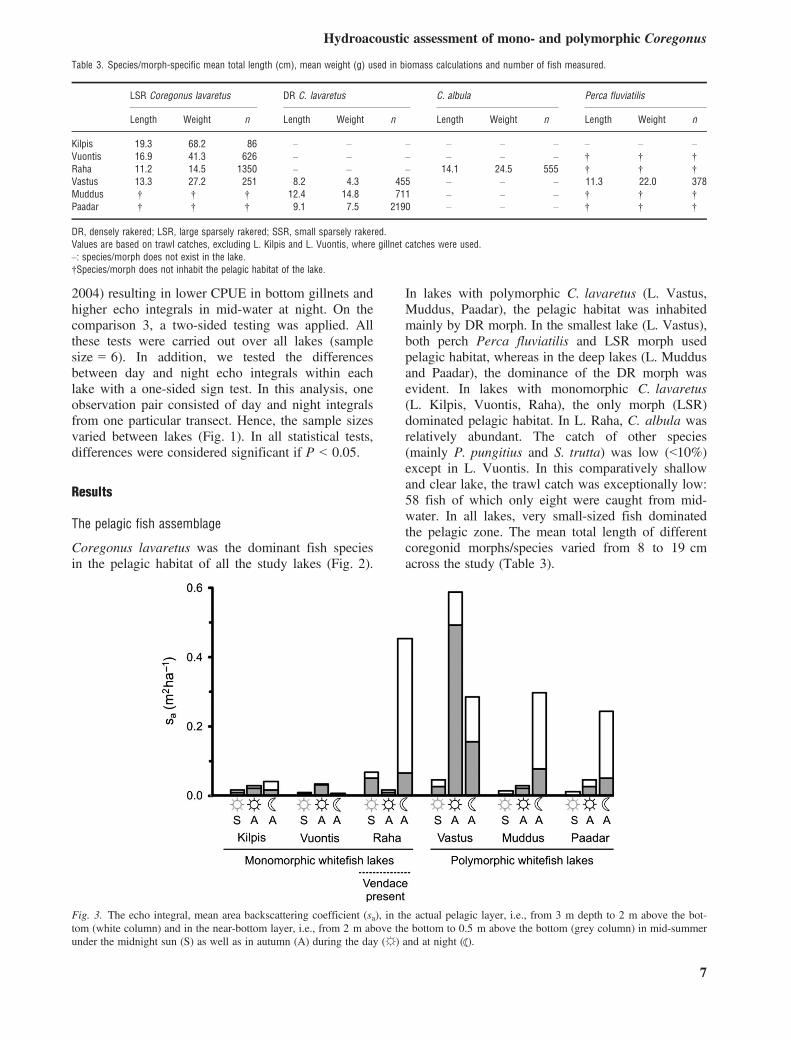
Fig. 3. The echo integral, mean area backscattering coefficient (sa), in the actual pelagic layer, i.e., from 3 m depth to 2 m above the bot-tom (white column) and in the near-bottom layer, i.e., from 2 m above the bottom to 0.5 m above the bottom (grey column) in mid-summerunder the midnight sun (S) as well as in autumn (A) during the day (☼) and at night ( ).
7
Hydroacoustic assessment of mono- and polymorphic Coregonus

The most suitable survey occasion
The autumn night-time echo integrals were signifi-cantly greater than the light summer night integrals(Mann–Whitney U-test, P < 0.05). This was true forboth the total integrals (actual pelagic layer + near-bottom layer) and the integrals for the actual pelagiclayer (Mann–Whitney U-test, P < 0.05). Theseresults showed that more fish used the pelagic layerduring autumn than mid-summer. Hence, autumn wasthe best occasion for conducting hydroacoustic sur-veys to estimate coregonid populations in our studylakes.During autumn, the night-time echo integral was
higher than the daytime integral in four of six cases
(Fig. 3). In analysis over all lakes, however, thesedifferences were not significant (Mann–WhitneyU-test, P > 0.05). In L. Vuontis, the LSR morphdid not utilise pelagic habitat at night based onextremely low trawl catch (eight individuals), whilein L. Vastus, relatively large fish shoals detected indaytime resulted in a high integral value. Accordingto pelagic trawl and bottom gillnet catches, theseshoals were likely P. fluviatilis (Figs 2 and 4a). Awithin-lake analysis, however, revealed that in allother lakes (L. Kilpis, Raha, Muddus, Paadar) thenight integral (both the total integral and the integralin the pelagic layer) was significantly higher thanthe day integral (sign test, P < 0.05) suggesting forthese lakes that autumn nights are more suitable for
(a)
(b)
Fig. 4. The catch-per-unit-effort of species/Coregonus lavaretus morphs in bottom gillnets in September, (a) within the hydroacousticallysampled deep area during the day ( ) and at night ( ), (b) in shallow areas outside the hydroacoustic sampling at night. The number ofcatching periods and total catch in numbers (in parenthesis) are indicated above the columns. Error bars indicate standard errors.
8
Malinen et al.
hydroacoustic sampling than autumn days. Thesuperiority of night over daytime for hydroacousticsampling was also indicated by profundal gillnetcatches. The CPUE was higher during daytime thanat night (Mann–Whitney U-test, P < 0.05) support-ing the hypothesis that a considerable portion ofcoregonids ascended from the bottom dead zone tothe mid-water layer at dusk (Fig. 4a). The onlyexception was the deep lake (L. Kilpis), wherenight-time CPUE was slightly higher than daytimeCPUE. In conclusion, based on echo integrals andprofundal gillnet CPUEs, the most suitable occasionfor the hydroacoustic assessment of the coregonidpopulations in our study lakes was night-time duringautumn.
The accuracy of assessment
Relatively high night-time gillnet CPUEs in all lakessuggest that the inability to sample the acoustic deadzone may be a considerable source of error even
during autumn nights (Fig. 4a). The accuracy of thehydroacoustic assessment was worsened by the factthat coregonid populations also inhabited the shallowzone outside the acoustically sampled area in five ofsix lakes (Fig. 4b). L. Raha was an exception here:no C. albula and only a few LSR C. lavaretus werecaught from shallow area. The night-time gillnetCPUEs in shallow areas outside the hydroacousticsampling were in general lower than in profundalzone except in L. Paadar (Fig. 4). However, no statis-tically significant differences were found neitherbetween-lake nor within-lake comparisons (Mann–Whitney U-test, P > 0.05). The surface blind zoneseemed to be only a minor error source for popu-lation assessment. The approximate swept areaestimates by surface trawling were <10 fish�ha�1 inall lakes except in L. Muddus (20 fish�ha�1). Onlythe DR morph and a few P. pungitius were caughtfrom the surface layer. Although a low number ofhauls (2–6 within lake) did not make it possible toevaluate the precision of the swept area estimates,
(a)
(b)
Fig. 5. The mean density (a) and biomass estimates (b) of species/Coregonus lavaretus morphs in the pelagic area of study lakes accordingto hydroacoustics and trawling (gillnet fishing in L. Kilpis) and 95% confidence intervals of the total density and biomass estimates. Thehatched borders in (b) indicate high uncertainty in biomass estimates induced by using mean weights obtained from gillnet catches.
9
Hydroacoustic assessment of mono- and polymorphic Coregonus

such low numbers strongly suggested that densitiesin the surface blind zones were negligible. In L. Kil-pis, no trawling was conducted, but low CPUE insurface gillnets (0.7 fish�h�1, four gillnet series occa-sions) suggests that the bias induced by fish residingthe surface layer was also small.
Pelagic fish density and biomass
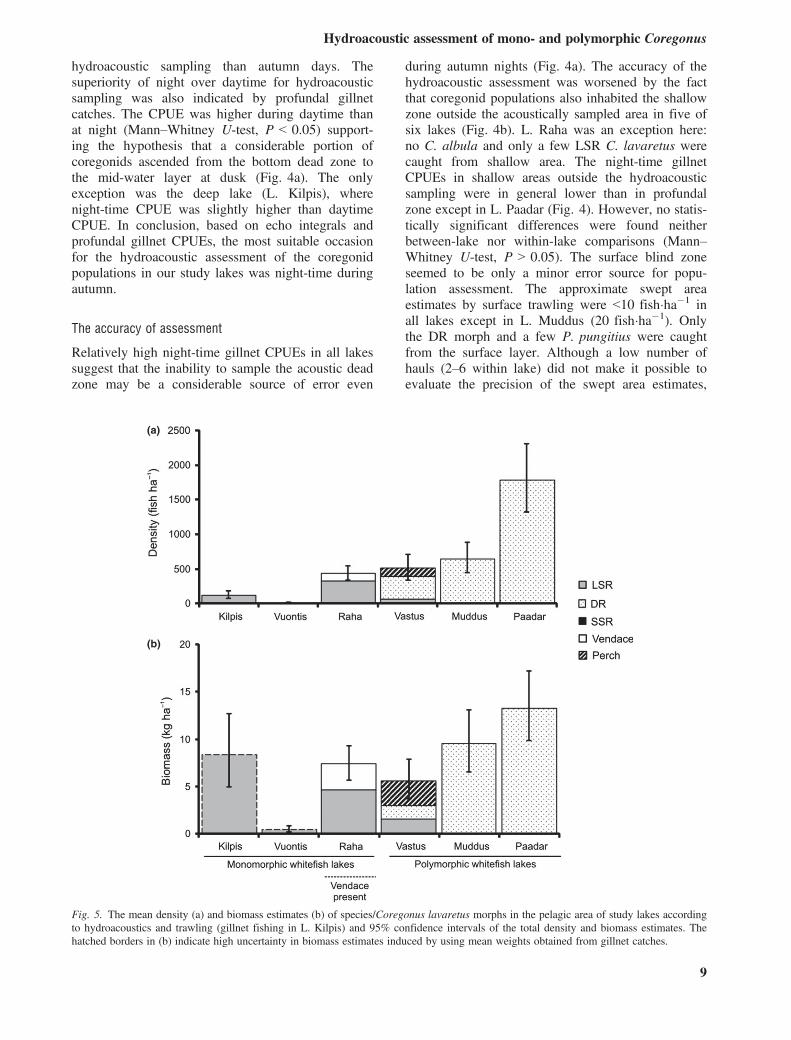
The pelagic fish density during autumn nights variedbetween lakes (Fig. 5a). In lakes with monomorphicLSR C. lavaretus (group A: Kilpis, Vuontis, Raha),the density of C. lavaretus varied from 10 to320 fish�ha�1, whereas in lakes with polymorphicC. lavaretus (group B: Vastus, Muddus, Paadar), thedensity of C. lavaretus varied from 390 to1780 fish�ha�1. The pelagic density of other specieswas negligible except C. albula in L. Raha(110 fish�ha�1) and P. fluviatilis in L. Vastus(120 fish�ha�1). In group B, the density was statisti-cally higher than in group A (Mann–Whitney U-test,P < 0.05) giving support to hypothesis that the pres-ence of specialised planktivore DR C. lavaretusyields higher overall pelagic fish density. Withingroup A, the density (LSR morph and C. albula) inL. Raha was clearly higher than in two other lakes asevidenced by nonoverlapping confidence limits.Within group B, the density was higher in L. Paadarthan in other lakes. The precision of density estimatesseemed to be acceptable based on relatively narrowconfidence limits.The between-lake variation of biomass was gener-
ally lower than that of density. In monomorphiclakes, the biomass estimate of C. lavaretus variedfrom 0.5 to 8.4 kg�ha�1, whereas in polymorphiclakes, it varied from 3.0 to 13.3 kg�ha�1 (Fig. 5b).The biomass estimates for L. Kilpis and L. Vuontisare uncertain and should be considered with cautionbecause mean weights were estimated from gillnetcatches. Thus, no statistical testing was applied forbiomass estimates. The biomass proportions of P. flu-viatilis and LSR morph in L. Vastus were higher thantheir density proportions due to their higher meanweight compared with DR morph (Table 3).
Size of coregonid populations in Lake Raha
The strong preference of coregonids for pelagic habi-tat indicated by very low gillnet catches in the shal-low areas enabled hydroacoustic estimation of theabundance of C. albula and LSR morph populationsin L. Raha. Assuming the density in <5 m deep areasto be negligible, the total population estimate in thelake was 574,000 individuals for LSR morph and196,000 individuals for C. albula. Correspondingbiomass estimates were 8300 kg for LSR morph and
4800 kg for C. albula. Hence, the total number ofcoregonids in L. Raha was ca. 0.77 million individu-als (95% confidence intervals 0.59–0.97 million indi-viduals) and the total biomass ca. 13,100 kg (95%confidence intervals 10,100–16,500 kg). These esti-mates should be considered as minimum values forpopulation sizes and biomasses, because it is obviousthat at least some fish occupied bottom dead zone ofechosounder and shallow areas.
Discussion
Hydroacoustics proved to be a suitable method forestimating density and biomass of pelagic plankti-vores (i.e., DR morph and C. albula) during autumnnights. During the daylight, and under the midnightsun, reliable estimation was not possible because cor-egonids avoided the pelagic habitat. In general, thepelagic fish density and biomass were higher in lakeswith polymorphic C. lavaretus including specialistplanktivore DR morph than in lakes inhabited bymonomorphic LSR C. lavaretus.The present study revealed the high variation of
pelagic fish density with season. In most lakes, theintegrals were much higher during autumn nightsthan light summer nights. The avoidance of the pela-gic habitat under the midnight sun is likely related tolow water temperature and zooplankton biomass(Kahilainen et al. 2005) and high predation risk dueto sufficient illumination for predators (Gjelland et al.2009; Kahilainen et al. 2009; Elliott 2011). The pela-gic fish backscattering (measured as echo integral)varied also between diel periods in September, beingsubstantially higher at night than during daytime.Thus, our hypothesis that a notable proportion of cor-egonids would inhabit the bottom dead zone during
Table 4. Summary on the accuracy of hydroacoustic density estimates ofdifferent coregonid populations applying classification by Malinen et al.(2005). Numbers indicate the most relevant bias sources.
Species/morph
LSRCoregonus lavaretus
DRC. lavaretus
SSRC. lavaretus
Coregonusalbula
Kilpis Low1 – – –Vuontis Estimation
not possible1,2– – –
Raha High – – HighVastus Low1,2,3 Low1,2,3 – –Muddus Estimation
not possible2Average1,2 Estimation
not possible1–
Paadar Estimationnot possible2
Average2 Estimationnot possible1
–
DR, densely rakered; LSR, large sparsely rakered; SSR, small sparselyrakered.1Bottom dead zone.2Shallow areas.3Confusion by other species.
10
Malinen et al.

daytime but ascend to mid-water at dusk was sup-ported. The effect of vertical migration on mid-waterfish density was pronounced in three lakes: L. Raha,Muddus and Paadar, and the echo integrals were 5–26 times higher at night than during the day. Thishighlights that in addition to well-documented verti-cal migration within mid-water (Kahilainen et al.2004, 2009; Gjelland et al. 2009), a broad switchoverfrom the profundal near-bottom layer to the mid-water layer also occurs.One could argue that horizontal migration from the
shallow area explains the increase in pelagic densityat night. However, the comparison of diurnal gillnetCPUEs in shallow areas did not give support to thispossibility (Tuomaala 2008). Because daytime surfacetrawling in L. Raha and Paadar indicated very lowfish density in the surface blind zone (T. Malinen,unpublished data), the vertical movement from thebottom dead zone is the only reasonable explanationfor the increase. From a population assessment pointof view, the situation was completely unsuitable indaytime, but adequate or even optimal (L. Raha) atnight. It should be noticed, however, that hydroacous-tics might be useful in estimation of pelagic fish den-sity even on occasion which is not representative forthe whole population. This kind of information isimportant, for instance, in studies concerning a habitatchoice or seasonal dynamics of a pelagic food web.The approximate indicators of different bias
sources made it possible to evaluate the degree ofbias in assessment of different coregonid populations(Table 4). In general, the accuracy was the highestfor C. albula and DR C. lavaretus populationsbecause of their preference for pelagic habitat. Themost suitable population for hydroacoustic assess-ment was the C. albula population in L. Raha due toits clear avoidance of the surface and bottom layersas well as shallow areas. The accuracy seemed to beacceptable also for DR morph populations of L.Muddus and L. Paadar. In these lakes, DR morphpopulation inhabits also the shallow area outside theacoustically sampled area and the bottom dead zoneof the echosounder. These potential sources of biasneed to be taken into account when interpreting thefindings of hydroacoustic surveys. Low accuracy wasfound for both benthivores (SSR and LSR morphs)with the exception of L. Raha, where the generalistLSR morph used pelagic habitat to greater extentthan other sparsely rakered populations. According tostable isotope results, monomorphic LSR C. lavar-etus in L. Kilpis and L. Vuontis used partly pelagicprey (Harrod et al. 2010), but our findings showedthey inhabited primarily the bottom layer. In lakeswith the sympatric DR morph, the LSR morph usesalmost exclusively the shallow littoral niche and thebenthivorous SSR morph the deep profundal niche
(Kahilainen et al. 2003). Both these habitats arepoorly sampled with hydroacoustics. The reliabilityof our accuracy analysis is weakened by the lowoverall benthic gillnet effort. Given the relativelylarge area of study lakes, much higher gillnet effortwould be needed if, for instance, one wanted to com-pare the degree of bias induced by shallow areas andbottom dead zone. Our data enabled only a roughcomparison of the importance of these bias sources inthe assessment of different coregonid populations.The presence of many species/morphs complicates
estimation as hydroacoustics is not suitable for theidentification of fish targets. In the present study, theproportions of species and morphs were determinedby a small pelagic trawl. As an active and relativelynonselective gear, it is thought to produce accurateinformation on species composition and sizedistributions (Olin & Malinen 2003; Olin et al. 2009;Emmrich et al. 2010). However, the present studyrevealed some circumstances, where the trawl data onspecies proportions were obviously biased. At first,in L. Vastus, many fish aggregations were observedrelatively close to the bottom along steep slopes,where trawling was practically impossible. It is likelythat in these aggregations, the proportions of P. flu-viatilis, DR morph and LSR morph were differentthan in the actual pelagic layer inducing some bias tospecies/morph-specific estimates. The bias in totaldensity and biomass estimates, however, should berelatively small. Secondly, in four lakes (L. Vuontis,Raha, Muddus and Paadar), P. pungitius was caughtonly occasionally with the trawl, but TS distributionsby echosounder revealed that these very small-bodiedfish (the average LT 33–38 mm in trawl catches)were actually highly abundant. Because P. pungitiusand coregonids produced clearly distinct peaks in TSdistributions and did not form schools at autumnnights, it was easy to exclude P. pungitius targetsbefore computing the estimate for coregonid density.When the accuracy of hydroacoustic estimation
was evaluated in estimation of total fish density andbiomass of each lake, it was considered high in L.Raha and average in L. Muddus and L. Paadar, butlow in the other three lakes. The sufficiently accurateestimates were 430 fish and 7.4 kg�ha�1 in L. Raha,640 fish and 9.5 kg�ha�1 in L. Muddus as well 1780fish and 13.3 kg�ha�1 in L. Paadar. The observeddensities in this study were generally lower than inC. albula-dominated Norwegian subarctic lakesSkrukkebukta and Vaggatem (2600–2700 fish�ha�1,Jensen et al. 2008), where biomass estimates werenot provided. It is obvious that the food resources ofthese lakes are more efficiently utilised by the intro-duced planktivore C. albula than in the lakes of thepresent study, where native C. lavaretus morphsdominate. Recent observations about the diet of
11
Hydroacoustic assessment of mono- and polymorphic Coregonus

C. albula in Lakes Skrukkebukta and Vaggatem,however, suggest that the carrying capacity of lakeshas been exceeded, and these populations may col-lapse (Liso et al. 2011) as a nearby C. albula popula-tion in Lake Inari did in 1990s (Salonen et al. 2007).In some subarctic lakes where coregonids are
missing, the pelagic habitat is dominated by theplanktivorous and small-sized morph of another poly-morphic species, S. alpinus. In more productive LakeThingvallavatn in Iceland, its density (450–1000 fish�ha�1) was comparable (Snorrason et al.1992a), but biomass clearly higher (70 kg�ha�1,Snorrason et al. 1992b) than in DR morph-dominatedstudy lakes. In southern Fennoscandian lakes, the fishdensity and biomass are naturally higher due tohigher productivity and longer growing season com-pared with subarctic lakes. In addition, southern lakeshave more diverse fish assemblages usually domi-nated by smelt Osmerus eperlanus (L.) and C. albu-la. These species have high seasonal and annualvariation of density having maximum autumn-timedensities up to 20,000 fish�ha�1 (Nyberg et al. 2001;Malinen & Tuomaala 2005) and biomasses up to75 kg�ha�1 (Jurvelius et al. 1988). The nearest com-parable estimates of C. lavaretus-dominated lakescome from Alpine and pre-alpine lakes. In large anddeep Lake Constance, C. lavaretus biomass levelswere estimated at 20–25 kg�ha�1 (Appenzeller 1996),whereas in smaller and shallower Austrian Lake Irr-see, the biomass was estimated to be ca. 35 kg�ha�1
(Gassner et al. 2003).To conclude, vertical echosounding provides a
robust tool for the density and biomass estimation ofpelagic planktivores, C. albula and DR morph ofC. lavaretus in subarctic lakes during the autumnnights, whereas for the benthivorous LSR and SSRmorphs of C. lavaretus, other estimation methodsshould be tested.
Acknowledgements
Authors would like to thank Ministry of Agriculture andForestry, County of Inari and European Regional DevelopmentFund (project A30205) as well as Jenny and Antti Wihuri,Finnish Cultural, Otto A. Malm and Emil Aaltonen Founda-tions for the financial support. Muddusj€arvi Research Stationand Kilpisj€arvi Biological Station provided excellent fieldfacilities. We are grateful to all people who participated infield work sessions during these years. We also thank Dr DanielL. Yule for constructive and valuable comments to themanuscript.
References
Ahrenstorff, T.D., Hrabik, T.R., Jacobson, P.C. & Pereira,D.L. 2013. Food resource effects on diel movements and
body size of cisco in north-temperate lakes. Oecologia: DOI:10.10077s00442-013-2719-3 (in press).
Amundsen, P.-A., Staldvik, F.J., Reshetnikov, Y.S., Kashulin,N., Lukin, A., Bøhn, T., Sandlund, O.T. & Popova, O.A.1999. Invasion of vendace Coregonus albula in a subarcticwatercourse. Biological Conservation 88: 405–413.
Amundsen, P.-A., Knudsen, R., Klemetsen, A. & Kristoffersen,R. 2004. Resource competition and interactive segregationbetween sympatric whitefish morphs. Annales ZoologiciFennici 41: 301–307.
Appenzeller, A.R. 1995. Hydroacoustic measurement ofspatial heterogeneity of European whitefish (Coregonuslavaretus) and perch (Perca fluviatilis) in Lake Constance.Archiv f€ur Hydrobiologie Special Issues Advanced Limnol-ogy 46: 265–275.
Appenzeller, A.R. 1996. Seasonal variability patterns of acous-tic estimates of fish biomass and fish size in a lake dominatedby coregonids (Lake Constance). Archiv f€ur HydrobiologieSpecial Issues Advanced Limnology 50: 227–236.
Bøhn, T. & Amundsen, P.-A. 2001. The competitive edge ofan invading specialist. Ecology 82: 2150–2163.
Bøhn, T., Amundsen, P.-A. & Sparrow, A. 2008. Competitiveexclusion after invasion? Biological Invasions 10: 359–368.
Clark, C.W. & Levy, D.A. 1988. Diel vertical migrations byjuvenile sockeye salmon and the antipredation window.American Naturalist 131: 271–290.
Cochran, W.G. 1977. Sampling techniques. New York, NY:John Wiley and Sons.
Conover, W.J. 1980. Practical nonparametric statistics. NewYork, NY: John Wiley and Sons.
Eckmann, R. 1991. A hydroacoustic study of the pelagicspawning behaviour of whitefish (Coregonus lavaretus) inLake Constance. Canadian Journal of Fisheries and AquaticSciences 48: 995–1002.
Eckmann, R. 1998. Allocation of echo integrator output tosmall larval insect (Chaoborus sp.) and medium-sized (juve-nile fish) targets. Fisheries Research 35: 107–113.
Elliott, J.M. 2011. A comparative study of the relationshipbetween light intensity and feeding ability in brown trout(Salmo trutta) and Arctic charr (Salvelinus alpinus). Fresh-water Biology 56: 1962–1972.
Emmrich, M., Helland, I.P., Busch, S., Schiller, S. & Mehner,T. 2010. Hydroacoustic estimates of fish densities in com-parison with stratified pelagic trawl sampling in two deep,coregonid-dominated lakes. Fisheries Research 105: 178–186.
Enderlein, O. & Appelberg, M. 1992. Sampling stocks ofCoregonus lavaretus and C. albula with pelagic gillnets,trawls and hydroacoustic gear. Polskie Archiwum Hydrobio-logii 39: 771–777.
Gassner, H., Tischler, G. & Wanzenb€ock, J. 2003. Ecologicalintegrity assessment of lakes using fish communities – sug-gestions of new metrics developed in two Austrian prealpinelakes. International Review of Hydrobiology 88: 635–652.
Gjelland, K.Ø. 2008. Ecological interactions, light responsesand vertical habitat use in a subarctic pelagic freshwatercommunity. PhD thesis. Department of Aquatic BioScienc-es. University of Tromsø. 68 pp.
Gjelland, K.Ø., Bøhn, T., Horne, J.K., Jensvoll, I., Knud-sen, F.R. & Amundsen, P.-A. 2009. Planktivore verticalmigration and shoaling under a subarctic light regime.
12
Malinen et al.

Canadian Journal of Fisheries and Aquatic Sciences 66:525–539.
Harrod, C., Mallela, J. & Kahilainen, K.K. 2010. Phenotype-environment correlations in a putative whitefish adaptiveradiation. Journal of Animal Ecology 79: 1057–1068.
Hendry, A.P. 2009. Ecological speciation! Or the lack thereof?Canadian Journal of Fisheries and Aquatic Sciences 66:1383–1398.
Jensen, H., Kahilainen, K.K., Amundsen, P.-A., Gjelland,K.Ø., Tuomaala, A., Malinen, T. & Bøhn, T. 2008. Preda-tion by brown trout (Salmo trutta) along a diversifying preycommunity gradient. Canadian Journal of Fisheries andAquatic Sciences 65: 1831–1841.
Jolly, G.M. & Hampton, I. 1990. Some problems in thestatistical design and analysis of acoustic surveys to assessfish biomass. Rapports et Proc�es-verbaux des R�eunionsConseil International pour l’Exploration de la Mer 189:415–420.
Jurvelius, J., Lindem, T. & Heikkinen, T. 1988. The size of avendace, Coregonus albula L., stock in a deep lake basinmonitored by hydroacoustic methods. Journal of Fish Biol-ogy 32: 679–687.
Kahilainen, K. & Lehtonen, H. 2002. Brown trout (Salmo tru-tta L.) and Arctic charr (Salvelinus alpinus (L.)) as predatorson three sympatric whitefish (Coregonus lavaretus (L.))forms in the subarctic Lake Muddusj€arvi. Ecology of Fresh-water Fish 11: 158–167.
Kahilainen, K. & Lehtonen, H. 2003. Piscivory and preyselection of four predator species in a whitefish domi-nated subarctic lake. Journal of Fish Biology 63: 659–672.
Kahilainen, K. & Østbye, K. 2006. Morphological differentia-tion and resource polymorphism in three sympatric whitefishCoregonus lavaretus (L.) forms in a subarctic lake. Journalof Fish Biology 68: 63–79.
Kahilainen, K., Lehtonen, H. & K€on€onen, K. 2003. Conse-quence of habitat segregation to growth rate of twosparsely rakered whitefish (Coregonus lavaretus (L.))forms in a subarctic lake. Ecology of Freshwater Fish 12:275–285.
Kahilainen, K., Malinen, T., Tuomaala, A. & Lehtonen, H.2004. Diel and seasonal habitat and food segregation ofthree sympatric Coregonus lavaretus forms in a subarcticlake. Journal of Fish Biology 64: 418–434.
Kahilainen, K., Alaj€arvi, E. & Lehtonen, H. 2005. Planktivoryand diet-overlap of densely-rakered Coregonus in a subarcticlake. Ecology of Freshwater Fish 14: 50–58.
Kahilainen, K.K., Malinen, T., Tuomaala, A., Alaj€arvi, E.,Tolonen, A. & Lehtonen, H. 2007. Empirical evaluation ofphenotype-environment correlation and trait utility with allo-patric and sympatric whitefish (Coregonus lavaretus (L.))populations in subarctic lakes. Biological Journal of the Lin-nean Society 92: 561–572.
Kahilainen, K.K., Malinen, T. & Lehtonen, H. 2009. Polarlight regime and piscivory govern diel vertical migrations ofplanktivorous fish and zooplankton in a subarctic lake. Ecol-ogy of Freshwater Fish 18: 481–490.
Kahilainen, K.K., Siwertsson, A., Gjelland, K.Ø., Knudsen,R., Bøhn, T. & Amundsen, P.-A. 2011a. The role of gillraker number variability in adaptive radiation of coregonidfish. Evolutionary Ecology 25: 573–588.
Kahilainen, K.K., Østbye, K., Harrod, C., Shikano, T.,Malinen, T. & Meril€a, J. 2011b. Species introductionpromotes hybridization and introgression in Coregonus: isthere sign of selection against hybrids? Molecular Ecology20: 3838–3855.
Linløkken, A. 1995. Monitoring pelagic whitefish (Coregonuslavaretus) and vendace (Coregonus albula) in a hydroelec-tric reservoir using hydroacoustics. Regulated Rivers-Research and Management 10: 315–328.
Liso, S., Gjelland, K.Ø., Reshetnikov, Y.S. & Amundsen, P.-A. 2011. A planktivorous specialist turns rapacious: pisci-vory in invading vendace Coregonus albula. Journal of FishBiology 78: 332–337.
MacLennan, D.N. & Simmonds, E.J. 1992. Fisheries acous-tics. London: Chapman & Hall.
MacLennan, D.N., Fernandes, P.G. & Dalen, J. 2002. A con-sistent approach to definitions and symbols in fisheriesacoustics. ICES Journal of Marine Science 59: 365–369.
Malinen, T. & Tuomaala, A. 2005. Comparison of day andnight surveys in hydroacoustic assessment of smelt (Osmer-us eperlanus) density in Lake Hiidenvesi. Archiv f€ur Hy-drobiologie Special Issues Advanced Limnology 59: 161–172.
Malinen, T., Tuomaala, A. & Peltonen, H. 2005. Vertical andhorizontal distribution of smelt (Osmerus eperlanus) andimplications of distribution patterns on stock assessment.Archiv f€ur Hydrobiologie Special Issues AdvancedLimnology 59: 141–159.
Marjom€aki, T.J. & Huolila, M. 1995. Monitoring the densityof Lake Puulavesi vendace (Coregonus albula (L.)) byhydroacoustics, catch per unit effort, virtual population andcatch per swept area. Archiv f€ur Hydrobiologie SpecialIssues Advanced Limnology 46: 267–276.
Mehner, T. 2012. Diel vertical migration of freshwater fishes– proximate triggers, ultimate causes and research perspec-tives. Freshwater Biology 57: 1342–1359.
Nyberg, P., Bergstrand, E., Degerman, E. & Enderlein, O.2001. Recruitment of pelagic fish in an unstable climate:studies in Sweden’s four largest lakes. Ambio 30: 559–564.
Olin, M. & Malinen, T. 2003. Comparison of gillnet and trawlin diurnal fish community sampling. Hydrobiologia 506–509: 443–449.
Olin, M., Malinen, T. & Ruuhij€arvi, J. 2009. Gillnet catch inestimating the density and structure of fish community –comparison of gillnet and trawl samples in eutrophic lakes.Fisheries Research 96: 88–94.
Østbye, K., Bernatchez, L., Næsje, T.F., Himberg, K.-J.M. &Hindar, K. 2005. The evolutionary history of Europeanwhitefish (Coregonus lavaretus L.) as inferred from mtDNAphylogeography and gillraker numbers. Molecular Ecology14: 4371–4387.
Østbye, K., Amundsen, P.-A., Bernatchez, L., Klemetsen, A.,Knudsen, R., Kristoffersen, R., Næsje, T.F. & Hindar, K.2006. Parallel evolution of ecomorphological traits in theEuropean whitefish Coregonus lavaretus (L.) species complexduring postglacial times. Molecular Ecology 15: 3983–4001.
Salonen, E., Amundsen, P.-A. & Bøhn, T. 2007. Invasion,boom and bust by vendace (Coregonus albula) in thesubarctic Lake Inari, Finland and the Pasvik watercourse,Norway. Archiv f€ur Hydrobiologie Special Issues AdvancedLimnology 60: 331–342.
13
Hydroacoustic assessment of mono- and polymorphic Coregonus

Sandlund, O.T., Næsje, T.F., Klyve, L. & Lindem, T. 1985.The vertical distribution of fish species in Lake Mjøsa,Norway, as shown by gill-net catches and echo sounder.Report of the Institute of Freshwater Research Drottning-holm 62: 136–149.
Scheuerell, M.D. & Schindler, D.E. 2003. Diel verticalmigration by juvenile sockeye salmon: empirical evidencefor the antipredation window. Ecology 84: 1713–1720.
Schluter, D. & McPhail, J.D. 1993. Character displacementand replicate adaptive radiation. Trends in Ecology and Evo-lution 8: 197–200.
Shotton, R. & Bazigos, G.P. 1984. Techniques and consider-ations in the design of acoustic surveys. Rapports etProc�es-verbaux des R�eunions Conseil International pourl’Exploration de la Mer 184: 34–57.
Siwertsson, A., Knudsen, R., Kahilainen, K.K., Præbel, K.,Primicerio, R. & Amundsen, P.-A. 2010. Sympatric diversi-fication influenced by ecological opportunity and historical
contingency in a young species lineage of whitefish. Evolu-tionary Ecology Research 12: 929–947.
Snorrason, S.S., J�onasson, P.M., Jonsson, B., Lindem, T.,Malmquist, H.J., Sandlund, O.T. & Sk�ulason, S. 1992a. Pop-ulation dynamics of the planktivorous arctic charr Salvelinusalpinus (“murta”) in Thingvallavatn. Oikos 64: 352–364.
Snorrason, S.S., Sandlund, O.T. & Jonsson, B. 1992b. Produc-tion of fish stocks on Thingvallavatn, Iceland. Oikos 64:371–380.
Tuomaala, A. 2008. Siikamuotojen habitaatin valinta ja kaik-uluotauksen soveltuvuus niiden kannan koon arviointiinsubarktisissa j€arviss€a. M.Sc. thesis. Department of Biologicaland Environmental Sciences. University of Helsinki. 45 pp(in Finnish).
Yule, D.L., Evrard, L.M., Cachera, S., Colon, M. & Guillard,J. 2013. Comparing two fish sampling standards over time:largely congruent results but with caveats. Freshwater Biol-ogy 58: 2065–2073.
14
Malinen et al.




















